
Abstract
Ancient DNA and branched glycerol dialkyl glycerol tetraethers are powerful tools to reconstruct past ecosystems and climate in high-latitude lakes, but often require contrasting redox conditions for reliable interpretations. Here, we present a new approach using adjacent oxic and seasonally suboxic lakes from Iceland to 1) better understand the impacts of redox potential on these molecular proxies, and 2) reconstruct Holocene climate, human settlement, and plant history. We observe that oxic lakes provide more reliable branched glycerol dialkyl glycerol tetraethers-based paleotemperature records, possibly due to the diminished presence of bacterial communities associated with reducing environments. On the other hand, oxic conditions in shallow lakes, which are often targeted for ancient DNA, can lead to poor preservation. By combining oxic and suboxic lake sediment records, we show that natural climate, and not humans, has controlled Holocene vegetation changes in northeast Iceland, and therefore provides ideal constraints for Earth system models.
Similar content being viewed by others

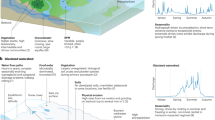

Introduction
Modern anthropogenic warming and enhanced moisture availability are reshaping high-latitude plant communities with important consequences for the climate system and regional biodiversity1,2. These changes include the northward expansion and increased height and density of woody vegetation3,4, which reduce surface albedo5 and increase atmospheric water vapor6, further amplifying high-latitude warming7. Changes in tundra ecosystems will also leave lasting impacts on regional biodiversity, food web structure, and nutrient availability4,8,9. Plant records from warm periods in Earth’s history can provide fundamental insight into the processes that may be expected in our near future and help constrain predictive models10,11. Some high-latitude regions, such as Iceland, offer the opportunity to examine the natural relationship between Holocene changes in climate and plants as humans only colonized Iceland ~1080 years before present (yr BP, Landnámabók). Geologic records that cover the settlement of Iceland also allow us to better understand the impact of pastoral activities on sensitive landscapes12. To this end, recent efforts have targeted high-resolution lake sedimentary records in Iceland13,14,15, but open questions remain on the evolution of Holocene plant communities as well as the relative impact of climate and humans on the landscape12.
Recent analytical advances provide new molecular tools to quantify changes in past temperature, plant communities, and human presence using lipid biomarkers and sedimentary ancient DNA (sedaDNA) in lake sediment. For lipids, the methylation number of branched glycerol dialkyl glycerol tetraethers (brGDGTs), a globally ubiquitous bacterial lipid class, shows strong empirical correlations to warm-season temperatures in high-latitude lakes16,17. For plant histories, sedaDNA metabarcoding provides more reliable records compared to other traditional proxies (i.e., pollen and macrofossils) due to its local source in lake catchments and generally good preservation in sediments18,19,20. Similarly, mammalian sedaDNA provides evidence for the presence of taxa, including those associated with animal husbandry21,22,23. However, brGDGT production and DNA preservation may be influenced by additional processes, including oxygen exposure (e.g., refs. 24,25), which is important to consider for high-latitude lakes that have a wide range of mixing and water column oxygen dynamics26. While the impact of oxygen can be inferred from several geochemical tools, such as archaeal isoprenoidal GDGT (isoGDGT) distributions as a proxy for methanogenesis in sediments and the overlying water column (e.g., refs. 27,28), oxygen exposure is rarely considered in paleoclimate studies.
In this study, we address these shortcomings in a natural laboratory experiment by comparing detailed multi-proxy paleoclimate records from two neighboring lakes in Iceland (separated by 0.6 km and at similar elevation, Fig. 1). Due to different lake morphologies (large and deep vs small and shallow), the two lakes feature contrasting concentrations of dissolved oxygen in their bottom water columns, and the lake’s proximity means that they experience the same climate. Relying on a suite of molecular proxies, including brGDGTs for temperature, sedaDNA for plant and mammal history, and isoGDGTs for redox potential, we answer the following questions. First, how does lake morphology and oxygen availability impact the fidelity of widely used brGDGT and sedaDNA proxies through time? Second, how can sediment records from morphologically contrasting lakes lead to more holistic paleoenvironmental reconstructions than from one site alone? Ultimately, our dual lake approach provides a model for identifying the impacts of oxygen availability when using molecular proxies in Arctic lakes and insight into high-latitude climate-driven ecosystem changes.

a Locations of Stóra and Litla Viðarvatn in northeast Iceland related to other terrestrial (yellow) and marine (blue) paleoclimate record sites. Central volcanoes (triangles) that produced tephra layers used in age models are also marked. b Close-up of Stóra and Litla Viðarvatn, their catchments (dotted lines), bathymetry (10-m isolines), and location of core sites for 20SVID-02 (refs. 14,30) and 20LVID-02/01 (this study). 2017 base image courtesy of Loftmyndir ehf.
Results
Modern seasonal lake stratification and oxygen availability
Stóra Viðarvatn (66.24°N, 15.84°W) is a relatively large (2.5 km2), deep lake (48 m), and Litla Viðarvatn (66.24°N, 15.81°W) is a relatively small (0.2 km2), shallow lake (2.5 m)—both located at 151 m asl and separated by 0.6 km in northeast Iceland (Fig. 1). iButton temperature loggers deployed from September 2019 to 2020 record seasonal fluctuations in lake water temperature in the two lakes, Stóra and Litla Viðarvatn (Fig. 2a, b). Due to the different lake volumes and energy required to heat water, peak summer temperature in Litla Viðarvatn (June, 14.7 °C) is earlier and higher than in Stóra Viðarvatn (August, 10.6 °C). In both lakes, water temperature begins to increase at the end of May, reflecting the seasonal melting of lake ice and overturning of the lake water columns before cooling at the end of August. Surface and bottom water temperatures at Stóra Viðarvatn’s sediment coring location, which is from 20 m depth and not the deepest portion of the lake (Fig. 1b), show minimal thermal stratification during summer and winter months. This is supported by vertically homogenous water quality measurements (i.e., dissolved oxygen, specific conductivity, and pH, ref. 29) in both September 2019 and February 2020 (Figs. 2c and S1). However, the same water quality measurements for Litla Viðarvatn show the development of seasonal stratification in February 2020 (Figs. 2d and S1), reflected by lower bottom water dissolved oxygen concentrations and an oxic water column in September 2019 (Fig. 2d).

a, b iButton temperature measurements from September 2019 to September 2020 for surface (red) and bottom water (blue) (ref. 16) and c, d Sonde dissolved oxygen concentrations (mg/L) from September 2019 (red) and February 2020 (blue) (ref. 29). See Supplementary Fig. 1 for seasonal pH and specific conductivity Sonde measurements.
Lake sediment age models
Sediment core images from Stóra and Litla Viðarvatn demonstrate similar stratigraphies, including visible tephra layers of known age (Supplementary Fig. 2). Based on tephra geochemical analyses in Stóra Viðarvatn14,30, we use the relatively thick and black G10ka Series (10,400–9900 yr BP, ref. 31) and light gray Hekla 3 tephra layers (Hekla 3, 3010 ± 54 yr BP, ref. 32) in Litla Viðarvatn’s age model (Fig. 3b), in addition to 11 AMS radiocarbon dates of plant macrofossils (Supplementary Table 1). Bayesian age models for both lake sediment records show relatively linear sedimentation rates throughout the Holocene, although there is slightly more variability in Litla Viðarvatn, possibly due to the lake’s smaller size and susceptibility to changes in sediment supply (Fig. 3). Stóra Viðarvatn’s record spans the last 10,950 yr BP (Fig. 3a, ref. 30) and Litla Viðarvatn’s record spans the last 9900 yr BP (Fig. 3b). Both age models feature a high density of chronological control points, including common tephra layers (Fig. 3), allowing for high-resolution and synchronized proxy record comparisons between the two lakes.

Age models for (a) Stóra Viðarvatn and (b) Litla Viðarvatn using tephra layers of known age (green) and radiocarbon dates of plant macrofossils (blue) constructed with the R package rbacon (Blaauw and Christen77), and the IntCal20 calibration curve (Reimer et al.78). The solid red line reflects the median of model iterations, and the outer gray lines reflect the 95% confidence envelope. Horizontal gray bars indicate periods of instantaneous tephra layer deposition. See Supplementary Table 1 for radiocarbon information.
Lake sediment organic matter sources
Today, the catchment cover of Stóra and Litla Viðarvatn is comprised of heath and grassland or sparsely vegetated habitats33. Holocene magnetic susceptibility (MS) records from the two lakes are similar, showing relatively high values near the base of each record and then relatively low values until increases beginning at 1150 yr BP in Stóra Viðarvatn and 1400 yr BP in Litla Viðarvatn (Fig. 4a, b), where higher MS values generally reflect the greater contribution of minerogenic material to the lake sediment. Elevated and decreasing MS at the base of the records reflects waning deglacial sediments, and subsequent decadal-scale MS spikes are due to tephra layers. Bulk organic geochemistry in Litla Viðarvatn is characterized by %TOC ranges from 0.19 to 10.7 %, C/N ranges from 3.86 to 11.0, δ13C ranges from −34.9 to −19.4‰, and BSi ranges from 21 to 128 FTIRs absorbance units (Fig. 4). In comparison to Stóra Viðarvatn’s bulk organic geochemistry14, Litla Viðarvatn’s higher %TOC (Fig. 4c, d) reflects a greater loading of organic matter in the sedimentary system, and δ13C values above −22‰ (Fig. 4g, h) indicates that this organic matter is predominately derived from aquatic sources (e.g., ref. 12). Importantly, bulk geochemistry trends for Stóra and Litla Viðarvatn indicate relatively stable sources of organic matter to both lakes following deglaciation until ~1500 yr BP (dashed gray line, Fig. 4), when terrestrial sources become more dominant. Finally, both lakes feature similar BSi records indicating relatively similar responses of diatom productivity through time (e.g., ref. 34). In both cases, the most notable BSi changes are persistent shifts to lower values beginning at 1800 yr BP and 1700 yr BP in Stóra and Litla Viðarvatn, respectively (Fig. 4i, j).

Orange bars reflect portions of the sediment cores likely impacted by low oxygen lake conditions based on isoGDGT-0/crenarchaeol ratios (see Fig. 5a, b). a, b Magnetic susceptibility (MS, SI × 10−5), c, d percent total organic carbon (TOC), e, f carbon to nitrogen ratios (C/N), g, h organic carbon stable isotopes (δ13C, ‰), i, j biogenic silica (BSi, FTIRs absorbance units). Vertical gray dashed line reflects the onset of soil erosion in both lakes at ~1500 BP. Note that for δ13C values, the y-axis is cut off at −28‰, and that lower values are found for Stóra Viðarvatn’s basal sediment (−31.2‰) and in Litla Viðarvatn at ~8700 BP (−34.9‰). Horizontal yellow line broadly separates aquatic and terrestrial organic matter based on δ13C values.
GDGT-inferred redox potential and temperature
Isoprenoid and branched GDGTs are present above the detection limit in all samples from Stóra and Litla Viðarvatn. The ratio of isoGDGT-0/crenarchaeol ranges from 0.88 to 75.6 in Stóra Viðarvatn and from 1.20 to 380 in Litla Viðarvatn (Fig. 5a, b). Stóra Viðarvatn’s isoGDGT-0/crenarchaeol ratio remains low (<10) for most of the record with a temporary relative increase identified between 2900 and 1000 yr BP (Fig. 5a). In contrast, Litla Viðarvatn’s isoGDGT-0/crenarchaeol ratio remains relatively elevated from 8000 yr BP onwards (Fig. 5b). Increased isoGDGT-0/crenarchaeol ratios likely indicate intervals of more reducing conditions (low oxygen concentrations) that promote archaeal methanogens27,35. These reducing conditions could result from increased organic matter deposition and respiration in the lake sediment and/or low dissolved oxygen concentrations in the water column27,35. While we cannot discern between these two possibilities with GDGTs, lake surface sediments are commonly anoxic due to organic matter respiration, particularly in more productive lakes36, whereas variations in water column stratification are a leading control in dissolved oxygen concentrations in lake systems37. Hence, we assume that changes in water column stratification and dissolved oxygen concentrations are more likely to influence varying isoGDGT-0/crenarchaeol ratios in our records. As modern seasonal water column oxygen concentrations differ substantially between the two lakes (Fig. 2c, d), it is reasonable to assume that similar differences in reducing conditions existed throughout the Holocene as well.

Orange bars reflect portions of the sediment cores likely impacted by low oxygen lake conditions based on isoGDGT-0/crenarchaeol ratios (a, b). c, d Total brGDGT concentrations (ng/g TOC), e, f MBT’5Me after De Jonge et al.39, g, h MAF after Raberg et al. (ref. 16) and Otiniano et al. (ref. 17) (°C), and i, j MST anomaly after Harning et al. (ref. 40) (°C). RMSE root mean square error, SE standard error are shown in (h, j).
In Stóra and Litla Viðarvatn sediments, the relative distributions of brGDGTs are similar (Supplementary Fig. 3a). When compared to those of modern Icelandic lake and soil samples, these distribution patterns suggest that brGDGTs in both lakes are generally lake derived while brGDGTs in Litla Viðarvatn may have a minor component from catchment soils (Supplementary Fig. 3a, ref. 38). In terms of temperature proxies (see Materials and Methods), relative MBT’5Me proxy values (Fig. 5e, f, ref. 39) and quantitative temperature for the months above freezing (MAF, refs. 16,17) indices are relatively flat in both Stóra and Litla Viðarvatn (Fig. 5g, h). However, mean summer lake temperature (MST, ref. 40) anomalies (relative to core top, i.e., modern) differ between the two lakes (Fig. 5i, j). MST anomalies from Stóra Viðarvatn show relatively high yet variable temperatures until 5600 yr BP, after which temperatures generally decline towards present. For Litla Viðarvatn, the highest MST anomalies are reached during the earliest portion of the record between 8700 and 8150 yr BP, before declining rapidly and fluctuating between ~0 and 1 °C relative to today for the remainder of the record (Fig. 5i, j).
Plant and mammalian sedaDNA
Of the 54 samples analyzed in Litla Viðarvatn, 47 yield amplifiable plant DNA using the trnL P6 loop primer set (see refs. 41, 30 for Stóra Viðarvatn details). The 7 samples that failed were the samples older than 8000 yr BP. Following data filtering, the trnL dataset yields 8,147,575 total assigned reads, with an average of 150,881 assigned reads per sample. The relative stability of qPCR cycle threshold (CT) values, which reflect PCR efficiency and the quantity of suitable target sequences for amplification, reveal relatively stable trends and indicate that the efficiency of PCR amplification of trnL targets is consistent throughout the record (Supplementary Fig. 4). Metabarcoding technical quality (MTQ) and analytical quality (MAQ) scores are below suggested low quality thresholds (0.75 and 0.1, respectively, ref. 42) in some samples during the Early and Middle Holocene (Supplementary Fig. 4). However, given that the quality scores correlate with species richness and species richness is always below 30 (Supplementary Fig. 4), the low MTQ and MAQ scores are likely an artifact of the requirement that the 10 best represented barcode sequences are required for calculation42, and not necessarily an indication of poor DNA preservation. However, the low MTQ and MAQ scores for the oldest 7 samples likely do reflect poor DNA preservation, as amplification of trnL targets resulted in no assigned reads. We identified 47 plant taxa across a range of plant functional groups throughout Litla Viðarvatn’s lake sediment record, and except for aquatic plants, species richness (calculated as total species identified per sample) generally increases throughout the Holocene in all plant functional groups (Fig. 6). Compared to Stóra Viðarvatn, Litla Viðarvatn has a higher richness of aquatic plants (Fig. 6). As PCR amplification results in sequence read abundances that may not reflect original relative abundances in a sample43 we focus our discussion on taxa presence/absence.

Top: gray diamonds denote where samples were taken and analyzed for DNA metabarcoding. Bubble plots reflect the presence/absence of select taxa, where the size of the bubble is proportional to the number of PCR replicates1–5. Bottom: species richness shown for the total number of taxa as well as four plant functional groups (i.e., aquatic, woody, graminoid, and forb). Orange bars reflect portions of the sediment cores likely impacted by low oxygen lake conditions based on isoGDGT-0/crenarchaeol ratios (see Fig. 5a, b).
For mammalian DNA, we experimented with several primers and approaches targeting mammals, including sheep—the most populous domesticated mammal in Iceland44—in a subset of Late Holocene sediment samples from both lakes (see Supplementary Methods and Discussion). Results using the mammP007 primer, which is commonly used to identify the presence of sheep in lake sediment records (e.g., ref. 22), failed to yield any positive identification of sheep or any other domesticated mammals in Stóra and Litla Viðarvatn. However, we did detect sheep in 5/5 replicates using mammP007 in two modern samples (lake surface sediment and soil) from Vestra Gíslholtsvatn, a lake in southwest Iceland with large modern sheep populations (Fig. 1a). Broadly, the other mammalian primer (16Smamm, ref. 45), sheep-specific primers (L15496 forward primer and H15661 reverse primer, ref. 46), and hybridization capture approaches tested also failed to yield identification of mammals in the lake sediment samples. As sheep are present in the catchments of both lakes today, these results imply that the density of domesticated mammal populations around Stóra and Litla Viðarvatn has been too low to leave a sedaDNA signal.
Discussion
Lake and catchment morphologies
Stóra and Litla Viðarvatn differ substantially in maximum lake depth (48 vs 2.5 m), surface area (2.51 vs 0.21 km2), and catchment area (16.6 vs 1.96 km2) (Fig. 1b), resulting in distinct seasonal physical-chemical properties of the lake water columns. Even though Stóra Viðarvatn’s sediment core and water property measurements were not taken from the deepest portion of the lake, which is located to the south (Fig. 1b), measurements taken at 20 m water depth indicate that the lake remains well mixed even at these greater depths throughout the year (Fig. 2a, c). In contrast, shallow Litla Viðarvatn develops an oxycline during winter, leading to the seasonal depletion of dissolved oxygen in its bottom water (Fig. 2d). We hypothesize that Litla Viðarvatn’s oxycline results from its comparatively smaller volume, where dissolved oxygen is more readily depleted during microbial respiration under winter lake ice cover (e.g., ref. 47). This is supported by water nutrient analyses that demonstrate Litla Viðarvatn has higher TOC/DOC than Stóra Viðarvatn in both summer and winter29. Given that Stóra and Litla Viðarvatn are only separated by 0.6 km (Fig. 1b), any differences in sedaDNA and GDGT proxy records must therefore be controlled by non-climate factors, such as lake morphology and/or physical-chemical properties.
Impact of redox potential on sedaDNA preservation
Our dual lake, multi-proxy approach suggests that Holocene oxygen availability differed substantially between the two lakes. In Stóra Viðarvatn, low isoGDGT-0/crenarchaeol values suggest a reduced presence of archaeal methanogens, except between 2900 and 1000 yr BP (Fig. 5a), implying that Holocene water column and/or sediment oxygen concentrations remained at levels above those supporting methanogenesis. In contrast, isoGDGT-0/crenarchaeol ratios in Litla Viðarvatn are relatively high from 8000 yr BP onwards (Fig. 5b), indicating the presence of reducing conditions that would favor methanogenesis. The proxy-based indications of past redox potential in the two lakes are consistent with our modern seasonal observations, where Stóra Viðarvatn is oxic and Litla Viðarvatn is seasonally reducing in bottom waters. Thus, we assume that Litla Viðarvatn’s reducing conditions from 8000 yr BP onwards likely reflect winter conditions as well.
SedaDNA quality metrics indicate that DNA is poorly preserved in Litla Viðarvatn prior to 8000 yr BP (Supplementary Fig. 4). Failure to identify plant sedaDNA in Litla Viðarvatn at this time is likely due to a lack of trnL amplification targets rather than inhibition, given the uniformity of cycle threshold (CT) values throughout the core. Based on the same timing of oxygen availability inferred from isoGDGT-0/crenarchaeol, we therefore infer that a more oxidizing environment accelerated DNA degradation in Litla Viðarvatn’s earliest record. Given that bulk geochemistry proxies from Litla Viðarvatn, such as δ13C, indicate that organic matter remained primarily aquatic through this interval (Fig. 4h), changes in organic matter source (e.g., terrestrial vs aquatic) are unlikely to explain the observed GDGT ratios. A step shift to high %TOC at the same time as increased isoGDGT-0/crenarchaeol (Fig. 4d) further supports a reduction in redox potential as reduced oxygen exposure is an important process that contributes to increased organic matter preservation36. The sediments of shallow Arctic lakes, like Litla Viðarvatn, are also sensitive to UV radiation due to low DOC and long summer days48 and temperature change due to their small volume (Fig. 2b). While we lack specific data on UV radiation from Litla Viðarvatn, experimental studies indicate that oxygen exposure, UV radiation, temperature, which are all impacted by lake depth, can influence DNA preservation24,48,49,50,51,52. With the available proxy data from our records, we currently infer that increased oxygen exposure and higher temperatures likely contributed to Early Holocene DNA degradation in Litla Viðarvatn.
Other factors beyond oxygen exposure, UV radiation, temperature, and lake depth are also known to impact DNA preservation in lake sediment, such as lake water chemistry (i.e., pH and conductivity), sediment mineralogy, and distance to shore52,53,54. For instance, ref. 52 suggests that sedimentary DNA is better preserved in sediments with lake water pH between 7 and 9 and conductivity between 100 and 500 μS cm−1. Both Stóra and Litla Viðarvatn have modern lake water pH values between 7 and 9, although conductivity for both lakes is considerably lower than 100 (<0.2 μS cm−1) (Supplementary Fig. 1). The identical water chemistries of the two lakes, despite low conductivity levels being potentially detrimental to DNA preservation, suggest that these factors cannot explain the preferentially poor DNA preservation observed in Litla Viðarvatn. Regarding sediment mineralogy, ref. 53 shows that DNA adsorbs best to clays minerals. While we lack detailed data on sediment mineralogy from Stóra and Litla Viðarvatn, both lakes have the same local geology. As a result, there is no expectation that sediment mineralogy would differ between the lakes and preferentially impact DNA preservation. Finally, ref. 54 demonstrates that DNA is more reliably detected closer to shore, particularly for terrestrial taxa. As Litla Viðarvatn’s sediment record is closer to shore than Stóra Viðarvatn’s (Fig. 1b), we would not expect better DNA preservation in Stóra Viðarvatn if distance to shore was an important factor at these lakes. Collectively, given the lack of evidence for the impact of these additional factors and the strong correspondence between independent redox proxies and poor DNA preservation in Litla Viðarvatn’s oldest sediments, a previously oxidizing environment is the simplest explanation.
In contrast to Litla Viðarvatn, Stóra Viðarvatn’s Holocene sedaDNA record is well preserved30, despite that the lake’s water column has been oxic throughout the Holocene (Fig. 5a). A survey of modern high-latitude lake surface sediments indicates that deep lakes generally promote DNA preservation, likely due to some combination of low bottom water temperatures and/or anoxia52. Given its more oxidizing environment, Stóra Viðarvatn’s sedaDNA preservation may therefore be elevated by lower seasonal temperatures (Fig. 2a). This suggests that the interplay of environmental variables that accelerate DNA degradation is complex and that identifying lakes that will yield valuable sedaDNA records may counter prevailing practices. For example, even though small high-latitude lakes are often targeted for sedaDNA studies (e.g., ref. 42), deeper lakes may be more suitable for such pursuits52. In addition, our results show that sediment cores retrieved from below an oxycline in shallow lakes will likely yield well-preserved sedaDNA records as well. As the pattern of postglacial plant colonization is one current focus in sedaDNA research13,30,55,56, our results highlight that reducing conditions and oxygen availability in lakes is an important consideration and that shallow and/or intermittently oxic lakes like Litla Viðarvatn may not reliably preserve the sedaDNA needed to address this fundamental question.
Impact of oxygen and climate on brGDGT-based temperature records
Our comparison of brGDGTs in two different lakes demonstrates how redox potential may impact the reliability of brGDGT-based temperature reconstructions. The similarity between brGDGT distributions in Stóra and Litla Viðarvatn compared to modern Icelandic lake sediments16 suggests in situ brGDGT production dominates in both lakes (Supplementary Fig. 3). Given Litla Viðarvatn’s relatively low brGDGT ΣIIIa/ΣIIa ratios (0.59 and 0.92, Supplementary Fig. 3e), some of the Litla Viðarvatn’s brGDGTs may also be derived from catchment soils57,58 and/or modified by oxygen availability and the composition of the microbial community59. As sediment bulk geochemistry from Litla Viðarvatn demonstrates that the organic matter is predominantly aquatic for the entire record (Fig. 4h), modification of brGDGT distributions under more reducing lake conditions is more likely. These reducing conditions can lead to changes in brGDGT distributions, potentially through the restructuring of the brGDGT-producing bacterial community25,59,60,61,62. However, the exact mechanisms are not currently clear, and oxygen is likely one of many factors affecting brGDGT distributions in high-latitude lakes63. Acknowledging these limitations, it is noteworthy that all of Litla Viðarvatn’s brGDGT inferred temperature histories are flat, in contrast to the established trends of Holocene temperature variability seen in other Icelandic lakes (e.g., ref. 40), when the lake is under reducing conditions. This suggests that changes in microbial community may overprint any brGDGT pattern influenced by temperature changes. Thus, at the very least, we do not currently consider Litla Viðarvatn’s mean summer lake temperature (MST) record to be reliable.
Low isoGDGT-0/crenarchaeol ratios in Stóra Viðarvatn indicate that the lake environment was more oxidizing (Fig. 5a) and that the application of lake-specific brGDGT temperature calibrations is therefore not likely compromised by low oxygen levels. Moreover, the relative stability of ΣIIIa/ΣIIa ratios (Supplementary Fig. 3), bulk geochemistry proxies (Fig. 4), and brGDGT concentrations (Fig. 5c) suggests that there have not been substantial changes in brGDGT or organic matter sources that could impact inferred temperature patterns prior to the onset of soil erosion at ~1500 BP. The qualitative MBT’5Me index, which has been widely used in various brGDGT lake temperature calibrations (e.g., refs. 64,65), yields a relatively flat record for Stóra Viðarvatn (Fig. 5e), similar to Skorarvatn, another oxic lake in northwest Iceland40. While the insensitivity of the MBT’5Me to temperature in Icelandic lake sediments is currently unclear, a recent lake brGDGT mesocosm study shows a limited sensitivity of MBT’5Me to low ambient temperatures66. Similarly, we find that global and Arctic months above freezing (MAF) lake temperature calibrations, which use a multilinear regression model16,17, also produce relatively flat records for Stóra Viðarvatn (Fig. 5g). In contrast to indices such as mean summer temperature (MST), MAF can be challenging to interpret as a short, warm season may have the same value as a relatively long, mild season that extends outside of the summer months. In addition to the fact that MST, where summer refers to JJA months, is a more important control on high-latitude plant communities (e.g., ref. 4), we do not use the MAF metric.
Here, we use an Icelandic lake brGDGT temperature cross-calibration that relies on the strong relationship between the unsaturation of haptophyte alkenones and MST40. The calibration was developed from sediments collected at 25 m water depth in Skorarvatn, an oxic lake in northwest Iceland40, and since it is based on photosynthetic algal lipids, it is taken to reflect summer lake water temperatures when most haptophyte productivity is expected to occur67. Our in situ temperature measurements from both lakes support this, as peak seasonal temperatures are recorded in JJA (Fig. 2a, b). Given the strong similarities in water chemistry and lake morphology between Skorarvatn and Stóra Viðarvatn, the application of this brGDGT temperature calibration anchored in an alkenone calibration is deemed appropriate. Stóra Viðarvatn MST anomalies (relative to modern) reveal a pattern consistent with qualitative and quantitative lake temperature histories in Iceland12,40. More specifically, we find the highest MST anomalies during the Early Holocene (+1.75 °C, Fig. 5i) followed by general summer cooling during the Middle and Late Holocene to the lowest MST anomalies (−2.0 °C, Fig. 5i). If temperatures from Litla Viðarvatn are considered prior to the onset of long-term oxygen limitation, peak Early Holocene temperatures for this region of Iceland may have been up to 2.1 °C higher than today (Fig. 5j). While water column dissolved oxygen does not generally affect Stóra Viðarvatn, elevated isoGDGT-0/crenarchaeol ratios between 2900 and 1000 yr BP suggest a possible period of more reducing conditions, making this interval currently questionable for temperature interpretation. In contrast to Litla Viðarvatn, where a persistent shift in oxygen conditions likely contributed to its relatively flat MST record over the last 8000 years, the brGDGT record from Stóra Viðarvatn highlights that lake sediment records from more oxidizing locations without substantial changes in organic matter source can generally yield reliable records of temperature variability. As commonly targeted shallow lakes are often stratified and seasonally anoxic in the Arctic26 and water column anoxia can impact brGDGT distribitions25,47,59,61,62,63, we recommend using lake sediments acquired from above the seasonal oxycline or an oxic lake like Stóra Viðarvatn for the optimal use of brGDGTs in paleotemperature reconstructions. Moreover, providing auxiliary datasets on potential confounding variables (e.g., sediment source and oxygen availability) will bolster any future applications of brGDGTs as a potential lake temperature proxy.
Climate-driven changes in Holocene plant assemblages
Our dual lake approach provides insight in regional paleoenvironmental history by providing a robust summer temperature record from an oxic lake and complementary vegetation records from both lakes. These records demonstrate that regional summer temperature has been the primary forcing behind broad Holocene vegetation patterns in northeast Iceland. Total species richness in Stóra and Litla Viðarvatn increases through the Holocene with a substantial increase at ~4000 yr BP in both lakes despite differences in identified taxa (Fig. 6 and Supplementary Fig. 5). The increase in total plant species richness at 4000 yr BP corresponds with a transition from woodland to heathland environment, marked by the consistent presence of Vaccinium and Carex, as well as cold- (e.g., Bistorta and Oreojuncus) and dry-tolerant taxa (e.g., Galium, Ranunculus, and Saxifraga) in both lakes (Fig. 6). Our independent MST anomaly estimates from Stóra Viðarvatn are anti-correlated to species richness (R2 = 0.45, p < 0.001), recording peak summer temperature anomalies during the Early Holocene (+1.75 °C and +2.1 °C in Litla Viðarvatn) when species richness is low and lower summer temperature anomalies (−2.0 °C) during the Late Holocene when species richness is high (Fig. 7a, b). Outside of Iceland, sedaDNA studies from Holocene lakes in northern Fennoscandia42 and Late Quaternary sites across the Arctic11 also show relatively increased plant species richness during the Late Holocene (4200 yr BP to present) and Last Glacial Maximum (26,500 to 19,000 yr BP), two periods that were broadly colder than today. The broad consistency across these Arctic and sub-Arctic sites demonstrates that summer temperature has been at least one important control on Holocene plant community patterns in the high latitudes.

a Stóra and Litla Viðarvatn MST anomalies (°C, this study) where orange portion between 2900 and 1000 BP reflects a potential influence of suboxic conditions and SE indicates MST calibration standard error, b Stóra and Litla Viðarvatn sedaDNA species richness (this study), c Ytra-Áland pollen (%TLP, total land pollen, ref. 69), d Stóra Viðarvatn pyroPAHs (ng/g sed, ref. 14), e eastern North Iceland Shelf sea surface (HM107-05, ref. 73) and subsurface temperatures (JR51-GC35, ref. 72), and f North Iceland Shelf sea ice records based on the IP25 biomarker (ng/g sed) from sites MD99−2269 and JR51-GC35 (ref. 71). Vertical blue bars highlight peaks in Ericales and heathland inferred from Ytra-Áland pollen. Dashed blue line at 4000 BP marks the onset of local heathland expansion, and dashed gray line at 1500 BP indicates the onset of local soil erosion.
Following the major change from woodland to heathland environments around Stóra and Litla Viðarvatn at 4000 yr BP, it is challenging to infer relative changes in plant abundance based on sedaDNA, which, for the metabarcoding approach used, is limited to presence/absence. Hence, we look to a Holocene pollen record from Ytra-Áland, a peat section located ~12 km east of Stóra and Litla Viðarvatn (Fig. 1a), to provide complementary information on past changes in plant communities and the factors that have controlled their local Holocene evolution. While pollen may not necessarily originate from the lake catchment like sedaDNA18,19, pollen does generally reflect taxa abundance68. From 9000 to 7900 yr BP and from 2900 to 1000 yr BP, the relative abundance of Betula pollen (green, Fig. 7c) decreases at the expense of Ericales (blue, Fig. 7c), reflecting periods of peak local heathland environments69, which could not be inferred from our sedaDNA records. Prior to 2900 yr BP, Ericales pollen begins to increase at 4000 yr BP (Fig. 7c), which is consistent with our two sedaDNA records and additional local peat pollen records reflecting the Late Holocene expansion of heathland (Fig. 670), the timing of increased sedaDNA species richness (Fig. 7b), declining MST anomalies (Fig. 7a), and increased sea ice presence on the North Iceland Shelf (Fig. 7f, ref. 71). Between 2900 and 1000 yr BP, increased fire frequency reconstructed from pryogenic polycyclic aromatic hydrocarbons (pyroPAHs) in Stóra Viðarvatn (Fig. 7d, ref. 14) further suggests a relatively dryer climate that may have preconditioned the environment to natural fires at this time. As Stóra Viðarvatn’s MST anomalies may be impacted by low oxygen conditions between 2900 and 1000 yr BP (Fig. 7a), we cannot infer quantitative changes in local temperature at this time. However, broadly, these climate changes follow a regional pattern of Late Holocene cooling observed in eastern North Iceland Shelf surface and subsurface temperatures72,73 and drift ice71 and reinforce our conclusions that temperature has been an important control on Holocene vegetation patterns in northeast Iceland. Future efforts to understand the potential impacts of oxygen on Stóra Viðarvatn’s Late Holocene MST record, as well as reconstruct precipitation patterns, will be instrumental.
While the sedaDNA total species richness records from Stóra and Litla Viðarvatn show similar patterns, one key difference between the two is the diversity of aquatic taxa (Supplementary Fig. 5). Bulk geochemical proxies (high δ13C and low C/N) indicate that organic matter in Litla Viðarvatn is predominately aquatic (Fig. 5f, h, ref. 12), which is consistent with higher diversity of aquatic macrophyte DNA (Fig. 6) and with the shallowness of the lake (2.5 m) that permits light penetration needed for higher photosynthetic rates. Litla Viðarvatn’s record includes two notable aquatic taxa not identified in Stóra Viðarvatn that inhabit clear and calm waters: Isoetes and Callitriche. Whereas some cosmopolitan aquatic taxa (e.g., Potamogetonaceae and Myriophyllum) are present throughout Litla Viðarvatn’s record, Isoetes and Callitriche disappear after 1450 and 2150 yr BP, respectively (Fig. 6). Based on bulk geochemistry records from Stóra and Litla Viðarvatn (δ13C and C/N), the onset of Late Holocene soil erosion begins at ~1500 yr BP (Fig. 5, ref. 14), which would have enhanced water turbidity and limited Isoetes productivity. A sharp decrease in Litla Viðarvatn’s BSi occurs at 1500 yr BP as well (Fig. 5d, j), suggesting diatom productivity may have also decreased due to increased soil erosion and more limited sunlight transmission. Considering the current debate on the onset and origins of soil erosion in Iceland (i.e., natural vs anthropogenic, refs. 12,14), our bulk geochemistry and plant sedaDNA evidence are consistent with the onset of persistent soil erosion prior to local human occupation (~1010 yr BP/940 CE, ref. 74). The coincidental timing of increased soil erosion in northeast Iceland and drift ice on the North Iceland Shelf at 1500 BP suggests that the rapid expansion of Polar water masses around Iceland and cooling climate played an important role in these natural environmental changes (Fig. 7f).
The presence and environmental impact of humans in Icelandic lake sediments has largely been assumed based on the timing of acknowledged Norse settlement (~1080 yr BP), although some proxies diagnostic of human presence have been proposed12. Of these proxies, fecal sterols (biomarkers produced in mammal intestinal tracts) and mammalian sedaDNA of humans and/or their domestic livestock offer the best promise as they are more direct indicators of human presence in the catchment21,22,23. In Iceland, fecal biomarker records from Stóra Viðarvatn only show concentrations above background levels in recent centuries, suggesting that following Norse settlement there was either 1) no substantial mammalian population or 2) fecal sterols were diluted in a large lake14. Even with Litla Viðarvatn’s considerably smaller size and higher TOC content (i.e., less minerogenic dilution), our DNA metabarcoding and target capture efforts, which are more source-specific than fecal biomarkers, failed to yield reads for domestic livestock in either lake. Coupled with the positive identification of sheep DNA in our test validation site (Vestra Gílsholtsvatn), the null results from Stóra and Litla Viðarvatn indicate that the lack of mammalian sedaDNA reads is likely due to low sheep populations (e.g., ref. 22). As we also observe no substantial changes in bulk geochemistry or sedaDNA taxa following local human settlement at ~1010 yr BP/870 CE, it is likely that humans and their livestock left a limited footprint on the local terrestrial environment (e.g., ref. 14). In contrast to other Icelandic sedaDNA records that report ecosystem disturbances following settlement (e.g., ref. 13), our climate and plant and mammalian DNA records indicate that northeast Iceland’s environment has been primarily forced by natural climate changes throughout the Holocene. Hence, the records from Stóra and Litla Viðarvatn can provide robust empirical constraints for climate and vegetation models.
In conclusion, to provide robust constraints for predictive models, reconstructing past climate-plant dynamics requires quality records of local climate and plant assemblage. However, two of the most promising proxy methods in the field (brGDGTs and plant sedaDNA) often require contrasting redox conditions for reliable paleoenvironmental interpretations. Consistent with the observed impact of anoxia on brGDGT distributions in other lake systems25,60,61,62, our results from Iceland demonstrate that brGDGTs are best applied in lakes that do not experience reducing conditions for reliable paleotemperature reconstructions. On the other hand, our results demonstrate that plant sedaDNA is best preserved in stratified, reducing settings. Collectively, this highlights that both climate and plant history can be challenging to reconstruct from just one lake site alone, which may be one reason why their complementary analysis from the sediment records is rare (e.g., ref. 55). As shallow lakes may also capture relatively greater proportions of aquatic plant DNA compared to deeper lakes20, we demonstrate that applying a dual lake approach, with geographically proximal oxic and low oxygen lakes, can overcome these hurdles and lead to more holistic and detailed records of climate-plant dynamics in sites across the high latitudes.
Methods
Modern water quality and water chemistry
Stóra Viðarvatn (66.24°N, 15.84°W) is a relatively large (2.5 km2), deep lake (48 m), and Litla Viðarvatn (66.24°N, 15.81°W) is a relatively small (0.2 km2), shallow lake (2.5 m)—both located at located at 151 m asl and separated by 0.6 km in NE Iceland (Fig. 1). We used iButtons loggers (Thermochron DS1925L, Maxim Integrated Products) to measure in situ surface and bottom water (20 m depth) temperatures in Stóra Viðarvatn and surface water temperature only in Litla Viðarvatn (due to the shallow water depth) at 6-h intervals between September 2019 and September 2020 (Fig. 2a, b, ref. 75). We also measured temperature, pH, dissolved oxygen, and conductivity with a multiparameter probe (HydroLab HL4, OTT HydroMet) at ~0.5-m increments along vertical profiles at each lake’s coring location in September 2019 and February 2020 (Figs. 2c, d and S1, ref. 29). We note that bottom water measurements taken for Stóra Viðarvatn are not from the deepest portion of the lake (48 m depth), which is located south of the sampling site (20 m depth) (Fig. 1b).
Lake sediment cores and chronology
In February 2020, we recovered an 8.93 m long core from Stóra Viðarvatn (20SVID-02) and a 7.25 m long core from Litla Viðarvatn (20LVID-02/01) atop lake ice platforms using Bolivia coring systems (Fig. 1b). 20SVID-02 was collected in 7 continuous drives, whereas 20LVID-02/01 was collected in 2 overlapping drives. Sediment sections were photographed and measured for magnetic susceptibility (MS) at the University of Minnesota’s Continental Scientific Drilling Facility, the latter of which was used to splice sections 20LVID-02 and 20LVID-01 into a single composite sediment record (Supplementary Fig. 2). Sealed core sections were stored at 4 °C until sampling was conducted for proxy analyses.
Bayesian age models for Stóra Viðarvatn have been previously published, which use 13 visible tephra layers of known age and their geochemical fingerprints14,30. For Litla Viðarvatn, age control is based on 11 radiocarbon ages from moss macrofossils (Supplementary Table 1) and 2 marker tephra layers identified visually (G10ka Series, 10,400–9900 yr BP, ref. 31, and Hekla 3, 3010 ± 54 yr BP, ref. 32). Radiocarbon samples were given an acid-base-acid pretreatment and graphitized at the University of Colorado Boulder, then measured by AMS at the University of California Irvine. The two marker tephra sequences/layers identified in Litla Viðarvatn are widely dispersed across Iceland31,76, form relatively thick, coarse-grained local deposits (Supplementary Fig. 2), and are geochemically confirmed in the sediment record from Stóra Viðarvatn14,30. We generated a Bayesian age model for Litla Viðarvatn using the R package rbacon, default model functions77, and the IntCal20 calibration curve78. For the 9-cm-thick Hekla 3 tephra layer, we used the “slump” function in rbacon to represent its instantaneous deposition (Fig. 3). For the G10ka Series, we only used the upper boundary dated to ~9900 yr BP as the Litla Viðarvatn sediment record’s bottom ends within the tephra unit and it is unclear if the entire tephra unit extending back to ~10,400 yr BP was recovered.
Bulk sediment geochemistry
Stóra Viðarvatn’s bulk geochemical record has been previously published and is based on 181 samples14. 150 samples were taken from Litla Viðarvatn and measured for total carbon (TC), total nitrogen (TN), and δ13C (relative to VPDB) on a PDZ Europa ANCA-GSL elemental analyzer interfaced to a PDZ Europa 20–20 isotope ratio mass spectrometer at the University of California Davis Stable Isotope Facility. We did not decalcify the samples from either lake sediment record due to the limited stock of inorganic carbon in and around the lakes, and therefore, take TC to reflect total organic carbon (TOC)14. For each of these samples, we also measured biogenic silica by diffuse reflectance Fourier Transform Infrared Spectrometry (FTIRS) on a Bruker Vertex 70 with a Praying Mantis diffuse reflectivity accessory (Harrick) at the University of Colorado Boulder. We report values in FTIR absorbance units.
Lipid biomarkers
At the University of Colorado Boulder Organic Geochemistry Laboratory, we freeze-dried 54 sediment samples from Litla Viðarvatn (~1–2 g) and extracted each two times on a Dionex accelerated solvent extractor (ASE 350) using dichloromethane (DCM):methanol (9:1, v/v) at 100 °C and 1500 psi. For Stóra Viðarvatn, we used 83 previously extracted samples14. A 5 or 10 % aliquot of total lipid extracts (TLE) was taken for glycerol dialkyl glycerol tetraether (GDGT) analysis, resuspended in n-hexane:isopropanol (99:1, v/v), sonicated, vortexed, and then filtered using a 0.45 µm polytetrafluoroethylene (PTFE) syringe filter. Prior to analysis, samples were spiked with 10 ng of the C46 GDGT internal standard79. GDGTs were identified and quantified via high-performance liquid chromatography–mass spectrometry (HPLC-MS) following modified methods of Hopmans et al. (ref. 80) on a Thermo Scientific Ultimate 3000 HPLC interfaced to a Q Exactive Focus Quadrupole-Orbitrap MS16. Isoprenoid and branched GDGTs were identified based on their characteristic masses and elution patterns.
To reconstruct past environmental conditions, we used a variety of published indices and temperature calibrations that rely on the distribution and fractional abundance of isoprenoid and branched GDGTs. While many regional and global temperature calibrations exist for lake sediment brGDGTs, we focus on those that are either local to Iceland40 or “global” and incorporate Icelandic lake sediment samples16,17. This rationale is supported by recent statistical analyses that demonstrate regional clustering of global brGDGT distributions and advocate for site-specific or regional calibrations81.
First, we used the ratio of isoGDGT-0/crenarchaeol as a proxy for the presence of archaeal methanogens27. For relative temperature, we used the MBT’5ME39, which while not including samples from Iceland, forms the basis of many lake brGDGT calibrations:
For quantitative temperature estimates, we used an in situ lake brGDGT calibration from Skorarvatn40, a lake in NW Iceland (Fig. 1a), that capitalizes on the strong relationship between the unsaturation of alkenones, a separate class of lipids produced by haptophyte algae (UK37), and mean summer lake temperature (MST, ref. 67):
While this MST calibration was developed for Skorarvatn’s sediment record specifically, we argue that the application for at least Stóra Viðarvatn is reasonable given the similar water depths of core sites (25 vs 20 m depth, respectively), and therefore, likely similar seasonal water properties. We assume that the reconstructed lake water temperatures reflect June, July, and August based on the peak lake water temperatures recorded in both lakes (Fig. 2) when most alkenone synthesis should occur during haptophyte algal blooms (e.g., ref. 67).
In addition, we used three global lake temperature calibrations that incorporate high-latitude and highly seasonal regions, including Iceland, and reconstruct temperature using the months above freezing (MAF) metric16,17. The first is based on the traditional calculation of brGDGT fractional abundance against all 15 major brGDGTs (i.e., Full Set, ref. 16):
The second is based on a revised fractional abundance calculation that isolates structural groups, such as methylation number and position, and cyclization number (i.e., Methylation Set, ref. 16):
Finally, we used an Arctic lakes temperature calibration that uses the traditional calculation of brGDGT fractional abundance against all 15 major brGDGTs (i.e., Full Set) and the months above freezing (MAF) metric17:
DNA metabarcoding
All DNA sampling was conducted in a dedicated clean lab facility with no PCR products in the University of Colorado Boulder Trace Metal Lab. Stóra Viðarvatn’s sedaDNA plant DNA record has been previously published and is based on 75 samples30. For Litla Viðarvatn, we took 54 samples immediately after splitting the sediment cores. Biomarker samples, as described above, were collected from the same intervals as DNA samples, ensuring that the two time series are time locked.
We performed sample extraction and processing in a dedicated ancient DNA laboratory at the University of California, Santa Cruz Paleogenomics Lab. Before sample extraction, we compared three sedimentary DNA extraction methods82,83,84 to evaluate their individual performance for Icelandic lake sediment (see Supplementary Methods). Based on these results, we extracted lake sediment samples following Rohland et al. (ref. 82). Complete methods for extraction, quantitative PCR (qPCR), trnL metabarcoding, sequencing, and bioinformatic processing are provided in the Supplementary Methods and are identical to those previously published for Stóra Viðarvatn30.
For mammalian DNA, we tested several metabarcoding primers and methods. We initially generated metabarcoding libraries for 16Smamm45 on the extraction comparison sample set, as well as two sheep-specific targeting primer sets46, and then focused efforts on the shorter mammP007 primer in tandem with previously described human blocking oligos22. The sheep-specific and mammP007 primers were tested on a subset of lake sediment samples from Stóra and Litla Viðarvatn. Complete methods for sequencing and bioinformatic processing are provided in the Supplementary Methods. Lastly, we generated single-stranded shotgun libraries from a subset of Litla Viðarvatn lake sediment samples using the Santa Cruz library preparation method85 and performed hybridization capture with a mammalian mitochondrial genome targeting Arbor mybaits set86 and modified to include additional taxa (see Supplementary Methods). To validate the mammalian DNA tests, we collected and analyzed two modern soil and lake surface sediment samples from south Iceland, where sheep are prevalent today (Vestra Gíslholtsvatn, Fig. 1a).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data associated with this manuscript are available at the Arctic Data Center87.
References
Myers-Smith, I. H. et al. Shrub expansion in tundra ecosystems: dynamics, impacts and research priorities. Environ. Res. Lett. 6, 045509 (2011).
Dial, R. J. et al. Arctic sea ice retreat fuels boreal forest advance. Science 383, 877–884 (2024).
Tape, K., Sturm, M. & Racine, C. The evidence for shrub expansion in Northern Alaska and the Pan-Arctic. Glob. Change Biol. 12, 686–702 (2006).
Elmendorf, S. C. et al. Plot-scale evidence of tundra vegetation change and links to recent summer warming. Nat. Clim. Change 2, 453–457 (2012).
Sturm, M., Douglas, T., Racine, C. & Liston, G. E. Changing snow and shrub conditions affect albedo with global implications. J. Geophys. Res. 110, 1–13 (2005).
Pearson, R. G. et al. Shifts in Arctic vegetation and associated feedbacks under climate change. Nat. Clim. Change 3, 673–677 (2013).
Thompson, A. J., Zhu, J., Poulsen, C. J., Tierney, J. E. & Skinner, C. B. Northern Hemisphere vegetation change drives a Holocene thermal maximum. Sci. Adv. 8, eabj6535 (2022).
Fauchald, P., Park, T., Tømmervik, H., Myneni, R. & Hausner, V. H. Arctic greening from warming promotes declines in caribou populations. Sci. Adv. 3, e1601365 (2017).
Collins, C. G., Stajich, J. E., Weber, S. E., Pombubpa, N. & Diez, J. M. Shrub range expansion alters diversity and distribution of soil fungal communities across an alpine elevation gradient. Mol. Ecol. 27, 2461–2476 (2018).
Crump, S. E. et al. Ancient plant DNA reveals High Arctic greening during the last interglacial. Proc. Natl. Acad. Sci. USA 118, 1–9 (2021).
Wang, Y. et al. Late Quaternary dynamics of Arctic biota from ancient environmental genomics. Science 600, 86–92 (2021).
Geirsdóttir, Á et al. Holocene history of landscape instability in Iceland: Can we deconvolve the impacts of climate, volcanism and human activity?. Quat. Sci. Rev. 249, 106633 (2020).
Alsos, I. G. et al. Ancient sedimentary DNA shows rapid post-glacial colonisation of Iceland followed by relatively stable vegetation until the Norse settlement (Landnám) AD 870. Quat. Sci. Rev. 259, 106903 (2021).
Ardenghi, N. et al. A Holocene history of climate, fire, landscape evolution, and human activity in Northeast Iceland. Clim. Past 20, 1087–1123 (2024).
Harning, D. J. et al. High-resolution Holocene record based on detailed tephrochronology from Torfdalsvatn, north Iceland, reveals natural and anthropogenic impacts on terrestrial and aquatic environments. Clim. Past 21, 795–815 (2025).
Raberg, J. H. et al. Revised fractional abundances and warm-season temperatures substantially improve brGDGT calibrations in lake sediments. Biogeosciences 18, 3579–3603 (2021).
Otiniano, G. A. et al. Reconstructing warm-season temperatures using brGDGTs and assessing biases in Holocene temperature records in northern Fennoscandia. Quat. Sci. Rev. 329, 105555 (2024).
Sjögren, P. et al. Lake sedimentary DNA accurately records 20th Century introductions of exotic conifers in Scotland. New Phytol. 213, 929–941 (2017).
Alsos, I. G. et al. Plant DNA metabarcoding of lake sediments: How does it represent the contemporary vegetation. PLoS ONE 13, e0195403 (2018).
Capo, E. et al. Lake sedimentary DNA research on past terrestrial and aquatic biodiversity: overview and recommendations. Quaternary 4, 6 (2021).
Curtin, L. et al. Sedimentary DNA and molecular evidence for early human occupation of the Faroe Islands. Commun. Earth Environ. 2, 253 (2021).
Giguet-Covex, C. et al. Long livestock farming history and human landscape shaping revealed by lake sediment DNA. Nat. Comm. 5, 3211 (2014).
Garcés-Pastor, S. et al. High resolution ancient sedimentary DNA shows that alpine plant diversity is associated with human land use and climate change. Nat. Comm. 13, 6559 (2022).
Lindahl, T. Instability and decay of the primary structure of DNA. Nature 362, 709–715 (1993).
Weber, Y. et al. Redox-dependent niche differentiation provides evidence for multiple bacterial sources of glycerol tetraether lipids in lakes. Proc. Natl. Acad. Sci. USA 115, 10926–10931 (2018).
Klanten, Y., Couture, R.-M., Christoffersen, K. S., Vincent, W. F. & Antoniades, D. Oxygen depletion in Arctic lakes: circumpolar trends, biogeochemical processes, and implications of climate change. Global Biogeochem. Cycles37, e2002GB007616 (2023).
Blaga, C. I., Reichart, G.-J., Heiri, O. & Sinninghe Damsté, J. S. Tetraether membrane lipid distribution in water-column particulate matter and sediments: a study from 47 European lakes along a north-south transect. J. Paleolimnol. 41, 535–540 (2009).
Schneider, T. et al. Tracing Holocene temperatures and human impact in a Greenlandic Lake: Novel insights from hyperspectral imaging and lipid biomarkers. Quat. Sci. Rev. 339, 108851 (2024).
Raberg, J., Harning, D., Geirsdóttir, Á., Sepúlveda, J., & Miller, G. H. Water chemistry profiles of lakes in Iceland (2019-2021). Arctic Data Center. https://doi.org/10.18739/A26688K7F (2023).
Harning, D. J. et al. Delayed postglacial colonization of Betula in Iceland and the circum North Atlantic. eLife 12, RP87749 (2023).
Óladóttir, B. A., Thordarson, T., Geirsdóttir, Á, Jóhannsdóttir, G. E. & Mangerud, J. The Saksunarvatn Ash and the G10ka series tephra. Review and current state of knowledge. Quat. Geochronol. 56, 101041 (2020).
Dugmore, A. J. et al. Radiocarbon dating tephra layers in Britain and Iceland. Radiocarbon 37, 379–388 (1995).
Icelandic Institute of Natural History. Vegetation map of Iceland. https://www.ni.is/en/resources/publications/maps/vegetation-maps (1998).
Conley, D. J. & Schelske, C. L. Biogenic silica. in Tracking Environmental Change Using Lake Sediments (eds Last, W. M. & Smol, J. P.) 281–293 (Springer Science & Business Media, 2002).
Naeher, S. et al. Sources of glycerol dialkyl glycerol tetraethers (GDGTs) in catchment soils, water column and sediments of Lake Rotsee (Switzerland)—Implications for the application of GDGT-based proxies for lakes. Org. Geochem. 66, 164–173 (2014).
Sobek, S. et al. Organic carbon burial efficiency in lake sediments controlled by oxygen exposure time and sediment source. Limnol. Oceanogr. 54, 2243–2254 (2009).
Jane, S. F. et al. Longer duration of seasonal stratification contributes to widespread increases in lake hypoxia and anoxia. Glob. Change Biol. 29, 1009–1023 (2023).
Raberg, J. H. et al. BrGDGT lipids in cold regions reflect summer soil temperature and seasonal soil water chemistry. Geochim. Cosmochim. Acta 369, 111–125 (2024).
De Jonge, C. et al. Occurrence and abundance of 6-methyl branched glycerol dialkyl glycerol tetraethers in soils: implications for palaeoclimate reconstruction. Geochem. Cosmochim. Acta 141, 97–112 (2014).
Harning, D. J. et al. Lipid biomarkers quantify Holocene summer temperature and ice cap sensitivity in Icelandic lakes. Geophys. Res. Lett. 47, 1–11 (2020).
Taberlet, P. et al. Power and limitations of the chloroplast trnL (UAA) intron for plant DNA barcoding. Nucleic Acids Res. 35, e14 (2007).
Rijal, D. P. et al. Sedimentary ancient DNA shows terrestrial plant richness continuously increased over the Holocene in northern Fennoscandia. Sci. Adv. 7, 1–16 (2021).
Nichols, R. V. et al. Minimizing polymerase biases in metabarcoding. Mol. Ecol. Res. 18, 927–939 (2018).
Ross, L. C. et al. Sheep grazing in the North Atlantic region: A long-term perspective on environmental sustainability. Ambio 45, 551–566 (2016).
Taylor, P. G. Reproducibility of ancient DNA sequences from extinct Pleistocene fauna. Mol. Biol Evol. 13, 283–285 (1996).
Cai, D. W., Han, L., Zhang, X. L., Zhou, H. & Zhu, H. DNA analysis of archaeological sheep remains from China. J. Archaeol. Sci. 34, 1347–1355 (2007).
Golosov, S. et al. Physical background of the development of oxygen depletion in ice-covered lakes. Oecologia 151, 331–430 (2007).
Vincent, W. F. & Pienitz, R. Sensitivity of high-latitude freshwater ecosystems to global change: temperature and solar ultraviolet radiation. Geosci. Canada 23, 231–236 (1996).
Lindahl, T. & Nyberg, B. Rate of depurination of native deoxyribonucleic acid. Biochemistry 11, 3610–3618 (1972).
Strickler, K. M., Fremier, A. K. & Goldberg, C. S. Quantifying effects of UV-B, temperature, and pH on eDNA degradation in aquatic microcosms. Biol. Conserv. 183, 85–92 (2015).
Mejbel, H. S., Dodsworth, W. & Rick, F. R. Effects of temperature and oxygen on cyanobacterial DNA preservation in sediments: a comparison study of major taxa. Environ. DNA 4, 717–731 (2022).
Jia, W. et al. Preservation of sedimentary plant DNA is related to lake water chemistry. Environ. DNA 4, 425–439 (2021).
Kanbar, H. J., Olajos, F., Englund, G. & Holmboe, M. Geochemical identification of potential DNA-hotspots and DNA-infrared fingerprints in lake sediments. Appl. Geochem. 122, 104728 (2020).
Wang, Y., Wessels, M., Pedersen, M. W. & Epp, L. S. Spatial distribution of sedimentary DNA is taxon-specific and linked to local occurrence at intra-lake scale. Comm. Earth Environ. 4, 172 (2023).
Crump, S. E. et al. Arctic shrub colonization lagged peak postglacial warmth: molecular evidence in lake sediment from Arctic Canada. Glob. Change Biol. 25, 4244–4256 (2019).
Alsos, I. G. et al. Postglacial species arrival and diversity buildup of northern ecosystems took millennia. Sci. Adv. 8, 1–14 (2022).
Xiao, W. et al. Ubiquitous production of branched glycerol dialkyl glycerol tetraethers (brGDGTs) in global marine environments: a new source indicator for brGDGTs. Biogeosciences 13, 5883–5894 (2016).
Martin, C. et al. Impact of human activities and vegetation changes on the tetraether sources in Lake St Front (Massif Central, France). Org. Geochem. 135, 38–52 (2019).
Zander, P. D. et al. Reconstruction of warm-season temperatures in central Europe during the past 60000 years from lacustrine branched glycerol dialkyl glycerol tetraethers (brGDGTs). Clim. Past 20, 841–864 (2024).
Baxter, A. J. et al. Disentangling influences of climate variability and lake-system evolution on climate proxies derived from isoprenoid and branched glycerol dialkyl glycerol tetraethers (GDGTs): the 250 kyr Lake Chala record. Biogeosciences 21, 2877–2908 (2024).
Colcord, D. E. et al. Assessment of branched GDGTs as temperature proxies in sedimentary records from several small lakes in southwestern Greenland. Org. Geochem. 82, 33–41 (2015).
Van Bree, L. G. J. et al. Seasonal variability and sources of in situ brGDGT production in a permanently stratified African crater lake. Biogeosciences 17, 5443–5463 (2020).
Raberg, J. H., de Wet, G., Geirsdóttir, Á., Sepúlveda, J., & Miller, G. H. Oxygen depletion in lake waters may skew brGDGT-inferred temperatures by more than 10 °C. Geophys. Res. Lett. 52, e2024GL113562 (2025).
Russell, J. M., Hopmans, E. C., Loomis, S. E., Liang, J. & Sinninghe Damsté, J. S. Distributions of 5- and 6-methyl branched glycerol dialkyl glycerol tetraethers (brGDGTs) in East African lake sediment: effects of temperature, pH, and new lacustrine paleotemperature calibrations. Org. Geochem. 117, 56–69 (2018).
Zhao, B. et al. Development of an in situ branched GDGT calibration in Lake 578, southern Greenland. Org. Geochem. 152, 104168 (2021).
Ajallooeian, F., Deng, L., Lever, M. A. & De Jonge, C. Seasonal temperature dependency of aquatic branched glycerol dialkyl glycerol tetraethers: a mesocosm approach. Org. Geochem. 189, 104742 (2024).
D’Andrea, W. J., Theroux, S., Bradley, R. S. & Huang, X. Does phylogeny control UK37-temperature sensitivity? Implications for lacustrine alkenone paleothermometry. Geochim. Cosmochim. Acta 175, 168–180 (2016).
Birks, H. J. B. et al. Does pollen-assemblage richness reflect floristic richness? A review of recent developments and future challenges. Rev. Palaeobot. Palynol. 228, 1–25 (2016).
Karlsdóttir, L. et al. Birch hybridization in Thistilfjördur, North-east Iceland during the Holocene. Ice. Agric. Sci. 27, 95–109 (2014).
Roy, N., Bhiry, N., Woollett, J. & Fréchette, B. Vegetation history since the mid-Holocene in northeastern Iceland. Écoscience 25, 109–123 (2018).
Cabedo-Sanz, P., Belt, S. T., Jennings, A. E., Andrews, J. T. & Geirsdóttir, Á Variability in drift ice export from the Arctic Ocean to the North Icelandic Shelf over the last 8000 years: a multi-proxy evaluation. Quat. Sci. Rev. 146, 99–115 (2016).
Harning, D. J. et al. Response of biological productivity to North Atlantic marine front migration during the Holocene. Clim. Past 17, 379–396 (2021).
Sha, L. et al. Diatom-reconstructed summer sea-surface temperatures and climatic events off North Iceland during the last deglaciation and Holocene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 602, 111154 (2022).
Roy, N. et al. Perspectives of landscape change following early settlement (landnám) in Svalbarðstunga, northeastern Iceland. Boreas 47, 671–686 (2018).
Raberg, J., Harning, D., Geirsdóttir, Á., Sepúlveda, J., & Miller, G. H. Soil and lake water temperatures of Iceland (2019-2021). Arctic Data Center. https://doi.org/10.18739/A2XP6V46R (2021).
Larsen, G. & Thorarinsson, S. H4 and other acid Hekla tephra layers. Jökull 27, 28–46 (1977).
Blaauw, M. & Christen, J. A. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 6, 457–474 (2011).
Reimer, P. J. et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0-55 cal kBP). Radiocarbon 62, 725–757 (2020).
Huguet, C. et al. An improved method to determine the absolute abundance of glycerol dibiphytanyl glycerol tetraether lipids. Org. Geochem. 37, 1036–1041 (2006).
Hopmans, E. C., Schouten, S. & Sinninghe Damsté, J. S. The effect of improved chromatography on GDGT-based palaeoproxies. Org. Geochem. 93, 1–6 (2016).
O’Beirne, M. D., Scott, W. P. & Werne, J. P. A critical assessment of lacustrine branched glycerol dialkyl glycerol tetraether (brGDGT) temperature calibration models. Geochim. Cosmochim. Acta 359, 100–118 (2023).
Rohland, N., Glocke, I., Aximu-Petri, A. & Meyer, M. Extraction of highly degraded DNA from ancient bones, teeth and sediments for high-throughout sequencing. Nat. Protoc. 13, 2447–2461 (2018).
Seersholm, F. V. et al. Rapid range shifts and megafaunal extinctions associated with late Pleistocene climate change. Nat. Comm. 11, 1–10 (2020).
Murchie, T. J. et al. Optimizing extraction and targeted capture of ancient environmental DNA for reconstructing past environments using the PalaeoChip Arctic-1.0 bait-set. Quat. Res. 99, 305–328 (2021).
Kapp, J. D., Green, R. E. & Shapiro, B. A fast and efficient single-stranded genomic library preparation method optimized for ancient DNA. J. Hered. 112, 241–249 (2021).
Slon, V. et al. Mammalian mitochondrial capture, a tool for rapid screening of DNA preservation in faunal and undiagnostic remains, and its application to Middle Pleistocene specimens from Qesem Cave (Israel). Quat. Int. 398, 210–218 (2016).
Harning, D. et al. Holocene climate and vegetation history from Stóra and Litla Viðarvatn, Iceland (2019-2024). Arctic Data Center. https://doi.org/10.18739/A2DB7VS14 (2025).
Acknowledgements
This study has been supported by the National Science Foundation (ARCSS #1836981). We kindly thank Sveinbjörn Steinþórsson and Þór Blöndahl for lake coring assistance, Prof. Thomas Marchitto for access to the Trace Metal Lab, Brooke Holman, Isiah Castro, and Nadia Dildar for lab assistance, and Rienk Smittenberg and an anonymous reviewer for improving the quality and clarity of the manuscript.
Author information
Authors and Affiliations
Contributions
G.H.M., Á.G., J.S., and B.S. funded the project; G.H.M., Á.G., D.J.H., and J.H.R. cored the lakes; J.H.R., D.J.H., and Á.G. collected modern lake quality data, and J.H.R. processed them; D.J.H., Á.G., and N.A. sampled lake sediment; D.J.H. generated age model, bulk geochemistry, and GDGT datasets; S.S. performed DNA lab work and experiments, and D.J.H. performed bioinformatics; D.J.H. wrote the manuscript with contributions from all co-authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth and Environment thanks Rienk Smittenberg and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer review was single-anonymous OR Peer review was double-anonymous. Primary Handling Editors: Ola Kwiecien, Somaparna Ghosh, Aliénor Lavergne.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Harning, D.J., Sacco, S., Raberg, J.H. et al. Both redox potential and climate control molecular proxies in Icelandic Holocene lake sediments. Commun Earth Environ 6, 763 (2025). https://doi.org/10.1038/s43247-025-02701-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-025-02701-7
