
Abstract
Postoperative cognitive dysfunction (POCD) is common in older surgical patients, yet its mechanistic underpinnings remain incompletely defined. Building on the premise that immune-cell metabolism shapes inflammatory signaling, we investigated whether microglia export metabolic instructions through extracellular vesicles (EVs) to remodel astrocytic function. We show that plasma EVs from POCD patients and EVs from activated microglia are enriched for ATP citrate lyase (ACLY) and are efficiently taken up by astrocytes. EV-delivered ACLY increases acetyl-CoA availability and enhances NF-κB p65 acetylation, which in turn suppresses the astrocytic glutamate transporter EAAT1 (SLC1A3). This program diminishes glutamate clearance, leading to synaptic dysfunction and cognitive impairment. In aged mice, systemic or hippocampal EV administration recapitulated microgliosis/astrogliosis, dendritic spine loss, impaired long-term potentiation, and deficits in the Morris water maze and novel object recognition. Chromatin immunoprecipitation confirmed acetyl-p65 occupancy at the SLC1A3 promoter, linking ACLY activity to direct transcriptional repression of EAAT1. Interventions that blocked EV biogenesis/uptake or reduced ACLY (genetically or pharmacologically) attenuated p65 acetylation, restored EAAT1 expression and glutamate uptake, and improved synaptic and behavioral outcomes. These findings identify a microglia-to-astrocyte ACLY–p65–EAAT1 axis that couples immunometabolism to excitatory neurotransmitter dysregulation and cognitive decline in POCD, highlighting ACLY and EV pathways as actionable therapeutic targets.

Similar content being viewed by others

Introduction
Postoperative cognitive dysfunction (POCD) represents a prevalent yet underrecognized neurocognitive complication following surgery, particularly prevalent among elderly patients1,2. Characterized by deficits in memory, attention, and executive function, which significantly prolong recovery and diminish quality of life3. Accumulating evidence implicates neuroinflammation as a central driver in POCD pathogenesis, with activated microglia playing a pivotal role in initiating and sustaining central nervous system (CNS) immune responses4,5,6,7. Despite extensive research, the precise molecular pathways through which microglia contribute to synaptic dysfunction and cognitive decline remain incompletely elucidated8.
Recent progress in immunometabolism highlights that metabolic rewiring of immune cells is not merely permissive but instructive for inflammatory phenotypes2. Among key metabolic enzymes, ATP citrate lyase (ACLY) serves as a nexus between mitochondrial metabolism and epigenetic regulation by catalyzing the conversion of citrate into acetyl-CoA9,10,11. Acetyl-CoA serves as an essential substrate for histone acetylation and transcription factor activation, including NF-κB—a master regulator of inflammatory responses12,13. Although ACLY has been implicated in peripheral inflammatory diseases14, its role in microglial-driven neuroinflammation and cognitive impairment, particularly in the context of POCD, remains unexplored.
Beyond metabolic reprogramming, intercellular communication between glial cell types is increasingly recognized as a critical component of the neuroinflammatory landscape15,16. Extracellular vesicles (EVs), including exosomes, have emerged as potent mediators of glia–glia and glia–neuron signaling networks17. These vesicles transport diverse bioactive cargoes—proteins, RNAs, and lipids—that modulate phenotype and function in recipient cells18. However, the specific EV cargoes responsible for triggering astrocytic reactivity and synaptic dysregulation in POCD remain largely unidentified19.
In parallel, astrocytic glutamate transporters are pivotal for synaptic glutamate clearance; downregulation of excitatory amino acid transporter-1 (EAAT1) impairs glutamate homeostasis and promotes excitotoxicity20,21. Whether microglia-derived signals directly suppress EAAT1 in POCD has remained an open question.
Here, building upon the concept that microglial metabolism can shape intercellular signaling, we hypothesized that activated microglia secrete EVs loaded with ACLY that are internalized by astrocytes, where ACLY enhances NF-κB p65 acetylation, suppresses EAAT1 transcription, and precipitates glutamate dysregulation and synaptic failure22. Using POCD patient plasma-derived EVs, aged-mouse POCD models, and glial cell systems, we identify a microglia-to-astrocyte ACLY–p65–EAAT1 axis that links immunometabolism to excitotoxic injury and cognitive decline. These findings nominate ACLY and its EV-mediated transfer as actionable nodes for therapeutic intervention in POCD.
Results
Plasma-derived EVs from POCD patients disrupt hippocampal neuronal function in aged mice
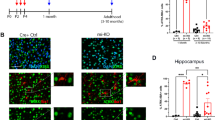
Plasma-derived extracellular vesicles from POCD patients (EVsPOCD) were successfully isolated and characterized, which was administrated to primary hippocampal neurons derived from neonatal mice and old mice (Fig. 1A). Transmission electron microscopy and nanoparticle tracking analysis confirmed the expected morphology and size distribution of EVsCon and EVsPOCD (Fig. 1B), while immunoblotting demonstrated enrichment of canonical EV markers (CD63, CD81) and absence of GM130 (Fig. 1C).

A Schematic of experimental design: EVs isolated from POCD and non-POCD patient(Control) plasma were applied to primary hippocampal neuron or injected into aged mice (five injections, every other day). Behavioral and electrophysiological assessments were conducted from day 11 onward. B, C Characterization of EVs by TEM, NTA, and immunoblotting. D Immunofluorescence analysis of hippocampal tissue showing elevated ACLY and Iba1 expression following EVsPOCD administration. n = 5 mice in each group, **p < 0.01, all values represent mean ± standard deviation. independent two-tailed Student’s t tests. E Co-localization of EAAT1 and p65 in EVs-treated mice. n = 5 mice in each group, *p < 0.05, **p < 0.01, all values represent mean ± standard deviation. independent two-tailed Student’s t tests. F Glutamate levels were assessed in hippocampal tissue after EVsPOCD treatment. *p < 0.05, n = 5 mice in each group/time point, all values represent mean ± standard deviation; two-way ANOVA with factors time and treatment with Newman–Keuls multiple comparison test. G, H Morris water maze performance showing impaired spatial learning and memory after EVsCon/EVsPOCD treatment. n = 15 mice in each group, *p < 0.05, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. I Fluoro-Jade B staining revealed increased degenerating neurons in EVsPOCD-treated hippocampi. n = 5 mice in each group, *p < 0.05, all values represent mean ± standard deviation. independent two-tailed Student’s t tests. J Electrophysiology showing attenuated fEPSPs and impaired LTP induction in EVsPOCD-treated slices. n = 10 mice in each group, **p < 0.01, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests.
Stereotactically administration of EVsPOCD into aged mice resulted in pronounced neuroinflammatory responses. Immunofluorescence revealed increased ACLY and IBA1 expression(Fig. 1D), as well as co-localization of EAAT1 and phosphorylated p65 in hippocampal regions and a concomitant reduction in EAAT1 immunoreactivity in hippocampal regions of EVsPOCD-treated mice, consistent with microglial activation and astrocytic dysfunction (Fig. 1E). In parallel, hippocampal glutamate concentrations were significantly elevated, showing a transient peak at 20–50 min post-stimulation in the EVsPOCD group compared with EVsCon (Fig. 1F).
Behavioral testing demonstrated marked cognitive impairments following EVsPOCD exposure. In the Morris water maze, EVsPOCD-treated mice exhibited longer escape latencies during training and reduced time spent in the target quadrant during the probe trial (Fig. 1G, H). Histological assessment using Fluoro-Jade B staining showed increased degenerating neurons in the hippocampus (Fig. 1I). Electrophysiological recordings revealed attenuated fEPSP slopes and impaired LTP induction in hippocampal slices from EVsPOCD-treated mice, further confirming synaptic dysfunction (Fig. 1J). Collectively, these results indicate that EVsPOCD trigger neuroinflammation, glutamate dysregulation, and synaptic plasticity impairment, leading to cognitive deficits in aged mice.
EVsPOCD impair dendritic spine morphology and neuronal connectivity
To further investigate the structural consequences of EVsPOCD exposure, we performed Golgi staining and scanning electron microscopy in hippocampal tissue of aged mice. Compared with EVsCon, EVsPOCD treatment led to a significant reduction in mushroom-type spines, total spine density, synapse numbers, and PSD thickness (Fig. 2A–D).

A Golgi staining analysis of hippocampal neurons from aged mice revealed reduced mushroom spines and spine density after EVsPOCD treatment. n = 30 dendritic segments analyzed from 5 mice per group. *p < 0.05, **p < 0.01, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. B–D TEM analysis of synapse number and PSD thickness after EVsPOCD treatment. n = 10 fields analyzed from 5 mice per group. *p < 0.05, **p < 0.01, all values represent mean ± standard deviation. independent two-tailed Student’s t tests. E, F Primary hippocampal neurons exposed to EVsPOCD exhibited decreased dendritic branching, reduced neurite length, and fewer intersections. n = 10 fields analyzed from 5 mice per group. **p < 0.01, ***p < 0.001, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests was used to analysis branches length and number of branches; two-way ANOVA with factors time and treatment, with Newman–Keuls multiple comparison test was used to analysis the number of intersections. G, H Immunofluorescence analysis demonstrated decreased PSD95 and Synapsin-1 expression in EVsPOCD-treated neurons. n = 5 mice per group. *p < 0.05, **p < 0.01, all values represent mean ± standard deviation. independent two-tailed Student’s t tests.
Consistent results were obtained in primary hippocampal neurons derived from neonatal mice. EVsPOCD exposure reduced dendritic branching, neurite length, and the number of Sholl intersections (Fig. 2E, F). Immunofluorescence further revealed decreased expression of PSD95 and Synapsin-1 in EVsPOCD-treated neurons (Fig. 2G, H). These findings suggest that EVsPOCD compromise both synaptic structure and functional connectivity in vivo and in vitro.
Pharmacological microglial depletion reverses ACLY-related neuroinflammation and cognitive dysfunction
To determine the causal role of microglial ACLY in POCD pathogenesis, we employed the CSF1R inhibitor PLX5622 to pharmacologically deplete microglia. PLX5622 treatment significantly attenuated the expression of both ACLY and IBA1 in hippocampal tissues (Fig. 3A), concomitantly reducing ac-p65 levels while restoring EAAT1 expression (Fig. 3B). These molecular changes were accompanied by improved neuronal integrity, as evidenced by preserved MAP2 staining (Fig. 3C), decreased the accumulation of extracellular glutamate (Fig. 3D), and enhanced cognitive performance in the Morris water maze test, with reduced escape latency and increased platform crossings. This restoration of synaptic function strongly correlates with and provides a mechanistic substrate for the improved spatial learning and memory observed in the Morris water maze (Fig. 3E, F). Furthermore, electrophysiological recordings demonstrated restored synaptic plasticity, characterized by increased fEPSP slopes and long-term potentiation (Fig. 3G–I), while dendritic spine analysis revealed increased total and mushroom spine densities (Fig. 3J–M). Collectively, these results implicate microglial ACLY as a key mediator of POCD-associated neurotoxicity and cognitive decline.

A Representative immunofluorescence images showing staining for ACLY (yellow), microglial marker IBA1 (red), and nuclei marker DAPI (blue) in 20-month-old POCD mice treated with or without PLX5622. Right panels: fluorescence intensity profiles for control and PLX5622-treated groups. Scale bar: 20 μm. n = 5 mice in each group, **p < 0.01, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. B Immunofluorescence staining depicting astrocytic marker GFAP (green), glutamate transporter EAAT1 (red), acetylated p65 (yellow), and nuclei marker DAPI (blue). Right panels: corresponding fluorescence intensity profiles comparing control and PLX5622-treated groups. Scale bar: 20 μm. n = 5 mice in each group, *p < 0.05, **p < 0.01, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. C Representative immunofluorescence images showing dendritic marker MAP2 (green), microglial marker IBA1 (red), and nuclei marker DAPI (blue). Intensity profiles are presented for both control and PLX5622-treated groups. Scale bar: 50 μm. n = 5 mice in each group, *p < 0.05, **p < 0.01, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. D Time course showing glutamate release (% baseline) in hippocampal slices from POCD mice treated with or without PLX5622. Arrows indicate stimulation. n = 5 mice in each group/time point, **p < 0.01, two-way ANOVA with Newman–Keuls multiple comparison test. E Representative swimming trajectories in Morris water maze tests for POCD mice treated with or without PLX5622. F Quantification of escape latency (left panel) and number of crossings over the platform location (right panel) in the Morris water maze test. n = 15 mice in each group, *p < 0.05, **p < 0.01; all values represent mean ± standard deviation. independent two-tailed Student’s t tests. G Representative field excitatory postsynaptic potentials (fEPSP) recorded from hippocampal slices (left inset indicates recording and stimulating electrode positions). Traces from control and PLX5622-treated POCD miceshown. “1” = baseline fEPSP trace before HFS (pre-LTP induction); “2” = fEPSP trace recorded at the late phase after HFS. H Quantification of normalized fEPSP slopes (left panel) and long-term potentiation (LTP) time course following high-frequency stimulation (HFS) (right panel). n = 10 mice in each group, **p < 0.01; all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. Golgi staining illustrating hippocampal dendritic spine morphology (J) and representative high-magnification images of dendritic spines (K). Scale bars are indicated. Quantitative analysis of spine density (L) and mushroom spine density (M) per 10 μm dendritic segment. n = 30 dendritic segments analyzed from 5 mice per group, **p < 0.01; all values represent mean ± standard deviation. Independent two-tailed Student’s t tests.
ACLY modulates astrocytic NF-κB acetylation and represses EAAT1 transcription
To elucidate the mechanistic link between ACLY activity and astrocytic dysfunction, we pharmacologically manipulated ACLY expression in C8-D1A astrocytes using pcDNA3.1(+)-Flag-ACLY for overexpression and SB (ACLY inhibitor), respectively. To establish a reliable model of microglial activation and the subsequent astrocytic response for mechanistic investigation, we utilized lipopolysaccharide (LPS) stimulation. To further and directly interrogate the dependency of EAAT1 suppression on NF-κB signaling, we employed the NF-κB inhibitor bortezomib (BZ). Activation of ACLY significantly increased the nuclear localization of acetylated NF-κB p65, whereas its inhibition abrogated p65 nuclear translocation (Fig. 4A). In silico analysis of the SLC1A3 promoter region revealed canonical NF-κB binding motifs (Fig. 4B), and using an antibody specific for acetyl p65 Lys310 (CST #12629), ChIP qPCR showed enriched occupancy at two NF-κB motifs within the SLC1A3 promoter (Fig. 4C). Furthermore, knockdown of p65 led to a significant increase in SLC1A3 mRNA levels (Fig. 4D), supporting a repressive outcome of Lys310 acetylated p65 at this locus, demonstrating that ACLY-mediated p65 acetylation directly represses EAAT1 transcription.

A Western blot analysis of ACLY, p65, ac-p65, and EAAT1 protein levels in nuclear (left part) and cytoplasmic fractions (right part) of C8-D1A cells treated with pcDNA3.1(+)-Flag-ACLY (ACLY) or ACLY inhibitor (SB) and stimulated with LPS or NF-κB inhibitor (BZ). Histone H3 and GAPDH served as nuclear and cytoplasmic loading controls, respectively. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, n = 5 independent biological replicates (independent cell cultures); all values represent mean ± standard deviation, one-way ANOVA with Newman–Keuls multiple comparison test. B Bioinformatics analysis identifying putative NF-κB p65 binding motifs within the SLC1A3 promoter region. Motifs 1 and 2 indicate potential binding sites relative to the transcription start site (TSS). C ChIP assay results indicating enhanced binding of acetylated p65 (ac-p65) to motifs 1 and 2 in the SLC1A3 promoter region compared to the negative control (NC). IgA/G serves as a control antibody. n = 4 independent biological replicates, *p < 0.05, values represent mean ± standard deviation. independent two-tailed Student’s t tests. D Quantitative PCR analysis demonstrating increased SLC1A3 mRNA expression in C8-D1A cells transfected with shp65/NF-κB compared to control (shNC). n = 4 independent biological replicates, *p < 0.05, values represent mean ± standard deviation. Independent two-tailed Student’s t tests.
Activated microglia impair astrocyte function through EV-mediated transfer
In order to examine the impact of microglial-derived EVs on astrocytic inflammation and apoptosis, we utilized a transwell co-culture system. C8-D1A astrocytes were cultured co-cultured with BV2/LPS-activated BV2 microglia (BV2M1)/ BV2 + GW4869(an inhibitor of extracellular vesicles, 10 mM, # D1692-5MG, Sigma, USA) for 48 h (Fig. 5A). As shown in Fig. 5B, the ACLY expression in BV2M1 groups was significantly upregulated, and this upregulation was decreased by GW4869. Flow cytometry analysis revealed no significant differences in early or late apoptosis (annexin V/PI) between these three groups, suggesting that the co-culture did not induce a substantial increase in apoptosis (Fig. 5C).

A Schematic illustration of experimental design for co-culture experiments. Astrocytic C8-D1A cells were co-cultured with BV2/BV2M1(activated with LPS, 2 μg/ml for 48 h)/BV2M1 + GW4869 for 48 h. The downstream analyses included Western blot, annexin V/PI assay and immunofluorescence staining for p65 and EAAT1. Created in BioRender. Ding, L. (2026) https://BioRender.com/g1hk8n9. B Western blot analysis comparing ACLY protein level in C8-D1A after co-cultured with BV2, BV2M1, or BV2M1 + GW4869. **p < 0.01, n = 5 independent biological replicates; all values represent mean ± standard deviation, one-way ANOVA with Newman–Keuls multiple comparison test. C Flow cytometry analysis (annexin V/PI) assessing cell viability and apoptosis in C8-D1A cells after co-cultured with BV2, BV2M1 or BV2M1 + GW4869. D Immunofluorescence staining of C8-D1A cells for NF-κB p65 (green), glutamate transporter EAAT1 (red), and nuclei (DAPI, blue) in different groups. Scale bar: 50 μm. E Fluorescence intensity profiles for p65, EAAT1 and nuclear-to-cytoplasmic ratio of p65 in C8-D1A co-cultured with BV2, BV2M1, or BV2M1 + GW4869. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.001, n = 5; all values represent mean ± standard deviation, one-way ANOVA with Newman–Keuls multiple comparison test.
Immunofluorescence staining further demonstrated that co-culture with BV2M1 induced nuclear translocation of NF-κB p65 (green), a hallmark of microglial-driven inflammation, and repression of the glutamate transporter EAAT1 (red) in astrocytes (Fig. 5D, F). However, treatment with the EV secretion inhibitor GW4869 effectively reversed these changes, suggesting that EVs from BV2M1 mediated the observed effects on astrocytic inflammation and EAAT1 expression (Fig. 5D, G). Intensity profiles across the immunofluorescence images confirmed these observations, with increased p65 and decreased EAAT1 fluorescence intensity in the co-cultured group, and partial restoration in the GW4869-treated group (Fig. 5E). Moreover, the p65 nuclear transferred rate was promoted by BV2M1, which was reversed by GW4869.
These data indicate that while co-culturing with LPS-activated microglia induces inflammatory changes in astrocytes, the impact on apoptosis is minimal. Moreover, our results indicate that EVs from activated microglia contribute to inflammatory responses in astrocytes, which are associated with p65 activation and concomitant downregulation of EAAT1.
EVs from activated microglia contain ACLY and induce astrocytic inflammation
To further investigate the mechanism of microglia-to-astrocyte signaling, we isolated EVs from control (EVsCon) and LPS-activated (EVsM1) microglial cultures. Transmission electron microscopy and nanoparticle tracking analysis confirmed typical EV morphology and size distribution (Fig. 6A–D). Western blot analysis revealed that EVsM1 were enriched in EV markers (CD63, CD81) and notably contained higher levels of ACLY, while lacking the Golgi marker GM130, confirming purity (Fig. 6C). Upon application to astrocytes and mice model, PKH67-labeled EVsM1 and Dir-labeled EVsM1 were efficiently internalized both in vitro and in vivo systems, respectively(Fig. 6F), resulting in elevated nuclear acetylated p65 and pronounced downregulation of EAAT1 expression (Fig. 6G–I). These findings demonstrate that EVs derived from activated microglia serve as carriers of ACLY, capable of reprogramming astrocytic inflammatory signaling and glutamate transporter expression.

A Schematic illustration depicting the experimental approach used to isolate and characterize EVs from LPS-stimulated BV2 microglial cells (EVsM1). EVs were analyzed using transmission electron microscopy (TEM), nanoparticle tracking analysis (NTA), and Western blot, followed by uptake assays and immunofluorescence staining for ac-p65 and EAAT1. B Representative TEM images showing morphology of EVs from control BV2 (EVsCon) and activated BV2 (EVsM1). Scale bar: 100 nm. C Western blot characterization of EV markers (CD63, CD81), GM130 (negative marker), and ACLY protein in lysates and isolated EVs. D NTA particle size distribution profiles for EVs isolated from control (EVsCon) and activated microglia (EVsM1). E Uptake analysis of PKH67-labeled EVs (green) by C8-D1A cells, co-stained with nuclei marker DAPI (blue). Scale bar, 50 μm. F PBS/EVsCon/ EVsM1 were labeled with the lipophilic fluorescent tracer Dir, and then were stereotactically injected into C57BL/6. After 24 h, the EVs distribution in vivo was visualized by an IVIS imaging system. G Immunofluorescence staining showing nuclear localization of acetylated p65 (ac-p65, green), EAAT1 expression (red), and nuclei (DAPI, blue) in C8-D1A cells treated with EVsCon or EVsM1. Scale bar, 50 μm. H, I Fluorescence intensity profiles of EAAT1 and ac-p65 staining in C8-D1A cells treated with EVsCon and EVsM1. n = 5, *p < 0.05, **p < 0.01, Values represent mean ± standard deviation. independent two-tailed Student’s t tests.
EVsM1 exacerbate neuroinflammation, synaptic loss, and cognitive deficits in POCD mice
To assess the functional consequences of EV-mediated ACLY transfer in vivo, we administered intra-hippocampal injections of EVsM1 into POCD mice. Treated animals exhibited markedly elevated expression of ACLY and IBA1 in the hippocampus (Fig. 7A), and immunofluorescence revealed that EVsM1 treatment increased the number of GFAP-positive astrocytes with intense nuclear ac-p65 staining, which was associated with a pronounced reduction in EAAT1 in the astrocytes (Fig. 7B). Neuronal degeneration was evidenced by diminished MAP2 immunoreactivity and increased Fluoro-Jade B-positive cells (Fig. 7C, D). Behavioral testing via the Morris water maze revealed significant impairments in spatial learning and memory (Fig. 7E). Structural analyses further demonstrated a substantial loss of dendritic spines (Fig. 7F, G) and synaptic degeneration as confirmed by transmission electron microscopy (Fig. 7H, I). These results underscore the pathogenic role of EVsM1 in amplifying neuroinflammation and precipitating structural and cognitive deficits characteristic of POCD.

A Immunofluorescence staining of ACLY (yellow), microglial marker IBA1 (red), and nuclei (DAPI, blue) in brain tissues from POCD mice treated with EVsCon or EVsM1(5 µg in 2 μl PBS). Fluorescence intensity profiles presented for each group. Scale bar:20 μm. n = 5 mice per group. *p < 0.05, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. B Immunofluorescence images of astrocytic marker GFAP (green), glutamate transporter EAAT1 (red), acetylated p65 (yellow), and nuclei (DAPI, blue). Corresponding fluorescence intensity profiles are shown for each condition. Scale bar: 20 μm. n = 5 mice per group. *p < 0.05, **p < 0.01, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. C Immunofluorescence staining of dendritic marker MAP2 (green), microglial marker IBA1 (red), and nuclei (DAPI, blue). Intensity profiles indicate distribution patterns. Scale bar: 50 μm. n = 5 mice per group. *p < 0.05, ***p < 0.001, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. D Fluoro-Jade B (FJB) staining of hippocampal regions to identify neuronal damage in POCD mice treated with EVsCon or EVsM1. Quantification of FJB-positive cells shown on the right. Scale bar: 100 μm. n = 5mice per group. **p < 0.01, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. E Morris water maze test assessing cognitive function in POCD mice treated with EVsCon or EVsM1. Representative swim paths, escape latency, and platform crossing times presented. n = 15mice in each group, *p < 0.05, all values represent mean ± standard deviation. independent two-tailed Student’s t tests. Golgi staining images illustrating hippocampal dendritic spine density (F) and high-magnification views of dendritic spines (G). Quantitative analysis of spine number and mushroom spine number per 10 μm dendritic segment shown. Scale bar indicated. n = 30 dendritic segments analyzed from 5 mice per group. *p < 0.05, ***p < 0.0001, all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. Transmission electron microscopy (TEM) images of synaptic ultrastructure, highlighting synaptic morphology and postsynaptic density (PSD) thickness (H), and quantification of synapse number per field (I). Scale bars: 500 nm. n = 5 mice in each group, *p < 0.05, all values represent mean ± standard deviation. independent two-tailed Student’s t tests.
Knockout of ACLY in EVs reverses astrocytic and cognitive pathology
To directly establish the pathogenic role of EV-associated ACLY, we generated ACLY-deficient EVs (EVsM1-ACLY KO) from LPS-stimulated microglia via gene silencing. Morphological and particle size analyses confirmed that EVsM1-ACLY KO retained typical vesicle characteristics (Fig. 8C). Compared to control EVs, treatment with EVsM1-ACLY KO led to reduced astrocytic ac-p65 accumulation and restored EAAT1 expression in vivo (Fig. 8D). Neuronal integrity was preserved, with significantly fewer FJB-positive degenerating cells (Fig. 8E), and cognitive performance markedly improved, as evidenced by Morris water maze metrics (Fig. 8F). Moreover, glutamate release profiles normalized (Fig. 8G), and electrophysiological recordings showed restored synaptic plasticity, including enhanced LTP and fEPSP slopes (Fig. 8H, I). These data confirm that ACLY is a critical cargo within microglial EVs responsible for mediating astrocytic dysfunction and cognitive decline in POCD.

A Western blot analysis verifying ACLY knockout in EVs derived from microglia. EV markers CD63 and CD81, and negative marker GM130 are included. B Representative TEM images illustrating the morphology of EVsM1-ACLY siRNA NC and EVsM1-ACLY KO. Scale bar: 100 nm. C Nanoparticle tracking analysis showing particle size distribution profiles of EVsM1-ACLY siRNA NC and EVsM1-ACLY KO. D Immunofluorescence staining for GFAP (astrocyte marker, green), EAAT1 (red), acetylated p65 (yellow), and DAPI (nuclei, blue) in brain tissues from POCD mice treated with EVsM1-ACLY siRNA NC or EVsM1-ACLY KO(5 µg in 2 μl PBS). Corresponding intensity profiles are presented. Scale bar: 20 μm. n = 5 mice per group. *p < 0.05, **p < 0.01, all values represent mean ± standard deviation. independent two-tailed Student’s t tests. E Fluoro-Jade B (FJB) staining assessing neuronal damage in the hippocampus of POCD mice receiving EVsM1-ACLY siRNA NC or EVsM1-ACLY KO treatment. Quantitative analysis presented. Scale bar, indicated. **p < 0.05, n = 5 mice per group; all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. F Morris water maze test depicting cognitive performance through representative swim paths, number of crossings over the previous platform location, and escape latency in POCD mice treated with EVsM1-ACLY siRNA NC or EVsM1-ACLY KO. *p < 0.05, n = 15 mice in each group; all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. G Time course of glutamate release (% baseline) measured in hippocampal slices following stimulation in POCD mice treated with EVsM1-ACLY siRNA NC or EVsM1-ACLY KO. *p < 0.05, **p < 0.01, n = 5 mice in each group/time point, all values represent mean ± standard deviation; two-way ANOVA with Newman–Keuls multiple comparison test. H, I Representative field excitatory postsynaptic potentials (fEPSP) recorded from hippocampal slices and quantification of normalized fEPSP slopes in POCD mice treated with EVsM1-ACLY siRNA NC or EVsM1-ACLY KO. **p < 0.01, n = 10; all values represent mean ± standard deviation. Independent two-tailed Student’s t tests. “1” = baseline fEPSP trace before HFS (pre-LTP induction); “2” = fEPSP trace recorded at the late phase after HFS.
EV-induced synaptic dysfunction progresses in a time-dependent manner
To characterize the temporal progression of EV-mediated neuropathology, we administered EVsM1 stereotactically into POCD mice at defined postoperative intervals (Fig. 9A). Field potential recordings revealed a progressive decline in fEPSP slopes with repeated EVsM1 injections, indicating cumulative synaptic dysfunction (Fig. 9B–D). Consistently, extracellular glutamate levels increased in a time-dependent manner (Fig. 9E), suggesting impaired glutamate clearance as an evolving pathological feature. Behavioral assessments mirrored these changes, with worsening spatial memory deficits as measured by platform crossing frequency and escape latency in the Morris water maze (Fig. 9F–H). These data highlight the dynamic and cumulative nature of EV-induced neurotoxicity and underscore the importance of early intervention to mitigate POCD-related synaptic and cognitive decline.

A Schematic timeline depicting experimental design, including stereotaxic injection of EVsCon or EVsM1 (5 µg in 2 μl PBS) every 2 days after POCD establishment, followed by assessment of LTP induction, extrasynaptic glutamate determination, and cognitive testing via Morris water maze. B, C Field excitatory postsynaptic potential (fEPSP) slope recordings and representative fEPSP traces from hippocampal slices of control and POCD mice injected with EVsCon or EVsM1 at days 2, 4, and 6 post-injection. Arrow indicates high-frequency stimulation (HFS). “1” = baseline fEPSP trace before HFS (pre-LTP induction); “2” = fEPSP trace recorded at the late phase after HFS. D Quantitative analysis of normalized fEPSP slopes among different experimental groups. ***p < 0.001, ****p < 0.0001, n = 10 mice in each group; all values represent mean ± standard deviation, one-way ANOVA with Newman–Keuls multiple comparison test. E Normalized glutamate release profiles from hippocampal slices of control and POCD mice treated with EVsCon or EVsM1 at the indicated time points. n = 5 mice in each group/time point, *p < 0.05,**p < 0.01, all values represent mean ± standard deviation, two-way ANOVA with Newman–Keuls multiple comparison test. F Representative swim trajectories of control and POCD mice treated with EVsCon or EVsM1 on days 2, 4, and 6 in Morris water maze tests. Quantification of the number of crossings over the previous platform location (G) and escape latencies (H) in Morris water maze tests among different groups. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. n = 15 mice in each group; all values represent mean ± standard deviation, one-way ANOVA with Newman-Keuls multiple comparison test.
Discussion
This study delineates a previously unrecognized metabolic-inflammatory communication axis in which microglial EVs deliver ACLY to astrocytes, augmenting NF-κB p65 acetylation, repressing EAAT1, and driving glutamate accumulation, synaptic dysfunction, and cognitive impairment in POCD. By integrating patient-derived EVs, aged-mouse behavior and electrophysiology, and targeted gain-/loss-of-function approaches, we provide multi-scale evidence that microglial ACLY is not only upregulated during POCD but is also selectively packaged into EVs to enact pathogenic reprogramming in recipient astrocytes.
A central finding of our work is the selective upregulation of ACLY in microglia during POCD. While prior studies have focused predominantly on cytokine signaling and surface markers to define microglial activation23,24,25, our data highlight metabolic reprogramming as a core feature of neuroinflammation. ACLY, which converts citrate into acetyl-CoA, serves as a critical link between cellular metabolism and epigenetic regulation via histone and transcription factor acetylation26,27. This aligns with recent work suggesting that immune cell metabolism not only supports energetic demands but also actively dictates inflammatory phenotypes through metabolite-derived signaling pathways28. Here, we extend this paradigm by demonstrating that microglial ACLY is not merely a marker of activation, but a functional driver of downstream neurotoxicity.
One of the most novel aspects of our study is the discovery that ACLY-driven acetylation of NF-κB p65 in astrocytes directly represses transcription of the glutamate transporter EAAT1. This mechanism establishes a previously unappreciated link between metabolic enzymes and excitatory neurotransmitter regulation. Although NF-κB signaling is well-established in neuroinflammation29, the role of acetylation in modulating its transcriptional targets in astrocytes has been largely overlooked. Our ChIP-qPCR data and p65 knockdown experiments confirm that acetylated p65 binds to the SLC1A3 promoter and suppresses EAAT1 expression. This transcriptional repression disrupts glutamate clearance, leading to accumulation of extra synaptic glutamate, synaptic degeneration, and memory impairment—hallmarks of POCD. These findings expand the functional repertoire of NF-κB acetylation and underscore its potential as a therapeutic target in glial dysfunction. While our study establishes acetylated NF-κB p65 as a key transcriptional regulator of EAAT1 in this context, it is plausible that other acetylation-dependent transcription factors implicated in astrocyte reactivity—such as STAT3, histone, or ac4C—could also be influenced by ACLY-derived acetyl-CoA and contribute to the overall reprogramming phenotype30,31,32. Future acetylome profiling of astrocytes upon EV-ACLY uptake will help delineate the full spectrum of targets and their hierarchical importance.
Another important innovation lies in the identification of EVs as functional carriers of ACLY between glial cell types. Previous studies have established that EVs can deliver cytokines, miRNAs, or proteins in neurodegenerative disease models33. However, to our knowledge, this is the first demonstration that EVs can transfer a metabolic enzyme capable of altering transcriptional regulation in recipient astrocytes. The uptake of ACLY-containing EVs by astrocytes triggered a cell-extrinsic epigenetic response that reprogrammed inflammatory gene expression and disrupted glutamate homeostasis. This EV-mediated mechanism represents a paradigm shift in our understanding of glial crosstalk—suggesting that microglial metabolism can exert long-range, vesicle-dependent effects on neighboring astrocytes and neurons.
In this study, we used ultracentrifugation to isolate extracellular vesicles from plasma. However, different methods for isolating EVs yield preparations that vary markedly in composition and thereby influence in vitro and in vivo functional outcomes34,35. In vitro, isolation by ultracentrifugation often gives high particle yields but brings along contaminating proteins or lipoproteins that may enhance cell-uptake, signaling or proliferation assays in an artifactual way. In contrast, size-exclusion chromatography (SEC) or density-gradient approaches produce purer vesicle populations, reducing confounding background activity and yielding more specific functional responses36,37. Immunoaffinity capture enriches for specific subpopulations, improving receptor-targeted assays but narrowing the breadth of vesicle phenotypes. In vivo, the isolation method affects biodistribution, clearance and organ-targeting: contaminant proteins or lipoproteins can change immune-recognition or clearance kinetics, while subpopulation selection skews delivery to particular tissues38. Recent guideline updates (MISEV2023) emphasize that EV functional data must be interpreted in light of isolation workflow, purity metrics and contaminant controls39.
Targeting the ACLY or EV biogenesis pathways presents novel therapeutic strategies for POCD. Pharmacological inhibitors of ACLY, such as SB-204990 used in this study and the clinically approved agent bempedoic acid, have demonstrated anti-inflammatory potential in preclinical models. However, their application in neurological contexts depends on blood-brain barrier penetration, the optimal therapeutic window, and potential off-target effects of systemic metabolic inhibition. Similarly, inhibiting EV release (GW4869) has been shown to mitigate neuropathology in animal models40. For POCD, the short-term perioperative use of such inhibitors to block the propagation of neuroinflammatory signaling is theoretically feasible but requires careful evaluation of its impact on systemic, EV-mediated physiological communication.
The ACLY–p65–EAAT1 axis elucidated here likely does not operate in isolation but may engage in complex crosstalk with other established pathological pathways in POCD. On one hand, NF-κB activation can prime the NLRP3 inflammasome by upregulating its components. On the other hand, ACLY-driven metabolic rewiring may impact mitochondrial function, potentially leading to reactive oxygen species (ROS) accumulation. ROS can both activate NF-κB and serve as a potent activator of the NLRP3 inflammasome41. This interplay could create a positive feedback loop, exacerbating neuroinflammation and excitotoxicity.
Strengths of this work include the convergence of human plasma EVs with aged-mouse and cellular models, quantitative electrophysiology, and structure-function readouts of synaptic integrity. Limitations should be acknowledged. First, while we identify astrocytes as primary recipients, EV biodistribution and possible neuronal or oligodendroglial uptake warrant deeper profiling. Second, although we used pharmacologic and genetic tools to manipulate ACLY, future studies should define the EV-packaging code for ACLY and map acetylome-wide consequences in astrocytes. Third, translation will require careful consideration of CNS penetration, dosing paradigms for ACLY/EV-pathway inhibitors, and perioperative safety in older patients. Finally, while our data strongly implicate microglia as a key source of pathogenic EVs, the heterogeneous nature of plasma EVs means they originate from multiple cell types. The inability to definitively assign the proportion of ACLY+ EVs in patient plasma that are of microglial origin. Future studies employing cell-type-specific EV markers, such as leveraging microglia-specific surface proteins (e.g., Tmem119) for immunocapture, could directly isolate and profile microglial EVs from patient blood, providing even more direct evidence.
Methods
Patient plasma collection and POCD mice model
A sample size estimate was conducted using an online sample size calculator (https://sample287size.net/). Peripheral blood was collected from POCD patients and matched non-POCD controls (Table S1) after written informed consent, under protocols approved by the institutional ethics committee. Twenty-month-old male C57BL/6 mice (20–25 g) were obtained from the Animal Center of Zhengzhou University, Zhengzhou, China, and maintained under a 12-h light/dark cycle with free access to food and water, room temperature (22 ± 1 °C) and 50 ± 10% humidity. The study protocol was approved by the Ethics Committee of the First Affiliated Hospital of Zhengzhou University (2021-KY-0771-002). Twenty-month-old male C57BL/6 mice were randomly(conducted with the random number table approach) divided into different groups (assigned a permanent numerical designation in the cages), and POCD was induced via laparotomy under isoflurane anesthesia as previously described2, all procedures were performed in accordance with the Guidelines for the Care and Use of Laboratory Animals by the National Institutes of Health (Bethesda, MD, USA).
Immunofluorescence staining
Brain tissues or cultured cells were fixed in 4% paraformaldehyde (Servicebio, Cat# G1101, China) at room temperature for 15 min and washed three times with phosphate-buffered saline (PBS). Tissues were permeabilized with 0.3% Triton X-100 (Sigma-Aldrich, Cat# X100, USA) in PBS for 10 min and blocked with 5% bovine serum albumin (BSA; Sigma-Aldrich, Cat# A7906, USA) for 1 h at room temperature. Samples were incubated overnight at 4 °C with primary antibodies: ACLY (1:200; Abcam, Cat# ab40793), IBA1 (1:300; Abcam, Cat# ab178846), GFAP 1:300; Abcam, Cat# ab7260), EAAT1 (1:200; Abcam, Cat# ab416), NF-kappaB p65 (1:200; Cell Signaling Technology, Cat# 8242), ac-p65 (1:200; Cell Signaling Technology, Cat# 3045), MAP2 (1:400; Abcam, Cat# ab32454). After washing with PBS three times, samples were incubated for 1 h at room temperature with Alexa Fluor-conjugated secondary antibodies (1:500; Cell Signaling Technology, Cat#4412, #8889, #4410, USA). Nuclei were stained with DAPI (Sigma-Aldrich, Cat# D9542, 1:1000 USA). Images were observed using a fluorescence microscope. The semi-quantitative fluorescence intensity was analyzed by ImageJ software (National Institutes of Health, Bethesda, USA). For quantification of fluorescence intensity, images from at least 5 biological replicates (mice) were analyzed. Using ImageJ, regions of interest were drawn around specific hippocampal regions (for tissue sections). The integrated density was measured for each channel. The nuclear translocation rate was quantified by calculating the nuclear-to-cytoplasmic (N/C) ratio of p65 (or ac-p65) fluorescence intensity. Briefly, regions of interest (ROIs) were defined for the nucleus (based on DAPI staining) and the surrounding cytoplasm using ImageJ. The mean fluorescence intensity (MFI) for p65/ac-p65 was measured in each ROI, and the N/C ratio was calculated for individual cells from multiple images across biological replicates.
Western blot analysis
Cells and tissues were lysed in RIPA buffer containing protease inhibitors (Beyotime, Cat# P0013B, China). Protein concentrations were quantified using the BCA assay (Thermo Fisher Scientific, Cat# 23227, USA). Equal amounts of proteins (30 μg per lane) were separated by SDS-PAGE and transferred onto PVDF membranes (Millipore, Cat# IPVH00010, USA). Membranes were blocked with 5% non-fat milk in TBST and incubated overnight at 4 °C with primary antibodies against ACLY, IBA1, CD63, CD81, GM130, GAPDH, Histone H3 (Cell Signaling Technology, Cat# 4499). The primary antibody for acetylated p65 (ac-p65) (Cell Signaling Technology, Cat# 3045) detects p65 acetylated at lysine 310. HRP-conjugated secondary antibodies (Cell Signaling Technology, Cat# 7074, 7076; 1:5000) were used for detection. Chemiluminescent signals were visualized using an ECL detection system (GE Healthcare, Cat# RPN2232) and analyzed with ImageJ software.
Extracellular vesicle isolation and characterization
BV2 cells were cultured in F12 DMEM (# PM150312, Procell, China) supplemented with 10% EV-depleted FBS (prepared by ultracentrifugation at 120,000 × g for 16 h) and 1% penicillin-streptomycin (#PB180120, Procell, China). EVs were isolated from patient plasma and from BV2 microglial culture media using ultracentrifugation (Beckman Coulter Optima XPN-100 Ultracentrifuge), following our established protocols42. Plasma/cell culture supernatants were centrifuged at 300 × g for 10 min, followed by 2000 × g for 20 min, and 10,000 × g for 30 min at 4 °C. Subsequently, supernatants were ultracentrifuged at 100,000 × g for 70 min. Pellets were resuspended in PBS, passed through a 0.22 µm filter and ultracentrifuged 100,000 × g for 70 min again. Finally, the resulting pellet was suspended in 200 μl of PBS and then stored at −80 °C.
Transmission electron microscopy (TEM)
Freshly isolated EVs were resuspended in cold phosphate-buffered saline (PBS) for TEM analysis. For negative staining, 20 µL of the EV suspension was loaded onto carbon-coated copper grids and allowed to adsorb for 5 min at room temperature. The grids were then stained with 2% phosphotungstic acid (Servicebio, China) for 1 min and blotted dry with filter paper. EV morphology was examined using a Hitachi HT7800 transmission electron microscope (Hitachi, Japan) at an acceleration voltage of 100 kV and a magnification of 100,000 times magnification.
For brain samples, the samples were fixed, dehydrated, and embedded in epoxy resin. Ultrathin sections (70 nm) were stained with uranyl acetate and lead citrate and imaged with transmission electron microscope.
Nanoparticle tracking analysis (NTA)
EVs particle size distribution and concentration were determined using nanoparticle tracking analysis (NTA) on a ZetaView system (Particle Metrix, Germany). Prior to measurements, the instrument was calibrated with 100 nm polystyrene beads (Sigma-Aldrich, Germany). EV samples were diluted 1:100 in PBS and analyzed at 25 °C. For each sample, five independent measurements (30 s each) were recorded.
EVs labeling and uptake identification using PKH26
PKH26-labeled EVs were prepared according to the manufacturer’s protocol (Sigma-Aldrich, St. Louis, USA). Fresh EVs samples (100–200 µL) were first mixed with 500 µL Diluent C and 6 µL PKH67, vortexed for 30 s, and incubated for 5 min in the dark at room temperature. To stop the staining reaction, 2 mL of 10% BSA in PBS was added. The mixture was then ultracentrifuged at 120,000 × g for 70 min at 4 °C to remove excess dye and isolate PKH67-labeled EVs, which were subsequently stored at −80 °C. The uptake test was performed as our previous publication2.
In vivo EVs distribution
To verify the distribution of EV particles in the mice, PBS, EVsCon and EVsM1 were labeled with the lipophilic fluorescent tracer Dir (#UR21017, Umibio, Shanghai, China) as previously described43. Then the labeled EVs (5 μg in 2 μl PBS/mice) were stereotactically injected into mice. As a control, 5 mice were injected with the same volume of labeled PBS. After 24 h, an IVIS imaging system (PerkinElmer; USA) was used for tracking EVs distribution in vivo.
Primary hippocampal neuron culture and EV treatment
Primary hippocampal neurons were isolated from neonatal C57BL/6 mice (postnatal day 1) using a papain-based dissociation system (Worthington Biochemical, LK003150). Cells were plated on poly-D-lysine-coated coverslips in Neurobasal-A medium supplemented with B27 (Gibco, 17504-044), GlutaMAX (Gibco, 35050-061), and penicillin-streptomycin. After 4 DIV, neurons were exposed to EVsPOCD(20 µg/ml) or EVsCon (20 µg/ml) for 48 h under serum-free conditions.
Aged male C57BL/6 mice received stereotactically injections of EVsPOCD or EVsCon(5 µg EV protein per injection) once every 2 days, for a total of 5 injections. Behavioral testing commenced on day 11 after the first injection, followed by sacrifice for histological and biochemical analyses.
Microglial depletion with PLX5622
To deplete microglia, 20-month-old male C57BL/6 mice were fed ad libitum with a chow diet containing 1200 ppm PLX5622 (#HY-114153, MedChemExpress, USA;) for 7 consecutive days. Control mice received an identical diet without PLX5622. Microglial depletion efficiency was confirmed via Immunofluorescence in a subset of mice at the endpoint.
ChIP assay
Chromatin immunoprecipitation (ChIP) assays were performed using a ChIP assay kit (Millipore, Cat# 17–295). Cells were cross-linked with 1% formaldehyde, sonicated, and immunoprecipitated overnight with anti-acetylated p65 antibody (Cell Signaling Technology, Cat# 8242). Immunoprecipitated DNA fragments were purified and analyzed by quantitative PCR using specific primers for SLC1A3 promoter regions.
Quantitative PCR analysis
Total RNA was extracted using TRIzol reagent (Thermo Fisher Scientific, Cat# 15596026) and reverse transcribed using a PrimeScript RT reagent kit (Takara, Cat# RR037A). Quantitative PCR was performed using SYBR Green Master Mix (Roche, Cat# 04913914001) on a Roche LightCycler 480 System. Primers specific for SLC1A3(5′-ACAGCGTCTAGACCGGTTAGCGCTAGCTCATTAC-3′ and 5′-CGACAGTTAGCCAGAGAGCTCGCGGCCGCCGCTGT-3′) and GAPDH (5′-AAC TTTGGCATTGTGGAAGG-3′ and 5′-ACACATTGGGGGTAGGAACA-3′) were synthesized by Sangon Biotech. Gene expression was normalized to GAPDH and analyzed by the 2−ΔΔCT method.
p65 knockdown in C8-D1A
To specifically inhibit NF-κB p65 expression in C8-D1A astrocytes, short hairpin RNA (shRNA)-mediated knockdown was performed. Cells were seeded in 6-well plates at a density of 2 × 105 cells per well and cultured until 60–70% confluence. Transfection complexes were prepared by mixing 2 µg of either p65-targeting shRNA(Genechem, Shanghai, China) or non-targeting control shRNA with 5 µL of Lipofectamine 3000 reagent (#L3000015, Invitrogen, USA) in 250 µL Opti-MEM medium. After 20 min incubation at room temperature, the complexes were added dropwise to the cells. The medium was replaced with complete DMEM 6 h post-transfection, and cells were cultured for an additional 48 h to allow efficient knockdown. Silencing efficiency was assessed by quantitative RT-PCR for p65 mRNA and by Western blot analysis of p65 protein levels.
BV2 microglial cell culture and inflammatory model establishment
BV2 microglial cells (#CL-0493, Procell, China) were maintained in Dulbecco’s Modified Eagle Medium (DMEM) containing 10% fetal bovine serum and 1% penicillin-streptomycin (Gibco, ThermoFisher, USA). To establish an inflammatory model, cells were activated by continuous stimulation with 2 µg/ml lipopolysaccharide (LPS, #L4391, Sigma-Aldrich, USA) for 48 h.
Inhibition of microglial EV secretion in BV2 cells using GW4869
To inhibit EVs secretion by BV2 microglial cells, BV2 cells were cultured in DMEM supplemented with 10% EV-depleted FBS and 1% penicillin-streptomycin. At 80% confluency, cells were treated with 10 mM GW4869 (an inhibitor of extracellular vesicles, 10 mM, # D1692-5MG, Sigma, USA) for 48 h.
ACLY overexpression and inhibition
ACLY overexpression was achieved by seeding C8-D1A cells at a density of 2 × 105 cells per well in a 6-well plate, cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin. After 24 h, when the cells reached 60–70% confluence, a transfection mixture was prepared using 5 μL of lipofectamine RNAiMAX transfection reagent (#11668019, Invitrogen, USA) and 2 μg of pcDNA3.1(+)-Flag-ACLY plasmid(Bio-Research Innovation Center, Suzhou, China) in 500 μL of Opti-MEM(#51985091, Gibco, USA). This mixture was incubated at room temperature for 20 min to form DNA-lipid complexes, then added to the cells and incubated at 37 °C in a 5% CO2 incubator for 6 h. The medium was subsequently replaced with fresh DMEM, and the cells were cultured for an additional 48 h to ensure adequate ACLY expression. Following overexpression, the cells were treated with 20 μM SB-204990 (#SML2829-5MG, Sigma, USA), an ACLY inhibitor, prepared from a 10 mM stock solution in dimethyl sulfoxide (DMSO) and diluted in DMEM to maintain a final DMSO concentration below 0.1%. The cells were incubated with SB-204990 at 37 °C in a 5% CO2 incubator for 24 h. This combined approach of overexpression and inhibition allows for the evaluation of ACLY’s role in cellular processes, with the outcomes assessed through Western blot analysis.
NF-κB inhibition
To block the NF-κB signaling pathway, C8-D1A cells were treated with bortezomib (BZ, #HY-100227, MedChem Express, USA), an NF-κB inhibitor. A 10 mM stock solution of BZ was prepared in DMSO and diluted to a working concentration of 10 μM in complete DMEM. When the cells in 6-well plates reached 70–80% confluence, the medium was replaced with the BZ-containing DMEM, and the cells were incubated at 37 °C in a 5% CO2 incubator for 24 h.
ACLY knockdown
ACLY-specific siRNA and control siRNA were obtained from Promega (Shanghai, China). A total of 2 × 105 LPS-pretreated BV2 cells were transfected with 10 µM ACLY siRNA or control siRNA using lipofectamine RNAiMAX transfection reagent (#11668019, Invitrogen, USA). Silencing efficiency was assessed 24 h post-transfection by qRT-PCR with ACLY-specific primers: forward 5′-TGCTCGATTATGCACTGGAAGT-3′, reverse 5′-ATGAACCCCATACTCCTTCCCAG -3′. Gene expression levels were normalized to GAPDH as an endogenous control.
Morris water maze test
Spatial learning and memory were assessed using a Morris water maze apparatus (Shanghai Xinruan Information Technology Co., Ltd). Mice were trained for 5 consecutive days with four trials per day. Escape latency and crossing numbers were recorded using ANY-maze tracking software (Stoelting Co.).
Golgi staining
Brain tissues were stained using the FD Rapid GolgiStain Kit (FD NeuroTechnologies, Cat# PK401). Neuronal dendritic spines were visualized under a Nikon Eclipse Ni-E microscope (Nikon Corporation). Spine density was quantified using ImageJ software.
Electrophysiological recording
Field excitatory postsynaptic potentials (fEPSPs) were recorded from hippocampal slices (400 μm thick) using Axon Multiclamp 700B amplifier (Molecular Devices). Slices were stimulated with a bipolar tungsten electrode and recordings analyzed using Clampfit software (Molecular Devices).
Flow cytometry (annexin V/PI)
Apoptosis of C8-D1A cells was evaluated using the Muse® Annexin V & Dead Cell Kit (Luminex, MCH100105) following treatment. Briefly, cells (1 × 105 in 50 μL PBS) were incubated with 20 μL of the reagent for 20 min at room temperature in the dark. Samples were then analyzed on the Guava Muse Cell Analyzer (Luminex), with 5000 events recorded per sample. Data acquisition and analysis were performed using the Muse software according to the manufacturer’s protocol.
Fluoro-Jade B (FJB) staining
Brain sections were stained with Fluoro-Jade B (#TR-150-FJB, Biosensis, Thebarton, Australia) to identify degenerating neurons, following the manufacturer’s instructions as we previously described42.
Glutamate concentration measurement
Glutamate concentration of hippocampal tissue was determined using a glutamate assay kit (#MAK004, Sigma, USA). Homogenize hippocampal tissue (50–100 mg) in 100 μL ice-cold Glutamate Assay Buffer, centrifuge (13,000 × g, 10 min, 4 °C), and collect supernatant. Filter through a 10 kDa spin filter if needed. Dilute the 0.1 M glutamate standard to 1 mM, add 0–10 μL to wells (0–10 nmol/well), adjust to 50 μL with buffer. Load 50 μL sample or standard per well, add 100 μL Reaction Mix (90 μL buffer + 8 μL developer + 2 μL enzyme mix). Incubate 30 min at 37 °C (dark), read absorbance at 450 nm. The glutamate production was measured according to the standard curve and the corresponding optical density.
Statistical analysis
For all in vivo experiments, including behavioral assessments and electrophysiological recordings, the experimenters were blinded to the treatment groups during both data acquisition and analysis. Statistical analysis was performed using Prism 10.1.2 software (GraphPad Software, USA), and data presented as means ± SD. Normality tests were performed first. Nonparametric tests were employed for data that did not follow a normal distribution. Comparisons between 2 groups were analyzed by independent two-tailed Student’s t tests, whereas those among more than 2 groups were analyzed using one-way ANOVA with Newman–Keuls multiple comparison test. Statistical significance was set at p < 0.05.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Conclusion
In summary, we propose a mechanistic model in which activated microglia export ACLY via EVs to astrocytes, where elevated p65 acetylation suppresses EAAT1, thereby impairing glutamate clearance and disrupting synaptic plasticity to produce cognitive deficits in POCD. This ACLY–p65–EAAT1 triad integrates metabolism, epigenetic control, and intercellular communication into a unified pathogenic circuit and highlights multiple points of therapeutic entry for postoperative neurocognitive disorders.
Data availability
The data used to support the findings of this study can be obtained from the Supplementary data.
References
Wu, W.-F. et al. The role of epigenetic modification in postoperative cognitive dysfunction. Ageing Res. Rev. 89, 101983 (2023).
Qi, Z. et al. Modulating neuroinflammation and cognitive function in postoperative cognitive dysfunction via CCR5-GPCRs-Ras-MAPK pathway targeting with microglial EVs. CNS Neurosci. Ther. 30, e14924 (2024).
He, L. et al. Unveiling the role of astrocytes in postoperative cognitive dysfunction. Ageing Res. Rev. 95, 102223 (2024).
Spiteri, A. G., Wishart, C. L., Pamphlett, R., Locatelli, G. & King, N. J. C. Microglia and monocytes in inflammatory CNS disease: integrating phenotype and function. Acta Neuropathol. 143, 179–224 (2022).
Yang, N.-S. -Y. et al. mtDNA-cGAS-STING axis-dependent NLRP3 inflammasome activation contributes to postoperative cognitive dysfunction induced by sevoflurane in mice. Int. J. Biol. Sci. 20, 1927–1946 (2024).
Kong, X. et al. Itaconate alleviates anesthesia/surgery-induced cognitive impairment by activating a Nrf2-dependent anti-neuroinflammation and neurogenesis via gut-brain axis. J. Neuroinflammation 21, 104 (2024).
Zhou, Y. et al. Tregs dysfunction aggravates postoperative cognitive impairment in aged mice. J. Neuroinflammation 20, 75 (2023).
Zhang, M. & Yin, Y. Dual roles of anesthetics in postoperative cognitive dysfunction: regulation of microglial activation through inflammatory signaling pathways. Front. Immunol. 14, 1102312 (2023).
Batchuluun, B., Pinkosky, S. L. & Steinberg, G. R. Lipogenesis inhibitors: therapeutic opportunities and challenges. Nat. Rev. Drug Discov. 21, 283–305 (2022).
Desjardins, E. M. et al. Combination of an ACLY inhibitor with a GLP-1R agonist exerts additive benefits on nonalcoholic steatohepatitis and hepatic fibrosis in mice. Cell Rep. Med. 4, 101193 (2023).
Kaymak, I. et al. ACLY and ACSS2 link nutrient-dependent chromatin accessibility to CD8 T cell effector responses. J. Exp. Med. 221, e20231820 (2024).
Shvedunova, M. & Akhtar, A. Modulation of cellular processes by histone and non-histone protein acetylation. Nat. Rev. Mol. Cell Biol. 23, 329–349 (2022).
Altea-Manzano, P. et al. A palmitate-rich metastatic niche enables metastasis growth via p65 acetylation resulting in pro-metastatic NF-κB signaling. Nat. Cancer 4, 344–364 (2023).
Schulz-Kuhnt, A. et al. ATP citrate lyase (ACLY)-dependent immunometabolism in mucosal T cells drives experimental colitis in vivo. Gut 73, 601–612 (2024).
Seguella, L. & Gulbransen, B. D. Enteric glial biology, intercellular signalling and roles in gastrointestinal disease. Nat. Rev. Gastroenterol. Hepatol. 18, 571–587 (2021).
Allen, N. J. & Lyons, D. A. Glia as architects of central nervous system formation and function. Science 362, 181–185 (2018).
Madhu, L. N. et al. Extracellular vesicles from human-induced pluripotent stem cell-derived neural stem cells alleviate proinflammatory cascades within disease-associated microglia in Alzheimer’s disease. J. Extracell. Vesicles 13, e12519 (2024).
Li, Z. et al. M2 microglial small extracellular vesicles reduce glial scar formation via the miR-124/STAT3 pathway after ischemic stroke in mice. Theranostics 11, 1232–1248 (2021).
Upadhya, R., Zingg, W., Shetty, S. & Shetty, A. K. Astrocyte-derived extracellular vesicles: neuroreparative properties and role in the pathogenesis of neurodegenerative disorders. J. Control. Release 323, 225–239 (2020).
Xie, Y. et al. Astrocyte-neuron crosstalk through hedgehog signaling mediates cortical synapse development. Cell Rep. 38, 110416 (2022).
Linnerbauer, M., Wheeler, M. A. & Quintana, F. J. Astrocyte crosstalk in CNS inflammation. Neuron 108, 608–622 (2020).
Guttenplan, K. A. et al. Neurotoxic reactive astrocytes drive neuronal death after retinal injury. Cell Rep. 31, 107776 (2020).
Wang, Y. et al. Caffeic acid phenethyl ester suppresses oxidative stress and regulates M1/M2 microglia polarization via Sirt6/Nrf2 pathway to mitigate cognitive impairment in aged mice following anesthesia and surgery. Antioxidants 12, 714 (2023).
Wang, J. et al. Irisin reprograms microglia through activation of STAT6 and prevents cognitive dysfunction after surgery in mice. Brain. Behav. Immun. 125, 68–91 (2025).
Li, S. et al. Microglia mediate memory dysfunction via excitatory synaptic elimination in a fracture surgery mouse model. J. Neuroinflammation 21, 227 (2024).
Lee, H.-G. et al. Disease-associated astrocyte epigenetic memory promotes CNS pathology. Nature 627, 865–872 (2024).
Thomas, B. L. et al. Molecular determinants of neutrophil extracellular vesicles that drive cartilage regeneration in inflammatory arthritis. Arthritis Rheumatol. 76, 1705–1718 (2024).
Wang, S. et al. Metabolic reprogramming of macrophages during infections and cancer. Cancer Lett. 452, 14–22 (2019).
Kumari, S. et al. The impact of cytokines in neuroinflammation-mediated stroke. Cytokine Growth Factor Rev. 78, 105–119 (2024).
Yang, L. et al. NAT10 inhibition alleviates astrocyte autophagy by impeding ac4C acetylation of Timp1 mRNA in ischemic stroke. Acta Pharm. Sin. B 15, 2575–2592 (2025).
Mi, Y. et al. Loss of fatty acid degradation by astrocytic mitochondria triggers neuroinflammation and neurodegeneration. Nat. Metab. 5, 445–465 (2023).
Lv, M. et al. PHGDH-mediated serine synthesis in astrocytes supports neuroinflammation by sustaining NADH level to promote histone acetylation. Cell Death Dis. 16, 397 (2025).
Giovannelli, L. et al. Mesenchymal stem cell secretome and extracellular vesicles for neurodegenerative diseases: risk-benefit profile and next steps for the market access. Bioact. Mater. 29, 16–35 (2023).
Boulestreau, J. et al. Salivary extracellular vesicles isolation methods impact the robustness of downstream biomarkers detection. Sci. Rep. 14, 31233 (2024).
Balbi, C. et al. Impact of isolation methods on extracellular vesicle functionality in vitro and in vivo. Adv. Biol. 8, e2300185 (2024).
Merij, L. B. et al. Density-based lipoprotein depletion improves extracellular vesicle isolation and functional analysis. J. Thromb. Haemost. 22, 1372–1388 (2024).
Robinson, S. D. et al. Confirming size-exclusion chromatography as a clinically relevant extracellular vesicles separation method from 1mL plasma through a comprehensive comparison of methods. BMC Methods 1, 7 (2024).
Hu, L. et al. Optimized AF4 combined with density cushion ultracentrifugation enables profiling of high-purity human blood extracellular vesicles. J. Extracell. Vesicles 13, e12470 (2024).
Welsh, J. A. et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J. Extracell. Vesicles 13, e12404 (2024).
Guo, M. et al. Exosome-mediated microglia-astrocyte interactions drive neuroinflammation in Parkinson’s disease with Peli1 as a potential therapeutic target. Pharmacol. Res. 219, 107908 (2025).
Wang, D. et al. Sweroside alleviates pressure overload-induced heart failure through targeting CaMKⅡδ to inhibit ROS-mediated NF-κB/NLRP3 in cardiomyocytes. Redox Biol. 74, 103223 (2024).
Qi, Z. et al. M1-type microglia-derived extracellular vesicles overexpressing IL-1R1 promote postoperative cognitive dysfunction by regulating neuronal inflammation. Inflammation 46, 2254–2269 (2023).
Li, S. et al. Low miR-182-5p expressing extracellular vesicles derived from human bone marrow stromal cells of subjects with steroid-induced osteonecrosis of the femoral head aggravate disease progression. J. Bone Miner. Res. 38, 976–993 (2023).
Acknowledgements
We thank the Biobank of the First Affiliated Hospital of Zhengzhou University (China Human Genetic Resources Preservation approval no. [2022]BC0079) for providing sample sectioning, scanning and staining equipments. This study was supported by the National Natural Science Foundation of China (82201331) and the Foundation of Henan Educational Committee (26A320016).
Author information
Authors and Affiliations
Contributions
Z.Q., L.D.: conceptualization, investigation, data curation, writing–original draft; Y.Z., G.C.: methodology, validation; B.C.: data curation; Y.S.: formal analysis; Q.X.: conceptualization; J.Y.: conceptualization, project administration, writing–review and editing. All authors have proof read the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Hyung W. Nam, Miho Terunuma, and the other anonymous reviewer for their contribution to the peer review of this work. Primary handling editor: Benjamin Bessieres. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Qi, Z., Ding, L., Zhao, Y. et al. Astrocytic EAAT1 suppression by EV-ACLY underlies glutamate imbalance and cognitive impairment in POCD. Commun Biol 9, 636 (2026). https://doi.org/10.1038/s42003-026-09888-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-026-09888-1
