
Abstract
Stand structure influences the distribution of light and water and thus affects the functioning of forest ecosystems. However, information on stand structure capturing both horizontal and vertical structures of forest canopies simultaneously is lacking. Here we use principal component analysis to derive a stand structural indicator (SSI) index based on four structural metrics derived from GEDI covering tropical forests and upscale the GEDI footprint-level SSI to a spatially continuous distribution using Sentinel-1&2 imagery. The majority of high SSI values representing higher forest height, density and diversity of canopy height, is found for all tropical forest areas in moist regions, natural forests, and regions with less fire activity. We show that SSI is positively correlated with primary productivity and that the sensitivity of productivity to SSI is larger in natural forests than in managed forests. Our results highlight synthesized stand structure information to support sustainable forest management and conservation.
Similar content being viewed by others

Introduction
Stand structure describes the horizontal and vertical arrangements of forest canopies and influences the distribution of both light and water and thus affects the functioning of forest ecosystems. For example, stand structural complexity can affect biogeochemical cycles and the energy balance by altering light absorption and precipitation interception1,2. Oppositely, these factors (e.g., light and water) also affect the development of stand structure3,4. Higher complex stand structure can also support a large variety of ecological niche space available for a given community5,6, by providing ample shading cover, nutrition for many species and individuals, and creating favorable microclimate benefiting understory diversity7. Moreover, complex stand structure has been documented to considerably influence the forest carbon cycle2,8,9,10,11, promote forest recovery from drought12, and reduce tree mortality during drought13. An improved understanding of the effects of stand structure on tropical forest productivity can thus be of great importance for the planning and sustainable management of tropical forest restoration14. This subject however remains under-studied due to the lack of detailed information (information at a high spatial resolution) on stand structure of forests across larger spatial scales.
Stand structure can be quantified with structural metrics such as canopy cover, leaf area index, tree height, stand density15, as well as structural complexity16, but the metrics used vary considerably among existing studies3,5,8,15. For example, an entropy-based canopy structural complexity index describing the degree of disorder, randomness of structure has been proposed to quantify structure of forest canopy16, while structural density derived from averaging several structural metrics has also been used to characterize forest structure3,5,8,15. Moreover, the structure measurements based on spaceborne, unmanned aerial vehicle/airborne and ground-based lidar laser scanners have resulted in incommensurable estimates of structure across different local-scale studies17,18,19. Considerable efforts have further been made to map spatial patterns of forest structural density and complexity at a regional or global scales, based on a climate data-driven modeling4, or by a simple aggregation of footprint-level structural metrics or complexity index based on spaceborne lidar of GEDI3,20 within gridded cells or from plot-level observations5. The accuracy of these derived maps of structural density or complexity, however, can be severely affected by the uneven distribution of footprints (Lidar shots) available in each gridded cell, while the sparse distributions of plot-level measurements also reduce the accuracy of data-driven structural complexity across large scales21. Ultimately, it is essential to unify the representativeness of the GEDI structural index metrics to quantify forest stand structure and generate wall-to-wall, high-resolution maps of stand structure information covering large spatial scales.
Tropical forests play a key role in mitigating climate change22, but are experiencing increases in disturbances including mortality from climate extremes, fires23 and human exploitation24 that result in carbon loss25 and fragmentation26. An improved understanding of the spatial patterns of forest stand structure over tropical regions and the possible contribution to ecosystem carbon uptake might benefit tropical forest management and restorations27. The GEDI instrument provides a unique opportunity to capture and quantify stand structure across all tropical forest areas, but due to the scattered nature of the footprints recorded, most land areas remain without observations, impeding accurate characterization of forest structural dynamics at local scales. Moreover, several GEDI based structural metrics, including tree height (RH100), total plant area index (PAI), foliage height diversity (FHD), and total canopy cover (CC) respectively quantify stand structure in vertical and horizontal dimensions15, but provide an incomplete assessment of stand structure in three-dimensions when applied individually. An explicit representation of stand structure that allows the integration of different individual structural metrics may better capture the structural variations in both vertical and horizontal dimensions and facilitate the incorporation of structural information into modeling and applications of ecological systems15.
In this study, we propose a means for quantifying spatial patterns of forest stand structure in both horizontal and vertical dimensions and investigate how such structural variations relate to primary productivity across all pantropical regions (Supplementary Fig. 1a). Based on a principal component analysis (PCA), we derive the major functional axis that dominates the variability of four key forest structural metrics (i.e., RH100, PAI, FHD and CC) readily available from the GEDI data collected (Supplementary Fig. 2 and Supplementary Table 1). This first principal component (PC1) captures the structural variations of forest in both vertical and horizontal dimensions simultaneously and is used as a stand structural indicator (SSI) (see “Methods”). In order to advance the spatially detailed monitoring capacity, we upscaled the footprint-level SSI to a spatial continuous distribution across the pantropical region using Sentinel-1 (synthetic aperture radar, SAR) and Seninel-2 imagery (optical images). The complementary use of multiple SAR and optical features based on Sentinel-1 and -2 offers great potential to overcome the limitations of each individual data source in quantifying stand structures28,29. We further tested the hypothesis that SSI is positively correlated with primary productivity (i.e., gross primary productivity, GPP) at the scale of the pantropical region using a structural equation modeling. We hypothesize that a complex stand structure can lead to greater primary productivity, as a more complex stand structure with multiple layers allows trees to make better use of the available light. Specifically, leaves occupy a larger range of light environments and span a wider physiological spectrum enabling to use light more efficiently1,9,10. By controlling for covarying socio-environmental factors (i.e., climate variables, fire activity, human population density, soil properties, and topography), the effects of SSI and tree species richness on primary productivity are expected to provide a better basis for understanding the functioning of tropical forest ecosystems in regards to carbon sequestration and biodiversity.
Results
GEDI based forest stand structure
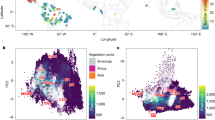
Four stand structural metrics, i.e., tree height (RH100), total plant area index (PAI), foliage height diversity (FHD), and total canopy cover (CC), each of them quantifying the multidimensional forest structure, were derived from the GEDI data and used together to capture structural variations in both horizontal and vertical dimensions. Due to the high collinearity among these structural metrics, we used a principal component analysis (PCA) method to reduce the dimensionality of the four metrics and found that the major axis (the first principal component, PC1) can capture 81.1% of variance in these metrics (Fig. 1a and Supplementary Fig. 3). Moreover, positive correlations between PC1 and the four metrics were observed, with relative contributions of PAI (25.4%), CC (25.0%), RH100 (24.6%) and FHD (24.2%) to PC1 (Fig. 1b). We thus rescaled PC1 to the range between 0 and 1 allowing global comparability and used it as a stand structural indicator (SSI) that captures structural variations in both horizontal and vertical dimensions simultaneously (see “Methods”). A higher value of SSI indicates a higher forest height, density, and foliage height diversity and thereby complex structure. We show that SSI is significantly correlated to two other commonly used structural complexity indices (Supplementary Fig. 4), the so-called entropy-based canopy structural complexity indices, quantifying the degree of disorder, randomness in the structure16.

a The explanatory power of individual principal components (PC1-4), explaining the variations of tree height (RH100, mean = 26.38, S.D. = 10.55), total plant area index (PAI, mean = 2.93, S.D. = 0.41), foliage height diversity (FHD, mean = 0.81, S.D. = 0.23), and total canopy cover (CC, mean = 81%, S.D. = 23%) of the GEDI data; the error bars represent one standard deviation. b The relative contributions of RH100, PAI, FHD and CC to the two axes of PC1 and PC2. c Frequency plots of stand structural indicator (SSI) values across all tropical regions. d Spatial distributions of GEDI footprint-level SSI across all tropical regions. The gray inserts show the frequencies of SSI distributions in the tropical regions of America, Africa and Asia, respectively.
We show left-skewed distributions of SSI based on the footprint-level observations of GEDI across the entire pantropical region and at the scale of each continent (Fig. 1c, d), indicating a generally higher proportion of forest areas having high SSI values as compared to areas having low SSI values. Spatially, SSI shows a latitudinal pattern with a higher SSI in tropical humid areas and lower SSI in tropical dry areas. To reduce potential dependence of variations in SSI on tree cover, we also analyzed the spatial variations in SSI in tropical areas with tree cover larger than 95% and observe spatially distinct distribution of SSI (Supplementary Fig. 5).
Spatial patterns of forest stand structure
To obtain spatially detailed information on stand structure, we upscaled the GEDI footprint-level SSI to a spatial resolution of 30 m × 30 m using Sentinel-1 based polarization features and Sentinel-2 based optical spectral features (Supplementary Table 2). We thus derived a spatial continuous map of SSI at a high level of accuracy of R2 = 0.84, rRMSE = 8% (Supplementary Figs. 6, 7), to characterize the spatial patterns of stand structure across the entire pantropical region. We generally observed left-skewed distributions of SSI values across all tropical continents, agreeing well with the patterns of GEDI footprint-level SSI (Fig. 2a and Fig. 1d), indicating a pronounced spatial coverage of complex forest structure characterized by a higher forest height, density, and foliage height diversity in all tropical regions. The zoom-in examples show distinct spatial variations in SSI, mostly with higher SSI in dense forests and lower SSI in forest edge areas (Fig. 2b and Supplementary Table 3). Considering SSI of three major forest management classes accounting for 96% of all tropical regions (Supplementary Fig. 1b), we show that natural forests without any signs of management (NF) characterized by primary forests, have the highest SSI, followed by natural forest with signs of management (MF) and agroforestry (AF) (Fig. 2c and see the spatial patterns of SSI in NF, MF and AF respectively in Supplementary Fig. 8).

a Spatial pattern of SSI at a spatial resolution of 30 m × 30 m across all tropical forest regions. b Zoom-in examples of SSI for tropical regions of different continents. c The distributions of SSI for different forest management classes including: naturally regenerating forest without any signs of management (NF), and regenerating forest with signs of management (MF), agroforestry (AF). The horizontal lines within the violin plot represent the median of SSI values and the vertical lines denote the first (25th) and third (75th) quartiles of the data distribution for each forest class. d Differences in SSI between dry and moist tropical regions across American, African and Asian tropical forest areas. Error bars denote the 95% confidence interval of SSI. e Differences in SSI between intact and non-intact forests in moist tropical regions across three different continents. In the boxplots, the whiskers represent the fifth and 95th percentiles of the SSI statistics, whereas the outer edges of the boxes and the horizontal lines within the boxes represent the 25th, 75th, and 50th percentiles of the SSI statistics. f Differences in SSI between dry and moist tropical regions against fire intensity indicated by percentage values of burned areas (%). Error bars represent the 95% confidence interval of SSI.
We further divided the pantropical region into dry and moist tropical regions (Supplementary Fig. 1a) and show that forests in moist regions generally have a higher SSI (Fig. 2d), with SSI in moist regions being 37%, 32% and 19% higher than in dry tropical regions of America, Africa and Asia, respectively. Moreover, we show that intact forests30 generally exhibit higher SSI values compared to non-intact forest areas (Fig. 2e). Larger differences in SSI between intact/non-intact forests were observed in America and Asian tropical forests than in the African tropics. Only intact and non-intact forests30 in moist tropical regions were considered, as very little intact forest areas have been preserved in dry tropical regions (Supplementary Fig. 1a). In addition, a higher intensity of fire activity was associated with a lower SSI, which was observed in both dry and moist tropical regions (Fig. 2f and Supplementary Fig. 9e). These results, all together, show the impacts of climate variations, fire disturbances, and human management on stand structure across all tropical forest regions.
The relative contributions of these relevant factors (see the collinearity between these factors in Supplementary Table 4) were further quantified by a boosted regression tree model (BRT), showing that precipitation dominates (43%) the spatial variations of SSI across all pantropical regions, followed by incoming solar radiation (11%) and soil cation exchange capacity (10%). All remaining individual factors including i.e., temperature, soil sand fraction, fire activity, soil organic carbon, topography, and human population density contributed less than 10% to the total variance (Supplementary Fig. 10). We also identified the relative importance of precipitation, temperature and human population as the most important factors influencing SSI in tropical areas with tree cover larger than 95% (Supplementary Fig. 11).
Stand structure in relation to forest primary productivity
To understand how stand structure affected the forest carbon cycle, we fitted the potential relationships among SSI, tree species richness, GPP, and environmental variables (i.e., climate, disturbance, soil properties and topography, see spatial patterns of these abiotic environmental variables in Supplementary Fig. 9) using a structural equation model (SEM) (see “Methods”).
The results show that SSI, tree species richness and environmental variables including climate, disturbances soil properties, and topography have significant (p < 0.01) direct effects on GPP, explaining together 67% of the spatial variability in GPP. Among them, SSI has the largest impact on GPP, with a standardized path coefficient (β) of 0.35, while species richness shows the second most important effect on GPP (β = 0.31). The significant association (r = 0.33, p < 0.01) between GPP and SSI is also observed in tropical areas with >95% tree cover where GPP is weakly correlated with tree cover (Supplementary Fig. 12a–c), which is stronger than the association between each individual structural metrics and GPP (Supplementary Fig. 12d–g). Climate factors (i.e. precipitation, incoming solar radiation, and air temperature) are found to have both direct effects on GPP and indirect effects through species richness (Fig. 3a). SSI is also found to be positively associated with tree species richness (correlation coefficient r = 0.39), however, the causal relationship between them remains a topic of ongoing debate2,31,32. We also observe that the sensitivity of GPP to SSI is highest in NF (slope = 0.411), followed by MF (0.323) and AF (0.273) (Fig. 3b–d), suggesting that the productivity of natural forests is more sensitive to complex structure compared to the more simplified or controlled environments of managed forests.

a Path diagrams showing the relationships by the standardized path coefficients (β) among environmental factors, tree species richness, SSI, and GPP in tropical forests. Only significant pathways are shown (***p < 0.001; **p < 0.01; *p < 0.05), where black and red arrows represent significantly positively and negatively relationships, respectively. Double-headed arrows indicate two-way correlation (0.39). R2 is the degree of SSI and GPP variability explained, respectively. Fisher’s C = 2.847; d.f. = 390; P = 0.241. SSI: stand structural indicator; Richness: tree species richness from Liang, et al.47. Climate includes mean annual temperature (TEM), precipitation (PRE), and incoming solar radiation (RAD). Disturbance factors include human population density (POP) and fire activity (FIRE). Soil and topography include topsoil cation exchange capacity (CEC), sand fraction (SAND) and a digital elevation model (DEM). b–d Partial dependency plots of GPP against SSI for different forest management classes (NF, MF and AF), based on a multiple linear regression (Supplementary Table 5) (n = 5920). Variables were normalized (mean = 0, SD = 1) prior to the analysis. Slope indicates the sensitivity of GPP to SSI and p indicates the significance level based on a t test. The shading denotes the 95% confidence intervals of the linear fitting.
Fire is observed to have a direct negative effect on SSI, species diversity and GPP. Topography shows a significant effect on SSI while soil properties (cation exchange capacity and sand fraction) show significantly but relatively weak effects on SSI and species richness (Fig. 3a). Consistent results were observed with solar-induced chlorophyll fluorescence (SIF) used as a proxy of GPP (Supplementary Fig. 13a) while accounting for the effects of spatial autocorrelation (Supplementary Fig. 14a), supporting the robustness of the relationships observed. We also show similar relationships in tropical areas with tree cover larger than 95% (Supplementary Fig. 13b, c).
Discussion
This study proposed a stand structural indicator (SSI) that captures structural variations in both horizontal and vertical dimensions simultaneously, based on four structural metrics from the GEDI instrument designed to measure and map Earth’s forests in high resolution. We used the key axis (i.e., PC1) of a PCA explaining the most variations in the four-dimensional spectrum of structure metrics to quantify forest stand structure in both horizontal and vertical dimensions33, being different from the structural complexity index developed to describe the degree of disorder or randomness of structure based on a entropy method16. Our approach can reduce the potential uncertainty attributed to the collinearity between different structural metrics and advance the representativeness of volumetric structure (in three dimensions). SSI has indeed integrated multidimensional structure information (e.g., PAI and foliage height diversity) and a higher SSI value can indicate a higher complex structure characterized by a higher forest height, density, and foliage height diversity. SSI is thus expected to lay the foundation for an improved understanding of forest stand structural variations and enrich forest structural monitoring of global forests diversity beyond focusing only on forest canopy height or complexity.
The GEDI footprint-level SSI was subsequently upscaled to a spatial continuous map at a fine spatial resolution of 30 m × 30 m across all tropical regions, by deploying satellite remote sensing imagery from Sentinel-1 and -2. The map represents a substantially enhanced spatial resolution of forest stand structural information (independent of climate variables) compared to large-scale maps generated in previous studies3,4, leveraging new possibilities for studying the underlying relationship between forest structure and ecosystem functioning. This is because that existing baseline maps characterizing forest structure were derived from averaging GEDI footprint-level structural metrics within grid cells20,21. Considerable uncertainty could therefore be introduced due to the uneven distribution of GEDI footprints across grid cells and due to the assumption of an equal weight of individual structural metrics in quantifying structure21.
We show that moist tropical regions, intact forests, and regions with less fire interventions generally have a higher SSI (representing a higher forest height, density, and foliage height diversity), which in turn suggests negative impacts of drying conditions and disturbed forests on structure. Widespread climate droughts have been documented to cause tree mortality and forest degradation in tropical regions23,34, while drought related tree mortality is expected to be worsened by rising atmospheric water stress35,36, and drought induced changing fire regimes (e.g., increasing fire frequency)37 and their interactions of drought and fire34. Also, management practices (e.g., logging, clear cuts, mining, urban expansion) and agroforestry have decreased stand structural complexity, as compared to a higher structural complexity observed in the tropical forests with less managements. These generally indicate the joint controls of climate, disturbances and human pressure on the structure3, and provide essential insights into the need for conservation and restoration of stand structural complexity when implementing tree plantations across tropical regions38. Restoring large tropical forest fragments particularly in primary forests could thereby be an efficient way to improve forest stand structural complexity.
We documented a significant positive relationship between SSI and GPP across tropical regions, supporting the key role of SSI in regulating forest primary productivity that has previously been observed at the local scale or site-level5,10,11,39. Our results have substantially extended the spatial scales for which already established relationships apply and further generalized the current understanding of the relationship between SSI and the carbon cycle2,5,11. Stand structure quantifies the distribution of vegetation canopies in the horizontal and vertical dimensions. A higher complex structure of forest features multilayered canopies that absorb more light (including direct and diffused light) and are thus expected to gain efficient use of light39, as well as water and other resources within the forest40. Additionally, a more structural complex forest can better promote niche partitioning and facilitation among diverse species. This, in turn, allows diverse forest stands to exploit available resources more efficiently than less diverse forest stands, thereby promoting highly functional species that use light more efficiently41,42. Moreover, complex stand structure was also more resilient to disturbances (e.g., drought or pests)12,13, maintaining productivity under conditions of environmental stress. These factors can well support an underlying close relationship between forest stand structure and productivity, particularly for natural forests. This complex structure enhances ecosystem functions (e.g., light interception, nutrient cycling, and microhabitat diversity) and complex ecological interactions among species (e.g., competition, facilitation) are more finely tuned to the existing complex structure, thereby leading to a higher sensitivity of productivity to structural variations in natural forests. In contrast, managed forests are often subject to practices such as thinning, clear-cutting, and planting of monocultures, resulting in a more homogeneous environment where structure has a less pronounced effect on productivity. We thereby highlight the important role of enhancing complex structure in forest management to improve carbon sequestration.
Our results are based on a transferable framework to map spatial details of forest stand structure in both horizontal and vertical dimensions that could help in refining the understanding of the spatial patterns of forest structure globally and uncovering the mechanism of which forest ecosystems are linked with environmental stressors (e.g., the role of structural complexity in enhancing forest resilience to disturbances43,44). We evidenced a positive association between SSI and primary productivity across the pantropical region. In particular, stand structure outperforms tree species richness in explaining the spatial variations in forest productivity, supported by previous studies conducted at site level5,11,39,45, indicating the importance of prioritizing stand structure in regulating carbon cycles. The impact of SSI may vary across forest biomes globally, e.g. the relation between primary productivity and structural complexity of boreal forest areas may be very different due to a less rich species diversity46,47. Conversely, productivity may also shape stand structure by influencing tree growth rates and species composition, which requires further investigation. The saturation effect of the GEDI signal in tall canopies is however likely to introduce an inherent uncertainty in the estimates of structural attributes, thereby affecting the relationships between SSI and productivity. Such saturation problems might be minimized or resolved by new techniques such as artificial intelligence-based methods48. Future studies are also called for to estimate the contribution of stand structure to forest biomass in different forest biomes, and to go beyond focusing only on the height signal2,9,11. In summary, enhancing our knowledge on the nature of global forest stand structure would improve insights into forest functions and guide forest management strategies towards achieving carbon neutrality by implementing forest conservation, restoration and afforestation measures.
Methods
GEDI data
GEDI (Global Ecosystem Dynamics Investigation) is a spaceborne Light Detection and Ranging (LiDAR) instrument mounted on the International Space Station (ISS) and is primarily designed for monitoring the three-dimensional structure of forests49. GEDI provides vertical profiles of the forest canopy structure, covering the Earth’s landmasses between approximately 51.6 degrees north and 51.6 degrees south latitude, making it particularly suitable for studying tropical forest resources. In this study, we collected four structural metrics including the tree height (RH100, mean = 26.38, S.D. = 10.55), total plant area index (PAI, mean = 2.93, S.D. = 0.41), foliage height diversity (FHD, mean = 0.81, S.D. = 0.23), and total canopy cover (CC, mean = 81%, S.D. = 23%) corresponding to ‘rh100’, ‘pai’, ‘fhd_normal’, and ‘cover’ in the GEDI Level 2B products during 2020–2022. For each year we excluded data during periods of leaf-off using the “leaf-off” flag provided as part of the dataset. The RH100 metric represents the height (in meters) at the 100th percentile of the returned laser’s energy relative to the ground. The PAI is one half of the total canopy structural elements per unit ground surfaces, which is estimated by counting the canopy directional gap probability. The FHD metric quantifies the vertical structure diversity in canopy heights within a given ecosystem. The CC is defined as percentage of the ground covered by the vertical projection of canopy structural material, such as leaves, branches and stems.
To ensure data quality, we used the sensitivity metric (i.e., beam sensitivity ≥0.98) and quality flag (i.e., quality flag = 1) provided with GEDI Level 2B products to select data with good quality. Beam sensitivity represents a measure of signal-to-noise related to the canopy through which the waveform has sufficient energy to reach the ground. A quality flag value of 1 in the GEDI Level 2B products typically signifies that the structure metrics for the land surface meet certain criteria such as waveform shot energy, sensitivity, and amplitude, indicating a high level of data quality and reliability49.
Sentinel-1 and -2 imagery
The Sentinel-1 synthetic aperture radar (SAR) and Sentinel-2 surface reflectance data from the Copernicus Open Access Hub were used to upscale forest stand structural indicator. The processing of the ground-range detected (GRD) Sentinel-1 A/B Interferometric Wide Swath (IW) σ 0 measurements in vertical-vertical (VV) and vertical-horizontal (VH) polarization were performed using Google Earth Engine (GEE) JavaScript API. We applied standard Sentinel-1 processing techniques, including thermal noise correction, radiometric calibration, and range-doppler terrain correction (Supplementary Table 2). We obtained Sentinel-2 imagery from the GEE archive, which provides atmospherically corrected surface reflectance images. For each image, we used the quality assurance band to identify and remove any bad-quality observations caused by clouds and snow/ice. Both Sentinel-1 and -2 imagery were resampled to a spatial resolution of 30 m × 30 m per pixel to match the size of GEDI footprints and the inputs (Supplementary Table 2) ingested into models were calculated based on average VV/VH polarization and optical reflectance bands of Sentinel-1 and -2 during 2020–2022.
Primary productivity
Gross primary productivity (GPP) is a measurement of the amount of organic carbon produced during photosynthesis. We obtained the GPP for the years from FluxSat v2.0 products and solar-induced chlorophyll fluorescence (SIF), a proxy of GPP, from the TROPOspheric Monitoring Instrument (TROPOMI)50,51. The FluxSat GPP were upscaled from selected FLUXNET 2015 eddy covariance tower sites based GPP, based on the MCD43C4v006 nadir bidirectional reflectance distribution adjusted reflectance (NBAR) product of the MODerate-resolution Imaging Spectroradiometer (MODIS) instruments on the NASA Terra and Aqua satellites and machine learning algorithms52. The annual average GPP spanning the period of 2018–2021 were used to indicate tropical forest primary productivity. TROPOMI SIF (RTSIF) were generated based on the Caltech TROPOMI SIF data, MODIS NBAR, land surface temperatures, etc51. The annual average RTSIF spanning the period of 2018–2020 were used to indicate tropical forest primary productivity.
Tree species diversity
We used the tree species richness map across global forests at a spatial resolution of 0.025° × 0.025° developed by Liang et al.47. This map was generated by training machine learning models with tree species richness measurements from 773,100 permanent sample plots, in combination with 47 relevant variables such as precipitation, temperature, canopy cover, elevation, slope and soil C/N ratio47.
Environmental data
We collected a set of potential environmental variables to explore the associations between forest stand structural indicator and environmental conditions. These variables were grouped into four categories: climate (i.e., precipitation, incoming solar radiation and temperature), disturbances (i.e., fire activity and human population density), soil properties (i.e., topsoil sand fraction, soil organic carbon and cation exchange capacity in the topsoil), and topography (Supplementary Fig. 9). Climate data including mean annual precipitation (PRE), incoming solar radiation (RAD), and air temperature (TMP) from 1981 to 2020 were obtained from the ECMWF Reanalysis v5 (ERA5)53. Sand fraction (SAND), soil organic carbon (SOC), and cation exchange capacity (CEC) of the clay fraction in the topsoil were obtained from the Regridded Harmonized World Soil Database v.1.254. We quantified the fire intensity using the monthly MODIS burned area product (MCD64A1) by summing the number of fire occurrences for each 500 m pixel from 2001 to 2020. We derived global human population density during 2020 at 30 arc-second (approximately 1 km) grid cells from the Gridded Population of World Version 4 (GPWv4) Revision 1155, which was used as an indicator of human pressure. A digital elevation model (DEM) with 30-m spatial resolution was acquired from the ASTER Global Elevation Model. These variables were resampled to a 500 m × 500 m spatial resolution using bilinear interpolation method.
Quantification of stand structure at GEDI footprint level
Forest stand structure has been quantified by either a single structural metrics (e.g., tree height, stand density, or a diameter at breast height) or a combination of multiple structural metrics. The combined use of RH100, PAI, FHD and CC derived from GEDI footprint-level records provide multidimensional quantifications on stand structure across the global scales (between 51.6° N and 51.6° S) and thereby has potential to improve the understanding of global forest stand structural variations, considering both vertical and horizonal dimensions, and its association with forest ecosystem functioning.
We gridded the study area into 5° × 5° to optimize the handling of large volumes of data and computation. Thus, we retrieved the GEDI Level 2B products during 2020–2022 in each grid cell using the GEE cloud-based platform56. For each laser footprint, four structural metrics, namely RH100, PAI, FHD, and CC, as well as the longitude, latitude and altitude, were extracted. In total, approximately 2.9 × 108 laser footprints, located within 266 grid cells covering the entire pantropical region, were obtained after data-quality filtering
To comprehensively synthetize the structural variations in both horizontal and vertical dimensions, we analyzed the orthogonal variations of the four structural metrics (i.e., RH100, PAI, FHD and CC), using a principal component analysis (PCA). We first normalized the structural metrics from the entire GEDI footprint dataset and computed the covariance matrix of the normalized data. The four forest structural metrics were then subjected to a PCA analysis to single out the variance explained by all metrics. We used the singular value decomposition method to examine the covariance between individual metrics and performed an eigenvalue decomposition of the covariance matrix to obtain the eigenvalues and corresponding eigenvectors. The eigenvector with the highest eigenvalue was PC1 that explained 81.1% of the variance in the four metrics, suggesting strong collinearities between them (Supplementary Table 6), and indicates that PC1 preserved most of the variations of the four original structural metrics. We extracted the variance explained (Fig. 1a) and the respective loadings for the first two principal components (PC1 and PC2) (Supplementary Fig. 3).
To provide a globally consistent, scalable, and continuous index of forest stand structural information, we rescaled the PC1 to the range between 0 and 1 and defined it as a forest stand structural indicator (SSI). The SSI thus captures most of the structural variations in both horizontal and vertical dimensions across the tropical forest regions. A higher value of SSI close to 1 indicates higher forest height, density, and foliage height diversity and thereby complex structure and vice versa. PC2 explains approximately 16% of the variance. The divergent relationships between PC2 and RH100, FHD (positive), and PAI, CC (negative) (Fig. 1b) lead to ambiguity in the interpretation of the structural complexity, and we therefore only investigated the relationships between GEDI footprint-level PC2 and primary productivity in the analysis presented in Supplementary Fig. 12h. The other remaining PCs account for less than 3% of the variance and have been excluded. Finally, we derived SSI as defined above, across the pantropical region based on GEDI footprint-level records, which further served as training data for the establishment of a predictive model used for upscaling.
Mapping SSI with Sentinel-1 and -2 imagery
SSI was upscaled with a Random Forest (RF) regression model with SSI as a response variable and Sentinel-1/2 polarization and optical spectral features (Supplementary Table 2) as explanatory variables. RF has been extensively used for regression, as it avoids overfitting and can incorporate non-parametric data57. We tuned the main hyperparameters of the RF, including number of decision trees, number of variables per split, nodes, input fraction, and randomization seed, to achieve the best prediction performance. To avoid model overfitting and reduce the time required for model training, we conducted subset features selection by eliminating the pairs of predictor variables with high collinearity determined by absolute Spearman’s correlation coefficients ( | ρ | ≥ 0.95). We excluded features that ranked low in the feature importance based on the variable’s permutation importance tests.
Model performance was evaluated using a ten-fold cross-validation method, which ensures that the validation set was independent and spans the entire range of data. We randomly divided the sample data (GEDI footprint-level SSI) into ten folds and performed ten iterations. In each iteration, we used the random 70% of training samples to train the model and the remaining 30% for validation. We calculated the root-mean-square error (RMSE) and coefficient of determination (R2) to assess model performance.
Statistical analysis
We used a boosted regression tree model (BRT) to assess the potential impacts of the explanatory variables (including temperature, precipitation, incoming solar radiation, human population density, fire activity, topography and soil properties) on the spatial variations of tropical forest stand structural indicator (SSI). BRT is an advanced algorithm of machine learning that iteratively fits and combines multiple regression tree models to improve predictive performance, and has shown to consistently outperform other computationally intensive methods such as bagged trees and random forest58. The process for each model was iterated 10 times to avoid contingency. Partial dependency plots resulting from BRT analysis were further obtained to describe how SSI responds to changes in each predictor independent of the other predictors.
To understand the relationships among the factors of SSI, we designed pathways among environmental variables, SSI, tree species richness, and primary productivity using the piecewiseSEM R package59. Our model includes three main pathways: (1) A direct effect of climate (PRE, RAD, TMP), disturbance (POP and FIRE), and soil and topography (DEM, SAND, CEC, SOC) on primary productivity, (2) An indirect effect of climate, soil and topography and disturbance on primarily productivity via its effect on SSI, and (3) An indirect effect of these environmental variables on primarily productivity via its effect on species richness. For simplicity, we only modeled the hypothesized effects, although the feedbacks can occur whereby ecosystems regulate climate changes or the occurrence of wildfire events60,61,62. We hypothesize that SSI will have a positive effect on primary productivity, because a more complex, multi-layered layers stand structure allows trees to make better use of available light as the leaves occupy a larger range of light environments and span a wider physiological spectrum allowing them to use light more efficiently9,10. Meanwhile, species diversity can promote complementary resource use (e.g., nitrogen) among species, and can also vary in their shade tolerance and photosynthetic light-response, allowing species to efficiently partition light gradients within canopies. This supports another hypothesis that species richness has a positive effect on primary productivity. We will also test the correlation between SSI and species richness considering the interactions between them2,31.
We have used samples with a distance of 50 km to minimize spatial autocorrelation between adjacent samples while at the same time retaining enough samples to train the models. The residuals of the modeling between variables based on Gaussian and Exponential correlation structures did not show much signs of spatial autocorrelation and its effect on the modeling is considered negligible. (Supplementary Fig. 14b, c). The goodness of fit of our SEM was assessed using Fisher’s C test statistic. A multiple linear regression model was further used to explore the association between primary productivity and the structural indicator, while controlling for other variables. All explanatory variables were normalized (mean = 0, SD = 1) prior to the analysis. All statistical analyses were performed in R-4.4.1 using the packages gbm63, tidyverse64, lme465 and ggplot266.
Data availability
All data used to support the findings of this study are publicly available. The GEDI data can be obtained at https://lpdaac.usgs.gov/. The Sentinel-1 and -2 imagery can be downloaded from https://dataspace.copernicus.eu/. The gross primary productivity based FluxSat v2.0 are available at https://doi.org/10.3334/ORNLDAAC/1835. RTSIF is available from https://figshare.com/articles/dataset/RTSIF_dataset/19336346/2. The ECMWF Reanalysis v5 (ERA5) data are available at https://cds.climate.copernicus.eu/cdsapp#!/home. The foret management classes map was available from https://zenodo.org/records/4541513. The fire frequency from MODIS (MCD64A1) is available from Google Earth Engine. The global human population density can be downloaded from https://sedac.ciesin.columbia.edu/data/set/gpw-v4-population-count-rev11. The World Soil Database are available at https://daac.ornl.gov/SOILS/guides/HWSD.html. The generated map of SSI was available at the Zenodo repository: https://zenodo.org/records/14270194.
Code availability
The codes that support the main findings in this study are available at the Zenodo repository: https://zenodo.org/records/14270194.
References
Atkins, J. W., Fahey, R. T., Hardiman, B. H. & Gough, C. M. Forest canopy structural complexity and light absorption relationships at the subcontinental scale. J. Geophys Res-Biogeo 123, 1387–1405 (2018).
Ray, T. et al. Tree diversity increases productivity through enhancing structural complexity across mycorrhizal types. Sci. Adv. 9, eadi2362 (2023).
Li, W. et al. Human fingerprint on structural density of forests globally. Nat. Sustainability 6, 368–379 (2023).
Ehbrecht, M. et al. Global patterns and climatic controls of forest structural complexity. Nat. Commun. 12, https://doi.org/10.1038/s41467-020-20767-z (2021).
LaRue, E. A. et al. Structural diversity as a reliable and novel predictor for ecosystem productivity. Front Ecol. Environ. 21, 33–39 (2023).
LaRue, E. A., Hardiman, B. S., Elliott, J. M. & Fei, S. L. Structural diversity as a predictor of ecosystem function. Environ. Res. Lett. 14, 114011 (2019).
Zellweger, F. et al. Forest microclimate dynamics drive plant responses to warming (vol 368, eabd3881, 2020). Science 369, 1066–1066 (2020).
Ma, Q. et al. The coordinated impact of forest internal structural complexity and tree species diversity on forest productivity across forest biomes. Fundamental Res. 4, 1185–1195 (2022).
Aponte, C. et al. Structural diversity underpins carbon storage in Australian temperate forests. Glob. Ecol. Biogeogr. 29, 789–802 (2020).
Hardiman, B. S., Bohrer, G., Gough, C. M., Vogel, C. S. & Curtis, P. S. The role of canopy structural complexity in wood net primary production of a maturing northern deciduous forest. Ecology 92, 1818–1827 (2011).
Crockett, E. T. H. et al. Structural and species diversity explain aboveground carbon storage in forests across the United States: Evidence from GEDI and forest inventory data. Remote Sens. Environ. 295, https://doi.org/10.1016/j.rse.2023.113703 (2023).
Yang, H. et al. Climatic and biotic factors influencing regional declines and recovery of tropical forest biomass from the 2015/16 El Nino. Proc. Natl. Acad. Sci. USA 119, https://doi.org/10.1073/pnas.2101388119 (2022).
Ma, Q. et al. Tree mortality during long-term droughts is lower in structurally complex forest stands. Nat. Commun. 14, https://doi.org/10.1038/s41467-023-43083-8 (2023).
Heinrich, V. H. et al. The carbon sink of secondary and degraded humid tropical forests. Nature 615, 436–442 (2023).
McElhinny, C., Gibbons, P., Brack, C. & Bauhus, J. Forest and woodland stand structural complexity: Its definition and measurement. For. Ecol. Manag 218, 1–24 (2005).
Liu, X. et al. A novel entropy-based method to quantify forest canopy structural complexity from multiplatform lidar point clouds. Remote Sens. Environ. 282, 113280 (2022).
Queinnec, M., White, J. C. & Coops, N. C. Comparing airborne and spaceborne photon-counting LiDAR canopy structural estimates across different boreal forest types. Remote Sens. Environ. 262, https://doi.org/10.1016/j.rse.2021.112510 (2021).
Brede, B. et al. Non-destructive tree volume estimation through quantitative structure modelling: Comparing UAV laser scanning with terrestrial LIDAR. Remote Sens. Environ. 233, https://doi.org/10.1016/j.rse.2019.111355 (2019).
Henning, J. G. & Radtke, P. J. Ground-based laser imaging for assessing three-dimensional forest canopy structure. Photogramm. Eng. Rem. S 72, 1349–1358 (2006).
Liu, X. et al. Enhancing ecosystem productivity and stability with increasing canopy structural complexity in global forests. Sci. Adv. 10, eadl1947 (2024).
Leiterer, R., Furrer, R., Schaepman, M. E. & Morsdorf, F. Forest canopy-structure characterization: A data-driven approach. For. Ecol. Manag 358, 48–61 (2015).
Zhang, W. M. et al. Recent decrease of the impact of tropical temperature on the carbon cycle linked to increased precipitation. Nat. Commun. 14, https://doi.org/10.1038/s41467-023-36727-2 (2023).
Berenguer, E. et al. Tracking the impacts of El Nino drought and fire in human-modified Amazonian forests. Proc. Natl. Acad. Sci. USA 118, https://doi.org/10.1073/pnas.2019377118 (2021).
Fagan, M. et al. The expansion of tree plantations across tropical biomes. Nat. Sustain 5, 681–688 (2022).
Gatti, L. V. et al. Amazonia as a carbon source linked to deforestation and climate change. Nature 595, 388 (2021).
Hansen, M. C. et al. The fate of tropical forest fragments. Sci. Adv. 6, eaax8574 (2020).
Heinrich, V. H. A. et al. The carbon sink of secondary and degraded humid tropical forests. Nature 615, 436 (2023).
Hyde, P. et al. Mapping forest structure for wildlife habitat analysis using multi-sensor (LiDAR, SAR/InSAR, ETM+, Quickbird) synergy. Remote Sens. Environ. 102, 63–73 (2006).
Nandy, S., Srinet, R. & Padalia, H. Mapping forest height and aboveground biomass by integrating ICESat‐2, Sentinel‐1 and Sentinel‐2 data using Random Forest algorithm in northwest Himalayan foothills of India. Geophys. Res. Lett. 48, e2021GL093799 (2021).
Potapov, P. et al. The last frontiers of wilderness: Tracking loss of intact forest landscapes from 2000 to 2013. Sci. Adv. 3, https://doi.org/10.1126/sciadv.1600821 (2017).
Ishii, H. T., Tanabe, S.-i & Hiura, T. Exploring the relationships among canopy structure, stand productivity, and biodiversity of temperate forest ecosystems. For. Sci. 50, 342–355 (2004).
Dănescu, A., Albrecht, A. T. & Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 182, 319–333 (2016).
Díaz, S. et al. The global spectrum of plant form and function. Nature 529, 167 (2016).
Brando, P. M. et al. Abrupt increases in Amazonian tree mortality due to drought-fire interactions. Proc. Natl. Acad. Sci. USA 111, 6347–6352 (2014).
Bauman, D. et al. Tropical tree mortality has increased with rising atmospheric water stress. Nature 608, 528 (2022).
Fang, Z. X., Zhang, W. M., Brandt, M., Abdi, A. M. & Fensholt, R. Globally increasing atmospheric aridity over the 21st century. Earths Fut. 10, https://doi.org/10.1029/2022EF003019 (2022).
Senande-Rivera, M., Insua-Costa, D. & Miguez-Macho, G. Spatial and temporal expansion of global wildland fire activity in response to climate change. Nat. Commun. 13, https://doi.org/10.1038/s41467-022-28835-2 (2022).
Fagan, M. E. et al. The expansion of tree plantations across tropical biomes. Nat. Sustainability 5, 681–688 (2022).
Gough, C. M., Atkins, J. W., Fahey, R. T. & Hardiman, B. S. High rates of primary production in structurally complex forests. Ecology 100, https://doi.org/10.1002/ecy.2864 (2019).
Ishii, H. T., Tanabe, S. & Hiura, T. Exploring the relationships among canopy structure, stand productivity, and biodiversity of temperature forest ecosystems. For. Sci. 50, 342–355 (2004).
Seidel, D. & Ammer, C. Towards a causal understanding of the relationship between structural complexity, productivity, and adaptability of forests based on principles of thermodynamics. Forest Ecol. Manag. 544, https://doi.org/10.1016/j.foreco.2023.121238 (2023).
Alton, P. B., North, P. R. & Los, S. O. The impact of diffuse sunlight on canopy light-use efficiency, gross photosynthetic product and net ecosystem exchange in three forest biomes. Glob. Change Biol. 13, 776–787 (2007).
Seidl, R. & Turner, M. G. Post-disturbance reorganization of forest ecosystems in a changing world. Proc. Natl. Acad. Sci. USA 119, https://doi.org/10.1073/pnas.2202190119 (2022).
Atkins, J. W. et al. Application of multidimensional structural characterization to detect and describe moderate forest disturbance. Ecosphere 11, https://doi.org/10.1002/ecs2.3156 (2020).
Danescu, A., Albrecht, A. T. & Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 182, 319–333 (2016).
Keil, P. & Chase, J. M. Global patterns and drivers of tree diversity integrated across a continuum of spatial grains. Nat. Ecol. Evolut. 3, 390 (2019).
Liang, J. J. et al. Co-limitation towards lower latitudes shapes global forest diversity gradients. Nat. Ecol. Evolut. 6, 1423–1437 (2022).
Reichstein, M. et al. Deep learning and process understanding for data-driven Earth system science. Nature 566, 195–204 (2019).
Duncanson, L. et al. Aboveground biomass density models for NASA’s Global Ecosystem Dynamics Investigation (GEDI) lidar mission. Remote Sens. Environ. 270, https://doi.org/10.1016/j.rse.2021.112845 (2022).
Guanter, L. et al. The TROPOSIF global sun-induced fluorescence dataset from the Sentinel-5P TROPOMI mission. Earth Syst. Sci. Data. 2021, 1–27 (2021).
Chen, X. et al. A long-term reconstructed TROPOMI solar-induced fluorescence dataset using machine learning algorithms. Sci. Data 9, 427 (2022).
Pastorello, G. et al. The FLUXNET2015 dataset and the ONEFlux processing pipeline for eddy covariance data. Sci. Data 7, https://doi.org/10.1038/s41597-020-0534-3 (2020).
Hersbach, H. et al. The ERA5 global reanalysis. Q J. R. Meteor Soc. 146, 1999–2049 (2020).
Wieder, W., Boehnert, J., Bonan, G. & Langseth, M. Regridded harmonized world soil database v1. 2. ORNL DAAC (2014).
Center for International Earth Science Information Network - CIESIN - Columbia University. (NASA Socioeconomic Data and Applications Center (SEDAC), Palisades, New York, 2018).
Gorelick, N. et al. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 202, 18–27 (2017).
Breiman, L. Random forests. Mach. Learn 45, 5–32 (2001).
Elith, J., Leathwick, J. R. & Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 77, 802–813 (2008).
Lefcheck, J. S. piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evolut. 7, 573–579 (2016).
Zhang, W. et al. Global quantification of the bidirectional dependency between soil moisture and vegetation productivity. Agric. For. Meteorol. 313, 108735 (2022).
Wei, F. et al. Nonlinear dynamics of fires in Africa over recent decades controlled by precipitation. Glob. Change Biol. 26, 4495–4505 (2020).
Forzieri, G., Alkama, R., Miralles, D. G. & Cescatti, A. Satellites reveal contrasting responses of regional climate to the widespread greening of Earth. Science 356, 1180–1184 (2017).
Ridgeway, G. Generalized boosted models: A guide to the gbm package. Update 1, 2007 (2007).
Wickham, H. et al. Welcome to the Tidyverse. J. Open Source Softw. 4, 1686 (2019).
Bates, D. et al. Package ‘lme4’. Convergence 12, 2 (2015).
Wickham, H., Chang, W. & Wickham, M. H. Package ‘ggplot2’. Create elegant data visualisations using the grammar of graphics. Version 2, 1–189 (2016).
Acknowledgements
W.Z. and R.F. acknowledge funding support from the Independent Research Fund Denmark (CLISA, grant number 2032-00026B). R.F. acknowledges funding by the research grant DeReEco (34306) from VILLUM FONDEN.
Author information
Authors and Affiliations
Contributions
W.Z. conceived the study. Y.X. and W.Z performed data analyses and drafted figures. W.Z. and Y.X. wrote the first draft of manuscript. M.B., C.R. and J.B. contributed to the discussions of the results. Q.M. and R.F. contributed to the review and editing of the text.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editors: Marie-Therese Noedl, Heike Langenberg, and Mengjie Wang. A peer review file is available
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhang, W., Xi, Y., Brandt, M. et al. Stand structure of tropical forests is strongly associated with primary productivity. Commun Earth Environ 5, 796 (2024). https://doi.org/10.1038/s43247-024-01984-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-024-01984-6
