
Abstract
We assess three words commonly used to represent the environmental dimensions of antimicrobial resistance (AMR) – ‘hotspot’, ‘reservoir’ and ‘pristine’ – through two questions: how are these terms used in published research; and how do these terms shape research being conducted? We advocate for the community to reflect on and improve its use of language, and suggest four potentially more productive and precise terms for AMR hazard: prevalence; transmission; evolution and connectivity.
Similar content being viewed by others

Introduction
Antimicrobial resistance (AMR) poses a serious global threat to human health1, with bacterial AMR being the central topic of scientific and regulatory interest2. The One Health paradigm has brought attention to AMR as an interlinked human, animal and environmental issue3. In this article we focus specifically on research related to the environmental dimensions of AMR. There are three major threats associated with AMR in the environment. First, from antimicrobial pollution, including both antibiotic4 and non-antibiotic chemicals5, especially in water environments6,7, creating selection8 and co-selection conditions9. Second, from the presence and transmission of mobile and mobilizable resistance gene(s)10,11,12 through horizontal gene transfer (HGT), ultimately into clinically important pathogens13,14,15. This includes the risk of HGT transferring previously undocumented antimicrobial resistance genes (ARGs)16, or giving rise to new combinations of ARGs, conferring and spreading multi-drug or pan-drug resistance17,18,19. Third, from the direct transfer of AMR pathogens to humans via contaminated water20, and food21 or inadvertent colonization of commensals containing ARGs through environmental exposure22.
The environmental dimensions of AMR poses a distinct set of challenges in comparison with the human and livestock contexts. With the latter, research and policy have drawn attention to failures of public and professional knowledge leading to the overuse and prescription of antibiotics, in turn presenting as a solution the promotion of ‘judicious’23, ‘prudent’24 or ‘rational’25 antibiotic prescribing and use behaviors26,27. In contrast, research on the environmental dimensions of AMR draws attention to a broader set of open-ended processes, flows and interactions between society, the environment, antimicrobial chemicals and bacterial ecosystems implicated in the rising prevalence of AMR28,29,30,31.
There are considerable challenges in determining how complex social-environmental systems can be made accessible to rigorous scientific study, as well as practically or politically manageable. It requires decisions about which environmental and AMR entities, interactions, and networks are most important targets for scientific analysis, across scales from molecular to the entire planet32. These decisions bring to the fore the importance of scientific language in framing, describing, and representing these processes.
The relationship between language, science, and society is a long-standing topic of research for social scientists and humanities scholars33. Within this work, language is a fundamental tool for thinking about and acting in the world. Language used by scientists influences scientific thought processes and designs, hypothesis development, methods and thus, the conclusions we can come to. How we represent the environmental dimensions of AMR thus influences how science is done and understood34. This includes what science is funded, where and what is sampled, choices over data processing and analysis, and consequently, the picture that is collectively developed through rigorous scientific research about the environmental dimensions of AMR, the challenges it poses to human and animal health, and the need for potential surveillance, mitigatory and regulatory actions.
A prominent example of the study of use of language in AMR is the use of war or eschatological metaphors such as the ‘war on superbugs’ or the ‘antibiotic apocalypse’ within public media and science communication35,36,37. But other terminology used to represent microbes and antimicrobial resistance, have largely escaped broader scrutiny38. In this piece we specifically study the terms ‘hotspot’, ‘reservoir’ and ‘pristine’ identified in previous work as significant representations in the field39. These words sit within a broad spectrum of descriptive scientific language that extends from the precise, e.g., ‘NDM-1 metallo-beta-lactamase’, to less-precise terminology, e.g., ‘hotspot’. Even widely used terms such as AMR, antibiotic resistance or drug resistance have different meanings, significance and framings depending on the community and context within which they are used40,41. For example, ‘resistance’ can be seen by medical professionals as treatment failure42, or by ecologists as a strain having selective advantage43. This imprecise language can focus attention towards areas that are thought to be most important, but risks overemphasis of certain sites or obscuring important dimensions of the phenomena and the hazards they present.
This Perspective is specifically aimed at natural scientists who work in the field of AMR in the environment. We wish to provide a launching point for considering the language used to represent the environmental dimensions of AMR and reflect on its broader consequences for both science and governance. This article assesses two key questions in relation to these terms: (1) how does the scientific community (of which we are part) use these terms in our publications; and (2) how do these terms shape research being conducted in the environment?
Unpacking environmental terminology
We start by providing an initial definition of each of the three terms. This is followed by an analysis and discussion of how they are used in the literature.
Hotspots
Since the 1950s, epidemiological approaches sought to identify spatial clusters or concentrations of illness or infectious disease44. In the 1980s and 90s, AMR research mainly referred to ‘genetic hotspots’45. This changed when ‘hotspot’ was more explicitly popularised around the early 2000s with calls to target infectious disease hotspots in global public health policy and research46. This concept is a spatial metaphor that represents environments where there is an elevated concentration of pollutants, infectious agents, or possibilities for disease transmission and thus with the highest risk of producing carcinogenic or pathogenic results46,47. Regarding the environmental dimensions of AMR, hotspots are areas in which the selection, accumulation, and transmission of AMR genes and bacteria are anticipated to be concentrated. The environmental AMR hotspot does not necessarily refer to heightened possibilities of infection per se, but also includes sites where there is an abundance of both microbial life and antimicrobial pollutants.
Reservoirs
In medicine, the notion of the disease ‘reservoir’ emerged around the 1900s and was linked to colonial authorities’ efforts to control disease, human populations, and relations between humans and animals48. It was more widely popularised in the 1930s due to the development of ecological approaches to understanding disease outbreaks44. Its use in relation to AMR dates back to at least the late 1950s49. The reservoir conceptualizes a ‘vessel’, or ‘source’, whether this is a population of organisms or an environment, which harbors pathogens and/or transmits them to an at-risk population. The pathogenic potential of the reservoir is always present but nascent, requiring a moment of transmission to mobilize that potential. In the context of AMR, the ‘reservoir’ partially differs, because it is often conceptualized in relation to genes, rather than pathogenic bacteria: non-pathogenic environmental bacteria are a source of diverse resistance genes that can be mobilized into bacterial pathogens39. This genetic pool is framed as a latent environmental threat50 or ‘resistome’.
Pristine environments
The idea of ‘pristine’ environments originated in late 19th century narratives about the Americas as having been largely pristine, uninhabited natural landscapes prior to European arrival51. Although this is a debunked proposition, the notion of pristine environments has since become important to environmental and biodiversity science52,53. ‘Pristine’ environments are thus imagined as untouched by human activity, and in the case of AMR, less exposed to antibiotic, bacterial or genetic pollutants that might result in elevated levels of AMR39. Pristine environments are positioned as a comparator, enabling assessment of a natural baseline, background or benchmark against which more contaminated areas can be compared54.
Literature Analysis of the use of Hotspot, Reservoir and Pristine
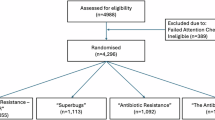
To assess how these terms are used within published research into the environmental dimensions of AMR, we used a mini-scoping review following PRISMA guidelines (Supplementary Note 1; Supplementary Figure 1; Supplementary Table 1). The review allowed a rapid assessment of appropriate studies suitable for the aims of the research, is transparent and reproducible, but is not comprehensive of the full set of terminology that could have been considered, nor does it consider (relevant) articles that use none of these terms. The mini-scoping review identified 60 articles containing at least one of the key search terms for detailed assessment (Supplementary Table 2).
Hotspot and reservoir are the most popular terms, often being used in the same papers, with pristine being used the least (Fig. 1). Out of the 60 papers 42 used hotspot at least once, 44 used the word reservoir at least once, and 12 used pristine at least once.

Hotspot and Reservoir are the more commonly used terms, with many papers making use of both terms.
Use of Hotspot in the Literature
‘Hotspot’ appeared in 42 of the 60 articles. The most common use of ‘hotspot’ is to describe a place or area55, including pharmaceutical manufacturing sites, sewage and wastewater56,57,58, wastewater treatment plants (WWTPs)59,60,61,62,63,64,65,66,67,68,69,70,71, biofilm72, aquaculture73, livestock farms74, paddy soils75, and slaughterhouses76. Thus the term hotspot encompasses a variety of spatial scales from aquaculture or farm sites to biofilms. However, the characteristics of the hotspot can vary: to justify research in a particular location e.g., in a ‘recognized’ hotspot such as a WWTP; as an objective of the work e.g., to identify hotspots within aquatic systems; or as a hypothesis to be tested e.g., slaughterhouses are a potential AMR hotspot77. The hotspot is, therefore, also a possible conclusion, the results of analysis confirming or refuting the hypothesis or establishing a new site as a potential hotspot.
Complicating the hotspot further is its use to describe processes. ‘Consumption’ was identified as a hotspot for AMR when, for example, products containing antimicrobials were used, or residues in food could encourage AMR when encountering the human microbiota55. ‘Disposal’ was another stage when antimicrobials could be introduced to soil, water and air, stimulating selection and transmission78. While these processes map onto the spatial hotspots identified above (farm or WWTP), the emphasis is shifted away from the spatial and towards a set of acts or practices.
In contrast, the notion of certain places as hotspots has been challenged by some scientists due to conflicting empirical results. Rather than confirming the provenance of a hotspot, the presentation of empirical findings from hypothesised ‘hotspots’ often complicates such ready characterization with ambiguous results79,80 and the presence of confounding factors81. Other work has been more explicit, arguing that WWTPs and animal slurries are not necessarily hotspots82,83.
Thus the ‘hotspot’ provides quite different ideas about where and what is important to sample and study. These ideas also communicate different responsibilities and priorities, e.g., emphasising those responsible for antimicrobial pollution, or on individuals entering areas of exposure. Including biofilms as hotspots72 raises challenging questions about the scale of study necessary to comprehend the hotspot accurately; is the challenge to comprehend AMR in relation to the farm as a whole, or just the biofilms on said farm? The inclusion of acts of antibiotic consumption and disposal is arguably an effort to refine the hotspot to identify times in which the concentration of antibiotics or potential for AMR selection or transmission is higher at a particular site, such as an aquaculture farm. However, this suggests that antibiotic concentration is the key factor in defining a hotspot, neglecting the enduring effects of consumption or disposal, including accumulation which could be just as significant.
Use of Reservoir in the Literature
‘Reservoir’ appeared in 44 of the 60 articles. Similar to the hotspot, the reservoir can be presented as a justification and an objective of the work e.g., to identify specific reservoirs within aquatic systems that might be of concern or require more research. Specifically, ‘reservoir’ is used to describe places and organisms where antimicrobial resistant and multi-drug resistant bacterial strains could be isolated in research and from which they could spread further into other environments84.
The reservoir is often situated as the background, the ubiquitous presence of microbes that could harbor ARGs, upon which hotspots which might mobilize these ARGS, could be identified. This is especially the case with aquatic systems61,85,86,87,88, commonly referred to as a reservoir because they harbor taxonomically large and genetically varied systems, with the potential for selection and subsequent transmission of ARGs that could contribute to AMR. Therefore, the task is to understand this aquatic reservoir as a whole and identify the ‘hotspots’ within it, e.g., aquaculture or WWTPs55,56,73. Following this, the reservoir is, therefore, sometimes a defined position within a broader system that is situated as a ‘distinct’ reservoir of interest: river sediments89,90, surface waters85, soil, animal manures and slurries75,77,91, or activated sludge87,92,93.
Wild animals were also categorized as reservoirs in these articles, be these wild birds94,95, wild owls96 or wild ungulates97,98. These organisms contain distinct microbiota and encounter diverse environments, including those polluted directly or indirectly with antimicrobial pollutants or ARGs. But in contrast to the above framing in which the reservoir is a place, animal reservoirs are organisms or living vectors that circulate ARGs and resistant bacteria between people, agricultural animals and the environment94.
A very different and widespread use for ‘reservoir’ is to discuss AMR at a microbiological level. Papers identified Phyla as reservoirs99, phages as reservoirs100, Bacteroides as reservoirs65, and specific microbial species as reservoirs101. The reservoir, therefore, attempts to communicate highly varied ideas about the nature of the environmental dimensions of AMR. Is the reservoir everywhere, or is it particular organisms, sites or microbes? Each of these raises quite distinct questions about the type of research necessary, the important dynamics of study and, following this, how the environmental dimensions might be practically managed and understood.
Use of Pristine in the Literature
Of the 12 articles that used the word ‘pristine’, 8 used it to designate a site to take samples from for comparison with places deemed contaminated59,90,91,95,102,103,104,105, including rivers or preserved or controlled soils, particularly those that have not been amended with manures containing antibiotics. The assumption is that pristine environments can offer a window into undisturbed microbial communities, allowing us to identify the changes in other locations due to human-caused pollution.
Consequently, pristine appeared most often in relation to methods and research design102. This is not to say that pristine means a lack of ARGs, though. Indeed, ARGs are expected to be naturally occurring as reflected in some conceptualizations of the reservoir. Following this, some studies used the concept of pristine to justify sampling in isolated places, for example, Antarctica soil and deep-sea sediments for comparison90.
The terminology of a pristine environment was used directly in scientific practice and data interpretation, specifically, how to establish proper comparators and identify control or blank samples. In contrast to the hotspot, which was re-established in new places in response to ambiguous results, the notion of a pristine environment faces a more fundamental challenge to its providence, namely, the difficulty of finding a genuinely pristine place in a polluted world.
The Uses of Hotspot and Reservoir in the Same Paper
‘Hotspot’ and ‘reservoir’ were both used in 24 papers, the largest point of overlap (Fig. 1). In some articles, reservoir and hotspot were used alongside each other, but are used to describe different spaces and concepts. For example one article85 described an aquatic environment as a reservoir with a distinct genetic community; but also as an important place for transmission of AMR genes, a characteristic more often attributed to the hotspot in literature39. Some articles have also used these terms to refer to different environments, for example calling a WWTP a hotspot and a biofilm a reservoir69; or identifying a clinical facility as a hotspot and describing surface waters as a reservoir74.
Other studies use hotspot and reservoir comparably. For example, in the same article, ’hotspot’ is first described as WWTPs90, but then biofilms are identified using both ‘hotspot’ and ‘reservoir’, which emphasizes how these terms are interlinked. Other papers use these two terms completely interchangeably57,106, e.g., to describe animal farms98. That paper, as with many others identified in this study, was written by scientists for whom English might be presumed to be an additional language, which raises additional questions about how these terms are used and understood in a global linguistic context.
Reflections and Recommendations
Our analysis highlights that there is significant diversity within the representation of the hotspot and the reservoir that is then folded into a singular specification. Indeed, their diverse and imprecise use emphasizes that the meaning and understanding of these terms can be specific to the author, leaving the reader the challenge of interpreting what the author is attempting to portray. Such flexibility is arguably part of the appeal of the words ‘hotspot’ and ‘reservoir’, and their utility for both justifying the locations of scientific study and interpreting conclusions. But it also collapses diverse phenomena with different temporal and spatial scales into a common representation that masks the ontological complexity of the problem of AMR in the environment. Furthermore, we note that although we have examined a sub-section of academic literature, these terms circulate more broadly in scientific and popular material, from newspaper articles107,108 to conference posters and presentations, as well as calls for funding.
These representations also seek to enclose the problem, so that AMR is seen as a problem that travels, rhetorically at least, between discrete places, out of an environmental hotspot or reservoir into (say) clinical settings or into otherwise pristine locations, whilst alluding to forms of spatial control that restrict flows between them as the evident solution. Our analysis of ‘pristine’ suggests the incorrectness of this framing, because said locations also consistently contain ARGs and ARBs. AMR is a phenomenon that is produced through interactions between numerous interrelated biological and social drivers109, and is therefore challenging to segregate via technological or managerial interventions that do not address these systemic drivers.
Furthermore, there is a risk that ingrained ways of thinking about the world limit our research and potential interventions through narrowed thought patterns arising from uncritical use of language; research based on these ideas may undoubtedly be of value, but we may miss other valuable approaches. One example of how these framings might make a practical difference is in surveillance: the hotspot/reservoir/pristine framing could lead to more focused surveillance on areas conceived as problem or comparator sites; alternative framings could lead to unbiased surveillance programmes e.g., based on a spatial grid. These two framings would produce different evaluations of the prevalence of AMR.
As we have shown, AMR is more complex and varied than can be reasonably described by the representations we have examined in this paper. Based on our analysis, we suggest four terms that could be used more precisely to describe the AMR hazard at sampling locations: prevalence, transmission, evolution, and connectivity (Table 1). Our contention is that these terms could be used either instead of the terms hotspot, reservoir and pristine, amongst other metaphors, or, alternatively, form the basis for justifying or clarifying the use of one or more of those terms, whilst also capturing key facets of AMR as a phenomenon and challenge.
To show how these terms might be used, we have taken a number of examples from our work and others’ that might fit the description of hotspot, reservoir or pristine, and assessed them against the five criteria we have suggested (Table 2).
Given the information in Table 2, if one wanted to refer to (say) the slurry tank as a ‘reservoir’, the low-income country water system as a ‘hotspot’ or the Arctic as ‘pristine’, this could now be done while attributing precise meanings to those terms. Alternatively, those terms could be avoided altogether, and replaced with precise terms with measurable attributes.
To conclude, the world is not composed of neatly bound spaces; it contains a continuous gradient of prevalence, hazard, transmission, evolution, and connectivity. Our intention is to establish some potential grounds for clarifying the dynamics of a particular place of study, and the basis for making judgements and assessments about where it sits on such a continuum.
Data availability
This article contains no new data. The list of papers retrieved in the scoping review and data extracted from them are available in Supplementary Table 2.
Change history
13 May 2025
A Correction to this paper has been published: https://doi.org/10.1038/s44259-025-00114-2
References
Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet Lond. Engl. 399, 629–655 (2022).
O’Neill, J. Tackling drug resistant infections globally: final report and recommendations. Technical report, (2016).
World Health Organization. WHO integrated global surveillance on ESBL-producing E. coli using a “One Health” approach: implementation and opportunities. (2021).
Kümmerer, K. Antibiotics in the aquatic environment–a review–part I. Chemosphere 75, 417–434 (2009).
Davies, R. & Wales, A. Antimicrobial resistance on farms: a review including biosecurity and the potential role of disinfectants in resistance selection. Compr. Rev. Food Sci. Food Saf. 18, 753–774 (2019).
Zhang, A. et al. β-lactam resistance genes in bacteriophage and bacterial DNA from wastewater, river water, and irrigation water in Washington State. Water Res. 161, 335–340 (2019).
Waśko, I., Kozińska, A., Kotlarska, E. & Baraniak, A. Clinically relevant β-lactam resistance genes in wastewater treatment plants. Int. J. Environ. Res. Public. Health 19, 13829 (2022).
Bengtsson-Palme, J. & Larsson, D. G. J. Concentrations of antibiotics predicted to select for resistant bacteria: proposed limits for environmental regulation. Environ. Int. 86, 140–149 (2016).
Arya, S. et al. Towards a general model for predicting minimal metal concentrations co-selecting for antibiotic resistance plasmids. Environ. Pollut. 275, 116602 (2021).
Poirel, L., Lartigue, M.-F., Decousser, J.-W. & Nordmann, P. ISEcp1B-mediated transposition of blaCTX-M in Escherichia coli. Antimicrob. Agents Chemother. 49, 447–450 (2005).
Jiao, Y.-N., Chen, H., Gao, R.-X., Zhu, Y.-G. & Rensing, C. Organic compounds stimulate horizontal transfer of antibiotic resistance genes in mixed wastewater treatment systems. Chemosphere 184, 53–61 (2017).
Wang, Y. et al. Non-antibiotic pharmaceuticals promote the transmission of multidrug resistance plasmids through intra- and intergenera conjugation. ISME J. 15, 2493–2508 (2021).
Pitout, J. D. D., Nordmann, P., Laupland, K. B. & Poirel, L. Emergence of Enterobacteriaceae producing extended-spectrum β-lactamases (ESBLs) in the community. J. Antimicrob. Chemother. 56, 52–59 (2005).
Duran, G. M. & Marshall, D. L. Ready-to-eat shrimp as an international vehicle of antibiotic-resistant bacteria. J. Food Prot. 68, 2395–2401 (2005).
Liu, Y.-Y. et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect. Dis. 16, 161–168 (2016).
Séveno, N. A. et al. Occurrence and reservoirs of antibiotic resistance genes in the environment. Rev. Res. Med. Microbiol. 13, 15 (2002).
Martínez, J. L. Antibiotics and antibiotic resistance genes in natural environments. Science 321, 365–367 (2008).
Singer, A. C., Shaw, H., Rhodes, V. & Hart, A. Review of antimicrobial resistance in the environment and its relevance to environmental regulators. Front. Microbiol. 7, 1728 (2016).
Wright, G. D. Antibiotic resistance in the environment: a link to the clinic? Curr. Opin. Microbiol. 13, 589–594 (2010).
Olanrewaju, O. S., Molale-Tom, L. G., Kritzinger, R. K. & Bezuidenhout, C. C. Genome mining of Escherichia coli WG5D from drinking water source: unraveling antibiotic resistance genes, virulence factors, and pathogenicity. BMC Genomics 25, 263 (2024).
Samtiya, M., Matthews, K. R., Dhewa, T. & Puniya, A. K. Antimicrobial resistance in the food chain: trends, mechanisms, pathways, and possible regulation strategies. Foods Basel Switz. 11, 2966 (2022).
Leonard, A. F., Morris, D., Schmitt, H. & Gaze, W. H. Natural recreational waters and the risk that exposure to antibiotic resistant bacteria poses to human health. Curr. Opin. Microbiol. 65, 40–46 (2022).
Hulscher, M. E. J. L., Grol, R. P. T. M. & van der Meer, J. W. M. Antibiotic prescribing in hospitals: a social and behavioural scientific approach. Lancet Infect. Dis. 10, 167–175 (2010).
McNulty, C. A. M., Boyle, P., Nichols, T., Clappison, P. & Davey, P. Don’t wear me out–the public’s knowledge of and attitudes to antibiotic use. J. Antimicrob. Chemother. 59, 727–738 (2007).
Holloway, K. A. Promoting the rational use of antibiotics. Reg. Health Forum 15, 122–130 (2011).
Podolsky, S. The Antibiotic Era: Reform, Resistance, and the Pursuit of a Rational Therapeutics. (John Hopkins University Press, Baltimore, MA, 2015).
Chandler, C. I. R. Current accounts of antimicrobial resistance: stabilisation, individualisation and antibiotics as infrastructure. Palgrave Commun. 5, 1–13 (2019).
Bengtsson-Palme, J., Kristiansson, E. & Larsson, D. G. J. Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol. Rev. 42, fux053 (2018).
Ding, G.-C. et al. Dynamics of soil bacterial communities in response to repeated application of manure containing sulfadiazine. PLoS ONE 9, e92958 (2014).
Greenfield, B. K. et al. Modeling the emergence of antibiotic resistance in the environment: an analytical solution for the minimum selection concentration. Antimicrob. Agents Chemother. 62, https://doi.org/10.1128/aac.01686-17 (2018).
Larsson, D. G. J., de Pedro, C. & Paxeus, N. Effluent from drug manufactures contains extremely high levels of pharmaceuticals. J. Hazard. Mater. 148, 751–755 (2007).
Bengtsson-Palme, J. et al. Towards monitoring of antimicrobial resistance in the environment: For what reasons, how to implement it, and what are the data needs? Environ. Int. 178, 108089 (2023).
McLeod, C. & Nerlich, B. Synthetic biology, metaphors and responsibility. Life Sci. Soc. Policy 13, 13 (2017).
van der Hel, S., Hellsten, I. & Steen, G. Tipping Points and Climate Change: Metaphor Between Science and the Media. Environ. Commun. 12, 605–620 (2018).
Nerlich, B. ‘The post-antibiotic apocalypse’ and the ‘war on superbugs’: catastrophe discourse in microbiology, its rhetorical form and political function. Public Underst. Sci. Bristol Engl. 18, 574–588 (2009).
Bouchoucha, S. L., Whatman, E. & Johnstone, M.-J. Media representation of the antimicrobial resistance (AMR) crisis: an Australian perspective. Infect. Dis. Health 24, 23–31 (2019).
Walker, I. F. Beyond the military metaphor: comparing antimicrobial resistance and the COVID-19 pandemic in the United Kingdom. Med. Anthropol. Theory 7, 261–272 (2020).
Kamenshchikova, A., Wolffs, P. F. G., Hoebe, C. J. P. A., Penders, J. & Horstman, K. Metaphors of foreign strangers: antimicrobial resistance in biomedical discourses. Sci. Cult. 32, 294–314 (2023).
Helliwell, R., Raman, S. & Morris, C. Environmental imaginaries and the environmental sciences of antimicrobial resistance. Environ. Plan. E Nat. Space 4, 1346–1368 (2021).
Wind, L. L. et al. Finding what is inaccessible: antimicrobial resistance language use among the one health domains. Antibiotics 10, 385 (2021).
Krockow, E. M., Cheng, K. O., Maltby, J. & McElroy, E. Existing terminology related to antimicrobial resistance fails to evoke risk perceptions and be remembered. Commun. Med. 3, 149 (2023).
Tsai, H.-C. et al. High rate of HIV-1 drug resistance in treatment failure patients in Taiwan, 2009-2014. Infect. Drug Resist. 10, 343–352 (2017).
Phan, M.-D. et al. Plasmid-mediated ciprofloxacin resistance imparts a selective advantage on Escherichia coli ST131. Antimicrob. Agents Chemother. 66, e02146-21.
Honigsbaum, M. ‘Tipping the Balance’: Karl Friedrich Meyer, latent infections, and the birth of modern ideas of disease ecology. J. Hist. Biol. 49, 261–309 (2016).
Murray, B. E. Problems and mechanisms of antimicrobial resistance. Infect. Dis. Clin. North Am. 3, 423–440 (1989).
Lessler, J., Azman, A. S., McKay, H. S. & Moore, S. M. What is a Hotspot Anyway? Am. J. Trop. Med. Hyg. 96, 1270 (2017).
Brown, H. & Kelly, A. H. Material proximities and hotspots: toward an anthropology of viral hemorrhagic fevers. Med. Anthropol. Q. 28, 280–303 (2014).
da Silva, M. A. D., French, O., Keck, F. & Skotnes-Brown, J. Introduction: disease reservoirs: from colonial medicine to one health. Med. Anthropol. (2023).
Knight, V., White, A., Hemmerly, T. & Martin, M. P. Studies on the Origin of Drug-Resistant Staphylococci in a Mental Hospital. Trans. Am. Clin. Climatol. Assoc. 70, 30–48 (1959).
Inda-Díaz, J. S. et al. Latent antibiotic resistance genes are abundant, diverse, and mobile in human, animal, and environmental microbiomes. Microbiome 11, 44 (2023).
Denevan, W. M. The Pristine Myth: The Landscape of the Americas in 1492. Ann. Assoc. Am. Geogr. 82, 369–385 (1992).
Clement, C. R. & Junqueira, A. B. Between a pristine myth and an impoverished future. Biotropica 42, 534–536 (2010).
Wooley, C. The Myth of the “Pristine Environment”: past human impacts in prince william sound and the Northern Gulf of Alaska. Spill Sci. Technol. Bull. 7, 89–104 (2002).
Rothrock, M. J. Jr. et al. How should we be determining background and baseline antibiotic resistance levels in agroecosystem research? J. Environ. Qual. 45, 420–431 (2016).
Brunton, L. A. et al. Identifying hotspots for antibiotic resistance emergence and selection, and elucidating pathways to human exposure: application of a systems-thinking approach to aquaculture systems. Sci. Total Environ. 687, 1344–1356 (2019).
Goh, S. G. et al. A new modelling framework for assessing the relative burden of antimicrobial resistance in aquatic environments. J. Hazard. Mater. 424, 127621 (2022).
Voigt, A. M. et al. Association between antibiotic residues, antibiotic resistant bacteria and antibiotic resistance genes in anthropogenic wastewater: an evaluation of clinical influences. Chemosphere 241, 125032 (2020).
Paulshus, E. et al. Diversity and antibiotic resistance among Escherichia coli populations in hospital and community wastewater compared to wastewater at the receiving urban treatment plant. Water Res. 161, 232–241 (2019).
Ewbank, A. C. et al. Seabirds as anthropization indicators in two different tropical biotopes: A One Health approach to the issue of antimicrobial resistance genes pollution in oceanic islands. Sci. Total Environ. 754, 142141 (2021).
Di Cesare, A. et al. Contribution of plasmidome, metal resistome and integrases to the persistence of the antibiotic resistome in aquatic environments. Environ. Pollut. Barking Essex 297, 118774 (2022).
Govender, R. et al. Identification, antibiotic resistance, and virulence profiling of Aeromonas and Pseudomonas species from wastewater and surface water. Environ. Monit. Assess. 193, 294 (2021).
Hassen, B. et al. Genetic characterization of ESBL-producing Escherichia coli and Klebsiella pneumoniae isolated from wastewater and river water in Tunisia: predominance of CTX-M-15 and high genetic diversity. Environ. Sci. Pollut. Res. 27, 44368–44377 (2020).
Kutilova, I. et al. Extended-spectrum beta-lactamase-producing Escherichia coli and antimicrobial resistance in municipal and hospital wastewaters in Czech Republic: Culture-based and metagenomic approaches. Environ. Res. 193, 110487 (2021).
Makowska, N., Koczura, R. & Mokracka, J. Class 1 integrase, sulfonamide and tetracycline resistance genes in wastewater treatment plant and surface water. Chemosphere 144, 1665–1673 (2016).
Manoharan, R. K., Srinivasan, S., Shanmugam, G. & Ahn, Y.-H. Shotgun metagenomic analysis reveals the prevalence of antibiotic resistance genes and mobile genetic elements in full scale hospital wastewater treatment plants. J. Environ. Manage. 296, 113270 (2021).
Furness, L. E., Campbell, A., Zhang, L., Gaze, W. H. & McDonald, R. A. Wild small mammals as sentinels for the environmental transmission of antimicrobial resistance. Environ. Res. 154, 28–34 (2017).
Lorenzo, P. et al. Antibiotic resistance in urban and hospital wastewaters and their impact on a receiving freshwater ecosystem. Chemosphere 206, 70–82 (2018).
Guo, J., Li, J., Chen, H., Bond, P. L. & Yuan, Z. Metagenomic analysis reveals wastewater treatment plants as hotspots of antibiotic resistance genes and mobile genetic elements. Water Res. 123, 468–478 (2017).
Brienza, M. et al. Reclaimed wastewater reuse in irrigation: Role of biofilms in the fate of antibiotics and spread of antimicrobial resistance. Water Res. 221, 118830 (2022).
Leroy-Freitas, D. et al. Exploring the microbiome, antibiotic resistance genes, mobile genetic element, and potential resistant pathogens in municipal wastewater treatment plants in Brazil. Sci. Total Environ. 842, 156773 (2022).
Rodríguez, E. A., Ramirez, D., Balcázar, J. L. & Jiménez, J. N. Metagenomic analysis of urban wastewater resistome and mobilome: a support for antimicrobial resistance surveillance in an endemic country. Environ. Pollut. Barking Essex 276, 116736 (2021).
Kaszab, E. et al. Groundwater, soil and compost, as possible sources of virulent and antibiotic-resistant Pseudomonas aeruginosa. Int. J. Environ. Health Res. 31, 848–860 (2021).
Lu, J. et al. Responses of sediment resistome, virulence factors and potential pathogens to decades of antibiotics pollution in a shrimp aquafarm. Sci. Total Environ. 794, 148760 (2021).
Hsu, C.-Y. et al. A potential association between antibiotic abuse and existence of related resistance genes in different aquatic environments. Water. Air. Soil Pollut. 226, 2235 (2014).
Cui, H. et al. Co-occurrence of genes for antibiotic resistance and arsenic biotransformation in paddy soils. J. Environ. Sci. 125, 701–711 (2023).
Savin, M. et al. Antibiotic-resistant bacteria and antimicrobial residues in wastewater and process water from German pig slaughterhouses and their receiving municipal wastewater treatment plants. Sci. Total Environ. 727, 138788 (2020).
Zhou, M. et al. Evolution and distribution of resistance genes and bacterial community in water and biofilm of a simulated fish-duck integrated pond with stress. Chemosphere 245, 125549 (2020).
Kunhikannan, S. et al. Environmental hotspots for antibiotic resistance genes. Microbiology Open 10, e1197 (2021).
Caucci, S. et al. Seasonality of antibiotic prescriptions for outpatients and resistance genes in sewers and wastewater treatment plant outflow. FEMS Microbiol. Ecol. 92, fiw060 (2016).
Flach, C.-F., Genheden, M., Fick, J. & Joakim Larsson, D. G. A comprehensive screening of Escherichia coli isolates from Scandinavia’s largest sewage treatment plant indicates no selection for antibiotic resistance. Environ. Sci. Technol. 52, 11419–11428 (2018).
Bengtsson-Palme, J. et al. Elucidating selection processes for antibiotic resistance in sewage treatment plants using metagenomics. Sci. Total Environ. 572, 697–712 (2016).
Quintela-Baluja, M. et al. Spatial ecology of a wastewater network defines the antibiotic resistance genes in downstream receiving waters. Water Res. 162, 347–357 (2019).
Baker, M. et al. Antimicrobial resistance in dairy slurry tanks: a critical point for measurement and control. Environ. Int. 107516 https://doi.org/10.1016/j.envint.2022.107516 (2022).
Waseem, H., Ali, J., Jamal, A. & Ali, M. I. POTENTIAL DISSEMINATION OF ANTIMICROBIAL RESISTANCE FROM SMALL SCALE POULTRY SLAUGHTERHOUSES IN PAKISTAN. Appl. Ecol. Environ. Res. 17, 3049–3063 (2019).
Asaduzzaman, M. et al. Spatiotemporal distribution of antimicrobial resistant organisms in different water environments in urban and rural settings of Bangladesh. Sci. Total Environ. 831, 154890 (2022).
Rowe, W. et al. Comparative metagenomics reveals a diverse range of antimicrobial resistance genes in effluents entering a river catchment. Water Sci. Technol. J. Int. Assoc. Water Pollut. Res. 73, 1541–1549 (2016).
Wang, M., Shen, W., Yan, L., Wang, X.-H. & Xu, H. Stepwise impact of urban wastewater treatment on the bacterial community structure, antibiotic contents, and prevalence of antimicrobial resistance. Environ. Pollut. Barking Essex 231, 1578–1585 (2017).
Callahan, M. T., Van Kessel, J. A. & Micallef, S. A. Salmonella enterica recovery from river waters of the Maryland Eastern Shore reveals high serotype diversity and some multidrug resistance. Environ. Res. 168, 7–13 (2019).
Devarajan, N. et al. Antibiotic resistant Pseudomonas spp. in the aquatic environment: a prevalence study under tropical and temperate climate conditions. Water Res. 115, 256–265 (2017).
Chen, H., Bai, X., Jing, L., Chen, R. & Teng, Y. Characterization of antibiotic resistance genes in the sediments of an urban river revealed by comparative metagenomics analysis. Sci. Total Environ. 653, 1513–1521 (2019).
He, L.-Y. et al. Discharge of swine wastes risks water quality and food safety: antibiotics and antibiotic resistance genes from swine sources to the receiving environments. Environ. Int. 92–93, 210–219 (2016).
Zhou, S., Zhu, Y., Yan, Y., Wang, W. & Wang, Y. Deciphering extracellular antibiotic resistance genes (eARGs) in activated sludge by metagenome. Water Res. 161, 610–620 (2019).
Cheng, X., Xu, J., Smith, G., Nirmalakhandan, N. & Zhang, Y. Metagenomic profiling of antibiotic resistance and virulence removal: activated sludge vs. algal wastewater treatment system. J. Environ. Manage. 295, 113129 (2021).
Torres, R. T., Cunha, M. V., Ferreira, H., Fonseca, C. & Palmeira, J. D. A high-risk carbapenem-resistant Pseudomonas aeruginosa clone detected in red deer (Cervus elaphus) from Portugal. Sci. Total Environ. 829, 154699 (2022).
Ewbank, A. C. et al. Extended-spectrum β-lactamase (ESBL)-producing Escherichia coli survey in wild seabirds at a pristine atoll in the southern Atlantic Ocean, Brazil: First report of the O25b-ST131 clone harboring blaCTX-M-8. Sci. Total Environ. 806, 150539 (2022).
Fuentes-Castillo, D. et al. Wild owls colonized by international clones of extended-spectrum β-lactamase (CTX-M)-producing Escherichia coli and Salmonella Infantis in the Southern Cone of America. Sci. Total Environ. 674, 554–562 (2019).
Torres, R. T. et al. Emergence of colistin resistance genes (mcr-1) in Escherichia coli among widely distributed wild ungulates. Environ. Pollut. Barking Essex 291, 118136 (2021).
Torres, R. T. et al. A walk on the wild side: wild ungulates as potential reservoirs of multi-drug resistant bacteria and genes, including Escherichia coli harbouring CTX-M beta-lactamases. Environ. Pollut. 306, 119367 (2022).
Borsetto, C. et al. Impact of sulfamethoxazole on a riverine microbiome. Water Res. 201, 117382 (2021).
Hayes, A. et al. Predicting selection for antimicrobial resistance in UK wastewater and aquatic environments: Ciprofloxacin poses a significant risk. Environ. Int. 169, 107488 (2022).
Roman, V. L. et al. Abundance and environmental host range of the SXT/R391 ICEs in aquatic environmental communities. Environ. Pollut. 288, 117673 (2021).
Muurinen, J. et al. Antibiotic resistomes and microbiomes in the surface water along the Code River in Indonesia reflect drainage basin anthropogenic activities. Environ. Sci. Technol. 56, 14994–15006 (2022).
Xiang, Q. et al. Spatial and temporal distribution of antibiotic resistomes in a peri-urban area is associated significantly with anthropogenic activities. Environ. Pollut. 235, 525–533 (2018).
Calero-Cáceres, W. & Balcázar, J. L. Antibiotic resistance genes in bacteriophages from diverse marine habitats. Sci. Total Environ. 654, 452–455 (2019).
Pruden, A., Arabi, M. & Storteboom, H. N. Correlation between upstream human activities and riverine antibiotic resistance genes. Environ. Sci. Technol. 46, 11541–11549 (2012).
Xin, H. et al. Animal farms are hot spots for airborne antimicrobial resistance. Sci. Total Environ. 851, 158050 (2022).
Harvey, F. Superbug hotspots emerging in farms across globe – study. The Guardian (2019).
Daniel, R. & Prickett, K. Rivers becoming ‘reservoirs of disease’ warn experts. BBC News (2024).
Rickard, H. et al. Antimicrobial resistance as a super wicked problem: how do we engage the public to be part of the solution. Infect. Prev. Pract. 5, 100314 (2023).
Acknowledgements
This work was supported by Antimicrobial Resistance Cross Council Initiative supported by the seven United Kingdom research councils (NE/N019881/1) and The Research Council of Norway (Project No. 320715).
Author information
Authors and Affiliations
Contributions
Conceptualization: R.H., C.M., S.R., D.J.S.; Investigation: I.E.; Writing—Original Draft: R.H., I.E., A.D.W., D.J.S.; Writing—Reviewing and Editing: R.H., A.D.W., C.M., A.S., D.T.L., D.J.S.; Visualization: I.E., D.J.S.; Supervision: C.M., S.R., D.J.S.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Helliwell, R., Ewin, I., Williams, A.D. et al. Rethinking the words hotspot reservoir and pristine in the environmental dimensions of antimicrobial resistance. npj Antimicrob Resist 3, 11 (2025). https://doi.org/10.1038/s44259-025-00080-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s44259-025-00080-9
