
Abstract
The Zambezian Biome of southern-central Africa was an important environmental setting for ancient human interactions and biogeographic dispersals. Here, we assess herbivore migratory behavior and habitat selection with a ~ 2,500 km2 isoscape of bioavailable strontium (87Sr/86Sr) for the Kasitu Valley of northern Malawi, and a probabilistic spatial assignment of fossil herbivore enamel from four sites dated to the last 25,000 calibrated years BP. The results show a very large range in bioavailable 87Sr/86Sr values in the study area (0.7248-0.8521) and reveal that foragers hunted small game locally and procured most large prey in the Afromontane grasslands to the southeast of the study area and along narrow riparian corridors. We found no evidence of migratory behavior in species that exhibit this behavior today. Ancient foragers likely hunted prey that were available year-round, consistent with zooarchaeological and genetic evidence for reduced mobility at the end of the Pleistocene.
Similar content being viewed by others

Introduction
The Zambezian biome (ZB) is an ecoregion of southern-central Africa, comprising 13% of the African continental land, that is characterized by biotic cohorts assembled on nutrient-poor soils derived from Precambrian geology. These soils support open-canopy woodlands and thickets locally known as miombo, seasonally flooded grasslands and reed swamps (dambos), seasonal gallery forests, and higher-elevation grasslands (generally above ~1600 masl) that contain patches of evergreen-dominated Afromontane forest1,2. Human foragers inhabited the ZB throughout the Pleistocene, but little is known about land use and subsistence strategies and how they differed compared to more comprehensively studied areas in eastern and southern Africa3,4. Previous archeological and paleoenvironmental research in Malawi showed an increase in human impact on the environment ~85,000 years (ka) ago, which may have accompanied an increase in regional population sizes5. aDNA of Late Pleistocene and early Holocene foragers from across the ZB has shown that foragers participated in mating networks connecting central, eastern and southern Africa between 20 and 80 ka ago, and that sometime after the Last Glacial Maximum (LGM, ending ~19.5 ka ago)6 long-distance genetic exchange transitioned to more locally restricted mating interactions at the scale of a few tens of km7. Local lifeways are also reflected in a focus on short-distance hunting of small ungulates at Hora 1 (HOR-1) rock shelter4. To understand the role of ecology in this major change in population connectivity during the LGM, it is necessary to reconstruct the mobility of people and their prey at a multi-site, landscape level. Understanding the seasonal distribution of prey on the landscape also informs hypotheses of forager socio-territorial organization, where seasonally predictable clustering of resources at key locations may have favored the establishment of more territorial behavior8,9,10.
Radiogenic strontium isotope ratios (87Sr/86Sr) in biological hard tissues are a common tool to estimate the provenience and movement history of individuals, since they reflect 87Sr/86Sr of the substrate with no trophic fractionation11,12,13. This approach requires an understanding of how environmental 87Sr/86Sr values vary spatially, which is best achieved with an isoscape, i.e., a predictive model of isotope values at unsampled locations14,15. Furthermore, when teeth are sampled sequentially along the growth axis, it is possible to obtain intra-annual time series reflecting migratory behavior16,17. For further details, see Supplementary Text 1. Here, we present a plant-based isoscape (Nplants = 196, plus 25 samples of dust and 12 gastropods) of bioavailable 87Sr/86Sr for an area of over 2500 km2 in the upper part of the Kasitu Valley in northern Malawi (Fig. 1). The area has been under archaeological field investigation by MALAPP (Malawi Ancient Lifeways and Peoples Project) between 2016 and 2023, which has led to the recovery of materials whose provenience can be tracked using a Sr isoscape (Supplementary Text 2), as well as Pleistocene human remains18 documenting shifts in population connectivity7. We applied Empirical Bayesian Kriging19 to generate the isoscape and used it to create probabilistic assessments of provenience for a sample of archaeological faunal remains (Nteeth = 45, Nsamples = 166; Table 1) and test the hypothesis that most prey hunted by humans lived within the confines of our study area during dental development, consistent with the aDNA evidence for reduced mobility and “local” lifeways over the last ~20 ka.

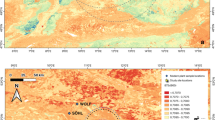
Left: digital elevation model of the study area showing the excavated sites, the three main rivers, and the sampling locations, color-coded by 87Sr/86Sr of the samples collected at each location. Color-coding of the samples follows natural breaks (jenks57) in the data. Note the transect of samples extending east from Hora Mountain to the Kasitu River. Right: map of Africa with Malawi indicated, showing the extent of the Zambezian biome (green shading). The study location is located in the north of the country.
We defined our study area as the landscape within a radius of ~25 km from Hora Mountain and encompassing all the archaeological sites investigated by MALAPP (Fig. 1). Such a radius should reasonably encompass most of the daily foraging activities of the people inhabiting these sites, based on ethnographic data on modern foragers from the ZB20,21. The overall landscape is dominated by Precambrian to Lower Paleozoic Basement Complex bedrock granitic gneiss inselbergs, as well as biotite and muscovite schists, and gneiss outcrops, interspersed between broad flat valleys filled with young (Quaternary) sandy sediments. The valleys also include alluvial and floodplain terrace dambos supporting “rivers of grass” with patches of reeds and sedges2,22 (Fig. 2). The southeastern quarter of our study area is occupied by a mosaic of Afromontane vegetation with grassland being the dominant component1 (Supplementary Fig. 1). The rest of the study area is mostly miombo woodland, which loses most of its understory grasses during the dry season (Supplementary Fig. 1).

A Open woodland; B dambo in the Kasitu Valley; C dambo in Vwaza marsh (close but outside of the study area); D Mountain grassland in the Nyika plateau. Credit for photos (A, C, D) to Jessica Thompson; credit for photo (B) to Chelsea Smith.
Results
Our plant sample records a very large range and variation of 87Sr/86Sr among collection sites (Fig. 3A and Supplementary Data). The range of 87Sr/86Sr in non-riparian plant samples is wider (0.7248–0.8521; Δ = 0.1273, x̄ = 0.7669) than that of riparian (including dambo) plant samples (0.7383–0.7860; Δ = 0.0477, x̄ = 0.7571). Land snail shell 87Sr/86Sr ranges from 0.7183 to 0.7861 (Δ = 0.0678). Although gastropods were collected opportunistically, they sample a variety of localities (Supplementary Data). Intra-plant 87Sr/86Sr variability (leaves 1 m above ground—leaves 2 m above ground; n = 34 pairs; Fig. 3B) averages 0.0007 ± 0.0011. Intra-locality 87Sr/86Sr variability (mixed grass—leaf average; n = 30 pairs; Fig. 3 C) averages 0.0029 ± 0.0033. 87Sr/86Sr differences between leaves and the dust deposited on them are high and average 0.0104 ± 0.0086 (n = 25 pairs; Fig. 3D). In 22 of 25 cases, dust had lower 87Sr/86Sr than the leaves it had coated.

A Distribution of plant 87Sr/86Sr values across bins 0.01 wide. Color-coding highlights natural breaks (jenks) in the data. Note that some natural breaks occur within bins. B Relationship between 87Sr/86Sr values in leaves collected 1 m above ground vs 2 m above ground, within individual trees. C Relationship between 87Sr/86Sr values in mixed grasses vs leaves collected at the same locality. D Relationship between 87Sr/86Sr values in leaves vs the dust deposited on them. Red dashed lines in plots (B, C, D) represent perfect correspondence (slope = 1) between the pairs of samples.
Concentrations of trace elements that might suggest diagenetic alteration (rubidium, uranium, vanadium, and rare earth elements)23 were higher than expected based on modern mammals in 20 enamel samples, but none of the 87Sr/86Sr ratios are outliers relative to other samples in the same tooth or in other teeth of the same taxon (Supplementary text 8). Therefore, all measurements are included in the analysis. Our enamel samples span 87Sr/86Sr from 0.7299 to 0.7813 (Δ = 0.0514; see Supplementary Data). Different taxa yield different ranges of 87Sr/86Sr ratios (Fig. 4). Zebras have the lowest ratios (0.7299–0.7654). Bovids have intermediate ratios (0.7390–0.7697), when excluding an individual reduncin (puku, 87Sr/86Sr = 0.7813) with a very high ratio. The average 87Sr/86Sr ratio for zebra (0.7482 ± 0.0103, 1 sigma) is not significantly lower than that of bovids (0.7570 ± 0.0005) (Wilcoxon rank sum test [two-tailed], W = 138, p = 0.1). Bushpigs (0.7509–0.7568) also have intermediate ratios, while warthogs (0.7587–0.7770) and rhinos (0.7473–0.7729) have higher ratios, except for one warthog with low ratios (0.7346-0.7393). Warthogs and rhinos are not statistically significantly different from zebras and bovids. Diachronically, average zebra 87Sr/86Sr is significantly higher 25–15 ka (0.7497 ± 0.0108) than 14–10 ka (0.7361 ± 0.0004) (W = 271, p = 0.02). The average zebra 87Sr/86Sr 6.5–0 ka (0.7468 ± 0.0025) is indistinguishable from the 25–15 ka average, but significantly different from the 14–10 ka average (W = 0, p < 0.001). Average small bovid 87Sr/86Sr decreases between the older time bins (25–15 ka: 0.7560 ± 0.0042; 10– 8 ka: 0.7615 ± 0.0055) and 6.5–0 ka (0.7500 ± 0.0062), but the difference is statistically significant only between the two Holocene bins (W = 15, p = 0.03). When pooling the data by archaeological site (Supplementary Fig. 2), the HOR-1 samples yield a significantly higher average (0.7588 ± 0.0069, Nsamples = 105) compared to the MAZ-1 samples (0.7437 ± 0.0089, Nsamples = 55) (W = 5355.5, p < 0.001). The sample sizes from HOR-5 and KAD-1 are too small for this comparison.

Note that the two Hippotragus sp. teeth sample the same individual. Silhouettes from phylopic.org.
Figures 5 and 6 show 87Sr/86Sr variation within individual sequentially sampled zebra Equus quagga and alcelaphin teeth, plotted against δ18O series from the same teeth as a proxy for season of formation (high δ18O values representing the dry season and low δ18O values representing the wet season; data from Bertacchi et al., in review). In most individuals, variation is small compared to the range of values measured in plant samples (Table 2), which is expected as plants sample point locations while mobile animals average across their range. Intra-tooth variability in 87Sr/86Sr in several individuals follows a quasi-sinusoidal pattern consistent with directional movement over several months (e.g., specimens #12711, #46459, #101126) but with very small overall change. Zebra #67935 stands out as the specimen with the largest intra-tooth variation (0.0121) and the pattern of change in 87Sr/86Sr is mirrored by changes in δ18O values, with a few mm of “temporal lag” between the twocf 24. This individual has higher 87Sr/86Sr associated with higher δ18O values and lower 87Sr/86Sr associated with lower δ18O values, but this intratooth profile likely does not capture a full seasonal cycle (based on the shape of the oxygen curve). Intra-tooth variation in other taxa is displayed in Supplementary Fig. 4. 87Sr/86Sr variation in Hippotragus sp. (0.0011) and Kobus cf. ellipsiprymnus (0.0001) is very small, consistently with limited seasonal mobility over a homogenous substrate. In contrast, 87Sr/86Sr variation in warthog Phacochoerus africanus (0.0047), kudu Tragelaphus strepsiceros (0.0056), and white rhino Ceratotherium simum (0.0029) is larger, suggesting either a larger home range or the use of habitats situated at the interface of different substrates.

δ18O values and ages from Bertacchi et al. (in review). Enamel goes from formed earlier to the left to formed later to the right (note axis inversion depending on preservation of the cervix). Calibrated ages: #11736: 3.8–3.6 ka cal BP; #9354: 12.5–12.0 ka cal BP; #41169: 15.7–15.3 ka cal BP; #12711: 16.6–16.3 ka cal BP; #46459: 22.0–21.5 ka cal BP; #82032: 17.4–17.1 cal BP; #67935: 17.0–16.6 cal BP. Silhouettes from phylopic.org.

δ18O values and ages from Bertacchi et al. (in review). Enamel goes from formed earlier to the left to formed later to the right (note axis inversion depending on preservation of the cervix). Calibrated ages: #152874.8: 10.7–10.5 ka cal BP; #75753: 15.3–15.0 ka cal BP; #101126: 16.9–16.5 ka cal BP; #153057.5: 15.4–15.0 ka cal BP. Specimens #67572, 64737, and #67485 are not directly dated. Silhouettes from phylopic.org.
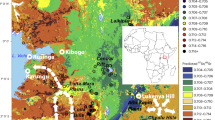
The EBK-generated isoscape is shown in Fig. 7. The standard error of the predicted 87Sr/86Sr is relatively high, especially at the undersampled margins of our study area. This highlights the heterogeneity of 87Sr/86Sr in, and the geological complexity of, the Kasitu Valley. Visual inspection suggests that several low values occur in the higher elevation southeastern portion of the study area. We assessed variations of this isoscape but found this version to perform better overall (Supplementary Text 7).

Left: predicted 87Sr/86Sr in the study area based on an Empirical Bayesian Kriging (EBK) model. Right: standard error of the predicted 87Sr/86Sr. Basemap from ESRI World Imagery.
Based on our isoscape, we generated posterior probability density maps representing spatial assignments for our faunal enamel samples. Most maps usefully show areas with high probability of origin, while also including very small pockets of probability around sampling points that are likely methodological artefacts. E. quagga from MAZ-1, dated to ~12.3 ka cal BP, likely originated from the area today occupied by Afromontane grasslands in the southeastern quadrant of our study area (Fig. 8). Conversely, 25–15 ka cal BP E. quagga from both MAZ-1 and HOR-1 appears to have had a narrower habitat preference along riparian corridors (Fig. 8). E. quagga from MAZ-1, dated to ~3.5 ka cal BP, possibly occupied both the southeastern quadrant and the upper Kasitu River (Fig. 8).The only E. quagga individual yielding large intratooth variation (#67935 from HOR-1, dated to ~16.7 ka cal BP) appears to have ranged along the upper Kasitu during a wetter part of the year and along a different riparian corridor during a drier part of the year (Supplementary Fig. 9). Alcelaphins from HOR-1 have an even more restricted likely provenience, with most posterior probability concentrated along the Luwewya River to the west of the site (Fig. 9). Among the other large bovids, all from MAZ-1, Hippotragus sp. dated to ~18.8 ka cal BP likely originates from the riparian habitats along the upper course of the Kasitu River or the area today occupied by Afromontane grassland around KAD-1; T. strepsiceros dated to ~10 ka cal BP originates from the southeastern quadrant; T. oryx dated to ~6.4 ka cal BP likely originates from a riparian habitat other than the Kasitu (Fig. 10). The likely provenience of reduncin bovids is shown in Supplementary Fig. 10. Some of the equifinality in the density maps can be resolved by considering that these antelopes are unlikely to have ranged far from dependable sources of water25. Thus, both Pleistocene Kobus vardonii from MAZ-1 and Kobus ellipsiprymnus from HOR-1 were likely procured from the upper Kasitu River, while 10–8 ka BP K. vardonii from HOR-1 derived from a secondary riparian corridor or dambo. Among the small bovids, duikers at KAD-1 and HOR-1 appear to have been procured from different localities (Supplementary Fig. 11). Precise proveniences are difficult to determine, but local procurement near the sites cannot be rejected. Klipspringers Oreotragus oreotragus from KAD-1 were likely procured locally near the site, while the provenience of the individuals from HOR-1 is more ambiguous (Supplementary Fig. 13). Equifinality in spatial assignments also affects other small bovids, but in general, local procurement remains the most parsimonious interpretation (Supplementary Fig. 16). 14–10 ka BP Ph. africanus from HOR-1 likely originated from the area that today is Afromontane grassland, while 6.5–0 ka BP individuals from HOR-1, HOR-5 and MAZ-1 all have their most likely origin in riparian corridors other than the Kasitu (Supplementary Fig. 18). Pleistocene Potamochoerus larvatus from HOR-1 also originated from a riparian habitat, but this is more likely to be the upper Kasitu (Supplementary Fig. 18). C. simum dated to 25–15 ka BP from HOR-1 and to 6.5–0 ka BP from MAZ-1 could have been procured from a relatively large area in the southeastern quarter of the study area, with clusters of probability in the upper Kasitu and in the habitats around KAD-1 (Supplementary Fig. 21) Conversely, 10–8 ka BP C. simum from HOR-1 likely inhabited lower elevation areas in the west or northeast of the study area. Using mean faunal 87Sr/86Sr ratios from HOR-1 and MAZ-1 as a proxy for the hunting catchment of each site, it appears that the foragers occupying both sites focused their foraging along the riparian corridors of the Luwewya and Kasitu, as well as the southeast of the study area (Fig. 11).


Posterior probability density maps overlain on satellite imagery of the study area showing the likely provenience of alcelaphins from MAZ-1 and HOR-1. The 25–15 ka map combines multiple individuals, for individual maps see Supplementary Fig. 9. Basemap from ESRI World Imagery.

Posterior probability density maps overlain on satellite imagery of the study area showing the likely provenience of tragelaphines and hippotragins from MAZ-1 and HOR-1. Basemap from ESRI World Imagery.

Posterior probability density maps overlain on satellite imagery of the study area using the mean 87Sr/86Sr ratios from archaeological fauna at HOR-1 and MAZ-1 as a proxy for the main catchment territory of each site. Basemap from ESRI World Imagery.
Discussion
The broad 87Sr/86Sr ranges and the high intra-locality variation recorded in the Kasitu Valley are in accordance with the global observation that regions with higher mean 87Sr/86Sr also have higher variability14. This is likely due to the differential weathering of different components of old rocks, which also weather overall slowly making the contribution of exogenous sources proportionally larger14. This is evident in our samples, where dust has lower 87Sr/86Sr than the plants it was deposited on, suggesting it originated from the erosion of rocks that were not as old and K-rich as local rocks. Since 23 of 25 leaf-dust pairs come from non-riparian or dambo localities and the upslope soils in our study area are acidic, it is also possible that the relatively large contribution of atmospheric Sr to the bioavailable pool is a product of low Sr concentration in those soils26. Intra-locality variability is also comparatively largecf 27, although not as high as the variability of 0.0052 noted by Snoeck et al. in plants from Ireland28. Non-riparian plants yielded a higher mean 87Sr/86Sr than riparian and dambo plants, likely because the latter average across different sources in their watershed. While our plant sample records a very large variability, the true range of bioavailable 87Sr/86Sr in the Kasitu Valley is likely to be even larger than the one captured by our sampling strategy. For instance, two coffee beans from Ludwing–Mzuzu (~39 km northeast from our study area) yielded 87Sr/86Sr lower than our lowest plant sample (0.7131 and 0.7148)29, although this might be influenced by fertilizers. The water of lake Malawi also has 87Sr/86Sr lower than that recorded in our plant sample (0.7166)30.
Our local-scale isoscape is broadly in agreement with a recent continental-scale isoscape computed through random forest regression31. The differences between the two isoscapes are mostly within ±0.01, especially along rivers and in the southeastern quadrant of the study area, whereas larger differences exist in undersampled areas (Supplementary Fig. 22). Although our range of 87Sr/86Sr in plants is greater than those directly measured at most other localities worldwide, an even larger range (0.7054–0.8780; Δ = 0.1726) is recorded in southern Zimbabwe, where plants growing on substrates with Archaean gneisses and massive granites yielded 87Sr/86Sr > 0.7552 (ref. 32). Much lower values are recorded in most regions of southern Africa—for instance the southern Cape (87Sr/86Sr < 0.7242)16,33 and the Maloti-Drakensberg mountains (87Sr/86Sr < 0.7173)34—and the Ca-rich volcanics of eastern Africa—for instance Kenya and northern Tanzania (87Sr/86Sr < 0.7280)15, and Olduvai Gorge (87Sr/86Sr < 0.7048)35. There are exceptions, such as the quartzites, basalts, and shales in the Sterkfontein valley (87Sr/86Sr up to 0.774)13 and the Archaean rocks of the Angola Block (87Sr/86Sr up to 0.7681)36. Trying to make inferences about the past circulation of people, animals, and goods between eastern and southern Africa, e.g.37, it is tempting to use the highly radiogenic 87Sr/86Sr observed in Malawi and Zimbabwe as a relatively strong proxy for provenience. For instance, a single Late Pleistocene ostrich eggshell bead from Sehonghong rock shelter in Lesotho yielded a 87Sr/86Sr of 0.7555 (ref. 34) and may have been imported from southeastern-central Africa. However, it is likely that the recorded 87Sr/86Sr variation across sub-Saharan Africa is biased by research being concentrated around famous paleoanthropological localities and future work will demonstrate more widespread equifinality in radiogenic Sr ratios, as predicted by the continental isoscape31. For instance, modern land snails from the Ntinini Valley in KwaZulu-Natal yielded unexpectedly high 87Sr/86Sr ranging from 0.7245 to 0.7843 (ref. 38). Further assessment of bioavailable Sr ratios will likely expose new areas with elevated 87Sr/86Sr.
Our spatial assignments were successful in revealing some patterns in the distribution of hunting territories of the foragers who inhabited the Kasitu Valley. First, Sr isotopes support the common assumption, e.g.39,40, that small game (here, small bovids) in archaeological assemblages represents short-distance hunting in the immediate vicinity of a site. Larger game (equids, suids, large antelopes) was likely hunted over a broader area, but we do not have evidence of long-distance hunting (i.e., extending beyond the confines of our study area). However, it is possible that some individuals were hunted further away, but their teeth were not transported back to the site. The LGM zebras in our sample were restricted to riparian habitats, while the postglacial and Holocene individuals ranged in the Afromontane grasslands further from rivers. This is consistent with oxygen isotopic evidence from the Kasitu Valley showing that the LGM was arid (Bertacchi et al., in review), thus forcing grazers closer to permanent water sources, while the Holocene was more humid, thus enabling herbivores to range more broadly. Alcelaphins are also bound to riparian environments, as these fresh grass grazers need high-quality forage and would not have survived on sparse, nutrient-poor grasses in the understory of the open woodland (see also Supplementary Text 9). Other large bovids, bushpig, white rhinoceros, and Holocene warthog also point at the importance of riparian habitats as focal points for ungulate hunting. A single 14–10 ka BP warthog may be an outlier in that it seems to originate from the Afromontane grassland at a time when aridity would have favored staying closer to the rivers. However, this individual is not directly dated and could have an age closer to 10 ka, or it could have occupied a small riparian pocket close to MAZ-1.
Today, plains zebra and blue wildebeest migrate long distances in the Serengeti-Mara41, the Kalahari42, and other areas in eastern and southern Africa43,44, but we do not find evidence of migration at any scale in our sample. At most, some of the individuals we sampled show some non-random variation in 87Sr/86Sr that is consistent with seasonal movement within the study area. For instance, a single zebra with relatively large intratooth variation in 87Sr/86Sr could have moved seasonally between the upper Kasitu and another undetermined riparian corridor. We cannot determine with the available data why migratory behavior was absent. This could be due to a lack of necessity, with not enough competition to favor long-distance movement. Alternatively, grassland habitats could have been too discontinuous and fragmented to allow obligate grazers to migrate. A recent 87Sr/86Sr study of Kenyan ungulates 115–12 ka similarly found little evidence of migratory behavior among plains zebra and extant blue wildebeest17. Multi-isotopic studies of faunas 160–40 ka from the southern Cape similarly could not prove migratory behavior16,45. While ours and these two studies are separated by thousands of km and cover different time spans, the evidence as it stands today suggests that Pleistocene African foragers were potentially less mobile than previously thought46, exploited a broad resource base including small game47,48,49,50 and were not specifically focused on migrating herds51. A zooarchaeological study of HOR-1 found broad-spectrum prey selection focusing on small taxa and limited evidence of selective transport of large bovids, both consistent with short-distance hunting4.
In conclusion, we cannot falsify the hypothesis that large game was available year-round “locally”, i.e., within 25–30 km of the sites. Late LGM and postglacial foragers in Malawi were less mobile than previously thought, practicing most of their subsistence activities at the scale of tens, not hundreds, of km. This spatial scale is consistent with the restricted size of the social and mating networks inferred from aDNA evidence7. Here we hint at a potential explanatory mechanism that links the local availability of abundant and predictable resources—large ungulates congregating near water courses—to the restriction of foragers subsistence, social, and mating landscapes, as predicted by behavioral ecological models8,46,52. Future work will expand the current sample of archaeological faunal remains to include periods older than the LGM and clarify the timing of the transition from long- to short-distance interactions. Our isoscape can also be used as a test for the local origin and small life-time mobility of post-LGM foragers suggested by aDNA studies7. Finally, our method can be applied to personal ornaments, particularly ostrich eggshell beads, to test whether the reduced size of the mating and subsistence network extended to other aspects of the social lives of the foragers who inhabited the Kasitu Valley.
Methods
We collected a total of 196 plant samples in the study area (Fig. 1 and Supplementary Text 4), including non-riparian localities (n = 135), non-dambo riparian localities (n = 33) and dambos (n = 28) (see Supplementary Data for plant taxa and details of the localities). Twelve unbroken shells of giant land snails of the family Achatinidae were opportunistically collected, all from non-riparian localities. Plant samples were air-dried in the field and shipped with shells to the Environmental Isotope Paleobiogeochemistry Laboratory of the University of Illinois in Urbana-Champaign (UIUC), where they were prepared as described in Supplementary Text 4. Elemental concentrations of Ca, Sr, Rb, and Ba were measured on dilute aliquots using an iCAP Q Inductively Coupled Plasma-Mass Spectrometer (ICP-MS) (Thermo Fisher) in the Department of Geology at UIUC. Sr was eluted from aliquots of stock solution by cation exchange chromatography using Sr-spec resin (Eichrom) and 87Sr/86Sr was measured using a Nu Instruments High Resolution Multi-Collector Inductively-Coupled-Plasma Mass-Spectrometer (MC-ICP-MS) as described in Supplementary Text 4. Duplicate measurements (n = 6) indicate a measurement precision of ±0.00003.
Fossil teeth were identified by visual comparison with reference specimens from the Yale Peabody Museum of Natural History (CT, USA), as well as published and unpublished atlases. We selected enamel samples as a subset of the samples analyzed for stable carbon and oxygen isotopes and reported in detail in Bertacchi et al. (in review). After the study, fossil specimens were deposited in the National Repository in Nguludi (near Blantyre, Malawi). In total, we analyzed 166 samples from 45 teeth (19 teeth sequentially sampled, 26 teeth bulk sampled from the total length of available enamel). Time-bin assignment (Table 1) is based on direct-dates (Bertacchi et al., in review) and stratigraphic association with dated charcoal, ostrich eggshell and land snail shell7,53,54. Further details are in Supplementary Text 5 and Supplementary Data. Enamel aliquots (~5 mg) were processed either at the Yale Metal Geochemistry Center (YMGC) or in the Department of Earth Science and Environmental Change at UIUC, as detailed in Supplementary Text 5. 87Sr/86Sr was measured either on a Thermo Neptune MC-ICP-MS at YMGC or on a Nu Instruments High Resolution MC-ICP-MS at UIUC. Measurement precision was estimated at ±0.00006 and ±0.000025, respectively (details in Supplementary Text 5).
EBK was computed in ArcGIS Pro v. 3.2.1 using a power variogram, splitting of the entire study area in 250 cells and allowing 10–15 neighbors per cell. Known values having the same coordinates (e.g., 1 m leaves and 2 m leaves from the same plant) were averaged, since the differences are mostly small (Fig. 2). The process was iterated 10,000 times, each time with a subset of 100 measurements. The standard error of the prediction was estimated through a leave-one-out procedure. We generated probabilistic spatial assignments of archaeofaunal remains using the package “assignR” v. 2.4.0 (ref. 55) in R v. 4.3.1 (ref. 56). For details, see Supplementary Text 6.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data used in this paper are included in the Supplementary Data spreadsheet. Additional metadata and raster GeoTiff are accessible in the online repository Zenodo (https://doi.org/10.5281/zenodo.11397396) alongside a copy of the Supplementary Data.
Code availability
The R code used in this paper is available in the online repository Zenodo (https://doi.org/10.5281/zenodo.11397396).
References
van Breugel, P. et al. Potential Natural Vegetation Map of Eastern Africa (Burundi, Ethiopia, Kenya, Malawi, Rwanda, Tanzania, Uganda and Zambia). Version 2.0. (Forest & Landscape Denmark and World Agroforestry Centre (ICRAF), (2015).
Happold, D. & Lock, J. The Biotic Zones of Africa. Mamm. Afr. 1, 57–74 (2013).
Thompson, J. C., Bertacchi, A., Keller, H. M., Hallett, E. Y. & Pobiner, B. The Zooarchaeology of Pleistocene Africa. in Handbook of Pleistocene Archaeology of Africa (eds Beyin, A., Wright, D. K., Wilkins, J. & Olszewski, D. I.) 1955–2087. https://doi.org/10.1007/978-3-031-20290-2_126 (Springer International Publishing, 2023).
Bertacchi, A., Kaliba, P. & Thompson, J. C. Short-distance hunting strategies of Late Quaternary foragers in the miombo woodlands of Malawi. J. Anthropol. Archaeol. 77, 101656 (2025).
Thompson, J. C. et al. Early human impacts and ecosystem reorganization in southern-central Africa. Sci. Adv. 7, eabf9776 (2021).
Clark, P. U. et al. The last glacial maximum. Science 325, 710–714 (2009).
Lipson, M. et al. Ancient DNA and deep population structure in sub-Saharan African foragers. Nature 1–7 https://doi.org/10.1038/s41586-022-04430-9 (2022).
Dyson-Hudson, R. & Smith, E. A. Human territoriality: an ecological reassessment. Am. Anthropol. 80, 21–41 (1978).
Ambrose, S. H. & Lorenz, K. G. Social and ecological models for the Middle Stone Age in southern Africa. in The emergence of modern humans: An archaeological perspective. (ed. Mellars, P.) 3–33 (Edinburgh University Press, 1990).
Wilmsen, E. N. Interaction, spacing behavior, and the organization of hunting bands. J. Anthropol. Res. 29, 1–31 (1973).
Capo, R. C., Stewart, B. W. & Chadwick, O. A. Strontium isotopes as tracers of ecosystem processes: theory and methods. Geoderma 82, 197–225 (1998).
Bentley, A. R. Strontium isotopes from the Earth to the archaeological skeleton: a review. J. Archaeol. Method Theory 13, 135–187 (2006).
Copeland, S. R. et al. Strontium isotope evidence for landscape use by early hominins. Nature 474, 76–78 (2011).
Bataille, C. P., Crowley, B. E., Wooller, M. J. & Bowen, G. J. Advances in global bioavailable strontium isoscapes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 555, 109849 (2020).
Janzen, A. et al. Spatial variation in bioavailable strontium isotope ratios (87Sr/86Sr) in Kenya and northern Tanzania: Implications for ecology, paleoanthropology, and archaeology. Palaeogeogr. Palaeoclimatol. Palaeoecol. 560, 109957 (2020).
Copeland, S. R. et al. Strontium isotope investigation of ungulate movement patterns on the Pleistocene Paleo-Agulhas Plain of the Greater Cape Floristic Region, South Africa. Quat. Sci. Rev. 141, 65–84 (2016).
O’Brien, K. et al. Limited herbivore migration during the Last Glacial Period of Kenya. Nat. Ecol. Evol. 1–8 https://doi.org/10.1038/s41559-024-02413-9 (2024).
Cerezo-Román, J. et al. Detailed recovery methods show the complexity of ancient mortuary practices in later Stone Age hunter-gatherers of Southern-Central Africa. Bioarchaeology Int. https://doi.org/10.5744/bi.2024.0027 (2025).
Gribov, A. & Krivoruchko, K. Empirical Bayesian kriging implementation and usage. Sci. Total Environ. 722, 137290 (2020).
Crader, D. Hunters in Iron Age Malawi: The Zooarchaeology of Chencherere Rockshelter (Department of Antiquities Publication, 1984).
Marks, S. A. Hunting behavior and strategies of the Valley Bisa in Zambia. Hum. Ecol. 5, 1–36 (1977).
DeBusk, G. H. A 37,500-year pollen record from Lake Malaŵi and implications for the biogeography of afromontane forests. J. Biogeogr. 25, 479–500 (1998).
Bertacchi, A. et al. Trace element concentrations as proxies for diagenetic alteration in the African archaeofaunal record: implications for isotope analysis. J. Archaeol. Sci. Rep. 53, 104403 (2024).
Yang, D. et al. Intra-tooth stable isotope analysis reveals seasonal dietary variability and niche partitioning among bushpigs/red river hogs and warthogs. Curr. Zool. zoae007 https://doi.org/10.1093/cz/zoae007 (2024).
Estes, R. D. The Behavior Guide to African Mammals: Including Hoofed Mammals, Carnivores, Primates (Univ of California Press, 2012).
Miller, O. L., Solomon, D. K., Fernandez, D. P., Cerling, T. E. & Bowling, D. R. Evaluating the use of strontium isotopes in tree rings to record the isotopic signal of dust deposited on the Wasatch Mountains. Appl. Geochem. 50, 53–65 (2014).
Knipper, C. et al. Mobility in Thuringia or mobile Thuringians: a strontium isotope study from early Medieval central Germany. Popul. Dyn. Prehistory Early Hist. New Approaches Using Stable Isot. Genet. Berl. Boston Gruyter 5, 287–310 (2012).
Snoeck, C. et al. Towards a biologically available strontium isotope baseline for Ireland. Sci. Total Environ. 712, 136248 (2020).
Rodrigues, C., Máguas, C. & Prohaska, T. Strontium and oxygen isotope fingerprinting of green coffee beans and its potential to proof authenticity of coffee. Eur. Food Res. Technol. 232, 361–373 (2011).
Palmer, M. R. & Edmond, J. M. The strontium isotope budget of the modern ocean. Earth Planet. Sci. Lett. 92, 11–26 (1989).
Wang, X. et al. Strontium isoscape of sub-Saharan Africa allows tracing origins of victims of the transatlantic slave trade. Nat. Commun. 15, 10891 (2024).
House, M., Sealy, J., Chirikure, S. & Roux, P. L. Investigating cattle procurement at Great Zimbabwe using 87Sr/86Sr. J. Afr. Archaeol. 19, 146–158 (2021).
Radloff, F. G. T., Mucina, L., Bond, W. J. & le Roux, P. J. Strontium isotope analyses of large herbivore habitat use in the Cape Fynbos region of South Africa. Oecologia 164, 567–578 (2010).
Stewart, B. A. et al. Ostrich eggshell bead strontium isotopes reveal persistent macroscale social networking across late Quaternary southern Africa. Proc. Natl. Acad. Sci. USA 117, 6453–6462 (2020).
Tucker, L. et al. Initial assessment of bioavailable strontium at Oldupai Gorge, Tanzania: potential for early mobility studies. J. Archaeol. Sci. 114, 105066 (2020).
Wang, X. et al. A bioavailable strontium isoscape of Angola with implications for the archaeology of the transatlantic slave trade. J. Archaeol. Sci. 154, 105775 (2023).
Miller, J. M. & Wang, Y. V. Ostrich eggshell beads reveal 50,000-year-old social network in Africa. Nature 601, 234–239 (2022).
Fowler, K. D., Yang, P. & Halden, N. M. The provisioning of nineteenth century Zulu capitals, South Africa: insights from strontium isotope analysis of cattle remains. J. Archaeol. Sci. Rep. 31, 102306 (2020).
Starkovich, B. M. Optimal foraging, dietary change, and site use during the Paleolithic at Klissoura Cave 1 (southern Greece). J. Archaeol. Sci. 52, 39–55 (2014).
Stiner, M. C. Thirty years on the “Broad Spectrum Revolution” and paleolithic demography. Proc. Natl. Acad. Sci. 98, 6993–6996 (2001).
Anderson, T. M. et al. Interplay of competition and facilitation in grazing succession by migrant Serengeti herbivores. Science 383, 782–788 (2024).
Verlinden, A. Seasonal movement patterns of some ungulates in the Kalahari ecosystem of Botswana between 1990 and 1995. Afr. J. Ecol. 36, 117–128 (1998).
Naidoo, R. et al. A newly discovered wildlife migration in Namibia and Botswana is the longest in Africa. Oryx 50, 138–146 (2016).
Harris, G., Thirgood, S., Hopcraft, J. G. C., Cromsigt, J. P. & Berger, J. Global decline in aggregated migrations of large terrestrial mammals. Endanger. Species Res. 7, 55–76 (2009).
Hodgkins, J. et al. An isotopic test of the seasonal migration hypothesis for large grazing ungulates inhabiting the Palaeo-Agulhas Plain. Quat. Sci. Rev. 235, 106221 (2020).
Marean, C. W. The transition to foraging for dense and predictable resources and its impact on the evolution of modern humans. Philos. Trans. R. Soc. B Biol. Sci. 371, 20150239 (2016).
Clark, J. L. The Howieson’s Poort fauna from Sibudu Cave: documenting continuity and change within Middle Stone Age industries. J. Hum. Evol. 107, 49–70 (2017).
Clark, J. L. The still bay and pre-still bay fauna from Sibudu cave: taphonomic and taxonomic analysis of the macromammal remains from the Wadley excavations. J. Paleolit. Archaeol. 2, 26–73 (2019).
Prendergast, M. E. et al. Small game forgotten: Late Pleistocene foraging strategies in eastern Africa, and remote capture at Panga ya Saidi, Kenya. Quat. Sci. Rev. 305, 108032 (2023).
Jones, M. B. The dik-diks of Guli Waabayo: late Pleistocene net-hunting and forager sociality in eastern Africa. Archaeol. Anthropol. Sci. 15, 203 (2023).
Marean, C. W. Hunter–gatherer foraging strategies in tropical grasslands: model building and testing in the East African Middle and Later Stone Age. J. Anthropol. Archaeol. 16, 189–225 (1997).
Willems, E. P., Hellriegel, B. & van Schaik, C. P. The collective action problem in primate territory economics. Proc. R. Soc. B Biol. Sci. 280, 20130081 (2013).
Skoglund, P. et al. Reconstructing prehistoric African population structure. Cell 171, 59–71.e21 (2017).
Miller, J. M., Keller, H. M., Heckel, C., Kaliba, P. M. & Thompson, J. C. Approaches to land snail shell bead manufacture in the Early Holocene of Malawi. Archaeol. Anthropol. Sci. 13, 37 (2021).
Ma, C., Vander Zanden, H. B., Wunder, M. B. & Bowen, G. J. assignR: an R package for isotope-based geographic assignment. Methods Ecol. Evol. 11, 996–1001 (2020).
R Core Team. R: a language and environment for statistical computing. (2021).
Jenks, G. F. The data model concept in statistical mapping. Int. Yearb. Cartogr. 7, 186–190 (1967).
Acknowledgements
We thank the Malawi Department of Museum and Monuments for granting research permission. Access was facilitated by local authorities Paramount Chief Inkosi ya Makosi M’Mbelwa V, Inkosi Chindi, Inkosi Kampingo Sibande, Inkosana Thomas Nkosi, and the Mzimba Heritage Association. We are grateful to the local community for welcoming us and participating in the fieldwork alongside international students and our long-term Malawian collaborators Joseph Tembo, Henry Kalinga, Daudi Mwangomba, and Samson Kanyika. We thank Menno Welling (African Heritage Ltd. – Research & Consultancy, Zomba, Malawi) for help with field logistics; Noah Planavsky (Yale University) for access to laboratory spaces; Jiuyuan Wang, Boriana Kalderon-Asael, and Alice Knaf (Yale University) for advice on the enamel portion of the project. A grant to S.A., A.Z., and J.C.T. from the National Science Foundation (Grant BCS# 1725123) funded the collection and processing of plant and soil samples for the Kasitu isoscape. Enamel processing was supported by grants to A.B. from the National Science Foundation (Grant BCS# 2211151), the Yale Institute of Biospheric Studies, and the Smith Fund. The broader field project was supported by grants to J.C.T. by the Wenner-Gren Foundation (Grant no. 9437), the National Geographic Society (NGS-53412R-18), Emory University, Yale University, and Hyde Family Foundations.
Author information
Authors and Affiliations
Contributions
Alex Bertacchi: conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, resources, software, visualization, writing—original draft preparation, writing—review and editing. Andrew Zipkin: conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, resources, writing—original draft preparation, writing—Review and editing. Stanley Ambrose: conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, resources, supervision, writing—original draft preparation, writing—review and editing. Beth Scaffidi: formal analysis, writing—original draft preparation, writing—review and editing. Dan Asael: methodology, writing—review and editing. Zachary Benmamoun: investigation, writing—review and editing. Jacob Davis: investigation, writing—review and editing. Jeremy Farr: investigation, writing—review and editing. Matthew Fort: investigation, writing—review and editing. Julia Giblin: methodology, supervision, writing—review and editing. Thomas Johnson: investigation, methodology, resources, writing—review and editing. Potiphar Kaliba: writing—review and editing. Craig Lundstrom: investigation, methodology, resources, writing—review and editing. Fredrick Mapemba: writing—review and editing. Rafael Mwafulirwa: writing—review and editing. Jessica Thompson: conceptualization, funding acquisition, project administration, writing—original draft preparation, resources, writing—review and editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth and Environment thanks Kaedan O’Brien and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Somaparna Ghosh A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Bertacchi, A., Zipkin, A., Ambrose, S. et al. Biogeochemical evidence for targeted landscape use in ancient foragers of Malawi. Commun Earth Environ 6, 818 (2025). https://doi.org/10.1038/s43247-025-02795-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-025-02795-z
