
Abstract
Intercropping, a cornerstone of ecological intensification for sustainable agriculture and biodiversity conservation, has not fulfilled its potential to support global food-security promises under growing land and climate constraints. A major barrier lies in the context-dependent and often unpredictable nature of yield benefits, which emerge from complex interspecific interactions that remain poorly characterized across agroecosystems. Through a novel global meta-analysis (4,195 partial Land Equivalent Ratio observations from 334 studies across 60 countries) coupled with machine learning to disentangle the roles of plant functional traits and species-specific interactions, we demonstrate a substantial untapped potential to increase the production of major cereals—maize ( + 62.4%), barley ( + 6.3%), and wheat ( + 0.1%)—solely through optimized deployment of intercropping on existing agricultural land. Crucially, despite reduced planting density in intercropping, the mean partial Land Equivalent Ratio (pLER) of 0.79 (95% CI: 0.76-0.82) indicates that yield reductions for component species are proportionally smaller than the decrease in density, revealing consistent beneficial interactions. Our quantitative framework identifies relative planting density (RD), temporal niche differentiation (TND), and relative height difference as key levers for optimizing intercropping performance. We further unveil a predictable trade-off governed by asymmetric competition: targeted manipulation of RD and TND can selectively benefit either taller or shorter species, providing mechanistic insights into how interspecific dynamics shape intercropping success. This finding offers a scalable and ecologically grounded pathway to increase global crop production without cropland expansion, advancing sustainable agricultural intensification.
Similar content being viewed by others
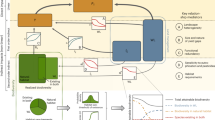
Introduction
Intercropping, the simultaneous cultivation of multiple crops within the same field during overlapping growth periods1, holds transformative potential for sustainable and resilient food systems. Yet, its performance is often unpredictable. Studies report widely varying effects of intercropping on the overall agronomic performance of the mixture2, with some leading to significant yield gains, improved resource efficiency3, reduced environmental impacts3,4 and restored biodiversity5, while others report opposite outcomes6,7. This variability reveals critical gaps in understanding interspecies interactions across crops and environmental contexts, limiting the development of robust and locally tailored intercropping strategies. Addressing these gaps is critical to unlock the full potential of intercropping and meet 21st-century agricultural demands while minimizing environmental impacts.
Crop associations determine resource partitioning and competition8. Geographic context shapes niche availability through edaphic and climatic constraints3. Agronomic practices, such as fertilization, directly alter resource competition, exemplified by nitrogen’s variable impact on maize-legume yields9,10. A system-level approach integrating these interactions is therefore crucial for predictive modeling and maximizing intercropping performance. This requires rigorous statistical analyses of key system indicators and characteristics5. Overlooking these contextual dependencies undermines intercropping potential for sustainable agriculture and limits the development of effective policies to harness its full benefits.
Functional traits distances -representing dissimilarity in species’ functional characteristics-offer thus promising framework for understanding intercropping variability by determining how species interact within mixtures11,12. Divergence in traits like root depth, leaf area, and canopy height shape species interactions, influencing competition and facilitation for resources such as water, nutrients, and light11. Complementary root structures enable species to exploit different soil niches, while variation in canopy architecture optimizes light capture, reducing competition, and enhancing productivity13,14. Agronomic practices, such as planting density and timing, modulate these interactions, with strategic management potentially reducing competitive pressures and improving yield15.
Although farmers consider various factors when deciding to intercrop-including pest and disease management and effects on succeeding crops-yield relative to monocropping systems remains a central concern. This practical concern is pivotal when assessing the feasibility of intercropping in terms of market demands, end-use quality, and economic profitability, which drive adoption rates and scaling of intercropping practices16. While the Land Equivalent Ratio (LER) provides insights, the partial Land Equivalent Ratio (pLER) better captures these dynamics17, but requires careful consideration of crop associations, geographic context, and agronomic practices3,18.
Here, we conducted a meta-analysis to quantify the global effect of intercropping on pLER. Based on machine learning algorithms, we identified the most influential explanatory variables among a wide range of potential factors related to pedoclimatic conditions, management practices, and crop characteristics. We then employed linear mixed models to dissect the individual and interactive functional effects of these key variables. Finally, we quantified the potential increase in global crop production of the five most popular mixtures without changing their respective areas. By shifting the focus from specific crop combinations to the functional diversity of species, we uncovered how differences in ecological niches of the species in the mixture drive variability in intercropping yields. Our extensive database, which included 2503 pairwise comparisons and 4195 pLER values from 334 primary studies across 60 countries, provides a comprehensive foundation for analysis (Fig. 1). With 50% of the crop pairs being cereal-legume associations, and maize dominating (e.g., maize-bean, maize-cowpea, and maize-soybean), our database accurately reflects global agricultural cultivation patterns. Additionally, it captures a diversity of cropping practices in intercropping systems, including variations in density, fertilization, and timing of sowing and harvesting—demonstrating the versatility of practices and the potential that such diversity could support yield optimization under various management scenarios. This approach refines our understanding of the mechanisms underlying intercropping performance and delivers actionable insights to farmers seeking to optimize yields in diverse agroecological contexts

Green areas represent the percentage of cropland based on Ramankutty et al.42.
Results and discussion
Relative density and temporal niche differentiation shape intercropping outcomes
Our global meta-analysis reveals that, on average, intercropping achieves a partial Land Equivalent Ratio (pLER) of 0.79 (95% bootstrap CIs: 0.76–0.82) (Figs. 1 and 2a). Random-forest analysis identified Relative Density (RD), Temporal Niche Differentiation (TND, i.e., differences in planting and harvesting times among species) and relative height difference as the most influential factors, which together accounted for 40% of the total variable importance (Fig. 2b). Increasing RD from 0.5 (50% of monocrop density) to 1 resulted in a 58% increase in pLER from 0.55 (CIs: 0.5-0.6) to 0.87 (CIs: 0.82–0.92), confirming that higher relative density generally enhances intercrop relative yield19. Yet, increasing the density of one species often leads to a reduction in the density of other species in the association19, as the total plant density is ultimately constrained by the available resources. Lower optimal densities, often observed in intercropping compared to monoculture20, are a direct result of balancing interspecific competition with complementary resource acquisition strategies.

a Histogram of relative yield (pLER). b Importance of moderators in determining the yield change of the intercropping system. The dashed line, solid red line and the shaded area in a represent pLER = 1, the mean effect size and the 95% bootstrap confidence intervals for the mean effect size. The numbers denote the total observations with the numbers of studies in parenthesis. The relative importance in (b) is quantified based on metaforest model and expressed as percentage relative to the top variable ‘Relative Density’, which is set to 100%.
Increasing temporal niche differentiation from 0.5 to a 50% growth period overlap—to 0.96—nearly distinct growth cycles—resulted in a mean pLER increase of 43%, from 0.44 (CIs: 0.34-0.52) to 0.63 (CIs: 0.43–0.82), highlighting the increased availability of resources due to longer growing seasons18. Yet, temporal niche differentiation is not always a flexible parameter; the local context, particularly the length of the growing season (Supplementary Fig. 1), can impose strong constraints, limiting the extent to which it can be managed. Complete temporal niche differentiation (TND = 1), akin to approaching sequential cropping, reduces the synergistic resource partitioning typical of relay intercropping (TND < 1), which has been shown to globally enhances light21 and nutrient use for higher yields22, and can also improve soil fertility, water retention, and pest control.
Predominance of asymmetric competition in intercropping systems for light
Asymmetric competition plays a critical role in intercropping systems, driven by the interplay of plant sizes, TND and RD (Fig. 3 and Supplementary Table 1). Plant height significantly modulates the impact of RD on pLER. Taller plants (height difference >0.5 m compared to their intercrop species) exhibit a greater pLER response to increasing RD (slope = 0.82, CIs: 0.73–0.91, p-value < 0.001) compared to smaller plants (slope = 0.21, CIs: 0.11-0.31, p-value < 0.001) (Fig. 3a). Increasing RD from 50% to 100% of monocropping density results in a 45% pLER increase for taller species and only a 19% increase for smaller species (Fig. 3a). A relative density of 0.9 is required to achieve a pLER of 1 for taller plants (TND at the mean observed value of 0.24), while this threshold is never reached for smaller plants (Fig. 3a). This process, often described as ‘size-asymmetric competition'23, can exacerbate yield disparities depending on species composition and environmental context, by limiting available photosynthetically active radiation, carbon assimilation and biomass allocation in suppressed plants. This phenomenon is evident in maize-soybean systems, where maize dominates light capture at the expense of soybean15, as well as in wheat-chickpea intercropping, where wheat delays chickpea development and reduces yield24.

Relationship between the pLER and the relative density (a, b) and the temporal niche differentiation (b). Colored lines represent the median response based on the best-fit linear mixed model in relation to RD or TND for each height difference (a, b) or TND category (c). Shaded lines indicate the 95% confidence intervals obtained via bootstrap. Example of interpretation for Fig. 3a (the same interpretation scheme applies to Fig. 3b, c): For the same mean TND of 0.24 and a relative density of 1, a crop with a relative height difference below -0.5 (meaning that the plant is smaller than its associated species) is predicted by our model to have a lower pLER than a crop with a relative height difference above 0.5 (meaning that the plant is taller than its associated species.
TND also mitigates asymmetric competition, particularly benefiting smaller plants, which showed a positive pLER response to increased TND (slope = 0.46, CIs: 0.27–0.64, p-value < 0.001), benefiting from reduced competition through temporal partitioning. Larger plants, conversely, exhibit a negative pLER response to TND (Fig. 3b slope = −0.1, CIs: −0.28 to 0.08, p-value < 0.001), likely reflecting their pre-established competitive dominance in the mixture. These results highlight the potential of strategic temporal adjustments to mitigate size-based competition18. For example, decreasing the co-growth duration in maize-soybean systems has been shown to enhance soybean biomass accumulation and competitive resilience, illustrating how strategic temporal adjustments can alleviate size-based competition15. Similarly, adjusting planting schedules in wheat-soybean intercrops optimizes resource capture by reducing overlap with the cereal crop25.
While plant height modulates the effects of both RD and TND, their combined manipulation also offers a promising approach to reduce competition and enhance resource use efficiency in intercropping systems. Low TND significantly reduces pLER, especially at relative density below 1. The pLER response to increasing RD is steeper under full temporal overlap (slope = 0.64, CIs: 0.56–0.72, p-value < 0.001) compared to weak temporal overlap (slope = 0.27, CIs: 0.11–0.43, p-value < 0.001) (Fig. 3c). At lower relative densities, pLER is higher with weak temporal overlap. For example, at RD = 0.5, pLER is 0.54 with full temporal overlap and 0.75 for weak temporal overlap. However, at RD = 1, pLER values converge across TND groups. The steeper pLER response to RD at low TND (slope = 0.64 vs. 0.27 for high TND) could reflect the limited capacity for phenotypic plasticity to offset resource competition under high relative density. At lower RD, plastic adjustments in biomass allocation and morphology mitigate competitive stress when TND is low, i.e., full temporal overlap. In contrast, when TND is high, competition for resources is inherently reduced, limiting the sensitivity of pLER to changes in RD. However, at high RD, resource availability becomes the dominant limiting factor, constraining compensatory responses and driving pLER convergence across TND levels. Thus, optimizing vertical canopy stratification and pairing erectophile and planophile leaf architectures, could minimize shading while maintaining light capture even under low TND and high total relative density26.
Effective management of planting density, spatial arrangement, and sowing timing can shift the balance from competition to facilitation. Low TND may intensify competition, but species-specific adjustments can foster positive interactions22, mitigating competitive stress. While Relative height difference emerged as a major factor influencing pLER, it does not fully capture the dynamic interactions between competition and facilitation during crop development13. For instance, taller plants may initially dominate light capture. This effect can be mitigated over time by belowground complementarity27, hydraulic lift28, or phenological shifts11. These processes help explain why systems with species exhibiting different root architectures or photosynthetic strategies tend to achieve more stable yields despite initial competition29, indicating that strategies should thus be tailored to specific crop types to maximize intercropping benefits. Using a Pareto front approach, optimal combinations (i.e., TND and RD for each species within the association) to maximize pLER for both species were identified (Fig. 4). While an equal relative density (1:1, i.e. additive intercropping) seems optimal for maize-soybean systems, other crop associations followed an augmentative pattern (total relative density between 1 and 2), where increasing the relative density of one species required reducing that of the other19. Consistent with previous research26, optimum relative densities for cereals never fell below 0.52, whereas for legumes the optimum could be lower, with faba bean reaching a relative density as low as 0.3 (Fig. 4). Additionally, each association exhibited multiple optimal configurations allowing for local adaptations based on environmental conditions (e.g., length of growing season) and production priorities.

The light blue area illustrates the possible space where the optimal relative densities of both species correspond to an augmentative total relative density (the sum of both densities between 1 and 2). The blue line represents a total relative density sum of 1, while the yellow line represents a sum of 2.
Scaling up intercropping for increasing global food production
Partially converting monoculture land to full overlapping intercropping systems (TND = 0), while optimizing relative density at the global scale, results in scenarios where production increase for all species in the five most frequent associations in our dataset. These increases ranged from +0.5 Mt for wheat in the wheat-faba bean intercropping to +701.9 Mt for maize in the maize-soybean intercropping (Fig. 5). The positive effects of intercropping resulted in increasing maize, barley, and wheat production by 62.4% (maize-soybean), 6.3% (barley-pea), and 0.1% (wheat-faba bean) respectively. These gains are far from negligible considering projected yield declines due to climate change- particularly for maize, which may decrease by up to 50% by the end of the century under the SSP585 scenario30. These gains are also particularly relevant considering projections that global food production will need to increase by approximately 70% by 2050 to adequately feed the growing world population31.

Total crop-specific land area remains constant. Dotted lines link crops within the same intercrop association. Bold black numbers indicate production gains (in Mt) compared to the 2013-2023 FAO 10-year average. Example of interpretation: Among a possible combination of optimal RD solutions identified using the Pareto front method (see Materials and Methods section), when the RD of faba bean = 0.29 and that of wheat = 0.74, the simulated pLER values obtained from our model (Eq. 9, Materials and Methods) are 0.73 for wheat and 0.46 for faba bean. If this combination (with these parameter values) were applied globally, it could lead to an increase in wheat production of 5.7 Mt and in faba bean production of 38.8 Mt relative to the 2013–2023 FAO 10-year average.
Furthermore, these overall benefits are primarily driven by the increased area allocated to intercropping (Supplementary Fig. 2), ranging from 9.4 million hectares for the Wheat-Faba bean association to 250 million hectares for Maize-Soybean association (Fig. 5). This is combined with near-unity pLER values for most cereals and lower, yet substantial, values for legumes. In detail, the lowest increase is observed for the Wheat-Faba bean combination with a LER of 1.19 (with pLER-wheat = 0.73 and pLER-faba bean = 0.46), and the highest for Maize-Soybean with a LER of 1.74 (with pLER-maize = 0.99 and pLER-soybean = 0.75) (Fig. 5).
Alleviating the constraints on full overlapping intercropping systems (TND > 0), combined with varying relative density values, resulted in a win-win scenario, leading to an overall increase in production (Supplementary Fig. 3). While the overall production increase remains, this growth is slightly lower for cereals in scenarios where practices prioritize the legumes (i.e., TND > 0 and higher relative density for legumes than for cereals) (Supplementary Fig. 3). This pattern is consistent with previous results of cereal competitive dominance with low TND. This highlights a flexibility that enables farmers to tailor intercropping configurations to local contexts, thereby supporting higher relative yields for legumes while still maintaining overall production increases.
Our global meta-analysis reveals that ecologically optimized intercropping offers substantial potential, boosting global cereal production by up to 51% on existing land and contributing to future food demands. However, association-specific partial Land Equivalent Ratios (pLERs)—0.46 to 0.99 across major combinations—highlight the context-dependent nature of yield benefits, necessitating a shift towards precision intercropping strategies. Indeed, strategically optimizing relative planting density (RD) and temporal niche differentiation (TND) consistently enhances productivity across these key associations under various constraints. This precision approach, informed by our quantitative framework linking functional traits and management to predict intercropping outcomes, provides a promising foundation for resilient and resource-efficient food systems crucial for global food security. While our analysis centered on yield, assessing economic profitability and ecosystem service contributions will be essential to fully evaluate the potential of intercropping systems in the future.
Methods
Data collection: Identification of field experiments
To compile a comprehensive set of primary studies, we first identified meta-analyses that assessed intercropping effects on yield. In January 2024, we conducted a comprehensive literature search to identify meta-analytical studies or databases that analyzed the impact of intercropping on yield. The search was performed using Web of Science with the following query: (((TS= (“intercrop*”OR “mixed crop” OR “crop mixture” OR “mixed cultivation” OR “cover crop*” OR “companion crop*”OR “mixed crop*” OR “relay crop*” OR “interplanting”)) AND TS = (“yield” OR “product”OR“harvest”)) AND TS = (“meta-analysis” OR “meta analysis” OR “global analysis”OR “systematic review”). In addition to this search, we included seven additional meta-analysis or databases identified through manual literature review. An initial screening was conducted to exclude narrative review and papers that did not specifically address the effect of intercropping on yield. To avoid redundancy, a secondary screening step was applied to exclude meta-analyses that re-analysed the same set of primary study; only the original meta-analysis was retained. Details of the data collection process are outlined in supplementary fig. 4.
From the selected meta-analyses, we identified key agronomic and environmental variables (referred to as moderators) reported as influencing yield outcomes in intercropping systems (supplementary Table 2). We assigned a score to each reference, assessing the presence (score = 0) or absence (score = 1) of moderating variables. The criteria for scoring and detailed description are provided in the supplementary files. Higher scores indicated absence in moderator data; we retained references with the scores below the median (see supplementary file). Finally, we checked the availability of each meta-analysis database; if a database was not freely available, the corresponding authors were contacted. If no response was received after one month and two follow-up reminders, the study was excluded. A total of four studies6,7,18,32 met all the inclusion criteria.
An Excel data file was compiled using the available database from the supplementary materials or repository provided by the authors, incorporating all primary data from this set of four meta-analyses. This dataset presents 3421 observations (i.e., rows) from 408 primary studies. We conserved information on all moderators or meta-data presented in the original databases. Inclusion criteria for primary studies within these meta-analyses were then applied based on the PICO-C framework: Population—annual cultivated plants (excluding perennials, except when grown as annuals); Intervention—intercropping with two plants; Comparator—monoculture; Outcome—yield comparison between monoculture and intercropping for at least one crop in the combination; and Context—all peer-reviewed scientific articles, regardless of year or language. In the end, 2503 observations from 334 primary studies were kept in our analysis.
Data collection: completion of the database for key agronomic and environmental variables
For each observation, the data from the original databases were thoroughly verified and corrected where necessary, leading to the correction of 117, 599 and 56 observations for nitrogen rate variable, density design and spatial arrangement design, respectively. Missing data for selected moderators was supplemented by referencing the primary studies. This process allowed for the completion of a portion of the data for the variables (see Supplementary Table 2). However, information remained unavailable for a percentage of the observation-moderator combinations. To address this, GPS coordinates were extracted (if missing) and used to retrieve the values of the moderators from publicly accessible databases. Soil pH and soil organic carbon at six different soil depths were obtained from the SoilGrids database at a resolution of 250 m. The value of soil organic matter was estimated by multiplying the soil organic carbon by a factor of 1.7233. The WorldClim database v 2.1 was used to derive cumulative rainfall and average temperature34,35. Unlike previous studies6,36,37, we did not rely on average annual rainfall or temperature values over the year but used the values over the crop cycle (i.e. the total growth time of the intercropped plants). To obtain these time-specific climate metrics, the sowing/planting and harvesting dates of the two intercropping plants were essential and were retrieved from the original articles when available. In cases where sowing or harvesting were not specified, estimates were made using global cultivation calendar30 or the FAO cultivation calendar (https://cropcalendar.apps.fao.org/#/home).
We also completed the database by linking the traits of the species for each observation using the Plant Trait Database (TRY) (https://www.try-db.org/TryWeb/Home.php)38. We then calculated the relative trait distances between continuous single traits of the two species in association, in particular plant height, leaf area and root depth, and multiple functional trait distances. Functional distance was calculated using the Gower distance (gowdis function from the FD R package), as it integrates both continuous traits (maximum height, maximum root depth and leaf area) and binary traits (C3 vs. C4 photosynthetic pathways and legume vs. non-legume) with equal weighting for all traits.
The relative distance of continuous single traits between the two species in association was calculated as follows:
With \({{Height}}_{a}\), \({{Leaf\; area}}_{a}\) and \({{Root\; depth}}_{a}\) respectively the maximum height, the leaf area and the maximum root depth for species a. A negative value indicates that the plant has a lower trait value (leaf area, height, or root depth) than the species it is associated with, and vice versa for a positive value.
The Temporal Niche Differentiation (TND), the Relative Sowing Time (RST) and the Relative Density (RD), were also calculated according to the formulas given in literature18,19 when they were missing:
With \({P}_{{overlap}}\) the overlapping growth period of the intercropped species and \({P}_{{system}}\) the total duration of the intercropped system. TND therefore varies between 0 and 1 (never reaching 1 in this case, as that would indicate double cropping). A TND of 0 means that both plants are sown and harvested at the same time.
With \({S}_{a}\) and \({S}_{b}\) the sowing times for species a and b respectively and \({H}_{a}\) the harvest time of species a. RST denotes the relative sowing time of species a in relation to species b, meaning that an RST of 0 indicates that both plants were sown at the same time, a negative RST means that species b was sown before species a, and vice versa.
With \({d}_{{ic},a}\) and \({d}_{{mc},a}\) the densities of species within the intercrop and the monocrop system, respectively. The density of a species is defined as the number of individuals of that species per unit area. It is generally provided by the authors or can be determined based on the row or plant arrangement39. The Total Relative Density is simply obtained by summing up the relative densities of the two intercropped species. When this value equals 1, it is referred to as a substitutive (or replacement) design; when it falls between 1 and 2, it is considered an augmentative design; and when it equals 2, it is classified as an additive design.
Finally, we verified the data using descriptive statistics (Supplementary Table 3). The distribution of each continuous variable was plotted to identify potential outliers. Any extreme value was either (i) corrected if it was an error, or (ii) retained if present as it was in the original article.
Data analysis: calculation of mean effect size
To evaluate the effect of intercropping on crop yields, the partial land equivalent ratio (relative yield) was used and calculated as follows:
Where \({Y}_{{ic}}\) and \({Y}_{{mc}}\) represent yield of a species i in intercropped and monocropped systems, respectively. Observations were excluded when the yield values for intercropping or monocropping were zero, indicating either crop failure or experimental error.
Data in meta-analyses are typically weighted using the inverse of their variance. However, this information is insufficiently reported in the dataset. Instead, the number of experimental replications was used to weight each observation:
Where \({Weight}\) is the weight associated with each observation, and \({N}_{t}\) and \({N}_{c}\) are the number of replicates for the intercropping and monocropping system treatments, respectively. If the number of replicates was not provided, it was assumed as 3.
Data analysis: identification of main moderators
To identify key moderators influencing intercropping yield, we employed a multi-step approach. First, we conducted a pairwise correlation analysis to mitigate potential multicollinearity issues. Moderators with correlation coefficients (Person’ r) below 0.7 were retained, while highly correlated pairs (r ≥ 0.7) were reduced to a single representative variable based on interpretability and agronomic relevance. We visualized the correlation matrix using the corrplot and ggplot2 packages in R (Supplementary Fig. 5). Second, we performed an exploratory search for relevant moderators (Supplementary Table 2) using the R package Metaforest, a machine-learning approach based on random forests40. A preselection step was applied using the preselect function, where a recursive algorithm with 10,000 iterations and 100 replications was used to assess variable importance. Variables with consistently negative importance were excluded, and those with positive importance were retained for further analysis. Next, we tunned the parameters of Metaforest using the train function from the caret package focusing on the minimum node size and the number of features selected for splitting nodes. The optimal model was selected based on the root mean square error (RMSE) from the 10-fold cross-validation resulting in a cross-validated R² of 0.35 and an out-of-bag (OOB) R² of 0.76. The final model had a minimum node size of 5, and a randomly selected number of features set at 3 for node splitting. Finally, the relative importance of the retained moderators was assessed using the varImp function.
From these analyses, we found that the intercropping effect was predominantly explained by the relative density (RD), the temporal niche differentiation (TND) and the relative height difference (H) over a broad range of diverse crop associations, pedo-climatic and agricultural management factors. Intercropping effect was thus modeled as follows:
Where \({\beta }_{0}\), \({\pi }_{{\rm{study}}}\), and \(\varepsilon\) are the mean effects, the random estimation error associated with the study, and sampling error, respectively. This linear mixed effect model (LMM) was fitted using the lme4 package. To address the hierarchical data structure, we tested multiple random effect configurations and ultimately retained the model with random effects due solely to study-level variability, selecting it based on the lowest Akaike Information Criterion (AIC)40 (Supplementary Table 4). As shown in Eq. 9, we did not include the full model containing all three key explanatory variables identified earlier and their interactions due to concerns about multicollinearity. Instead, we selected the model with the lowest Akaike Information Criterion (AIC), calculated using the dredge function (MuMIn package in R) for all possible combinations of explanatory variables (fixed effects) identified through the Random Forest analysis. To ensure the robustness of the final model (Eq. 9), multicollinearity was checked by calculating the variance inflation factor (VIF) using the vif function (car package in R), verifying that the maximum VIF did not exceed a threshold of 1041,42 (Supplementary Fig. 6).
Confidence intervals for the fitted coefficients of the final LMM (Eq. 9 and Supplementary Table 1) were calculated using a bootstrapping technique to ensure robust interval estimation and because the LMM violated the assumption of normality of model residuals. The procedure involved resampling the data with replacement and refitting the model for each of the resampled datasets. A total of 1000 bootstrap iterations were conducted using the bootMer function (lme4 package), resulting in 95% confidence intervals for each predictor. All data analyses were performed in R.4.3.2.
Practical application of the model for scaling up intercropping
The five most frequent species associations in our dataset were identified, all of which were cereal-legume associations. A full sampling of parameter combinations (TND, relative density of species a and b) was generated using the range of observed values by species within each association. To do this, we employed the chull function from the grDevices package and the point.in.polygon function from the sp package, which verifies whether a set of points falls within a given polygon.
The partial Land Equivalent Ratio (pLER) for each species was computed for each parameter combination using the previously established model (Eq. 9). The Pareto-optimal combinations of parameters (TND, relative density of species a and b) were then identified for each species based on these pLER. Among these optimal solutions, we selected the combination that yielded the smallest possible TND (TND = 0 or TND = 0.18 for the Wheat-Faba bean association) to align as closely as possible with conventional intercropping practices typically employed by farmers. We also selected the highest relative density for the cereal species to promote cereal production, which is generally the fastest-growing sector in the 2050 scenarios and is at the core of current food security policies.
For harvested area, yield, and production data at the current period, we extracted and averaged over a 10-year period (2013–2023) FAOSTAT data on each species.
For this optimal solution, new production levels were estimated under the assumption that the cropland area of one species in the association is maximized in intercropping (the legume here, due to its smaller area), while maintaining the actual total cultivated area. The cultivated area was adjusted to account for both monoculture and intercropped surfaces, with relative density values determining the land share (\({{LS}}_{a}\) or \({{LS}}_{b}\)), representing the proportion of the intercropped area occupied by each species. The area in intercrop (AIC) was calculated using the following relation:
Where \({A}_{{mc},a,{FAO}}\) represents 100% of the FAO-cultivated area, assuming that the intercropped area is maximized for legumes (i.e., total conversion of the current area). We assumed that the FAO data for the cultivated area is entirely in monoculture. The relationship \({{LS}}_{a}+{{LS}}_{b}=1\) holds, and the constraint \({A}_{{ic}}\le {A}_{{mc},a,{FAO}}+{A}_{{mc},b,{FAO}}\) is applied. Thus, we computed:
The yield in intercropping was recalculated using the calculated pLER values and monoculture yield data from FAOSTAT. The yield in intercropping \({Y}_{{ic},a}\) for species “a” was calculated as:
where \({Y}_{{mc},a,{FAO}}\) is the yield from FAOSTAT, \({{pLER}}_{a}\) is the pLER calculated using our model (Eq. 9), and the optimal parameters identified and selected previously. The total production \({P}_{a}\) was then computed as:
where \({Y}_{{ic},a}\) is the intercropping yield (Eq. 12), \({A}_{{ic}}\) is the total area in intercropping, and \({A}_{{mc},a}\) is the new monoculture area of species “a” (similarly calculated for species “b,” the cereal). Finally, to assess the gain in production due to intercropping, we calculated the percentage change for each species in each association relative to the baseline production data provided by FAOSTAT.
Data availability
All raw data used in this manuscript are publicly available in the cited references. Additionally, the data supporting the findings of this study are available in the supplementary files.
Code availability
The underlying code for this study is not publicly available but may be made available to qualified researchers on reasonable request from the corresponding author.
References
Mead, R. & Willey, R. W. The Concept of a ‘Land Equivalent Ratio’ and Advantages in Yields from Intercropping. Exp. Agric. 16, 217–228 (1980).
Li, C. et al. The productive performance of intercropping. Proc. Natl. Acad. Sci. USA 120, e2201886120 (2023).
Li, C. et al. Syndromes of production in intercropping impact yield gains. Nat. Plants 6, 653–660 (2020).
Huss, C. P., Holmes, K. D. & Blubaugh, C. K. Benefits and risks of intercropping for crop resilience and pest management. J. Econ. Entomol. 115, 1350–1362 (2022).
Brooker, R. W. et al. Improving intercropping: a synthesis of research in agronomy, plant physiology and ecology. N. Phytol. 206, 107–117 (2015).
Viaud, P. et al. Sugarcane yield response to legume intercropped: a meta-analysis. Field Crop. Res. 295, 108882 (2023).
Carrillo-Reche, J. et al. Finding guidelines for cabbage intercropping systems design as a first step in a meta-analysis relay for vegetables. Agric. Ecosyst. Environ. 354, 108564 (2023).
Hauggaard-Nielsen, H. & Jensen, E. S. Evaluating pea and barley cultivars for complementarity in intercropping at different levels of soil N availability. Field Crops Res. 72, 185–196 (2001).
Xu, Z. et al. Intercropping maize and soybean increases efficiency of land and fertilizer nitrogen use; a meta-analysis. Field Crop. Res. 246, 107661 (2020).
Feng, C. et al. Maize/peanut intercropping increases land productivity: a meta-analysis. Field Crop. Res. 270, 108208 (2021).
MacLaren, C. et al. Predicting intercrop competition, facilitation, and productivity from simple functional traits. Field Crops Res. 297, 108926 (2023).
Mahaut, L. et al. Beyond trait distances: functional distinctiveness captures the outcome of plant competition. Funct. Ecol. 37, 2399–2412 (2023).
Wang, Z. et al. Competition for light drives yield components in strip intercropping in the Netherlands. Field Crops Res. 320, 109647 (2025).
Mao, L. et al. Crop growth, light utilization and yield of relay intercropped cotton as affected by plant density and a plant growth regulator. Field Crops Res. 155, 67–76 (2014).
Ahmed, S. et al. Optimized planting time and co-growth duration reduce the yield difference between intercropped and sole soybean by enhancing soybean resilience toward size-asymmetric competition. Food Energy Security 9, e226 (2020).
Martin-Guay, M.-O., Paquette, A., Dupras, J. & Rivest, D. The new Green Revolution: Sustainable intensification of agriculture by intercropping. Sci. Total Environ. 615, 767–772 (2018).
Van der Werf, W. et al. Comparing performance of crop species mixtures and pure stands. Front. Agric. Sci. Eng. 8, 481–489 (2021).
Yu, Y., Stomph, T.-J., Makowski, D. & van der Werf, W. Temporal niche differentiation increases the land equivalent ratio of annual intercrops: A meta-analysis. Field Crops Res. 184, 133–144 (2015).
Yu, Y., Stomph, T.-J., Makowski, D., Zhang, L. & van der Werf, W. A meta-analysis of relative crop yields in cereal/legume mixtures suggests options for management. Field Crop. Res. 198, 269–279 (2016).
Wang, Q. et al. Does reduced intraspecific competition of the dominant species in intercrops allow for a higher population density?. Food Energy Security 10, e270 (2021).
Zhang, L. et al. Light interception and utilization in relay intercrops of wheat and cotton. Field Crops Res. 107, 29–42 (2008).
Yamane, K., Ikoma, A. & Iijima, M. Performance of double cropping and relay intercropping for black soybean production in small-scale farms. Plant Production Sci. (2016).
Schwinning, S. & Weiner, J. Mechanisms determining the degree of size asymmetry in competition among plants. Oecologia 113, 447–455 (1998).
Banik, P., Midya, A., Sarkar, B. K. & Ghose, S. S. Wheat and chickpea intercropping systems in an additive series experiment: advantages and weed smothering. Eur. J. Agron. 24, 325–332 (2006).
Yu, J., Rezaei, E. E., Reckling, M. & Nendel, C. Winter wheat–soybean relay intercropping in conjunction with a shift in sowing dates as a climate change adaptation and mitigation strategy for crop production in Germany. Field Crops Res. 322, 109695 (2025).
Lebreton, P. et al. Optimal species proportions, traits and sowing patterns for agroecological weed management in legume–cereal intercrops. Eur. J. Agron. 159, 127266 (2024).
Mao, L. et al. Yield advantage and water saving in maize/pea intercrop. Field Crops Res. 138, 11–20 (2012).
Sekiya, N. & Yano, K. Do pigeon pea and sesbania supply groundwater to intercropped maize through hydraulic lift?—Hydrogen stable isotope investigation of xylem waters. Field Crops Res. 86, 167–173 (2004).
Homulle, Z., George, T. S. & Karley, A. J. Root traits with team benefits: understanding belowground interactions in intercropping systems. Plant Soil 471, 1–26 (2022).
Jägermeyr, J. et al. Climate impacts on global agriculture emerge earlier in new generation of climate and crop models. Nat. Food 2, 873–885 (2021).
Roy, A., Moradkhani, H., Mekonnen, M., Moftakhari, H. & Magliocca, N. Towards strategic interventions for global food security in 2050. Sci. Total Environ. 954, 176811 (2024).
Paut, R., Garreau, L., Ollivier, G., Sabatier, R. & Tchamitchian, M. A global dataset of experimental intercropping and agroforestry studies in horticulture. Sci. Data 11, 5 (2024).
Pribyl, D. W. A critical review of the conventional SOC to SOM conversion factor. Geoderma 156, 75–83 (2010).
Fick, S. E. & Hijmans, R. J. WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315 (2017).
Harris, I., Osborn, T. J., Jones, P. & Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 7, 109 (2020).
Chen, G. et al. Potential crop yield gains under intensive soybean/maize intercropping in China. Plant Soil https://doi.org/10.1007/s11104-023-06423-7 (2023).
Muoni, T. et al. Effects of management practices on legume productivity in smallholder farming systems in sub-Saharan Africa. Food Energy Secur 11, e366 (2022).
Kattge, J. et al. TRY plant trait database–enhanced coverage and open access. Glob. Change Biol. 26, 119–188 (2020).
Li, C. et al. Yield gain, complementarity and competitive dominance in intercropping in China: A meta-analysis of drivers of yield gain using additive partitioning. Eur. J. Agron. 113, 125987 (2020).
Van Lissa, C. J. Small sample meta-analyses: Exploring heterogeneity using MetaForest. In Small Sample Size Solutions: A Guide for Applied Researchers and Practitioners (edsVan de Schoot, R. & Miočević, M.) (CRC Press, 2020). https://doi.org/10.4324/9780429273872-16.
Gareth, J., Witten, D., Hastie, T. & Tibshirani, R. An Introduction to Statistical Learning with Applications in R: By Gareth James, Daniela Witten, Trevor Hastie (New York, 2014).
Ramankutty, N., Evan, A. T., Monfreda, C. & Foley, J. A. Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Global Biogeochemical Cycles 22 (2008).
Acknowledgements
The authors would like to thank the Horizon Europe 2020 project and CIRAD (French Agricultural Research Centre for International Development) for their financial support. We would also like to express our sincere thanks to Patrice Dumas and Pierre-Marie Aubert for their constructive comments and enriching discussions during the preparation of this study.
Author information
Authors and Affiliations
Contributions
D.B., R.P. and M.R. conceived and designed the research. M.R. collected data, performed the statistical analysis and wrote the first draft of the manuscript. D.B., R.P. and M.R. wrote interactively through multiple rounds of revisions. R.P. supervised the work. All the authors reviewed and approved the final paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ruillé, M., Beillouin, D. & Prudhomme, R. Ecological drivers of intercropping performance for enhanced global crop production. npj Sustain. Agric. 4, 8 (2026). https://doi.org/10.1038/s44264-025-00110-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s44264-025-00110-z
