
Abstract
The FOX transcription factor family plays a pivotal role in the malignant progression of tumors. We propose a hypothesis that FOXQ1 recruits p300 and BRD4 to super-enhancer regions. Our findings indicate that p300 acetylates Lys190 of FOXQ1, resulting in its recognition and binding by BRD4. Subsequently, BRD4 recruits RNA-Pol II to form a “FOXQ1-p300-BRD4-RNA Pol II” complex, which then binds to the super-enhancers of target genes. Meanwhile, acetylation at Lys190 of FOXQ1 directly enhances its binding affinity to super-enhancers. Consequently, more target oncogenes can be transcribed to promote CRC proliferation and metastasis. Our results suggest that FOXQ1 acts as a key regulator of super-enhancers, providing insights into its role in CRC and highlighting its potential as a therapeutic target.
Similar content being viewed by others
Introduction
The FOX transcription factor family comprises about 50 members and is divided into 19 sub-cohorts, labeled FOXA to FOXS in humans1,2. These proteins all possess a DNA-binding domain, known as the forkhead domain, or winged helix3. The FOX transcription factor family members paly roles in various cellular processes such as cell differentiation, proliferation, apoptosis, aging, embryonic development, metabolism, and immune regulation4. Despite ongoing research, the specific biological functions of mammalian forkhead transcription factors of Q class are still not yet clear.
FOXQ1 is the only member of the forkhead transcription factors of Q class. The FOXQ1 gene is situated on human chromosome 6p25.35, and consists of three structural domains: the alanine and glycine enrichment region, the forkhead box domain (FHD), and the proline enrichment region. The FHD, also known as winged helix domain, serves as a DNA-binding domain4. The other two domains are responsible for binding transcriptional activators/repressors and DNA repair complexes4. FOXQ1 functions as a transcription factor and plays a vital role in various biological processes such as inhibiting smooth muscle differentiation, promoting epithelial differentiation, controlling mucin gene expression, regulating granule content in stomach surface mucous cells, activating T cells and autoimmunity6,7,8. However, further research is needed to fully understand the detailed mechanism of its function. Despite abnormal regulation of FOXQ1 in various tumors, the specific mechanism of FOXQ1 in tumorigenesis and progression remains incompletely elucidated.
An increasing number of studies have demonstrated the significant regulatory role of acetylation in various genes. Specifically, AGO2 was found be acetylated by p300/CBP at K355, K493, and K720, which enhanced its oncogenic activity. HDAC7 was identified as a blocker of this acetylation process9. Similarly, SKP2 was found to be acetylated at K68 and K71 by p300, and deacetylated by SIRT3. This acetylation of SKP2 was shown to interfere with CDH1-mediated proteolysis and promote cancer cells migration, partially involving E-cadherin, ultimately enhancing SKP2’s oncogenic potential10. The impact of acetylation on the DNA-binding capacity of FOXO1 has also been documented11, a phenomenon that aligns with our findings on FOXQ1.
Super-enhancers, unlike ordinary enhancers, are characterized by their large size, spanning over 8 kb, and their ability to interact with a diverse range of tissue-specific transcription factors (such as Oct4, Sox2, and Nanog in embryonic stem cells), mediator complex (Med), chromatin regulator and RNA polymerase II complex in cells12. The concentration of those transcription factors is significantly higher than ordinary enhancers, resulting in a greater capacity to enhance the transcription of target genes13.
In our study, we observed that Lys190 of FOXQ1 is acetylated by p300, leading to its recognition and binding by BRD4. Our findings indicate that BRD4 facilitates the recruitment of RNA-Pol II, resulting in the formation of the “FOXQ1-p300-BRD4-RNA-Pol II” complex. This complex can target the super enhancers of various oncogenes, ultimately boosting their transcription and contributing to the malignant progression of CRC. Through our investigation, we delved into the impact of FOXQ1 on CRC proliferation and metastasis, as well as explored the clinicopathological significance of FOXQ1 in CRC progression.
Results
Abnormally high expression of FOXQ1 in CRC
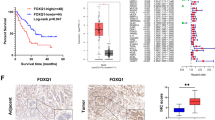
Transcriptome sequencing results revealed FOXQ1 as the most significantly up-regulated transcription factor within FOX transcription factor family in CRC tissues compared to normal colorectal tissues (Fig. 1A). Immunohistochemistry (IHC) staining confirmed a significant upregulation of FOXQ1 in CRC (Fig. 1B), with higher expression levels observed in CRC tissues compared to their normal counterparts (n = 415, pairs) (Fig. 1C). Analysis of clinicopathological characteristics in CRC patients (summarized in Supplementary Table 1) indicated that 55% (231/415) exhibited high FOXQ1 expression, while 45% (184/415) had low expression levels. High FOXQ1 expression was found to be correlated positively with tumor size (p < 0.001), TNM stage (p < 0.001), lymph node metastasis (p = 0.001), and depth of invasion (p = 0.014), but not with age (p = 0.068), sex (p = 0.093), or tumor differentiation (p = 0.568). Survival analysis showed that patients with high FOXQ1 expression demonstrated worse Overall Survival (OS) and Disease-free survival (DFS) compared to those with low FOXQ1 expression (Fig. 1D, E). Univariate analysis revealed a significant association between FOXQ1 and poor OS (hazard ratio [HR], 2.432; 95% confidence interval [CI], 1.649–3.587; p < 0.001) (Supplementary Table 2), While multivariate analysis identified FOXQ1 expression as an independent prognostic marker for OS (HR, 2.111; 95%CI, 1.529–3.533; p < 0.001) (Supplementary Table 3).

A 6 pairs of colorectal cancer (C) and non-cancerous (N) tissues were collected from the Jiangsu Cancer Hospital. RNA-seq was used to analyze the mRNA expression levels of FOX family in colorectal cancer and non-cancerous tissues. The RNA-seq analysis methodologies are detailed in the Materials and Methods section. B Immunohistochemistry images of FOXQ1 protein expression in carcinoma and non-cancerous tissues in CRC patients collected from the Affiliated Hospital of Xuzhou Medical University. C Staining intensities of FOXQ1 in colorectal cancer tissues compared with paired adjacent noncancerous tissues collected from the Affiliated Hospital of Xuzhou Medical University. D, E Kaplan–Meier survival curves depicting overall survival (OS) and Disease-free survival (DFS) in colorectal cancer tissues collected from the Affiliated Hospital of Xuzhou Medical University. F Analysis of genes positively or negatively correlated with FOXQ1 expression in TCGA database. G Biological process (BP), Cellular component (CC), Molecular function (MF) and KEGG analysis results of genes positively expressed with FOXQ1 in TCGA database. Data are mean ± SDs. *p < 0.05, **p < 0.01, ***p < 0.001.
Next, we utilized the LinkedOmics (https://www.linkedomics.org/login.php) database to analyze genes related to FOXQ1 (Fig. 1F). Genes exhibiting positive correlation with FOXQ1 were selected for Biological Process (BP), Cellular Component (CC), Molecular Function (MF), and KEGG analysis (Fig. 1G). These genes were enriched in oncogenic functions such as cell-substrate adhesion, response to transforming growth factor beta, and various signaling pathways linked to tumorigenesis and tumor development, including Wnt, Hippo, and TGF-beta. These findings supported the potential of FOXQ1 expression as an independent prognostic factor for patients with CRC.
FOXQ1 is acetylated at Lys190 within its FHD domain by p300
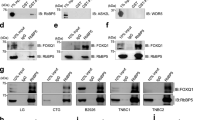
Acetylation of certain transcription factors have been shown to enhance their ability in promoting tumor progression14. In our study, we observed that endogenous FOXQ1 underwent acetylation modification in various CRC cell lines (Fig. 2A). Through FpClass (http://dcv.uhnres.utoronto.ca/FPCLASS/ppis/) analysis, we found that the combination score between p300 and FOXQ1 was the highest (Fig. 2B). Furthermore, Co-immunoprecipitation (CoIP) analysis of total proteins from CRC cells showed that FOXQ1 and p300 were efficiently co-immunoprecipitated with each other, indicating their physical association in vivo (Fig. 2C). In addition, only p300 overexpression significantly increased the acetylation of FOXQ1, but not other acetyltransferases, such as GCN5, Tip60, and PCAF (Fig. 2D). Treatment with A485, a specific inhibitor of p300, or p300 knockdown both caused a decrease in endogenous FOXQ1 acetylation level (Fig. 2E–G). In vitro experiments also proved that p300 directly acetylated FOXQ1 (Fig. 2H). These results suggest that p300 acts as a physiological acetyltransferase for FOXQ1.

A In HCT-116, Lovo, DLD1 and SW480 cells, FOXQ1 and pan-acetylated-lysine were immunoprecipitated (IP) and examined by western blotting. B Analysis of proteins potentially binding to FOXQ1 using FpClass. C CoIP was used to detect interactions between FOXQ1 and p300. D FOXQ1 was immunoprecipitated and detected for acetylation levels in HEK-293T cells overexpressing p300, GCN5, PCAF and Tip60. E, F HCT-116 and Lovo cells were treated with DMSO or indicated concentration of A485 for 4 h, then acetylation changes of FOXQ1 were detected by immunoprecipitation and western blotting. G Anti-FOXQ1 IPs derived from HCT-116 cells transfected with sip300 or siCTR as negative control, and then the acetylation of FOXQ1 was examined by western blotting. H In vitro acetylation assay using purified Flag-FOXQ1 incubated with p300 in the presence or absence of Acetyl-CoA or A485. I IB analysis of IP products derived from HEK-293T cells transfected with FOXQ1 or KR mutants. J In vitro acetylation dot blotting assay using purified peptides incubated with p300 in the presence or absence of Acetyl-CoA or A485. K In vitro acetylation assay using purified Flag-FOXQ1 incubated with p300 in the presence or absence of Acetyl-CoA to examine the acetylation changes of FOXQ1 Lys190. L Flag-FOXQ1(WT) and Flag-FOXQ1(K190R) were transfected, respectively, into HCT-116 cells. The cytoplasm and nucleus were isolated, and the localization of both in the cells was detected. M DNA pull down was used to detect the effect of K190R on its affinity to the specific binding sequence (TGTTTATGTTTAGTTT) of FOXQ1. N Flag-foxQ1 (WT) and FLAG-K190R (K190R) constructs were transiently transfected into HCT-116 cells, respectively. Subsequently, chromatin immunoprecipitation (ChIP) assays were performed using anti-Flag antibodies to investigate the impact of mutation of FOXQ1-K190 on its interaction with the promoters of SIX2, ZEB1, and STAT2. O The effects of both the acetylated FOXQ1-K190 peptide and the non-acetylated FOXQ1-K190 peptide on their ability to bind to the promoters of SIX2, ZEB1, and STAT2 were investigated using the ChIP assay.
Utilizing data from PhosphoSitePlus (https://www.phosphosite.org), deep-PLA (http://deeppla.cancerbio.info/webserver.php) and CUCKOO (http://biocuckoo.c/) databases, our analysis predicted the potential acetylation modifications sites of FOXQ1 as Lys2, Lys190, Lys266, and Lys360. Substituting these lysine (K) residues with arginine (R) we observed that K190R, situated in the DNA binding domain (119-214), and K360R showed the more reduced level of acetylation compared with FOXQ1-WT (Fig. 2I). Through homology alignment analysis, we found that Lys190 of FOXQ1 has higher conservation than Lys360 (Supplementary Fig. 1A). Using a synthetic peptide of FOXQ1 amino acids 185–195 (SRPWGKDNYWM), we confirmed the acetylation of Lys190 by p300 via dot blot analysis (Fig. 2J). In vitro acetylation experiment showed that p300 can directly acetylate Lys190 using specific acetylation antibody (Fig. 2K). Subcellular fractionation analysis demonstrated that the FOXQ1-K190 mutation did not alter its cellular localization (Fig. 2L). Next, we examined whether the FOXQ1-K190 acetylation modulates its DNA-binding capacity. Consistent with prior studies, FOXQ1 has been shown to target the promoters of SIX2, ZEB1, and STAT2, thereby promoting their transcription15. DNA pull down results showed that mutation of Lys190 of FOXQ1 to arginine weakened its affinity to the specific binding sequence (TGTTTATGTTTAGTTT) (Fig. 2M). Compared with FOXQ1-WT, FOXQ1-K190R exhibited reduced recruitment of target gene promoters (Fig. 2N) and the non-acetylated FOXQ1-K190 peptide (CRDPSRPWGKDNYWM) bound significantly less FOXQ1 target gene promoters compared to the acetylated FOXQ1-K190 peptide (CRDPSRPWGK(ac)DNYWM) (Fig. 2O). Taken together, FOXQ1 Lys190 can be acetylated by p300, and this acetylation promotes FOXQ1’s ability to bind to DNA.
BRD4 can recognize and bind to Lys190-acetylated FOXQ1
To identify FOXQ1-interacting proteins in CRC, we transfected HCT-116 cells with Flag-FOXQ1, and performed Flag-FOXQ1 complexes purification, and mass spectrometry (Fig. 3A). One of the most prevalent post-translational modifications in proteins is lysine acetylation (Kac). A specific class of “reader” proteins, which contain bromodomain (BD) motifs, recognize and bind to acetylated lysine residues. Among the 46 BD-containing proteins identified in humans, BRD2, BRD3, BRD4, and Bromodomain testis-specific protein (BRDT) have been extensively investigated as members of the BET family in relation to their roles in physiology and disease16,17,18. Through mass spectrometry results, we identified that among the BET family members, only BRD4 has the potential to interact with FOXQ1. Subsequent CoIP analysis confirmed the interaction between endogenous FOXQ1 and BRD4, as well as the acetylation of endogenous FOXQ1 in HCT-116 and Lovo cells. Interestingly, FOXQ1 exhibited higher acetylation levels and stronger interaction with BRD4 when treated with the Trichostatin (TSA, histone deacetylase inhibitor) (Fig. 3B). Additionally, the ectopic expression of p300 in HCT-116 and Lovo cells led to enhanced acetylation of FOXQ1 and improved association of FOXQ1 with BRD4 even in the absence of TSA, whereas p300 knockdown yielded the opposite effects even in the presence of TSA (Fig. 3C). Notably, the K190R recruited less BRD4 compared to the wild type (Fig. 3D). The results above indicated that BRD4 specifically recognizes and binds to Lys190-acetylated FOXQ1.

A Schematic illustration for the detection of FOXQ1-binding proteins by immunoprecipitation and protein mass spectrometry. B HCT-116 cells were treated with TSA as in Fig. 2E. FOXQ1, endogenous BRD4 and acetylated FOXQ1 were immunoprecipitated and examined by western blotting. C Ectopic expression of p300 or knockdown of endogenous p300 were performed in HCT-116 cells. After endogenous FOXQ1 was immunoprecipitated, acetylation of FOXQ1 and the bound BRD4 were examined by western blotting. D HEK-293T cells were transfected with Vector, FOXQ1-wild-type (WT) or FOXQ1-K190R, then Flag tag was immunoprecipitated, followed by Flag-FOXQ1 and the bound BRD4 were examined by western blotting. E Schematic depiction of the functional domains and deletion constructs used for BRD4 and FOXQ1. F, G. Flag-tagged WT or deletion mutants of BRD4 and FOXQ1 were expressed in HEK293 cells. After being immunoprecipitated with Flag antibody, the bound FOXQ1, BRD4 or p300 was examined by western blotting. H HEK-293T cells were transfected with Vector, Flag-tagged FOXQ1-WT or FOXQ1-K190R and lysed. GST-BD1 was incubated with cell lysate. The binding of GST-BD1 to FOXQ1-WT or FOXQ1-K190R was detected by western blotting.
To identify the interaction domain of the FOXQ1-p300-BRD4 complex, we constructed protein deletion constructs of BRD4 and FOXQ1 (Fig. 3E), which were transfected into HCT-116 cells. Further CoIP analysis indicated that FOXQ1 binds to the BD1 domain of BRD4 (Fig. 3F). BRD4 binds to the FHD domain, while p300 can bind to both FHD and pro rich domains of FOXQ1 (Fig. 3G). Moreover, Lys190R mutant impaired the strong binding between FOXQ1 and BD1 domain (Fig. 3H). Taken together, these results demonstrated that BRD4 recognizes and binds to Lys190 acetylated FOXQ1.
HDAC1 specially interacts with and deacetylates FOXQ1
Based on the mass spectrometry data, HDAC1 was found to potentially bind to FOXQ1 (Supplementary Fig. 2A), acting as an “eraser” for acetylation19. Physical interaction between FOXQ1 and HDAC1 was observed in HCT-116 and Lovo cell lines (Supplementary Fig. 2B, C). Meanwhile, HDAC1 knockdown or TSA treatment increased the acetylation of FOXQ1 (Supplementary Fig. 2D, E). In vitro acetylation-deacetylation experiments showed that p300-induced acetylation of Lys190 was reversed by HDAC1 (Supplementary Fig. 2F). These findings suggested that HDAC1 functions as a deacetylase for FOXQ1 at Lys190.
FOXQ1 complex binds to super-enhancers of a cohort of oncogenes
To investigate the function of FOXQ1 complex in CRC development, we analyzed the distribution and densities of FOXQ1 on chromatin (Fig. 4A, B). To substantiate the argument that the “FOXQ1-p300-BRD4” is localized at super-enhancers, Rank Ordering of Super-Enhancers (ROSE) was utilized to identify super-enhancers. With a ROSE-estimated optimal cutoff of the FOXQ1, a total of 68 super-enhancers were identified (Fig. 4C). FOXQ1 super-enhancer, p300 and BRD4 peaks are intersected, and the intersecting peaks are annotated (Fig. 4D). FCGR2A and MIR21 are among the annotated genes. FCGR2A is identified as an oncogene in various tumor types20,21,22. Utilizing the TCGA database, we observed a higher expression level of FCGR2A in colorectal cancer tissues compared to adjacent tissues (Supplementary Fig. 3A). Furthermore, there was a positive correlation between FCGR2A expression and stage T, but not stage N or M (Supplementary Fig. 3B-D). Patients exhibiting elevated FCGR2A expression experienced poorer overall survival rates (OS) (Supplementary Fig. 3E). These findings strongly suggest that FCGR2A may exert a promoting role in colorectal cancer development. The expression of MIR21 is also significantly upregulated in colorectal cancer tissues compared to paracancer tissues (Supplementary Fig. 3F), and it has been reported as an oncogene in colorectal cancer23. The mRNA levels of FCGR2A and MIR21 were significantly upregulated upon overexpression of FOXQ1, but this promoting effect was partially counteracted by simultaneous knockdown of p300 or BRD4 (Fig. 4E). In summary, p300 and BRD4 may collaboratively regulate super-enhancers with FOXQ1.

ChIP-seq reads were mapped to the hg19 human genome using bowtie2. Samtools was used to convert sam files to bam format. The significance of enriched ChIP regions was evaluated by using MACS2. Intervene was used for determining the overlapped peaks. The R package ChIPseeker were used to analyze FOXQ1 peaks distribution (A) and Tag densities (B) in chromatin. C Super-enhancers based on FOXQ1 occupancy using the ROSE algorithm. D The Venn diagram of overlapping peaks targeted by FOXQ1 super enhancers, p300 and BRD4 using the VennDiagram. E HCT-116 cells were transfected with siRNAs of control, p300, or BRD4 together with empty vector or FOXQ1 expression constructs. The mRNA level of FCGR2A and MIR21 were measured by real-time RT-PCR. Data are mean ± SDs. *p < 0.05, **p < 0.01, ***p < 0.001.
FOXQ1 acetylation at Lys190 is required for its binding to super-enhancers
In order to further study the regulatory function of FOXQ1 on super-enhancer, the chromatin distribution of FOXQ1 and H3K27ac was analyzed, considering that H3K27ac is a well-established marker for super-enhancers. Importantly, both FCGR2A and MIR21 super-enhancer regions [particularly FCGR2A: constituents Enhancer 1 (E-1) and Enhancer 2 (E-2), MIR21: constituents Enhancer A (E-A) and Enhancer B (E-B)] were co-occupied by FOXQ1 and H3K27ac (Fig. 5A). ChIP-qPCR and ChIP-reChIP experiments demonstrated the enrichment of FOXQ1, p300, and BRD4 can be observed in E-1, E-2, E-A, and E-B (Fig. 5B, Supplementary Fig. 4A–C). We also found that the enrichment of FOXQ1 at the super-enhancers (E-1, E-2, E-A, and E-B) was significantly reduced following p300 silencing, while depletion of HDAC1 was associated with enhanced recruitment of FOXQ1 at super-enhancers (Fig. 5C). Luciferase reporter assays revealed a significant reduction in enhancer activity after knocking down p300, whereas silencing of HDAC1 resulted in the opposite effect (Fig. 5D). Compared with FOXQ1-WT, K190R exhibited less enrichment at the super-enhancers (Fig. 5E) and less promoted the activity of the super-enhancers (Fig. 5F).

A Occupancy profiles of FOXQ1 and H3K27ac at the FCGR2A and MIR21 super-enhancer regions. B Occupancy of the E-1, E-2, E-A and E-B by FOXQ1, p300 and BRD4. Soluble chromatin from MCF-7 cells was prepared for ChIP and Re-ChIP assays with antibodies against the indicated proteins. C HCT-116 cells transfected with the siCTR, sip300 or siHDAC1 were collected and ChIP-qPCR experiments were performed with FOXQ1 antibody. D E-1, E-A, E-A and E-B activities are measured in HCT-116 cells transfected with the Vector, FOXQ1-WT or FOXQ1-K190R by luciferase reporter assays. (E) HCT-116 cells transfected with Vector, Flag-tagged FOXQ1-WT or FOXQ1-K190R were collected and ChIP-qPCR experiments were performed with Flag antibody. F E-1, E-A, E-A and E-B activities are measured in HCT-116 cells transfected with the Vector, FOXQ1-WT or FOXQ1-K190R by luciferase reporter assays. G Assessment of the effect of endogenous BRD4 knockdown or overexpression on the association of FOXQ1 and RNA-Pol II in HCT-116 cells. H HCT-116 cells transfected with the siCTR or siBRD4 were collected and ChIP-qPCR experiments were performed with RNA-Pol II antibodies. Data are mean ± SDs. *p < 0.05, **p < 0.01, ***p < 0.001.
Recruitment and activation of RNA-Pol II at gene promoters and enhancers are known as two key steps required for productive transcription24. Our study implied that the recruitment of RNA-Pol II is facilitated in part by BRD4. CoIP analysis showed a decrease in the amount of RNA-Pol II bound by FOXQ1 after BRD4 knockdown (Fig. 5G). Furthermore, the enrichment of RNA-Pol II at both super-enhancers decreased significantly upon silencing BRD4 (Fig. 5H). Taken together, these results supported the hypothesis that FOXQ1 acetylation at K190 is required for its binding with super enhancers, thereby facilitating the transcription of target genes.
Acetylation at Lys190 in FOXQ1 promotes the proliferation and metastasis of CRC partly though a cohort of oncogenes
A mutation at FOXQ1-K190 or treatment of A485 also weakened the promotion effect of FOXQ1 on migration, invasion, proliferation, and EMT in HCT-116 and Lovo cell lines (Fig. 6A–C). Subsequent in vivo experiments were conducted to validate the aforementioned findings. Utilizing subcutaneous HCT-116 tumor-bearing nude mouse model, it was observed that overexpression of FOXQ1 promoted tumor growth, while a mutation at Lys190 attenuated this effect (Fig. 6D–F). In a lung metastasis mouse model, overexpression of FOXQ1 significantly increased the number of lung metastases, which was partially reversed by a mutation at Lys190 (Fig. 6G, H). These results collectively suggested that FOXQ1 promotes the proliferation and metastasis of CRC in part through acetylation at Lys190.

A Assessment of cells motility by migration assays, invasion assays in HCT-116 and Lovo cells transfected with Vector, FOXQ1-WT, FOXQ1-K190R or FOXQ1 with A485 treatment (1 μM, 24 h), scale bar 200 μm. B Assessment of cells proliferation by colon formation in HCT-116 and Lovo cells transfected with Vector, FOXQ1-WT, FOXQ1-K190R or FOXQ1 with A485 treatment (1 μM, 24 h). (C). EMT markers were detected in HCT-116 and Lovo cells transfected with Vector, FOXQ1-WT, FOXQ1-K190R or FOXQ1 with A485 treatment (1 μM, 24 h) by western blotting. D–F 106 HCT-116 cells stably expressing Vector, FOXQ1-WT or FOXQ1-K190R were subcutaneously injected into the abdominal flanks of each mouse. n = 5 for each group. Tumor volume (E) and weight (F) were measured after 4 weeks. G–H Number of lung metastases was examined detected by H&E staining after 8 weeks. The black arrows denote the metastatic nodules, scale bar 50 μm, n = 5 for each group. Data are mean ± SDs. *p < 0.05, **p < 0.01, ***p < 0.001.
Clinicopathological relevance of FOXQ1 in CRC
To examine the clinicopathological significance of FOXQ1 in CRC, tumor tissues were obtained from 26 CRC patients for IHC staining. The samples were then categorized based on the IHC score of Ac-K190 FOXQ1. The results revealed a negative association between the levels of Ac-K190 FOXQ1 and E-cadherin, while a positive correlation was observed with vimentin, MMP2, and Ki67(Fig. 7A–E), indicating that Ac-K190 acetylated FOXQ1 is positively correlated with EMT, metastasis, and proliferation of CRC.

A Representative immunohistochemistry images of E-Cadherin, Vimentin, MMP2 and Ki67 protein expression according to the level of Ac-K190 FOXQ1 in CRC patients (n = 26) collected from the Affiliated Hospital of Xuzhou Medical University, scale bars, 100 μm. B–E The correlation between the expression of Ac-K190 FOXQ1 and E-Cadherin, Vimentin, MMP2 and Ki67 were detected by immunohistochemistry in CRC patients (n = 26) collected from the Affiliated Hospital of Xuzhou Medical University. F In the TCGA database, the expression of FOXQ1 in non-cancerous tissue and carcinoma tissues in CRC patients by pair comparison. G The correlation of FOXQ1 expression and pathological stage using the TCGA data for CRC. H The correlation of FOXQ1 expression and pathological carcinoembryonic antigen (CEA) level for CRC using the TCGA data. I–J Kaplan–Meier survival curves depicting overall survival and disease-free survival in patients with CRC in TCGA database stratified by FOXQ1 protein expression levels in CRC tissues. K Schematic for the mechanisms of this project. Data are mean ± SDs. *p < 0.05, **p < 0.01, ***p < 0.001.
Further analysis of a public dataset on FOXQ1 expression using TCGA for CRC supported the finding that the expression of FOXQ1 was higher in cancer tissues compared to their paired paracancerous tissues (Fig. 7F). Meanwhile, FOXQ1 expression level is positively correlated with the pathological stage of tumors (Fig. 7G) and carcinoembryonic antigen level (Fig. 7H). CRC patients with high FOXQ1 expression had worse OS and disease specific survival (DSS) compared with those with low FOXQ1 expression (Fig. 7I, J). Collectively, these results supported our observations that FOXQ1 act as a transcription activator, promoting the development and progression of CRC.
Discussion
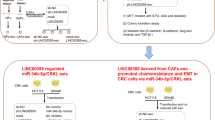
FOXQ1 has been shown to promote the proliferation, invasion, metastasis, and tumorigenesis of colorectal cancer25,26,27. Additionally, the Wnt/beta-catenin and TGF-beta signaling pathways can enhance FOXQ1 expression at the transcriptional level28,29. p53 is capable of inhibiting FOXQ1 transcription30. However, at the protein level, FOXQ1 does not interact with either β-catenin or p53 (Supplementary Fig. 5A). Acetylation exerts a profound influence on multiple functions of FOX family proteins. For instance, the acetylation of FOXP1 has been shown to inhibit the self-renewal capability of mesenchymal stem cells31. Additionally, acetylation of FOXO3a contributes to drug resistance in breast cancer32. Disruption of spermine synthase in CRC cells results in spermidine accumulation, which inhibits FOXO3a acetylation and promotes its nuclear translocation, thereby inducing Bim expression33. In our study, acetylation of Lys190 enhances FOXQ1’s ability to activate transcription in two mechanisms. Firstly, FOXQ1 binds to DNA through its FHD domain, which contains Lys190. The mutation at this site impairs its ability to locate super enhancers, indicating that Lys190 acetylation enhances the binding affinity of FOXQ1 to DNA regulatory elements. Previous studies have shown that FOXQ1 binds to the promoter of NRXN3 to activate its expression, thus promoting the glioma proliferation34. It can also target the promoter of ZEB2 to active its transcription, leading to increased invasion and metastasis in HCC35,36,37,38. With great possibility, Lys190 acetylation of FOXQ1 may play a role in these above-mentioned processes. Secondly, p300 acetylates FOXQ1 at Lys190, enabling recognition and binding by BRD4, which then recruits RNA-Pol II to form the “FOXQ1-p300-BRD4-RNA Pol II” complex for transcriptional regulation of target genes. Thus, the acetylation of FOXQ1 at Lys190 is indispensable for its transcriptional regulation ability (Fig. 7K).
The role of p300 in gene transcription involves direct acetylation of histones or transcription factors39, as well as indirect promotion of BRD4 binding to super enhancers40. Despite BRD4’s significance as a transcription factor, it lacks a DNA-binding domain41. Therefore, the mechanism by which BRD4 interacts with proteins in the transcription complex and subsequently binds to specific gene super enhancers requires further investigation. Previous studies have reported that FOXQ1 has potential combination with p300 and BRD442. We found that the FOXQ1 can be acetylated by p300 at Lys190, and then recognized and bound by BRD4-BD1, which allows FOXQ1 to recruit BRD4 to the super enhancers of the target genes. This finding enriches our understanding of the collaboration between p300 and BRD4 in transcriptional regulation.
BRD4 contains two bromodomains, BD1 and BD2, each characterized by a conserved fold consisting of four antiparallel helices (αZ, αA, αB, and αC) and two loop regions (ZA and BC loops)43. The four α-helical structural motifs are linked by two loops that act as readers, recognizing and binding to acetylated lysine residues on histones or transcription factors44,45,46,47. Despite the high sequence homology between BD1 and BD2, they exhibit differences in recognizing various forms of acetylation48. BD2 demonstrates broader recognition of acetylated substrates, whereas BD1 shows a preference for acetylation of H448. BRD4 has been reported to bind with acetylated Twist1 through its BD2 domain, enhancing the stemness of breast cancer41. Meanwhile, BD1 can interact with acetylated RELA to promote the invasion and metastasis of triple-negative breast cancer49. At present, several inhibitors targeting BD1 and BD2 have been developed to inhibit tumor initiation and progression, such as GSK77850 (BD1 inhibitor), ABBV-74451 (BD2 inhibitor), and JQ1(BD1 and BD2 inhibitor)52. In our study, BRD4-BD1 could recognize and bind to acetylated FOXQ1, synergistically promoting the proliferation and metastasis of CRC, which indicated that targeted disruption of their interaction may offer novel therapeutic strategies for CRC.
Super-enhancers belong to a cluster of enhancers that play crucial role in regulating transcription53. In tumor cells, super-enhancers can efficiently drive the transcription of oncogenes, such as KLF5, USP12, PARD6B, and MYC, thus promoting the malignant progression of tumors54. Similar to enhancers, super-enhancers can not only function as a gene regulatory element, but also recruit high-density key transcription factors, transcription cofactors, histones with specific epigenetic modifications (H3K4me1 and H3K27ac), and RNA Pol II complex12,55,56,57. However, the binding density of these factors in super-enhancers is approximately 10 times higher than that in traditional enhancers58. Accordingly, super-enhancers can markedly increase the transcription levels of target genes59,60. In our study, we identified a “FOXQ1-p300-BRD4-RNA-Pol II” complex, which was derived by Lys190 acetylation of FOXQ1 and targeted the super-enhancer of FCGR2A and MIR21. Knocking down p300 or introducing a Lys190 mutation resulted in decreased binding of FOXQ1 at super-enhancers, while knocking down the deacetylase HDAC1 led to increased enrichment of FOXQ1 at super-enhancers. Since Lys190 is located in the FOXQ1 DNA binding domain (119-214), acetylation at this site not only facilitates the formation of “FOXQ1-p300-BRD4-RNA-Pol II” complex but also potentially alters the conformation of FOXQ1, thereby enhancing its affinity for DNA.
In conclusion, our results confirmed that the acetylation of FOXQ1 promotes the formation of “FOXQ1-p300-BRD4-RNA Pol II” complex, which subsequently regulates the super enhancers of series of oncogenes. By targeting this complex, we can develop small molecule inhibitors that specifically interfere with the protein-protein interactions within it, thereby preventing its pro-oncogenic effects. Additionally, considering the critical role of FOXQ1 acetylation in the assembly of this complex, the acetylation status of FOXQ1 is anticipated to serve as a valuable diagnostic biomarker for colorectal cancer. Further assessment is required to fully elucidate the regulatory role of FOXQ1 on DNA regulatory element in various tumor types. Nevertheless, our findings highlight FOXQ1 as a key regulator of super enhancers, contributing to a deeper comprehension of FOXQ1-related tumors and bolstering the notion that FOXQ1 could be a potential target for CRC treatment.
Materials and methods
Cell lines
HCT-116, Lovo, DLD1, SW480 and 293 T cells were obtained from the Chinese Academy of Sciences (Shanghai, China) and maintained in DMEM (Sigma) with 10% FBS (Excell) and 1% penicillin-streptomycin solution (TIANGEN). All cell lines were stored at 37 °C under a humidified atmosphere with 5% CO2 and then trypsinized and passaged every two days.
Antibodies and plasmids
FOXQ1 (Proteintech, 23718-1-AP, IP: 1:200, WB: 1:1000, IHC: 1:200), p300 (Abcam, ab275378, IP: 1:200, WB: 1:1000), BRD4 (Abcam, ab314432, WB: 1:1000), HDAC1 (Abcam, ab280198, IP: 1:300, WB:1:1000), Flag (Proteintech, 20543-1-AP, WB: 1:10000), Pan Acetylation (Proteintech, 66289-1-Ig, WB:1:1000), RNA-Pol ll (Abcam, ab5408, IP: 1:100, WB:1:1000, ChIP: 1:100), GCN5 (Proteintech, 66575-1-Ig, WB:1:2000), Tip60 (Proteintech, 10827-1-AP, WB:1:2000), PCAF (Proteintech, 28770-1-AP, WB:1:2000), GST (Proteintech, 66001-2-Ig, WB:1:1000), N-Cadherin (Proteintech, 22018-1-AP, WB: 1:2000, IHC: 1:300), Vimentin (Proteintech, 60330-1-Ig, WB: 1:2000, IHC: 1:400), E-Cadherin (Proteintech, 20874-1-AP, WB: 1:2000, IHC: 1:50), Ac-K190 (HUABIO, IHC: 1:200, WB: 1:1000), Ki67 (Proteintech, 27309-1-AP, IHC: 1:200) and Tubulin (Proteintech, 80762-1-RR, WB: 1:20000). Flag-FOXQ1 and Flag-FOXQ1 mutants were all plasmids were synthesized at JinWeizhi company and the sequence information of FOXQ1 was supplied in Supplementary Table 6.
Tissue specimens
Colorectal cancer and adjacent normal tissues were collected from the Affiliated Hospital of Xuzhou Medical University. All samples were collected with approval from the Ethics Committee of the Affiliated Hospital of Xuzhou Medical University, and all patients provided informed consent (XYFY2016-JS026-01). The research was performed in compliance with the Helsinki declaration and internationally accepted ethical rights for health studies done on humans. Proper ethical rules concerning participation of human subjects in research were ensured to be adhered to.
RNA-seq experiment
For RNA-Seq, colorectal cancer tissues and adjacent non-tumor tissues were ground, and their RNA was extracted and purified. A total of 1 μg RNA was used to prepare libraries with NEBNext Ultra RNA Library Prep Kit (New England Biolabs, E7530) and sequenced on the NextSeq Illumina platform. Each sample had six biological replicates. RNA-seq data were aligned to human genome using STAR with ENCODE options. Transcript quantification was done with RSEM, and differential expression analysis with DESeq2.
Western blotting
Cell lysates were isolated utilizing RIPA lysis buffer supplemented with 1% PMSF and 1% protease inhibitor cocktail, incubated for 30 minutes at 4 °C. Subsequently, protein concentrations were accurately measured with a BCA assay kit (Thermo). These samples were then separated via 10% SDS-PAGE electrophoresis and efficiently transferred onto PVDF membrane. Blocking was meticulously performed using NcmBlot buffer, followed by an overnight incubation period with specific primary antibodies. After incubation with secondary antibodies for 1 hour at room temperature, the membranes were clearly visualized using BeyoECL Plus (Beyotime) on the Tanon 4600 system.
Pull down and silver staining
Lysates from HCT-116 cells were transfected with Flag-FOXQ1 and prepared using 0.3% Nonidet P-40 lysis buffer (0.2 mM EDTA; 50 mM Tris-HCl, pH 7.4; 150 mM NaCl; 0.3% Nonidet P-40) containing the protease inhibitor cocktail (Roche). Anti-Flag tag (L5) affinity beads (BioLegend) were used to incubate with the cell extracts for 12 h at 4°C. After binding, the beads were washed with the above-mentioned lysis buffer. Flag peptide (Sigma) was then applied to the beads to elute the Flag protein complex as per the manufacturer’s instructions. The eluents were collected and visualized via 10% SDS-PAGE followed by silver staining with Fast Silver Stain Kit (Beyotime). Distinct protein bands were retrieved and analyzed by LC-MS/MS.
Co-Immunopurification (Co-IP)
For Co-IP, 800 μg of protein was incubated with 2 μg of antibodies for 12 h at 4 °C with constant rotation and cellular extracts at 5% concentration were used for analysis. Subsequently, 80 μL of 50% protein A/G agarose beads were added, and incubation was continued for an additional 2 h. The beads were then washed five times by lysis buffer. In each wash interval, the beads were collected by centrifugation at 2000 rpm for 5 min at 4 °C. The precipitated proteins were eluted from the beads by resuspending them in 2 × SDS-PAGE loading buffer and boiling for 10 min. The resultant proteins from CoIP or the cell lysates were separated by Western blotting.
GST pull-down assay
GST fusion constructs were expressed in BL21 Escherichia coli, and crude bacterial lysates were prepared by sonication in 1× phosphate-buffered saline (PBS) with protease inhibitor mixture. Equal amounts of GST fusion proteins were immobilized on 50 μL of 50% glutathione Sepharose 4B slurry beads. After incubation for 1 h at 4 °C with rotation, the beads were washed three times with PBS and resuspended in cell lysates for 8 h at 4 °C with constant rotation. Subsequently, the beads underwent four washes with the binding buffer. The bound proteins were eluted by boiling in 30 μL of 2×sample loading buffer and separated by Western blotting.
Antibody production and purification
The acetylated FOXQ1-K190 site-specific antibody was obtained from HUABIO company. The production and purification of the antibody were performed by this company. Specifically, the antibody was generated by immunizing rabbits with a synthesized peptide CRDPSRPWGK(ac)DNYWM conjugated to keyhole limpet hemocyanin (KLH). Serum was collected after 2 immunizations. The antibody was tested using ELISA and FOXQ1-K190 site mutation. The antibody was tested using dot blot and FOXQ1-K190 site mutation (Supplementary Fig. 1B, C).
In vitro acetylation/deacetylation assay
For acetylation reaction, 0.5 μg of p300 protein (Active Motif, 81158) and Flag-FOXQ1, produced by transcription and translation in vitro (Promega), along with anti-Flag tag (L5) affinity beads (or peptides) along with 10 µM of AC-CoA and A485, were added to the reaction buffer (50 mM Tris-CI, pH 8.0, 10% glycerol, 0.1 mM EDTA, and 50 ng/μl BSA). For deacetylation reaction, Flag-FOXQ1 was first acetylated according to the acetylation reaction as described above. Following this, 0.5 μg of HDAC1 protein (Active Motif, 31504), 2 µM of TSA, and acetylated Flag-FOXQ1 along with Anti-Flag tag (L5) affinity beads were added to the reaction buffer according to different groups. The reaction mixtures were incubated at 30 °C for 1 h. The resultant materials were separated by Western blotting.
DNA pull down
Briefly, the biotin-labeled FOXQ1 binding sequence (TGTTTATGTTTAGTTT) was incubated with Streptavidin Magnetic Beads (MCE, HY-K0208). The non-biotinylated FOXQ1 binding sequence was used as the negative control. Flag-FOXQ1 (WT) and Flag-FOXQ1 (K190R) were immunoaffinity-purified from 293 T and eluted with FLAG peptide. Proteins from the input group were used as the positive control. The biotinylated DNA was incubated overnight with protein lysates at 4 °C. After washing and separation, bead-DNA-bound proteins were subjected to Western blotting.
Dot blot
The denatured protein sample with FOXQ1 peptide (SRPWGKDNYWM) was gently applied to the nitrocellulose membrane and allowed to air-dry at room temperature. Following this, the membrane was incubated in a blocking solution containing 5% bovine serum albumin (BSA) for 1 h at room temperature. Next, the membrane was incubated overnight with primary antibodies at 4 °C. Subsequently, the membrane was incubated with secondary antibodies for 1 h at room temperature and visualized using NcmECL Ultra on the Tanon 4600 Image Analysis System.
ChIP
ChIP assays were performed using the ChIP assay kit (Beyotime). DNA was purified using the PCR Clean Up Kit (Beyotime). ChIP-qPCR assay was performed using the UltraSYBR One Step RT-qPCR Kit (CWBio).
Peptide pull-down ChIP
The biotin-labeled peptides (10 mM, AceK190: RDPSRPWGK(ac)DNYWM, NonAceK190: RDPSRPWGKDNYWM) were incubated with 40 μL of Streptavidin Magnetic Beads at 4 °C for 6 hours. This step is analogous to the conjugation of antibodies with protein A/G beads. Following this, the promoter region of the FOXQ1 target gene was enriched using the ChIP Assay Kit (Beyotime).
ChIP-seq analysis
ChIP-seq reads were mapped to the hg19 human genome using bowtie2. Samtools was used to convert sam files to bam format. The significance of enriched ChIP regions was evaluated by using MACS2. Intervene was used for determining the overlapped peaks. The R package ChIPseeker and VennDiagram were used to analyze peak distribution in chromatin and gene annotation. the ROSE (Rank Ordering of Super-Enhancers) algorithm was used to identify super-enhancers. ROSE was run by allowing enhancers within 12,500 bp to be stitched together.
Luciferase reporter assay
E-1, E-2, E-A, and E-B sequences are cloned into pGL3-promotor, and the sequence information was supplied in Supplementary Table 6. Cells in 12-well plates were transfected with luciferase reporter (pGL3-promotor- E-1/ E-2/ E-A / E-B, Control: pGL3-promotor), Renilla, and indicated expression constructs. Two days after transfection, the firefly luciferase and Renilla luciferase were assayed according to the manufacturer’s instructions (Promega), and the firefly luciferase activity was normalized to Renilla luciferase. Each experiment was performed in triplicate and repeated at least three times.
Animal work
HCT-116 cells were infected with lentiviruses carrying vector (pLenti-CMV-GFP), FOXQ1 (pLenti-CMV-GFP-FOXQ1), and K190R (pLenti-CMV-GFP-K190R). Eight-week-old female NOD/SCID mice were randomly assigned to three groups, with five mice in each group. To minimize potential confounding factors, all grouped animals were treated concurrently. Similarly, all animal studies were approved by the Institutional Animal Care and Use Committee of Xuzhou Medical University (No. 202309T032). We have complied with all relevant ethical regulations for animal use. For the tumor formation assay, HCT-116 cells expressing the specified plasmids were prepared in the medium and mixed with Matrigel at a 1:9 ratio. In the subcutaneous tumor model, 106 cells were injected subcutaneously into NOD/SCID mice (n = 5/group). After 4 weeks, the animals were humanely euthanized in accordance with ethical guidelines and standard protocols776, and the tumor volume (V = a × (b × b)/2, where a is the largest diameter, and b is the smallest diameter) and weight were measured. In the lung metastasis model, HCT-116 cells (5 × 106) were injected into the mice via the caudal vein (n = 5/group). All the above-mentioned animal experiments were conducted in SPF-level animal house. After 8 weeks, the number of tumor metastases was determined using H&E staining.
Immunohistochemistry assay
The tissues were incubated with xylene for deparaffinization and decreasing concentrations of ethanol for rehydration. Next, 3% hydrogen peroxide was added to block endogenous peroxidase activity. The microwave antigen retrieval technique was used for antigen retrieval. After blocking, the samples were incubated with the primary antibodies overnight at 4 °C. PBS was used instead of the primary antibodies in the negative cohort. The secondary antibody was subsequently added using a horseradish peroxidase–polymer anti-mouse/rabbit IHC kit (Maixin Biotech) followed by incubation for 1 h at room temperature. Next, the samples were developed using diaminobenzidine reagent, counterstained with hematoxylin, and mounted with Permount.
Quantitative RT-PCR
Total RNA was isolated using the RNA isolater Total RNA Extraction Reagent (Vazyme), followed by reverse transcription with HiScrip III All-in-One RT SuperMix (Vazyme). Quantitative real-time PCR was conducted using UltraSYBR Mixture (CWBIO) on either the QuantStudio Flex 6 or LightCycler 96 system (Roche), with GAPDH serving as the internal control. Each sample was analyzed in triplicate to ensure reproducibility.
Transwell assay
The Transwell assay was employed to assess cellular invasion and migration. Cells were seeded onto an 8 μm polyethylene terephthalate filter membrane, which was coated with matrix gel for invasion assays or uncoated for migration assays. The lower chamber contained approximately 500 μL of medium. Following incubation, cells were fixed with 4% paraformaldehyde for 30 minutes and subsequently stained with 0.1% crystal violet for 20 minutes. Images of the invaded cells were captured using an inverted microscope at 100x magnification, and cell numbers were manually quantified.
Clone formation
HCT-116 or LoVo cells were cultured in 6-well plates for 14 days, fixed with 4% paraformaldehyde, and stained with 0.1% crystal violet for colony observation. Colonies were counted using a light microscope. Each experiment was performed in triplicate and repeated at least three times.
Statistics and reproducibility
Data from all biological triplicate experiments are presented with error bars as mean ± SD. Two-tailed unpaired Student’s t test was used to compare the two cohorts of data. One-way ANOVA or two-way ANOVA was performed to compare multiple cohorts of data. P value of less than 0.05 was considered statistically significant. All statistical analyses were performed using GraphPad Prism 10.
Data availability
The GEO Number of FOXQ1, p300, BRD4 and H3K27ac ChIP-seq date are GSM7255349, GSM1240110, GSM2058664 and GSM2265671. The raw data of RNA sequencing was deposited in figshare (https://figshare.com/) with doi: 10.6084/m9.figshare.29311406. The raw data of mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium (https://proteomecentral.proteomexchange.org) via the iProX partner repository with the dataset identifier PXD064807. All source data are included in the “Supplementary data 1–6” file linked to this manuscript. The RNA sequencing data and proteomics results data were deposited in Supplementary Data 5 and Supplementary Data 1. Supplementary Fig. 6 lists all Weatern blot experiments in the manuscript in uncropped and unedited forms. The data which are used to substantiate the findings of this study are retrievable from the author at a reasonable request.
References
Benayoun, B. A., Caburet, S. & Veitia, R. A. Forkhead transcription factors: key players in health and disease. Trends Genet. 27, 224–232 (2011).
Kaestner, K. H., Knochel, W. & Martinez, D. E. Unified nomenclature for the winged helix/forkhead transcription factors. Genes Dev. 14, 142–146 (2000).
Wang, J. et al. Members of FOX family could be drug targets of cancers. Pharm. Ther. 181, 183–196 (2018).
Carlsson, P. & Mahlapuu, M. Forkhead transcription factors: key players in development and metabolism. Dev. Biol. 250, 1–23 (2002).
Katoh, M. & Katoh, M. Human FOX gene family (Review). Int J. Oncol. 25, 1495–1500 (2004).
Hannenhalli, S. & Kaestner, K. H. The evolution of Fox genes and their role in development and disease. Nat. Rev. Genet. 10, 233–240 (2009).
Hoggatt, A. M., Kriegel, A. M., Smith, A. F. & Herring, B. P. Hepatocyte nuclear factor-3 homologue 1 (HFH-1) represses transcription of smooth muscle-specific genes. J. Biol. Chem. 275, 31162–31170 (2000).
Jonsson, H. & Peng, S. L. Forkhead transcription factors in immunology. Cell Mol. Life Sci. 62, 397–409 (2005).
Zhang, H. et al. Acetylation of AGO2 promotes cancer progression by increasing oncogenic miR-19b biogenesis. Oncogene 38, 1410–1431 (2019).
Inuzuka, H. et al. Acetylation-dependent regulation of Skp2 function. Cell 150, 179–193 (2012).
Matsuzaki, H. et al. Acetylation of Foxo1 alters its DNA-binding ability and sensitivity to phosphorylation. Proc. Natl Acad. Sci. USA 102, 11278–11283 (2005).
Jia, Q., Chen, S., Tan, Y., Li, Y. & Tang, F. Oncogenic super-enhancer formation in tumorigenesis and its molecular mechanisms. Exp. Mol. Med 52, 713–723 (2020).
Jia, Y., Chng, W. J. & Zhou, J. Super-enhancers: critical roles and therapeutic targets in hematologic malignancies. J. Hematol. Oncol. 12, 77 (2019).
Shi, J., Cao, J. & Zhou, B. P. Twist-BRD4 complex: potential drug target for basal-like breast cancer. Curr. Pharm. Des. 21, 1256–1261 (2015).
Mitchell, A. V. et al. FOXQ1 recruits the MLL complex to activate transcription of EMT and promote breast cancer metastasis. Nat. Commun. 13, 6548 (2022).
Dey, A., Chitsaz, F., Abbasi, A., Misteli, T. & Ozato, K. The double bromodomain protein Brd4 binds to acetylated chromatin during interphase and mitosis. Proc. Natl Acad. Sci. USA 100, 8758–8763 (2003).
Jones, M. H., Numata, M. & Shimane, M. Identification and characterization of BRDT: A testis-specific gene related to the bromodomain genes RING3 and Drosophila fsh. Genomics 45, 529–534 (1997).
Shang, E., Nickerson, H. D., Wen, D., Wang, X. & Wolgemuth, D. J. The first bromodomain of Brdt, a testis-specific member of the BET sub-family of double-bromodomain-containing proteins, is essential for male germ cell differentiation. Development 134, 3507–3515 (2007).
Dunaway, L. S. & Pollock, J. S. HDAC1: an environmental sensor regulating endothelial function. Cardiovasc. Res. 118, 1885–1903 (2022).
He, J. et al. Genetic polymorphisms of FCGR2A, ORAI1 and CD40 are associated with risk of lung cancer. Eur. J. Cancer Prev. 31, 7–13 (2022).
Wang, W. et al. FCGR2A and FCGR3A Genotypes Correlate with Farletuzumab Response in Patients with First-Relapsed Ovarian Cancer Exhibiting Low CA125. Cytogenet. Genome Res. 152, 169–179 (2017).
Ying, H. Q. et al. FCGR2A, FCGR3A polymorphisms and therapeutic efficacy of anti-EGFR monoclonal antibody in metastatic colorectal cancer. Oncotarget 6, 28071–28083 (2015).
Lampis, A. et al. MIR21-induced loss of junctional adhesion molecule A promotes activation of oncogenic pathways, progression and metastasis in colorectal cancer. Cell Death Differ. 28, 2970–2982 (2021).
Zhou, Q., Li, T. & Price, D. H. RNA polymerase II elongation control. Annu. Rev. Biochem 81, 119–143 (2012).
Abba, M. et al. Unraveling the role of FOXQ1 in colorectal cancer metastasis. Mol. Cancer Res. 11, 1017–1028 (2013).
Weng, W. et al. FOXM1 and FOXQ1 Are Promising Prognostic Biomarkers and Novel Targets of Tumor-Suppressive miR-342 in Human Colorectal Cancer. Clin. Cancer Res. 22, 4947–4957 (2016).
Kaneda, H. et al. FOXQ1 is overexpressed in colorectal cancer and enhances tumorigenicity and tumor growth. Cancer Res. 70, 2053–2063 (2010).
Peng, X. et al. FOXQ1 mediates the crosstalk between TGF-beta and Wnt signaling pathways in the progression of colorectal cancer. Cancer Biol. Ther. 16, 1099–1109 (2015).
Christensen, J., Bentz, S., Sengstag, T., Shastri, V. P. & Anderle, P. FOXQ1, a novel target of the Wnt pathway and a new marker for activation of Wnt signaling in solid tumors. PLoS ONE 8, e60051 (2013).
Pizzolato, G. et al. The tumor suppressor p53 is a negative regulator of the carcinoma-associated transcription factor FOXQ1. J. Biol. Chem. 300, 107126 (2024).
Ling, S. et al. Deacetylation of FOXP1 by HDAC7 potentiates self-renewal of mesenchymal stem cells. Stem Cell Res. Ther. 14, 188 (2023).
Liu, J. et al. Targeting the BRD4/FOXO3a/CDK6 axis sensitizes AKT inhibition in luminal breast cancer. Nat. Commun. 9, 5200 (2018).
Guo, Y. et al. Spermine synthase and MYC cooperate to maintain colorectal cancer cell survival by repressing Bim expression. Nat. Commun. 11, 3243 (2020).
Sun, H. T., Cheng, S. X., Tu, Y., Li, X. H. & Zhang, S. FoxQ1 promotes glioma cells proliferation and migration by regulating NRXN3 expression. PLoS ONE 8, e55693 (2013).
Xia, L. et al. Forkhead box Q1 promotes hepatocellular carcinoma metastasis by transactivating ZEB2 and VersicanV1 expression. Hepatology 59, 958–973 (2014).
Sanchez-Tillo, E. et al. Expanding roles of ZEB factors in tumorigenesis and tumor progression. Am. J. Cancer Res. 1, 897–912 (2011).
Xiong, H. et al. Roles of STAT3 and ZEB1 proteins in E-cadherin down-regulation and human colorectal cancer epithelial-mesenchymal transition. J. Biol. Chem. 287, 5819–5832 (2012).
Cong, N. et al. Downregulated microRNA-200a promotes EMT and tumor growth through the wnt/beta-catenin pathway by targeting the E-cadherin repressors ZEB1/ZEB2 in gastric adenocarcinoma. Oncol. Rep. 29, 1579–1587 (2013).
Kim, H. J., Moon, S. J., Hong, S., Won, H. H. & Kim, J. H. DBC1 is a key positive regulator of enhancer epigenomic writers KMT2D and p300. Nucleic Acids Res. 50, 7873–7888 (2022).
Li, X. et al. Autophagy enhanced by curcumin ameliorates inflammation in atherogenesis via the TFEB-P300-BRD4 axis. Acta Pharm. Sin. B 12, 2280–2299 (2022).
Shi, J. et al. Disrupting the interaction of BRD4 with diacetylated Twist suppresses tumorigenesis in basal-like breast cancer. Cancer Cell 25, 210–225 (2014).
Pizzolato, G., Moparthi, L., Soderholm, S., Cantu, C. & Koch, S. The oncogenic transcription factor FOXQ1 is a differential regulator of Wnt target genes. J. Cell Sci. 135 https://doi.org/10.1242/jcs.260082 (2022).
Jang, M. K. et al. The bromodomain protein Brd4 is a positive regulatory component of P-TEFb and stimulates RNA polymerase II-dependent transcription. Mol. Cell 19, 523–534 (2005).
Vollmuth, F., Blankenfeldt, W. & Geyer, M. Structures of the dual bromodomains of the P-TEFb-activating protein Brd4 at atomic resolution. J. Biol. Chem. 284, 36547–36556 (2009).
Crowe, B. L. et al. Structure of the Brd4 ET domain bound to a C-terminal motif from gamma-retroviral integrases reveals a conserved mechanism of interaction. Proc. Natl Acad. Sci. USA 113, 2086–2091 (2016).
Lin, Y. J. et al. Solution structure of the extraterminal domain of the bromodomain-containing protein BRD4. Protein Sci. 17, 2174–2179 (2008).
Wu, S. Y. & Chiang, C. M. The double bromodomain-containing chromatin adaptor Brd4 and transcriptional regulation. J. Biol. Chem. 282, 13141–13145 (2007).
Filippakopoulos, P. et al. Histone recognition and large-scale structural analysis of the human bromodomain family. Cell 149, 214–231 (2012).
Yang, G. J. et al. An optimized BRD4 inhibitor effectively eliminates NF-kappaB-driven triple-negative breast cancer cells. Bioorg. Chem. 114, 105158 (2021).
Liu, C. S. et al. Selective inhibitors of bromodomain BD1 and BD2 of BET proteins modulate radiation-induced profibrotic fibroblast responses. Int J. Cancer 151, 275–286 (2022).
Faivre, E. J. et al. Selective inhibition of the BD2 bromodomain of BET proteins in prostate cancer. Nature 578, 306–310 (2020).
Delmore, J. E. et al. BET bromodomain inhibition as a therapeutic strategy to target c-Myc. Cell 146, 904–917 (2011).
Bal, E. et al. Super-enhancer hypermutation alters oncogene expression in B cell lymphoma. Nature 607, 808–815 (2022).
Zhang, X. et al. Identification of focally amplified lineage-specific super-enhancers in human epithelial cancers. Nat. Genet. 48, 176–182 (2016).
Sengupta, S. & George, R. E. Super-Enhancer-Driven Transcriptional Dependencies in Cancer. Trends Cancer 3, 269–281 (2017).
Liu, M. et al. Super enhancer regulation of cytokine-induced chemokine production in alcoholic hepatitis. Nat. Commun. 12, 4560 (2021).
Thandapani, P. Super-enhancers in cancer. Pharm. Ther. 199, 129–138 (2019).
Loven, J. et al. Selective inhibition of tumor oncogenes by disruption of super-enhancers. Cell 153, 320–334 (2013).
Chen, X., Ma, Q., Shang, Z. & Niu, Y. Super-enhancer in prostate cancer: transcriptional disorders and therapeutic targets. NPJ Precis. Oncol. 4, 31 (2020).
Tang, F., Yang, Z., Tan, Y. & Li, Y. Super-enhancer function and its application in cancer targeted therapy. NPJ Precis. Oncol. 4, 2 (2020).
Acknowledgements
This work was supported by the Natural Science Foundation of China (82103141), the Postdoctoral Science Foundation Funded Project of China (2021M702755), and the Outstanding Youth Foundation of Jiangsu Province (BK20200046).
Author information
Authors and Affiliations
Contributions
W.D.Y., J.B., H.M.Y., D.J., and M.Y.Z. contributed to study design. W.D.Y., Z.Y.S., and K.S. collected, analyzed and interpreted the data. W.D.Y., Z.H.Z., Y.F.M., Q.S., M.R.L., M.H.C., S.F.C., S.M., J.Y.X., and K.S. performed the experiments. J.B., W.D.Y., Z.Y.S., and D.J. wrote the manuscript. All authors approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests
Peer review
Peer review information
Communications Biology thanks Stefan Koch and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Silvia Belluti and Kaliya Georgieva.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yang, WD., Zhang, ZH., Zhao, MY. et al. P300-dependent acetylation of the FOXQ1 complex activates super-enhancers to promote colorectal cancer proliferation and metastasis. Commun Biol 8, 1016 (2025). https://doi.org/10.1038/s42003-025-08430-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-025-08430-z
