
Abstract
Wildfires are increasing in severity, frequency and size, potentially threatening freshwater species that adapted under different disturbance regimes. However, few wildfire studies have comprehensively evaluated freshwater populations and assemblages following wildfire over broad spatial scales while accounting for post-fire salvage practices in the watershed. We reveal that stream vertebrate assemblages across thirty 4th order streams, spanning a range of both watershed fire severity and post-fire forest management extent, were minimally influenced by immediate effects of fire alone (absent of channel reorganization events). Greater total taxa, total fish, and adult trout densities were associated with streams draining more severely burned watersheds, whereas sculpin, amphibian and crayfish densities appeared uninfluenced by burn severity. Greater extent of post-fire watershed salvage practices was negatively associated with frog densities and positively with young-of-year trout and crayfish densities. Our findings indicate fishes thrive, and amphibians and crayfish persist despite experiencing high-severity megafires in the western Cascades of Oregon.
Similar content being viewed by others
Introduction
A combination of increased temperatures, shifts in precipitation, higher fuel aridity, and prior forest management is resulting in greater frequency, size, intensity, and season lengths of wildfires in many regions globally, including the Pacific Northwest, USA1,2,3. This region has recently experienced megafires which are extreme fire events with high area burned, intensity, and impacts beyond suppression capabilities4,5,6. Wildfire can alter the structure and function of riparian and freshwater ecosystems by influencing a suite of physical and biological habitat variables that control stream vertebrate populations7,8,9,10. Habitat responses can sometimes threaten fish, amphibians and crayfish that are cold-water species or that have adapted under different disturbance regimes. Fire can induce short-term (less than five years) changes in inputs of wood, sediment, ash, nutrients, and organic material, changes to stream food webs11,12 and stream temperatures13 with resulting effects on stream fauna ranging from extirpations to increases in populations12,14,15. However, many aquatic taxa populations may be adapted to fire-induced habitat changes or can withstand a disturbance or recover over time. Responses depend on population connectivity, habitat range and degradation, invasion by non-native species, physiological tolerances and capacity for mitigative behavior, and fire characteristics14,16. Additionally, wildfires followed by high precipitation events or post-fire forest management (e.g., salvage practices such as harvest and replanting) may further alter stream ecosystems, complicating our understanding of effects on stream vertebrates that are due to wildfire alone17,18,19.
Population responses of stream fish, amphibians and crayfishes have exhibited mixed short term responses in temperate forests. Fish populations post-fire have increased14,20, decreased, or had no observable change post-fire15,21,22. Stream breeding amphibian responses to wildfire have received less attention than fish, but observed responses were mixed and variable over time23. Different species and age classes of fish, and potentially also of amphibians, and crayfishes may vary in response to fire-induced changes in physical habitat conditions and resource availability due to varying physiologies, habitat use, or life history strategies24,25. For example, studies on trout species have observed changes in body size, earlier maturity, or faster growth rates after fire24,26,27. Extirpations and population decreases are most commonly due to impacts from subsequent landslides and channel reorganization events post-fire10,28,29, but population declines in systems unaccompanied by post-fire channel reorganization events have also been attributed to elevated stream temperatures causing stressful conditions for biota24,28,30.
Elevated stream temperatures have the potential to influence populations of cold-water adapted species (e.g. salmonids) and stream vertebrate assemblages due to physiological limits31, sublethal effects on behavior32, and competitive interactions that favor species with warmer tolerances33. Stream temperatures 1–3 years post-fire across 31 sites in the Pacific Northwest increased daily maximum temperatures by up to 8.5 °C34. Modelling studies suggest post-fire stream warming can decrease fish populations24,35,36, although empirical studies have demonstrated the persistence of fish populations despite high post-fire stream temperature increases37,38. Further, different age classes may respond differently to warm post-fire temperature conditions as previous work has found differences in age distributions with fewer age 1+ rainbow trout in burned streams24, and higher densities and larger individuals of YOY (young-of-year/age-0) coastal cutthroat trout in burned than unburned streams26,27.
While links between burn severity and streamflow, sediment, and stream temperature are well studied10,19,39, the influences of burn severity on fish, amphibians and vertebrate assemblages are not. Greater increases in summer stream temperatures are associated with greater burn severity28,34. However, most studies simply compare burned sites (any portion of the watershed burned) to unburned sites24,27,40, but where burn severity is evaluated as driver of aquatic biota, watersheds are often categorized as a coarse categorical variable (low and high, or low, moderate, and high severity)12,41, and rarely has watershed-scale fire severity been evaluated as a predictor of aquatic response to fire across many sites (except see Coble et al. 38). Even within vast megafires that encompass entire watersheds, a range of fire severities can occur, and while coarse categories are simple to communicate, they are unlikely to capture variation needed to gage ecological effects39. By using a continuous metric of fire severity that accounts for the range of conditions across watersheds, we can begin to evaluate not only the extent of fire in a watershed but also the role of severity and how this may impact fish, amphibians and assemblages.
Few wildfire studies have comprehensively evaluated freshwater vertebrate populations and assemblages likely reflecting a focus of prior research on headwater systems with limited vertebrate species richness. Stream vertebrate and invertebrate assemblages can serve as indicators of ecological change with shifts in composition (e.g., increased percentage of tolerant taxa) observed in response to stressors such as pollutants, loss of habitat complexity, or alterations in watershed hydrology42,43,44, but stream vertebrate assemblages have seldom been evaluated in response to wildfire. Evaluations of responses to wildfire in the Pacific Northwest region of North America have primarily focused on socio-economically important salmonids or other target species reflecting their imperiled status. This emphasis on salmonids overlooks other critical ecosystem responses including stream assemblage composition and less commonly evaluated fish species, amphibians and crayfish, that may have important ecosystem functions14,40,45.
Post-fire harvest followed by state-required replanting (hereafter “salvage practices”), are common management activities that can also strongly alter ecosystems, yet little empirical data exists evaluating potential influences on aquatic ecosystems and biota46,47. Salvage practices aim to: 1) extract economically valuable timber and encourage vegetation re-establishment by replanting desired species and 2) enhance soil stability and provide shade for more rapid thermal recovery48. While these concepts are often described as rationales for post-fire riparian management (active vs. natural trajectory of recovery), others have argued salvage practices delay or prevent natural recovery by damaging soils, increasing runoff and erosion, and removing shade and wood that would have contributed to complex stream habitat49,50,51. However, experimental evaluation of hypothesized responses of habitat variables and stream vertebrates is lacking in the scientific literature46,47, so the persistence of stream vertebrate assemblages and populations after post-fire management practices remains unclear.
To evaluate effects of watershed-scale fire severity and extent of salvage practices in the watershed on freshwater vertebrate assemblages and densities, we examined fish, amphibians, and crayfish in thirty 4th-order sites in the first three years following megafires in the western Cascades of Oregon in 2020 (Fig. 1). Streams in this region can contain fishes (salmonids, dace, sculpin, and lamprey), amphibians (salamanders and frogs), and crayfishes that are often ignored in many salmonid focused ecological studies. The lack of landslides and channel reorganization events in our focal watersheds following these fires52 allowed us to evaluate effects of wildfire unconfounded by effects of post-fire channel reorganization events. We expected that within the first three years post-fire: 1) the structures of fish, amphibian and crayfish assemblages are driven by burn severity and exhibit more interannual variability in more severely burned watersheds (as has been observed for macroinvertebrates53); 2) lower fish, amphibian and crayfish densities are associated with warmer more severely burned systems and greater extent of salvage practices in the watershed; and 3) size of young-of-the-year (YOY) trout would be greater in more severely burned systems with warmer stream temperatures. With the anticipated increases in fire size and severity as climate continues to change54, understanding stream ecosystem responses to wildfire is essential to inform risks to biodiversity and listed species especially in light of the paucity of research exploring the influence of differing post-fire management strategies.

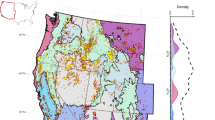
Study sites in the Riverside, Beachie Creek and Holiday Farm Fires (black outlines, listed from north to south) in western Oregon. Watershed burn severity (mean watershed basal area mortality, %) for each site is shown with a red color gradient in the watershed boundary and the extent of salvage practices in the watershed (%) is represented by the size of blue circles at the site location. 24 of the 30 sites were sampled in 2021, 2022 and 2023. McRae Creek (unburned) was only sampled in 2022 and 2023, and Cedar, Sardine, Copper, NF Gate and SF Gate Creeks were only sampled in 2023. Basemap from ESRI109.
Results
Stream temperature
Mean weekly maximum stream temperature (MWMT, °C) represents the hottest week of stream temperatures with a value for each site each year. MWMTs across all sites in the study (including unburned references) ranged from 15.3 to 27.4 °C during the first three summers post-fire (2021–2023). Greater MWMTs were strongly positively correlated with greater watershed burn severity each year and trends were similar all three years (Fig. 2). Unburned reference sites were generally below 20 °C, whereas sites with greater than 40% watershed burn severity were greater than 20 °C. Sites with 5-40% watershed burn severity were highly variable, with the greatest MWMT reaching 27.4 °C (Coal, 30% burn severity).

Stream temperatures (MWMTs, °C) were positively correlated with watershed burn severity (mean watershed basal area mortality, %) in the first three years post-fire. There were 21, 25 and 29 sites for 2021, 2022 and 2023, respectively.
Vertebrate and crayfish assemblages
We evaluated the effects of watershed fire severity and extent of salvage practices on stream vertebrate assemblages and taxa densities and body sizes (see Methods). We captured and identified 18 species of fishes, amphibians and crayfish that we grouped into 11 taxa. The most abundant vertebrate taxa were coastal giant salamanders (Dicamptodon tenebrosus) closely followed by YOY trout (Fig. 3, Table S1). YOY trout are comprised of both native coastal cutthroat trout (Oncorhynchus clarkii clarkii) and native rainbow or steelhead trout (Oncorhynchus mykiss) because we cannot distinguish between species for YOY individuals in the field. We refer to age 1+ rainbow and steelhead trout as O. mykiss because we cannot distinguish between resident rainbow trout and anadromous steelhead in the field. Nonnative brook trout (Salvelinus fontinalis) were the least abundant taxa (only two individuals total). Nonnative juvenile coho (Oncorhynchus kisutch) salmon were found at 2 of 30 sites and juvenile Chinook salmon (Oncorhynchus tshawytscha) were found at 1 of 30 sites. Speckled dace (Rhinichthys osculus) and longnose dace (Rhinichthys cataractae) and Western River lamprey (Occidentis ayresii) and Pacific lamprey (Entosphenus tridentatus) had limited presence (2 of 30 sites) and were more common at lower elevations except at Cook where both speckled and longnose dace species were present despite the relatively high elevation (Fig. 3). Frogs, which were mainly comprised (96%) of coastal tailed frogs (Ascaphus truei) but also a few northern red-legged frogs (Rana aurora), Cascades frogs (Rana cascadae), western toads (Anaxyrus boreas), and Pacific chorus frogs (Pseudacris regilla), were present at 26 sites and more common at higher elevation sites. Sculpin spp. (Cottus beldingii, Cottus rhotheus) and crayfish (Pacifastacus leniusculus) were more common at lower elevation sites.

Relative densities of fish, amphibian, and crayfish taxa at each site in the first 3 years post-fire, 2021–2023. Sites (shown and labeled in Fig. 1) are ordered from low to high burn severity and labeled with site name and burn severity (mean watershed basal area mortality, %). YOY coastal cutthroat trout and YOY Oncorhynchus mykiss are represented in YOY trout, and coastal cutthroat trout and Oncorhynchus mykiss taxa refer to age 1+ individuals. Brook trout and Coho salmon are (included in the YOY salmon taxa) are non-native.
Based on our Non-metric multidimensional scaling (NMDS) analysis of the relationships between stream vertebrate assemblages and environmental variables (Fig. 4b, Table S1), stream temperature (which is influenced by both landscape variables and fire) and variables unrelated to fire or post-fire forest management were the strongest drivers of assemblage structure. Vertebrate assemblages were most strongly correlated with stream gradient (channel steepness, r = 0.50, p = 0.001), stream temperature (r = 0.44, p = 0.001), followed by watershed area (r = 0.43, p = 0.001), elevation (r = 0.38, p = 0.001), and pre-fire stand age (r = 0.31, p = 0.001). Wildfire severity and extent of salvage practices were slightly less correlated with assemblage structure than variables related to landscape position or pre-fire conditions (extent of salvage practices (r = 0.34, p = 0.001), watershed burn severity (r = 0.29, p = 0.001), canopy cover (r = 0.22, p = 0.001)). Other habitat variables related to wood and sediment were not strongly correlated with assemblages (Table S1).

NMDS ordinations indicating the structure of stream assemblages of fish, amphibians, and crayfish species across all study sites over three years in western Oregon, USA (n = 79). a Significantly correlated environmental variables (p < 0.005) are indicated by vectors where the vector length represents the Pearson’s correlation (r) between quantitative variables along ordination axes. b Significantly correlated species (p < 0.005) are indicated by vectors where the vector length represents the Pearson’s correlation (r) between species abundances along ordination axes. c Vectors indicating assemblage shifts over time within sites.
The relative abundances of YOY trout were most closely related to greater stream temperature, but also correlated with lower elevation, greater watershed burn severity and greater extent of salvage practices (Fig. 4a, b). Relative abundances of the dace species (speckled and longnose dace) were positively associated with stream temperature, watershed burn severity, extent of salvage practices, and negatively associated with elevation. Coastal tailed frogs were most strongly positively associated with higher elevation and greater pre-fire stand age. Age 1+ coastal cutthroat trout most aligned with greater canopy cover and higher elevation, although at the two highest elevation sites (Hugh and Norhorn) we did not detect age 1+ coastal cutthroat trout (but did detect age 1 + O. mykiss). Sculpin spp. were closely aligned with greater watershed area.
Variation in assemblage structure among sites was generally greater than interannual variation within sites suggesting relative stability over time across the gradient of post-fire and salvaged conditions in the first three years post-fire (Fig. 3c). Shifts in assemblages from 2021 to 2022 were relatively consistent, trending in the negative direction along NMDS axis 2 (Fig. 4c). This reflects, in part, shifts in relative abundance toward fewer YOY trout in 17 of 24 sites between these two sampling years (Figs. 3 and 4a). Changes from 2022 to 2023 were more variable but trended positively along both axes, with assemblages at 13 of 24 sites shifting towards those more typical of sites that were warmer or more severely burned (Fig. 4b). Despite these within-site trends, sites were relatively stable over time (within years 1–3 post-fire) when considered in context of variation in assemblage structure across sites. Further, yearly variation (quantified by Bray-Curtis dissimilarity) was not related to watershed burn severity (Fig. S1).
Taxa densities
Linear mixed effects models identified at least one significant predictor for each taxa, with the most important predictor varying among taxa (Fig. 5). Evaluating the three post-fire years collectively, watershed burn severity was significantly (95% confidence interval did not overlap 0) positively associated with densities of total taxa, all fish, and native adult trout but was not included in top ranked models for other taxa densities. This is likely in part because less common taxa were not present at each site, so the analysis for taxa subgroups often represented less than 30 sites (Fig. 5). The extent of salvage practices in the watershed was significantly positively associated with YOY trout and crayfish densities but negatively associated with frog densities, and was not included in best models for total taxa, total fish, native adult trout, sculpin, and salamanders (Fig. 5). Watershed burn severity and extent of salvage practices had similar effects on taxa biomass as were observed for densities (Fig. S3).

Predictors of taxa densities (abundance per wetted area) included: mean watershed burn severity, extent of salvage practices in the watershed, mean bankfull width and elevation. Taxa densities are shown in Fig. S2. Predictors were considered significant when 95% CI did not overlap 0.
Bankfull width was included in top models for total taxa, total fish, native adult trout, YOY trout, and salamander densities, but was not significant for total taxa and total fish densities. Bankfull width was significantly negatively associated with native adult trout, YOY trout, salamander densities. Elevation was significantly positively associated with native adult trout, salamander and frog densities and negatively associated with sculpin and crayfish densities (Fig. 5).
YOY trout size
Mean YOY trout weight was significantly positively correlated with MWMT (°C) in each of the first three summers post-fire (2021-2023) (Fig. 6). Mean weights and correlations with stream temperatures were similar all three years with the strongest correlation in 2023 (R2 = 0.25, p = 0.009). The two heaviest individual mean trout weights were at sites with moderate stream temperatures within the range observed at our sites.

YOY trout mass (g) was positively correlated with stream temperatures (MWMT, °C). There were 21, 25 and 29 sites with stream temperature data available and YOY trout present for 2021, 2022 and 2023, respectively.
Discussion
We evaluated the effects of watershed burn severity and extent of salvage practices in the watershed on stream vertebrate and crayfish assemblages and densities using a spatially extensive dataset for the first three years immediately following three 2020 megafires in western Oregon. The lack of channel reorganizing events in these systems following these fires allowed us to evaluate wildfire effects unconfounded by post-fire channel reorganization events which are known to affect fish and amphibian densities in other regions25,55,56,57. Assemblages were correlated with stream temperature, watershed burn severity and extent of salvage practices, however elevation and watershed area were also strongly correlated with assemblages making the influences of fire and post-fire management difficult to disentangle from landscape position variables within this study. Contrary to our expectations, burn severity was not associated with yearly changes in stream vertebrate assemblage structure over time, nor did burn severity negatively affect biota for any taxa group. Rather, burn severity was positively associated with total taxa densities and fish densities, but did not affect amphibian or crayfish densities. Extent of salvage practices was associated with greater densities of YOY trout and crayfish, but lower densities of frogs. Lastly, as expected, larger YOY trout were found in warmer post-fire sites. Results from this observational post-fire study suggest that fishes appear to thrive and amphibians and crayfish persist in the three years after megafires in western Oregon.
Despite stream temperatures that often exceeded 20 °C and reached up to 27.4 °C, warmer sites with greater burn severity had higher fish populations and larger YOY trout in the first three years post wildfire, and stream temperature was a strong predictor of assemblage structure. A global review found mixed fish density responses to fire10, but our findings agree with other recent research in our study region27,37. Although direct mortality due to immediate temperature increases during wildfire may be rare19, sub-lethal effects can result from stressful conditions and changes in behavior and metabolic demand58,59,60.
We hypothesize that greater food availability may aid in supporting high densities of fish, amphibians and crayfish in warm burned systems where metabolic demand is elevated. In early years after fire, elevated light from reduced canopy cover may drive greater autotrophic production11,12 and macroinvertebrate production41,61,62. Shifts towards greater autotrophic resources in the stream food web have been observed post-fire via isotopic studies11,18. Greater food availability during the warm summer conditions may help accommodate increased summer metabolic demand, but greater food resources (and potentially greater consumption) may also be available during other seasons.
We found that high stream temperatures were linked with greater YOY trout size, and greater YOY trout densities were found in systems with greater extent of watershed salvage practices. Other post-fire studies have also found larger juvenile fish in warmer conditions26,27,63. While changes in food availability may be an important factor, we suspect that this finding is at least in part due to earlier emergence of YOY trout in warmer sites which is well documented for poikilothermic organisms. Greater densities and larger sizes of YOY trout cohorts observed in the first 3 post-fire years could potentially carry over into adult populations, warranting further investigation of long-term fish population responses to megafires. Our study was unique as it focused on higher order, larger stream systems than many fire studies, which resulted in the vertebrate community including fish species beyond salmonids such as lamprey, sculpin and dace. Lamprey were only found at the two lowest elevation sites (Coal and Clear), so we were not able to evaluate effects of wildfire or salvage practices on lamprey. Sculpin were found at 19 of the 30 sites, but did not seem to be influenced by burn severity or extent of salvage practices. While sculpin are often present in sites in other published post-fire studies, their results are rarely specified. However, they may respond differently to riparian disturbances due to differences in foraging, microhabitat requirements, and thermal preferences than salmonids64. Dace were found at only 5 of the 30 sites but contributed substantially to total fish and total taxa densities at 3 of the sites. Despite dace having warmer thermal tolerances than most other species at these sites, with so few sites with dace present and no pre-fire data, we cannot clearly demonstrate whether their density is associated with fire. Much of the findings about dace responses to fire have been from the southwestern US where post-fire monsoons are primary controls on stream fish assemblages55,65. Further research is warranted on lamprey, sculpin, and dace responses, and how additional species influence stream ecosystem function in changing conditions.
Crayfish have received less attention in wildfire research than fish or amphibians. One post-fire study found no change in abundance of non-native Virile crayfish (Faxonius virilis) to a wildfire in New Mexico55, while another in Australia observed declines in abundance of four native species post-fire compared to pre-fire66. As omnivores, crayfish responses to disturbance may be important for understanding aquatic food webs postfire as they utilize resources from more than one trophic level and contribute to detrital and periphyton processing via consumption and bioturbation67,68,69. However, documentation of crayfish responses to fire is relatively limited.
In the Pacific Northwest, most work on post-wildfire amphibian responses has focused on coastal tailed frogs which are a species of concern in Oregon. Coastal tailed frogs have low temperature tolerances and life-history traits considered to confer vulnerability to disturbance such as low vagility, long larval period, delayed sexual maturity and low fecundity70,71. Of the frog species we collected, 96% were coastal tailed frogs, of which 90% were larval or metamorphic stages (e.g., aquatic). Although declines in tadpole abundances in the two years following fire were shown in northwestern Montana with the closely related Rocky Mountain tailed frog72; tadpoles in our study were not negatively associated with fire severity despite coastal tailed frogs being generally restricted to stream temperatures below 16⁰C73,74, while Rocky Mountain tailed frogs were found at post-fire sites with maximum stream temperatures up to 26.6 °C in Idaho28. As with fish, under increased stream temperatures and therefore greater metabolic demand, food availability may also be an important driver of frog population persistence post-fire especially for tadpoles which graze on periphyton that can also increase post-fire11,12.
Terrestrial salamanders (often plethodontid) are often negatively affected by wildfire in part because of their lungless anatomy and terrestrial habitat needs23,75, but less is known about fire effects on stream-living salamanders like those in this study that depend on aquatic habitats for most or all of their developmental stages. Our findings indicated a lack of influence of fire severity or salvage practices on stream dwelling salamander densities, which were comprised of coastal giant salamanders (Dicamptodon tenebrosus). In contrast, greater burn severity led to lower populations of adult male long toed salamanders (Ambystoma macrodactylum) in Montana, although a pond breeding species76. A replicated before-after control-impact study at three sites in the Holiday Farm Fire (one of the wildfires examined in our study) and three reference sites just outside of the fire boundary reported small declines in coastal giant salamander densities due to fire27 and a 38-year salamander dataset from Mack Creek, OR, which is a similar size stream to those in the present study, shows a drastic decline in coastal giant salamander densities the first year after the 2023 Lookout fire (I. Arismendi, personal communication). Salamander responses to fire are complicated by low capture probabilities, high error on population estimates and their metamorphosis to terrestrial life stages. However, understanding salamander responses is important for stream ecosystem function because they are often more abundant and have greater biomass than salmonid fishes in headwaters77.
We documented the complete species assemblages of stream vertebrates and crayfish for three years post-fire in thirty 4th order streams in the Pacific Northwest. Prior work in the region related to wildfire effects has focused mainly on how salmonid populations respond to fire14,20,24. The lack of focus on non-salmonid taxa may be, in part, because post-fire research has primarily focused on lower order, higher elevation streams near headwaters10 where few other fishes are present and assemblages are more simple78,79. However, large wildfires which have occurred frequently in the past several years can encompass more of the landscape80,81 including higher order systems which host more species, and often species with potentially both higher thermal and lower water quality tolerances82.
Varying life history strategies or habitat requirements can result in species-specific responses to stream warming83,84,85, and post-fire community composition may shift to an increase in relative abundance of disturbance-adapted taxa8,61,86. However, in our study, fire severity did not supersede effects of non-fire related variables. We also did not observe substantial post-fire shifts (quantified by assemblage dissimilarity) in assemblages in sites with greater burn severity. Factors characterizing landscape position that are fixed (gradient, elevation, watershed size) were important controls on assemblage structure in our models and are not influenced by wildfire or post-fire management. Stream temperature, also an important predictor of assemblage structure, is influenced by both landscape position and wildfire/management factors, and as such are difficult to separate from fire-related variables39.
In the first three years after wildfire, we expected habitat and stream vertebrate assemblages in more severely burned stream systems to change more than less burned or unburned systems, due to recovery from disturbance. While this difference has been observed for macroinvertebrate assemblages53, we did not find vertebrate assemblages in more burned systems to be more dynamic among the first three years post-fire (Fig. 4c). However, this may be at least in part due to the lack of large precipitation events87,88 or landslides in these watersheds52 in the first three years of this study which may have limited changes to physical habitat conditions. Channel reorganization events primarily occur while the landscape is less stable in the first six months to three years post-fire28,86,89,90,91 but landslides and geomorphic events could still occur in these systems in the near future with unknown implications for stream vertebrates. Fire followed by flooding may lead to greater effects on vertebrate and crayfish assemblages in montane streams20,62 therefore considering geomorphic context92 and distinguishing wildfire effects from wildfire-induced geomorphic events on stream ecosystems is critical.
Conceptual reviews have suggested salvage practices may have negative effects on freshwater ecosystems46,47, but empirical data are limited. Salvage extent was positively associated with YOY trout densities for which the effect of burn severity was also positive, suggesting a potential link to warmer stream temperatures or to increased habitat area from potentially increased flows. We did not observe negative associations between extent of salvage practices and fish, crayfish, or salamanders. Although, frog densities (found at 26 of the 30 sites) were negatively associated with greater extent of salvage practices in the watershed and also positively associated with elevation. Salvage extent across our sites was negatively related to elevation38 as private lands in this region are located at low to mid elevation with federal lands more commonly at high elevations.
Lower frog densities in streams do not necessarily indicate lower populations of frogs in the watersheds because of complicating factors influencing timing of their metamorphosis and persistence in their terrestrial life stage. Tadpoles undergo metamorphosis earlier in response to a myriad of environmental factors93. Elevation, which was positively associated with frogs in our study, has been shown to affect tadpole timing of metamorphosis and growth94. For example, coastal tailed frog tadpoles had longer larval periods in higher elevations and increased growth rates with increasing elevations in northern California94. Similarly, presence of adult coastal tailed frogs is complicated by elevation, breeding and other movement patterns as well as habitat type95,96. Further research disentangling elevation and salvage effects and including evaluation of all life stages and presence in aquatic and terrestrial environments is required to understand how the extent of salvage practices influences frog populations.
Our study was limited to the first three years after the wildfire, and salvage effects may become more apparent after longer timescales, such as decades. Systems with salvage practices are likely to experience alternate recovery trajectories of riparian regeneration resulting in potentially different large wood and other terrestrial inputs to streams46,48. Additionally, a lack of an effect of extent of salvage practices on vertebrates may be in part due to the lack of landslides and channel reorganization events in these watersheds52, which likely limited effects of erosion and channel reorganization previously found to be the result of salvage harvest49,50,51,97. Our findings are supported by a recent study that also observed persistence of salmonids in high densities one year after severe wildfire followed by intensive salvage practices but no winter flooding37. Other work has demonstrated that the extent of salvage practices in watersheds is correlated with watershed burn severity, and that smaller-diameters of large wood that tend to predominate in salvaged watersheds38 also depend on disturbance history46. However, there is still very limited specific information on the influence of salvage practices on aquatic ecosystems47, and therefore, studies with pre- and post-fire data and larger sample sizes and including multiple years of population data are needed to begin to address the influence of salvage practices on stream vertebrates and crayfish.
Conclusions
Despite concern, especially for cold-water adapted species, our findings show that fishes thrived, and amphibians and crayfish persisted in post-fire and post-fire salvaged sites in the western Cascade Range in Oregon within the first three years post-fire. Greater total fish, amphibian and crayfish densities were found in sites with greater fire severity. We did not detect effects on vertebrates due to salvage practices in the watershed, with the exception of frogs whose densities were associated with elevation and extent of watershed salvage practices, but frog populations were dominated by tadpoles whose presence may be complicated by metamorphosis, as timing and tadpole growth can vary with elevation. While our study includes a large number of sites and post-fire conditions, the long-term influences and dynamic aspects of wildfire and post-fire forest management on aquatic ecosystems remain uncertain and must be considered in combination with the evolutionary mechanisms of native species in these systems. These findings expand our understanding of the persistence of freshwater fauna after fire and salvage practices separate from the often-confounding effects of flooding and geomorphic changes.
Methods
Site description and study design
Our study was conducted in the moist conifer forests on the west slope of the Cascade Range in Oregon, USA. Forest composition across these locations included, but were not limited to, Douglas-fir (Pseudotsuga menziesii), red alder (Alnus rubra), western red cedar (Thuja plicata), bigleaf maple (Acer macrophyllum), and western hemlock (Tsuga heterophylla). This region has a Mediterranean climate with long dry summer periods. In the three years of the study period annual precipitation was 1790 mm, 2165 mm and 1617 mm for water years 2021, 2022 and 2023 respectively at Cougar Dam a monitoring station near the Holiday Farm Fire (USC00351914, https://www.ncdc.noaa.gov/cdo-web/). Mean annual precipitation was 1873 mm from WY2007-WY2022. The combination of a rapid drying period and strong easterly-wind event in 2020 allowed wildfires to burn 340,000 ha in western Oregon, with almost as much forest area burned within 48 hours as in the last 30 years and at high severities98. We selected three of these wildfires in Oregon: the Beachie Creek, Riverside, and Holiday Farm Fires (Fig. 1). These fires spanned federal, state and private ownerships, wholly encompassing a range of pre- and post-fire forest management strategies.
In our study, we extend initial results from a prior study38 to include multiple years of vertebrate assemblages and effects of watershed fire severity and extent of salvage practices in the watershed on specific taxa densities. Our study evaluated sites consisting of 40 m vertebrate sampling reaches of thirty 4th order streams (Strahler stream order, Strahler 1957), as defined by National Hydrography Dataset V2 Plus, at the base of watersheds within 6 km of the three fire boundaries (Fig. 1, Table S2). In 2021, twenty-four 4th order fish-bearing stream reaches were selected as sites using a stratified random sampling design within 6 km of the footprint of the Riverside, Beachie Creek and Holiday Farm fires to ensure selected sites encompassed a range of pre-fire watershed stand age and watershed fire severity (including unburned sites where watershed burn severity equals 0) (see Coble et al. 38). We added an additional unburned site (McRae) in 2022, and we added five sites with high watershed burn severity (defined as greater than 50% mean watershed basal area mortality as noted below) in 2023, the latter to better represent sites with high burn severity and greater extent of salvage practices in the watershed. We sampled each site (stream reaches within each watershed) between 5 July and 24 August each year (2021: 7/13-8/23, 2022: 7/5-8/19, 2023: 7/6-8/24). The only exception was Elkhorn in 2021, which we sampled on 13 September because stream temperatures in July and August exceeded permitted sampling levels for fish.
Watershed burn severity
To evaluate the influence of burn severity on freshwater responses, we summarized percent basal area mortality at the watershed level. We used percent basal area mortality (%) data from the Rapid Assessment of Vegetation Condition after Wildfire (RAVG) program from the U.S. Department of Agriculture Forest Service Geospatial Technology and Applications Center (https://burnseverity.cr.usgs.gov/ravg/data-access). This is a satellite derived product that provides an immediate assessment of fire effects on vegetation after wildfires, reporting multiple metrics at 30 meter pixels including basal area mortality. We calculated mean raster values of basal area mortality (%) values across all pixels within each watershed area and refer to this metric as watershed burn severity (%).
Watershed salvage practices
We defined the extent of salvage practices in the watershed as the area within a watershed that was harvested post-wildfire for merchantable timber and then replanted. Salvage harvest is generally completed during the first-year post-fire to avoid wood decay that reduces wood quality for market. In our study area, most salvage harvest began in 2020 and continued through 2022. Stands that are salvage harvested have merchantable timber remaining post-fire, so are generally older than 30 years. Tree removal for other management actions such as road safety (e.g. hazard trees to prevent road and vehicle damage) or culvert damage prevention are not included in a salvage harvest estimate although they may have consequential short- and long-term effects on riparian and stream ecosystems, especially when modifications occur near streams. In Oregon, replanting is required after salvage harvest99, so all salvaged area was immediately replanted, and we refer to the collective treatment as watershed salvage practices.
We received information on the area within our watersheds with salvage practices from federal, state, and private forest landowners as the most up-to-date record of area of land salvaged within their ownership that intersected our watersheds. For the five sites added in 2023, we did not receive information directly from two landowners, so we obtained area salvaged and replanted from Oregon Department of Forestry’s Forest Activity Electronic Reporting and Notification System (FERNs; https://ferns.odf.oregon.gov/E-Notification/). Although FERNs data may over or under-estimate actual salvaged and replanted area, this was the best available information for these areas.
Stream temperature
Stream temperature was recorded throughout each summer (2021–2023) at 15-minute intervals using sensors (miniDOT, Precision Measurement Engineering, Vista, CA, with accuracy of ± 0.1 °C) placed in the center of the stream on the bed covered in polyvinyl chloride housing at the downstream end of the vertebrate sampling reaches at each site. We report stream temperatures summarized as mean weekly maximum stream temperature (MWMT, C°) for each year at each site.
Stream vertebrates and crayfish
Locations of sampling reaches were selected from the base (pour point) of each watershed selected by the burn severity criteria (mentioned above). At each site, we conducted triple-pass depletion electroshocking surveys and collected fish, aquatic amphibians, and crayfish in two adjoining 20-m reaches with no space in between. We deployed a block net at the upper and lower end of each reach to prevent movement of individuals in or out of the sample area. Each captured individual was anesthetized (buffered MS-222), weighed (wet weight; nearest 0.1 g) and measured for total length (nearest 1 mm), and snout-to-vent length for amphibians (nearest 1 mm). Individuals were allowed to recover for a minimum of one hour from anesthetization in buckets and then returned to their stream habitat within the sampling reach. We calculated population estimates and 95% confidence intervals using maximum likelihood estimates100 in the Fisheries Stock Analysis package (Ogle 2022) in R (R Core Team 2021).
Sculpin species can be difficult to distinguish, so we combined all of them into Cottus spp. (sculpin) taxa. YOY Oncorhynchus mykiss and Oncorhynchus clarkii clarkii (Coastal cutthroat trout) are difficult to distinguish visually in the field. Therefore, we combined these species into young-of-the-year trout (YOY trout). Within frog species, individuals were noted as tadpoles, adults, or froglet. We combined all frog species in tadpole, froglet and adult forms together into the frog taxa for population estimates. The vertebrate assemblage analysis uses species level groupings and the population estimates are done for the taxa groupings. We calculated taxa biomass (g) estimates by multiplying population estimates with mean weight (g) of each taxa or age class. We normalized abundance and biomass estimates by wetted area (wetted width * reach length) to calculate density (individuals m−2) and biomass (g m−2) estimates. We estimated total taxa density and biomass per area by adding population estimates from all taxa and age groups. We averaged YOY trout lengths (mm) and weights (g) at each site.
Covariates
We collected covariates at each site (stream reach or watershed where the metric is at the watershed level) to evaluate correlations with the vertebrate assemblages. Vertebrate sampling was performed in 40 meters of stream reach at the downstream end of 100 m stream reaches, so additional reach length for covariates was upstream. Covariates included: mean canopy cover (%, collected via a spherical densiometer measured every 20 meters), photosynthetically active radiation (PAR, mol m−2 s−1, collected every 20 meters with a mini PAR (Precision Measurement Engineering, Vista, CA) logging at 1-minute intervals held vertically facing upstream for a minimum of 4 minutes), mean watershed pre-fire stand age (yrs), stream gradient (extracted from NHD High Res for the stream segment), means of bankfull widths and wetted widths (m, collected every 20 m to the nearest 0.1 m), stream depths (collected from 5 equidistant locations along each wetted width), sediment size as d50 (mm), percent embeddedness (%, collected via visual estimations of substrate embeddedness), large wood was measured for each reach as total count for the given 100 m of stream reach (n), diameter (m), and volume (m3). Further detailed methods of covariate data collection can be found in a previous study evaluating these systems38.
Statistical methods
Stream temperature
We evaluated yearly relationships between watershed burn severity (%) and summer mean weekly maximum stream temperatures (MWMT, °C) with linear models and used the p-value and R2 statistics to evaluate correlations. Stream temperature data from 2021 were missing at three sites (Elkhorn, Coal, and Ella) and in 2023 data were missing from one site (Sardine).
Assemblage analysis
To examine relationships in vertebrate and crayfish assemblage structure across watersheds over three years, we used a matrix of relative abundances of all vertebrate and crayfish species. We used non-metric multidimensional scaling (NMDS) to display assemblage Bray-Curtis dissimilarity. We removed taxa present in less than 5% of the samples from the analysis. We evaluated species and covariates correlated with assemblage structure. We compared correlations between all covariates including watershed burn severity and extent of watershed salvage practices (all numeric) with commonly occurring taxa. We transformed covariates for normality with arc sine transformation for variables that are percentages (canopy cover, watershed burn severity, watershed salvage practices and proportion of fine sediment) or log 10 for PAR and pre-fire stand age. We conducted all multivariate analyses using the vegan package in R101.
We also evaluated Bray-Curtis dissimilarity (a measure of β diversity) of assemblages between years versus watershed burn severity by quantifying the Bray-Curtis distances from the same sites over different years (2021, 2022, 2023) for sites with three years of data (24 sites). Using the Bray-Curtis distances between points of the same site for 2021 to 2022, 2022 to 2023 and sum of the Bray-Curtis distances from 2021 to 2022, 2022 to 2023 and 2021 to 2023, we used linear regression to evaluate the effect of watershed burn severity on dissimilarity of sites over time.
Taxa densities
We quantified the effects of watershed burn severity and extent of salvage practices in the watershed on stream vertebrate and crayfish population densities. We expected landscape variables such as stream size and position, measured here as bankfull width and elevation (of the stream reach sampling location), to be strong controls on vertebrate densities102,103,104 and therefore also accounted for them in our statistical model along with our main variables of interest (watershed burn severity and extent of watershed salvage practices). We did not include stream temperature in the statistical model because our goal was to evaluate effects of watershed burn severity and extent of salvage practices on biota, and including stream temperature would confound effects of burn severity because stream temperatures are affected by fire13,105,106. We compared linear mixed effects models of all possible combinations of models with independent terms for watershed burn severity, extent of watershed salvage practices, elevation and bankfull width as fixed effects and year (2021, 2022, or 2023) as a random effect. We used the corrected Akaike Information Criterion to select top ranked models to predict densities of each taxa using the AICcmodavg package in R107. While the extent of watershed salvage practices and watershed burn severity are correlated because salvage practices can only occur after fire, we evaluate both variables independently. Within linear models, a correlation between fixed effect estimates does not result in biased estimates but can result in less precision of estimates only if very strongly correlated and inclusion of interaction terms between correlated variables can lead to multicollinearity potentially confounding the individual contributions of the variables108. Using the top ranked model, we extracted the standardized model coefficients of the variables. Explanatory variables were standardized for comparability such that the distributions have a mean equal to zero and a standard deviation equal to one. Therefore, one standard deviation of the effect size of the coefficient is equal to 27.5% for watershed burn severity, 3.7 m for bankfull width is 148.1 m for elevation, and 10.9% for extent of watershed with salvage practices area. We evaluated effect sizes of the explanatory variables by assessing whether the 95% confidence intervals around the model estimates overlapped zero.
YOY trout size
We evaluated yearly relationships between summer mean weekly maximum stream temperatures (MWMT °C) and mean YOY trout size with linear models and used the p-value and r2 statistics to evaluate correlations. We use the mean weight of individuals at each site to indicate size.
Data availability
All data supporting the findings of this study are published online at the Oregon State University library archive at https://doi.org/10.7267/rx914027s.
References
Jolly, W. M. et al. Climate-induced variations in global wildfire danger from 1979 to 2013. Nat. Commun. 6, 7537 (2015).
Abatzoglou, J. T. et al. Projected increases in western US forest fire despite growing fuel constraints. Commun. Earth Environ. 2, 1–8 (2021).
Jones, M. W. et al. Global and Regional Trends and Drivers of Fire Under Climate Change. Rev. Geophys. 60, e2020RG000726 (2022).
Linley, G. D. et al. Megafire’—You May Not Like It, But You Cannot Avoid It. Glob. Ecol. Biogeogr. 34, e70032 (2025).
Tedim, F. et al. Defining extreme wildfire events: Difficulties, challenges, and impacts. Fire 1, 9 (2018).
Williams, J. et al. Findings and implications from a coarse-scale global assessment of recent selected mega-fires. in Proceedings of the 5th International Wildland Fire Conference, Sun City, South Africa 9–13 (2011).
Minshall, G. W., Brock, J. T. & Varley, J. D. Wildfires and Yellowstone’s Stream Ecosystems. Biosci 39, 707–715 (1989).
Mihuc, T. B. & Minshall, G. W. The trophic basis of reference and post-fire stream food webs 10 years after wildfire in Yellowstone National Park. Aquat. Sci 67, 541–548 (2005).
Pettit, N. E. & Naiman, R. J. Fire in the riparian zone: Characteristics and ecological consequences. Ecosyst 10, 673–687 (2007).
Erdozain, M., Cardil, A. & de-Miguel, S. Fire impacts on the biology of stream ecosystems: A synthesis of current knowledge to guide future research and integrated fire management. Glob. Chang. Biol. 30, e17389 (2024).
Spencer, C. N., Gabel, K. O. & Hauer, F. R. Wildfire effects on stream food webs and nutrient dynamics in Glacier National Park, USA. For. Ecol. Manag. 178, 141–153 (2003).
Cooper, S. D. et al. Physicochemical and biological responses of streams to wildfire severity in riparian zones. Freshw. Biol. 60, 2600–2619 (2015).
Koontz, E. D., Steel, E. A. & Olden, J. D. Stream thermal responses to wildfire in the Pacific Northwest. Freshw. Sci. 37, 731–746 (2018).
Dunham, J. B., Young, M. K., Gresswell, R. E. & Rieman, B. E. Effects of fire on fish populations: Landscape perspectives on persistence of native fishes and nonnative fish invasions. For. Ecol. Manag. 178, 183–196 (2003).
Rieman, B., Gresswell, R. & Rinne, J. Fire and fish: A synthesis of observation and experience. USDA Forest Service - General Technical Report RMRS-GTR 159–175 (2012).
Jager, H. I. et al. Resilience of terrestrial and aquatic fauna to historical and future wildfire regimes in western North America. Ecol. Evol. 11, 12259–12284 (2021).
Burton, T. A. Fish and stream habitat risks from uncharacteristic wildfire: Observations from 17 years of fire-related disturbances on the Boise National Forest, Idaho. For. Ecol. Manag. 211, 140–149 (2005).
Silins, U. et al. Five-year legacy of wildfire and salvage logging impacts on nutrient runoff and aquatic plant, invertebrate, and fish productivity. Ecohydrology 7, 1508–1523 (2014).
Gomez Isaza, D. F., Cramp, R. L. & Franklin, C. E. Fire and rain: A systematic review of the impacts of wildfire and associated runoff on aquatic fauna. Glob. Change Biol. 28, 2578–2595 (2022).
Rieman, B. E. & Lee, D. Does wildfire threaten extinction for salmonids: responses of redband trout and bull trout following recent large fires on the Boise National Forest. Proceedings - Fire Effects on Rare and Endangered Species and Habitat Conference 16, 47–57 (1997).
Jones, R. D., Carty, D. G., Gresswell, R. E., Hudson, C. J. & Mahony, D. L. Fishery and aquatic management program in Yellowstone National Park. US Fish and Wildlife Service, Technical Report (1981).
Jakober, M. J. A Summary of Fish Habitat and Population Changes in Flat Creek, Upper Selway River Drainage One Year after the 2000 Wilderness Complex Fires, Bitterroot National Forest. A summary of fish habitat and population changes in Flat Creek, Upper Selway River Drainage one year after the 2000 Wilderness Complex fires, Bitterroot National Forest [Bitterroot National Forest] https://doi.org/10.5962/BHL.TITLE.124546, (2002).
Hossack, B. R. & Pilliod, D. S. Amphibian responses to wildfire in the western united states: Emerging patterns from short-term studies. Fire Ecol 7, 129–144 (2011).
Rosenberger, A. E., Dunham, J. B., Neuswanger, J. R. & Railsback, S. F. Legacy effects of wildfire on stream thermal regimes and rainbow trout ecology: an integrated analysis of observation and individual-based models. Freshw. Sci. 34, 1571–1584 (2015).
Hedden, C., Propst, D. L., Gido, K. B., Hedden, S. C. & Whitney, J. E. Differential Responses of Native Fishes in Two Headwater Tributaries of the Gila River Following Severe Wildfires. West. N. Am. Nat. 82, 201–207 (2022).
Heck, M. P. Effects of Wildfire on Growth and Demographics of Coastal Cutthroat Trout in Headwater Streams. (Oregon State University, 2006).
Swartz, A. & Warren, D. Wildfire in western Oregon increases stream temperatures, benthic biofilms, and juvenile coastal cutthroat trout size and densities with mixed effects on adult trout and coastal giant salamanders. https://doi.org/10.1139/CJFAS-2022-0053 (2022).
Dunham, J. B., Rosenberger, A. E., Luce, C. H. & Rieman, B. E. Influences of wildfire and channel reorganization on spatial and temporal variation in stream temperature and the distribution of fish and amphibians. Ecosyst 10, 335–346 (2007).
Lyon, J. P. & O’connor, J. P. Smoke on the water: Can riverine fish populations recover following a catastrophic fire-related sediment slug? Austral Ecol 33, 794–806 (2008).
Sestrich, C. M., McMahon, T. E. & Young, M. K. Influence of fire on native and nonnative salmonid populations and habitat in a western Montana basin. Trans. Am. Fish. Soc. 140, 136–146 (2011).
Pörtner, H. O. & Farrell, A. P. Physiology and climate change. Science 322, 690–692 (2008).
Meeuwig, M. H., Dunham, J. B., Hayes, J. P. & Vinyard, G. L. Effects of constant and cyclical thermal regimes on growth and feeding of juvenile cutthroat trout of variable sizes. Ecol. Freshw. Fish 13, 208–216 (2004).
McMahon, T. E., Zale, A. V., Barrows, F. T., Selong, J. H. & Danehy, R. J. Temperature and competition between bull trout and brook trout: a test of the elevation refuge hypothesis. Trans. Am. Fish. Soc. 136, 1313–1326 (2007).
Beyene, M. T. & Leibowitz, S. G. Heterogeneity in post-fire thermal responses across Pacific Northwest streams: A multi-site study. J. Hydrol. 23, 100173 (2024).
Beakes, M. P., Moore, J. W., Hayes, S. A. & Sogard, S. M. Wildfire and the effects of shifting stream temperature on salmonids. Ecosphere https://doi.org/10.1890/ES13-00325.1 (2014)
Roon, D. A. et al. Linking Fire, Food Webs, and Fish in Stream Ecosystems. Ecosyst 28, 1–22 (2025).
Warren, D. R., Roon, D. A., Swartz, A. G. & Bladon, K. D. Loss of riparian forests from wildfire led to increased stream temperatures in summer, yet salmonid fish persisted. Ecosphere 13, e4233 (2022).
Coble, A. A., Penaluna, B. E., Six, L. J. & Verschuyl, J. Fire severity influences large wood and stream ecosystem responses in western Oregon watersheds. Fire Ecol 19, 34 (2023).
Chen, J. & Chang, H. A review of wildfire impacts on stream temperature and turbidity across scales. Prog. Phys. Geogr. 47, 369–394 (2023).
Preston, D. L. et al. Short-term effects of wildfire on high elevation stream-riparian food webs. Oikos 2023, e09828 (2023).
Malison, R. L. & Baxter, C. V. The fire pulse: Wildfire stimulates flux of aquatic prey to terrestrial habitats driving increases in riparian consumers. Can. J. Fish. Aquat. Sci. 67, 570–579 (2010).
Whittier, T. R., Hughes, R. M., Lomnicky, G. A. & Peck, D. V. Fish and Amphibian Tolerance Values and an Assemblage Tolerance Index for Streams and Rivers in the Western USA. Trans. Am. Fish. Soc. 136, 254–271 (2007).
Lücke, J. D. & Johnson, R. K. Detection of ecological change in stream macroinvertebrate assemblages using single metric, multimetric or multivariate approaches. Ecol. Indic. 9, 659–669 (2009).
Hellawell, J. M. Biological Indicators of Freshwater Pollution and Environmental Management. (Springer Science & Business Media, 2012).
Neville, H., Dunham, J., Rosenberger, A., Umek, J. & Nelson, B. Influences of Wildfire, Habitat Size, and Connectivity on Trout in Headwater Streams Revealed by Patterns of Genetic Diversity. Trans. Am. Fish. Soc. 138, 1314–1327 (2009).
Reeves, G. H., Bisson, P. A., Rieman, B. E. & Benda, L. E. Postfire Logging in Riparian Areas. Conserv. Biol. 20, 994–1004 (2006).
Barrett, S. W. & Reilly, M. Effects of salvage logging on riparian zones in coniferous forests of eastern Washington and adjacent regions. Washington State Forest Practices Adaptive Management Program CMER# 17–100, (2017).
Lindenmayer, D. B. & Noss, R. F. Salvage Logging, Ecosystem Processes, and Biodiversity Conservation. Conserv. Biol. 20, 949–958 (2006).
Beschta, R. L. et al. Postfire management on forested public lands of the western United States. Conserv. Biol. 18, 957–967 (2004).
Lindenmayer, D. B. et al. Salvage Harvesting Policies After Natural Disturbance. Science 303, 1303–1303 (2004).
Karr, J. R. et al. The Effects of Postfire Salvage Logging on Aquatic Ecosystems in the American West. Biosci 54, 1029–1033 (2004).
Selander, B. D. et al. Inventory of debris flows in burned (2020-2022) and unburned (1995-2020) areas in the western Cascade Range of Oregon. https://doi.org/10.5066/P13TPP8J (2024).
Minshall, G. W., Robinson, C. T., Lawrence, D. E., Andrews, D. A. & Brock, J. T. Benthic macroinvertebrate assemblages in five central Idaho (USA) streams over a 10-year period following disturbance by wildfire. Int. J. Wildland Fire 10, 201–213 (2001).
Higuera, P. E. & Abatzoglou, J. T. Record-setting climate enabled the extraordinary 2020 fire season in the western United States. Glob. Chang. Biol. 27, (2021).
Whitney, J. E., Gido, K. B., Pilger, T. J., Propst, D. L. & Turner, T. F. Consecutive wildfires affect stream biota in cold- and warmwater dryland river networks. Freshw. Sci. 34, 1510–1526 (2015).
Gido, K. B. et al. Pockets of resistance: Response of arid-land fish communities to climate, hydrology, and wildfire. Freshw. Biol. 64, 761–777 (2019).
Reale, J. K. et al. Differential effects of a catastrophic wildfire on downstream fish assemblages in an aridland river. Aquat. Ecol. 55, 483–500 (2021).
Schulte, P. M. The effects of temperature on aerobic metabolism: towards a mechanistic understanding of the responses of ectotherms to a changing environment. J. Exp. Biol. 218, 1856–1866 (2015).
Chabot, D., McKenzie, D. J. & Craig, J. F. Metabolic rate in fishes: definitions, methods and significance for conservation physiology. J. Fish Biol. 88, 1–9 (2016).
Fitzgerald, A. M., John, S. N., Apgar, T. M., Mantua, N. J. & Martin, B. T. Quantifying thermal exposure for migratory riverine species: Phenology of Chinook salmon populations predicts thermal stress. Glob. Chang. Biol. 27, 536–549 (2021).
Minshall, G. W. Responses of stream benthic macroinvertebrates to fire. For. Ecol. Manag. 178, 155–161 (2003).
Malison, R. L. Effects of wildfire on aquatic insect assemblages, emerging adult insects, and riparian consumers in a wilderness watershed. (Idaho State University, 2008).
Armstrong, J. B. et al. The importance of warm habitat to the growth regime of cold-water fishes. Nat. Clim. Chang. 11, 354–361 (2021).
Benjamin, J. R. et al. Pathways of productivity and influences on top consumers in forested streams. For. Ecol. Manag. 508, 120046 (2022).
Whitney, J. E., Gido, K. B., Pilger, T. J., Propst, D. L. & Turner, T. F. Metapopulation analysis indicates native and non-native fishes respond differently to effects of wildfire on desert streams. Ecol. Freshw. Fish. 25, 376–392 (2016).
Johnston, K., Matthews, T., Robson, B. & Chester, E. Impacts of extreme events on southeastern Australian freshwater crayfish. Freshw. Crayfish 20, 61–72 (2014).
Parkyn, S. M., Collier, K. J. & Hicks, B. J. New Zealand stream crayfish: functional omnivores but trophic predators? Freshw. Biol. 46, 641–652 (2001).
Momot, W. T., Gowing, H. & Jones, P. D. The dynamics of crayfish and their role in ecosystems. Am. Midl. Nat. 10, 35 (1978).
Usio, N. & Townsend, C. R. Functional significance of crayfish in stream food webs: roles of omnivory, substrate heterogeneity and sex. Oikos 98, 512–522 (2002).
Hossack, B. R. & Honeycutt, R. K. Declines revisited: Long-term recovery and spatial population dynamics of tailed frog larvae after wildfire. Biol. Conserv. 212, 274–278 (2017).
Corn, P. S., Bury, R. B. & Hyde, E. J. Conservation of North American stream amphibians. in 24–36 (2003).
Hossack, B. R., Corn, P. S. & Fagre, D. B. Divergent patterns of abundance and age-class structure of headwater stream tadpoles in burned and unburned watersheds. Can. J. Zool. 84, 1482–1488 (2006).
Welsh, H. H. & Lind, A. J. Multiscale Habitat Relationships of Stream Amphibians in the Klamath-Siskiyou Region of California and Oregon. J. Wildl. Manage. 66, 581–602 (2002).
Pilliod, D. S., Bury, R. B., Hyde, E. J., Pearl, C. A. & Corn, P. S. Fire and amphibians in North America. For. Ecol. Manag. 178, 163–181 (2003).
Gade, M. R., Gould, P. R. & Peterman, W. E. Habitat-dependent responses of terrestrial salamanders to wildfire in the short-term. For. Ecol. Manag. 449, 117479 (2019).
Hossack, B. R., Lowe, W. H., Honeycutt, R. K., Parks, S. A. & Corn, P. S. Interactive effects of wildfire, forest management, and isolation on amphibian and parasite abundance. Ecol. Appl. 23, 479–492 (2013).
Bury, R. B., Corn, P. S., Aubry, K. B., Gilbert, F. F. & Jones, L. L. Aquatic Amphibian Communities in Oregon and Washington. (1991).
Rieman, B. & Clayton, J. Wildfire and Native Fish: Issues of Forest Health and Conservation of Sensitive Species. Fisheries 22, 6–15 (1997).
Rieman, B. et al. Status of native fishes in the western United States and issues for fire and fuels management. For. Ecol. Manag. 178, 197–211 (2003).
Flannigan, M. D., Krawchuk, M. A., De Groot, W. J., Wotton, B. M. & Gowman, L. M. Implications of changing climate for global wildland fire. Int. J. Wildland Fire 18, 483–507 (2009).
Burke, M. et al. The changing risk and burden of wildfire in the United States. Proc. Natl. Acad. Sci. USA. 118, (2021).
Huff, D. D., Hubler, S. L. & Borisenko, A. N. Using Field Data to Estimate the Realized Thermal Niche of Aquatic Vertebrates. N. Am. J. Fish. Manag. 25, 346–360 (2005).
Eaton, J. G. & Scheller, R. M. Effects of climate warming on fish thermal habitat in streams of the United States. Limnol. Oceanogr. 41, 1109–1115 (1996).
Isaak, D. J. et al. Effects of climate change and wildfire on stream temperatures and salmonid thermal habitat in a mountain river network. Ecol. Appl. 20, 1350–1371 (2010).
Ebersole, J. L., Quiñones, R. M., Clements, S. & Letcher, B. H. Managing climate refugia for freshwater fishes under an expanding human footprint. Front. Ecol. Environ. 18, 271–280 (2020).
Verkaik, I. et al. Fire as a disturbance in mediterranean climate streams. Hydrobiologia 719, 353–382 (2013).
Bush, S. A., Johnson, S. L., Bladon, K. D. & Sullivan, P. L. Stream chemical response is mediated by hydrologic connectivity and fire severity in a Pacific Northwest forest. Hydrol. Process. 38, e15231 (2024).
Kang, H., Cole, R. P., Miralha, L., Compton, J. E. & Bladon, K. D. Hydrologic responses to wildfires in western Oregon. USA. J. Hydrol. 639, 131612 (2024).
Minshall, G. W., Brock, J. T., Andrews, D. A. & Robinson, C. T. Water quality, substratum and biotic responses of five central Idaho (USA) streams during the first year following the Mortar Creek fire. Int. J. Wildland Fire 10, 185–199 (2001).
Vieira, N. K. M., Clements, W. H., Guevara, L. S. & Jacobs, B. F. Resistance and resilience of stream insect communities to repeated hydrologic disturbances after a wildfire. Freshw. Biol. 49, 1243–1259 (2004).
Romme, W. H. et al. Twenty Years After the 1988 Yellowstone Fires: Lessons About Disturbance and Ecosystems. Ecosyst 14, 1196–1215 (2011).
Montgomery, D. R. Process domains and the river continuum. JAWRA 35, 397–410 (1999).
Nizam, K. S., Ismail, N. A., Farinordin, F. A. & Izam, N. A. M. Frog Metamorphosis: A Review of Metamorphic Stages, Developmental Progression, and Influential Factors. Bioresources and Environment 1, 120–155 (2023).
Macedo, A. D., Demianew, J. A., Garwood, J. M. & Reiss, J. O. Larval Life History of Coastal Tailed Frogs (Ascaphus truei) Across an Elevational Gradient in Northern California: Implications for a Changing Climate. Journal of Herpetology 57, 52–59 (2023).
Matsuda, B. M. & Richardson, J. S. Movement patterns and relative abundance of coastal tailed frogs in clearcuts and mature forest stands. Canadian Journal of Forest Research 35, 1131–1138 (2005).
Hayes, M. P. et al. Dispersion of Coastal Tailed Frog (Ascaphus Truei): An hypothesis relating occurrence of frogs in non–fish-bearing headwater basins to their seasonal movements. Journal of Herpetology 40, 531–543 (2006).
Beschta, R. L. et al. Recommendations for ecologically sound postfire salvage management and other postfire treatments on federal lands in the west. (1995).
Reilly, M. J. et al. Cascadia Burning: The historic, but not historically unprecedented, 2020 wildfires in the Pacific Northwest. USA. Ecosphere 13, e4070 (2022).
OAR 629-610-0020 – Reforestation Stocking Standards. https://oregon.public.law/rules/oar_629-610-0020.
Carle, F. L. & Strub, M. R. A New Method for Estimating Population Size from Removal Data. Biometrics 34, 621 (1978).
Oksanen, J. et al. vegan: Community Ecology Package. (2024).
Waite, I. R. & Carpenter, K. D. Associations among Fish Assemblage Structure and Environmental Variables in Willamette Basin Streams, Oregon. Trans. Am. Fish. Soc. 129, 754–770 (2000).
Grenouillet, G., Pont, D. & Hérissé, C. Within-basin fish assemblage structure: the relative influence of habitat versus stream spatial position on local species richness. Can. J. Fish. Aquat. Sci. 61, 93–102 (2004).
Smith, T. A. & Kraft, C. E. Stream Fish Assemblages in Relation to Landscape Position and Local Habitat Variables. Trans. Am. Fish. Soc. 134, 430–440 (2005).
Rhoades, C. C., Entwistle, D. & Butler, D. The influence of wildfire extent and severity on streamwater chemistry, sediment and temperature following the Hayman Fire, Colorado. Int. J. Wildland Fire 20, 430–442 (2011).
Sanders, A. M. et al. Heat and smoke from wildfires influence water temperature and dissolved oxygen levels in headwater streams. Freshw. Sci. 41, 665–679 (2022).
Mazerolle, M. J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). (2023).
Schielzeth, H. et al. Robustness of linear mixed-effects models to violations of distributional assumptions. Methods Ecol. Evol. 11, 1141–1152 (2020).
ESRI. World Topographic Map. (2012).
Acknowledgements
We thank funding partners, including NCASI, the Pacific Northwest Research Station, U.S.D.A. Forest Service, the U.S. Environmental Protection Agency, and Weyerhaeuser Co. We thank D. Leer, B. Laufer, T. Archer, W. Arthurs, A. Borquez, M. Dirling, T. Harron, J. Ikoma, W. Kiridena, R. Kuemmerlin, C. Lindelien, H. Lossiah, R. Ragsdale, S. Richardson, A. Salyer, M. Weaver, J. White and A. Wilson for assistance in the field and laboratory and Benthic Aquatic Research Services for macroinvertebrate identification. We thank forest landowners for providing land access and assistance: Bureau of Land Management, Campbell Global, LLC, Giustina Land and Timber, Giustina Resources, Oregon Department of Forestry, Port Blakely, Rayonier, United States Department of Agriculture Forest Service, and Weyerhaeuser Company. We thank D. Miller and J. Verschuyl for providing constructive comments that improved the manuscript. Scientific take permits: NOAA permit #24235, #26102, #26942, ODFW 097-21, OFW 097-22, ODFW 102-23, USFWS TE74340D-0.
Author information
Authors and Affiliations
Contributions
A.C., B.P. contributed to conceptualization, methodology and project administration. A.S. performed formal analysis and writing of the original draft. A.C., B.P., R.F., J.E., and M.K. contributed to conceptualization, funding acquisition and review and editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
The views expressed in this article are those of the author(s) and do not necessarily represent the views or policies of the U.S. Environmental Protection Agency.
Peer review
Peer review information
Communications Earth and Environment thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editors: Mengjie Wang. [A peer review file is available].
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Swartz, A.G., Coble, A.A., Penaluna, B.E. et al. Following megafires fishes thrive and amphibians persist even in severely burned watersheds. Commun Earth Environ 6, 945 (2025). https://doi.org/10.1038/s43247-025-02893-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-025-02893-y
