
Abstract
Many European lakes have suffered oxygen declines from recent eutrophication raising the question as to which conditions are representative of their pre-impact state. Here we present a palaeoecological record revealing that deepwater oxygen in small lakes can be variable over long timescales and subject to human influence for millennia. Remains of chironomids and other invertebrates indicate that Bichelsee (Switzerland) shifted from a stable, oxygen-rich to a dynamic, oxygen-poor state before major human activity. Initially, increased human activity and clearcutting coincided with improved oxygen conditions for deepwater midge larvae. However, from the Roman Age onwards, similar disturbances were associated with low oxygen and reduced deepwater invertebrate populations. These results demonstrate that small European lakes can have multiple, contrasting pre-impact reference states, and that ecological effects of human activity, even their directionality, may shift over time. Furthermore, repeated cycles of impact and recovery can occur over millennia, complicating assumptions about their baseline conditions.
Similar content being viewed by others

Introduction
Lake ecosystems are important biodiversity hotspots, freshwater reservoirs and provide a wide range of ecosystem services such as habitats for endangered species, recreational areas and access to fisheries1. However, many lakes have been heavily impacted and modified in past decades and centuries by human activities leading to eutrophication and associated anoxia with detrimental effects on their flora and fauna2,3. To counter this, major efforts have been made to slow down or reverse eutrophication and restore European lakes4,5. When assessing the extent that lake ecosystems have been disturbed by human activities, theoretical or reconstructed reference conditions play an important role to determine the deviation of affected ecosystems from a state with minimal human impacts. For example, the European Water Framework Directive (WFD), the primary environmental legislature for European lakes, assesses ecological status of aquatic ecosystems relative to a hypothetical reference state defining their conditions under minimal human impact6. The concept of natural state also plays an important role in more general ecological discussions on the extent that modern ecosystems deviate from their undisturbed, pristine state and have been modified by human impact7.
Palaeoenvironmental reconstructions based on sediment analyses provide information on past conditions in aquatic ecosystems and can therefore play a key role in discussions of pre-impact and reference states of impacted lakes8 and marine ecosystems9,10. Lake sediment studies focussing on environmental change during the recent past, e.g., during 20th century eutrophication or acidification, often assume that human impact was negligible or not relevant before the industrial period (i.e., 1850 CE)11. However, in regions like Central Europe, agriculture and other human activities have had distinct influences on the environment since the Neolithic period12. Individual studies have shown that such early human activities may already have had noticeable effects on lake ecosystems13,14,15. However, long-term and particularly prehistoric human impacts are rarely considered in discussions on natural background conditions and reference states of lakes. Remains of chironomid larvae and other aquatic invertebrates are sensitive indicators for reconstructing lake ecosystem dynamics and particularly changes in deepwater oxygen concentrations. The presence of benthic invertebrates such as chironomids in deepwater environments of lakes requires sufficient oxygen concentrations for their survival and therefore represents an important indicator of lake ecological status. For example, in Switzerland, the Water Protection Ordinance (WPO) prescribes that oxygen concentrations at any given water depth throughout the year should be at minimum 4 mg/L, a threshold considered necessary to allow benthic invertebrates to exist, unless unusual natural conditions favour anoxic waters16.
With this study we present a well-dated and exceptionally highly resolved aquatic invertebrate record from a small Swiss lowland lake (Bichelsee). Remains of chironomid larvae and other invertebrates in this record allow us to reconstruct major variations in invertebrate assemblages and deepwater oxygen availability, determine when deepwater hypoxia first developed, describe the dynamics of changes in deepwater environments and assess how early phases of hypoxia compare with the most recent change in oxygen due to 20th century eutrophication. Our results indicate a stable oxygen-rich phase in the early and mid-Holocene and a more dynamic, hypoxic phase after the expansion of beech (Fagus sylvatica) and closing of forests ca. 7100 years ago. Phases of human activities initially led to improved conditions for deepwater invertebrates, presumably caused by the opening of local forests, but from the Roman Age onwards were associated with more extreme phases of hypoxia. Our results challenge the concept of stable, oxygen-rich pre-impact and reference states for small Central European lowland lakes. They indicate that for such lakes, multiple pre-impact states may be possible, that hypoxia can develop well before noticeable local human impact and that phases of increased human activity can have both positive and negative effects on the deepwater fauna, prevailing oxygen conditions and ecological status.
Results
Chronology and sediment core lithology
The Bichelsee sediment record covers the last 13,500 years. With 72 radiocarbon dates and 22 samples analysed for 210Pb and 137Cs (Fig. 1; Supplementary Fig. 1; Supplementary Data), it is one of the best dated lake sediment records in southwestern Central Europe allowing a detailed comparison of our results with archaeological and historical evidence. The precise dating also enables us to calculate influx values (remains deposited per year and cm2) of chironomid and other aquatic invertebrate remains. In contrast to percentages, which reflect changes in the relative abundances of different taxonomic groups, influx can track changes in absolute abundances of sensitive indicator groups, such as deepwater taxa, and therefore provides additional information on past changes in the Bichelsee fauna.

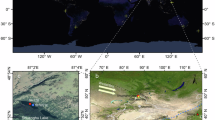
A Elevation map of Switzerland showing the location of Bichelsee (red dot) and close-up map showing the vegetation height around Bichelsee and the streams (drawn from data of the Federal Office of Topography swisstopo). B Aerial photo of Bichelsee (Photo credit: René Thoma, r). C Age-depth model calculated for the Bichelsee sediment core analysed in the present study. Black dots indicate the calibrated radiocarbon ages, the red line shows the most likely age-depth relationship and the grey area the 95 % age confidence interval. A detailed version of the age-depth relationship with calibrated age probability curves of individual 14C dates is available in the online supporting material (Supplementary Table 1 and Supplementary Fig. 1).
Chironomids and aquatic invertebrates
Subfossil remains of a large range of aquatic invertebrates preserve well in lake sediments and can be analysed to reconstruct past invertebrate assemblage composition through time17. Our study focuses on chitinous remains larger than 100 μm, which include head capsules and mouthparts of aquatic insect larvae, statoblasts of bryozoans, exoskeletons of oribatid mites and ephippia of water fleas (see Online Methods). At Bichelsee, 257 samples were analysed for aquatic invertebrate remains of which chironomids were the most abundant with 9266 remains compared to 7720 other aquatic invertebrate remains. Chironomid larvae are typically the most abundant macroinvertebrate group in lakes18 with many indicator taxa sensitive to changes in trophic state and oxygen availability19,20. Deepwater chironomids, mainly Tanytarsus lugens-type and Chironomus anthracinus-type, dominated most of the record with exception of the youngest section (340 calibrated 14C years BP (cal. BP) to present) where a more diverse chironomid fauna was present (Supplementary Fig. 2). Chironomid and overall aquatic invertebrate records clearly show two distinct phases in the development of Bichelsee. The first stable phase (13,500–7100 cal. BP), dominated by deepwater chironomids (average: 95 %; Supplementary Fig. 2) with some occurrences of Chaoborus flavicans-type, Daphnia and Plumatella (Supplementary Fig. 3), is typical for lakes with high deepwater oxygen availability21. The second much more dynamic phase (7100 cal. BP to present) is characterised by variable but overall lower chironomid percentages (Fig. 2A), a situation usually encountered in lakes with hypoxic deepwater environments21. During the first stable phase, Tanytarsus lugens-type, typical of oxygen-rich deepwater environments of oligo- to mesotrophic lakes22, dominated the chironomid assemblages (average: 66 %). In the second, dynamic phase, the abundance of Tanytarsus lugens-type was very variable but generally lower (average: 24 %). Chironomus anthracinus-type joined and sometimes replaced Tanytarsus lugens-type as dominant taxon (average: 31 %). This latter taxon, starting to increase from ca. 6500 cal. BP, is typically found in lakes with seasonal oxygen depletion23. The co-occurrence of Tanytarsus lugens-type and Chironomus anthracinus-type indicates the coexistence of deepwater environments with both relatively high (e.g., in the upper profundal) and low oxygen concentration (e.g., in the deepest parts) during summer stratification. The dynamic phase of the record was also characterised by high rates of change between adjacent chironomid samples. This suggests that chironomid assemblages in Bichelsee were extremely variable during this time interval (Supplementary Fig. 2), although changes in sedimentation rates, bioturbation and associated changes in temporal resolution may have also contributed to the high between-sample variability (Fig. 2B). Although oxygen concentrations were overall low in this phase, there were nevertheless major decadal- to centennial-scale changes in oxygen availability for benthic invertebrates. Percentages of non-chironomid remains increased from relatively low values before ca. 7100 cal. BP (average: 5 %) to relatively high values after ca. 7100 cal. BP (average: 56 %) suggesting lower deepwater oxygen availability21. Particularly, remains of planktonic chaoborids, a group of invertebrates able to profit from deepwater hypoxia and often dominating invertebrate assemblages under low deepwater oxygen concentrations24, increased ca. 7100–5000 cal. BP (average: 13 %; Supplementary Fig. 3). At the same time, chironomid percentages remained consistently low (average: 30 %; Supplementary Fig. 2) suggesting a long phase with oxygen depleted deepwater at Bichelsee. From ca. 5000 cal. BP, percentages of chironomids became highly variable (11–84 %) implying decadal- to centennial-scale variability of deepwater oxygen in the lake. This is also supported by passively plotting downcore samples from the Bichelsee record in a Canonical Correspondence Analysis (CCA) of modern aquatic invertebrate assemblages of small Swiss lakes covering a wide range of elevation, nutrient and deepwater oxygen concentrations21. In this analysis, CCA axis 1 closely correlates with summer deepwater oxygen concentration21 and the axis 1 scores of the Bichelsee invertebrate samples confirm the high variability of oxygen availability in this part of the record (Fig. 2C).

A Percentages of chironomids relative to all aquatic invertebrate remains. B Log-transformed chironomid influx at high resolution (grey) and with a 9-sample running average (black). C Scores of the first axis of a CCA calculated as in ref. 21 in orange and log-transformed Chaoborus-to-chironomid ratio in blue24. D Rates of change between the chironomid samples. E Selected tree pollen percentages (Alnus glutinosa-type, Fagus sylvatica). The vertical shading shows the three major phases of oxygen availability in Bichelsee (oxygen-rich, hypoxic and variable hypoxic).
Chironomid influx values, available at higher temporal resolution than percentage data, reveal high abundances of chironomid larvae at the coring location in the early Holocene (8400–7600 cal. BP; average: 4.7 remains cm−2 yr−1) followed by a dramatic decrease ca. 7100 cal. BP (0.3 remains cm−2 yr−1), also apparent in the percentage data (Fig. 2A, B). During the following dynamic phase, aquatic invertebrate assemblage percentages changed regularly but the chironomid influx stayed relatively low (average: 0.5 remains cm−2 yr−1) with some few exceptions with higher values at ca. 3300 cal. BP (ca. 1.4 remains cm−2 yr−1), 500 cal. BP (ca. 8.0 remains cm−2 yr−1) and 90–50 years ago (ca. 1.3 remains cm−2 yr−1). When influx values of deepwater and littoral chironomid taxa are examined separately during this dynamic phase, it becomes apparent that the influx of the deepwater chironomids is consistently higher (Supplementary Fig. 4). This is expected as remains of littoral taxa have to be transported over longer distances to reach the coring site whereas deepwater taxa can colonise the surroundings of the coring location if sufficient oxygen is available. Overall, variations in influx of deepwater taxa are much more pronounced than of littoral taxa confirming that the variability in the chironomid assemblages in Bichelsee is mainly driven by variations in population size of oxygen-sensitive deepwater chironomid groups. Local minima in total and deepwater chironomid influx are visible at ca. 6000–5600, 4400–4200, 3000–2600, 900–700, 400–100 cal. BP and during the last 50 years, whereas local maxima are apparent at ca. 5100–4500, 3800–3100, 1800–1000 and 600–400 cal. BP (Fig. 3A, B). The most recent decrease in chironomid influx and shift of invertebrate indicators to values typical for oxygen-poor systems coincides with the 20th century eutrophication of the lake, which at Bichelsee is recorded by an increase of the nutrient-indicating diatom taxon Stephanodiscus binatus & S. rugosus (Fig. 4C)25.

A Log-transformed total chironomid (black) and deepwater chironomid (darkblue) influx. B Log-transformed deepwater chironomid influx (darkblue) and log-transformed percentages of herb pollen (green). C Aquatic plant pollen percentages with Cyperaceae included (orange) and excluded (brown). D Cultural plant pollen percentages (Anethum, Cerealia-type, Fagopyrum esculentum, Linum usitatissimum, Pisum sativum, Secale cereale, Vicia faba, Zea mays). E Organic matter accumulation rate. F Pollen percentages of Alnus glutinosa-type and Cannabis sativa. G Sedimentation rate. Curves in panels (A–D) and (F) are smoothed with a 9-sample running average. ME: Modern Era; MA: Middle Ages; RA: Roman Age; IA: Iron Age; BA: Bronze Age; N: Neolithic. Letters (a–e) in panel B indicate local maxima in the herb pollen record as referred to in the manuscript.

A Percentages of chironomids (black), Chaoborus flavicans-type (blue) and Plumatella (red). B Scores of the first axis of a CCA calculated as in ref. 21 in orange and log-transformed Chaoborus-to-chironomid ratio in blue24. C Percentages of Pantocsekiella ocellata (blue) and Stephanodiscus binatus & S. rugosus (brown) in the uppermost sections of the record25.
Discussion
The Bichelsee chironomid and aquatic invertebrate records indicate that the lake was characterised over thousands of years by stable profundal environments with a well-established benthic fauna and large populations of deepwater chironomid larvae (Fig. 2). These conditions prevailed even under the relatively warm summers of the early and mid-Holocene26, which contributed to stable summer stratification of the lake and favoured late summer anoxia compared to the cooler summers characterising the region during the late Holocene, after ca. 5000–4000 cal. BP. A major decrease in deepwater chironomid populations is registered ca. 7500–7100 cal. BP, suggesting a massive depletion of oxygen at least in the deepwater environments (Fig. 2). This interpretation is consistently supported by the observed decrease in chironomid influx, strongly reduced chironomid percentages in the overall invertebrate assemblages, increase in Chaoborus flavicans-type remains, the ratio of chaoborids relative to chironomids, as well as the strong decrease in CCA axis 1, which all shift towards values typically found in small lakes with anoxic bottom waters21,24. The shift in invertebrate assemblages coincided with a major change in vegetation on the Swiss Plateau. Around 8000 cal. BP, the shade tolerant tree Fagus sylvatica (European beech) started to expand in the region to reach 50 % in the Bichelsee pollen assemblages by ca. 7100 cal. BP (Fig. 2E), replacing mixed oak forests dominated by Corylus avellana, Ulmus, Quercus robur-type, Fraxinus excelsior and Tilia27. This rapid expansion of Fagus sylvatica, together with the expansion of other shade-tolerant trees such as Taxus baccata (yew), and in other parts of the Swiss Plateau, Abies alba (silver fir), suggests denser, dark and mesophyllous forests in the region from ca. 7600 cal. BP onwards27,28,29. The expansion of Fagus sylvatica and Abies alba has generally been considered to be related to a shift in moisture availability in southwestern Central Europe that enabled these trees to expand at the expense of previously dominating tree species30, although at later stages Fagus sylvatica may also have been favoured by human activity during the Neolithic period28,29. However, at Bichelsee no clear indications of major human activities or agriculture around the lake are apparent before ca. 6000 cal. BP27. In addition, increasing abundances of Alnus glutinosa-type pollen and macrofossils in the record suggest that the vegetation belt around the lake may also have become denser during the transition to Fagus-dominated forests with maximum values reached ca. 7100 cal. BP (Fig. 2E). Alnus glutinosa is favoured by wet habitats and near Bichelsee will predominantly have grown along the shores27. Changes in the density of the forest vegetation around small lakes in Central Europe have previously been suggested to have a strong impact on deepwater oxygen conditions in these ecosystems31. The density of vegetation around small lakes may contribute to sheltering them from wind32 and therefore reduce mixing during the main circulation periods in spring and autumn33,34. In addition, the deciduous forests around Bichelsee during this period, and especially the more abundant Alnus glutinosa trees along the shoreline, will have provided important contributions of terrestrial organic matter, in form of coarse organic debris, to the lake31,35, enhancing the oxygen demand of both littoral and deepwater sediments. Variations in the organic carbon accumulation rate indicate a moderate increase in organic carbon availability during this phase of the lake’s history compared with the period before the expansion of mixed Fagus sylvatica forests (Fig. 3E). Our evidence, therefore, strongly suggests that the shift from oxygen-rich to hypoxic deepwater conditions for Bichelsee in the mid-Holocene was related to the closing of the forests, the expansion of Fagus sylvatica and, along the shorelines, of Alnus glutinosa.
For several thousand years after the expansion of mixed Fagus sylvatica forests, our invertebrate assemblages consistently indicate low deepwater oxygen availability in Bichelsee. However, minor variations in the influx of deepwater chironomids, percentages of chironomid remains and CCA axis 1 scores indicate decadal- to centennial-scale phases of increased populations of deepwater chironomids from ca. 5000 cal. BP onwards. The first such phase is centred on ca. 4800 cal. BP, during the Neolithic, with only a relatively minor increase in deepwater chironomid influx. This coincides with a minor but distinct increase in pollen of herbs (a in Fig. 3B) and cultural indicators such as Cerealia-type and Plantago lanceolata as well as a distinct rise in the influx of charcoal and spores of dung-loving fungi in the sediment of Bichelsee, indicating increasing human activity, agriculture and forest opening around the lake27. The second phase with increased chironomid influx values takes place in the Bronze Age ca. 3400 cal. BP (b in Fig. 3B), when clear signs of more intense human activities are seen in the pollen data, with again a decadal-scale increase in indicators such as herbs, Cerealia-type and Plantago lanceolata pollen27. It therefore seems that Neolithic and Bronze Age agriculture activities led to an opening of the forests around Bichelsee and clearly detectable increases in deepwater oxygen availability for invertebrates in the lake. Similar increases in inferred oxygen concentrations associated with mid- to late-Holocene deforestation of catchments have been reported from several other small lakes on the Swiss Plateau33,36.
From the early Iron Age and Roman Age onwards, the reaction of the deepwater chironomid fauna to phases with more intense land use changed dramatically. Phases of more open vegetation now coincide with reduced abundances of deepwater chironomid remains, as indicated by local minima in deepwater chironomids during episodes of high abundances of herb pollen and cultural plants during the Roman Age (ca. 1900 cal. BP; c in Fig. 3B), the High Medieval Period (ca. 1000–800 cal. BP; d in Fig. 3B) and the Modern Era (ca. 400–100 cal. BP; e in Fig. 3B), whereas chironomid influx indicates that deepwater oxygen became more available when local agriculture was reduced between the maxima of cultural plant pollen, especially if these phases coincided with periods of forest recovery (Fig. 3B). These recovery phases coincided with the crisis of the Roman Empire from the third century CE onwards (ca. 1700 cal. BP; Fig. 3B), the Migration Period (ca. 1600–1300 cal. BP; Fig. 3B), and the Medieval Economic Crisis centred on ca. 1350 CE (ca. 600 cal. BP; Fig. 3B), which in the region also coincided with major plague epidemics and local wars. Cultural indicators, Alnus glutinosa-type, Cannabis sativa and aquatic plants show that human activity, and its impact on Bichelsee, changed considerably from the late Iron Age and Roman Age onwards, with more open vegetation, intensified cultivation of the landscape, and deforestation of the Alnus vegetation belt along the shore. This shift was associated with an important increase in pollen of aquatic plants (particularly Nymphaea and Myriophyllum spicatum), possibly a combined effect of higher nutrient availability due to more intense agriculture and higher light availability in shallow sections of the lake due to the cutting of the riparian tree and shrub belt. Organic matter accumulation rate in the sediments of Bichelsee also began to increase from this period onwards (Fig. 3B), indicating that agriculture, land use and other human activities started to have a distinct effect on the lake’s carbon cycle, either by increasing lake productivity and/or increasing organic matter input from the catchment. The presence of the profundal chironomid Chironomus plumosus-type (Supplementary Fig. S3), indicative of eutrophied lakes37, supports our interpretation that increasing human impact altered organic carbon availability and habitats for chironomid larvae. In the Medieval Period abundant Cannabis sativa pollen suggests that cultivation and retting of hemp became an important agricultural activity around Bichelsee. The increase in Cannabis sativa correlates well with the decrease of Alnus glutinosa-type pollen in this period, suggesting that the shoreline vegetation of Bichelsee was cleared to obtain better access to the lake shore for this activity. At the same time, sedimentation rates start to increase indicating that the Medieval rise in agricultural activities, clearcutting of the catchment vegetation and hemp retting favoured erosion. After the short-term recovery of the deepwater chironomid fauna during the Medieval Economic Crisis ca. 600 cal. BP, cultural indicators again start to increase in the pollen record and influx of deepwater chironomid remains decreases again. Our evidence therefore indicates that the effect and directionality of human impact on the lake changed in the course of the Holocene. After hypoxic conditions were established around 7100 cal. BP, first phases of increased clearcutting and agriculture around the lake were associated with decadal- to centennial-scale phases of increased oxygen availability. With the progressive clearing and more intense agriculture from the Roman Age onwards, phases of enhanced human activity were instead corresponding to periods with reduced oxygen availability, presumably associated with higher nutrient or organic matter influx into the lake due to higher population levels and intensive colonisation, agriculture and animal husbandry in the vicinity of Bichelsee. In contrast, major economic disturbances and societal crises such as the fall of the Western Roman Empire and the following Migration Period, as well as the Medieval Economic Crisis and plague epidemics in the 14th century CE corresponded with a distinct recovery of oxygen conditions and deepwater invertebrate populations. The 20th century eutrophication of the lake was again associated with major changes in the abundances of invertebrate indicators of deepwater oxygen availability (Fig. 4) suggesting a renewed decrease in oxygen availability during the past ca. 50 years.
Climatic changes can have considerable impacts on lake ecosystems38. In hydrologically closed basins, variations in hydroclimate can lead to changes in lake levels, which in turn can influence lake size and depth, shift the ratio between littoral and deepwater habitats and modify the proportion of the lake basin affected by deepwater anoxia in stratified lakes. Temperature changes can also influence many catchment processes as well as conditions within lakes, such as plant productivity in both littoral and pelagic habitats, the stability of summer stratification and consequently the availability of oxygen in deepwater environments38,39. In our study region, decadal- to centennial-scale Holocene cold events40 and phases of changing lake levels41 have been described, which have a similar pacing as variations in deepwater oxygen availability in Bichelsee as reconstructed by aquatic invertebrate remains. However, several arguments lead us to conclude that climatic changes only had minor impacts on aquatic invertebrate assemblages in Bichelsee compared with the major changes in vegetation, land use and human impact in the lake catchment. First, water level changes in other studied lakes in southwestern Central Europe and adjacent regions only show relatively minor Holocene variations, typically in the order of ± 1 m (Supplementary Fig. S6). Water level changes of this amplitude would have only been associated with relatively minor changes in deepwater environments of Bichelsee. Assuming a water table close to the present conditions (with a water depth of 7 m) and the infilling of the lake basin during the past millennia, Bichelsee would have been ca. 11–14 m deep during the middle and early late Holocene (Fig. 1). In such small, deep and stratified lakes, it is unlikely that minor changes in lake level will have led to major shifts in summer stratification patterns or deepwater oxygen availability. Second, if climatic variations would have caused major water table changes in Bichelsee, several aquatic indicators, such as influx of littoral chironomid remains (Supplementary Fig. S5) or percentages of aquatic macrophytes (Fig. 3C), would be expected to show major and consistent variations with reconstructed phases of lake level changes in the region. However, before the distinct increase of human impacts during the Roman Age, these indicators do not show variations that would agree with changing water table and associated shifts in habitat availability or summer stratification. Third, and most importantly, a detailed comparison of our record with reconstructed phases of elevated lake levels in other lakes41, climatic variations for the Alpine area40 (Supplementary Fig. S6) and climate events during the last 2000 years42 (Supplementary Fig. S7) only shows a poor agreement in the pacing and timing of climate events and our reconstructed variations in deepwater oxygen availability in Bichelsee. For example, a few increases in lake levels on the Swiss Plateau41 (ca. 1700 and 1300–100 cal. BP) seem to coincide with phases of increased deepwater oxygen availability in Bichelsee. However, the majority of lake level increases do not agree with reconstructed phases of minimum or maximum deepwater oxygen availability and instead occur during periods of relatively stable deepwater oxygen conditions (ca. 6300–5900, 5600–5200, 2800–2400 cal. BP). A similar pattern emerges when we compare our record with cold events in the Swiss Alps (Supplementary Fig. S6) or climatic events during the last 2000 years (Supplementary Fig. S7). This mismatch between the Bichelsee invertebrate record and climate reconstructions contrasts strongly with the close temporal agreement between variations in deepwater oxygen availability and changes in forest cover, agriculture and land use in the catchment (Figs. 2, 3).
Our results challenge the concept of a clearly definable, natural pre-impact state for small Central European lakes such as Bichelsee. First, our record shows that Bichelsee experienced two very different Holocene ecosystem states in regard to oxygen well before any detectable phases of early human impact around the lake. The first phase, lasting until ca. 7400 cal. BP, was characterised by stable oxygen-rich deepwater conditions, as demonstrated by high abundances of deepwater chironomid larvae and stable fossil chironomid assemblage composition. The second phase was associated with a major transition of forest composition in the lake catchment and development of denser vegetation along the lakeshore coinciding with oxygen-poor and, on multidecadal to centennial timescales, very variable and unstable conditions for invertebrates and other organisms living in the deepwater environments. Some earlier studies have also shown that hypoxic conditions can develop in small Central European lakes under densely forested conditions, in some cases with hypoxia reported for most of the early and mid-Holocene36, in other cases with distinct increases in productivity and decreases in mixing and oxygenation reported during the mid-Holocene, as we find in our study33. However, these studies were not able to provide information on the presence and abundance of deepwater invertebrate populations at the surveyed lakes. At Bichelsee, hypoxic conditions only established with the expansion of Fagus sylvatica forests in the catchment, indicating that both oxygen-rich and oxygen-poor states are possible in such lakes under relatively stable Holocene conditions and in absence of important human activities. Second, our results show that, depending on the overall climatic and environmental background, small Central European lakes can be characterised by very stable, but also very variable deepwater environments on decadal time scales. At Bichelsee, the abundances of different invertebrate groups, and the chironomid assemblage composition were more variable from ca. 7100 cal. BP onwards than before the transition to hypoxic conditions. Third, our results strongly suggest that from ca. 4800 cal. BP onwards, oxygen availability and deepwater invertebrate assemblages were regularly affected by decadal- to centennial-scale phases of human activities in the lake catchment, related to phases of increased clearcutting, agricultural activity and overall human presence. However, unexpectedly our results indicate that the directionality of the effects of these human activities on Bichelsee changed in the course of the late Holocene, first promoting benthic invertebrate assemblages and increased deepwater oxygen availability in the lake, later coinciding with decreased deepwater invertebrate populations and oxygen availability. Finally, our results show that in regions where landscapes and vegetation have a long history of human impacts, sensitive aquatic ecosystems, such as Bichelsee, may have experienced a series of complex responses and recoveries related to increasing and decreasing human pressures. The transition to more oxygen-poor conditions due to the mid-20th century eutrophication (Fig. 4) only represents the most recent of a series of shifts that started 4800 years ago. Defining any particular time interval within the past millennia as representing the natural, undisturbed reference conditions, such as the period immediately preceding the Industrial Era eutrophication (e.g., the interval around 1850 CE as used in some other palaeolimnological studies) can only be considered as arbitrary for such lakes.
Palaeolimnological reconstructions covering environmental changes over millennia have to take into account the slowly decreasing depth of lake basins due to sediment infilling, a process that may change the bathymetry and overall depth of small lakes considerably. In the case of Bichelsee, the current lake is shallower (7 m) than the expected depth during the mid- and early late-Holocene (11–14 m; assuming no sediment compaction and similar water table as present). Water depth is one criterion regularly used to assign possible reference conditions for impacted lake ecosystems, and, as a consequence, the deeper mid-Holocene conditions may not necessarily reflect a realistic restoration aim for Bichelsee. However, this does not influence the conclusions listed above as they are not constrained to lakes with similar depths as Bichelsee is characterised by at present but based on the reconstructed ecosystem dynamics of the deeper lake during the mid- and late-Holocene. For example, our conclusions that Bichelsee was characterised by two different natural pre-impact states prior to any major human impacts or that lakes such as Bichelsee during the mid- to late Holocene may experience rapid transitions between hypoxic and oxygen-rich ecosystem states as a consequence of changing human activities can be taken independently of the long-term infilling of the lake. Similarly, the finding that the directionality of these shifts can change in the course of a lake’s history, depending on the exact nature of human activities, is independent of the reduced depth of the lake in the recent past.
The possibility of multiple, distinct pre-impact ecosystem states, changes in the stability of deepwater environments and a potentially long history of human impacts on lake ecosystems, as well as a changing directionality of their effects on deepwater oxygen conditions, should be considered when defining natural and undisturbed reference conditions for small Central European lakes such as Bichelsee. These considerations are relevant when defining reference states that should represent the expected status of evaluated, modern lake ecosystems under minimal human activity, as required by water protection legislation such as the European Water Framework Directive. However, they should also be considered in more general scientific discussions on the expected conditions in European freshwater ecosystems under minimal human impact, on how strongly these ecosystems presently deviate from these conditions, and since when, to what extent and with which dynamics human activities affected freshwater environments and led to their presently observed state in the modern environment.
Methods
Study site
Bichelsee is a small lake (9.7 ha) located on the north-eastern Swiss Plateau (47°27.466‘ N, 8°54.027‘ E, 591 m a.s.l.) in a meltwater channel formed during the last glaciation (ca. 20,000 years ago). At present, the maximum water depth is 7 m and the lake has a simple, oval shape with the deepest section in its centre43. Limnological measurements are limited but indicate that under modern conditions the water column is thermally stratified and bottom water is hypoxic during summer (<0.2 mg/L O2 below 4 m water depth in August 2025; Data from the Amt für Umwelt Kanton Thurgau; Supplementary Fig. 7). The lake is surrounded by steep hills presently covered by mixed beech forests and agricultural lands (Fig. 1)43.
Core sampling and chronology
Parallel sediment cores were retrieved in November 2019 and 2021 with a UWITEC piston corer whereas the uppermost sediments were sampled with a UWITEC gravity corer and hammer action44. During the first campaign only 6.25 m of sediment were retrieved, and therefore a second campaign was necessary. The different core segments were correlated in the laboratory based on conspicuous layers and colour changes resulting in a continuous sediment section of 10.85 m44. The sediment core is composed of calcareous gyttja and silt from 10.85–9.18 m, darkbrown gyttja from 9.18–9.09 m and brown gyttja between 9.09–0 m (Supplementary Fig. S2). Detailed descriptions of the coring, sediment composition and geochemical analyses (XRF, organic matter and carbonate content) are available in ref. 44.
Seventy-two plant macrofossil remains were selected for radiocarbon dating using accelerator mass spectrometry (AMS) at the Laboratory for the Analysis of Radiocarbon with AMS (LARA) of the University of Bern44. In addition, 22 samples from the topmost 101 cm of sediment were analysed for 210Pb and 137Cs activities with a Broad Energy Germanium detector (Canberra GmbH, Rüsselsheim, Germany)44. Changes in 137Cs activity were used to constrain the first increase in nuclear bomb tests in 1955 CE, the maximum 137Cs input in 1963 CE from nuclear bomb testing and the Chernobyl reactor accident in 1986 CE. The modelling of the relationship between sediment depth and age for the uppermost sediments followed the Piecewise Constant Rate of Supply Model45. With this approach, ages were estimated based on 210Pb measurements assuming a constant rate of supply but constrained to intercept with the horizons dated based on 137Cs. The 72 14C ages and the ages based on 210Pb and 137Cs were combined to calculate an overall age-depth relationship for the sequence using the programme Bacon46. For these final calculations two of the 72 14C measurements were excluded as outliers. The age model for the Bichelsee record is described in more detail in ref. 44.
Aquatic invertebrate analysis
A total of 257 samples were selected for aquatic invertebrate analysis of the Bichelsee sediment core. A first, coarse-resolution chironomid record for the site was provided in ref. 47. For the present study the sampling resolution was more than trebled to allow high resolution analyses of chironomid influx variations throughout the Holocene, reveal decadal-scale variability in the invertebrate populations, assess rates of change along the record and enable comparisons with other high-resolution records from Bichelsee. A continuous subsampling strategy was adopted for the topmost 500 cm (ca. 4000 cal. BP) with 3 cm thick subsamples of 6 cm3 (average temporal resolution: 21.6 years). For the rest of the record, samples were selected at lower temporal resolution (average temporal resolution: 121.6 years; 3 to 23 cm between adjacent samples), with a volume ranging from 1 to 4 cm3 and thickness between 1 to 3 cm depending on the concentration of aquatic invertebrate remains. The samples were gently sieved under water through a 100 µm mesh-size sieve following standard methods48. After that, chironomids and other aquatic invertebrate remains were hand-picked from the sieved residue using fine forceps and a Bogorov tray under a stereomicroscope (20–50x magnification) and permanently mounted on a microscope slide using Euparal. Chironomids were identified under a compound microscope (100–400x magnification) based on refs. 48,49,50. Head capsules were enumerated as follows: as one when more than half of the mentum was present, as half when half of the mentum was present and as zero when less than half of the mentum was present. For graphical and statistical presentation, chironomid remains not identified to the highest taxonomic resolution (e.g., due to missing mouthparts or antennal pedestal) were assigned to the more detailed taxonomic categories based on the ratio of identified specimens in the same sample51. Cladoceran ephippia present in the sieved residue prepared for chironomid analyses were identified following refs. 52,53, insect mandibles based on ref. 17, Chaoborus mandibles using ref. 54, bryozoan statoblasts with ref. 55 and oribatid mites with ref. 56. Due to the low concentration of remains in some parts of the record, adjacent samples were grouped together to reach at least 20 identified chironomids and 80 aquatic invertebrates (chironomids included) per sample to perform numerical analyses (e.g., calculation of percentages, CCA, Chaoborus-to-chironomid ratio). Influx values of chironomid and invertebrate remains were calculated based on the concentration of each taxon divided by the accumulation rate of sediment inferred from the age-depth model. Influx values of total chironomid and deepwater chironomid remains were calculated for all 257 samples without pooling adjacent samples. As in most other Holocene sediment records from Swiss lowland lakes, preservation of chironomid and other aquatic invertebrate remains in the sediment was excellent, with no evidence of degradation.
Invertebrate indicators of deepwater oxygen availability and numerical analyses
For most numerical analyses, the pooled dataset of 123 samples was used, whereas influx values of total and deepwater chironomid taxa are also presented based on the full record of 257 samples. The dissimilarity between samples, which can also be considered as the rate of change between adjacent samples, was calculated as percentage dissimilarity index following ref. 57 for the chironomid dataset. We used several approaches to assess whether invertebrate assemblages in Bichelsee changed towards states typical for higher or lower oxygen availability in small Swiss lakes. Higher abundances of Chaoborus remains and a higher ratio of Chaoborus relative to chironomid remains have been indicated to be more typical for lower deepwater oxygen concentrations in small stratified lakes21,24. Higher percentages of chironomids relative to other aquatic invertebrate remains can be indicative for high bottom water oxygen availability in small Swiss lakes21. For a more quantitative assessment as to the extent that fossil invertebrate assemblages change from states representative for oxygen-rich conditions to states typical for low oxygen availability, and vice versa, we plotted fossil invertebrate samples passively in a CCA of invertebrate assemblages in lake surface sediment of 36 small Swiss lakes covering a wide range of elevations (403–2065 m a.s.l.), water depths (<1–38 m) and deepwater oxygen concentrations (0–105.6 %)21. Deepwater oxygen availability was a major determinant of invertebrate assemblage composition in this dataset when all the examined environmental variables were assessed together21 (Supplementary Fig. 8A). For the present study, we calculated a CCA with deepwater oxygen concentration as the only constraining variable, and changes along CCA axis 1 are therefore constrained to reflect changes in deepwater oxygen availability (Supplementary Fig. 8B). Variations of CCA axis 1 scores of the passively added Bichelsee samples can therefore be interpreted as indicating shifts towards invertebrate assemblage states more typical for high or low oxygen availability in small Swiss lakes. Analyses were based on square-root transformed percentages of the overall aquatic invertebrate dataset with chironomids grouped in one category (123 samples)21. The CCA was calculated with the function cca from the vegan package version 2.6–4 in R Studio version 2024.12.1.563. Finally, influx values of total chironomid and deepwater chironomid remains were calculated as an indicator of the abundance of benthic invertebrates and oxygen availability in the deepwater environments of Bichelsee. In sediment cores taken from the deepest sections of lakes, both of these indicators are expected to closely track the abundance of chironomid larvae living close to the coring location, which will closely relate to the availability of oxygen in water layers above the sediment. For these analyses, we considered chironomids that can colonise the profundal of thermally stratified Central European lakes as deepwater taxa37,58. In the Bichelsee record, these included Chironomus anthracinus-type, Chironomus plumosus-type, Heterotrissocladius grimshawi-type, Heterotrissocladius marcidus-type, Macropelopia, Micropsectra contracta-type, Micropsectra insignilobus-type, Paracladius, Procladius, Sergentia coracina-type, Tanytarsus lugens-type and Tanytarsus mendax-type. Chironomid influx values, Chaoborus-to-chironomid ratios and, for comparison with the chironomid influx record, also herb pollen percentages were plotted on a log – or if the records contained zero values – a log(x + 1)-scale to visualise both relatively subtle changes at low values and major shifts at high values in these records. Untransformed data for these variables are provided in the online supporting data.
Pollen analysis
The pollen analysis of the Bichelsee record followed standard methods that are described in detail in ref. 27. A total of 706 1 cm3 sediment samples were treated with HCl 10 %, KOH 10 %, HF 40 % and acetolysis following ref. 59. Pollen slides were mounted in glycerine and then identified under a compound microscope at 400x magnification. We compare the Bichelsee chironomid and invertebrate records with variation in the percentages of selected plant taxa and indicator groups including overall herbs, Fagus sylvatica, Alnus glutinosa-type, Cannabis sativa pollen and aquatic plant pollen and palynomorphs (Alisma, Cladium mariscus, Menyanthes trifoliata, Myriophyllum spicatum, Myriophyllum verticillatum, Oenanthe fistulosa-type, Polygonum amphibium, Potamogeton, Ranunculus aquatilis-type, Rumex aquaticus, Scheuchzeria palustris, Sparganium-type, Typha latifolia-type, Utricularia pollen, Ceratophyllum spines, Nymphaeaceae base cells and hairs). The aquatic plants are shown both including and excluding Cyperaceae pollen, since Cyperaceae can be expected to also be a common component of terrestrial ecosystems on the lake shores. In addition, we show the sum of cultural pollen types, which includes Anethum, Cerealia-type, Fagopyrum esculentum, Linum usitatissimum, Pisum sativum, Secale cereale, Vicia faba and Zea mays. Following other pollen records on the Swiss Plateau, total aquatic plant and Cannabis sativa pollen were not included in the pollen sum to calculate pollen percentages as they do not represent the terrestrial vegetation around the lake60. Cannabis sativa pollen deposited in sediments of small European lakes during the Roman Age, Medieval Period and Modern Era may originate to considerable extents from hemp retting in the water along the lake shores rather than the local vegetation around the lake and are therefore regularly excluded from the pollen sum60,61.
Diatom analysis
Diatoms were poorly preserved along large sections of the Bichelsee sediment profile and it was therefore not possible to produce a continuous, high resolution diatom record for this lake25. However, diatom preservation was better and continuous in the uppermost sediment sections (from ca. 1940 CE) and variations in diatom assemblage composition track the peak industrial-age eutrophication and recovery of the lake since the 1950s. Here we only show variations in percentages of Pantocsekiella ocellata, a diatom occurring in a large range of trophic conditions62, but more frequently found in small and eutrophic lakes, and Stephanodiscus binatus & S. rugosus, a diatom group that is favoured by high nutrient concentrations25,63,64, in these uppermost sediment layers. At Bichelsee, high percentage values of the latter taxon represent the peak eutrophication phase of the lake between 1970–1990 CE. Full details on diatom analyses in the Bichelsee sediment record can be found in ref. 25.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Data supporting the findings of this manuscript (chironomid and aquatic invertebrate counts, percentages, influxes, 210Pb and 137Cs measurements, radiocarbon dates) are available in the Supplementary Data associated with this manuscript and on Dryad at https://doi.org/10.5061/dryad.gb5mkkx47. Data published in previous studies used to draw the figures can be found in the Supplementary Data and Dryad (https://doi.org/10.5061/dryad.gb5mkkx47) or in the respective studies cited in the manuscript.
Code availability
The code used to calculate the CCAs is available in Dryad at https://doi.org/10.5061/dryad.gb5mkkx47.
References
Heino, J. et al. Lakes in the era of global change: moving beyond single-lake thinking in maintaining biodiversity and ecosystem services. Biol. Rev. 96, 89–106 (2021).
Bhateria, R. & Jain, D. Water quality assessment of lake water: a review. Sustain. Water Resour. Manag. 2, 161–173 (2016).
Alexander, T. J., Vonlanthen, P. & Seehausen, O. Does eutrophication-driven evolution change aquatic ecosystems? Philos. Trans. R. Soc. B Biol. Sci. 372, 20160041 (2017).
Søndergaard, M. et al. Lake restoration: successes, failures and long-term effects. J. Appl. Ecol. 44, 1095–1105 (2007).
Kristensen, P. et al. European waters assessment of status and pressures 2018. EEA Report No 7/2018 (85 pp.), European Environment Agency (2018).
Hering, D. et al. The European Water Framework Directive at the age of 10: A critical review of the achievements with recommendations for the future. Sci. Total Environ. 408, 4007–4019 (2010).
McNellie, M. J. et al. Reference state and benchmark concepts for better biodiversity conservation in contemporary ecosystems. Glob. Change Biol. 26, 6702–6714 (2020).
Bennion, H. & Battarbee, R. The European Union Water Framework Directive: opportunities for palaeolimnology. J. Paleolimnol. 38, 285–295 (2007).
Jokinen, S. A. et al. A 1500-year multiproxy record of coastal hypoxia from the northern Baltic Sea indicates unprecedented deoxygenation over the 20th century. Biogeosciences 15, 3975–4001 (2018).
Osterman L. E. et al. The last 1000 years of natural and anthropogenic low-oxygen bottom-water on the Louisiana shelf, Gulf of Mexico. Marine Micropaleontol. 66, 291–303 (2008).
Bennion, H. et al. Defining reference conditions and restoration targets for lake ecosystems using palaeolimnology: a synthesis. J. Paleolimnol. 45, 533–544 (2011).
Lang, G. et al. Quaternary Vegetation Dynamics of Europe (Haupt Verlag, 2023).
Dubois, N. et al. First human impacts and responses of aquatic systems: A review of palaeolimnological records from around the world. Anthr. Rev. 5, 28–68 (2018).
Tóth, M. et al. Pronounced early human impact on lakeshore environments documented by aquatic invertebrate remains in waterlogged Neolithic settlement deposits. Quat. Sci. Rev. 205, 126–142 (2019).
Zapolska, A. et al. More than agriculture: Analysing time-cumulative human impact on European land-cover of second half of the Holocene. Quat. Sci. Rev. 314, 108227 (2023).
The Swiss Federal Council, 2025. 814.201 Waters Protection Ordinance of 28 October 1998 (WPO). https://www.fedlex.admin.ch/eli/cc/1998/2863_2863_2863/en.
Courtney-Mustaphi, C. J. et al. Aquatic invertebrate mandibles and sclerotized remains in Quaternary lake sediments. J. Paleolimnol. 71, 45–83 (2024).
Armitage, P. D., Cranston, P. S. & Pinder, L. C. V. The Chironomidae: The biology and ecology of non-biting midges (Springer Netherlands, 1995).
Jyväsjärvi, J., Nyblom, J. & Hämäläinen, H. Palaeolimnological validation of estimated reference values for a lake profundal macroinvertebrate metric (Benthic Quality Index). J. Paleolimnol. 44, 253–264 (2010).
Verbruggen, F. et al. Subfossil chironomid assemblages in deep, stratified European lakes: relationships with temperature, trophic state and oxygen: Chironomid assemblages in deep, stratified lakes. Freshw. Biol. 56, 407–423 (2011).
Ursenbacher, S., Stötter, T. & Heiri, O. Chitinous aquatic invertebrate assemblages in Quaternary lake sediments as indicators of past deepwater oxygen concentration. Quat. Sci. Rev. 231, 106203 (2020).
Lotter, A. F. et al. Modern diatom, cladocera, chironomid, and chrysophyte cyst assemblages as quantitative indicators for the reconstruction of past environmental conditions in the Alps. I. Climate. J. Paleolimnol. 18, 395–420 (1997).
Hamburger, K., Dall, P. C. & Lindegaard, C. Energy metabolism of Chironomus anthracinus (Diptera: Chironomidae) from the profundal zone of Lake Esrom, Denmark, as a function of body size, temperature and oxygen concentration. Hydrobiologia 294, 43–50 (1994).
Quinlan, R. & Smol, J. P. Use of subfossil Chaoborus mandibles in models for inferring past hypolimnetic oxygen. J. Paleolimnol. 44, 43–50 (2010).
Grudzinska, I. & Heiri, O. Kieselalgen in den Sedimenten des Bichelsees und des Hüttwilersees. Klima, Umwelt, Mensch im Thurgau (KUMIT) – hochaufgelöste Umweltrekonstruktionen der letzen 17’000 Jahre anhand von Sedimentkernen aus dem Bichelsee und dem Hüttwilersee, Kanton Thurgau (Schweiz) (eds Geisser, H. & Leuzinger, U.) 117–136 (Mitteilungen der Thurgauischen Naturforschenden Gesellschaft 72, 2024).
Heiri, O. et al. Stacking of discontinuous regional palaeoclimate records: chironomid-based summer temperatures from the Alpine region. Holocene 25, 137–149 (2015).
Wick, L., Rey, F. & Heiri, O. Pollenanalyse und Pflanzenmakroreste. Klima, Umwelt, Mensch im Thurgau (KUMIT) – hochaufgelöste Umweltrekonstruktionen der letzen 17’000 Jahre anhand von Sedimentkernen aus dem Bichelsee und dem Hüttwilersee, Kanton Thurgau (Schweiz) (eds Geisser, H. & Leuzinger, U.) 73–116 (Mitteilungen der Thurgauischen Naturforschenden Gesellschaft 72, 2024).
Rey, F. et al. Causes and mechanisms of synchronous succession trajectories in primeval Central European mixed Fagus Sylvatica forests. J. Ecol. 107, 1392–1408 (2019).
Rey, F. et al. Neolithic land use and forest dynamics on the Swiss Plateau (southwestern Central Europe). Quat. Sci. Rev. 360, 109372 (2025).
Tinner, W. & Lotter, A. Holocene expansions of Fagus silvatica and Abies alba in Central Europe: Where are we after eight decades of debate? Quat. Sci. Rev. 25, 526–549 (2006).
Żarczyński, M., Wacnik, A. & Tylmann, W. Tracing lake mixing and oxygenation regime using the Fe/Mn ratio in varved sediments: 2000 year-long record of human-induced changes from Lake Żabińskie (NE Poland). Sci. Total Environ. 657, 585–596 (2019).
Becu, M. H. J., Michalski, T. A. & Richardson, J. S. Forest harvesting impacts on small, temperate zone lakes: a review. Environ. Rev. 31, 376–402 (2023).
Makri, S. et al. Early human impact in a 15,000-year high-resolution hyperspectral imaging record of paleoproduction and anoxia from a varved lake in Switzerland. Quat. Sci. Rev. 239, 106335 (2020).
Klaus, M., Karlsson, J. & Seekell, D. Tree line advance reduces mixing and oxygen concentrations in arctic–alpine lakes through wind sheltering and organic carbon supply. Glob. Change Biol. 27, 4238–4253 (2021).
Czarnecka, M. Coarse Woody debris in temperate littoral zones: implications for biodiversity, food webs and lake management. Hydrobiologia 767, 13–25 (2016).
Lotter, A. F. The palaeolimnology of Soppensee (Central Switzerland), as evidenced by diatom, pollen, and fossil-pigment analyses. J. Paleolimnol. 25, 65–79 (2001).
Sæther, O. A. Chironomid communities as water quality indicators. Holarct. Ecol. 2, 65–74 (1979).
Woolway, R. I. et al. Global lake responses to climate change. Nat. Rev. Earth Environment 1, 388–403 (2020).
Eggermont, H. & Heiri, O. The chironomid-temperature relationship: expression in nature and palaeoenvironmental implications. Biol. Rev. 87, 430–456 (2012).
Haas, J. N. et al. Synchronous Holocene climatic oscillations on the Swiss Plateau and at timberline in the Alps. Holocene 8, 301–309 (1998).
Magny, M. Holocene climate variability as reflected by mid-European lake-level fluctuations and ist probable impact on prehistoric human settlements. Quaternary Int. 113, 65–79 (2004).
Wanner, W., Pfister, C. & Neukom, R. The variable European Little Ice Age. Quaternary Sci. Rev. 287, 107531 (2022).
Leuzinger, U. & Heiri, O. Hüttwilersee und Bichelsee – zwei Datenarchive für die Rekonstruktion von Klima, Umwelt und menschlichem Einfluss durch die Jahrtausende im Kanton Thurgau. Klima, Umwelt, Mensch im Thurgau (KUMIT) – hochaufgelöste Umweltrekonstruktionen der letzen 17’000 Jahre anhand von Sedimentkernen aus dem Bichelsee und dem Hüttwilersee, Kanton Thurgau (Schweiz) (eds Geisser, H. & Leuzinger, U.) 19–26 (Mitteilungen der Thurgauischen Naturforschenden Gesellschaft 72, 2024).
Heiri, O. et al. Beprobung, Sedimentabfolge, XRF-Analysen und Datierung. Klima, Umwelt, Mensch im Thurgau (KUMIT) – hochaufgelöste Umweltrekonstruktionen der letzen 17’000 Jahre anhand von Sedimentkernen aus dem Bichelsee und dem Hüttwilersee, Kanton Thurgau (Schweiz) (eds Geisser, H. & Leuzinger, U.) 51–72 (Mitteilungen der Thurgauischen Naturforschenden Gesellschaft 72, 2024).
Bruel, R. & Sabatier, P. serac: an R package for ShortlivEd RAdionuclide chronology of recent sediment cores. J. Environ. Radioact. 225, 106449 (2020).
Blaauw, M. & Christen, J. A. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal 6, 457–474 (2011).
Lapellegerie, P., Breu, S. & Heiri, O. Aquatische Invertebraten. Klima, Umwelt, Mensch im Thurgau (KUMIT) – hochaufgelöste Umweltrekonstruktionen der letzen 17’000 Jahre anhand von Sedimentkernen aus dem Bichelsee und dem Hüttwilersee, Kanton Thurgau (Schweiz) (eds Geisser, H. & Leuzinger, U.) 137–153 (Mitteilungen der Thurgauischen Naturforschenden Gesellschaft 72, 2024).
Brooks, S. J., Langdon, P. G. & Heiri, O. The Identification and Use of Palaearctic Chironomidae Larvae in Palaeoecology (Quaternary Research Association, 2007).
Wiederholm, T. Chironomidae of the Holarctic region. Keys and diagnoses. Part 1. Larvae (Entomologica Scandinavica, 1983).
Schmid, P. E. A key to the larval Chironomidae and their instars from Austrian Danube region streams and rivers (Federal Institute for Water Quality, 1993).
Bolland, A. et al. Summer temperature development 18,000–14,000 cal. BP recorded by a new chironomid record from Burgäschisee, Swiss Plateau. Quat. Sci. Rev. 243, 106484 (2020).
Vandekerkhove, J. et al. Use of ephippial morphology to assess richness of anomopods: potentials and pitfalls. J. Limnol. 63, 75 (2004).
Szeroczynska, K. & Sarmaja-Korjonen, K. Atlas of Subfossil Cladocera from Central and Northern Europe (Friends of the Lower Vistula Society, 2007).
Uutala, A. J. Chaoborus (Diptera: Chaoboridae) mandibles – paleolimnological indicators of the historical status of fish populations in acid-sensitive lakes. J. Paleolimnol. 4, 139–151 (1990).
Wood, T. S. & Okamura, B. A new key to the freshwater bryozoans of Britain, Ireland and continental Europe, with notes on their ecology (Freshwater Biological Association, 2005).
Solhøy, I. W. & Solhøy, T. The fossil oribatid mite fauna (Acari: Oribatida) in late-glacial and early-Holocene sediments in Kråkenes Lake, western Norway. J. Paleolimnol. 23, 35–47 (2000).
Kiernan, D. D. Natural resources biometrics (Open SUNY Textbooks, 2014).
Janececk, B., Moog, O. & Orendt, C. Diptera: Chironomidae (Non-Biting Midges). Fauna Aquatica Austriaca (eds Moog, O. & Hartmann, A.) (BMLFUW, Wien, Austria, 2017).
Moore, P. D., Webb, J. A. & Collinson, M. E. Pollen analysis (Blackwell, 1991).
Rey, F. et al. Climate impacts on vegetation and fire dynamics since the last deglaciation at Moossee (Switzerland). Clim. Past 16, 1347–1367 (2020).
Ranalli, P. & Venturi, G. Hemp as a raw material for industrial applications. Euphytica 140, 1–6 (2004).
Houk, V., Klee, R. & Tanaka, H. Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A: Cyclotella, Tertiarius, Discotella (Czech Phycological Society, 2010).
Houk, V., Klee, R. & Tanaka, H. Atlas of freshwater centric diatoms with a brief key and descriptions. Part IV. Stephanodiscaceae B: Stephanodiscus, Cyclostephanos, Pliocaenicus, Hemistephanos, Stephanocostis, Mesodictyon & Spicaticriba (Czech Phycological Society, 2014).
Rott, E. & Kofler, W. Planktonic centric diatoms from the Eastern Alps: morphology, biogeography and ecology. Plant Fungal Syst 66, 1–36 (2021).
Acknowledgements
We would like to thank the owners of Bichelsee, the Aktiengesellschaft J. Kindlimann and the Genossenschaft Badi Bichelsee for supporting and approving the sediment sampling. Furthermore, we would like to thank Richard Niederreiter, Martin Niederreiter, Daniel Niederreiter and Günther Praunias from UWITEC for their help during the coring. Further thanks go to Roman Schawalder, Heinz Ehmann and the Amt für Umwelt (Kanton Thurgau) for the water chemistry measurements performed in August 2025 as well as René Thoma for providing the aerial photo of Bichelsee and Dominik Schmed for helping during the aquatic invertebrate sample preparation. We also thank the three reviewers for their valuable comments on the manuscript. This work was funded by the Walter-Enggist Fonds.
Author information
Authors and Affiliations
Contributions
Pierre Lapellegerie: Conceptualisation, Data Curation, Formal Analysis, Investigation, Visualisation, Writing - Original Draft, Writing - Review & Editing. Simona Breu: Investigation, Writing - Review & Editing. Lucia Wick: Conceptualisation, Investigation, Writing - Review & Editing. Fabian Rey: Conceptualisation, Investigation, Visualisation, Writing - Review & Editing. Ieva Grudzinska: Investigation, Writing - Review & Editing. Nathalie Dubois: Writing - Review & Editing. Soenke Szidat: Investigation, Writing - Review & Editing. Sandra Camara-Brugger: Writing - Review & Editing, Visualisation. Urs Leuzinger: Conceptualisation, Funding Acquisition, Writing - Review & Editing. Oliver Heiri: Conceptualisation, Formal Analysis, Funding Acquisition, Visualisation, Writing - Original Draft, Writing - Review & Editing.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth and Environment thanks Guangjie Chen and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Somaparna Ghosh [A peer review file is available].
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lapellegerie, P., Breu, S., Wick, L. et al. Dynamic deepwater invertebrate populations challenge the concept of oxygen-rich reference conditions for European lakes. Commun Earth Environ 7, 301 (2026). https://doi.org/10.1038/s43247-026-03284-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-026-03284-7
