
Abstract
Indoor vertical farming emerges as a sustainable paradigm, enabling crop cultivation in vertically layered ecosystems under precisely controlled environmental parameters. This innovative approach not only bolsters climate resilience but also minimizes the strain on precious arable land and water resources, aligning with sustainable principles. While posited as a transformative solution to global food security challenges and the limitations of traditional agriculture, a pivotal facet within indoor farming necessitates sustainable optimization: the crops cultivated within. Conventional commercial crop varieties utilized in indoor vertical farming lack tailored adaptations to controlled environments, underscoring the imperative to cultivate crops endowed with distinct indoor-farming traits. These include better performance under energy-efficient artificial lighting, high growth rate, and unique flavours. In this review, we highlight recent technological advancements in crop breeding and elaborate these coveted indoor-farming traits. In addition, we illuminate potential genetic workflows essential for breeders to strategically refine crops as a foundation for the long-term sustainability of indoor farming.
Similar content being viewed by others
Introduction
With the world’s population projected to grow from 8 billion in 2022 to 9.6 billion by 2050, there is an increasing need for food security to ensure sufficient and nutritious food at all times1. This global challenge is exacerbated by the rising frequency of weather extremes and the limitations of expanding arable land. In response, the agricultural sector is pivoting towards sustainable practices that optimize operating conditions, including light, temperature, humidity, and nutrients, through controlled environments. In addition, the accelerating urbanization trend necessitates a shift towards producing food closer to urban consumers, a concept that aligns with sustainability goals and minimizes transportation emissions. Thus, indoor vertical farming that grows crop plants in a vertically layered system in urban settings has emerged to be one of the promising options of modern agriculture as recent technological advances have made it possible to grow crops with precisely controlled growing parameters. This will allow crops to be produced all year round as they are no longer reliant on weather patterns and disrupted by weather abnormalities. This model of controlled environment agriculture also provides reproducibility in yield, rapid turnover, scalability in having more farms in land-scarce urban precincts, and shorter food supply chain with lower carbon footprint.
While new engineering technologies for lighting, climate control and automation have enabled the production of different types of crops in an indoor environment2, there remains a notable lag in the innovation of the plant itself, necessitating the generation of crops and varieties tailored to thrive in controlled environments. The genetic background and the agronomic characteristics of a crop largely influence the space requirements, inputs and processes needed for optimal production. Currently, as indoor vertical farming is at its nascent stage, commercial varieties are primarily designed for outdoor environments and lack traits optimized for controlled indoor settings3. To address this challenge, there is a compelling need to develop new crop varieties tailored specifically for indoor vertical farming. These varieties should exhibit traits, such as improved performance under artificial lighting, optimized plant architecture, and enhanced nutritional profiles.
Genetic breeding via choosing parents with specific characteristics of interest to generate offspring with desirable traits has been continuously practised since domestication4. Over thousands of years, various approaches have been developed to permit farmers to improve the crop productivity greatly with high yielding, increased resistance to diseases and pests, higher nutritional contents, and better adaptation to different ecological conditions5. In today’s technological era, we are better equipped with advanced breeding tools to combine genomics and phenomics with computational biology to generate novel crop varieties in a shorter period. In particular, gene editing technology is becoming a driving force in quick production of desired plants for controlled environment. These advances, together with an increasing diversity in consumer preferences, will promote genetic breeding of crops with niche characteristics under controlled environments as an integral part of the transition towards sustainable agriculture.
The long-term success of indoor vertical farming hinges on genetic improvements that align with sustainability goals. In this review, we present an overview of recent technological advancements in crop breeding and discuss the essential traits and potential breeding workflows required to develop crops suitable for sustainable indoor vertical farming practices.
Indoor vertical farming is shaping the future of resilient cities
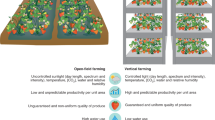
Urban farming refers to the practice of cultivating, processing and distributing food in or around urban areas6. It has become an increasingly popular trend in cities around the world7. With projections indicating that nearly 70% of the global population will be urban dwellers by 2050, the need for food security in these densely populated regions is paramount. However, this urban shift also presents a promising opportunity to integrate urban agriculture into the global economy and supply chain1. There are various types of urban farming, such as backyard gardens, open rooftop farms, greenhouses, shipping containers and indoor vertical farms. Indoor vertical farming, in particular, stands out as a high-tech, smart farming approach that integrates data science, engineering, and plant sciences into precision agriculture2,8,9. This innovative farm strategy usually deploys artificial light sources, such as light-emitting diodes (LEDs), in vertically stacked layers within a controlled environment10,11.
The potential benefits of indoor vertical farming extend across environmental, social, and economic dimensions, yielding positive and sustainable outcomes for local communities (Fig. 1). From the environmental perspective, this approach minimizes the ecological footprint of food production by conserving land, water, fertilizers, and pesticides while reducing the need for long-distance food transportation. This will help conserve natural resources and improve air quality in cities by reducing greenhouse gas emission12. From the social perspective, the setting up of urban farms provides employment opportunities in different sectors of the industry. In addition to roles directly involved in farming such as agronomists, farm managers, and crop technicians, there are opportunities in infrastructure development, including construction workers, architects, and engineers tasked with designing and building the farm structures. The installation and maintenance of specialized equipment like growing systems, LED lighting, and automated irrigation systems also require skilled technicians and electricians. The stable supply of food also helps to improve the nutrition for people and strengthen food security. From the economic perspective, urban farming generates a higher crop yield per unit area via using vertical space (intensive farming) in a year-round operation mode13. The scalability and proximity to the consumer market allows for efficient matching between supply and demand, thus reducing the risk of over-production and minimizing wastage.

The establishment of an indoor vertical farm in an urbanized area delivers economic, social, and environmental advantages to the local community, fostering sustainability and enhancing the well-being of residents.
Notably, indoor vertical farming acts as a catalyst for agricultural innovation, fostering the development and adoption of new technologies and services. It also undoubtedly contributes to the growth of human capital in the agricultural sector while promoting sustainable practices through research and extension activities. In essence, indoor vertical farming is not only a means of urban food production, but also a transformative force driving sustainable urban agriculture into the future14.
Technology innovations for urban farmers and breeders
The phenotype, or the observable traits of a plant, is an outcome of the intricate interplay between its genotype and the growing environment. Technological innovations in the fields of engineering, data science and genetics have brought about significant advances in the way farmers and breeders grow and improve crop ideotypes for different environments (Fig. 2). These innovations, encompassing phenotyping capabilities, precision agriculture, and next-generation sequencing (NGS), have not only revolutionized the analysis and selection of crops, but also propelled agriculture towards greater efficiency, productivity, and sustainability15.

The combination of next-generation sequencing (NGS), precision agriculture, and phenotyping capability has revolutionised the way how breeders generate new crop varieties with traits suitable for indoor vertical farming.
Next-generation sequencing (NGS) is a cutting-edge technology that enables breeders to analyse the genomes or transcriptomes of crops in unprecedented detail16,17,18 (Fig. 2). By decoding the entire DNA sequence or transcriptomes of a plant associated with the phenotypes, breeders can identify the molecular markers or genes underpinning specific traits, such as increased yield or disease resistance. This wealth of genetic information empowers breeders to develop new crop varieties precisely adapting to their environments. The cost of performing NGS has decreased considerably over the years, allowing researchers to perform studies with a bigger sample sizes19. This has enabled breeders to perform genomics-assisted breeding by analysing the crop genetic background much more accurately than was previously possible20. The introduction of Single Molecule Real Time (SMRT) sequencing, nanopore sequencing and optical mapping enables long DNA fragments to be sequenced and assembled together for comprehensive genome analysis21,22,23. Recent plant genome sequencing projects have revealed that single reference genomes do not represent the diversity within a species24. Thus, the concept of pan-genomes is proposed to represent the genomic diversity of a species to determine its evolution through the presence and absence of genes25. For example, investigation of pan-genomes in Brassica oleracea has revealed around sixty genes linked to fungal diseases resistance26. The use of pan-genomes allows for phenotypic data to be linked to different types of genetic variations, thus, increasing the accuracy and efficiency of plant breeding27.
Precision agriculture is a technology-driven approach that empowers farmers with real-time control over environmental factors28 (Fig. 2). By using sensors, mapping, and other technologies, farmers can monitor and optimise essential variables, such as measuring nutrient contents of the soil and optimizing the use of water and fertilizers, thus resulting in more efficient and sustainable agriculture29,30. For breeders, precision farming provides invaluable insights into crop growth and development under specific environmental parameters, facilitating informed decisions about trait selection in breeding programmes. Further integration of IoT (Internet of Things) sensors and machine augments agricultural robustness by generating predictive models for diverse environmental and crop parameters31,32,33.
Phenotyping capability refers to the quantitative and qualitative assessment of the traits of a given plant in a chosen environment34 (Fig. 2). Plant phenotyping is an emerging field that links observable plant characteristics to the genetic and genomic background35,36. This involves the measurement of various traits, such as growth rate, yield, disease resistance, and nutritional contents, to identify the appropriate plants for breeding purposes. Currently, rapid development is taking place particularly in the area of non-destructive, imaging-based platforms for the evaluation of plant morphology, growth attributes and biochemical composition36. These imaging platforms include visible imaging (machine vision), imaging spectroscopy (multispectral and hyperspectral remote sensing), thermal infrared imaging, fluorescence imaging, 3D imaging, and tomographic imaging37,38. Digital phenotyping, the use of high-throughput imaging sensors for large-scale studies, has enabled breeders to efficiently and accurately analyse thousands of crop images in computerised processes, thus facilitating effective selection of the most promising plants from a huge population for breeding programmes38,39,40,41. By reducing the cost and time associated with digital phenotyping, breeding programmes can be further expanded, accelerating breeding cycles and enhancing the rate of genetic gain42.
The convergence of NGS, precision agriculture, and phenotyping capability is revolutionizing the way breeders work, enabling them to develop crops in an efficient, productive, and sustainable way under different environments (Fig. 2). These technologies empower breeders to pinpoint superior plants for their breeding programmes, monitor real-time crop growth and development, and scrutinize crop genetics in unprecedented detail, ultimately resulting in more effective and sustainable breeding efforts.
Optimization of traits suitable for indoor vertical farming
Indoor vertical farming has been gaining popularity as a more efficient and sustainable method of agriculture, particularly in urban areas with high population density43. However, despite its promise, its widespread adoption has been hindered by several economic and technical challenges. The profitability and market penetration of indoor vertical farming face challenges attributed to high upfront costs, energy-intensive lighting requirements, and a nascent market with a shortage of skilled labour44.
The controlled environment, limited space, and vertical orientation in indoor vertical farming present unique challenges and opportunities, along with the special benefits offered to the local community (Fig. 1). Genetic breeding, particularly through the lens of translational genomics stands out as a pivotal factor in enhancing the economic viability of indoor vertical farming45. By employing selective breeding and genetic engineering, crops can be customized to excel in the specific conditions of controlled indoor environments, leading to improved economic outcomes. Through these genetic interventions, plants can be tailored for traits such as compact growth, optimized nutrient absorption, and adaptability to artificial lighting, all of which contribute to increased yield and efficiency. This genetic optimization reduces dependence on external inputs, such as energy, water and fertilisers, leading to lowered operational costs. In addition, accelerated breeding programmes enable the swift development of crop varieties tailored to market demands, ensuring a competitive edge in the evolving landscape of indoor farming. The economic benefits extend further as genetic enhancements streamline cultivation processes, minimizing labour costs, and maximizing the efficient use of limited space. While some traits relevant to traditional farming practices, such as yield and growth rate are still applicable, many other traits take on greater significance in the context of indoor vertical farming.
Here, we have strategically clustered these indoor farming traits into three different groups, namely for maximising productivity, resource requirements/utilisation and add value to consumers (Fig. 3). The first group is meticulously curated for maximizing productivity, focusing on traits such as high yield and rapid growth rates to ensure efficient crop turnover and increased overall output/productivity. The second group is designed to enhance resource management, featuring traits like compact architecture that optimize space utilization, streamline operational processes including mechanised harvesting. Lastly, the third group is tailored to cater to consumer preferences spotlighting features like unique flavours, visual enhancement, and nutritional enrichment, which collectively elevate the market appeal and quality of the produce. This triad of groups reflects a comprehensive approach, where traits can be harmonized together to cultivate a thriving and economically viable indoor vertical farming ecosystem.

The controlled environmental parameters, confined spatial constraints, and vertical orientation pose distinctive challenges, while concurrently affording opportunities for the selective breeding of crops bearing desirable traits amenable to indoor vertical farming practices with a sustainability focus.
Vertical farming encompasses a diverse array of cultivation systems, each tailored to specific crop requirements and environmental conditions46. Hydroponic systems, for instance, suspend plant roots in a nutrient-rich solution, allowing for precise control over nutrient delivery and water uptake. This method is particularly suited to crops like lettuce, spinach, and herbs, which thrive in oxygenated water environments. Aeroponic systems, on the other hand, deliver a fine mist or air directly to plant roots, promoting rapid growth and efficient nutrient absorption. These systems are ideal for crops requiring high oxygen levels, such as leafy greens and strawberries47. In addition, soil-based vertical farming utilizes traditional soil substrates to support plant growth, providing essential nutrients and microbial interactions crucial for the development of crops with complex root systems. Each cultivation method offers distinct advantages and challenges, requiring careful consideration of crop adaptability, genetic traits, and agronomic practices to optimize yield and sustainability in vertical farming operations.
Currently, some crop types, such as leafy greens, fruits, microgreens and herbs13, are cultivated in indoor vertical systems with satisfactory growth and yield. Leafy greens, such as lettuce are suitable for indoor farming as they are more compact in size with high growth rate and yield per unit area2. Fruits, such as tomatoes and strawberries are considered to be high-value crops and increasingly adopted to indoor vertical farming9. The controlled environment has ensured stable and predictable yields without production disruptions because of abnormalities in weather patterns and biotic stresses. In addition, the control of light and temperature enables the fruiting process to be synchronised. Microgreens and herbs are rich in phytonutrients, such as polyphenols, terpenoids and N-containing metabolites, and can be harvested in short periods of time with good yields and high growth rates48,49. Other crops, such as maize, wheat and rice, can also be grown in indoor vertical farms. Traits that contribute to higher yields, such as rapid growth rate, high biomass accumulation, and multiple harvest cycles per year, are essential for maximizing output within the limited vertical farming space. Breeding high-yielding varieties is identified as a major contributor for improving both economic viability and environmental impact in indoor vertical farming50. While the theoretical yields of grain crops can reach up to 200–600 times of the current yield per area, the indoor wheat farming has been predicted to be less economically competitive than field farming because of the long cycling time51. Thus, whether grain crops could be the main crop type for indoor vertical farming is still questionable. In tomato, the flowering gene SINGLE FLOWER TRUSS increases yield by up to 60% through heterosis and overdominance52. In lettuce, the knockout of flowering time-related transcription factors LsSOC1 and LsSPL13 increases lettuce yield significantly53,54.
Given the heavy reliance on artificial lighting in indoor vertical farms, traits that enhance light absorption, such as high photosynthetic efficiency, prove invaluable in reducing energy consumption and operational costs55,56. In indoor vertical farming, electricity usage is approximately 250 kWh/kg/year, while traditional farming methods typically require only about 0.3 kWh/kg/year. Thus, there is a multitude of research on increasing the resource use efficiency through higher efficiency LED, shifting electrical demand and generating plants with high photosynthesis rate with lower energy spectrum11,57,58. The growth rate of crops emerges as an important factor in maximizing productivity. A rapid and vigorous growth rate directly influences the crop turnover for herbs and leafy vegetables, enabling more frequent planting and harvesting cycles. This accelerated growth trajectory aligns with the goal of optimizing resource utilization and space efficiency, resulting in increased overall productivity and profitability. The expeditious maturation of plants not only allows for a quicker return on investment but also enhances the adaptability of indoor farming operations to market demands.
In the pursuit of enhancing resource management in indoor vertical farming, the utilization of limited space becomes a critical consideration. By growing plants in stacked layers or vertical racks, traits that promote compact architecture, such as shorter internode lengths and dense foliage, enable more plants to be cultivated within a given area, maximizing productivity and yield. For example, in cucurbit crops, such as cucumber, pumpkin and watermelon, an enhanced translation of a YABBY1 transcription factor shortens the internode length and increases the compactness of plant without compromising yield59,60. In addition, the timing of flowering significantly impacts plant architecture, as demonstrated in tomatoes through the gene editing of SELF PRUNING (SP), SELF PRUNING 5G (SP5G), and tomato ERECTA (SlER)3. This genetic manipulation induces early flowering, encourages compact stem growth, and regulates shoot development, ultimately aiming to optimize plant architecture for vertical farming3. Ease of harvest holds the key to a spectrum of advantages. Crops designed for easy harvest, characterized by traits like uniform growth and readily detachable fruits, usher in operational efficiencies by simplifying the harvesting process. This streamlined approach translates to reduced labour intensity, enabling a more efficient use of human resources. In addition, the ease of harvest associated with these traits minimize the risk of crop damage, ensuring the delivery of a higher-quality harvest. The decrease in time needed for harvesting will foster a shorter cycle of planting and harvest, which directly contributes to heightened overall productivity. SP orthologs in Solanaceae plants, such as tomatoes, emerge as prime candidates for targeting fruit ripening synchronization due to their role in regulating determinate growth habits and axillary shoot development61. This approach holds promise for streamlining fruit harvesting processes with minimal handling.
Traits that promote self-pollination, such as self-fertile flowers or flowers with anthers and stigmas in proximity, enable plants to reproduce without external assistance. Indoor farms often face limitations in attracting pollinators because of the controlled environment and restricted access by natural pollination agents. Self-pollinating crops ensure reliable pollination and consistent yields, eliminating the need for manual pollination or reliance on pollinators62. In commercial fruit production, pollination is a limiting factor in yield and productivity63. In addition, in some crop species such as Brassicaceae and Solanaceae, the occurrence of self-incompatibility (SI) involves various genes and one of the most well-known gene families associated with SI is the S-locus gene family64,65. These genes play a crucial role in the recognition and rejection of self-pollen, preventing self-fertilization and promoting outcrossing. In many species, the S-locus contains multiple genes, including the S-RNase gene, which encodes a ribonuclease enzyme that degrades pollen RNA, and the S-locus F-box gene (SLF), which is involved in pollen recognition66. In addition, other genes involved in SI pathways include those encoding receptor-like kinases (RLKs) and other signalling proteins that mediate the pollen-pistil interactions necessary for SI recognition67.
Additional traits, such as enhanced nutrition, and unique colours and flavours (Fig. 3), hold significant influence for indoor vertical farming as they carry economical value to consumers68,69. Selecting and breeding plants with traits like enhanced nutrition, unique colours (e.g. purple leaf and purple-head trait in the leafy vegetable Chinese cabbage70,71), or novel flavours under the controlled environment could enhance the health benefits or sensory experience for consumers and open up opportunities for niche markets.
Notably, selection of the above traits suitable for indoor vertical farming will be based on various controlled growth conditions, which contrasts with breeding for traditional farming using various stresses or selective pressure mostly under outdoor conditions. This also ensures that the performance of the novel varieties selected for indoor farming can be reproduced under similar growth conditions.
Breeding of indoor farming crops by forward genetic approaches
Genetic breeding of indoor farming crops may follow the same forward genetic approaches for breeding field crops by selecting and crossing individuals with desirable traits suitable for indoor vertical farming72 (Fig. 4). These approaches could be integrated with various modern genetic and technological tools to create novel crop varieties bearing combined or improved beneficial traits in the context of indoor vertical farming with shortened breeding time and fewer resources required3,73.

In forward genetic approaches, the combination of next-generation sequencing and high-throughput phenotyping under controlled environmental conditions facilitates breeders to identify ideal parental lines to generate and select desirable hybrids through GWAS, eQTL analysis or genomic selection. In reverse genetic approaches, target genes are selected based on past functional genomics studies to generate the genetically modified lines with desirable traits through various mutation or silencing methods, such as the advanced CRISPR-Cas9 genome editing. RILs recombinant inbred lines, GWAS genome-wide association study, eQTL expression quantitative trait loci.
Germplasm or gene pool play a vital role in genetic breeding by forward genetic approaches, as they provide a source of genetic diversity required for improving the quality of crops74. Collection and exploration of wild and primitive cultivars permit identification of desirable traits that can be transferred to modern crops. These precious cultivars have evolved over thousands of years to adapt to different environmental conditions and various biotic stresses. As a result, they possess genetic diversity and unique traits that are not found in modern cultivated varieties. For example, some wild tomato cultivars contain higher levels of lycopene, a potent antioxidant linked to reduced risk of cancer and heart disease, than modern commercial varieties75. In general, modern cultivated varieties used in traditional farming usually have a narrow genetic base because of intensive selection and breeding that have taken place over the years76,77. This limits their ability to adapt to changing environmental conditions. By introducing desirable traits or genes from wild and primitive cultivars, breeders can broaden the genetic base of modern varieties suits to the microclimatic conditions of indoor farming. Careful management of germplasm resources is essential to ensure their continued availability for breeding of crops with desirable traits for indoor vertical farming78.
The selection of elite breeds has been largely influenced by recent technological advances from gathering genetic information to phenotypic characterization. The main breakthrough from these technologies is to establish and uncover new links between genotypes and phenotypes. The development of statistical methods and bioinformatic tools has laid the foundation for understanding how genetic differences can determine the performance of individual plants under different environment conditions79. As the phenotype is dependent on the interaction between genotype and environment (Fig. 2), it is important to cultivate genetically stable lines to generate progenies of desired genetic makeup in a shorter period. In particular, the use of double haploids or recombinant inbred lines complements in the genetic breeding studies involving wild accessions and enables more breeders to join into a concerted effort to improve crops80,81.
A powerful tool for identifying the genetic basis of complex traits in a population of plants is genome-wide association study (GWAS)82,83, which involves the analysis of large sets of genetic and phenotypic data, typically obtained from a diverse population of plants through high-throughput sequencing and phenotyping technologies (Fig. 2). The data are used to identify single nucleotide polymorphisms (SNPs) or other genetic markers across the genome. Statistical methods are applied to identify the association between genotypes and phenotypes of interest through comparing the frequency of each genetic marker/variant in individuals with or without the trait. One of the key advantages of GWAS lies in its ability to identify genetic variants responsible for complex traits, which are often controlled by multiple genes84. The power of the GWAS analysis depends on the number of accessions included in the study and the extent of genetic variations in the population85. Larger sample sizes are associated with higher statistical power as they allow for more precise estimates of effect sizes and reduce the impact of random variations. So far, GWASs have been routinely performed to generate more productive and higher quality crops through identifying genes associated with important traits, such as yield and disease tolerance85,86. In addition, GWASs could also be applied to plants mutagenized by chemicals and radiation, although this process is random in nature and relies on a large pool of population87.
Expression quantitative trait loci (eQTLs) refers to genomic loci that affect gene expression levels88. eQTL studies analyse genomic and transcriptomic datasets from the same individuals and allows breeders to understand how genetic variations influence the expression of genes, which in turn affects the relevant phenotypes89. Trait-associated SNPs are more likely to be relevant to differential gene expression90. eQTL analysis provides an additional dimension to understand how genetic variants affect phenotypes and contributes to identifying potential loci for breeding91. Recently, eQTL analysis has been combined with GWAS to provide increased statistical power and mapping resolution for uncovering novel genes, such as those affecting the initiation of secondary cell wall development in cotton, tocochromanol levels in maize grain, and flavours in strawberries92,93,94.
Genomic selection is routinely performed after genomic data analysis to generate crops with desired traits95. In this approach, the genotype and phenotype data are combined to generate a prediction model that links genetic markers with specific traits. Thereafter, the model is used to select the best lines for breeding based on the predicted performance with the combination of genetic variants. By using a large number of genetic markers covering the whole genome, genomic selection permits capturing more genetic variations that underlie a target trait96,97,98. This method is different from GWAS and eQTL analysis as the latter aims to identify individual genetic variants associated with the trait for marker-assisted breeding99. Thus, genomic selection can accelerate genetic gain and select superior individuals with a higher probability of success100,101. Moreover, it also minimizes the risk of negative traits being introduced into the breeding programmes.
After the selection of the parental lines through GWAS, eQTL analysis or genomic selection, the introduction of speed breeding can accelerate the breeding cycle of crops by manipulating the growth conditions to promote growth and development102. This approach is particularly useful for rapid generation cycling through single seed descent or crops with long breeding cycles, such as wheat, rice and tomatoes. For example, by extending the light duration to almost the whole day, wheat crops are able to mature after eight weeks, which is three times as fast as the traditional approach102. Optimizing speed breeding under various controlled environments can significantly shorten the overall duration of genetic breeding for indoor crops.
Breeding of indoor farming crops by reverse genetic approaches
Reverse genetic approach is commonly used to understand the function of a gene by analysing the phenotypes resulting from its disruption or mutation103. When this approach is applied to breeding of indoor farming crops, the candidate genes whose homologs and/or orthologs have been well characterized by functional genomics in model plants and other crop species104 can be selected for targeted mutagenesis by various mutation or silencing methods, such as the advanced CRISPR-Cas9 genome editing, to confer specific traits of interest (Fig. 4). Functional genomic studies in various plant species have established numerous connections between genome sequences and biological processes, enabling the selection and manipulation of potential candidate genes corresponding to desirable traits suitable for indoor vertical farming105. The reverse genetics approach has the advantage of rapid generation of modified lines with the traits of interest and bearing a relatively low number of deleterious alleles.
Among the various mutation and silencing approaches, CRISPR-Cas9 is a powerful molecular tool that has gained popularity in recent years for its capacity to alter the target DNA with high precision106,107. CRISPR, which stands for Clustered Regularly Interspaced Short Palindromic Repeats, is a natural defence mechanism found in bacteria and archaea that allows them to target and destroy invading viruses by cutting their DNA108. Cas9 is a dual RNA-guided DNA endonuclease enzyme associated with the CRISPR and acts as a molecular scissor that cuts the DNA at a specific location with the guidance of a guide RNA (gRNA). CRISPR-Cas9 has been used in many crop species to edit specific genes identified though GWAS and eQTL mapping as well as functional genomic studies109,110,111. In comparison to genomic selection, CRISPR-Cas9 is usually more effective for generating targeted traits caused by a small number of genes. This technique has been used in many plant species, such as leafy vegetables, tomatoes and even on fruit trees54,112,113, and is preferred particularly when cross breeding is not possible. In addition, speed breeding can be coupled with CRISPR-Cas9 genome editing to shorten the time for obtaining the mutants before phenotypic characterisation (Fig. 4). Commercially, modified plants generated using this approach is considered as not genetically modified (non-GMO) and can be readily introduced into the market114.
Concluding remarks
To realize the environmental, social, and economic potentials of indoor vertical farming in urban cities, it is critical to establish genetic breeding platforms to select and generate crop varieties bearing suitable traits for the controlled environment of indoor vertical farming. Technological innovations, such as next-generation sequencing, precision agriculture, and advanced phenotyping capabilities, have significantly accelerated the genetic breeding efforts for indoor farming. These tools enable breeders to analyse and understand the complex relationships between genotypes and phenotypes. Here, we highlight desirable traits with its potential genetic pathways to enable breeders to maximize productivity and quality within the limited vertical farming space and prepare for the evolving needs and preferences of consumers. The use of both forward and reverse genetic approaches combined with speed breeding has empowered breeders to identify potential genetic loci and generate targeted varieties with desirable traits in a shorter timeframe. Thus, the combination of advanced technologies, innovative breeding approaches, and the selection of traits best suited for controlled environments is paving the way for more efficient, sustainable, and resilient urban farming systems. In summary, genetic breeding of indoor farming crops stands as a pivotal component of sustainable agriculture, addressing the urgent challenges of global food security and sustainability in an increasingly urbanized world.
References
Economic, U. N. D. o. & Social Affairs, P. D. World Population Prospects 2022: Summary of Results. UN DESA/POP/2022/TR/NO. 3 (2022).
Benke, K. & Tomkins, B. Future food-production systems: vertical farming and controlled-environment agriculture. Sustainability: Sci. Prac. Policy 13, 13–26 (2017).
Kwon, C.-T. et al. Rapid customization of Solanaceae fruit crops for urban agriculture. Nat. Biotechnol. 38, 182–188 (2020).
Zamir, D. Improving plant breeding with exotic genetic libraries. Nat. Rev. Genet. 2, 983–989 (2001).
Brummer, E. C. et al. Plant breeding for harmony between agriculture and the environment. Front. Ecol. Environ. 9, 561–568 (2011).
Mougeot, L. J. Growing better cities: urban agriculture for sustainable development. (IDRC, 2006).
Pérez-Urrestarazu, L. et al. Vertical greening systems and sustainable cities. J. Urban Technol. 22, 65–85 (2015).
Wolfert, S. et al. Big data in smart farming—a review. Agric. Syst. 153, 69–80 (2017).
O’Sullivan, C. A. et al. Vertical farms bear fruit. Nat. Biotechnol. 38, 160–162 (2020).
SharathKumar, M. et al. Vertical farming: moving from genetic to environmental modification. Trends Plant Sci. 25, 724–727 (2020).
Wong, C. E. et al. Seeing the lights for leafy greens in indoor vertical farming. Trends Food Sci. Technol. 106, 48–63 (2020).
Kc, K. B. et al. When too much isn’t enough: does current food production meet global nutritional needs? PLoS ONE 13, e0205683 (2018).
Payen, F. T. et al. How much food can we grow in urban areas? Food production and crop yields of urban agriculture: a meta-analysis. Earth’s. Future 10, e2022EF002748 (2022).
Yoon, B. K. et al. Entrepreneurial talent building for 21st century agricultural innovation. ACS Nano 15, 10748–10758 (2021).
Krug, A. S. et al. The next era of crop domestication starts now. Proc. Natl Acad. Sci. USA 120, e2205769120 (2023).
He, J. et al. Genotyping-by-sequencing (GBS), an ultimate marker-assisted selection (MAS) tool to accelerate plant breeding. Front. Plant Sci. 5, 484 (2014).
Poland, J. A. & Rife, T. W. Genotyping-by-sequencing for plant breeding and genetics. Plant Genome 5, 92–102 (2012).
Varshney, R. K. et al. Next-generation sequencing technologies and their implications for crop genetics and breeding. Trends Biotechnol. 27, 522–530 (2009).
Kumar, K. R. et al. Next-generation sequencing and emerging technologies. Semin. Thromb. Hemost. 45, 661–673 (2019).
Varshney, R. K. et al. Genomics-assisted breeding for crop improvement. Trends Plant Sci. 10, 621–630 (2005).
Rhoads, A. & Au, K. F. PacBio sequencing and its applications. Genom. Proteom. Bioinform. 13, 278–289 (2015).
Branton, D. et al. The potential and challenges of nanopore sequencing. Nat. Biotechnol. 26, 1146–1153 (2008).
Tang, H. et al. Optical mapping in plant comparative genomics. GigaScience 4, s13742–13015 (2015).
Bayer, P. E. et al. Plant pan-genomes are the new reference. Nat. Plants 6, 914–920 (2020).
Tettelin, H. et al. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: implications for the microbial “pan-genome”. Proc. Natl Acad. Sci. USA 102, 13950–13955 (2005).
Bayer, P. E. et al. Variation in abundance of predicted resistance genes in the Brassica oleracea pangenome. Plant Biotechnol. J. 17, 789–800 (2019).
Della Coletta, R. et al. How the pan-genome is changing crop genomics and improvement. Genome Biol. 22, 1–19 (2021).
Gebbers, R. & Adamchuk, V. I. Precision agriculture and food security. Science 327, 828–831 (2010).
Cisternas, I. et al. Systematic literature review of implementations of precision agriculture. Comput. Electron. Agric. 176, 105626 (2020).
Shafi, U. et al. Precision agriculture techniques and practices: from considerations to applications. Sensors 19, 3796 (2019).
Shi, X. et al. State-of-the-art Internet of Things in protected agriculture. Sensors 19, 1833 (2019).
Sharma, A. et al. Machine learning applications for precision agriculture: a comprehensive review. IEEE Access 9, 4843–4873 (2020).
Siregar, R. R. A. et al. Vertical farming perspectives in support of precision agriculture using artificial intelligence: a Review. Computers 11, 135 (2022).
Pieruschka, R. & Schurr, U. Plant phenotyping: past, present, and future. Plant Phenomics 2019, 7507131 (2019).
Fiorani, F. & Schurr, U. Future scenarios for plant phenotyping. Annu. Rev. Plant Biol. 64, 267–291 (2013).
Watt, M. et al. Phenotyping: new windows into the plant for breeders. Annu. Rev. Plant Biol. 71, 689–712 (2020).
Dhondt, S. et al. Cell to whole-plant phenotyping: the best is yet to come. Trends Plant Sci. 18, 428–439 (2013).
Li, L. et al. A review of imaging techniques for plant phenotyping. Sensors 14, 20078–20111 (2014).
Yang, W. et al. Crop phenomics and high-throughput phenotyping: past decades, current challenges, and future perspectives. Mol. Plant 13, 187–214 (2020).
Lee, U. et al. An automated, high-throughput plant phenotyping system using machine learning-based plant segmentation and image analysis. PLoS ONE 13, e0196615 (2018).
Walter, J. et al. High-throughput field imaging and basic image analysis in a wheat breeding programme. Front. Plant Sci. 10, 449 (2019).
Araus, J. L. et al. Translating high-throughput phenotyping into genetic gain. Trends Plant Sci. 23, 451–466 (2018).
Al-Kodmany, K. The vertical farm: a review of developments and implications for the vertical city. Buildings 8, 24 (2018).
Song, S. et al. Comparison of vegetable production, resource-use efficiency and environmental performance of high-technology and conventional farming systems for urban agriculture in the tropical city of Singapore. Sci. Total Environ. 807, 150621 (2022).
Passioura, J. B. Translational research in agriculture. Can we do it better? Crop Pasture Sci. 71, 517 (2020).
Beacham, A. M. et al. Vertical farming: a summary of approaches to growing skywards. J. Hortic. Sci. Biotechnol. 94, 277–283 (2019).
Fussy, A. & Papenbrock, J. An overview of soil and soilless cultivation techniques—chances, challenges and the neglected question of sustainability. Plants 11, 1153 (2022).
Rajan, P. et al. Advancement in indoor vertical farming for microgreen production. Am. J. Plant Sci. 10, 1397 (2019).
Galieni, A. et al. Sprouts and microgreens: trends, opportunities, and horizons for novel research. Agronomy 10, 1424 (2020).
Song, S. et al. How crop breeding programs can improve plant factories’ business and environmental sustainability: Insights from a farm level analysis. Sustain. Prod. Consum. 44, 298–311 (2024).
Asseng, S. et al. Wheat yield potential in controlled-environment vertical farms. Proc. Natl Acad. Sci. USA 117, 19131–19135 (2020).
Krieger, U. et al. The flowering gene SINGLE FLOWER TRUSS drives heterosis for yield in tomato. Nat. Genet 42, 459–463 (2010).
Beracochea, V. et al. CRISPR/Cas9-mediated knockout of SPL13 radically increases lettuce yield. Plant Cell Rep. 42, 645–647 (2023).
Choi, S. H. et al. Development of late-bolting plants by CRISPR/Cas9-mediated genome editing from mesophyll protoplasts of lettuce. Plant Cell Rep. 41, 1627–1630 (2022).
Long, S. P. et al. Can improvement in photosynthesis increase crop yields? Plant Cell Environ. 29, 315–330 (2006).
Zhu, X.-G. et al. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol. 61, 235–261 (2010).
Avgoustaki, D. D. & Xydis, G. Energy cost reduction by shifting electricity demand in indoor vertical farms with artificial lighting. BioSyst. Eng. 211, 219–229 (2021).
Avgoustaki, D. D. & Xydis, G. How energy innovation in indoor vertical farming can improve food security, sustainability, and food safety? Adv. Food Secur. Sustain. 5, 1–51 (2020).
Wang, S. et al. Architecture design of cucurbit crops for enhanced productivity by a natural allele. Nat. Plants 8, 1394–1407 (2022).
Dong, N.-Q. & Lin, H.-X. Compact plants enhance productivity. Nat. Plants 8, 1335–1336 (2022).
Pnueli, L. et al. The SELF-PRUNING gene of tomato regulates vegetative to reproductive switching of sympodial meristems and is the ortholog of CEN and TFL1. Development 125, 1979–1989 (1998).
Wurz, A. et al. Hand pollination of global crops—a systematic review. Basic Appl. Ecol. 56, 299–321 (2021).
Benjamin, F. E. & Winfree, R. Lack of pollinators limits fruit production in commercial blueberry (Vaccinium corymbosum). Environ. Entomol. 43, 1574–1583 (2014).
Moreels, P. et al. Intra-and inter-specific reproductive barriers in the tomato clade. Front. Plant Sci. 14, 1326689 (2023).
Schopfer, C. R. et al. The male determinant of self-incompatibility in Brassica. Science 286, 1697–1700 (1999).
McClure, B. et al. Compatibility and incompatibility in S-RNase-based systems. Ann. Bot. 108, 647–658 (2011).
Shiu, S.-H. & Bleecker, A. B. Plant receptor-like kinase gene family: diversity, function, and signaling. Sci.’s. STKE 2001, re22–re22 (2001).
Saltzman, A. et al. Availability, production, and consumption of crops biofortified by plant breeding: current evidence and future potential. Ann. N. Y. Acad. Sci. 1390, 104–114 (2017).
Garcia-Casal, M. N. et al. Staple crops biofortified with increased vitamins and minerals: considerations for a public health strategy. Ann. N. Y. Acad. Sci. 1390, 3–13 (2017).
He, Q. et al. The novel gene BrMYB2, located on chromosome A07, with a short intron 1 controls the purple-head trait of Chinese cabbage (Brassica rapa L.). Horti. Res. 7, 97 (2020).
Wang, W. et al. Mapping the BrPur gene for purple leaf color on linkage group A03 of Brassica rapa. Euphytica 199, 293–302 (2014).
Acquaah, G. Principles of Plant Genetics and Breeding (Wiley, 2012).
Cobb, J. N. et al. Next-generation phenotyping: requirements and strategies for enhancing our understanding of genotype–phenotype relationships and its relevance to crop improvement. Theor. Appl. Genet. 126, 867–887 (2013).
Smith, S. et al. Genetic diversity and modern plant breeding. Genet. Divers. Erosion in Plants: Indicators and Prev. 55–88 (2015).
Kaur, C. et al. Functional quality and antioxidant composition of selected tomato (Solanum lycopersicon L) cultivars grown in Northern India. LWT-Food Sci. Technol. 50, 139–145 (2013).
Cooper, H. D. et al. Broadening the genetic base of crops: an overview. Broadening the Genet. Base of Crop Prod. 1–23 (2001).
Hyten, D. L. et al. Impacts of genetic bottlenecks on soybean genome diversity. Proc. Natl Acad. Sci. USA 103, 16666–16671 (2006).
Tanksley, S. D. & McCouch, S. R. Seed banks and molecular maps: unlocking genetic potential from the wild. Science 277, 1063–1066 (1997).
Zhu, C. et al. Status and prospects of association mapping in plants. Plant Genome 1, 5–20 (2008).
Burr, B. & Burr, F. A. Recombinant inbreds for molecular mapping in maize: theoretical and practical considerations. Trends Genet 7, 55–60 (1991).
Smith, J. et al. Use of doubled haploids in maize breeding: implications for intellectual property protection and genetic diversity in hybrid crops. Mol. Breed. 22, 51–59 (2008).
Tibbs Cortes, L. et al. Status and prospects of genome-wide association studies in plants. Plant Genome 14, e20077 (2021).
Wang, Q. et al. Advances in genome-wide association studies of complex traits in rice. Theor. Appl. Genet. 133, 1415–1425 (2020).
Zhu, Z. et al. Integration of summary data from GWAS and eQTL studies predicts complex trait gene targets. Nat. Genet. 48, 481–487 (2016).
Huang, X. et al. Genome-wide association studies of 14 agronomic traits in rice landraces. Nat. Genet. 42, 961–967 (2010).
Huang, X. & Han, B. Natural variations and genome-wide association studies in crop plants. Annu. Rev. Plant Biol. 65, 531–551 (2014).
Oladosu, Y. et al. Principle and application of plant mutagenesis in crop improvement: a review. Biotechnol. Biotechnol. Equip. 30, 1–16 (2016).
Le Nguyen, K. et al. Next-generation sequencing accelerates crop gene discovery. Trends Plant Sci. 24, 263–274 (2019).
Zhang, L. et al. Genome-wide analysis of expression quantitative trait loci (eQTLs) reveals the regulatory architecture of gene expression variation in the storage roots of sweet potato. Hortic. Res. 7, 90 (2020).
Nicolae, D. L. et al. Trait-associated SNPs are more likely to be eQTLs: annotation to enhance discovery from GWAS. PLoS Genet 6, e1000888 (2010).
Miculan, M. et al. A forward genetics approach integrating genome-wide association study and expression quantitative trait locus mapping to dissect leaf development in maize (Zea mays). Plant J. 107, 1056–1071 (2021).
Fan, Z. et al. A multi-omics framework reveals strawberry flavor genes and their regulatory elements. N. Phytol. 236, 1089–1107 (2022).
Li, Z. et al. Combined GWAS and eQTL analysis uncovers a genetic regulatory network orchestrating the initiation of secondary cell wall development in cotton. N. Phytol. 226, 1738–1752 (2020).
Wu, D. et al. Combining GWAS and TWAS to identify candidate causal genes for tocochromanol levels in maize grain. Genetics 221, iyac091 (2022).
Xu, Y. et al. Genomic selection: A breakthrough technology in rice breeding. Crop J. 9, 669–677 (2021).
Yadav, S. et al. Accelerating genetic gain in sugarcane breeding using genomic selection. Agronomy 10, 585 (2020).
Werner, C. R. et al. How population structure impacts genomic selection accuracy in cross-validation: implications for practical breeding. Front. Plant Sci. 11, 592977 (2020).
Stewart-Brown, B. B. et al. Genomic selection for yield and seed composition traits within an applied soybean breeding program. G3. Genes Genomes Genet 9, 2253–2265 (2019).
Collard, B. C. & Mackill, D. J. Marker-assisted selection: an approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. Lond., Ser. B: Biol. Sci. 363, 557–572 (2008).
Voss-Fels, K. P. et al. Accelerating crop genetic gains with genomic selection. Theor. Appl. Genet. 132, 669–686 (2019).
Xu, Y. et al. Enhancing genetic gain through genomic selection: from livestock to plants. Plant Commun. 1, 100005 (2020).
Watson, A. et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 4, 23–29 (2018).
Jankowicz-Cieslak, J. & Till, B. J. Forward and reverse genetics in crop breeding. Adv. Plant Breed. Strategies: Breeding, Biotechnol. Mol. Tools, 215–240 (2015).
Bevan, M. W. et al. Genomic innovation for crop improvement. Nature 543, 346–354 (2017).
Zhang, H. et al. Genome editing—principles and applications for functional genomics research and crop improvement. Crit. Rev. Plant Sci. 36, 291–309 (2017).
Hsu, P. D. et al. Development and applications of CRISPR-Cas9 for genome engineering. Cell 157, 1262–1278 (2014).
Doudna, J. A. & Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 346, 1258096 (2014).
Wiedenheft, B. et al. RNA-guided genetic silencing systems in bacteria and archaea. Nature 482, 331–338 (2012).
Ma, X. et al. CRISPR/Cas9 platforms for genome editing in plants: developments and applications. Mol. Plant 9, 961–974 (2016).
Liu, Q. et al. Application of CRISPR/Cas9 in crop quality improvement. Int. J. Mol. Sci. 22, 4206 (2021).
Bao, A. et al. The CRISPR/Cas9 system and its applications in crop genome editing. Crit. Rev. Biotechnol. 39, 321–336 (2019).
Nishitani, C. et al. Efficient genome editing in apple using a CRISPR/Cas9 system. Sci. Rep. 6, 31481 (2016).
Ueta, R. et al. Rapid breeding of parthenocarpic tomato plants using CRISPR/Cas9. Sci. Rep. 7, 507 (2017).
Gupta, S. et al. Genetically modified crop regulations: scope and opportunity using the CRISPR-Cas9 genome editing approach. Mol. Biol. Rep. 48, 4851–4863 (2021).
Acknowledgements
We apologize to those authors whose research could not be cited due to space limits. Preparation of this review was supported by Singapore Food Story R&D Program (SFS_RND_SUFP_001_04), the Reimagine Research Grant from the National University of Singapore, and the intramural research support from Temasek Life Sciences Laboratory.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Teo, Z.W.N., Yu, H. Genetic breeding for indoor vertical farming. npj Sustain. Agric. 2, 13 (2024). https://doi.org/10.1038/s44264-024-00021-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s44264-024-00021-5
This article is cited by
-
The emergence of indoor agriculture as a driver of global energy demand
npj Sustainable Agriculture (2025)
-
Vertical farming of duckweed: advancing edible vaccine production for global health
Discover Biotechnology (2025)
