
Abstract
Neuronal remodeling is essential for the precise formation of mature nervous systems. Drosophila serves to study neuronal remodeling; however, existing experimental systems are insufficient for examining circuit-level neuronal remodeling and its effect on behavior. We present a new model system to investigate neuronal remodeling. We show that the Moonwalker SEZ neuron, which elicits backward walking in adult flies, is part of a conserved circuit persisting from larva to adult. Utilizing the system, we examine developmental neuronal remodeling, describe a gene-regulatory mechanism controlling outgrowth, and uncover a causal relation between remodeling and behavior. The well-characterized connectivity of the circuit, low number of elements, direct control of the motor output, and availability of specific driver lines, provide an appealing system to study developmental remodeling of neuronal circuits and their impact on behavior. Hence, we establish a new model system in Drosophila to investigate neural circuit remodeling and its relation to behavior.
Similar content being viewed by others
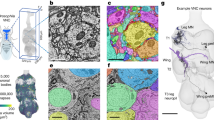

Introduction
The formation of precise neural circuits is crucial for the circuits’ function and the animals’ behavior. A key step in the establishment of refined neural circuits, in both vertebrate and invertebrates, is complex neuronal remodeling that includes both degenerative and regenerative events1,2,3,4,5. Various neurodevelopmental disorders have been associated with dysregulated postnatal neuronal remodeling6,7,8,9,10,11,12. In this context, holometabolous insects that undergo complete metamorphosis, involving large-scale remodeling, by which they transform from larvae to adult insects13, are useful model systems to study mechanisms underlying neuronal remodeling and its effects on behavior14,15,16,17. In particular, the Drosophila model system, which offers sophisticated genetic tools allowing for manipulation and visualization at a single cell resolution18,19, is considered an attractive model system to delineate the basic principles governing developmental neuronal remodeling3,4. Two major systems in the Drosophila nervous system have been extensively utilized to unravel key molecular and cellular mechanisms underlying spatiotemporal control of postnatal neuronal remodeling3,4,20,21: γ Kenyon cells (KCs) of the mushroom body (MB), and class IV dendritic arborization (C4da) neurons.
During metamorphosis, some neurons undergo trans-differentiation as their assumed larval identity is different from their terminal adult identity (that is, they are re-specified for adult functions), while other neurons maintain their original larval identity also during adulthood22,23. Of particular interest among the latter are recently identified command-like neurons that directly control a specific motor program in both larvae and adult flies24,25. The ability of specific neurons to drive analogous behavioral outputs in two substantially different neuronal architectures is striking, since it suggests that neuronal preservation is not merely limited to individual neurons persisting from larval to adult stages but rather exists on a much broader scale in the form of conserved neuronal circuits. However, our current understanding of neuronal remodeling at the circuit level, which ultimately shapes adult behavior, is still very limited.
Moreover, the existence of individual persistent neurons regulating a complete motor behavior in both larval and adult forms, presents an attractive opportunity to directly examine causality between developmental neuronal remodeling and behavior.
To experimentally explore neuronal remodeling at the circuit level, one should examine whether a specific neuron maintains functional connections with both of its pre- and post- synaptic partners following metamorphosis. Accordingly, a model system composed of a functional neuronal circuit of at least three elements, is essential. Much effort has been invested in characterizing the stereotypic remodeling of MB intrinsic neurons3,4,20,21,26. It was shown that the anterior paired lateral neuron (APL), a MB modulatory neuron, undergoes massive developmentally-regulated neuronal reorganization that is tightly coordinated with γ KC remodeling27. Moreover, it was recently found that ~37% of the different types of adult mushroom body input neurons (MBINs) and mushroom body output neurons (MBONs) originate from remodeled MB larval neurons23. Thus, these observations can support the notion that persistent MB microcircuit motifs function in both larval and adult nervous systems. However, it was discovered that neuronal pairs with maintained larval connections are sporadic in the adult MB, and that larval microcircuits mediating MB learning and memory processes do not persist into adulthood23. Furthermore, surprisingly, preventing γ KCs neuronal remodeling had only a mild behavioral effect, with a slight impairment in differential aversive olfactory learning, but not appetitive learning, or other MB-related functions28. These results thus indicate that although some of the neural components of the MB persist into adulthood, its core functional larval circuits are not maintained but rather reorganized during the transition from larva to adult. In addition, while some of C4da downstream targets in the larval nervous system have been identified29,30,31,32,33, the metamorphic fates of these neuronal elements in the adult nervous system are still unknown. Thus, C4da neurons are also not well-suited for probing inter-neuronal interactions during development. Taken together, it appears that an alternative system for studying circuit-level developmental neuronal remodeling with a clear and profound effect on the behavioral output is required.
The moonwalker descending neurons (MDNs)34, a bilateral pair of descending neurons which serve as the main neuronal hub for backward walking in the adult fly brain34,35,36, were found to exist also in larvae where they promote backward crawling25. MDNs undergo profound neuronal remodeling during metamorphosis25, nevertheless, they are capable of eliciting backward locomotor responses in both limbless larvae25, which translocate by repeated wave-like contractions of their body wall muscles, and in limbed adult flies34, which walk by coordinated contractions of their leg muscles. Furthermore, MDN-postsynaptic partner Pair1, a subesophageal zone (SEZ) descending neuron inhibiting forward locomotion25, serves a similar function in both larvae and adult flies24, and also undergoes neuronal remodeling during metamorphosis24. Notably, MDN-Pair1 synaptic connectivity, which is eliminated during early stages of metamorphosis due to MDN axonal- and Pair1 dendritic- pruning, is re-formed before pupal eclosion24. Recently, we reported the identification of a novel pair of bilateral SEZ neurons, moonwalker SEZ neurons (MooSEZs), which synapse onto MDNs and elicit backward walking in adult flies37. Thus, the backward locomotion neural circuit, composed of MooSEZ and its downstream partners MDN and Pair1, which were shown to persist through development, may serve to study remodeling of an entire circuit.
Here we focus on MooSEZ, the first neural element of a three-layered command circuit. We show that it also exists in larvae and drives larval backward crawling as well as adult backward walking. As MooSEZ is a synaptic partner of MDN at both larval and adult stages, our findings thus reveal a conserved neuronal network; in this circuit, at least one neuronal element (MDN) maintains functional connections with both of its pre- and post-synaptic partners (MooSEZ and Pair1, respectively), during the transformation from larva to adult. Furthermore, we utilize MooSEZ as a novel experimental system to mechanistically explore neuronal remodeling and probe causality between remodeling and behavior. Using this system, we find that activation of ecdysteroid-dependent genes, typically associated with neurite pruning, also regulates neurite outgrowth. Moreover, we show that developmental remodeling of a neuronal component within a conserved functional circuit is crucial for maintaining the behavioral output, and perturbation to neuronal remodeling results in a robust phenotype. Hence, the current study employs a new model system within the Drosophila nervous system, the motor circuit regulating backward locomotion, to reveal a novel gene-regulatory mechanism for neurite outgrowth during remodeling as well as preservation of a functional circuit motif whose adult functionality is causally linked to developmental neuronal remodeling.
Results
Optogenetic activation of olfactory PN driver lines drives larval backward crawling
It was previously shown that activation of MDNs drives backward locomotion in larvae25 and adult flies34. As MooSEZs are presynaptic partners of adult MDNs and trigger backward walking in adult flies37, we tested whether optogenetic activation of the broad olfactory projection neuron (PN) driver lines, GH146II-, NP225-, and NP5288-GAL4, which cover MooSEZs, could also induce a similar motor response of backward retreat among larvae. Optogenetic activation of larval neurons expressing UAS-CsChrimson38 under the control of GH146II-, NP225-, and NP5288-GAL4 driver lines resulted in a pronounced and sustained backward crawling throughout the duration of the light pulse (Fig. 1A, B, Supplementary Movie 1). Anatomical analysis of GH146II-, NP225- and NP5288-GAL4 larval expression patterns revealed that the three driver lines label a few neurons in the SEZ, in addition to antennal lobe (AL) PNs, and about a dozen of paired neurons in the ventral nerve cord (VNC) (Fig. 1C). The phenotypic similarities between adult and larva retreat responses following optogenetic activation of GH146II-, NP225- and NP5288-GAL4, and the fact that these driver lines target MooSEZs which trigger robust backward walking in adult flies37, suggest that MooSEZs may survive metamorphosis and mediate backward locomotion not only in adult flies but also in larvae. However, since the tested PN driver lines are broad, other larval neurons targeted by them could also underlie the observed backward crawling phenotype.

A Examples of single larva crawling trajectories for GH146II-, NP225-, and NP5288-GAL4 driving UAS-CsChrimson (top) and their respective parental controls (bottom) before, during, and after optogenetic stimulation. 9-s red light pulse is designated by yellow, blue denotes backward crawling, and brown forward motion. The (0, 0) coordinate represents larvae’s trajectory onset at t = 0. A 9-s optogenetic activation of larvae expressing CsChrimson using GH146II-, NP225- or NP5288-GAL4 driver lines elicits motor response of persistent backward retreat which is analogous to the motor pattern observed for adult flies of the same genotype. In contrast, light stimulation of the respective parental control larvae does not lead to backward crawling. B Top, mean translational velocity of larval crawling during a 9-s red light pulse for GH146II-, NP225-, and NP5288-GAL4 driving UAS-CsChrimson. Pronounced backward locomotion is observed during the 9-s light pulse for PN driver lines expressing CsChrismon, but not for their respective parental controls (n for GH146II>CsChrimson, GH146II, NP225>CsChrimson, NP225, NP5288>CsChrimson, NP5288, CsChrimson: 12, 11, 13, 10, 13, 11, 11 respectively, ***p < 0.001, ****p < 0.0001 for pairwise comparisons with respective parental controls, Kruskal-Wallis test followed by Dunn’s post-hoc tests). Bottom, number of peristaltic backward waves during a 9-s red light pulse of larvae expressing UAS-CsChrimson under the control of GH146II-, NP225-, and NP5288-GAL4. Number of peristaltic waves of muscle contractions traveling from anterior to posterior abdominal segments of the larva is significantly higher in GH146II-, NP225-, and NP5288-GAL4 driving CsChrimson compared with their respective parental controls (n for GH146II>CsChrimson, GH146II, NP225>CsChrimson, NP225, NP5288>CsChrimson, NP5288, CsChrimson: 12, 11, 13, 10, 13, 11, 11 respectively, ***p < 0.001, ****p < 0.0001 for pairwise comparisons with respective parental controls, Kruskal-Wallis test followed by Dunn’s post-hoc tests). C Top, Expression pattern in larvae of GH146II-, NP225-, and NP5288-GAL4. Maximum intensity projections of 95 confocal sections (1 µm) through the central brain and VNC are presented. Bottom, close-up of larval SEZ neurons labeled by GH146II-, NP225-, and NP5288-GAL4. Maximum intensity projections of 48-69 confocal sections (1 µm) through the central brain and VNC are presented. UAS-CsChrimson.mVenus was used to label the cells. Larval SEZ neurons are designated with red arrowheads. Scale bars apply to all images within the same row. D Top, mean translational velocity of larval crawling during a 9-s red light pulse for GH146II-GAL4 with subtracted GH146-QF. GH146-QF was used to drive QUAS-GAL80 to limit the expression pattern of GH146II-GAL4 driving UAS-CsChrimson. Pronounced backward locomotion is observed during the 9-s light pulse for GH146II-GAL4 intersection larvae, but not for their respective parental control larvae (n for GH146II-GAL4 > UAS-CsChrimsonՈGH146-QF > QUAS-GAL80, UAS-CsChrimson, GH146II-GAL4: 8, 11, 11 respectively, *p < 0.05, ****p < 0.0001 for pairwise comparisons with respective parental controls, Kruskal-Wallis test followed by Dunn’s post-hoc tests). Bottom, number of peristaltic backward waves during a 9-s red light pulse of larvae expressing CsChrimson under the control of GH146II-GAL4 and in the presence of GH146-QF driving QUAS-GAL80. Number of peristaltic waves of muscle contractions traveling from anterior to posterior abdominal segments of the larva is significantly higher in GH146II-GAL4 intersection larvae than in their respective parental control larvae (n for GH146II-GAL4 > UAS-CsChrimsonՈGH146-QF > QUAS-GAL80, UAS-CsChrimson, GH146II-GAL4: 8, 11, 11 respectively, **p < 0.01, ****p < 0.0001 for pairwise comparisons with respective parental controls, Kruskal-Wallis test followed by Dunn’s post-hoc tests). E Top, expression pattern of the intersection between GH146II-GAL4 driving UAS-CsChrimson.mVenus, and GH146-QF driving the GAL4 inhibitor QUAS-GAL80 in larvae. Maximum intensity projection of 95 confocal sections (1 µm) through the central brain and VNC is presented. Bottom, close-up of larval SEZ neurons labeled in GH146II-GAL4 intersection larvae. Larval SEZ neurons are designated with red arrowheads. Maximum intensity projection of 55 confocal sections (1 µm) through the central brain and VNC is presented. UAS-CsChrimson.mVenus was used to label the cells. Expression in the SEZ is maintained in GH146II-GAL4 intersection larvae. F Top left, translational velocity ± SEM (shading) of larvae treated (dark blue) or untreated (light blue) with RU-486 following a 9-s optogenetic stimulation. RU-486 driven recombinase (hPR:FLP) permanently marks larval GH146II-GAL4 neurons by flipping-out the FRT-flanked stop cassette in act5C-FRT-STOP-FRT-LexA. The 9-s light pulse is labeled by light red. Top right, mean translational velocity during the 9-s light pulse obtained from traces on the left. Optogenetic activation of RU-486 treated larvae induces pronounced larval backward locomotion which is not observed for RU-486 untreated larvae (n for -RU486, +RU486: 11, 14 respectively, ****p < 0.0001, Mann-Whitney test). Bottom left, translational velocity ± SEM (shading) of adult flies developed from larvae presented above either exposed (dark blue) or unexposed (light blue) to RU-486 during larval stages following a 1.5-s optogenetic stimulation. The 1.5-s light pulse is labeled by light red. Bottom right, mean translational velocity during the 1.5-s light pulse obtained from traces on the left. Larvae with immortalized GH146II expression maintain their backward locomotion phenotype into adulthood. By contrast, larvae unexposed to RU-486 show a consistent lack of backward locomotion through development (n for -RU486, +RU486: 11, 14, respectively, ****p < 0.0001, Mann-Whitney test). G Top left, translational velocity ± SEM (shading) of mosaic larvae generated with the SPARC method expressing CsChrimson::tdTomato in stochastically distributed subsets of neurons within the GH146II-GAL4 driver line during a 9-s red light stimulation. Larvae were grouped into backward crawlers (dark blue) and non-backward crawlers (light blue). The 9-s light pulse is labeled by light red. Top right, mean translational velocity during the 9-s light pulse obtained from traces on the left (n for NO BW, BW: 7, 13 respectively, ****p < 0.0001, Mann-Whitney test). Bottom left, translational velocity ± SEM (shading) of SPARC mosaic adult flies that were categorized as either backward crawlers (dark blue) or non-backward crawlers (light blue) during larval stages following a 2-s red light pulse. The 2-s light pulse is labeled in light red. Bottom right, mean translational velocity during the 2-s light pulse obtained from traces on the left. Adult flies classified as backward crawlers during larval stages walked backwards upon optogenetic stimulation, whereas adult flies classified as non-backward crawlers during larval stages did not (n for NO BW, BW: 7, 13 respectively, ***p < 0.001, Mann-Whitney test). For details of statistical analysis, see Supplementary Data 1.
We previously showed that activation of neurons labeled by GH146II-GAL4 but not GH146-QF39 of adult flies elicits pronounced backward walking through activation of MooSEZs37. To further investigate which neuronal subset is responsible for GH146II-, NP225-, and NP5288-GAL4-triggered backward crawling, we tested whether activation of larval neurons expressing CsChrimson under the control of GH146II-GAL4 with subtracted GH146-QF37 results in backward crawling. Indeed, we observed a clear and sustained backward retreat (Fig. 1D). In addition, subtracting GH146-QF expression pattern from GH146II-GAL4 expression pattern resulted in labeling of a significantly lower number of larval neurons. Yet, GAL4 expression in larval SEZ was still maintained (Fig. 1E). Thus, these results raise the possibility that MooSEZs exist already in the larva and can drive backward crawling. However, it is also possible, though less likely, that other neurons labeled in the subtracted GH146II-GAL4 subset underlie the observed backward crawling phenotype.
MooSEZ survives metamorphosis
The results presented hitherto indicate that the driver line, which labels MooSEZs in adult flies (GH146II-GAL4), also targets a similar neuron in larvae that controls backward crawling (Fig. 1A, B, D). Thus, it seems likely that MooSEZs persist from larval stages into adulthood. However, one cannot preclude the possibility that GH146II-GAL4 driver line may label a non-MooSEZ neuron driving larval crawling. Hence, the GAL4 expression of GH146II may be switched off in this neuron right before or during metamorphosis and switched on in the newly-formed MooSEZ only during metamorphosis or right after eclosion. To conclusively examine whether MooSEZ indeed survives metamorphosis, we used immortalization genetics23,24,25,40 to permanently label larval neurons in a temporally-controlled manner, and assay their presence and function. Mifepristone (RU-486) inducible flippase (UAS-hPR:FLP) was used along with GH146II-GAL4, act5C-FRT-STOP-FRT-LexA, and LexAop-CsChrimson to irreversibly label larval GH146II-GAL4 neurons. Upon RU-486 application during larval stages, hPR:FLP immortalized CsChrimson expression in GH146II-GAL4 larval neurons by inducing constitutive LexA expression (Supplementary Fig. 1A). Optogenetically activating larvae treated with RU-486 resulted in backward crawling. Notably, this behavioral phenotype persisted into adulthood, as respective adult flies also walked backwards upon optogenetic stimulation (Fig. 1F, Supplementary Fig. 1B). Importantly, MooSEZ labeling was consistently detected in the brains of these flies (Supplementary Fig. 1C). By contrast, larvae unexposed to RU-486 showed lack of backward locomotion at both larval and adult stages upon light activation (Fig. 1F, Supplementary Fig 1B). In line with this, MooSEZ was not targeted in these fly brains; nonetheless, it is to be noted that some CsChrimson expression was also observed for RU-486 untreated flies (Supplementary Fig. 1C), probably due to leaked hPR:FLP activity. Taken together, the immortalization experimental results unequivocally demonstrate that MooSEZ is a developmentally conserved larval neuron.
MooSEZ elicits larval backward crawling
The above results show that MooSEZ survives metamorphosis. However, they do not conclusively demonstrate that it indeed underlies larval backward crawling, since immortalized CsChrimson was not selectively detected in MooSEZ, but rather observed in a relatively broad manner across flies (Supplementary Fig. 1C). In theory, the results could be explained by the activation of two independent GH146II-GAL4 neurons controlling backward locomotion: a non-MooSEZ neuron during larval stages, and MooSEZ during adulthood. That is, it is possible that MooSEZ is a trans-differentiating neuron eliciting adult yet not larval backward locomotion.
To determine whether MooSEZ maintains its functionality through development, we took advantage of the sparse predictive activity through recombinase competition (SPARC) genetic technique41. Although the applicability of the SPARC approach was originally demonstrated only for the adult Drosophila brain, it can also be used effectively for genetic manipulations in larvae, as recent studies have successfully demonstrated42,43,44. As opposed to the immortalization approach, which labeled a rather uniform neuronal population (Supplementary Fig. 1C), this method targeted sparse and stochastic subsets of neurons within GH146II-GAL4 expression pattern across individuals, allowing us to examine the correlation between MooSEZ expression and behavior along development. Combining GH146II-GAL4 transgene with SPARC2-I-CsChrimson::tdTomato variant and nSyb-PhiC31 recombinase, produced mosaic larvae with stochastic expression of CsChrimson::tdTomato. We tested these larvae for backward locomotion and divided them into groups of backward crawling and non-backward crawling larvae. We reasoned that if MooSEZ elicits larval backward crawling and thus maintains its functionality through development, larval and adult behavioral phenotypes will be coordinated. Indeed, larval and adult behavioral responses were highly correlated, as adult flies classified as backward crawling larvae also walked backwards, whereas adult flies classified as non-backward crawling larvae showed a lack of backward locomotion upon optogenetic activation (Fig. 1G). Moreover, MooSEZ labeling was exclusively detected in the brains of adult flies classified as backward crawling larvae (Supplementary Fig. 1D). Hence, these results indicate that MooSEZ induces not only adult but also larval backward locomotion. Taken together with the above immortalization results (Fig. 1F, Supplementary Fig. 1B, C), MooSEZ is a developmentally persistent neuron with maintained larval functionality.
MooSEZ rotational component is adult-specific
We have previously demonstrated that short asymmetric activation of adult MooSEZs drives pronounced ipsiversive backward turning37. Having established MooSEZ’s ability to control larval backward crawling (Fig. 1G, Supplementary Fig. 1D), we asked whether MooSEZ’s rotational component is also a preserved larval motor feature. To determine this, we compared the behavioral responses of individual GH146II-GAL4 SPARC mosaic larvae that crawled backwards to the motor responses of their respective adult flies following optogenetic activation. We then dissected, stained, and examined the expression of CsChrimson in MooSEZs of each tested fly. We found that larvae performed straight backward locomotion that was independent of MooSEZ’s side of activation. In contrast, adult flies demonstrated ipsiversive backward rotation upon MooSEZ asymmetric activation, and walked backward in a straight fashion only if both MooSEZs were co-activated (Fig. 2A–C, Supplementary Movie 2). Moreover, we also examined whether long bilateral activation of MooSEZs, which was shown to evoke sustained rotational movements among adult flies37, triggers similar larval angular motions. Consistent with the results obtained for unilateral MooSEZ activation, long bilateral activation of MooSEZs induced notable angular motions only among adult flies (Fig. 2D, E). Together, the differential rotational responses observed for larva and adult following both activation protocols, suggest that the neuronal circuitry by which MooSEZ controls angular locomotion does not exist in larva and is being established only during metamorphosis.

A Example trajectories of mosaic larvae (top) and their matched adult flies (bottom) during optogenetic activation (larvae, 9-s; adults, 2-s) of CsChrimson in right (purple), left (yellow), or both MooSEZs (black). CsChrimson was expressed unilaterally or bilaterally using the SPARC “intermediate” effector and under the control of the GH146II-GAL4 driver line. MooSEZ unilateral activation drives straight backward locomotion in larvae but ipsilateral backward rotation in adult flies. B Single traces (light) of cumulative angular change and their respective means (dark) of larvae (top) and their matched adult flies (bottom) in which MooSEZs were optogenetically activated (larvae, 9-s; adults, 2-s) unilaterally (right, purple; left, yellow) or bilaterally (black). The light pulse is labeled in light red. CsChrimson was expressed unilaterally or bilaterally using the SPARC “intermediate” effector and under the control of the GH146II-GAL4 driver line. C Mean angular velocity during optogenetic stimulation (larvae, 9-s; adults, 2-s) of the right (purple), left (yellow), or both MooSEZs (black) for the single larvae and their matched adult flies presented in (B). Rotational backward locomotion upon MooSEZ unilateral activation is observed only following metamorphosis. D Left, angular speed ± SEM (shading) of larvae (light blue) and adult flies (dark blue) following a 9-s long bilateral optogenetic stimulation of CsChrimson in MooSEZs driven by GH146II-GAL4 with subtracted GH146-QF. GH146-QF was used to drive QUAS-GAL80 to limit the expression pattern of GH146II-GAL4 driving UAS-CsChrimson. The light pulse is labeled by light red. Continuous symmetric MooSEZ activation evokes rotational movements only among adult flies. Right, mean angular speed during the 9-s light pulse obtained from traces on the left. A significant difference is observed between the mean angular speed of larvae and adult flies upon prolonged bilateral light stimulation (n for larva, adult: 8, 18, respectively, ****p < 0.0001, Mann-Whitney test). E Single fly (dark blue) and larva (light blue) traces (light) of absolute cumulative angular change and their respective means (dark) following a 9-s long bilateral optogenetic stimulation of CsChrimson in MooSEZs driven by GH146II-GAL4 with subtracted GH146-QF. GH146-QF was used to drive QUAS-GAL80 to limit the expression pattern of GH146II-GAL4 driving UAS-CsChrimson. The light pulse is labeled by light red. Prolonged bilateral MooSEZ activation elicits sustained adult-specific backward rotation. Right, absolute total angular change at the end of the 9-s light pulse obtained from traces on the left. A significant difference is observed between the absolute total angular change of larvae and adult flies (n for larva, adult: 8, 18, respectively, ****p < 0.0001, Mann-Whitney test). For details of statistical analysis, see Supplementary Data 1.
AMB driver lines trigger backward locomotion in adult flies
Next, we sought to investigate MooSEZ remodeling during the transition from larva to adult. GH146II-, NP225-, and NP5288-GAL4 are all broad labeling driver lines37 unsuitable for precise visual inspection of MooSEZ spatiotemporal dynamics through development. Hence, we searched for an alternative driver line that selectively labels MooSEZ and also maintains its GAL4 expression throughout development.
It was recently shown that a pair of bilateral larval neurons, the ascending moonwalker-like backward neurons (AMBs), mediate backward locomotion via activation of MDNs45. Similarly to MooSEZs, AMBs reside in the SEZ brain region45. Notably, optogenetic activation of the AMBs, using the specific AMB split GAL4 driver line45, elicited robust larval backward crawling that was markedly similar to the motor responses observed for GH146II-, NP225-, NP5288, and GH146II-GAL4 with subtracted GH146-QF larvae (compare Supplementary Fig. 2A with Fig. 1A, B, D). Moreover, anatomy analysis revealed that the somata of the AMBs and the SEZ neurons labeled by GH146II-GAL4 with subtracted GH146-QF reside within the same region of the larval SEZ (compare Supplementary Fig. 2B with Fig. 1E). Thus, these results raise the possibility that AMB and MooSEZ are identical neurons and that the AMB split GAL4 driver could be used to study MooSEZ remodeling through development. Nevertheless, it is also possible that MooSEZ and AMB are non-identical cells in the same cluster with overlapping larval functionality, and hence the AMB split GAL4 driver is not appropriate for MooSEZ targeting.
To gain further insight into whether the AMB split GAL4 driver labels MooSEZs, we next asked whether this driver is capable of eliciting backward locomotion not only in larvae but also in adult flies, analogously to the driver lines GH146II-, NP225-, and NP5288-GAL4. However, upon optogenetic excitation with CsChrimson, no backward walking was observed (Fig. 3A). Correspondingly, CsChrimson expression was not detected in the adult SEZ brain region where MooSEZs reside. Rather, a single bilateral neuron located in the lateral horn (LH) brain region was sparsely labeled (Supplementary Fig. 2B). Hence, the combined adult anatomical and behavioral results indicate that the AMB split GAL4 driver line is not suitable for assessing MooSEZ expression pattern through development due to a lack of GAL4 expression in the neuron of interest.

A Left, translational velocity ± SEM (shading) of adult flies following a 2-s optogenetic activation of AMB split GAL4 driver line, its respective GAL4 parental drivers, and GH146II-GAL4 with subtracted GH146-QF as designated. Optogenetic stimulation of AMB split GAL4 adult flies does not evoke backward locomotion, in contrast to optogenetic activation of adult AMB-GAL4 parental drivers, and adult GH146II-GAL4 with subtracted GH146-QF. The split GAL4 hemidrivers R11E07-p65.AD and R60F09-GAL4.DBD were used to drive expression of CsChrimson for the AMB split GAL4 driver, R60F09- and R11E07-GAL4 drivers were used to drive expression of CsChrimson for the GAL4 parental drivers, and GH146-QF was used to drive QUAS-GAL80 to restrict the expression pattern of GH146II-GAL4 driving UAS-CsChrimson in GH146II-GAL4 intersection flies. The 2-s light pulse is labeled by light red. Right, mean translational velocity during the 2-s light pulse obtained from the traces on the left. Optogenetic stimulation of adult AMB GAL4 parental drivers triggers backward walking, which is similar to the motor response observed for light activation of GH146II-GAL4 intersection flies. In contrast, optogenetic activation of the adult AMB split GAL4 line does not elicit backward walking (n for R60F09-GAL4 > UAS-CsChrimson, R11E07-GAL4 > UAS-CsChrimson, R11E07-p65.ADՈR60F09-GAL4.DBD > UAS-CsChrimson, GH146II-GAL4 > UAS-CsChrimsonՈGH146-QF > QUAS-GAL80: 21, 11, 10, 32 respectively, *p < 0.05, ****p < 0.0001 for pairwise comparisons of GH146II-GAL4 with subtracted GH146-QF with each experimental group, Kruskal-Wallis test followed by Dunn’s post-hoc tests). B Left, expression pattern of R11E07-GAL4 driving UAS-CsChrimson.mVenus in adult flies. Maximum intensity projections of 140 confocal sections (1 µm) through the central brain and VNC are presented. Scale bar applies to both images. Right, Schematic drawing of the expression pattern of the driver line presented on the left. R11E07-GAL4 driver line targets cells in both the central brain and VNC of adult flies. C Left, expression pattern of R60F09-GAL4 driving UAS-CsChrimson.mVenus in adult flies. Maximum intensity projections of 135 confocal sections (1 µm) through the central brain and VNC are presented. Scale bar applies to both images. Right, Schematic drawing of the expression pattern of the driver line presented on the left. R60F09-GAL4 driver line labels cells primarily in the central brain of adult flies. D Expression pattern of R60F09-LexA (left), GH146II-GAL4 (middle), and both driver lines (right) in the adult fly brain. R60F09-LexA was used to drive LexAop-CsChrimson.mVenus, and GH146II-GAL4 was used to drive UAS-CsChrimson.mCherry. A clear overlap between the two driver lines is observed in a paired lateral SEZ neuron. White arrowheads mark the cell bodies of the paired lateral SEZ neuron targeted by both driver lines. Maximum intensity projections of 20 confocal sections (1 µm) through the central brain are presented. Scale bar applies to all images. E Left, translational velocity traces (left) ± SEM (shading) of RU-486-treated (dark red) or untreated (light red) larvae following a 9-s optogenetic stimulation and their respective means during the presented light pulse (right). RU-486-driven recombinase (hPR:FLP) permanently marks larval R60F09-GAL4 neurons by excising the FRT-flanked stop cassette in act5C-FRT-STOP-FRT-LexA. The 9-s light pulse is labeled in light red. Optogenetic activation of larvae exposed to RU-486 induces pronounced larval backward locomotion, which is not observed for RU-486 untreated larvae (n for -RU486, +RU486: 12, 11, respectively, ****p < 0.0001, Mann-Whitney test). Right, translational velocity traces (left) ± SEM (shading) of adult flies developed from larvae presented on the left either exposed (dark red) or unexposed (light red) to RU-486 during larval stages following a 1.5-s optogenetic stimulation and their respective means during the presented light pulse (right). The 1.5-s light pulse is labeled in light red. Larvae with immortalized R60F09 expression maintain their backward locomotion phenotype into adulthood. By contrast, larvae unexposed to RU-486 show a consistent lack of backward locomotion through development (n for -RU486, +RU486: 12, 11, respectively, ****p < 0.0001, Mann-Whitney test). F Left, schematic drawing of the expression pattern of the intersection between the activation domain (AD) GAL4 hemidriver VT032768, and the DNA binding domain (DBD) GAL4 hemidriver R60F09. Middle, expression pattern of VT032768-p65.ADՈR60F09-GAL4.DBD driving UAS-CsChrimson.mVenus. Arrowheads mark MooSEZ neurites. Maximum intensity projections of 110 confocal sections (1 µm) through the central brain and VNC are presented. Scale bar applies to both images. Right, close-up of VT032768-p65.ADՈR60F09-GAL4.DBD anterior (top) and posterior (bottom) central brain expression pattern. Arrowheads mark MooSEZ soma and neurites. Maximum intensity projections of 65 confocal sections (1 µm) are presented. Scale bar applies to both images. Combining GAL4 hemidrivers VT032768 and R60F09 leads to a specific GAL4 expression in adult MooSEZs and a bilateral pair of SEZ descending neurons. G Left, schematic drawing of the expression pattern of the intersection between the activation domain (AD) GAL4 hemidriver VT020742, and the DNA binding domain (DBD) GAL4 hemidriver R60F09. Middle, expression pattern of VT020742-p65.ADՈR60F09-GAL4.DBD driving UAS-CsChrimson.mVenus. Arrowheads mark MooSEZ neurites. Maximum intensity projections of 100 confocal sections (1 µm) through the central brain and VNC are presented. Scale bar applies to both images. Right, close-up of VT020742-p65.ADՈR60F09-GAL4.DBD anterior (top) and posterior (bottom) central brain expression pattern. Arrowheads mark MooSEZ somata and neurites. Maximum intensity projections of 50 confocal sections (1 µm) are presented. Scale bar applies to both images. Combining GAL4 hemidrivers VT020742 and R60F09 leads to GAL4 expression exclusively in adult MooSEZs. For details of statistical analysis, see Supplementary Data 1.
Since split GAL4 drivers do not always reproduce their parental drivers’ overlapping expression patterns46,47, we also examined the parental GAL4 driver lines, R60F09 and R11E07. Surprisingly, optogenetic activation of these drivers did induce robust adult backward locomotion (Fig. 3A, Supplementary Movie 3). Furthermore, both temporal and spatial motor features of the evoked behavioral responses were markedly similar to the motor dynamics observed for GH146II-GAL4 with subtracted GH146-QF as flies walked backwards in a persistent manner along the entire 2-s light stimulation window (Fig. 3A, Supplementary Fig. 2C). Hence, the parental larval AMB driver lines, R60F09- and R11E07-GAL4, may label MooSEZ of adult flies and thus might be utilized to assess MooSEZ expression pattern during development.
R60F09-GAL4 driver line targets adult MooSEZ
To find whether R60F09- and R11E07-GAL4 target the adult MooSEZ, we conducted anatomical analysis. Both lines exhibit broad expression patterns; while R11E07-GAL4 labels a significant amount of neurons in both the central brain and VNC (Fig. 3B), R60F09-GAL4 predominantly marks neurons within the central brain (Fig. 3C). Thus, it is possible that these drivers do not label MooSEZ, but rather other backward walking inducing neurons. To address this option, we conducted a series of behavioral and anatomical experiments. Since MooSEZ is a central brain neuron, we focused our experimental efforts on R60F09-GAL4.
We first performed a set of behavioral experiments to explore MooSEZ involvement in R60F09-GAL4-evoked backward locomotion phenotype. We optogenetically stimulated the neurons labeled by R60F09-LexA using CsChrimson while simultaneously silencing GH146II-GAL4 neurons by optogenetically activating the blue-gated Guillardia theta anion channelrhodopsin (GtACR2)48. GtACR2 activation in GH146II-GAL4 neurons eliminated the backward walking response evoked by activation of R60F09-LexA neurons (Supplementary Fig. 2D). In a complementary experiment, we blocked the activity of R60F09-LexA neurons by expressing tetanus toxin (TNT) light chain49, while optogenetically stimulating the neurons labeled by GH146II-GAL4 using CsChrimson. Inhibiting R60F09-LexA neurons strongly suppressed GH146II-GAL4-evoked backward locomotion (Supplementary Fig. 2E). The results of the reciprocal excitation-inhibition set of experiments demonstrate that neurons within R60F09 are functionally related to MooSEZs.
Next, we tested whether neurons within the SEZ, the brain region in which MooSEZ resides, underlie the observed behavioral effect. To this end, we utilized the Otd-nls:FLPo transgene50 that expresses flippase (FLP) in all central brain regions except for the gnathal ganglia in the SEZ51. Expressing Otd-nls:FLPo transgene along with R60F09-GAL4, UAS-CsChrimson, and tub-FRT-STOP-FRT-GAL80 significantly decreased CsChrimson expression in the adult nervous system except for the SEZ and VNC as the GAL4 repressor GAL8052,53 was expressed in the central brain except for the gnathal ganglia (Supplementary Fig. 2F). Optogenetic activation of this restricted neuronal subset led to pronounced backward locomotion that was similar to R60F09-GAL4 neuronal activation (Supplementary Fig. 2G), suggesting that neurons within the adult SEZ mediate R60F09-GAL4 backward walking phenotype.
Finally, we examined whether GH146II-GAL4 and R60F09 driver lines indeed target the same SEZ neuron in adult flies. Expressing mVenus in the cells covered by R60F09-LexA and mCherry in the neurons labeled by GH146II-GAL4 revealed co-expression in a single pair of lateral SEZ neurons (Fig. 3D). Taken together with the fact that MooSEZ, a bilateral SEZ neuron, exclusively mediates GH146II-GAL4-triggered backward walking37, the combined anatomical and behavioral results strongly indicate that the larval AMB driver line R60F09 labels MooSEZ in adult fly brains. Recalling that adult MooSEZ driver lines elicit larval backward crawling (Fig. 1A, B, D), and that MooSEZ already exists in larva (Fig. 1F, Supplementary Fig. 1B, C) and drives larval backward locomotion (Fig. 1G, Supplementary Fig. 1D), these results also imply that AMB and larval MooSEZ are identical.
R60F09-GAL4 driver line labels larval MooSEZ
To directly examine whether R60F09-GAL4 indeed labels larval MooSEZs and can thus be further utilized to explore MooSEZ developmental remodeling we used as above immortalization genetics. Applying RU-486 to larvae carrying UAS-hPR:FLP, act5C-FRT-STOP-FRT-LexA and LexAop-CsChrimson enabled us to irreversibly label R60F09-GAL4 larval neurons. Optogenetically stimulating RU-486-treated larvae elicited backward locomotion, which also persisted into adulthood as respective adult flies walked backwards upon light activation (Fig. 3E, Supplementary Fig. 3A). Conversely, RU-486 unexposed larvae did not perform backward locomotion at both larval and adult stages following optogenetic stimulation (Fig. 3E, Supplementary Fig. 3A). Remarkably, adult MooSEZ labeling was detected only in the brains of flies subjected to RU-486 during larval stages (Supplementary Fig. 3B). Since MooSEZ could have been labeled only during larval stages following RU-486 administration, we conclude that R60F09-GAL4 labels larval MooSEZ. Additionally, as AMB was shown to underlie R60F09-GAL4-evoked larval backward crawling45, these results indicate that AMB and larval MooSEZ are essentially the same neuron. Hence, the immortalization experimental results clearly demonstrate that R60F09-GAL4 can be further employed as a genetic tool to investigate MooSEZ developmental remodeling as it marks both larval and adult MooSEZ.
Constructing MooSEZ split-GAL4 driver lines and selectively manipulating MooSEZ activity
Thus far, we have found that R60F09-GAL4 is a broad driver line (Fig. 3C) that targets both larval and adult MooSEZ (Fig. 3D, E, Supplementary Fig. 2D-G). To gain selective and precise spatial access to MooSEZ, we sought to design split-GAL454,55 driver lines. To do so, we exploited NeuronBridge56,57 to query GAL4 drivers58 that cover MooSEZ, yet present minimal overlapping expression with all other neurons labeled by R60F09-GAL4 driver line. By screening through hundreds of potential driver lines suggested by the color depth searching method of NeuronBridge46, two candidate driver lines were identified: VT03276859- and VT020742-GAL459. The combination of R60F0947 and VT03276859 drives expression in adult MooSEZs and additional bilateral pair of descending neurons with somata at the medial anterior part of the SEZ (Fig. 3F). The combination of R60F0947 and VT02074259 drives selective expression in adult MooSEZs (Fig. 3G). Importantly, this combination also drives specific expression in larval MooSEZs (Supplementary Fig. 3C). The fantastic spatial specificity of the MooSEZ split GAL4 driver lines enabled us precise genetic access to manipulate MooSEZ activity and thus directly test its contribution to larval and adult behavioral output. Optogenetically activating MooSEZ using MooSEZ split GAL4 driver lines resulted in robust larval and adult backward locomotion (Supplementary Fig. 3D, E, Supplementary Movie 4). These results further confirm our earlier conclusion that MooSEZ is a preserved larval neuron with maintained locomotor functionality (Fig. 1F, G, Supplementary Fig. 1). Moreover, as MooSEZ split GAL4 driver lines had not been available thus far, repeating some of the experiments that had led to key findings presented in our previous work37 had a valuable importance. Consistent with our recent results37, selectively activating MooSEZ in flies with inhibited MDN resulted in decreased yet significant backward walking (Supplementary Fig. 3F). This result thus corroborates our original finding that MooSEZ elicits backward locomotion via both MDN-dependent and -independent pathways37. Furthermore, long optogenetic activation of MooSEZ in these flies generated sustained rotational responses, as previously observed for the GH146II-GAL4 driver line37, further validating MooSEZ's capacity to trigger robust MDN-independent turning responses (Supplementary Fig. 3G, H).
Tracking MooSEZ developmental trajectory
Having designed a GAL4 driver line that reliably and persistently labels MooSEZ at a single-cell resolution (Fig. 3G, Supplementary Fig. 3C), we conducted a detailed anatomical analysis of MooSEZ spatiotemporal dynamics through larval stages and subsequent metamorphosis. MooSEZ appears to be an embryonic-born neuron as it is detected already at the very early developmental stage of the 1st instar larva (L1) (Fig. 4A, A’). Its soma is located within the lateral aspect of the SEZ, with ipsilateral dendritic arborizations in both the SEZ and the thoracic segments of the VNC, and ipsilateral ascending axonal projections terminating dorso-posteriorly of the AL of the larval central brain. Hence, anatomy indicates that larval MooSEZ is an ascending neuron. As the larva progresses through development, MooSEZ exhibits a significant size expansion that is correlated with larval growth. Nonetheless, it maintains its distinct early larval morphological features (Fig. 4A–E’). However, by 8 hr after puparium formation (APF), the dendritic processes innervating the SEZ and VNC undergo dramatic pruning as indicated by their pronounced fragmentation (Fig. 4F’, F”). This regressive process continues at 18 hr APF (Fig. 4G’, G”) and appears to be completed by 24 hr APF (Fig. 4H’, H”). By contrast, the axonal processes ascending into the central brain seem to undergo only mild degeneration (Fig. 4F, F”, G, G”, H, H”). Thus, MooSEZ developmental neurite pruning is differentially modulated by neuronal compartmentalization. Regeneration of MooSEZ neurites commences between 18 hr and 24 hr APF as evident by increased innervations in the adult-forming SEZ and lateral accessory lobe (LAL) brain regions (Fig. 4G–H”). Notably, MooSEZ re-growing neurites do not reinvade the VNC. Hence, following initial pruning events and subsequent regrowth, the ascending larval MooSEZ transforms into a central brain intrinsic neuron. By 48 hr APF MooSEZ has acquired its final adult configuration, displaying elaborated arborizations in the SEZ and supraesophageal zone (SPZ) (Fig. 4I–L”). As previously reported37, MooSEZ adult morphology suggests dense dendritic arborizations in the SEZ, and presynaptic arborizations, organized in a vertical plane ascending anteromedially, along the frontal LAL. In addition, MooSEZ innervates the crepine (CRE), wedge (WED), vest (VES), and inferior posterior slope (IPS) brain regions. Noteworthily, MooSEZ’s time course of degenerative and subsequent regenerative events during metamorphosis largely corresponds to previously reported time frames observed for other well-established neuronal remodeling systems in Drosophila4,23.

A–E’ Top, expression pattern of MooSEZ at different larval developmental time points (A–D) and during puparium formation (E). CsChrimson.mVenus was selectively expressed in MooSEZ using the MooSEZ-specific split GAL4 driver line (VT020742-p65.ADՈR60F09-GAL4.DBD). Maximum intensity projections of 44-89 confocal sections (1 µm) are presented. Scale bar applies to all images. Bottom, schematic illustration of MooSEZ morphology at different larval developmental time points (A’–D’) and during puparium formation (E’) as presented above. F–L” Top and middle, expression pattern of MooSEZ at different developmental time points along pupal development (F–K’) and at the final adult stage (L-L’). CsChrimson.mVenus was selectively expressed in MooSEZ using the MooSEZ-specific split GAL4 driver line (VT020742-p65.ADՈR60F09-GAL4.DBD). Maximum intensity projections of 30-60 confocal sections (1 µm) through anterior (top) and posterior (middle) coronal stacks are presented. Purple arrows indicate VNC dendritic processes undergoing pruning. Dark and light red arrowheads indicate re-growing neurites in the SEZ and SPZ, respectively. Scale bar applies to all images. Bottom, schematic illustration of MooSEZ morphology at different pupal developmental time points (F”–K”) and at the final adult stage (L”) as presented above.
Cell-autonomous ecdysone signaling partially mediates MooSEZ pruning
The steroid hormone ecdysone (20-hydroxyecdysone) is a key regulator of insect developmental remodeling15,60. It mediates remodeling of various neuronal populations within the Drosophila nervous system26,27,61,62,63,64,65 via the nuclear heterodimer of Ultraspiracle (USP) and ecdysone receptor (EcR)66,67,68. Unliganded USP-EcR complex binds co-repressors and thereby inhibits transcription of downstream ecdysteroid-responsive genes. In contrast, the liganded complex drives gene expression by either activation (exchanging co-repressors with co-activators) or de-repression (co-repressors release)69,70. Interestingly, neurite pruning is mostly associated with ecdysteroid-induced gene activation, whereas subsequent outgrowth is predominantly linked to repression relief69,70.
We first sought to investigate whether ecdysteroid-mediated gene expression is required for MooSEZ pruning. To do so, we selectively expressed a dominant negative form of EcR, which cannot bind steroids due to a point mutation in its ligand-binding domain (EcRDN-B1(W650A))71 in MooSEZ. Importantly, EcRDN-B1(W650A) cannot support either gene activation or de-repression as it functions as a constitutive repressor69,70. Surprisingly, we were unable to detect any pruning defects in MooSEZ adult expression pattern, as unpruned neurites were not observed in both SEZ and VNC (Supplementary Fig. 4A). To exclude possible EcR isoform-specific effect, we also expressed in MooSEZ a different EcR isoform with identical mutation (EcRDN-A(W650A))70. In accordance with the results obtained for EcRDN-B1(W650A), no persisting larval neurites were identified in MooSEZ adult form (Supplementary Fig. 4B). This apparent lack of effect in adulthood has led us to monitor MooSEZ pruning throughout pupal development in a temporally-controlled manner. Strikingly, following EcRDN-B1(W650A) expression, larval branches within the VNC did exhibit local degeneration during early pupal stages (Fig. 5A’-A”). Nevertheless, the pruning phase was significantly extended due to slow fragmentation of MooSEZ processes (compare Figs. 4G’-G” with 5B’-B”, and 4H’-H” with 5C’-C”). By 48 hr APF MooSEZ completed its pruning (Fig. 5D’-D”) and maintained its anatomical structure throughout the remaining pupal stages (Fig. 5E’-F”). Taken together, these results demonstrate that expression of EcRDN-B1(W650A) does not block MooSEZ neurite pruning but rather alters the temporal dynamics of this process. Moreover, they also suggest that MooSEZ pruning is guided not only by cell-autonomous ecdysteroid signaling. Thus, MooSEZ pruning could be mediated via other interacting cells within MooSEZ vicinity27, as well as through steroid-independent mechanisms72.

A–F” Top and middle, expression pattern of MooSEZ expressing CsChrimson.mVenus and EcRDN-B1(W650A) at different developmental time points along pupal development (A–F’). MooSEZ-specific split GAL4 driver line (VT020742-p65.ADՈR60F09-GAL4.DBD) was used to drive UAS-CsChrimson.mVenus and UAS-EcRDN-B1(W650A). Maximum intensity projections of 30–45 confocal sections (1 µm) through anterior (top) and posterior (middle) coronal stacks are presented. Purple arrows indicate VNC dendritic processes undergoing pruning. Scale bar applies to all images. Expression of a dominant negative form of the ecdysone receptor B1 (W650A) in MooSEZ does not eliminate neurite pruning. Nevertheless, the pruning phase in which MooSEZ undergoes dendritic fragmentation is extended compared to wt genotype. Bottom, schematic illustration of MooSEZ morphology at different pupal developmental time points (A”–F”) as presented above. G-K’ Top, expression pattern of adult MooSEZ expressing CsChrimson.mVenus (G) and either EcRDN-B1(W650A) (H), EcRDN-A(W650A) (I), EcRDN-B1(F645A) (J), or EcRDN-A(F645A) (K). MooSEZ-specific split GAL4 driver line (VT020742-p65.ADՈR60F09-GAL4.DBD) was used to drive UAS-CsChrimson.mVenus and one of the UAS-EcR dominant negative constructs (when required) as designated. Maximum intensity projections of 65-100 confocal sections (1 µm) through the central brain are presented. Bottom, three-dimensional reconstructions of MooSEZ morphologies (G’-K’) obtained by tracing MooSEZ neurites in the confocal stacks used for constructing the above projection images. Size scale bar in (G) applies to all projection images (G-K) and three-dimensional reconstructions (G’-K’). Cyan gradient scale bar in (G’) applies to all three-dimensional reconstructions (G’-K’). L–N Analysis of the following quantitative metrics of adult morphology: cable length (L), number of branch points (M) and convex hull size (N) for MooSEZ expressing CsChrimson.mVenus in the absence or presence of either EcRDN-B1(W650A), EcRDN-A(W650A), EcRDN-B1(F645A), or EcRDN-A(F645A). MooSEZ-specific split GAL4 driver line (VT020742-p65.ADՈR60F09-GAL4.DBD) was used to drive UAS-CsChrimson.mVenus and one of the UAS-EcR dominant negative constructs (when required) as designated. MooSEZ displays severe outgrowth defects following expression of either of EcR dominant negative variants (n for each genotype = 10, *p < 0.05, **p < 0.01, ****p < 0.0001 for pairwise comparisons of wt with each experimental group, Kruskal-Wallis test followed by Dunn’s post-hoc tests). O Left, translational velocity ± SEM (shading) of adult flies expressing CsChrimson in the absence or presence of either EcRDN-B1(W650A), EcRDN-A(W650A), EcRDN-B1(F645A), or EcRDN-A(F645A) in MooSEZ following optogenetic activation. MooSEZ-specific split GAL4 driver line (VT020742-p65.ADՈR60F09-GAL4.DBD) was used to drive UAS-CsChrimson and one of the UAS-EcR dominant negative constructs (when required) as designated. The 2-s light pulse is labeled by light red. Expressing either of the EcR dominant negative variants abolishes MooSEZ-mediated adult backward walking. Right, mean translational velocity during the 2-s light pulse obtained from traces on the left. Manipulating ecdysone signaling by disrupting both activation and de-repression (EcRDN-B1(W650A) and EcRDN-A(W650A)), or only activation (EcRDN-B1(F645A) and EcRDN-A(F645A)) of ecdysone downstream genes eliminates MooSEZ-induced adult backward walking (n for VT020742-p65.AD&R60F09-GAL4.DBD > UAS-CsChrimson (II), VT020742-p65.AD&R60F09-GAL4.DBD > UAS-CsChrimson (III), VT020742-p65.AD&R60F09-GAL4.DBD > UAS-CsChrimson (II) UAS-EcRDN-B1(W650A), VT020742-p65.AD&R60F09-GAL4.DBD > UAS-CsChrimson (III) UAS-EcRDN-B1(F645A), VT020742-p65.AD&R60F09-GAL4.DBD > UAS-CsChrimson (III) UAS-EcRDN-A(W650A), VT020742-p65.AD&R60F09-GAL4.DBD > UAS-CsChrimson (III) UAS-EcRDN-A(F645A): 25, 33, 34, 33, 31, 29 respectively, ***p < 0.001, ****p < 0.0001 for pairwise comparisons of experimental groups with respective controls lacking EcRDN expression in MooSEZ, Kruskal-Wallis test followed by Dunn’s post-hoc tests). P Mean translational velocity following optogenetic activation (larvae, 9-s; adults, 2-s) of single larvae and their matched adult flies expressing CsChrimson in the presence (blue) or absence (brown) of EcRDN-B1(W650A) in MooSEZ. MooSEZ-specific split GAL4 driver line (VT020742-p65.ADՈR60F09-GAL4.DBD) was used to drive UAS-CsChrimson and UAS-EcRDN-B1(W650A) dominant negative construct (When required). Individual larvae and adult flies are matched by their identity. Backward crawling larvae expressing EcRDN-B1(W650A) do not persist into backward walking adult flies upon optogenetic activation of MooSEZ (n for each genotype (larva or adult) = 16, **p < 0.01, ****p < 0.0001 for main effect for genotype, and genotype X developmental stage interaction effect, respectively, repeated measures two-way ANOVA). For details of statistical analysis, see Supplementary Data 1.
MooSEZ outgrowth requires activation of ecdysteroid-responsive genes
Interestingly, expression of EcRDN-B1(W650A) resulted in severe outgrowth defects throughout development (Fig. 5A–F). We therefore next moved to explore the relationship between ecdysteroid signaling and MooSEZ outgrowth. For this experiment, we employed an additional set of EcR dominant negative isoforms: EcRDN-B1(F645A)71 and EcRDN-A(F645A)70 in addition to EcRDN-B1(W650A) and EcRDN-A(W650A). As these two additional receptors are not mutated in their ligand-binding domain, they can bind ecdysone and thus elicit de-repression through the release of co-repressors. However, they do have a point mutation in the co-activator binding site and hence cannot support gene activation. Accordingly, these receptors act as conditional repressors69,70. Since gene de-repression is presumed to underlie neurite outgrowth69,70, we hypothesized that expression of both EcRB1-W650A and EcRDN-A(W650A) would drastically impair MooSEZ outgrowth, whereas expression of both EcRDN-B1(F645A) and EcRDN-A(F645A) would leave MooSEZ outgrowth largely unaffected. However, to our surprise, expression of either EcR dominant negative variants resulted in notable remodeling defects; MooSEZ characteristic dendritic arborizations within the SEZ, and stereotypic presynaptic vertical sheet normally located posteriorly of the AL, were largely eliminated. Instead, ectopic thick branches with aberrant protrusions and reduced arborizations were detected in the SEZ and SPZ (Fig. 5G–K). We further traced MooSEZ neurites (Fig. 5G’–K’) and quantified the obtained anatomical results by computing the following metrics: cable length, number of branch points, and convex hull size (Fig. 5L–N, Supplementary Fig. 4C). Consistent with anatomy, quantitative analysis indicated that MooSEZ adult morphology was significantly disrupted not only for EcRDN-B1(W650A) and EcRDN-A(W650A) mutants, but also for EcRDN-B1(F645A) and EcRDN-A(F645A) mutants. The combined results thus indicate that ecdysteroid-mediated gene activation is indispensable for MooSEZ outgrowth. By contrast, de-repression, typically associated with neurite outgrowth69,70, does not seem to play a dominant role in MooSEZ outgrowth. Hence, by employing MooSEZ as a remodeling system, we identify a shared gene expression mechanism regulating both pruning and outgrowth.
Developmental neuronal remodeling is causally linked to MooSEZ-induced behavior
Developmental neuronal remodeling is considered crucial for the formation of mature functional circuits, which ultimately shape adult behavioral output. To probe causality, one should manipulate remodeling under experimentally-controlled conditions and closely monitor the behavioral outcome. However, thus far, there is no clear experimental evidence demonstrating a robust causal link between developmental remodeling and behavior. Previous studies have tested whether different aspects of adult MB-mediated functions require γ KC remodeling. Yet, impairing γ KCs remodeling resulted in only partial or weak effects on the behavioral output28,73. Additionally, a recent comprehensive anatomical analysis found developmentally segregated, rather than preserved, MB functional circuits for memory encoding in larva and adult23. Contrary to MB circuitry, MooSEZ and its downstream neural partners establish a persistent functional neural circuit that directly regulates a specific motor behavior at both larva and adult stages24,25,37. Furthermore, MooSEZ undergoes well-characterized stereotypic neuronal remodeling (Fig. 4F–L”) that can be precisely manipulated (Fig. 5A–N) using MooSEZ split GAL4 drivers (Fig. 3F, G, Supplementary Fig. 3C).
Hence, we sought to investigate causality between neuronal remodeling and behavior by utilizing MooSEZ as an experimental remodeling system. To test this, we selectively expressed CsChrimson in conjunction with each of the four EcR dominant negative variants used above using MooSEZ specific split GAL4 driver. Perturbing ecdysteroid signaling within MooSEZ virtually eliminated adult fly optogenetically-induced backward locomotion for all tested dominant negative variants (Fig. 5O, Supplementary Movie 5). These results directly show that ecdysteroid signaling is essential for MooSEZ-mediated backward walking. However, the above experiment does not unequivocally demonstrate that MooSEZ neuronal remodeling is causally related to behavior since the expression of EcR dominant negative variants was not temporally controlled. Theoretically, overexpression of an EcR dominant negative construct could affect MooSEZ functionality before metamorphosis since ecdysone is already secreted during embryogenesis and larval stages69. Thus, the observed behavioral phenotype could potentially arise from EcRDN-mediated perturbations unrelated to remodeling.
To conclusively determine whether MooSEZ remodeling is causally linked to adult behavior, we combined behavioral and anatomical approaches. We first compared between larval morphologies of MooSEZs that selectively expressed EcRDN-B1(W650A) and their wild type (wt) counterparts and found that their anatomical expression patterns were indistinguishable (compare Supplementary Fig. 4D with Supplementary Fig. 3C). Then, we compared the extent to which single larvae and their respective adult flies performed backward locomotion following optogenetic stimulation of MooSEZ in the presence or absence of EcRDN-B1(W650A). While larval backward locomotion phenotype was independent of EcRDN-B1(W650A) expression in MooSEZ, adult behavioral phenotype was abolished upon EcRDN-B1(W650A) expression (Fig. 5P). That is, backward crawling larvae expressing EcRDN-B1(W650A) in MooSEZ did not persist into backward walking adult flies following metamorphosis, as opposed to control larvae that did preserve their backward locomotion phenotype during adulthood. Hence, these behavioral results comply with our larval anatomical results (Supplementary Fig. 3C, Supplementary Fig. 4D) and indicate that remodeling defects occurring specifically during metamorphosis underlie the robust behavioral phenotype observed for EcRDN-B1(W650A) and most probably also for all other examined EcR dominant negative variants. Taken together, the combined results clearly show that cell-autonomous ecdysteroid signaling is crucial for regulating MooSEZ remodeling, which is essential for the behavioral output of the backward locomotion neural circuitry. In particular, these results demonstrate a clear causal relation between MooSEZ remodeling and adult behavioral response.
Discussion
In this study, we showed that the recently-reported MooSEZ persists through metamorphosis and can drive backward locomotion not only in adults but also in larvae (Fig. 1). AMBs, which were shown to elicit backward crawling in larvae45, are essentially the larval form of MooSEZs (Fig. 3). MooSEZ induces backward locomotion via direct connections with MDN in both larva and adult37,45. Whereas unilateral or long bilateral activation of larval MooSEZs drives symmetrical body wall muscle contractions and thus straight backward crawling, unilateral or long bilateral stimulation of adult MooSEZs triggers substantial backward turning (Fig. 2). In addition, using a new split GAL4 driver line, we selectively monitored MooSEZ spatiotemporal dynamics through postnatal development. We found that MooSEZ undergoes dramatic developmental reorganization as it transforms from a larval ascending neuron into an adult central brain intrinsic neuron (Fig. 4). We further revealed that MooSEZ pruning is partly mediated by cell-autonomous ecdysteroid-signaling and that MooSEZ outgrowth is largely regulated through gene activation rather than de-repression (Fig. 5). Finally, we demonstrated that MooSEZ ecdysteroid-mediated remodeling is causally linked to its adult functionality (Fig. 5).
Our work presents a new model system for studying developmental neuronal remodeling and its effect on behavior. The system is composed of a persistent neural network of remodeled neurons (MooSEZ37, MDN25,34 and Pair124,25) that directly regulates the motor output. We focused on a specific element within this system - MooSEZ, characterized its developmental trajectory, and discovered that it undergoes a unique developmental reorganization in which it transforms from a larval ascending neuron into an adult central brain neuron. Strikingly, the adult MooSEZ maintains its larval functional role. Moreover, using this system, we revealed a shared gene expression mechanism underlying neurite pruning and outgrowth. Furthermore, we found a clear causal relationship between developmental neuronal remodeling and behavior, as selective perturbation of MooSEZ remodeling resulted in pronounced behavioral deficits.
It is to be noted that in the current work we did not directly address MooSEZ-MDN connectivity. However, detailed EM connectivity analysis performed in FlyEM hemibrain74,75 and FlyWire76,77,78,79 demonstrated direct synaptic connections between MooSEZ and each of the four MDNs in the adult fly brain37. In addition, recent study found that the AMB, i.e., larval MooSEZ, forms direct synaptic connections with the larval MDN45. Thus, MooSEZ is a synaptic partner of MDN in both larvae and adults. Similarly, MDN and Pair1 neurons were also shown to be synaptic partners during larval stages25 and adulthood24. Their synaptic connectivity is first established at the late embryo stages, then eliminated ~24 hr APF, and finally re-formed ~48 hr APF24. Hence, it will be interesting to examine in future research whether the temporal dynamics characterizing the elimination and re-establishment of MooSEZ-MDN synaptic connectivity are similar to those previously described for the MDN-Pair1 neuronal pair.
Our results, combined with recent work showing that MDN and its postsynaptic partner Pair1 preserve their larval connectivity also in the adult24, indicate that MDN re-forms its larval synaptic connections with its pre- and post-synaptic partners (MooSEZ and Pair1, respectively) during the metamorphic transition. Collectively, our study demonstrates that a microcircuit motif, composed of a neuron and its pre- and post-synaptic partners, can maintain its connectivity and functionality despite dramatic postnatal neuronal modifications (Supplementary Fig. 5). Together, the current study utilizes a new model system within the Drosophila nervous system, the motor circuit regulating backward locomotion, to reveal preservation of a functional circuit motif.
Interestingly, contrary to the persistent motor phenotypes characterizing MDN (backward locomotion) and Pair1 neuron (forward locomotion inhibition), the motor program evoked by MooSEZ is differentially modulated by development (Supplementary Fig. 5); while larvae crawled in a consistent straight fashion, adults performed pronounced backward rotations following unilateral activation or long bilateral activation (Fig. 2). This result implies that lateralized and straight backward locomotions in larva are probably driven by segregated neuronal circuits, as opposed to a shared adult neuronal circuit underlying lateralized and straight backward locomotions. Hence, it appears that as the nervous system goes through profound reorganization during the transition from larva to adult, motor-related circuit motifs acquire additional functionalities to support a versatile and developmentally-relevant behavioral output.
Prior studies have identified several neuronal pairs that preserve their larval connectivity also in the adult (e.g., MDN and Pair124, MBON 01 and MBON 0923, OA-Vum2a and MBON 2223). However, thus far, it remained unknown whether a persistent neuron can maintain functional connections with both of its input and output partners. In the present study, we extended recent findings of Lee & Doe24, by showing that the MDN not only interacts with its postsynaptic partner Pair1, but also receives input from its presynaptic partner MooSEZ, to elicit backward locomotion in both larva and adult. Our work thus utilizes the motor circuit regulating backward locomotion in Drosophila to reveal circuit-level wiring of persistent functional connections.
MooSEZ synapses onto MDN, which interacts with VNC premotor neurons, inducing backward locomotion in larva24,25 and adult80. In addition, larval MDN also activates Pair1 neuron, which inhibits a premotor neuron involved in forward locomotion25. Although Pair1 and MDN re-form synaptic connections with each other, they do not seem to re-establish connectivity with their larval downstream targets in the adult VNC24. By contrast, the synaptic connectivity of MDN with its larval presynaptic partner MooSEZ is re-formed during the transition from larva to adult (Supplementary Fig. 5). Together, these observations imply that synapse re-establishment mechanisms between persistent neurons may be used more commonly in central brain circuits than in sensorimotor circuits located in the peripheral VNC. In line with this notion, a recent connectome-based study performed in Caenorhabditis elegans suggested that while peripheral neurons are dynamically remodeled during development, core motor circuits comprised of inter-connected interneurons are conserved and retain their functional roles81. The Drosophila brain is composed of a large number of intricately connected interneurons performing higher brain processing74,82, many of whom do not directly receive sensory inputs or send motor commands. If maintenance of inter-neuron connectivity is indeed employed by Drosophila as an adaptive neural mechanism, it is plausible to assume that stable connections between synaptically-coupled neurons exist on a much broader scale than has been documented to date. Thus, we speculate that additional persistent local central brain neurons may also function through conserved microcircuits. Notably, recent developments in connectomics now enable detailed synaptic-resolution mapping of circuits of interest in both larva83 and adult74,75,76,77 nervous systems. In this regard, it would be interesting to find whether other larval interneurons interacting with MDN and MooSEZ also persist into adulthood and re-establish their synaptic connections following metamorphosis. Revealing whether these neurons also play a role in the conserved neuronal circuit regulating backward locomotion may further our understanding of whether neural circuit maintenance is indeed used as a common strategy to maintain stable behavioral output following the transformation from larva to adult. Moreover, in a wider sense, broad maintenance of components and their respective connections within functional neural circuits may suggest that abstract drives (e.g., moving backwards) are encoded independent of developmental form (crawling larva or walking adult fly)84.
Our study reveals that cell-autonomous ecdysteroid signaling has only a moderate effect on MooSEZ pruning (Fig. 5A-F”, Supplementary Fig 4A, B). It was previously shown that the APL neuron undergoes ecdysteroid-mediated remodeling through interactions with its synaptic partner γ KC27. As MooSEZ and MDN are conserved synaptic partners, it will be interesting to uncover whether MooSEZ pruning is affected by MDN remodeling (and vice-versa) or any other synaptically-coupled neurons. In addition, it was shown that glial signaling is required for both ecdysteroid-dependent69,85,86 and -independent72 neuronal pruning. Hence, future research could examine whether glia cells mediate MooSEZ neurite fragmentation and depend upon ecdysteroid signaling. In contrast to the relatively mild effect observed for pruning, neurite outgrowth was dramatically affected by manipulating cell-autonomous ecdysteroid signaling in MooSEZ (Fig. 5G–N). Contrary to previous reports69,70, we find that gene activation rather than de-repression plays a central role in MooSEZ outgrowth. Future research could examine the molecular components underlying this mechanism and its prevalence across other remodeling systems.
Finally, our work presents a new model system within the Drosophila nervous system, the motor circuitry controlling backward locomotion, to investigate neuronal remodeling and its effect on behavior. The well-characterized connectivity of the circuit, combined with a low number of elements per neuronal subtype (MooSEZ-237; MDN-434; Pair1-225), and availability of subtype-specific driver lines24,25,34,87, provide an appealing system to mechanistically study developmental remodeling at both cellular and circuit levels. Cell-intrinsic and -extrinsic factors, previously-described in long-established remodeling systems in Drosophila4,27,88,89,90,91, could be further studied using this model system. Ultimately, as each of the persistent neuronal elements of the backward locomotion circuitry directly controls the motor output of the adult fly (as well as of the larva), the impact of developmental neuronal remodeling on the behavioral outcome, reflected by the outputted motor response, could be readily tested by employing this system.
Methods
Fly strains
Fly strains (see below) were reared at 25 °C on cornmeal agar under a 12 h light/12 h dark cycle. The following transgenes were used: GH146II-GAL4 (Bloomington, #30026), NP5288-GAL4 (KYOTO Stock Center, #104937), NP225-GAL4 (KYOTO Stock Center, #112095), GH146-QF (Bloomington, #30015), UAS-CsChrimson.mCherry (attP5) VT044845-LexAp65 (attP40) (MDN-LexA)36, nSyb-IVS-PhiC31 (attP18) (Bloomington, #84151), SPARC2-I-CsChrimson::tdTomato (Bloomington, #84144), UAS-CsChrimson.mVenus (attP40) (Bloomington, #55135), UAS-CsChrimson.mVenus (attP2) (Bloomington, #55136), UAS-GtACR2.EYFP (attP2) (Bloomington, #92987), tub-FRT-STOP-FRT-GAL80 (Bloomington, #39213), LexAop-CsChrimson.mVenus (attP40) (Bloomington, #55138), LexAop-CsChrimson.mVenus (attP2) (Bloomington, #55139), Otd-nls:FLPo50, QUAS-GAL8092, LexAop-TNT::HA (VK00033)93, R11E07-p65.AD (attP40) (Bloomington, #75953), R60F09-GAL4.DBD (attP2) (Bloomington, #75644), GMR60F09-GAL4 (attP2) (Bloomington, #39255), GMR60F09-LexA (attP40) (Bloomington, #61576), GMR11E07-GAL4 (attP2) (Bloomington, #48460), UAS-CsChrimson.mCherry (attP1) (Bloomington, #82182), VT020742-p65.AD (attP40) (Bloomington, #73460), VT032768-p65.AD (Bloomington, #71327), Actin5C-FRT > -dSTOP-FRT > -LexAp::65 in su(Hw)attP540, pJFRC108-20XUAS-IVS-hPR::Flp-p10 in VK0000540, UAS-EcRDN-A(W650A) (Bloomington, #9451), UAS-EcRDN-A(F645A) (Bloomington, #9452), UAS-EcRDN-B1(F645A) (Bloomington, #6869), UAS-EcRDN-B1(W650A) (VK00017)27.
Behavioral assays
Behavioral experiments were performed in either an open arena94 with a diameter of 60 mm composed of polyoxymethylene for testing adult flies or a 1% agarose arena with a diameter of 85 mm for testing larvae. The open arena was designed with sloped walls that limited flies to a shallow volume of space, and covered with a transparent acrylic plastic coated with Sigmacote (SL2; Sigma-Aldrich) to allow accurate and reliable measurement of locomotor behaviors94. The experimental arenas were illuminated from the bottom by a high-intensity 810 nm IR LED (SFH 4786S; OSRAM), while behavioral responses were recorded from above at 832×832 pixel resolution and 20 frames per second using a camera (PL-D795MU-T; PIXELINK) coupled to 16 mm focal length lens (NMV-16M11; NAVITAR) connected to 800 nm long-pass filter (LP800; MIDOPT). Optogenetic experiments were performed by illuminating either 617 nm red light pulse (M617L3 LED; THORLABS) for CsChrimson activation, or 470 nm blue light pulse (M470L3 LED; THORLABS) for GtACR2 activation. Larvae were illuminated with a red light intensity of either ~0.7 mW/cm2 (for activation of broad PN driver lines, and immortalized GH146II-GAL4 neurons) or ~26 mW/cm2 (in all other experiments); flies were illuminated with a red light intensity of either ~8 mW/cm2 (for immortalized GH146II-GAL4 neurons) or ~26 mW/cm2 (in all other experiments). For GtACR2 activation, flies were illuminated with a blue light intensity of ~24 mW/cm2. Pixelink Capture OEM software was used for video recordings. A custom-written software (LabVIEW 7.1, National Instruments) controlled the delivery of light by the LEDs and recording of videos by the camera via TTL inputs.
Adult flies
Behavioral experiments were performed on ~5-10 days post-eclosion male and female flies, except for SPARC41 experiments in which only female flies were tested (see below). Flies grown on standard food (cornmeal agar) in the dark were collected ~2–5 days post-eclosion and reared for additional ~3–5 days on 1 mM all-trans retinal (R2500; Sigma-Aldrich) supplemented food before experimental testing. Flies were examined either individually (for experiments in which larva and adult were matched by identity) or in groups of 3-15 flies. Tested flies were placed in the open arena and allowed to walk freely for ~1–3 minutes before the start of an experimental trial in which a light pulse was delivered.
Larvae
Behavioral experiments were performed on third instar larvae of both sexes, except for SPARC41 experiments in which only female larvae were tested (see below). Larvae were reared on 1 mM all-trans retinal (R2500; Sigma-Aldrich) supplemented food during development. For immortalization23,40 experiments, RU-486 was added to larval food vials 24–48 hr before experimental testing (see below). Larvae were examined either individually (for experiments in which larva and adult were matched by identity) or in groups of 3-8 larvae. Larvae grown in complete darkness throughout development were transferred to an 85 mm diameter 1% agarose arena and crawled freely before the start of an experimental trial, in which a light pulse was delivered.
Immortalization experiments
Mifepristone (RU-486) inducible flippase (UAS-hPR:FLP) was used along with act5C-FRT-STOP-FRT-LexA, and LexAop-CsChrimson to permanently mark either GH146II-GAL4 or R60F09-GAL4 larval neurons. During larval development, 60 µl of 10 mM RU-486 (M8046; Sigma-Aldrich) stock solution was added to the cornmeal agar that larvae were fed on. At 24–48 hours after RU-486 application, single larvae were transferred to the agarose arena and tested for backward crawling upon 9-s red light stimulation. Individual larvae were then grown to adulthood and retested for adult backward locomotion following a 1.5-s light stimulation in the open arena behavioral assay. Finally, adult fly brains were dissected, stained, and imaged to examine LexAop-CsChrimson expression in MooSEZ.
SPARC experiments
The SPARC41 genetic method was used to sparsely and stochastically target fractions of the neurons labeled by GH146II-GAL4 driver in female larvae. SPARC mosaic larvae were placed in the agarose arena and exposed to a 9-s red light stimulation. Following metamorphosis, adult female flies were loaded in the open arena and retested for backward locomotion using a 2-s red light pulse. Finally, their brains were dissected, stained, and imaged to examine CsChrimson expression in MooSEZ.
MooSEZ split GAL4 driver lines design
NeuronBridge56,57 was used to search for split GAL4 fragments that best combine with the R60F09 driver line to target MooSEZ (FlyEM hemibrain74 fragment #581304618537). From hundreds of candidate GAL4 driver lines58 suggested by the color-depth algorithm implemented in the NeuronBridge search tool46, two driver lines were visually identified as potential matches: VT02074259, and VT03276859. Both lines demonstrated overlapping expression with R60F0947 in MooSEZ and little to no overlap in off-target neurons. Hence, two split GAL4 combinations for MooSEZ selective labeling were created: VT020742-p65.ADՈR60F09-GAL4.DBD, and VT032768-p65.ADՈR60F09-GAL4.DBD.
Structural imaging
Dissections, fixation, and immunostaining of larva, pupa, and adult nervous systems were conducted as previously described37,95. Briefly, dissected brains and VNCs were fixed in 4% [w/v] paraformaldehyde in PBS for at least 20 minutes at room temperature. Samples were then washed for 20 minutes 3 times with PBS containing 0.3% (v/v) Triton-X-100 (PBT) and blocked for at least 35 minutes using 5% normal goat serum. Following incubation with primary antibodies for at least 2 days, samples were washed three times with PBT and incubated for at least 2 additional days with secondary antibodies. Finally, samples were washed in PBT and embedded in Vectashield plus (Vector Laboratories H-1900). Primary antibodies used: rabbit anti-GFP (Thermo Fisher Scientific A-11122, 1:250), rabbit anti-RFP (Abcam ab62341, 1:250), mouse anti-Bruchpilot (DSHB, 1:100). Secondary antibodies used: Alexa Fluor 488 goat anti-rabbit (Thermo Fisher Scientific A-11034, 1:250), Alexa Fluor 568 goat anti-rabbit (Abcam ab175471, 1:500), Alexa Fluor 647 goat anti-mouse (Abcam ab150115, 1:500). Larval, pupal, and adult samples were mounted between tape spacers, enabling bi-directional scanning. Imaging stacks were obtained using Leica TCS SP5 confocal (LAS AF software v2.7.3.9723) with HCX PL APO lambda blue 20.0 × 0.70 IMM UV objective, and further processed in Fiji/ImageJ96.
Three-dimensional light microscopy reconstruction
Tracing of adult MooSEZ neurites was performed semi-automatically using SNT toolbox97 in Fiji/ImageJ96. Three-dimensional reconstructions of MooSEZ traced structures were visualized using SNT’s reconstruction viewer.
Quantification of behavioral experiments
Both larva and fly behavioral responses were analyzed with Ctrax98, which allows to track the motion of objects along a recorded video. Following tracking with Ctrax, tracking errors were fixed using the “FixErrors Matlab GUI”. Only flies and larvae that moved in an unconstrained manner, exhibited coordinated locomotor patterns, and were successfully tracked by Ctrax were further used for analysis. The script “compute_perframe_stats” contained within the Ctrax “BehavioralMicroarray” MATLAB toolbox was utilized to compute per-frame statistics of locomotor activity. Final analysis was performed on the parameters outputted by “compute_perframe_stats” script using custom MATLAB scripts (except for larval number of peristaltic backward waves, see below). Translational velocity was defined as the velocity vector’s projection on the anterior-posterior orientation axis of the tracked animal. Mean translational velocity was computed as the average across animals of per-animal translational velocity mean values during the light stimulation window. Cumulative angular change was computed by cumulatively integrating the angular velocity as a function of time. Angular velocity was defined as an orientation angle change in relation to a global orientation coordinate system. Left and right rotations were defined as positive and negative angular velocity values, respectively. Accordingly, positive and negative values of cumulative angular change signified continuous leftward and rightward movements, respectively. Mean angular velocity was computed as the average across animals of per-animal angular velocity mean values during the light stimulation window. Accordingly, positive and negative values of mean angular velocity signified average leftward and rightward movements, respectively. Angular speed was calculated as the absolute value of angular velocity. Mean angular speed was computed as the average across animals of per-animal angular speed mean values during the light stimulation window. Absolute cumulative angular change was computed by cumulatively integrating the angular speed as a function of time. Absolute total angular change was computed by integrating the angular speed as a function of time during the light stimulation window. Larval number of peristaltic backward waves was quantified by visually counting backward waves generated by each larva during the light stimulation window. Mean larval number of peristaltic backward waves was computed as the average across animals of larval number of peristaltic backward wave values during the light stimulation window.
Quantification of MooSEZ adult morphology
Quantitative analysis of MooSEZ adult morphology was performed with SNT toolbox97 in Fiji/ImageJ96. Cable length metric was computed as the total length of all traced branches. Mean cable length was computed as the average across animals of cable length values. Number of branch points metric was computed as the total count of branch points along MooSEZ-traced segments. Mean number of branch points was computed as the average across animals of number of branch point values. Convex hull size was computed as the smallest convex bounding volume of the MooSEZ traced structure. Mean convex hull size was computed as the average across animals of convex hull size values.
Statistics and reproducibility
Statistical testing and parameter extraction were carried out in GraphPad Prism 10.2.0. Details of all statistical tests are specified in Supplementary Data 1. Sample sizes were based on typical sample sizes in the field and prior experience with similar behavioral and imaging paradigms. Measurements were taken from distinct samples except for samples in which each animal was tested twice (at both larval and adult stages). Significance was defined as a p-value smaller than 0.05. Statistical significance was determined using two-sided nonparametric tests except for repeated measures two-way ANOVA performed on standardized z-scores following normality testing (Fig. 5P, Supplementary Figs. 1B, 3A). Multiple comparisons were corrected by controlling the false discovery rate (FDR)99. Result graphs were generated using custom MathWorks MATLAB R2021b codes and GraphPad Prism 10.2.0. For presentation of translational velocity and angular speed traces with shaded error bars, the boundedline MATLAB function was used (https://github.com/kakearney/boundedline-pkg). Adobe illustrator 29.5.1 was used for final figure visual editing.
Movie clips
Video clips were edited using the video editing software Kapwing (https://www.kapwing.com).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data used to generate Figs. 1–5, supplementary Figs. 1–5, and Supplementary Movies 1–5 are available in Figshare with identifier: https://doi.org/10.6084/m9.figshare.29950172100. Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article. The study did not generate any new unique reagents, code, or dataset. Requests for fly stocks used in the study or for additional information required to reanalyze the data reported in the paper should be directed to and will be fulfilled by the corresponding author, Moshe Parnas (mparnas@tauex.tau.ac.il).
References
Innocenti, G. M. & Price, D. J. Exuberance in the development of cortical networks. Nat. Rev. Neurosci. 6, 955–965 (2005).
Luo, L. & O’Leary, D. D. M. Axon Retraction and Degeneration in Development and Disease. Annu. Rev. Neurosci. 28, 127–156 (2005).
Yu, F. & Schuldiner, O. Axon and dendrite pruning in Drosophila. Curr. Opin. Neurobiol. 27, 192–198 (2014).
Yaniv, S. P. & Schuldiner, O. A fly’s view of neuronal remodeling. WIREs Dev. Biol. 5, 618–635 (2016).
Innocenti, G. M. Growth and reshaping of axons in the establishment of visual callosal connections. Science 212, 824–827 (1981).
Tang, G. et al. Loss of mTOR-Dependent Macroautophagy Causes Autistic-like Synaptic Pruning Deficits. Neuron 83, 1131–1143 (2014).
Faust, T. E., Gunner, G. & Schafer, D. P. Mechanisms governing activity-dependent synaptic pruning in the developing mammalian CNS. Nat. Rev. Neurosci. 22, 657–673 (2021).
Cocchi, E., Drago, A. & Serretti, A. Hippocampal Pruning as a New Theory of Schizophrenia Etiopathogenesis. Mol. Neurobiol. 53, 2065–2081 (2016).
Sekar, A. et al. Schizophrenia risk from complex variation of complement component 4. Nature 530, 177–183 (2016).
Yilmaz, M. et al. Overexpression of schizophrenia susceptibility factor human complement C4A promotes excessive synaptic loss and behavioral changes in mice. Nat. Neurosci. 24, 214–224 (2021).
Thomas, M. S. C., Davis, R., Karmiloff-Smith, A., Knowland, V. C. P. & Charman, T. The over-pruning hypothesis of autism. Dev Sci. 19, 284–305 (2016).
Penzes, P., Cahill, M. E., Jones, K. A., VanLeeuwen, J.-E. & Woolfrey, K. M. Dendritic spine pathology in neuropsychiatric disorders. Nat. Neurosci. 14, 285–293 (2011).
Truman, J. W. The Evolution of Insect Metamorphosis. Curr. Biol. 29, R1252–R1268 (2019).
Consoulas, C., Duch, C., Bayline, R. J. & Levine, R. B. Behavioral transformations during metamorphosis: remodeling of neural and motor systems. Brain Res. Bull. 53, 571–583 (2000).
Levine, R. B., Morton, D. B. & Restifo, L. L. Remodeling of the insect nervous system. Curr. Opin. Neurobiol. 5, 28–35 (1995).
Truman, J. W. Metamorphosis of the central nervous system of Drosophila. J. Neurobiol. 21, 1072–1084 (1990).
Tissot, M. & Stocker, R. F. Metamorphosis in Drosophila and other insects: the fate of neurons throughout the stages. Prog. Neurobiol. 62, 89–111 (2000).
Owald, D., Lin, S. & Waddell, S. Light, heat, action: neural control of fruit fly behaviour. Philos. Trans. R. Soc. B: Biol. Sci. 370, 20140211 (2015).
Luo, L., Callaway, E. M. & Svoboda, K. Genetic Dissection of Neural Circuits: A Decade of Progress. Neuron 98, 256–281 (2018).
Furusawa, K. & Emoto, K. Spatiotemporal regulation of developmental neurite pruning: Molecular and cellular insights from Drosophila models. Neurosci. Res. 167, 54–63 (2021).
Schuldiner, O. & Yaron, A. Mechanisms of developmental neurite pruning. Cell. Mol. Life Sci. 72, 101–119 (2015).
Veverytsa, L. & Allan, D. W. Subtype-specific neuronal remodeling during Drosophila metamorphosis. Fly https://doi.org/10.4161/fly.23969 (2013).
Truman, J. W., Price, J., Miyares, R. L. & Lee, T. Metamorphosis of memory circuits in Drosophila reveals a strategy for evolving a larval brain. eLife 12, e80594 (2023).
Lee, K. & Doe, C. Q. A locomotor neural circuit persists and functions similarly in larvae and adult Drosophila. eLife 10, e69767 (2021).
Carreira-Rosario, A. et al. MDN brain descending neurons coordinately activate backward and inhibit forward locomotion. Elife 7, e38554 (2018).
Lee, T., Marticke, S., Sung, C., Robinow, S. & Luo, L. Cell-Autonomous Requirement of the USP/EcR-B Ecdysone Receptor for Mushroom Body Neuronal Remodeling in Drosophila. Neuron 28, 807–818 (2000).
Mayseless, O. et al. Developmental Coordination during Olfactory Circuit Remodeling in Drosophila. Neuron 99, 1204–1215.e5 (2018).
Poppinga, H. et al. Pruning deficits of the developing Drosophila mushroom body result in mild impairment in associative odour learning and cause hyperactivity. Open Biol. 12, 220096 (2022).
Yoshino, J., Morikawa, R. K., Hasegawa, E. & Emoto, K. Neural Circuitry that Evokes Escape Behavior upon Activation of Nociceptive Sensory Neurons in Drosophila Larvae. Curr. Biol. 27, 2499–2504.e3 (2017).
Imambocus, B. N. et al. A neuropeptidergic circuit gates selective escape behavior of Drosophila larvae. Curr. Biol. 32, 149–163.e8 (2022).
Ohyama, T. et al. A multilevel multimodal circuit enhances action selection in Drosophila. Nature 520, 633–639 (2015).
Burgos, A. et al. Nociceptive interneurons control modular motor pathways to promote escape behavior in Drosophila. eLife 7, e26016 (2018).
Hu, C. et al. Sensory integration and neuromodulatory feedback facilitate Drosophila mechanonociceptive behavior. Nat. Neurosci. 20, 1085–1095 (2017).
Bidaye, S. S., Machacek, C., Wu, Y. & Dickson, B. J. Neuronal control of Drosophila walking direction. Science 344, 97–101 (2014).
Sen, R., Wang, K. & Dickson, B. J. TwoLumps Ascending Neurons Mediate Touch-Evoked Reversal of Walking Direction in Drosophila. Curr. Biol. 29, 4337–4344.e5 (2019).
Sen, R. et al. Moonwalker Descending Neurons Mediate Visually Evoked Retreat in Drosophila. Curr. Biol. 27, 766–771 (2017).
Israel, S., Rozenfeld, E., Weber, D., Huetteroth, W. & Parnas, M. Olfactory stimuli and moonwalker SEZ neurons can drive backward locomotion in Drosophila. Curr. Biol. 32, 1131–1149.e7 (2022).
Klapoetke, N. C. et al. Independent optical excitation of distinct neural populations. Nat. Methods 11, 338–346 (2014).
Potter, C. J., Tasic, B., Russler, E. V., Liang, L. & Luo, L. The Q System: A Repressible Binary System for Transgene Expression, Lineage Tracing, and Mosaic Analysis. Cell 141, 536–548 (2010).
Harris, R. M., Pfeiffer, B. D., Rubin, G. M. & Truman, J. W. Neuron hemilineages provide the functional ground plan for the Drosophila ventral nervous system. eLife 4, e04493 (2015).
Isaacman-Beck, J. et al. SPARC enables genetic manipulation of precise proportions of cells. Nat. Neurosci. 23, 1168–1175 (2020).
Wang, J. et al. Drosophila Larval Light-Avoidance is Negatively Regulated by Temperature Through Two Pairs of Central Brain Neurons. Neurosci. Bull. 38, 200–204 (2022).
Zhao, Q. et al. A two-layer neural circuit controls fast forward locomotion in Drosophila. Curr. Biol. 34, 3439–3453.e5 (2024).
Lehman, M. et al. Neural circuits underlying context-dependent competition between defensive actions in Drosophila larvae. Nat. Commun. 16, 1120 (2025).
Omamiuda-Ishikawa, N., Sakai, M. & Emoto, K. A pair of ascending neurons in the subesophageal zone mediates aversive sensory inputs-evoked backward locomotion in Drosophila larvae. PLOS Genet. 16, e1009120 (2020).
Otsuna, H., Ito, M. & Kawase, T. Color Depth MIP Mask Search: A New Tool to Expedite Split-GAL4 Creation. 318006 (2018) https://doi.org/10.1101/318006.
Dionne, H., Hibbard, K. L., Cavallaro, A., Kao, J.-C. & Rubin, G. M. Genetic Reagents for Making Split-GAL4 Lines in Drosophila. Genetics 209, 31–35 (2018).
Mohammad, F. et al. Optogenetic inhibition of behavior with anion channelrhodopsins. Nat. Methods 14, 271–274 (2017).
Sweeney, S. T., Broadie, K., Keane, J., Niemann, H. & O’Kane, C. J. Targeted expression of tetanus toxin light chain in Drosophila specifically eliminates synaptic transmission and causes behavioral defects. Neuron 14, 341–351 (1995).
Asahina, K. et al. Tachykinin-Expressing Neurons Control Male-Specific Aggressive Arousal in Drosophila. Cell 156, 221–235 (2014).
Aranha, M. M. et al. apterous Brain Neurons Control Receptivity to Male Courtship in Drosophila Melanogaster Females. Sci. Rep. 7, 46242 (2017).
Ma, J. & Ptashne, M. The carboxy-terminal 30 amino acids of GAL4 are recognized by GAL80. Cell 50, 137–142 (1987).
Suster, M. L., Seugnet, L., Bate, M. & Sokolowski, M. B. Refining GAL4-driven transgene expression in Drosophila with a GAL80 enhancer-trap. Genesis 39, 240–245 (2004).
Luan, H., Peabody, N. C., Vinson, C. R. & White, B. H. Refined Spatial Manipulation of Neuronal Function by Combinatorial Restriction of Transgene Expression. Neuron 52, 425–436 (2006).
Pfeiffer, B. D. et al. Refinement of Tools for Targeted Gene Expression in Drosophila. Genetics 186, 735–755 (2010).
Clements, J. et al. NeuronBridge: an intuitive web application for neuronal morphology search across large data sets. BMC Bioinformatics 25, 114 (2024).
Bogovic, J. A. et al. An unbiased template of the Drosophila brain and ventral nerve cord. PLOS ONE 15, e0236495 (2020).
Meissner, G. W. et al. A searchable image resource of Drosophila GAL4 driver expression patterns with single neuron resolution. eLife 12, e80660 (2023).
Tirian, L. & Dickson, B. J. The VT GAL4, LexA, and split-GAL4 driver line collections for targeted expression in the Drosophila nervous system. Preprint at https://doi.org/10.1101/198648 (2017).
Thummel, C. S. Flies on steroids — Drosophila metamorphosis and the mechanisms of steroid hormone action. Trends Genet. 12, 306–310 (1996).
Marin, E. C., Watts, R. J., Tanaka, N. K., Ito, K. & Luo, L. Developmentally programmed remodeling of the Drosophila olfactory circuit. Development 132, 725–737 (2005).
Kuo, C. T., Jan, L. Y. & Jan, Y. N. Dendrite-specific remodeling of Drosophila sensory neurons requires matrix metalloproteases, ubiquitin-proteasome, and ecdysone signaling. Proc. Natl. Acad. Sci. 102, 15230–15235 (2005).
Schubiger, M., Wade, A. A., Carney, G. E., Truman, J. W. & Bender, M. Drosophila EcR-B ecdysone receptor isoforms are required for larval molting and for neuron remodeling during metamorphosis. Development 125, 2053–2062 (1998).
Sakamura, S. et al. Ecdysone signaling determines lateral polarity and remodels neurites to form Drosophila’s left-right brain asymmetry. Cell Rep. 112337 (2023) https://doi.org/10.1016/j.celrep.2023.112337.
Wong, K. K. L. et al. Origin of wiring specificity in an olfactory map revealed by neuron type–specific, time-lapse imaging of dendrite targeting. eLife 12, e85521 (2023).
Hall, B. L. & Thummel, C. S. The RXR homolog Ultraspiracle is an essential component of the Drosophila ecdysone receptor. Development 125, 4709–4717 (1998).
Yao, T.-P., Segraves, W. A., Oro, A. E., McKeown, M. & Evans, R. M. Drosophila ultraspiracle modulates ecdysone receptor function via heterodimer formation. Cell 71, 63–72 (1992).
Yao, T.-P. et al. Functional ecdysone receptor is the product of EcR and Ultraspiracle genes. Nature 366, 476–479 (1993).
Truman, J. W. & Riddiford, L. M. Drosophila postembryonic nervous system development: a model for the endocrine control of development. Genetics 223, iyac184 (2023).
Brown, H. L. D., Cherbas, L., Cherbas, P. & Truman, J. W. Use of time-lapse imaging and dominant negative receptors to dissect the steroid receptor control of neuronal remodeling in Drosophila. Development 133, 275–285 (2006).
Cherbas, L., Hu, X., Zhimulev, I., Belyaeva, E. & Cherbas, P. EcR isoforms in Drosophila: testing tissue-specific requirements by targeted blockade and rescue. Development 130, 271–284 (2003).
Lehmann, K. S. et al. Astrocyte-dependent local neurite pruning in Beat-Va neurons. J. Cell Biol. 224, e202312043 (2024).
Redt-Clouet, C. et al. Mushroom body neuronal remodelling is necessary for short-term but not for long-term courtship memory in Drosophila. Eur. J. Neurosci. 35, 1684–1691 (2012).
Scheffer, L. K. et al. A connectome and analysis of the adult Drosophila central brain. eLife 9, e57443 (2020).
Plaza, S. M. et al. neuPrint: An open access tool for EM connectomics. Front. Neuroinformatics 16, (2022).
Dorkenwald, S. et al. Neuronal wiring diagram of an adult brain. Nature 634, 124–138 (2024).
Zheng, Z. et al. A Complete Electron Microscopy Volume of the Brain of Adult Drosophila melanogaster. Cell 174, 730–743.e22 (2018).
Buhmann, J. et al. Automatic detection of synaptic partners in a whole-brain Drosophila electron microscopy data set. Nat. Methods 18, 771–774 (2021).
Heinrich, L., Funke, J., Pape, C., Nunez-Iglesias, J. & Saalfeld, S. Synaptic Cleft Segmentation in Non-isotropic Volume Electron Microscopy of the Complete Drosophila Brain. in Medical Image Computing and Computer Assisted Intervention – MICCAI 2018 (eds Frangi, A. F., Schnabel, J. A., Davatzikos, C., Alberola-López, C. & Fichtinger, G.) 317–325 (Springer International Publishing, Cham, 2018). https://doi.org/10.1007/978-3-030-00934-2_36.
Feng, K. et al. Distributed control of motor circuits for backward walking in Drosophila. Nat. Commun. 11, 6166 (2020).
Witvliet, D. et al. Connectomes across development reveal principles of brain maturation. Nature 596, 257–261 (2021).
Lin, A. et al. Network statistics of the whole-brain connectome of Drosophila. Nature 634, 153–165 (2024).
Winding, M. et al. The connectome of an insect brain. Science 379, eadd9330 (2023).
Büschges, A. & Ache, J. M. Motor control on the move: from insights in insects to general mechanisms. Physiol, Rev. 105, 975–1031 (2025).
Awasaki, T. & Ito, K. Engulfing Action of Glial Cells Is Required for Programmed Axon Pruning during Drosophila Metamorphosis. Curr. Biol. 14, 668–677 (2004).
Awasaki, T., Huang, Y., O’Connor, M. B. & Lee, T. Glia instruct developmental neuronal remodeling through TGF-β signaling. Nat. Neurosci. 14, 821–823 (2011).
Tastekin, I. et al. Sensorimotor pathway controlling stopping behavior during chemotaxis in the Drosophila melanogaster larva. eLife 7, e38740 (2018).
Riccomagno, M. M. & Kolodkin, A. L. Sculpting Neural Circuits by Axon and Dendrite Pruning. Annu Rev. Cell Dev. Biol. 31, 779–805 (2015).
Rumpf, S., Wolterhoff, N. & Herzmann, S. Functions of Microtubule Disassembly during Neurite Pruning. Trends Cell Biol. 29, 291–297 (2019).
Meltzer, H. & Schuldiner, O. With a little help from my friends: how intercellular communication shapes neuronal remodeling. Curr. Opin. Neurobiol. 63, 23–30 (2020).
Mayseless, O., Shapira, G., Rachad, E. Y., Fiala, A. & Schuldiner, O. Neuronal excitability as a regulator of circuit remodeling. Curr. Biol. 33, 981–989.e3 (2023).
Potter, C. J. & Luo, L. Using the Q system in Drosophila melanogaster. Nat. Protoc. 6, 1105–1120 (2011).
Karuppudurai, T. et al. A Hard-Wired Glutamatergic Circuit Pools and Relays UV Signals to Mediate Spectral Preference in Drosophila. Neuron 81, 603–615 (2014).
Simon, J. C. & Dickinson, M. H. A New Chamber for Studying the Behavior of Drosophila. PLOS ONE 5, e8793 (2010).
Wu, J. S. & Luo, L. A protocol for dissecting Drosophila melanogaster brains for live imaging or immunostaining. Nat. Protoc. 1, 2110–2115 (2006).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Arshadi, C., Günther, U., Eddison, M., Harrington, K. I. S. & Ferreira, T. A. SNT: a unifying toolbox for quantification of neuronal anatomy. Nat. Methods 18, 374–377 (2021).
Branson, K., Robie, A., Bender, J., Perona, P. & Dickinson, M. High-throughput Ethomics in Large Groups of Drosophila. Nat. Methods 6, 451–457 (2009).
Benjamini, Y. & Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc.: Ser. B (Methodol.) 57, 289–300 (1995).
Israel, S. & Parnas, M. Experimental data of the manuscript: ‘A new system for studying developmental neuronal remodeling and its relation to behavior in Drosophila’ by Israel and Parnas, 2025. figshare https://doi.org/10.6084/m9.figshare.29950172 (2025).
Acknowledgements
We thank Dr. James W. Truman, Dr. Barry J. Dickson, Dr. Adam Claridge-Chang, Dr. Andrew Lin, Dr. Stephen F. Goodwin, Dr. Oren Schuldiner, Dr. Marta Zlatic, the Bloomington Drosophila Stock Center (BDSC), and the Department of Drosophila Genomics and Genetic Resources (DGGR) at Kyoto Institute of Technology for fly strains. We also thank Dr. Wolf Huetteroth for advice on analysis of larval anatomy. This work was supported by the European Research Council (ERC, 101085605, MP), United States-Israel binational Science Foundation (BSF, 2020636, MP) and Israel Science Foundation (ISF, 404/23, MP).
Author information
Authors and Affiliations
Contributions
S.I.: Initiated the project, conceptualization, methodology, investigation, formal analysis, software, writing–original draft, writing–review & editing, visualization. M.P.: Initiated the project, conceptualization, methodology, writing–original draft, writing–review & editing, visualization, supervision, and funding acquisition.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Tamara Boto and the other anonymous reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Dr Jun Wei Pek and Dr Ophelia Bu. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Israel, S., Parnas, M. A new system for studying neuronal remodeling and its relation to behavior in Drosophila. Commun Biol 8, 1830 (2025). https://doi.org/10.1038/s42003-025-09344-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-025-09344-6
