
Abstract
Meiotic homologous recombination plays a central role in creating genetic variability, making it an essential biological process relevant to evolution and crop breeding. In this study, we used pollen-specific fluorescent tagged lines (FTLs) to measure male meiotic recombination frequency during the development of Arabidopsis thaliana. Interestingly, a subset of pollen grains consistently shows loss of fluorescence expression in tested lines. Using nine independent FTL intervals, the spatio-temporal dynamics of male recombination frequency was assessed during plant development, considering both shoot type and plant age as independent parameters. In most genomic intervals assayed, male meiotic recombination frequency is highly consistent during plant development, showing no significant change between different shoot types and during plant aging. However, in some genomic regions, such as I1a and I5a, a small but significant effect of either developmental position or plant age were observed, indicating that the meiotic CO frequency in those intervals varies during plant development. Furthermore, from an overall view of all nine genomic intervals assayed, both primary and tertiary shoots show a similar dynamics of increasing recombination frequency during development, while secondary and lateral shoots remain highly stable. Our results provide new insights in the dynamics of male meiotic recombination frequency during plant development.
Similar content being viewed by others
Introduction
During plant sexual reproduction, parental genomes reshuffle through a specialized type of cell division, e.g. meiosis, which reduces the somatic chromosome number to yield haploid cells and produces gametes with varying combinations of parental genetic information. A key event in the first meiotic cell division is the formation of crossovers (COs) between homologous parental chromosomes, which is tightly regulated by a series of consecutive events; including homolog recognition, pairing, synapsis, and recombination1. Meiotic CO formation is not only necessary for creating new allele combinations and increasing genetic diversity, but also is essential to ensure accurate chromosome segregation during meiosis I. Each CO acts as a tether between a pair of homologs, ensuring proper alignment and balanced segregation of resulting bivalents at metaphase I and anaphase I, respectively, by regulating correct attachment of corresponding kinetochores to the MI spindle2,3,4,5. Due to its central role in creating genetic variability, meiotic homologous recombination is considered crucial for plant evolution and speciation, and hence also constitutes an essential biological process for the development of novel crop varieties.
Experimentally, several methods exist for assessing patterns and rates of meiotic recombination. In general, they can be classified into two major categories: (1) cytological analysis of meiocytes using microscopy and (2) population-wide segregation analysis of genetic markers6,7,8,9. As an alternative method, Francis et al.10 developed a series of pollen-specific fluorescent markers (FTLs; fluorescent tagged lines) in the model plant Arabidopsis thaliana, that upon combination in a hemizygous state, allow for high-throughput quantification of male meiotic recombination in corresponding genomic intervals11,12. The FTLs contain linked transgenes (T-DNA inserts) that encode either DsRed, eYFP and eCFP in mature pollen grains through activation by the post-meiotic pollen-specific LAT52 promoter10,12,13. Moreover, since FTLs are designed to contain two or three different linked fluorescent markers in a hemizygous state, the segregation pattern of fluorescent markers in resulting pollen grains also allows for the detection and quantification of CO interference and events of gene conversion11. Pollen-based analysis of male meiotic recombination using FTLs is generally performed using one of two following methods; namely (1) tetrad-based analysis using epi-fluorescence microscopy10,11,14 and (2) single-pollen analysis using flow cytometry12. For tetrad-based analysis, the methodology takes the advantage of the Arabidopsis quartet1-2 (qrt1-2) mutant (Figure S1), in which the four pollen grains remain physically attached to each other (tetrad) throughout the whole process of male gametogenesis due to lack of the pectin methylesterase enzyme15. Alternatively, FTL-based quantification of meiotic recombination frequency can also be performed based on the fluorescent analysis of single pollen grains by flow cytometry (Figure S1). Using this methodology, data acquisition speed can be increased up to 150-fold compared to tetrad analysis, allowing rapid quantification of male meiotic CO rates12. Both methods have their own specific advantages and disadvantages for meiotic recombination analysis. Tetrad-based analysis enables the detection of unviable or ectopically non-fluorescent pollen grains and thus allows for the elimination of related biases in meiotic CO analysis. At the downside, this method is highly laborious and time-consuming, and may suffer from detection errors due to manual counting or interpretation. In contrast, single pollen grain analysis using flow cytometry allows high-throughput quantification of meiotic CO rates and excludes the possibility of human errors. Moreover, this method generates far more data points, improving statistical reliability. As a major drawback, flow cytometry-based FTL analysis does not allow for detection of alterations in pollen viability and related changes in fluorescence intensity, putatively leading to non-registered biases in recombination frequency measurement. Currently, there are 113 single FTL markers available, including 35 DsRed, 41 eYFP and 37 eCFP fluorescent markers, which are randomly spread across the five Arabidopsis chromosomes11. By combining these fluorescent markers in double or triple ‘linked’ configurations, this extensive set of FTLs provides a flexible way to assay the male meiotic recombination rate in different intervals of the Arabidopsis genome.
In Arabidopsis, meiotic recombination frequency varies during plant development16. Pollen tetrad analysis in the genomic intervals I1a and I3a revealed that crossover frequency in the second and third branches are significantly higher than that of the main shoot, which indicates that developmental position can affect crossover frequencies during male gametogenesis10. In addition, a whole-genome level study of meiotic CO frequency in Arabidopsis thaliana revealed that aging affects CO frequency during megaspore formation (increased 29.86%), but not during pollen formation17. By analyzing the data of main and lateral shoots, it was indicated that this age effect correlates to the developmental age of each shoot rather than that of the whole plant17. In Arabidopsis, flowers develop on different shoot types, such as primary shoot, secondary shoot, tertiary shoot and lateral shoot (Fig. 1), in which meiotic recombination occurs independently. Up till now, only for interval I1a and I3a, the developmental position was shown to have a significant effect on the meiotic CO frequency10. However, in general, there is no clear insight into whether there exists variability in the male meiotic CO frequency during plant aging and shoot development. In this study, we use nine independent FTLs intervals (I1a, I1b, I2a, I2b, I3c, CEN3, I5a, I5c and I5d) to measure the spatio-temporal dynamics of male CO frequency in flowers isolated from primary, secondary, tertiary and lateral shoots, in order to provide an extensive overview of the spatio-temporal dynamics of male meiotic recombination frequency in the genomic intervals delineated by a set of widely used pollen FTL markers.

Primary shoot is the main shoot of plant branching into secondary shoots which in turn branch into tertiary shoots. The lateral shoot develops from the axils of rosette leaves.
Results
Fluorescent Tagged Lines show partial loss of pollen-specific fluorescence
FTL-based quantification of meiotic recombination frequency in Arabidopsis relies on the segregation analysis of two or three linked hemizygous fluorescent markers (DsRed, eYFP and eCFP) in mature pollen grains. Due to their transgenic nature, these recombinant transgene encoding proteins may be prone to epigenetic silencing, reducing the accuracy of associated meiotic CO read-out. In order to assess putative losses or biases in pollen fluorescence of the widely used FTL marker combinations (including I1a, I1b, I2a, I2b, I3c, CEN3, I5a, I5c and I5d), we monitored DsRed, eYFP and eCFP expression in tetrad configured pollen grains isolated from quartet1-2−/− (qrt1-2−/−) plants containing homozygous FTL marker combinations.
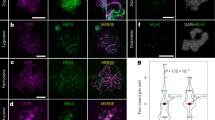
Theoretically, in the homozygous FTL tetrad-pollen, all four pollen grains enclosed should express the corresponding fluorescent markers. However, in all lines analyzed, we observed a combination of fluorescent and non-fluorescent pollen grains, suggesting that some pollen grains were either dead or suffered from a suppressed expression of the reporter construct. In our study, pollen grains are classified ‘dead’ when all fluorescent markers enclosed fail to express, irrespective whether the pollen is shriveled up or fully developed. In contrary, pollen showing specific loss of one or two fluorescent reporter(s), whereas the remaining other(s) are still expressed, are classified as ‘loss of fluorescence’. Based on these definitions, we quantified the marker-specific loss of pollen fluorescence and found that all FTL lines examined show a certain level of fluorescence ‘silencing’ in corresponding DsRed, eYFP and eCFP markers (Fig. 2, Table 1 and S1). Although the level of fluorescence ‘silencing’ in most FTL markers are generally low (ranging from 0.09% to 3.77%), some FTL markers show a more extreme loss of fluorescence. For example, the I1a line shows an extremely high level of fluorescent loss in DsRed marker (19.0% ± 3.68%).

(A) A tetrad-pollen show full expression of DsRed, eYFP, and eCFP fluorescence; (B) A tetrad-pollen show fluorescent loss of eCFP and weak DsRed expression; (C) A tetrad-pollen exhibit fluorescent loss of DsRed and weak eYFP expression; (D) A tetrad-pollen show weak DsRed expression in one pollen grain, and fluorescent loss of eYFP and eCFP in another pollen grain. The white circles indicate the pollen grains which lost fluorescence. Scale bar also indicated in 50 μm.
Male meiotic CO rate dynamics varies between genomic regions during plant development
We measured male meiotic recombination frequency at nine different genomic intervals in different shoot types at different time points during Arabidopsis plant development (Figure S2 and Table S2). Since the nine intervals comprise different sized genomic regions, we used ‘CO rate per standard unit DNA sequence’ (cM/Mb) to calculate the recombination frequency respective to the genomic size. To examine whether plant age, independent of shoot type, influences male meiotic recombination frequency, we compared the average recombination frequency of all shoots present at the same time during four subsequent weeks (Fig. 3). The results show that the genetic distance in four of the nine genomic intervals significantly varies according plant age [I1a (ANOVA: F = 6.5510, p = 0.0151), I1b (ANOVA: F = 19.8920, p = 0.0005), I3c (ANOVA: F = 28.1260, p = 0.0001) and I5a (ANOVA: F = 70.7510, p = 0.0001), p value was calculated with post-hoc Tukey HSD test (α = 0.05)]. There show a gradual increase of recombination rate during flowering in I1b and I3c interval, but is not that clear in the I1a and I5a. The other five intervals do not display any significant variation in male meiotic recombination frequency during plant aging. This indicates that plant age selectively affects male meiotic recombination frequency only in specific genomic regions, and that this age effect is rather minor.

The ‘number of weeks, in the legend refers to the number of week after flowering induction. Statistical significance was analyzed based on one-way ANOVA [I1a (F = 6.5510, p = 0.0151), I1b (F = 19.8920, p = 0.0005), I2a (F = 1.3280, p = 0.3314), I2b (F = 1.8500, p = 0.2164), I3c (F = 28.1260, p = 0.0001), CEN3 (F = 2.5310, p = 0.1306), I5a (F = 70.7510, p = 0.0001), I5c (F = 0.6430, p = 0.6088) and I5d (F = 1.7630, p = 0.2317), p value was calculated with post-hoc Tukey HSD test (α = 0.05)].
Male meiotic CO rate in most genomic regions are not influenced by shoot type
To assess the effect of shoot type on the male meiotic recombination rate in different genomic regions, the mean CO rate during the entire development of each shoot type was assessed in nine different genomic intervals (Fig. 4). Comparative analysis revealed that three intervals exhibit a significant difference in CO rate between the four shoot types [I1a (ANOVA: F = 12.8200, p = 0.0020), I2a (ANOVA: F = 5.5220, p = 0.0238) and I5a (ANOVA: F = 17.6830, p = 0.0007), p value was calculated with post-hoc Tukey HSD test (α = 0.05)]. In the genomic interval I1a, for example, the male meiotic recombination frequency in the tertiary shoot is significantly higher than that of other shoot types. These results indicate that the developmental position (shoot type) affects male meiotic recombination frequency in specific genomic regions during Arabidopsis male sporogenesis. However, this is not common to all genomic regions since the other six intervals tested do not show any significant difference in the male meiotic CO rate between the different shoot types.

Statistical significance was analyzed based on one-way ANOVA [I1a (F = 12.8200, p = 0.0020), I1b (F = 1.9040, p = 0.2074), I2a (F = 5.5220, p = 0.0238), I2b (F = 0.4580, p = 0.7193), I3c (F = 1.2450, p = 0.3561), CEN3 (F = 1.3560, p = 0.3236), I5a (F = 17.6830, p = 0.0007), I5c (F = 0.9230, p = 0.4725) and I5d (F = 0.7060, p = 0.5746), p value was calculated with post-hoc Tukey HSD test (α = 0.05)].
Male meiotic CO rate increase in primary and tertiary shoots during plant development
In order to have an extensive overview on the spatio-temporal dynamics of male meiotic recombination frequency in all nine genomic intervals, we next compared the mean CO rate (cMall/Mball) of all nine intervals between four different shoot types (primary shoot, secondary shoot, tertiary shoot and lateral shoot, Fig. 1) at four time points during plant development (Fig. 5). For both primary and tertiary shoots, the averaged CO frequency significantly increased over time [ANOVA: F = 6.7630, p = 0.0001, p value was calculated with post-hoc Tukey HSD test (α = 0.05)]. In the tertiary shoot, this CO increase amounted up to approximately 32% from the third to the fourth week upon flowering induction. In contrast, the averaged CO rate remained highly stable during plant aging in both secondary and lateral shoots, showing a constant rate around 2.5 cM/Mb. Strikingly, at the third week following flower induction, tertiary shoot show a significant lower recombination frequency compared to that of other three shoot types. Moreover, a similar decrease in CO rate was also observed in the primary shoot at the first week following flower induction. As such, these data reveal that both primary and tertiary shoots show similar dynamics in recombination frequency during shoot development, with young shoot displaying a generally lower recombination frequency that gradually increase during plant aging. In contrast, the overall male meiotic recombination frequency in secondary and lateral shoots remain highly stable during the whole period of flowering.

The mean recombination frequency calculated as cMall/Mball. The individual of each interval is shown in Figure S2. Statistical significance was analyzed based on one-way ANOVA [F = 6.7630, p = 0.0001, p value was calculated with post-hoc Tukey HSD test (α = 0.05)].
Variation in age-dependent recombination rate dynamics in different genomic intervals is not correlated with their distance from the centromere
Our analyses reveal variation in the age-dependent CO frequency dynamics in the different genomic intervals tested, with some intervals showing an increase in CO rate during plant aging whereas other intervals displaying a rather constant CO rate over time. This variation suggests that a regulatory relationship between chromosome structure or genomic location and CO stability control in Arabidopsis. To explore this possibility, we next assessed the correlation between the meiotic recombination frequency change (ΔRF, ‘+’: increase/‘−’: decrease) of each shoot type for all intervals with the corresponding distance (Δ Distance) of their genomic location to the centromere (Table 2). Although in most shoot types there was a tendency for a positive relationship between the level of recombination frequency variation (ΔRF) and genomic distance of location to centromere (Δ Distance), the regression analysis revealed no correlation. Therefore, other regulatory factors may be involved in the determination of CO rate variability over time.
Discussion
The pollen FTL system provides a fast and powerful tool to measure meiotic recombination frequency in Arabidopsis male sporogenesis12. As this system relies on the segregation analysis of hemizygous fluorescent markers in mature pollen grains, it is important that the fluorescent markers of each interval are correctly expressed. Considering their transgenic nature, we found that all FTL lines examined show a certain level of fluorescent loss in the corresponding DsRed, eYFP or eCFP fluorescent markers. Although the loss of fluorescent expression was generally low in most FTL intervals (around 1–4%), some intervals exhibited a much larger proportion of pollen that lost fluorescence. For example, the DsRed marker enclosed in the I1a interval displays a fluorescent loss rate of approximately 20%. This is potentially due to the location of the DsRed reporter near the centromeric region, putatively affecting the expression of transgenic reporter through epigenetic silencing.
Importantly, the inadvertent loss of fluorescence in each FTL interval may generate biases in the deduced recombination frequency, since this is calculated based on the quantification of pollen containing two or more fluorescent reporters10. The problem of fluorescence loss and associated biases in CO calculation can be bypassed by deploying the FTL system in the quartet1-2 (qrt1-2) mutant background, which allows to ignore tetrads containing non-fluorescent microspores, thereby effectively eliminating the erroneous segregation patterns from the analysis. By and large, we found that single-pollen analysis via flow cytometry is however most suitable for measuring recombination frequency for most FTL lines investigated, which allows high throughput fluorescent pollen analysis and generates much more data points to achieve statistically reliable analysis.
The frequency of meiotic recombination is strictly controlled at the molecular level18,19,20. In addition, recent studies have revealed that it is also influenced by both endogenous and external factors, including heterozygosity21,22,23, sex determination7,17,24 and ambient temperature10,25,26. Here, we show that the plant age and developmental position (shoot type) also influence the frequency of male meiotic recombination. More specifically, our results demonstrated that the recombination frequency significantly varies by plant age in four of the nine genomic intervals tested, with the other five intervals showing a similar trend of increased CO frequency during plant aging. Moreover, in line with the previous study using Arabidopsis FTL lines10, we found that interval I1a shows a significant increase in meiotic CO frequency in higher order branches. Strikingly, in six of the nine intervals tested, this positive correlation between branch order and male meiotic CO frequency was not observed, indicating that different genomic regions may show variable CO rate dynamics during shoot development. Furthermore, from the whole nine intervals view, both the primary and tertiary shoots show similar increasing dynamics of recombination frequency during shoot development. While in contrast, both secondary and lateral shoots remain highly stable during the whole period of flowering. All together, these findings suggest that there may exist inherent structural or regulatory differences (in cis or in trans) between different genomic regions that influence the dynamics of corresponding male meiotic recombination frequency during shoot branching and development. However, up till now, it is unclear which genomic or chromosomal features may underlie this region-specific variability in CO dynamics and stability stringency.
To determine genome-wide meiotic recombination frequencies in Arabidopsis, multiple regions in each of the five chromosomes need to be monitored. The FTL system, in this case, is an efficient method compared with other conventional methods, which are generally based on the segregation analysis of polymorphic molecular markers in a pool of individual progeny plants that result from a backcrossing of a hybrid to one of the corresponding parental lines. These genetic methods usually do not generate sufficiently large data sets to allow accurate determination of recombination frequency. Furthermore, thorough investigation of interactions between heterozygosity and meiotic recombination revealed that Arabidopsis F1 hybrids, resulting from the intercross between a Colombia-0 line and other 32 different accessions, show a significant variation in the male meiotic CO rate compared to their homozygote counterparts27. On the genomic level, it has been demonstrated that heterozygous regions show an increase meiotic CO rate when juxtaposed to homozygous regions27. All together, these findings demonstrate that genetic maps (i.e. representing the population-wide meiotic CO pattern) strongly depend on the genetic make-up and heterozygosity level of the plant(s) used for analysis. In contrast, the FTL system for male meiotic recombination measurement can be implemented in a fully homozygous background, and thus fully eliminates adverse effects of allelic diversity and heterozygosity.
In brief, our results provide new insights into the dynamics of male meiotic recombination frequency across different orders of inflorescence shoot branches. The results re-appraise the use of the FTL system, and provide a valuable data set that will be a benchmark for analyzing male meiotic recombination frequency in various genotypes or obtained under varying environmental conditions.
Methods
Plant material and growth conditions
Arabidopsis thaliana wild type accessions (Colombia-0 background) and qrt1-2 (Colombia-3 background) were obtained from the Nottingham Arabidopsis Stock Center (NASC). All FTL lines were provided by G. P. Copenhaver upon request, all containing homozygous fluorescent markers in the qrt1-2−/− background. For male meiotic recombination frequency analysis, hemizygous fluorescent marker lines were created by intercrossing homozygous FTL marker lines (either two or three fluorescent reporters linked on the same chromosome) with either Col-0 or qrt1-2−/− diploid plants for either single pollen analysis or tetrad-based pollen analysis, respectively.
Arabidopsis seeds were vernalized at 4 °C for 2 days and germinated on K1 medium in vitro for 8 days at 20 °C under a 12 h/12 h (light/dark) photoperiod regime. Then, germinated seedlings were transferred to soil and cultivated in a controlled climate chamber under the same growth conditions (20 °C, 12 h/12 h light/dark). In order to stimulate flowering induction, photoperiod was changed into 16 h/8 h (light/dark) under the same conditions at the beginning of flower initiation.
Pollen isolated from flowers located at different shoot types were analyzed by microscopy or flow cytometry during plant development. For each shoot type, at least 200 tetrads were scored by epi-fluorescent microscopy or a minimum number of 2,000 single pollen grains was used for flow cytometry-based analysis. All analyses were performed in triplicate; i.e. each analysis included three biological replicates with pollen isolated from plants grown at the same time and under the same growth conditions.
FTL genomic intervals
Nine independent FTL intervals (I1a, I1b, I2a, I2b, I3c, CEN3, I5a, I5c, and I5d) were used in this study to monitor male meiotic recombination frequency in Arabidopsis thaliana. These nine intervals span a series of genomic regions that range from 0.59 to 5.40 megabases (Mb) and that are randomly distributed along four of the five Arabidopsis chromosomes. All together, they cover 15.89% of the total genome (19.81 Mb), and they are located in either sub-telomeric, interstitial or centromeric regions (Fig. 6 and Table 3).

The genomic location of the transgenic reporters encoding red (DsRed), yellow (eYFP), and cyan (eCFP) fluorescent proteins are indicated by filled circles on the five Arabidopsis chromosomes (green bars). The FTLs covering nine different genetic intervals (I1a, I1b, I2a, I2b, I3c, CEN 3, I5a, I5c and I5d) used in this study are delineated with brackets. The Arabidopsis Information Resource (TAIR) ‘chromosome map tool’ was used to scale the chromosomes and place the transgene insertion points on the physical map.
Microscopy
Bright field and fluorescence microscopy and image capturing was performed using an Olympus IX81 inverted fluorescence microscope equipped with an X-Cite Series 120Q UV lamp and an XM10 camera (http://www.olympus.com/). Image processing and Z-stack projections were conducted using Olympus OlyVIA 2.4 software (http://www.olympus-sis.com).
Flow cytometry
Single pollen analysis was performed using the BD FACSVerse flow cytometer equipped with a 488-nm and a 633-nm laser and eYFP and DsRed fluorescence was detected using 527/32-nm and 586/42-nm band-pass filters, respectively, based on the high-throughput fluorescent pollen analysis protocol of Yelina et al.12. The data were analyzed by BD FACSuite software and Microsoft Excel 2013.
Recombination frequency (RF) quantification
Calculation of the Meiotic recombination frequency in each interval based on tetrad or single pollen grain analysis were performed by the following formulas:

Where ‘TT’ is the number of tetratype, ‘NPD’ is the number of non-parental ditype and ‘Total’ is the number of all counted tetrads. The letter codes (a-l) for classification of tetrads used in this research are the same as those used in Copenhaver’s tetrad analysis, with FTL markers combined in a red–yellow–cyan configuration11.

Where ‘G’ is the number of green-alone fluorescent pollen, ‘R’ is the number of red-alone fluorescent pollen and ‘GR’ is the number of pollen showing both green and red fluorescence.
Data analysis
Data analysis and statistics were performed by Microsoft Excel 2013 and Data Processing System (DPS 7.05)28. One-way ANOVA with post-hoc Tukey HSD test was used for mean comparison of either two or multiple samples at the 5% significance level (α = 0.05).
Additional Information
How to cite this article: Li, F. et al. Dynamics of male meiotic recombination frequency during plant development using Fluorescent Tagged Lines in Arabidopsis thaliana. Sci. Rep. 7, 42535; doi: 10.1038/srep42535 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Lu, P., Wijeratne, A. J., Wang, Z., Copenhaver, G. P. & Ma, H. Arabidopsis PTD is required for type I crossover formation and affects recombination frequency in two different chromosomal regions. J Genet Genomics 41, 165–175, doi: 10.1016/j.jgg.2014.02.001 (2014).
Ma, H. Molecular genetic analyses of microsporogenesis and microgametogenesis in flowering plants. Annu. Rev. Plant Biol. 56, 393–434 (2005).
Cnudde, F. & Gerats, T. Meiosis: inducing variation by reduction. Plant Biol (Stuttg) 7, 321–341, doi: 10.1055/s-2005-865655 (2005).
De Storme, N. & Geelen, D. The Arabidopsis mutant jason produces unreduced first division restitution male gametes through a parallel/fused spindle mechanism in meiosis II. Plant physiology 155, 1403–1415 (2011).
Zamariola, L., Tiang, C. L., De Storme, N., Pawlowski, W. & Geelen, D. Chromosome segregation in plant meiosis. Frontiers in plant science 5 (2014).
López, E. et al. Looking for natural variation in chiasma frequency in Arabidopsis thaliana . Journal of experimental botany err319 (2011).
Giraut, L. et al. Genome-wide crossover distribution in Arabidopsis thaliana meiosis reveals sex-specific patterns along chromosomes. PLoS Genet 7, e1002354, doi: 10.1371/journal.pgen.1002354 (2011).
Salomé, P. et al. The recombination landscape in Arabidopsis thaliana F2 populations. Heredity 108, 447–455 (2012).
Ferdous, M. et al. Inter-homolog crossing-over and synapsis in Arabidopsis meiosis are dependent on the chromosome axis protein AtASY3. PLoS Genet 8, e1002507 (2012).
Francis, K. E. et al. Pollen tetrad-based visual assay for meiotic recombination in Arabidopsis . Proc Natl Acad Sci USA 104, 3913–3918, doi: 10.1073/pnas.0608936104 (2007).
Berchowitz, L. E. & Copenhaver, G. P. Fluorescent Arabidopsis tetrads: a visual assay for quickly developing large crossover and crossover interference data sets. Nat Protoc 3, 41–50, doi: 10.1038/nprot.2007.491 (2008).
Yelina, N. E. et al. High-throughput analysis of meiotic crossover frequency and interference via flow cytometry of fluorescent pollen in Arabidopsis thaliana . Nat Protoc 8, 2119–2134, doi: 10.1038/nprot.2013.131 (2013).
Twell, D., Yamaguchi, J. & McCORMICK, S. Pollen-specific gene expression in transgenic plants: coordinate regulation of two different tomato gene promoters during microsporogenesis. Development 109, 705–713 (1990).
Sun, Y. et al. Deep genome-wide measurement of meiotic gene conversion using tetrad analysis in Arabidopsis thaliana . PLoS Genet 8, e1002968, doi: 10.1371/journal.pgen.1002968 (2012).
Francis, K. E., Lam, S. Y. & Copenhaver, G. P. Separation of Arabidopsis pollen tetrads is regulated by QUARTET1, a pectin methylesterase gene. Plant Physiol 142, 1004–1013, doi: 10.1104/pp.106.085274 (2006).
Sanchez-Moran, E., Armstrong, S., Santos, J., Franklin, F. & Jones, G. Variation in chiasma frequency among eight accessions of Arabidopsis thaliana . Genetics 162, 1415–1422 (2002).
Toyota, M., Matsuda, K., Kakutani, T., Terao Morita, M. & Tasaka, M. Developmental changes in crossover frequency in Arabidopsis . Plant J 65, 589–599, doi: 10.1111/j.1365-313X.2010.04440.x (2011).
Crismani, W. et al. FANCM limits meiotic crossovers. Science 336, 1588–1590, doi: 10.1126/science.1220381 (2012).
Bauer, E. et al. Intraspecific variation of recombination rate in maize. Genome biology 14, 1 (2013).
Campos, J. L., Halligan, D. L., Haddrill, P. R. & Charlesworth, B. The relation between recombination rate and patterns of molecular evolution and variation in Drosophila melanogaster. Molecular biology and evolution 31, 1010–1028 (2014).
Jeffreys, A. J. & Neumann, R. Factors influencing recombination frequency and distribution in a human meiotic crossover hotspot. Human molecular genetics 14, 2277–2287 (2005).
Baudat, F. & De Massy, B. Cis-and trans-acting elements regulate the mouse Psmb9 meiotic recombination hotspot. PLoS Genet 3, e100 (2007).
Cole, F., Keeney, S. & Jasin, M. Comprehensive, fine-scale dissection of homologous recombination outcomes at a hot spot in mouse meiosis. Molecular cell 39, 700–710 (2010).
Drouaud, J. et al. Sex-specific crossover distributions and variations in interference level along Arabidopsis thaliana chromosome 4. PLoS Genet 3, e106, doi: 10.1371/journal.pgen.0030106 (2007).
Modliszewski, J. L. & Copenhaver, G. P. Meiotic recombination heats up. New Phytologist 208, 295–297 (2015).
Phillips, D. et al. The effect of temperature on the male and female recombination landscape of barley. New Phytol 208, 421–429, doi: 10.1111/nph.13548 (2015).
Ziolkowski, P. A. et al. Juxtaposition of heterozygous and homozygous regions causes reciprocal crossover remodelling via interference during Arabidopsis meiosis. Elife 4, e03708 (2015).
Tang, Q. Y. & Zhang, C. X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Science 20, 254–260 (2013).
Acknowledgements
This research was supported by the China Scholarship Council (CSC) to Fan Li, and the Fund for Scientific Research (FWO-Flanders, FWO16/PDOH1/049) to Nico De Storme. We gratefully acknowledge and appreciate Gregory. P. Copenhaver for providing FTL lines. We wish to thank the editor and reviewers for insightful comments.
Author information
Authors and Affiliations
Contributions
Fan Li completed the experiments, performed data analyses and wrote manuscript. Danny Geelen and Nico De Storme conceptualized the project, provided overall supervision and guidance, and set up FTL analysis platforms. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Li, F., De Storme, N. & Geelen, D. Dynamics of male meiotic recombination frequency during plant development using Fluorescent Tagged Lines in Arabidopsis thaliana. Sci Rep 7, 42535 (2017). https://doi.org/10.1038/srep42535
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep42535
