
Abstract
Although protected areas can lead to recovery of overharvested species, it is much less clear whether the return of certain predator species or a diversity of predator species can lead to re-establishment of important top-down forces that regulate whole ecosystems. Here we report that the algal recovery in a Mediterranean Marine Protected Area did not derive from the increase in the traditional strong predators, but rather from the establishment of a previously unknown interaction between the thermophilic fish Thalassoma pavo and the seastar Marthasterias glacialis. The interaction resulted in elevated predation rates on sea urchins responsible for algal overgrazing. Manipulative experiments and field observations revealed that the proximity of the seastars triggered an escape response in sea urchins, extending their tube feet. Fishes exploited this behavior by feeding on the exposed tube feet, thus impairing urchin movement and making them vulnerable to predation by the seastars. These findings suggest that predator diversity generated by MPA establishment can activate positive interactions among predators, with subsequent restoration of the ecosystem structure and function through cascading consumer impacts.
Similar content being viewed by others
Introduction
Marine communities are rapidly changing due to anthropogenic impacts. In this context, depletion of predators due to overfishing may lead to degradation of ecosystems through trophic cascades1,2,3,4. Recent global climate change is causing shifts in the biogeographical ranges of some species5, providing novel assortments of species with potentially new and unexpected effects on ecosystems6. New interactions among species and the strengthening of existing ones may lead to unexpected consequences for ecosystem function7,8.
Banning human activities in Marine Protected Areas (MPAs) usually promotes recovery of targeted predators, especially strong predators such as large fishes and lobsters9,10,11 which, indirectly, leads to ecosystem restoration through trophic cascades. Such effects are by no means certain to occur in all locations however12,13,14,15 and responses to protection from fishing may be variable in magnitude, direction and degree of stability16. Sometimes no recovery is seen in higher predators on timescales of years, suggesting that ecosystem functioning within MPAs can follow different paths11,14,17,18. The establishment of direct and indirect interactions among different predator species may provide an alternative mechanism for controlling prey populations19. The potential for new interactions to occur is likely promoted by the increase in diversity across taxa and trophic groups after protection enforcement10,20. In particular, facilitative interactions among different predators can enhance the predation rate on common species21,22,23,24,25,26,27,28. Through synergistic facilitation19,29, weak predators can then control prey populations and restore the ecosystem even in the absence of strong predators. This process may be favored by the arrival of new species, creating novel species assemblages, due to range shifts prompted by climate change30.
The control of sea urchin populations by key predators, prompting a regime shift from grazed “barrens” to kelp forest, has been particularly well studied in temperate rocky reef MPAs31,32,33,34,35,36. In the Mediterranean Sea, recovery of sea bream populations (genus Diplodus) promotes algal forest growth by predation on the sea urchins Paracentrotus lividus and Arbacia lixula17,36. However, in some cases spatial protection alone is not sufficient to restore Diplodus populations37,38. This is the case in the “Ustica Island” MPA, where protection was enforced in 1991. Unlike other Mediterranean MPAs17,36, at Ustica the upper infralittoral zone developed as a barren following protection enforcement39. Until 2007, the sea urchins P. lividus and A. lixula and encrusting corallines such as Lithophyllum spp., Pseudolithophyllum expansum, Lithothamnium spp. and Mesophyllum coralloides dominated the shallow rocky bottom assemblages40. In recent years, sea urchins abundance decreased, the forest restored41, yet the population of Diplodus remained unexpectedly low14, following a steady long-term decline38. Sea urchin depletion was instead accompanied by an exceptional increase in the density of the seastar Marthasterias glacialis in the sea urchin-barren areas14 and by an emergent, facultative synergistic interaction between the seastar and the thermophilic fish Thalassoma pavo, herein described.
The wrasse T. pavo is known to consume small molluscs, crustaceans and juvenile sea urchins42. Although this small fish is unable to overcome the defences of adult sea urchins43, the congeneric T. noronhanum is reported to occasionally tear off parts of the sea urchin tube feet44. At the “Ustica Island” MPA, T. pavo is abundant on shallow reefs45, since it is favoured by the warm water and the steep shores of the island46,47,48,49 and possibly also by protection. The seastar M. glacialis is usually rare on Mediterranean shallow reefs, being more common in deeper reefs and has so far been overlooked as a predator of sea urchins due to its preference for bivalves42. Sea urchins are able to perceive seastars from a distance of up to 50 cm and successfully flee, hence reducing the success of attacks50,51. The predatory strength of M. glacialis on sea urchins is thus usually weak, but in Ustica MPA, M. glacialis is particularly abundant in the barren areas and is an important predator of sea urchins14,52.
Field observations showed that, in the proximity of M. glacialis, sea urchins escape their potential predator by extending their tube feet, lowering their spines and fleeing away from the seastar. At this point, several individuals of T. pavo usually gather and repeatedly bit off urchin tube feet. Mutilated sea urchins are then impaired in their movement capacity and become vulnerable to seastar predation.
We hypothesize that, after MPA implementation, an emergent, synergistic facilitative interaction between the seastar and the fish makes sea urchins vulnerable to seastar predation, thus contributing to population control of sea urchins at “Ustica Island” MPA allowing for consequent recovery of algal forests.
Results
Long-term data showed a clear trend with densities of both P. lividus and A. lixula enormously increasing after protection enforcement (Fig. 1). This trend reversed from 2000 to 2010 (Fig. 1). The decrease of sea urchins was accompanied by an increase in the population of M. glacialis and an extension of the algal coverage. The ecosystem state eventually shifted from barren to Cystoseira spp. forest (Fig. 1).

Conceptual model of trophic interactions and their effect on habitat state in Mediterranean rocky reefs.
(A) Interactions among the seastar M. glacialis, the ornate wrasse T. pavo, the sea urchins P. lividus and A. lixula and the algae Cystoseira spp. Dotted arrows indicate indirect interactions, whereas solid arrows indicate direct interactions. +/− symbols indicate positive or negative effect of the interaction on the species pointed by the arrow. In Ustica Island the interaction between fish and seastar (1) facilitates predation on sea urchins (2) by the seastar. Urchins are in turn strong grazers and are able to alter algal coverage (3). (B) Temporal trends in M. glacialis densities, sea urchin densities and habitat state in Ustica Island MPA. Red line: P. lividus. Black line: A. lixula. Blue bars: M. glacialis. Green line: algal coverage. Images in figure by F. Di Trapani and P. Gianguzza.
The success of the fish-seastar interaction was in part reflected by the dominance of sea urchins in the diet of large seastars (>27 cm total length). P. lividus accounted for 51.3% of captured prey observed and A. lixula for 28.2% (Supplementary Table 1). Similarly, tube feet of sea urchins were frequently found in large quantities (mean ± SE = 16.5 ± 4.8) in the stomachs of T. pavo (33% of fishes), indicating that the interaction is common throughout the MPA. Through direct observation and video recording, we established that the wrasse removed 39–88% of tube feet from P. lividus and 58–84% of tube feet from A. lixula during seastar attacks (Supplementary Fig. 1).
The 3-way ANOVA analysis showed a significant effect of the term “Treatment X Habitat” (Table 1). SNK a posteriori tests were run to investigate the differences found (Table 1, Fig. 2). In the forest, M. glacialis was always faster than P. lividus and A. lixula. We observed that the seastar was able to “float” above the forest thanks to its long arms, while urchin movement was impeded by their spines tangling among the thalli of erect algae.

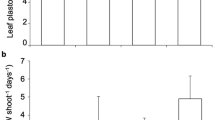
Attack speed (M. glacialis) and escape speed (P. lividus and A. lixula) measured in intact individuals (normal), control and treated individuals (with 50% and 70% tube feet removed) in forest (green bars) and barren (empty bars).
A 3-way ANOVA was performed and significant effect of the term “Treatment X Habitat” was found (Table 1). SNK a posteriori tests were run to investigate the differences found (Table 1); similar letters (a, b, c, d) indicate groups of data not significantly different. In forest, M. glacialis was faster than P. lividus and A. lixula (normal, intact, control and treated). In barren, M. glacialis was faster than P. lividus and A. lixula when 70% and 50% tube feet, respectively, were removed (Table 1). We observed that M. glacialis is able to “float” above the forest thanks to its long arms, while urchin movement was affected by their spines tangling among the thalli of erect algae (Table 1).
In the barrens habitat, the speed of sea urchins was negatively affected only when at least 70% and 50% of tube feet were removed from P. lividus and A. lixula, respectively (Table 1; Fig. 2).
The removal of tube feet slowed the movement speed of sea urchins, allowing M. glacialis – which otherwise moved more slowly than the urchins in barrens habitat – to overtake and capture them (Fig. 3).

Simulation of the effect of tube feet removal on escape velocity and predation outcome on the barren habitat based on experimental data (Table 1, Fig. 2).
V = Mean velocity (±s.e.). of attacking M. glacialis and escaping sea urchins (A. lixula and P. lividus). Removal of 70% and 50% of tube feet respectively in P. lividus and A. lixula allows the predator M. glacialis to reach and consume them. Simulated reaction distance of sea urchins is 10 cm (images in figure by F. Di Trapani).
Discussion
The hypothesis that a synergistic facilitative interaction between the seastar M. glacialis and the fish T. pavo makes sea urchins vulnerable to seastar predation was corroborated by the results of this study. Sea urchin resulted the preferred food item of M. glacialis while sea urchin tube feet were frequently found in the T. pavo gut contents.
The experimental removal of sea urchin tube feet was quantitatively similar to the action of wrasses during the escape behaviour triggered by the seastar. Our results suggested that T. pavo facilitated M. glacialis in the barren habitat by removing tube feet and reducing escape speeds of sea urchins allowing higher rates of capture by M. glacialis. At the same time, T. pavo seems to have benefited from the escape response of sea urchins triggered by the seastar, since extended tube feet became accessible to the fish. However, in our study we lacked the data to demonstrate whether the absence of seastars affects the ability of T. pavo to capture sea urchin tube feet.
The establishment of a facilitative interaction between T. pavo and M. glacialis is particularly interesting since these species usually occupy non-overlapping habitats and neither of them generally feed on adult of sea urchins42,43. The simultaneous occurrence of sea urchins, M. glacialis and T. pavo in the shallow rocky shore of Ustica, where high densities of the seastar occurred14, likely created the condition for the described interaction to happen.
Species of the Thalassoma genus display different feeding strategies. This fish can use stones as anvils to crash bivalve that are too large to swallow; it acts as facultative cleaner-fish; it follows “nuclear fish ” eating particles stirred up from the bottom44; and it follows divers to obtain food53. Importantly, wrasses are able to remember what, where and when food can be found54. Data from gut contents of T. pavo at Ustica clearly showed that sea urchin tube feet are an important component of its diet. Hence, T. pavo may have “learned” to follow the attacks of M. glacialis on sea urchins in order to easily feed on the sea urchin tube feet. This is of particular interest since T. pavo is a thermophilic species and its Mediterranean range has moved progressively northward, following increases in water temperature55. It is possible that interactions between T. pavo and seastars or other weak sea urchin predators will arise in other regions, with important consequences on sea urchin populations and benthic community structure.
The correlation between the decrease in sea urchin abundance, the increase in seastar population and the recovery in algae forest (Fig. 1) indicates that the high predation on sea urchins by M. glacialis, facilitated by T. pavo, is a key process for the recovery of the algae forest at Ustica island14,40.
The establishment of MPAs is a widespread tool for marine managers to fight the detrimental effect of multiple stressors, such as loss of species and climate change, on coastal systems worldwide11. Several studies indicate a positive effect of protection on targeted species within MPAs and, in some cases, the recovery of former ecosystems (e.g., kelp forest) through trophic cascades33,35,36. In other cases, however, the response of communities to protection seems to be idiosyncratic10. The variability in the effects of protection could depend on the time scale considered, as both direct and indirect effects of protection are slow, unstable and asynchronous16. Moreover, even if the goal of MPAs is the protection of the whole assemblages of organisms and their interactions, monitoring programs often focus only on targeted and conspicuous species, or limited assemblages (e.g., fish assemblages). The potential role of other species and the unique assemblages that may result due, in part, to their establishment, on the dynamics of protected ecosystems is thus largely overlooked.
Predator diversity may enhance the strength of trophic cascades by providing alternative consumers when environmental conditions change (i.e., Insurance Hypothesis)56,57. However, many systems are characterized by key predators and weak alternative consumers, whose effect on prey populations is mild58,59. In other cases, multi-predator assemblages may alter the strength of trophic control on preys by the so-called emergent multi-predator effect. Different predators can establish synergistic interactions, either positive (i.e., facilitation) or negative (i.e., interference), affecting predation rate on common prey19,59.
Here we presented a case where the paucity of traditional key predators is compensated by an unexpected interaction between two weak predators (a seastar and a fish), leading to the ecosystem restoration (Fig. 4). More broadly, our study represents an example where predator assemblage diversity strengthened the trophic control of keystone grazers, allowing the recovery of the algal forest habitat (i.e. the three-dimensional component).

Conceptual model of the trophic cascades without the presence of MPAs, in their presence and in the Ustica case study.
In areas where fishing is allowed, low densities of Diplodus causes the outbreak of sea urchin and consequently barren grounds are common. MPAs with their no-take policy are instead characterized by high densities of Diplodus that control urchins and allow algal canopy to return. The peculiar case of the Ustica MPA consists instead of low densities of Diplodus but, at the same time, extended algal coverage and low densities of sea urchins. In this case the seastar M. glacialis interacts with the ornate wrasse T. pavo and thus is able to heavily and successfully prey upon urchins species P. lividus and A. lixula and control their populations (images in figure by C. Bonaviri, F. Di Trapani and P. Gianguzza).
Our results confirm the importance of long term studies for proper assessment of the direct and indirect effects of protection on ecosystems. This study is also a reminder of the importance of field observation for revealing arising interactions among novel species assortments in a changing community.
Methods
Study area
“Ustica Island” MPA is located on a small volcanic island 80 km off the northern coast of Sicily, covering 16,000 ha, of which 65 are devoted to a no-take area along the northwestern part of the island. Like other volcanic islands, Ustica is characterized by steep cliffs, which favor the abundance of the thermophilic wrasse T. pavo within its distribution range48. The density of T. pavo is high at Ustica with estimates ranging from 48.6 mean individuals/250 m2 (Galasso unpublished data) to 140.33 mean individuals/250 m2 60. In the framework of scientific monitoring projects on the benthic megafaunal assemblages (2003-2010), an interaction between T. pavo and M. glacialis feeding on sea urchins was repeatedly observed. Ustica also has an unusually high abundance of the seastar M. glacialis with 1.2 mean individuals/250 m2 unlike other Mediterranean areas14 ranging from 0 to 0.1 mean individuals/250 m2 14.
Long-term monitoring of algae and sea urchins
Erect macroalgae cover (i.e., filamentous, foliose, corticated foliose, corticated macrophytes, leathery macrophytes and articulated calcareous) was visually estimated by either stereomicroscope analysis of slides taken by an analogue camera at high resolution, or computer analysis of images captured by a digital camera. A grid of 24 small quadrats was superimposed over each image and the abundance of each taxon assessed by giving a score from 0 to 4 to each subquadrat61. The total density of sea urchins (>1 cm test diameter) was estimated by visual counts within 1 m2 quadrats39. We analyzed 40 images for erect macroalgae cover and 40 replicates of each species of sea urchin per year.
Analysis of T. pavo gut content
In order to evaluate the frequency of tube feet ingestion by fishes, we collected 40 T. pavo individuals between June and July 2008 and analyzed their gut contents in the laboratory. Total fish length was measured to the nearest millimeter and stomachs were removed and fixed in 5% buffered formalin for gut content analysis. For those stomachs containing food, we calculated the percentage frequency of those containing tube feet and then the number of tube feet were quantified under a binocular microscope62. Fish size ranged from 8 to 12.5 cm (10.5 ± 1.3 cm, mean ± s.d.). Only four individuals had an empty stomach.
Prey of M. glacialis
M. glacialis, like many other seastar species, consumes its prey by extruding its stomach to envelop prey and digesting the soft tissues. We estimated M. glacialis prey frequencies in the field by lifting individuals, turning them upside-down and identifying the prey item to the lowest possible taxonomic level63. We conducted observations in 50 × 5 m random transects at ca. ≈5 m depth. Each individual showing feeding activity was measured using a Vernier caliper (maximum tip-to-tip diameter or total length, TL). We observed 241 M. glacialis, of which 118 were feeding upon 11 different prey items (Supplementary Table 1). Surveys were performed from June 2008 to June 2010. Data were grouped into 9 size classes: I (16–17 cm), II (17.1–20 cm), III (20.1–22 cm), IV (22.1–25 cm), V (25.1–27 cm), VI (27.1–29 cm), VII (29.1–32 cm), VIII (32.1–34 cm) and IX (>34 cm TL). To evaluate the effect of seastar size on its diet, we performed cluster analysis on a Bray Curtis similarity resemblance matrix of the arcsine-transformed data (Primer E Software v.6 64). Cluster analysis showed a clear division of seastars into two groups, larger and smaller than 27 cm TL(Supplementary Table 1), depending on their diet.
Field observation of tube feet removal by T. pavo
The number of urchin tube feet removed by T. pavo during the attacks by M. glacialis was estimated in the field by video analysis of predation experiments. For these experiments we placed seastars in the field where sea urchins were naturally present and abundant. Seastar individuals had been previously starved for at least two weeks. We used two different methods to video-record the attacks. In one case, three video cameras (JVC EVERIO GZ-MG330 with 30 GB internal hard-drive, equipped with a modified waterproof case connected to long-life battery packs) were secured through tripods on the sea floor, for remote recording. Alternatively, cameras were directly operated by scuba divers. The number of urchin tube feet removed by T. pavo during each seastar attack on a sea urchin was subsequently evaluated by computer video analysis. The duration of the attacks ranged from a number of seconds to a few minutes and terminated when the seastar reached the sea urchin or spontaneously stopped. Experiments were run in summer 2009.
Effect of habitat on attack speed of M. glacialis and on escape speed of treated and untreated sea urchins
To test whether the removal of tube feet effectively compromised the sea urchin movement, we simulated the effect of the wrasse bite experimentally, by removing tube feet from individuals collected in the field. We then measured the attack speed of M. glacialis and the escape speed of both intact and impaired sea urchins (P. lividus and A. lixula) in the field. The tested habitats were barren ground, where the four species are abundant and algal forest. In order to test the effect of the habitat on the speed of sea urchins and M. glacialis and the effect of tube feet removal on sea urchins speeds we manipulated the number of sea urchins tube feet in laboratory. Individuals of both P. lividus (n = 48) and A. lixula (n = 48) were placed in a glass tank in the laboratory. When the sea urchins were properly attached and adhered to the substratum, we removed the tube feet by rapidly pulling the urchins off the tanks. The detached tube feet remained on the wall of the tank and were easily counted. Using the equation of Santos & Flammang65 for estimating the total number of tube feet present in an urchin and by repeating the operation described above, we proceeded with the removal of 50% and 70% of tube feet for both P. lividus and A. lixula. Manipulated urchins were then used in the field to run the tests. To check for differences between treated and non-treated urchins due to handling or transportation stress, we also ran control tests on non-manipulated sea urchins. We performed a 3-way ANOVA analysis considering three factors66. Treatment, fixed with 8 levels: untreated M. glacialis, untreated P. lividus, untreated A. lixula, P. lividus with 50% of tube feet removed, A. lixula with 50% of tube feet removed, P. lividus with 70% of tube feet removed, control for P. lividus and control for A. lixula; Habitat, fixed with 2 levels: forest and barren; Site, with 2 levels, random and nested in Habitat. We ran 6 replicates for each experimental block. The sizes of the experimental animals were as follows: 4.28 ± 0.4 cm (mean ± s.d.) for P. lividus, 4.36 ± 5 cm (mean ± s.d.) for A. lixula (mean ± s.d.) and 32.2 ± 8 (mean ± s.d.) cm for M. glacialis. Experiments were run in summer 2009.
Ethic statement
For sea urchin and seastar collection and manipulation, no permits were required according to national laws (Italian Legislative decree n. 116/1992). All experimental protocols were approved by the Department of Hearth and Marine Science (DiSTeM) of the University of Palermo. This research was conducted under a research permit from MPA local authority (0041110-26/05/2010 – Harbour Office of Palermo, MPA management body of the Ustica Island).
Additional Information
How to cite this article: Galasso, N. M. et al. Fish-seastar facilitation leads to algal forest restoration on protected rocky reefs. Sci. Rep. 5, 12409; doi: 10.1038/srep12409 (2015).
References
Jackson, J. B. C. et al. Historical overfishing and the recent collapse of coastal ecosystems. Science 293, 629–638, 10.1126/science.1059199 (2001).
Scheffer, M., Carpenter, S., Foley, J. A., Folke, C. & Walker, B. Catastrophic shifts in ecosystems. Nature 413, 591–596, 10.1038/35098000 (2001).
Ling, S. D., Johnson, C. R., Frusher, S. D. & Ridgway, K. R. Overfishing reduces resilience of kelp beds to climate-driven catastrophic phase shift. Proceedings of the National Academy of Sciences of the United States of America 106, 22341–22345, 10.1073/pnas.0907529106 (2009).
Estes, J. A. et al. Trophic Downgrading of Planet Earth. Science 333, 301–306, 10.1126/science.1205106 (2011).
Harley, C. D. G. Climate Change, Keystone Predation and Biodiversity Loss. Science 334, 1124–1127, 10.1126/science.1210199 (2011).
Nogues-Bravo, D. & Rahbek, C. Communities Under Climate Change. Science 334, 1070–1071, 10.1126/science.1214833 (2011).
Walther, G. R. Community and ecosystem responses to recent climate change. Philosophical Transactions of the Royal Society B-Biological Sciences 365, 2019–2024, 10.1098/rstb.2010.0021 (2010).
Cardinale, B. J., Palmer, M. A. & Collins, S. L. Species diversity enhances ecosystem functioning through interspecific facilitation. Nature 415, 426–429, 10.1038/415426a (2002).
Micheli, F., Halpern, B. S., Botsford, L. W. & Warner, R. R. Trajectories and correlates of community change in no-take marine reserves. Ecological Applications 14, 1709–1723, 10.1890/03-5260 (2004).
Lester, S. E. et al. Biological effects within no-take marine reserves: a global synthesis. Marine Ecology Progress Series 384, 33–46, 10.3354/meps08029 (2009).
Edgar, G. J. et al. Global conservation outcomes depend on marine protected areas with five key features. Nature 506, 216-+, 10.1038/nature13022 (2014).
Micheli, F. et al. Cascading human impacts, marine protected areas and the structure of Mediterranean reef assemblages. Ecological Monographs 75, 81–102, 10.1890/03-4058 (2005).
Shears, N. T., Babcock, R. C. & Salomon, A. K. Context-dependent effects of fishing: variation in trophic cascades across environmental gradients. Ecological Applications 18, 1860–1873, 10.1890/07-1776.1 (2008).
Bonaviri, C. et al. Fish versus starfish predation in controlling sea urchin populations in Mediterranean rocky shores. Marine Ecology Progress Series 382, 129–138, 10.3354/meps07976 (2009).
Eklof, J. S. et al. How effective are MPAs? Predation control and ‘spill-in effects’ in seagrass-coral reef lagoons under contrasting fishery management. Marine Ecology Progress Series 384, 83–96, 10.3354/meps08024 (2009).
Babcock, R. C. et al. Decadal trends in marine reserves reveal differential rates of change in direct and indirect effects. Proceedings of the National Academy of Sciences of the United States of America 107, 18256–18261, 10.1073/pnas.0908012107 (2010).
Sala, E., Boudouresque, C. F. & Harmelin-Vivien, M. Fishing, trophic cascades and the structure of algal assemblages: evaluation of an old but untested paradigm. Oikos 82, 425–439, 10.2307/3546364 (1998).
Gilby, B. L. & Stevens, T. Meta-analysis indicayes habitat-specific alterations to primary producer and herbivore communities in marina protected areas. Global Ecology and Conservation 2, 289–299 (2014).
Bruno, J. F. & Cardinale, B. J. Cascading effects of predator richness. Frontiers in Ecology and the Environment 6, 539–546, 10.1890/070136 (2008).
Clemente, S., Hernandez, J. C., Rodriguez, A. & Brito, A. Identifying keystone predators and the importance of preserving functional diversity in sublittoral rocky-bottom areas. Marine Ecology Progress Series 413, 55–67, 10.3354/meps08700 (2010).
Soluk, D. A. & Collins, N. C. Synergistic interactions between fish and stoneflies facilitation and interference among stream predators. Oikos 52, 94–100, 10.2307/3565987 (1988).
de Goeij, P., Luttikhuizen, P. C., van der Meer, J. & Piersma, T. Facilitation on an intertidal mudflat: the effect of siphon nipping by flatfish on burying depth of the bivalve Macoma balthica. Oecologia 126, 500–506, 10.1007/s004420000526 (2001).
Mouritsen, K. N. Intertidal facilitation and indirect effects: causes and consequences of crawling in the New Zealand cockle. Marine Ecology Progress Series 271, 207–220, 10.3354/meps271207 (2004).
Meyer, J. J. & Byers, J. E. As good as dead? Sublethal predation facilitates lethal predation on an intertidal clam. Ecology Letters 8, 160–166, 10.1111/j.1461-0248.2004.00700.x (2005).
Bshary, R., Hohner, A., Ait-El-Djoudi, K. & Fricke, H. Interspecific communicative and coordinated hunting between groupers and giant moray eels in the Red Sea. Plos Biology 4, 2393–2398, 10.1371/journal.pbio.0040431 (2006).
Steinmetz, J., Soluk, D. A. & Kohler, S. L. Facilitation between herons and smallmouth bass foraging on common prey. Environmental Biology of Fishes 81, 51–61, 10.1007/s10641-006-9165-6 (2008).
Fodrie, F. J., Kenworthy, M. D. & Powers, S. P. Unintended facilitation between marine consumers generates enhanced mortality for their shared prey. Ecology 89, 3268–3274, 10.1890/07-1679.1 (2008).
Goyert, H. F., Manne, L. L. & Veit, R. R. Facilitative interactions among the pelagic community of temperate migratory terns, tunas and dolphins. Oikos 123, 1400–1408, 10.1111/oik.00814 (2014).
Sih, A., Englund, G. & Wooster, D. Emergent impacts of multiple predators on prey. Trends in Ecology & Evolution 13, 350–355, 10.1016/s0169-5347(98)01437-2 (1998).
Kordas, R. L., Harley, C. D. G. & O’Connor, M. I. Community ecology in a warming world: The influence of temperature on interspecific interactions in marine systems. Journal of Experimental Marine Biology and Ecology 400, 218–226, 10.1016/j.jembe.2011.02.029 (2011).
Pinnegar, J. K. et al. Trophic cascades in benthic marine ecosystems: lessons for fisheries and protected-area management. Environmental Conservation 27, 179–200, 10.1017/s0376892900000205 (2000).
Shears, N. T. & Babcock, R. C. Marine reserves demonstrate top-down control of community structure on temperate reefs. Oecologia 132, 131–142, 10.1007/s00442-002-0920-x (2002).
Shears, N. T. & Babcock, R. C. Continuing trophic cascade effects after 25 years of no-take marine reserve protection. Marine Ecology Progress Series 246, 1–16, 10.3354/meps246001 (2003).
Sonnenholzner, J. I., Ladah, L. B. & Lafferty, K. D. Cascading effects of fishing on Galapagos rocky reef communities: reanalysis using corrected data. Marine Ecology Progress Series 375, 209–218, 10.3354/meps07890 (2009).
Ling, S. D. & Johnson, C. R. Marine reserves reduce risk of climate-driven phase shift by reinstating size- and habitat-specific trophic interactions. Ecological Applications 22, 1232–1245 (2012).
Guidetti, P. Marine reserves reestablish lost predatory interactions and cause community changes in rocky reefs. Ecological Applications 16, 963–976, 10.1890/1051-0761(2006)016[0963:mrrlpi]2.0.co;2 (2006).
Sala, E. et al. The Structure of Mediterranean Rocky Reef Ecosystems across Environmental and Human Gradients and Conservation Implications. Plos One 7, 13, 10.1371/journal.pone.0032742 (2012).
Mirasole, A., Gianguzza, P., Badalamenti, F. & Bonaviri, C. Dinamica dell’ecosistema roccioso dell’AMP “Isola di Ustica” nell’ultimo cinquantennio attraverso la conoscenza dei veterani del mare in Ecology for a sustainable blue and green growth. (https://congressosite2013.files.wordpress.com/2013/09/site_abstract-book_definitivo_stampa.pdf.) (Ancona 2013).
Gianguzza, P. et al. The effects of recreational Paracentrotus lividus fishing on distribution patterns of sea urchins at Ustica Island MPA (Western Mediterranean, Italy). Fisheries Research 81, 37–44, 10.1016/j.fishres.2006.06.002 (2006).
Bonaviri, C., Fernandez, T. V., Fanelli, G., Badalamenti, F. & Gianguzza, P. Leading role of the sea urchin Arbacia lixula in maintaining the barren state in southwestern Mediterranean. Marine Biology 158, 2505–2513, 10.1007/s00227-011-1751-2 (2011).
Gianguzza, P. et al. Macroalgal assemblage type affects predation pressure on sea urchins by altering adhesion strength. Marine Environmental Research 70, 82–86, 10.1016/j.marenvres.2010.03.006 (2010).
Guidetti, P. Consumers of sea urchins, Paracentrotus lividus and Arbacia lixula, in shallow Mediterranean rocky reefs. Helgoland Marine Research 58, 110–116, 10.1007/s10152-004-0176-4 (2004).
Sala, E. Fish predators and scavengers of the sea urchin Paracentrotus lividus in protected areas of the north-west Mediterranean Sea. Marine Biology 129, 531–539, 10.1007/s002270050194 (1997).
Sazima, C., Bonaldo, R. M., Krajewski, J. P. & Sazima, I. The Noronha wrasse: a “jack-of-all-trades” follower. Aqua (Miradolo Terme) 9, 97–108 (2005).
Milazzo, M., Badalamenti, F., Fernandez, T. V. & Chemello, R. Effects of fish feeding by snorkellers on the density and size distribution of fishes in a Mediterranean marine protected area. Marine Biology 146, 1213–1222, 10.1007/s00227-004-1527-z (2005).
Guidetti, P. & Sala, E. Community-wide effects of marine reserves in the Mediterranean Sea. Marine Ecology Progress Series 335, 43–56, 10.3354/meps335043 (2007).
Ojeda-Martinez, C., Bayle-Sempere, J. T., Sanchez-Jerez, P., Forcada, A. & Valle, C. Detecting conservation benefits in spatially protected fish populations with meta-analysis of long-term monitoring data. Marine Biology 151, 1153–1161, 10.1007/s00227-006-0557-0 (2007).
Milazzo, M. et al. Vertical distribution of two sympatric labrid fishes in the Western Mediterranean and Eastern Atlantic rocky subtidal: local shore topography does matter. Marine Ecology-an Evolutionary Perspective 32, 521–531, 10.1111/j.1439-0485.2011.00447.x (2011).
Milazzo, M., Mirto, S., Domenici, P. & Gristina, M. Climate change exacerbates interspecific interactions in sympatric coastal fishes. Journal of Animal Ecology 82, 468–477, 10.1111/j.1365-2656.2012.02034.x (2013).
Urriago, J. D., Himmelman, J. H. & Gaymer, C. F. Responses of the black sea urchin Tetrapygus niger to its sea-star predators Heliaster helianthus and Meyenaster gelatinosus under field conditions. Journal of Experimental Marine Biology and Ecology 399, 17–24, 10.1016/j.jembe.2011.01.004 (2011).
Manzur, T. & Navarrete, S. A. Scales of detection and escape of the sea urchin Tetrapygus niger in interactions with the predatory sun star Heliaster helianthus. Journal of Experimental Marine Biology and Ecology 407, 302–308, 10.1016/j.jembe.2011.06.025 (2011).
Gianguzza, P., Bonaviri, C. & Guidetti, P. Crushing predation of the spiny star Marthasterias glacialis upon the sea urchin Paracentrotus lividus. Marine Biology 156, 1083–1086, 10.1007/s00227-009-1153-x (2009).
Pasko, L. Tool-Like Behavior in the Sixbar Wrasse, Thalassoma hardwicke (Bennett, 1830). Zoo Biology 29, 767–773, 10.1002/zoo.20307 (2010).
Salwiczek, L. H. & Bshary, R. Cleaner Wrasses Keep Track of the ‘When’ and ‘What’ in a Foraging Task. Ethology 117, 939–948, 10.1111/j.1439-0310.2011.01959.x (2011).
Lejeusne, C., Chevaldonne, P., Pergent-Martini, C., Boudouresque, C. F. & Perez, T. Climate change effects on a miniature ocean: the highly diverse, highly impacted Mediterranean Sea. Trends in Ecology & Evolution 25, 250–260 (2010).
Loreau, M. et al. Ecology - Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 294, 804–808, 10.1126/science.1064088 (2001).
Griffin, J. N. & Silliman, B. R. Predator diversity stabilizes and strengthens trophic control of a keystone grazer. Biology Letters 7, 79–82, 10.1098/rsbl.2010.0626 (2011).
Navarrete, S. A. & Menge, B. A. Keystone predation and interaction strength: Interactive effects of predators on their main prey. Ecological Monographs 66, 409–429, 10.2307/2963488 (1996).
Guidetti, P. Predator diversity and density affect levels of predation upon strongly interactive species in temperate rocky reefs. Oecologia 154, 513–520, 10.1007/s00442-007-0845-5 (2007).
Vacchi, M., Bussotti, S., Guidetti, P. & La Mesa, G. Study of the coastal fish assemblage in the marine reserve of the Ustica Island (southern Tyrrhenian Sea). Italian Journal of Zoology 65, 281–286 (1998).
Dethier, M. N., Graham, E. S., Cohen, S. & Tear, L. M. Visual versus random-point percent cover estimations - objective is not always better. Marine Ecology Progress Series 96, 93–100, 10.3354/meps096093 (1993).
Hyslop, E. J. Stomach contents analysis a review of methods and their application. Journal of Fish Biology 17, 411–430, 10.1111/j.1095-8649.1980.tb02775.x (1980).
Tuya, F. & Duarte, P. Role of food availability in the bathymetric distribution of the starfish Marthasterias glacialis (Lamk.) on reefs of northern Portugal. Scientia Marina 76, 9–15, 10.3989/scimar.2012.76n1009 (2012).
Clarke, K. R. & Gorley, R. N. (Plymouth, UK, 2006).
Santos, R. & Flammang, P. Intra- and interspecific variation of attachment strength in sea urchins. Marine Ecology Progress Series 332, 129–142, 10.3354/meps332129 (2007).
Underwood, A. J. Experiments in Ecology: Their Logical Design and Interpreation Using Analysis of Variance. (Cambridge University Press, 1997).
Acknowledgements
Dedicated to Diego and Lorenzo. The authors wish to thank Giorgio Aglieri, Manfredi Di Lorenzo, Giovanni Fanelli, Giacomo Milisenda, Alice Mirasole, Simona Noè, Claudia Scianna and Tomas Vega Fernández for their assistance in the field experiments and data collection. This study was funded by the MPA “Isola di Ustica” and the Italian Ministry of the Environment and Protection of the Territory and the Sea (MATTM). CB was supported by the Italian project FIRB (contract no.RBFR12RXW).
Author information
Authors and Affiliations
Contributions
F.B. conceived the project; C.B. and N.M.G. wrote the paper; D.A., C.B., F.B., F.D.T., N.M.G., P.G. and M.P. collected the data; D.A., C.B., F.B., F.D.T., N.M.G., P.G. and M.P. contributed to analyses and interpretation.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Galasso, N., Bonaviri, C., Trapani, F. et al. Fish-seastar facilitation leads to algal forest restoration on protected rocky reefs. Sci Rep 5, 12409 (2015). https://doi.org/10.1038/srep12409
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep12409
This article is cited by
-
Alterations in sea urchin (Mesocentrotus nudus) microbiota and their potential contributions to host according to barren severity
npj Biofilms and Microbiomes (2023)
-
Monitoring marine fishes using underwater video techniques in the Mediterranean Sea
Reviews in Fish Biology and Fisheries (2023)
-
Unveiling the diet of the thermophilic starfish Ophidiaster ophidianus (Echinodermata: Asteroidea) combining visual observation and stable isotopes analysis
Marine Biology (2020)
-
Biodiversity loss and turnover in alternative states in the Mediterranean Sea: a case study on meiofauna
Scientific Reports (2016)
-
Size-dependent predation of the mesopredator Marthasterias glacialis (L.) (Asteroidea)
Marine Biology (2016)
