
Abstract
From pathogens and computer viruses to genes and memes, contagion models have found widespread utility across the natural and social sciences. Despite their success and breadth of adoption, the approach and structure of these models remain surprisingly siloed by field. Given the siloed nature of their development and widespread use, one persistent assumption is that a given contagion can be studied in isolation, independently from what else might be spreading in the population. In reality, countless contagions of biological and social nature interact within hosts (interacting with existing beliefs, or the immune system) and across hosts (interacting in the environment, or affecting transmission mechanisms). Additionally, from a modeling perspective, we know that relaxing these assumptions has profound effects on the physics and translational implications of the models. Here, we review mechanisms for interactions in social and biological contagions, as well as the models and frameworks developed to include these interactions in the study of the contagions. We highlight existing problems related to the inference of interactions and to the scalability of mathematical models and identify promising avenues of future inquiries. In doing so, we highlight the need for interdisciplinary efforts under a unified science of contagions and for removing a common dichotomy between social and biological contagions.
Similar content being viewed by others


The science of contagions
Arguably, no scientific concept encapsulates the human experience as much as contagions. That is in large part because the situations in which people use the word “contagion” are so varied, referring to any process where a property is passed from one or many agents to others. Mathematically, the concept of contagion then inspires different forms of branching processes1, dynamical systems2, cascade models3, and network systems4. These are used to model human genealogies5, teaching6,7, culture and language8, viral trends9,10, scientific ideas11, innovation12, rumors9,13, misinformation14,15, social movements16,17, and obviously infectious diseases2. Although all of these phenomena interact to shape human life, they are unfortunately often studied in isolation, one at a time. Even simple classifications, such as distinguishing biological and social contagions, may create a false dichotomy and obfuscate their complexity. For instance, epidemics are generally shaped by multiple factors drawn from biological sciences (pathogens, genetics, microbiome) to social sciences (information, culture, behavior) and anywhere in between (nutrition, life history, living environment). All of these factors can affect both the local mechanisms by which a contagion is transmitted and its global, population-level presentation that we get to observe.
How do contagions spread? For biological contagions like infectious diseases, our intuition tends to be that if a pathogen occurs in every positive case and is not found in negative cases, it is causally responsible for the contagion as it gets transmitted from one individual to the next. Memes, as self-replicating cultural elements, might play a similar role in social contagions18. This intuition is the basis for the 19th-century postulates of Robert Koch’s19, which established both microbiologic methods and the necessary scientific framework of causality. This framework ushered in the golden age of microbial discovery in which many of the pathogens responsible for humanity’s worst diseases (tuberculosis, cholera, anthrax, rabies, diphtheria) were identified. While initially useful, the one-pathogen, one-disease model ceases to accurately describe the way in which pathogens interact not only with each other but with the host, environment, and societies in which they exist.
Koch’s postulates are too simplistic as they imply that every contagion is caused by a single, sufficient, and necessary pathogen, which is contradicted by empirical evidence20,21. For example, many microbiota-related bacterial populations can improve immune defense and prevent infection by intestinal pathogens22. Similarly, genetic conditions can spread vertically (from parent to offspring) but also interact with pathogens transmitted horizontally (within a generation), such as resistance conferred against malaria by the gene responsible for sickle haemoglobin23. Pathogens might therefore be found in negative cases because some other spreading process is interacting with the contagion. Conversely, one can imagine that pathogens can also be found in negative cases because it has yet to interact with some other synergistic contagion; this is the case, for instance, with opportunistic pathogens24. We detail documented biological mechanisms of pathogen interactions further in Box 1.
Similar arguments can explain why the modeling of social contagions should also move away from the assumption that one pathogen equals one contagion25. Obviously, no single piece of information, no single meme, and no single idea spreads in a vacuum. Different people can adopt the same ideology for different reasons. Information, ideas, or behaviors are related to each other, and they interact with each other, obfuscating classical notions of causality. For example, can a single piece of information cause anyone to develop an anti-vaccination sentiment? Probably not26,27, especially since the discourse surrounding epidemics can be quite complicated and nuanced28.
There is a dire need for unifying the different contagion frameworks and moving beyond the paradigm of “isolated contagion” and the classic distinction between “simple” and “complex” contagions. Indeed, modeling of biological contagions often uses a paradigm called “simple contagions” which assumes a linear relationship between exposure and transmission, while modeling of social processes often assumes more general “complex contagions”. These complex contagions take different nonlinear functional forms to relate exposure to transmission29, and they can also rely on mechanisms from social diffusion12, sensing30, adaptation31, or learning32. This conventional approach to modeling biological or social contagions creates a false dichotomy; contagions almost always interact, and these interactions blur the distinction between contagions of different natures (and models thereof). Regardless of its nature and transmission mechanisms, a contagion is often shaped by multiple factors that can be biological, ecological, or social. The effects we wish to consider can be direct interactions between different contagious processes (e.g., two infectious diseases or rumors) or indirect interactions through important covariates (e.g., education or norms). These different types of effects are not always easy to distinguish, given that covariates like education, norms, or culture can themselves be considered as contagious at some level. We provide examples of how contagions interact across scales and scientific domains in Box 2.
The renewed interest in interacting contagions brings more to the table than new keywords like “syndemics” for synergistic epidemics and “infodemics” for epidemics of (mis-)information33. It also brings a fresh post-disciplinary perspective on the problem and a call for integrated efforts34.
In what follows, we do a targeted review of the techniques for modeling interaction contagions in an attempt to highlight some key elements from that literature and map a way forward. In the section “The physics of interacting contagions,” we first explain how, following lessons learned from other complex systems35,36, “more contagions are different contagions.” We discuss how the behavior of interacting contagions differs from the conventional wisdom built on models of individual contagions. This makes contagion forecasting, an inherently noisy process37, even more intractable in the face of unknown interactions. In the section “Modeling social contagions and their interactions,” we then review novel empirical and data-driven efforts, mostly coming from interacting cascades of social contagions on social media and in the science of stories. In the section “The spread of beliefs as interacting contagions,” we give one particularly potent example of how contagions can interact within hosts using the study and dynamics of beliefs. Finally, in the section “The ecology of interacting contagions,” we attempt to outline promising ecological perspectives that might help the science of interacting contagions avoid the trap of high model dimensionality.
The physics of interacting contagions
Mathematical modeling of contagions has a long history going back at least to the 1700s, with the creative work of Daniel Bernoulli38 among others. In Bernoulli’s work, a mathematical system of smallpox dynamics is developed, but already, the text highlights the interaction of the disease with indirect factors such as the age of individuals and the dynamics of inoculation as a method of prevention. Most modern mathematical models borrow a structure and a set of assumptions from a model called Susceptible-Infected-Removed (or susceptible-infectious-recovered, the SIR model) published in 1927 by Kermack and McKendrick39. In this model, infected agents transmit the contagion to their susceptible neighbors at a given rate and are removed from the dynamics through death or immunity at some other rate. One can easily generate variants of this model. For example, the SIS process assumes that the disease does not confer long-lasting immunity, such that recovered individuals return directly to the susceptible state. The SI process, in turn, assumes that there is no recovery at all.
Early studies of multiple contagions built on the SIR and SIS foundations to couple the dynamics of contagion with the evolutionary dynamics or cross-immunity between biological pathogens40,41 or the social process of disease awareness42. These new models often assumed that contagions were unaffected by each other 40, or that one was necessary for the other43, or that they were competing for a chance to spread41,44,45,46,47,48,49,50, or directly in opposition to each other42,51,52,53,54. These efforts have previously been reviewed in detail55. And while there have been studies on a broader definition of interactions between contagions56,57,58,59,60,61,62, the mathematical modeling community has not quite moved beyond the simplest case of two contagions interacting in simple, often symmetric, and deterministic ways. This is due in part to the richness of the behaviors that emerge even in this simple case, and in part because considering more interacting contagions makes the dimensionality of our models grow exponentially.
Importantly, the physics of interacting contagions is more than that of the simple sum of independent contagions, especially when contagions interact synergistically as they spread63,64,65,66,67,68,69,70. In a simple contagion model, say the SIR model described above, there is a monotonic relationship between the transmission rate and the final size of the contagion2. Interestingly, depending on the density of contacts in the population, there exists a critical value of transmissibility below which the expected size of the contagion is zero, and above which it increases monotonically. But importantly, this phase transition between a contagion-free state and an endemic contagion is typically continuous. That is not the case for synergistic64 or cooperative65 contagions, where two spreading processes can increase each other’s transmissibility. These models can assume, for example, that a contact between a contagious agent and a susceptible agent transmits at a fixed rate λ, or a higher rate \(\lambda ^{\prime} > \lambda\) if a second contagion is involved. This effect might be the same if the second contagion affects the contagious agent (e.g., a superspreader) or the susceptible agent (e.g., increased susceptibility). Regardless of the details, the system can get into a frustrated state where many contagious agents can only transmit the contagion to their susceptible neighbors if a second contagion also reaches them (see Fig. 1A). These frustrated transmissions are similar to latent heat in physical systems, leading to discontinuous phase transitions. Or, in this case, the discontinuous emergence of a large contagion as we tune the transmission rate (see Fig. 1B).

A Schematic representation of two synergistic contagions (blue and orange) spreading synergistically through a network. Co-infections, shown in pink, are needed to sustain the contagion and are aided by clustering. B Phase transition of various types of contagions. While independent contagions display continuous transitions, synergistic contagions can build up transmission potential, which leads to discontinuous transitions reminiscent of physical systems where latent heat accumulates. C Growth rate of synergistic (pink), independent (black), and antagonistic (green) contagions. Synergistic contagions tend to grow superexponentially since they get more likely to interact as they spread further.
There are other elements of conventional wisdom from simple independent contagions that do not transfer to the dynamics of interacting contagions. Features of the structure of the underlying contact network that tend to slow down contagions, like triangles or any form of clustering, can now hasten the spread of synergistic contagions64. The intuition behind this phenomenon is simple. For an independent contagion, the optimal structure on which to spread is a tree-like network where it is impossible to backtrack when starting from a single patient zero. In that case, all connections lead to new, and therefore susceptible, agents. Clustering of connections can instead trap a contagion and cause transmission efforts to be “wasted” on already contagious agents. However, synergistic contagions benefit from being kept together. On a tree-like network, a co-infected patient zero might transmit one contagion to one branch of the tree and another to a different branch. These contagions might then not interact with one another for a long time and, therefore, not benefit from their synergy. A small amount of clustering helps them stay together.
A similar effect is found in parasitic contagions, where one contagion benefits from the presence of another contagion but hinders its transmission in return. This is the case, for example, of positive messaging around a negative contagion, like the spread of preventive awareness against an infectious disease. The awareness is more likely to spread when the disease is spreading, and individuals exposed to the disease are more likely to be receptive to relevant information. Conversely, the preventive message is likely to reduce the transmission risk of the disease. The awareness is thus a parasitic contagion to the epidemic71. With asymmetric interactions, clustering can hurt awareness more than the disease, leading again to a larger epidemic than expected without clustering (Table 1).
Finally, synergistic contagions can also spread superexponentially since the more contagions spread, the more likely they are to interact and benefit from their synergy, thus further accelerating their spread (see Fig. 1C). This dynamic clashes with most mechanistic prediction models69. As contagions spread and the number of contagious agents grows, the more likely co-infections become, such that positive interactions become more frequent and important. For a time series of either contagion, this simple statistical effect leads to what looks like an accelerating spread or a transmission rate that increases with time. Altogether, basic intuition built by focusing on a single pathogen might not hold when looking at an ecology of interacting contagions. New modeling approaches are needed. Thankfully, ideas from the social sciences might again come into play.

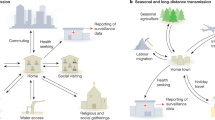
Biological and social contagions (here represented by colored viruses and different light bulbs, respectively) spread through contact networks and shared physical or digital environments. Contagions can interact directly in the environment or during contact. When an agent is exposed to new viruses or ideas, the potential transmission is mediated through other layers of interactions, now internal. New beliefs or ideas interact with an existing belief system. Likewise, new pathogens interact with an existing microbiome and immune system. These multi-layered interactions can all drive the observed dynamics of contagions195,196.
All of the features described above are also signatures of complex contagions, as introduced in the previous section. In complex contagions, also called frequency-dependent models, transmission depends nonlinearly on the number or frequency of exposures such that the growth rate of a contagion can vary as it spreads12,29. Recent results show that synergistic contagions can be mathematically indistinguishable from these nonlinear effects in complex contagions, such as the psychological phenomenon of social reinforcement69. Like peer pressure, this phenomenon means that a second exposure to an idea or behavior through a second source is more effective than the first exposure29,72. For example, ten friends telling you to read an article are more likely to convince you than one friend telling you the same thing ten times. That is usually not true for biological contagions, where our models assume a linear relationship between infection rate and exposure regardless of the source. Indeed, researchers often use simple linear contagions for biological applications and complex nonlinear contagions with reinforcement for social applications. Studies suggest the distinction breaks down once contagions interact.
In recent years, modeling studies have also broadened the concept of interaction structure and focused on networks with different (multilayer) interactions73 or with group (higher-order) interactions74. There has therefore been a rapid influx of studies generalizing these previous models. Given that nonlinear group interactions alone create apparent complex contagions, the phenomenology of higher-order and interacting contagions is only richer. Studies have looked at competing contagions in multilayer networks where different contagions spread on different sets of contact53,54,75,76, through higher-order dynamics77,78,79, and even both80,81,82,83,84. These extensions are natural, and in fact, some of the early studies on cooperating epidemics also assumed a multilayer structure85 or a group structure64.
The rich dynamics of interacting contagions pose an important challenge: Is it possible to measure contagion mechanisms without explicitly controlling for all possible interactions and covariates? Social reinforcement can be defined and measured by the increase in infection probability caused by repeated exposures compared to a null model of equivalent exposures. However, this measurement might be impossible to distinguish from interactions among pathogens. Did someone share a meme because many of their friends liked it, or had they previously shared a related meme? The flip side of this challenge is that measuring social reinforcement can then be used as a proxy for interactions between contagions. Recent work has taken this perspective to justify the need for tractable nonlinear models86. New effective models for interacting contagions are needed, especially since we know that countless contagions of biological and social nature interact at all times and that the dimensionality of our current models grows exponentially with the number of contagions involved.
Modeling social contagions and their interactions
Attempts to operationalize and model social contagion processes first appeared in the second half of the 20th century. In the 1950s, Katz and Lazarsfeld87 published “Personal Influence”87, a book which would become highly influential about the nature of influence. Their finding were based on their in-the-field study of decision making by women in the city of Decatur, Illinois. Their major contribution was to introduce the “two-step” model of influence, whereby media reached local opinion leaders who then influenced their friends and acquaintances. Though Katz and Lazarfeld were clear that their two-step model was a non-universal approximation of true social influence, the idea that there exists a class of special, influential people—and consequently the importance of identifying and engaging them for the means of large-scale social change to whatever ends—would become a hardened, enduring, and broadly popularized concept12,88,89. As the real-data-informed field of network science developed in the late 1990s, the notion of opinion leaders would be challenged with more sophisticated social network models90. Nevertheless, the two-step model moved the understanding of media influence beyond the traditional “hypodermic model,” revealing and elevating the role of social networks in shaping collective and individual opinions and beliefs.
In the 1960s, early considerations of how to mathematically operationalize social contagion fell to simply porting across the SIR model of mathematical epidemiology39. In a 1964 Nature article, Goffman and Newill explicitly cast the spread of an idea as akin to the spread of a disease91. It is worth reflecting on the scope and strength of their framing:
“For example, consider the development of psychoanalysis in the early part of this century. Freud was no less host to the infectious material of the ‘disease’ of psychoanalysis than the person carrying the organism capable of transmitting a cold, nor is his writing less of a ‘vector’ carrying the ‘infectious material’ than the mosquito as a carrier of malaria. Jung might represent an example of acquired resistance to the disease while the resistance of the medical community of Vienna could represent innate immunity. The development of the psychoanalytic movement …was in its way no less an ‘epidemic’ than the outbreak of influenza in 1917 and 1918.
One can argue similarly that Darwin and evolution, Cantor and set theory, Newton and mechanics, and so on, were examples of ‘epidemics’ in the world of scientific thought which were instigated by the introduction of a single infective into a population. The analogy is not restricted to science; for examples such as Christ, Buddha, Moses and Mohammed can be cited in the religious field …”
Of course, social contagions are inherently different from biological contagions as there are no “cultural pathogens” easily identified and cultured92. Instead, as recent studies suggest, social contagions are always shaped by other effects such as homophily or cultural93, human biases and demographic heterogeneity15, or because of dislike and distrust between subpopulations94.
Clear movement beyond SIR models began (at least) in the early 1970s, when Schelling introduced his physical, checkerboard-based model of self-organizing neighborhood segregation processes95. Schelling operationalized social contagion with the concept of thresholds: Individuals adopt (or reject) a characteristic (action, belief, behavior, etc.) based on the fraction of those around them with that characteristic or some other influential attribute. While simple to run, Schelling’s model was non-trivial to address analytically. In 1978, Granovetter showed that, absent any interaction structure, a mean-field threshold model produced informative stories of social contagion96,97,98,99. Like Schelling’s model, a seemingly moderate population could universally adopt a behavior. These models showed mechanistically how collective uniformity could arise not from individual uniformity but rather social following.
It is worth noting that while seemingly distinct, the essential models of biological and social contagion can be reconciled. For example, a generalized contagion model incorporating memory successfully interpolates between SIR-type models and threshold models100,101.
Abstract modeling of contagions, regardless of their nature, can only get us so far. Turning to data to validate these theories in specific domains is critical. A major, enduring problem with all empirical social network contagion is missing data, either through limited sampling of a social network (hence missing links and interactions) or due to interactions occurring outside of the sphere of observation (e.g., through direct text messages between individuals, or influence from other media across scales).
Modern social media platforms can control and monitor what people see and how they behave with high precision. But the algorithms behind major platforms are proprietary and hidden. Observational analyses of online behavior and properly formalized academic experiments promise an ethical way forward. Because of the massive data on offer, social media provides a unique lab bench to both characterize social contagion and test theoretical models.
As a first simple proxy for social contagion, one can look at cascades of re-sharing of the same content on social media (re-sharing a post, keyword, or a given URL). Under that lens, while co-infection data regarding biological contagions can be rare, social media are essentially a messy soup of countless interaction contagions102 shaped by each other and by the social networks that support them103. Therefore, it is not surprising that online social networks inspired some early studies of multiple spreading processes104. These processes can interact by competing or complementing each other105, and telling these mechanisms apart from noisy data can be subtle but is not impossible106,107. For example, Zarezade et al.107 highlight that incorporating interactions into models enhances their predictive accuracy of social media cascades, even when accounting for the added model parameters. In their model, a contagion spreads to susceptible individuals at a rate specified by a Hawkes process around each susceptible individual108, a self-exciting process that relates the transmission rate of a contagion to the sum of recent exposures to the contagion itself and to related contagions. Related work focused on competition between memes obtained similar results, showing that the competition for potential “hosts” and their attention is sufficient to qualitatively explain the broad diversity in popularity, lifetime, and activity of memes109. In that model, competition between memes occurs through a mechanism of limited attention, through which a susceptible agent is only really exposed to a finite number of memes in its recent memory. The general idea being that signatures of online behavior are a reflection of recent exposure to contagions, both online and in the real world, and both social and biological. In fact, a whole subfield of digital disease surveillance has also looked at the potential of using social media data as a proxy to track the spread of infectious diseases through keywords, posts, and searches related to a disease or its symptoms110.
Despite this abundance of data, monitoring of social contagions is complicated by the fact that the signature of any contagion can mutate quickly111. Early methods often focused on tracking specific hyperlinks or proper names112,113, which is appropriate for short timescale before new hyperlinks or names start describing the same information. Other approaches tend to aggregate many cascades, such as mixture models or dynamic topic models114, which by analogy would be similar to tracking respiratory illnesses without tracking individual virus families115. To follow a social contagion as it evolves, it is possible to track specific short phrases that are unlikely to mutate111, but this approach is hard to generalize to visual non-text memes. More recent work has therefore moved to multi-modal deep learning models to identify memes116, classify them in families117, and attempt to understand their relations118 or predict their potential for virality119.
Beyond the spreading of internet memes and the modeling of social contagions lies a developing, data-driven science of stories and beliefs. That stories matter profoundly to people and societies seems to be both unclear and obvious: Stories are portrayed by some as being just for entertainment, while by others as the core of being human120,121,122,123,124. Even so, the centrality of stories has gained ground, as has the conception and possibility of measuring stories through distant reading125,126,127,128,129,130,131,131,132. Understanding how stories develop, spread, interact, and compete with each other is of utmost importance to understanding social phenomena; from myths and conspiracy theories to hate speech and counter speech133. Operationalizing the measurement of stories and interactions between story elements is critical to advancing these aims (see Box 3).

The spread of beliefs as interacting contagions
Interacting contagions in social networks also differ from biological contagions as they often spread in signed social networks that explicitly distinguish between positive links (representing trust and favoritism, generally toward in-group members) and negative links (representing distrust and enmity, generally toward out-group members). Positive links facilitate the spread of contagions within the group of like-minded individuals, while negative links can lead to rejection or counter-adoption of ideas from distrusted sources. This sign-based framework provides essential structure for understanding how contagions compete across partisan divides. Recent dynamical modeling134 reveals that the relative strength of positive versus negative links determines whether the social network converges toward consensus or diverges toward polarization, explaining the rapid polarization of stance toward masking and lockdowns during the COVID-19 pandemic135.
While much of the work we have surveyed so far emphasizes the spread of contagions across agents, beliefs rarely spread in isolation from other beliefs and related cognitions such as knowledge, social norms, and emotions136,137. Therefore, beliefs and opinions (which we here use somewhat interchangeably138) spread not only across social networks but also within internal belief systems (Fig. 2). This interaction of contagions across levels makes the study of beliefs a prominent and powerful example of interacting contagions.
In social networks, the spread of some beliefs is often easier after a group has already been “infected” with a related set of beliefs. During the COVID-19 pandemic, people who grew up in the former East Germany and were thus more accustomed to the idea that the government can enforce certain behaviors were more likely to support the idea of mandatory vaccination139. Beliefs about whether abortion should be banned or not are strongly related to prior “infections” of a group by particular political and religious beliefs140. Beliefs about what is normal and desired in a group can change in line with shifting perceptions of what the majority of group members believe or do141, with examples ranging from the support for gay rights142 to the support for extreme political views143.
Beliefs also spread within individual minds, affecting existing beliefs and related cognitions and clearing the path for “invasions” of a belief system with other novel beliefs. For example, when one becomes skeptical about the safety of vaccinations, this can open doors to the development of skepticism for those supporting vaccination, such as scientists and the government. This distrust may make one more likely to accept further related beliefs and conspiracies144. Emotions can further facilitate belief spread within an individual’s mind. Even a temporary “infection” with fear of death can lead one to adopt dislike and prejudice towards beliefs and groups different than one’s own145.
Models of belief dynamics could profit from incorporating the effects of interacting socio-cognitive contagions. Many existing belief dynamics models focus on the spread of only one belief at a time138,146,147,148. To model the spread of several related beliefs or cognitions more broadly, one can proceed in at least two ways149.
One way to model the dynamics of several beliefs is to assume that each one is affected by a summary of all the others. Many plausible summary measures have been proposed. Some are normative, such as Bayesian reasoning150,151 or logic152. Others are more descriptive, aiming to mimic actual cognitive processes. Examples are averaging strategies153, frequency-based strategies such as plurality or tallying154, birth-death dynamics like Moran processes155, or various non-compensatory strategies such as deciding based on the most important belief or consideration156.
Another way to model interacting belief contagions is to model the whole network of beliefs. This idea is not new157, but formal models of such networks that can enable analyses of interacting contagions within belief systems have started to be developed only recently. In these network models, nodes are typically beliefs, and edges represent influence between them158,159,160,161. However, nodes can also represent concepts, and edges represent beliefs about the relationship between them162. Empirical work has been done to assess the assumptions of these models and their predictions, but this is still a developing area of research163,164,165.
Modeling interactions as a network of contagions may provide a unified way to explore a wide spectrum of contagion dynamics as emergent behaviors. For example, models of interacting beliefs have been shown to break the dichotomy between simple and complex contagions and integrate both dynamics under a single framework166. Linear, simple contagion dynamics occur when the existing belief system is primed to accept a new belief, while complex contagion dynamics occur when a new belief challenges the coherence of the existing belief system, echoing the interacting spreading processes discussed in the section “The physics of interacting contagions.”
Recent work has coupled the dynamics of disease spread with a spectrum of internal states that can represent awareness or behavior and can vary according to social contacts through some social sensing process167,168,169, Incorporating the richness of internal belief interactions is a promising direction for further study of the multiple interactions that occur between and within individuals during social and biological contagions.
The ecology of interacting contagions
Across biological contagions, abstract models, stories, and beliefs, the previous sections have hinted at two important challenges in the modeling of interacting contagions. First, the number of states that agents can take grows exponentially with the number of contagions spreading in a population. Therefore, the dimensionality of our models also tends to grow exponentially with the number of contagions we wish to consider170. Second, contagions can interact on multiple scales. Interactions can occur internally, within agents, mediated by an agent’s immune system (biological contagions) or cognition (social contagions). Interactions can also occur across agents, mediated by the environment, transmission pathways, or local culture and norms.
Despite these challenges, there is an obvious need to consider interactions between large numbers of contagions, in the thousands or more, that shape our everyday lives. Therefore, we need to be able to go beyond individual-based models whose dimensionality grows exponentially and start thinking in terms of an ecosystem of contagions. This call for an ecology of contagion is a recent development29, and we here outline some promising directions.
There already exist models to infer interactions in large numbers of contagions from detailed time series of social data106 and biological data171. More rarely, we also sometimes have access to co-infection data, often static counts of individuals infected or involved in multiple contagions. From co-infection data, previous studies have built inference frameworks based on permutation tests172 or joint-species distribution models173.
Biologically, emerging technologies such as multiplex PCR panels, high-throughput sequencing (metagenomics, metabolomics, single-cell sequencing), and organoid model systems can allow us to begin to better understand the rich interrelated dynamics occurring across microbial communities at pathogen-host interfaces. Integrating these datasets and their insights into contagion models can enhance their accurate reflection of the true biology at play. In the social sciences, it is also a challenge to build broad datasets that include multiple beliefs in a time-resolved manner. Large language models are providing new methods for stance detections and belief quantification that could also resolve interrelated social dynamics174.
That being said, co-occurrence alone is not evidence of interactions175, regardless of how many covariates are included in the analyses. The ideal dataset would, of course, be a time series of individual co-infections, where these existing methods could be combined. Despite the scarcity of such data, a significant number of approaches have already been developed176. A more spatial and ecological understanding of co-infections could also help improve inference. This question has been tackled in models170,177, but separating spatial correlations from actual contagion is not a simple task178. For social contagions, surveys can help sort these different effects92. For biological contagions, the majority of studies of co-infections rely on observational data or case notes, but a minority of studies attempt to survey specific populations179.
On the modeling side, it is unclear what the ecology of contagions should look like. Can models of food web stability inspire a new generation of models of contagions? Recent models attempt to study the endogenous emergence of interactions between contagions using a game-theoretic perspective180, evolutionary models181, and co-evolutionary interactions with measures to promote or hinder these contagions182. These models encourage us to think of interactions as endogenous or emergent features of contagions and not just as fixed parameters or model mechanisms.
The field of ecology itself is challenged by modeling interacting contagions and currently faces significant analytical limitations related to the issues reviewed above. Levins’ metapopulation models, for instance, are very similar to standard epidemiological models183, representing the dynamics of individual patches that could either be empty (susceptible) or occupied (infected) by a given species, and studied for the conditions allowing species persistence at the regional level (outbreaks). Such models have been extended to multiple interacting species184, but ecologists lost track of the rapidly growing number of potential community states185. Solving this modeling challenge may require a fundamental shift in state variables; instead of representing the dynamics of a set of individuals, which could either be susceptible or infected, perhaps a solution would be to represent the dynamics of the growth rates themselves as functions of the entire community of contagious agents. Focusing directly on the contagions rather than on their hosts might help relax assumptions about the binary nature of contagious states or about the different relative timescales of biological, social, and evolutionary contagious forces. With a proper formulation of the dynamical functions, the extensive toolbox of community ecology models could then be used to investigate problems of coexistence, feasibility, stability, and higher-level interactions among contagions.
We thus join our voices to the many recommending ecological models of infectious diseases186 and contagions29. This perspective is necessary to appreciate the intricate and dynamic web of interactions between viruses, animals, parasites, humans, behaviors, and beliefs. Furthermore, under this lens, the study of emerging beliefs, stories, or epidemics can then borrow from known theories regarding invasive species in classic ecological modeling187.
We end with a question not often posed in the context of these increasingly detailed frameworks. To what end do we pursue these models? The ecological and holistic approach argued above is an increasingly common one, as different fields attempt to produce spatial and population-level understanding in times where social, ecological, and biological variables are in constant flux188. Is our goal with these frameworks to actually predict and forecast? Is that truly a litmus test of our understanding of these rich, stochastic, and inherently noisy processes? Even then, what concrete observables are we aiming to predict? New cases of a disease? New believers in a conspiracy theory? Emergence of new contagions or stories? If we adopt an ecological perspective, our goal should probably be to better understand the structure of interactions and to predict their impact on the stability and hierarchy of existing contagions. Importantly, this new objective aims for an ecological science of interacting contagions where we can study contagions as a system in and of themselves, and not just through their individual parts of pathogens and hosts.
Data availability
No datasets were generated or analyzed during the current study.
References
Harris, T. E. et al. The Theory of Branching Processes Vol. 6 (Springer Berlin, 1963).
Anderson, R. M., May, R. M. & Anderson, B. Infectious Diseases of Humans: Dynamics and Control Vol. 28 (Wiley Online Library, 1991).
Zhou, F., Xu, X., Trajcevski, G. & Zhang, K. A survey of information cascade analysis: models, predictions, and recent advances. ACM Comput. Surv. 54, 1–36 (2021).
Porter, M. A. & Gleeson, J. P. Dynamical systems on networks. Front. Appl. Dyn. Syst. Rev. Tutor. 4, 29 (2016).
Watson, H. W. & Galton, F. On the probability of the extinction of families. J. Anthropol. Inst. Gt. Br. Irel. 4, 138–144 (1875).
Morone, P. & Taylor, R. Knowledge diffusion dynamics and network properties of face-to-face interactions. J. Evolut. Econ. 14, 327–351 (2004).
Kiss, I. Z., Broom, M., Craze, P. G. & Rafols, I. Can epidemic models describe the diffusion of topics across disciplines? J. Informetr. 4, 74–82 (2010).
Labov, W. Transmission and diffusion. Language 83, 344–387 (2007).
Jin, F., Dougherty, E., Saraf, P., Cao, Y. & Ramakrishnan, N. Epidemiological modeling of news and rumors on Twitter. In Proc. 7th Workshop on Social Network Mining and Analysis 1–9 (Association for Computing Machinery, 2013).
Weng, L., Menczer, F. & Ahn, Y.-Y. Virality prediction and community structure in social networks. Sci. Rep. 3, 1–6 (2013).
Bettencourt, L. M., Cintrón-Arias, A., Kaiser, D. I. & Castillo-Chávez, C. The power of a good idea: quantitative modeling of the spread of ideas from epidemiological models. Phys. A. 364, 513–536 (2006).
Rogers, E. M. Diffusion of Innovations 5th edn (Free Press, 2003).
Dietz, K. Epidemics and rumours: a survey. J. R. Stat. Soc. Ser. A 130, 505–528 (1967).
Van Der Linden, S. Misinformation: susceptibility, spread, and interventions to immunize the public. Nat. Med. 28, 460–467 (2022).
Lovato, J. et al. Diverse misinformation: impacts of human biases on detection of deepfakes on networks. npj Complex. 1, 5 (2024).
McAdam, D. & Paulsen, R. Specifying the relationship between social ties and activism. Am. J. Sociol. 99, 640–667 (1993).
Oliver, P. E. & Myers, D. J. Diffusion models of cycles of protest as a theory of social movements. In Proc. Congress of the International Sociological Association. https://users.ssc.wisc.edu/~oliver/PROTESTS/ArticleCopies/diffusion_models.pdf (1998).
Marsden, P. Memetics and social contagion: two sides of the same coin. J. Memet. Evolut. Models Inf. Transm. 2, 171–185 (1998).
Rivers, T. M. Viruses and Koch’s postulates. J. Bacteriol. 33, 1 (1937).
Byrd, A. L. & Segre, J. A. Adapting Koch’s postulates. Science 351, 224–226 (2016).
Pirofski, L.-A. & Casadevall, A. Q&A: What is a pathogen? A question that begs the point. BMC Biol. 10, 6 (2012).
Buffie, C. G. & Pamer, E. G. Microbiota-mediated colonization resistance against intestinal pathogens. Nat. Rev. Immunol. 13, 790–801 (2013).
Williams, T. N. et al. Sickle cell trait and the risk of plasmodium falciparum malaria and other childhood diseases. J. Infect. Dis. 192, 178–186 (2005).
Brown, S. P., Cornforth, D. M. & Mideo, N. Evolution of virulence in opportunistic pathogens: generalism, plasticity, and control. Trends Microbiol. 20, 336–342 (2012).
Lehmann, S. & Ahn, Y.-Y. Complex Spreading Phenomena in Social Systems (Springer, 2018).
Kata, A. Anti-vaccine activists, web 2.0, and the postmodern paradigm—an overview of tactics and tropes used online by the anti-vaccination movement. Vaccine 30, 3778–3789 (2012).
Klein, O. & Yzerbyt, V. The Psychology of Vaccination (Taylor & Francis, 2024).
Chen, E., Lerman, K. & Ferrara, E. Tracking social media discourse about the COVID-19 pandemic: development of a public coronavirus Twitter data set. JMIR Public Health Surveill. 6, e19273 (2020).
Guilbeault, D., Becker, J. & Centola, D. Complex contagions: a decade in review. in Complex Spreading Phenomena in Social Systems 3–25 (Springer, 2018).
Hammond, R. A. & Epstein, J. M. Exploring price-independent mechanisms in the obesity epidemic. Center on Social and Economic Dynamics Working Paper. (2007).
Alshamsi, A., Pianesi, F., Lepri, B., Pentland, A. & Rahwan, I. Beyond contagion: reality mining reveals complex patterns of social influence. PLoS ONE. 10, e0135740 (2015).
Galesic, M. et al. Beyond collective intelligence: collective adaptation. J. R. Soc. Interface 20, 20220736 (2023).
Briand, S. C. et al. Infodemics: a new challenge for public health. Cell 184, 6010–6014 (2021).
Bedson, J. et al. A review and agenda for integrated disease models including social and behavioural factors. Nat. Hum. Behav. 5, 834–846 (2021).
Anderson, P. W. More is different: broken symmetry and the nature of the hierarchical structure of science. Science 177, 393–396 (1972).
Hébert-Dufresne, L., Allard, A., Garland, J., Hobson, E. A. & Zaman, L. The path of complexity. npj Complex. 1, 4 (2024).
Krapivsky, P. & Redner, S. Epidemic forecast follies. npj Complex. 1, 7 (2024).
Bernoulli, D. Chapelle, D. Essai d’une nouvelle analyse de la mortalité causée par la petite vérole, et des avantages de l’inoculation pour la prévenir. Mémoires de Mathématique et de Physique, tirés des registres de l'Académie Royale des Sciences de l'année 1760. 1–45 https://inria.hal.science/hal-04100467 (2023).
Kermack, W. O. & McKendrick, A. G. A contribution to the mathematical theory of epidemics. Proc. R. Soc. Lond. A 115, 700–721 (1927).
May, R. M. & Nowak, M. A. Coinfection and the evolution of parasite virulence. Proc. R. Soc. Lond. Ser. B 261, 209–215 (1995).
Andreasen, V., Lin, J. & Levin, S. A. The dynamics of cocirculating influenza strains conferring partial cross-immunity. J. Math. Biol. 35, 825–842 (1997).
Epstein, J. M., Parker, J., Cummings, D. & Hammond, R. A. Coupled contagion dynamics of fear and disease: mathematical and computational explorations. PloS One. 3, e3955 (2008).
Newman, M. E. & Ferrario, C. R. Interacting epidemics and coinfection on contact networks. PLoS ONE. 8, e71321 (2013).
Gog, J. R. & Grenfell, B. T. Dynamics and selection of many-strain pathogens. Proc. Natl. Acad. Sci. USA 99, 17209–17214 (2002).
Karrer, B. & Newman, M. E. Competing epidemics on complex networks. Phys. Rev. E. 84, 036106 (2011).
Bansal, S. & Meyers, L. A. The impact of past epidemics on future disease dynamics. J. Theor. Biol. 309, 176–184 (2012).
Miller, J. C. Cocirculation of infectious diseases on networks. Phys. Rev. E 87, 060801 (2013).
Poletto, C., Meloni, S., Colizza, V., Moreno, Y. & Vespignani, A. Host mobility drives pathogen competition in spatially structured populations. PLoS Comput. Biol. 9, e1003169 (2013).
Gleeson, J. P., Ward, J. A., O’Sullivan, K. P. & Lee, W. T. Competition-induced criticality in a model of meme popularity. Phys. Rev. Lett. 112, 048701 (2014).
Williams, B. J., Ogbunugafor, C. B., Althouse, B. M. & Hébert-Dufresne, L. Immunity-induced criticality of the genotype network of influenza A (H3N2) hemagglutinin. PNAS Nexus 1, pgac143 (2022).
Newman, M. E. Threshold effects for two pathogens spreading on a network. Phys. Rev. Lett. 95, 108701 (2005).
Funk, S., Gilad, E., Watkins, C. & Jansen, V. A. The spread of awareness and its impact on epidemic outbreaks. Proc. Natl. Acad. Sci. USA 106, 6872–6877 (2009).
Funk, S. & Jansen, V. A. Interacting epidemics on overlay networks. Phys. Rev. E 81, 036118 (2010).
Marceau, V., Noël, P.-A., Hébert-Dufresne, L., Allard, A. & Dubé, L. J. Modeling the dynamical interaction between epidemics on overlay networks. Phys. Rev. E 84, 026105 (2011).
Wang, W., Liu, Q.-H., Liang, J., Hu, Y. & Zhou, T. Coevolution spreading in complex networks. Phys. Rep. 820, 1–51 (2019).
Vasco, D. A., Wearing, H. J. & Rohani, P. Tracking the dynamics of pathogen interactions: modeling ecological and immune-mediated processes in a two-pathogen single-host system. J. Theor. Biol. 245, 9–25 (2007).
Abu-Raddad, L. J., Van der Ventel, B. & Ferguson, N. M. Interactions of multiple strain pathogen diseases in the presence of coinfection, cross immunity, and arbitrary strain diversity. Phys. Rev. Lett. 100, 168102 (2008).
Hébert-Dufresne, L., Patterson-Lomba, O., Goerg, G. M. & Althouse, B. M. Pathogen mutation modeled by competition between site and bond percolation. Phys. Rev. Lett. 110, 108103 (2013).
Campbell, E. & Salathé, M. Complex social contagion makes networks more vulnerable to disease outbreaks. Sci. Rep. 3, 1905 (2013).
De Domenico, M., Granell, C., Porter, M. A. & Arenas, A. The physics of spreading processes in multilayer networks. Nat. Phys. 12, 901 (2016).
Fu, F., Christakis, N. A. & Fowler, J. H. Dueling biological and social contagions. Sci. Rep. 7, 43634 (2017).
Liu, Q.-H., Zhong, L.-F., Wang, W., Zhou, T. & Eugene Stanley, H. Interactive social contagions and co-infections on complex networks. Chaos 28, 013120 (2018).
Chen, L., Ghanbarnejad, F., Cai, W. & Grassberger, P. Outbreaks of coinfections: the critical role of cooperativity. Europhys. Lett. 104, 50001 (2013).
Hébert-Dufresne, L. & Althouse, B. M. Complex dynamics of synergistic coinfections on realistically clustered networks. Proc. Natl. Acad. Sci. USA 112, 10551–10556 (2015).
Cai, W., Chen, L., Ghanbarnejad, F. & Grassberger, P. Avalanche outbreaks emerging in cooperative contagions. Nat. Phys. 11, 936 (2015).
Chen, L., Ghanbarnejad, F. & Brockmann, D. Fundamental properties of cooperative contagion processes. New J. Phys. 19, 103041 (2017).
Soriano-Paños, D., Ghanbarnejad, F., Meloni, S. & Gómez-Gardeñes, J. Markovian approach to tackle the interaction of simultaneous diseases. Phys. Rev. E 100, 062308 (2019).
Grassberger, P., Chen, L., Ghanbarnejad, F. & Cai, W. Phase transitions in cooperative coinfections: simulation results for networks and lattices. Phys. Rev. E 93, 042316 (2016).
Hébert-Dufresne, L., Scarpino, S. V. & Young, J.-G. Macroscopic patterns of interacting contagions are indistinguishable from social reinforcement. Nat. Phys. 16, 426–431 (2020).
Lamata-Otín, S., Gómez-Gardeñes, J. & Soriano-Paños, D. Pathways to discontinuous transitions in interacting contagion dynamics. J. Phys. Complex. 5, 015015 (2024).
Hébert-Dufresne, L., Mistry, D. & Althouse, B. M. Spread of infectious disease and social awareness as parasitic contagions on clustered networks. Phys. Rev. Res. 2, 033306 (2020).
Centola, D. The spread of behavior in an online social network experiment. Science 329, 1194–1197 (2010).
de Arruda, G. F., Rodrigues, F. A. & Moreno, Y. Fundamentals of spreading processes in single and multilayer complex networks. Phys. Rep. 756, 1–59 (2018).
Wang, W. et al. Epidemic spreading on higher-order networks. Phys. Rep. 1056, 1–70 (2024).
Bródka, P., Musial, K. & Jankowski, J. Interacting spreading processes in multilayer networks: a systematic review. IEEE Access 8, 10316–10341 (2020).
Huang, H., Chen, Y. & Ma, Y. Modeling the competitive diffusions of rumor and knowledge and the impacts on epidemic spreading. Appl. Math. Comput. 388, 125536 (2021).
Li, W., Xue, X., Pan, L., Lin, T. & Wang, W. Competing spreading dynamics in simplicial complex. Appl. Math. Comput. 412, 126595 (2022).
Nie, Y., Li, W., Pan, L., Lin, T. & Wang, W. Markovian approach to tackle competing pathogens in simplicial complex. Appl. Math. Comput. 417, 126773 (2022).
Li, W. et al. Coevolution of epidemic and infodemic on higher-order networks. Chaos Solit. Fractals 168, 113102 (2023).
Fan, J., Yin, Q., Xia, C. & Perc, M. Epidemics on multilayer simplicial complexes. Proc. R. Soc. A 478, 20220059 (2022).
You, X., Zhang, M., Ma, Y., Tan, J. & Liu, Z. Impact of higher-order interactions and individual emotional heterogeneity on information-disease coupled dynamics in multiplex networks. Chaos Solit. Fractals 177, 114186 (2023).
Liu, L., Feng, M., Xia, C., Zhao, D. & Perc, M. Epidemic trajectories and awareness diffusion among unequals in simplicial complexes. Chaos, Solit. Fractals 173, 113657 (2023).
Hong, Z., Zhou, H., Wang, Z., Yin, Q. & Liu, J. Coupled propagation dynamics of information and infectious disease on two-layer complex networks with simplices. Mathematics 11, 4904 (2023).
Zhu, Y., Li, C. & Li, X. Epidemic spreading on coupling network with higher-order information layer. New J. Phys. 25, 113043 (2023).
Min, Y. et al. Diversity of multilayer networks and its impact on collaborating epidemics. Phys. Rev. E 90, 062803 (2014).
Nguyen, M. M. Upper bounds on overshoot in sir models with nonlinear incidence. npj Complex. 1, 11 (2024).
Katz, E. & Lazarsfeld, P. F. Personal Influence (The Free Press, 1955).
Weimann, G.The Influentials: People Who Influence People (State University of New York Press, 1994).
Gladwell, M. The Tipping Point (Little, Brown and Company, 2000).
Watts, D. J. & Dodds, P. S. Influentials, networks, and public opinion formation. J. Consum. Res. 34, 441–458 (2007).
Goffman, W. & Newill, V. A. Generalization of epidemic theory: an application to the transmission of ideas. Nature 204, 225–228 (1964).
Cavalli-Sforza, L. L., Feldman, M. W., Chen, K.-H. & Dornbusch, S. M. Theory and observation in cultural transmission. Science 218, 19–27 (1982).
Ananthasubramaniam, A., Jurgens, D. & Romero, D. M. Networks and identity drive the spatial diffusion of linguistic innovation in urban and rural areas. npj Complex. 1, 14 (2024).
Lerman, K., Feldman, D., He, Z. & Rao, A. Affective polarization and dynamics of information spread in online networks. npj Complex. 1, 8 (2024).
Schelling, T. C. Dynamic models of segregation. J. Math. Sociol. 1, 143–186 (1971).
Granovetter, M. Threshold models of collective behavior. Am. J. Sociol. 83, 1420–1443 (1978).
Granovetter, M. S. & Soong, R. Threshold models of diffusion and collective behavior. J. Math. Sociol. 9, 165–179 (1983).
Granovetter, M. S. & Soong, R. Threshold models of interpersonal effects in consumer demand. J. Econ. Behav. Organ. 7, 83–99 (1986).
Granovetter, M. & Soong, R. Threshold models of diversity: Chinese restaurants, residential segregation, and the spiral of silence. Sociol. Methodol. 18, 69–104 (1988).
Dodds, P. S. & Watts, D. J. Universal behavior in a generalized model of contagion. Phys. Rev. Lett. 92, 218701 (2004).
Dodds, P. S. & Watts, D. J. A generalized model of social and biological contagion. J. Theor. Biol. 232, 587–604 (2005).
Guille, A., Hacid, H., Favre, C. & Zighed, D. A. Information diffusion in online social networks: a survey. ACM Sigmod Rec. 42, 17–28 (2013).
Lerman, K. & Ghosh, R. Information contagion: an empirical study of the spread of news on Digg and Twitter social networks. In Proc. International AAAI Conference on Web and Social Media 4, 90–97 (Association for the Advancement of Artificial Intelligence, 2010).
Pathak, N., Banerjee, A. & Srivastava, J. A generalized linear threshold model for multiple cascades. In Proc. 2010 IEEE International Conference on Data Mining 965–970 (IEEE, 2010).
Lu, W., Chen, W. & Lakshmanan, L. V. From competition to complementarity: comparative influence diffusion and maximization. Proc. VLDB Endow. 9, 60–71 (2015).
Myers, S. A. & Leskovec, J. Clash of the contagions: cooperation and competition in information diffusion. In Proc. 2012 IEEE 12th International Conference on Data Mining 539–548 (IEEE, 2012).
Zarezade, A., Khodadadi, A., Farajtabar, M., Rabiee, H. & Zha, H. Correlated cascades: compete or cooperate. In Proc. AAAI Conference on Artificial Intelligence 31 (Association for the Advancement of Artificial Intelligence, 2017).
Hawkes, A. G. Spectra of some self-exciting and mutually exciting point processes. Biometrika 58, 83–90 (1971).
Weng, L., Flammini, A., Vespignani, A. & Menczer, F. Competition among memes in a world with limited attention. Sci. Rep. 2, 335 (2012).
Althouse, B. M. et al. Enhancing disease surveillance with novel data streams: challenges and opportunities. EPJ Data Sci. 4, 1–8 (2015).
Leskovec, J., Backstrom, L. & Kleinberg, J. Meme-tracking and the dynamics of the news cycle. In Proc. 15th ACM SIGKDD International Conference on Knowledge Discovery and Data Mining 497–506 (Association for Computing Machinery, 2009).
Gruhl, D., Guha, R., Liben-Nowell, D. & Tomkins, A. Information diffusion through blogspace. In Proc. 13th International Conference on World Wide Web 491–501 (Association for Computing Machinery, 2004).
Adar, E. & Adamic, L. A. Tracking information epidemics in blogspace. In Proc. 2005 IEEE/WIC/ACM International Conference on Web Intelligence (WI’05) 207–214 (IEEE, 2005).
Kleinberg, J. Bursty and hierarchical structure in streams. In Proc. Eighth ACM SIGKDD International Conference on Knowledge Discovery and Data Mining 91–101 (Association for Computing Machinery, 2002).
Mandl, K. D. et al. Implementing syndromic surveillance: a practical guide informed by the early experience. J. Am. Med. Inform. Assoc. 11, 141–150 (2004).
Dubey, A., Moro, E., Cebrian, M. & Rahwan, I. Memesequencer: Sparse matching for embedding image macros. In Proc. 2018 World Wide Web Conference 1225–1235 (Association for Computing Machinery, 2018).
Afridi, T. H., Alam, A., Khan, M. N., Khan, J. & Lee, Y.-K. A multimodal memes classification: a survey and open research issues. In Proc. Innovations in Smart Cities Applications Volume 4: The 5th International Conference on Smart City Applications 1451–1466 (Springer, 2021).
Beskow, D. M., Kumar, S. & Carley, K. M. The evolution of political memes: detecting and characterizing internet memes with multi-modal deep learning. Inf. Process. Manag. 57, 102170 (2020).
Ling, C. et al. Dissecting the meme magic: understanding indicators of virality in image memes. Proc. ACM Hum. Comput. Interact. 5, 1–24 (2021).
Sartre, J.-P. La Nausée (Éditions Gallimard, 1938).
Ranke, K. Kategorienprobleme der Volksprosa. Fabula 9, 4 (1967).
Fisher, W. R. Narration as a human communication paradigm: the case of public moral argument. Commun. Monogr. 51, 1–22 (1984).
Gould, S. J. So near and yet so far. N. Y. Rev. Books 41, 26 (1994).
Ferrand, N. & Weil, M. Homo narrativus: Dix ans de recherche sur la topique romanesque (Université Paul-Valéry de Montpellier, 2001).
Bruner, J. S. Making Stories: Law, Literature, Life (Harvard University Press, 2003).
Boyd, B. On the Origin of Stories: Evolution, Cognition, and Fiction (Belknap Press, 2010).
Moretti, F. Distant Reading (Verso, 2013).
Dodds, P. S. Homo Narrativus and the Trouble with Fame. Nautilus Magazine (2013).
Gottschall, J. The Storytelling Animal: How Stories Make Us Human (Mariner Books, 2013).
Maruna, S. Foreword: narrative criminology as the new mainstream. in Narrative Criminology vii–x (New York University Press, 2015).
Shiller, R. J. Narrative economics. Am. Econ. Rev. 107, 967–1004 (2017).
Puchner, M. The Written World: How Literature Shaped Civilization (Random, 2017).
Garland, J., Ghazi-Zahedi, K., Young, J.-G., Hébert-Dufresne, L. & Galesic, M. Impact and dynamics of hate and counter speech online. EPJ Data Sci. 11, 3 (2022).
Nettasinghe, B., Percus, A. G. & Lerman, K. How out-group animosity can shape partisan divisions: a model of affective polarization. PNAS Nexus 4, pgaf082 (2025).
Nettasinghe, B., Rao, A., Jiang, B., Percus, A. & Lerman, K. In-group love, out-group hate: a framework to measure affective polarization via contentious online discussions. In Proc. Web Conference (Association for Computing Machinery, 2025).
Ajzen, I. The theory of planned behavior. Organ. Behav. Hum. Decis. Process. 50, 179–211 (1991).
Monroe, B. M. & Read, S. J. A general connectionist model of attitude structure and change: the ACS (attitudes as constraint satisfaction) model. Psychol. Rev. 115, 733 (2008).
Olsson, H. & Galesic, M. Analogies for modeling belief dynamics. Trends Cogn. Sci. 28, 907–923 (2024).
Schmelz, K. & Bowles, S. Overcoming COVID-19 vaccination resistance when alternative policies affect the dynamics of conformism, social norms, and crowding out. Proc. Natl. Acad. Sci. USA 118, e2104912118 (2021).
Pew Research Center. Public opinion on abortion. https://www.pewresearch.org/religion/fact-sheet/publicopinion-on-abortion/(2024).
Festinger, L. A theory of social comparison processes. Hum. Relat. 7, 117–140 (1954).
Brenan, M. Same-sex relations, marriage still supported by most in U.S. (Gallup, 2024).
Ebner, J. From margins to mainstream: How extremism has conquered the political middle. (International Centre for Counter-Terrorism, 2023).
Pertwee, E., Simas, C. & Larson, H. J. An epidemic of uncertainty: rumors, conspiracy theories and vaccine hesitancy. Nat. Med. 28, 456–459 (2022).
Solomon, S., Greenberg, J. & Pyszczynski, T. Pride and prejudice: fear of death and social behavior. Curr. Dir. Psychol. Sci. 9, 200–204 (2000).
Castellano, C., Fortunato, S. & Loreto, V. Statistical physics of social dynamics. Rev. Mod. Phys. 81, 591–646 (2009).
Flache, A. et al. Models of social influence: towards the next frontiers. J. Artif. Soc. Soc. Simul. 20, 2 (2017).
Redner, S. Reality-inspired voter models: a mini-review. C. R. Phys. 20, 275–292 (2019).
Galesic, M., Olsson, H., Dalege, J., Van Der Does, T. & Stein, D. L. Integrating social and cognitive aspects of belief dynamics: towards a unifying framework. J. R. Soc. Interface 18, 20200857 (2021).
Chater, N., Tenenbaum, J. B. & Yuille, A. Probabilistic models of cognition: conceptual foundations. Trends Cogn. Sci. 10, 287–291 (2006).
Bikhchandani, S., Hirshleifer, D. & Welch, I. A theory of fads, fashion, custom, and culturalchange as informational cascades. J. Pol. Econ 100, 992–1026 (1992).
Van Benthem, J. Logical Dynamics of Information and Interaction (Cambridge University Press, 2011).
Friedkin, N. E. & Johnsen, E. C. Social influence and opinions. J. Math. Sociol. 15, 193–206 (1990).
Einhorn, H. J. & Hogarth, R. M. Unit weighting schemes for decision making. Organ. Behav. Hum. Perform. 13, 171–192 (1975).
Fic, M. & Gokhale, C. S. Catalysing cooperation: the power of collective beliefs in structured populations. npj Complex. 1, 6 (2024).
Gigerenzer, G. & Goldstein, D. G. Reasoning the fast and frugal way: models of bounded rationality. Psychol. Rev. 103, 650 (1996).
Converse, P. E. The nature of belief systems in mass publics. in Ideology and Its Disconte (ed. Apter, D. E.) 206–261 (The Free Press, 1964).
Dalege, J. et al. Toward a formalized account of attitudes: the Causal Attitude Network (CAN) model. Psychol. Rev. 123, 2 (2016).
Boutyline, A. & Vaisey, S. Belief network analysis: a relational approach to understanding the structure of attitudes. Am. J. Sociol. 122, 1371–1447 (2017).
Brandt, M. J., Sibley, C. G. & Osborne, D. What is central to political belief system networks? Personal. Soc. Psychol. Bull. 45, 1352–1364 (2019).
Vlasceanu, M., Dyckovsky, A. M. & Coman, A. A network approach to investigate the dynamics of individual and collective beliefs: advances and applications of the bending model. Perspect. Psychol. Sci. 19, 444–453 (2024).
Rodriguez, N., Bollen, J. & Ahn, Y.-Y. Collective dynamics of belief evolution under cognitive coherence and social conformity. PLoS ONE 11, e0165910 (2016).
Brandt, M. J. & Vallabha, S. Inter-attitude centrality does not appear to reduce persuasion for political attitudes. Eur. J. Soc. Psychol. 53, 1342–1358 (2023).
Turner-Zwinkels, F. M. & Brandt, M. J. Belief system networks can be used to predict where to expect dynamic constraint. J. Exp. Soc. Psychol. 100, 104279 (2022).
Dalege, J., Galesic, M. & Olsson, H. Networks of beliefs: an integrative theory of individual-and social-level belief dynamics. Psychol. Rev. 132, 253–290 (2024).
Aiyappa, R., Flammini, A. & Ahn, Y.-Y. Emergence of simple and complex contagion dynamics from weighted belief networks. Sci. Adv. 10, eadh4439 (2024).
Maghool, S., Maleki-Jirsaraei, N. & Cremonini, M. The coevolution of contagion and behavior with increasing and decreasing awareness. PLoS ONE 14, e0225447 (2019).
O’Gara, D., Kasman, M., Hébert-Dufresne, L. & Hammond, R. A. Adaptive behaviour during epidemics: a social risk appraisal approach to modelling dynamics. J. R. Soc. Interface 22, 20240363 (2025).
Khanjanianpak, M., Azimi-Tafreshi, N., Arenas, A. & Gómez-Gardeñes, J. Emergence of protective behaviour under different risk perceptions to disease spreading. Phil. Trans. R. Soc. A 380, 20200412 (2022).
Stanoev, A., Trpevski, D. & Kocarev, L. Modeling the spread of multiple concurrent contagions on networks. PLoS ONE 9, e95669 (2014).
Shrestha, S., King, A. A. & Rohani, P. Statistical inference for multi-pathogen systems. PLoS Comput. Biol. 7, e1002135 (2011).
Colgate, E. R. et al. Network analysis of patterns and relevance of enteric pathogen co-infections among infants in a diarrhea-endemic setting. PLOS Comput. Biol. 19, e1011624 (2023).
Fountain-Jones, N. M. et al. Endemic infection can shape exposure to novel pathogens: pathogen co-occurrence networks in the Serengeti lions. Ecol. Lett. 22, 904–913 (2019).
Gül, I., Lebret, R. & Aberer, K. Stance detection on social media with fine-tuned large language models. Preprint at arXiv, https://doi.org/10.48550/arXiv.2404.12171 (2024).
Blanchet, F. G., Cazelles, K. & Gravel, D. Co-occurrence is not evidence of ecological interactions. Ecol. Lett. 23, 1050–1063 (2020).
Powell-Romero, F., Wells, K. & Clark, N. J. A systematic review and guide for using multi-response statistical models in co-infection research. R. Soc. Open Sci. 11, 231589 (2024).
Chen, L. Persistent spatial patterns of interacting contagions. Phys. Rev. E 99, 022308 (2019).
Waring, T. M. et al. Operationalizing cultural adaptation to climate change: contemporary examples from United States agriculture. Proc. R. Soc. B 378, 20220397 (2023).
Griffiths, E. C., Pedersen, A. B., Fenton, A. & Petchey, O. L. The nature and consequences of coinfection in humans. J. Infect. 63, 200–206 (2011).
Su, Y., Zhang, X., Liu, L., Song, S. & Fang, B. Understanding information interactions in diffusion: an evolutionary game-theoretic perspective. Front. Comput. Sci. 10, 518–531 (2016).
Ghanbarnejad, F., Seegers, K., Cardillo, A. & Hövel, P. Emergence of synergistic and competitive pathogens in a coevolutionary spreading model. Phys. Rev. E 105, 034308 (2022).
Khazaee, A. & Ghanbarnejad, F. Effects of measures on phase transitions in two cooperative susceptible-infectious-recovered dynamics. Phys. Rev. E 105, 034311 (2022).
Levins, R. Some demographic and genetic consequences of environmental heterogeneity for biological control. Bull. Entomol. Soc. Am. 15, 237–240 (1969).
Hanski, I. Coexistence of competitors in patchy environment. Ecology 64, 493–500 (1983).
Cazelles, K., Mouquet, N., Mouillot, D. & Gravel, D. On the integration of biotic interaction and environmental constraints at the biogeographical scale. Ecography 39, 921–931 (2016).
Hassell, J. M. et al. Towards an ecosystem model of infectious disease. Nat. Ecol. Evol. 5, 907–918 (2021).
Vilà, M. et al. Viewing emerging human infectious epidemics through the lens of invasion biology. BioScience 71, 722–740 (2021).
Lasky, J. R., Hooten, M. B. & Adler, P. B. What processes must we understand to forecast regional-scale population dynamics? Proc. R. Soc. B 287, 20202219 (2020).
Fleming, D. T. & Wasserheit, J. N. From epidemiological synergy to public health policy and practice: the contribution of other sexually transmitted diseases to sexual transmission of HIV infection. Sex. Transm. Infect. 75, 3–17 (1999).
Mehta, R. S. & Rosenberg, N. A. Modelling anti-vaccine sentiment as a cultural pathogen. Evolut. Hum. Sci. 2, e21 (2020).
Lian, S., Liu, J., Wu, Y., Xia, P. & Zhu, G. Bacterial and viral co-infection in the intestine: competition scenario and their effect on host immunity. Int. J. Mol. Sci. 23, 2311 (2022).
Sznajd-Weron, K. & Sznajd, J. Who is left, who is right? Phys. A 351, 593–604 (2005).
Seekatz, A. M., Safdar, N. & Khanna, S. The role of the gut microbiome in colonization resistance and recurrent Clostridioides difficile infection. Ther. Adv. Gastroenterol. 15, 17562848221134396 (2022).
Tambuscio, M., Ruffo, G., Flammini, A. & Menczer, F. Fact-checking effect on viral hoaxes: A model of misinformation spread in social networks. In Proc. 24th International Conference on World Wide Web 977–982 (Association for Computing Machinery, 2015).
Bauch, C. T. & Galvani, A. P. Social factors in epidemiology. Science 342, 47–49 (2013).
Magarey, R. D. & Trexler, C. M. Information: a missing component in understanding and mitigating social epidemics. Humanit. Soc. Sci. Commun. 7, 1–11 (2020).
Negro, F. & Lok, A. S. Hepatitis D: a review. JAMA. 330, 2376–2387 (2023).
Rowe, H. M. et al. Direct interactions with influenza promote bacterial adherence during respiratory infections. Nat. Microbiol. 4, 1328–1336 (2019).
Martinson, M. L. & Lapham, J. Prevalence of immunosuppression among US adults. JAMA. 331, 880–882 (2024).
Wallace, B. I. et al. Prevalence of immunosuppressive drug use among commercially insured US adults, 2018-2019. JAMA Netw. Open 4, e214920–e214920 (2021).
Morales, F., Montserrat-De la Paz, S., Leon, M. J. & Rivero-Pino, F. Effects of malnutrition on the immune system and infection and the role of nutritional strategies regarding improvements in children’s health status: a literature review. Nutrients 16, 1 (2023).
Bell, L. C. & Noursadeghi, M. Pathogenesis of HIV-1 and Mycobacterium tuberculosis co-infection. Nat. Rev. Microbiol. 16, 80–90 (2018).
Do Nascimento, I. J. B. et al. Infodemics and health misinformation: a systematic review of reviews. Bull. World Health Organ. 100, 544 (2022).
Jin, S. L. et al. Social histories of public health misinformation and infodemics: case studies of four pandemics. Lancet Infect. Dis. 24, e638–e646 (2024).
Dyer, O. Philippines measles outbreak is deadliest yet as vaccine scepticism spurs disease comeback. BMJ. 364, l739 (2019).
Normile, D. Safety concerns derail dengue vaccination program. Science 358, 1514–1515 (2017).
BBC News. How a wrong injection helped cause Samoa’s measles epidemic. BBC https://www.bbc.com/news/world-asia-50625680. (2019).
Champredon, D., Shoukat, A., Singer, B. H., Galvani, A. P. & Moghadas, S. M. Curbing the 2019 Samoa measles outbreak. Lancet Infect. Dis. 20, 287–288 (2020).
Grant-Alfieri, A., Schaechter, J. & Lipshultz, S. E. Ingesting and aspirating dry cinnamon by children and adolescents: the “cinnamon challenge”. Pediatrics 131, 833–835 (2013).
Breakey, W., Crowley, T. P. & Alrawi, M. Salt and ice, a challenge not to be taken lightly. J. Burn Care Res. 36, e230–e230 (2015).
Drug Safety Communication. FDA warns about serious problems with high doses of the allergy medicine diphenhydramine (Benadryl). Food and Drug Administration. https://www.fda.gov/drugs/drugsafety-and-availability/fda-warns-about-serious-problems-high-doses-allergy-medicine-diphenhydramine-benadryl (2020).
Merchant, R. M., South, E. C. & Lurie, N. Public health messaging in an era of social media. JAMA. 325, 223–224 (2021).
Hyland-Wood, B., Gardner, J., Leask, J. & Ecker, U. K. Toward effective government communication strategies in the era of COVID-19. Humanit. Soc. Sci. Commun. 8, 30 (2021).
Labov, W. & Waletzky, J. Narrative analysis: oral versions of personal experience. JNLH 7, 3–38 (1997).
Barkun, M. A Culture of Conspiracy: Apocalyptic Visions in Contemporary America Vol. 15 (University of California Press, 2013).
Holur, P., Wang, T., Shahsavari, S., Tangherlini, T. & Roychowdhury, V. Which side are you on? Insider-Outsider classification in conspiracy-theoretic social media. In Proc. 60th Annual Meeting of the Association for Computational Linguistics Vol. 1, 4975–4987 (Association for Computational Linguistics, 2022).
Holur, P., Chong, D., Tangherlini, T. & Roychowdhury, V. My side, your side and the evidence: discovering aligned actor groups and the narratives they weave. In Proc. 61st Annual Meeting of the Association for Computational Linguistics 8938–8952 (Association for Computational Linguistics, 2023).
Tangherlini, T. R. et al. "Mommy blogs” and the vaccination exemption narrative: results from a machine-learning approach for story aggregation on parenting social media sites. JMIR Public Health Surveill. 2, e6586 (2016).
Greimas, A. J. Éléments pour une théorie de l’interprétation du récit mythique. Communications 8, 28–59 (1966).
Lehnert, W. G. Plot units and narrative summarization. Cogn. Sci. 5, 293–331 (1981).
Tangherlini, T. R., Shahsavari, S., Shahbazi, B., Ebrahimzadeh, E. & Roychowdhury, V. An automated pipeline for the discovery of conspiracy and conspiracy theory narrative frameworks: Bridgegate, Pizzagate and storytelling on the web. PLoS ONE 15, e0233879 (2020).
Boole, G. An investigation of the Laws of Thought: On Which are Founded the Mathematical Theories of Logic and Probabilities Vol. 2 (Walton and Maberly, 1854).
Eskeröd, A. Ĺrets äring: etnologiska studier i skördens och julens tro och sed. PhD thesis, Nordisk museet (1947).
Anderson, W. Kaiser und Abt: Die Geschichte eines Schwanks Vol. 42 (Suomalainen Tiedeakatemia, Academia Scientiarum Fennica, 1923).
Acknowledgements
This work is based on the workshop “Dynamics of interacting contagions” held at the Santa Fe Institute. The authors acknowledge financial support from the Santa Fe Institute, the Vermont Complex Systems Institute (award #2242829 from the National Science Foundation), and from the Translational Global Infectious Diseases Research Center of Biomedical Research Excellence (award P20GM125498 from the National Institute of General Medical Sciences).
Author information
Authors and Affiliations
Contributions
L.H.-D. oversaw the writing. L.H.-D., P.S.D., and M.G. wrote the initial draft. All authors reviewed and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
L.H.-D. is the Editor-in-Chief of the journal. A.A. is an Associate Editor of the journal. M.G. is a member of the journal’s Editorial Board, and the authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Hébert-Dufresne, L., Ahn, YY., Allard, A. et al. One pathogen does not an epidemic make: a review of interacting contagions, diseases, beliefs, and stories. npj Complex 2, 26 (2025). https://doi.org/10.1038/s44260-025-00050-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s44260-025-00050-2
This article is cited by
-
Homophily within and across groups
Nature Communications (2025)
