
Abstract
The human helicase senataxin (SETX) has been linked to the neurodegenerative diseases amyotrophic lateral sclerosis (ALS4) and ataxia with oculomotor apraxia (AOA2). Here we identified a role for SETX in controlling the antiviral response. Cells that had undergone depletion of SETX and SETX-deficient cells derived from patients with AOA2 had higher expression of antiviral mediators in response to infection than did wild-type cells. Mechanistically, we propose a model whereby SETX attenuates the activity of RNA polymerase II (RNAPII) at genes stimulated after a virus is sensed and thus controls the magnitude of the host response to pathogens and the biogenesis of various RNA viruses (e.g., influenza A virus and West Nile virus). Our data indicate a potentially causal link among inborn errors in SETX, susceptibility to infection and the development of neurologic disorders.
Similar content being viewed by others

Main
The innate immune response to infection represents the first line of cellular defense against invading pathogens. This response is orchestrated by a complex cohort of genes that are specifically induced upon infection1. Despite the protective roles of their products, prolonged or aberrant expression of such genes can be deleterious to the host2,3,4. Indeed, dysregulated expression of many such pro- and anti-inflammatory mediators can contribute to deaths caused by septic shock2 and by pandemic infection with influenza virus3 and has been linked to the etiology of several life-threatening diseases4. The expression of antiviral mediators is thus strictly regulated, with subsets of their genes expressed in distinct temporal patterns during infection5.
A classic example of the temporal regulation of infection-induced gene expression is the induction of genes encoding type I interferons and interferon-stimulated genes (ISGs) following viral infection. Upon entry of the virus into the cell, dedicated cytosolic receptors (including RIG-I, LGP2 and Mda5) recognize virus-derived nucleic acids and initiate a cascade of events that ultimately lead to activation of the transcription factor IRF3 ('interferon-regulatory factor 3') and its translocation to the nucleus6. Activated IRF3 is then recruited to the promoters of early-response genes, including IFNB1 (which encodes interferon-β (IFN-β)), and induces their transcription6. IFN-β subsequently induces the expression of ISGs via a second, interferon-dependent wave of transcription that relies on signaling via the kinase Jak and STAT transcription factors5. Notably, beyond IFN-β, the early interferon-independent wave of transcription also allows the concomitant expression of other transcription factors (for example, IRF7) and antiviral mediators (for example, IFIT1 and RIG-I), which establishes a feed-forward loop that probably potentiates the antiviral response during the course of infection. Light has been shed on the cellular signaling pathways that culminate in the early induction of genes encoding antiviral molecules, but detailed mechanistic insight into how chromatin-regulatory factors control this response is lacking7.
Stimulus-induced transcription is a tightly regulated, multi-step process that is controlled by a plethora of protein complexes and through multiple mechanisms acting at the level of initiation, pause-release, elongation and termination by RNA polymerase II (RNAPII)8. Studies have highlighted important factors and rate-limiting steps9,10 of these processes that operate on chromatin, which functions as a signal-integration platform during the cell's response to various stimuli11. For example, the initiation of transcription and progression to the elongation and termination phases of transcription12 are heavily influenced by dynamic changes to post-translational modifications on histone amino-terminal domains and the carboxy-terminal domain of RNAPII (ref. 13). The recognition of these chemical marks by dedicated proteins controls transcriptional activity14, co-transcriptional events15,16 and, ultimately, cell responses17. Thus, the ability to target key regulators of the chromatin environment presents a likely means by which to manipulate the transcriptional response to stimuli18,19.
Through the use of genomics, cell biology and genetic approaches, we found here that the human ATP-dependent helicase senataxin (SETX) exerted an inhibitory effect on the transcriptional response to viral infection. We showed that SETX promoted early termination by RNAPII at IRF3-dependent genes encoding antiviral products. This effect was dependent on the RNA-binding and ATPase activity of SETX and resulted in the inhibition of IRF3-dependent expression of genes encoding antiviral molecules. After viral infection, we observe higher expression of antiviral mediators in cells depleted of SETX and SETX-deficient cells, as well as in vivo in Setx−/− mice, than in SETX-sufficient cells or wild-type mice, respectively. In addition, reduced cellular levels of SETX rendered cells more resistant to viral infection and suppressed viral biogenesis. Notably, SETX-deficient cells derived from patients with ataxia with oculomotor apraxia (AOA2) displayed higher expression of antiviral mediators during infection and inhibition of viral replication than that of control cells from healthy donors. Our results describe a previously unknown function for SETX as a negative regulator of the antiviral response and may have relevant implications for neurological disorders characterized by SETX mutations.
Results
SETX inhibits expression of genes encoding antiviral factors
We were interested in identifying factors that might affect the transcriptional response to viruses. The putative ATP-dependent helicase SETX and its yeast ortholog Sen1 are reported to be positive regulators of basal gene transcription20,21,22. Sen1 has also been shown to negatively regulate the expression of genes induced by cell-wall perturbations23, a system functionally equivalent to the cellular response to stimuli in mammals24. Those findings, along with the possibility of a connection between viral infection and AOA2 or amyotrophic lateral sclerosis (ALS4), led us to hypothesize that SETX might regulate the transcriptional response of mammalian cells to viral infection1,5.
We first investigated the subcellular localization of SETX in the human lung epithelial cell line A549 before and after infection with the influenza A virus strain A/Puerto Rico/8/1934(ΔNS1) (PR8ΔNS1). We used SETX-sufficient ('wild-type') and SETX-deficient human fibroblasts25 as controls for antibody staining. Immunohistochemistry showed a nuclear distribution of SETX in PR8ΔNS1-infected A549 cells and PR8ΔNS1-infected wild-type control fibroblasts (Supplementary Fig. 1a), which indicated that the subcellular localization of SETX was unaffected by infection. SETX eluted together with high-molecular-weight nuclear complexes positive for RNAPII, by glycerol gradient fractionation (Supplementary Fig. 1b). The proposal of an association with the transcriptional machinery was further supported by the finding that SETX immunoprecipitated together with RNAPII (Supplementary Fig. 1c), similar to what has been observed with other cell types21. Together these results were consistent with a potential role for SETX in the regulation of transcription after infection.
To substantiate that hypothesis, we profiled the global mRNA-expression patterns of human A549 cells treated with SETX-specific small interfering RNA (siRNA) (siSETX) and nontargeting control siRNA (siCtrl) before and 4 h after infection with PR8ΔNS1. The depletion of SETX (via siSETX) was specific and resulted in more upregulation of genes encoding antiviral products than that of cells left untransfected or treated with siCtrl (P = 1.72 × 10−23 (hypergeometric test); Fig. 1a, Table 1, Supplementary Fig. 1d and Supplementary Table 1), with negligible effects on the expression of 'housekeeping' genes (Supplementary Fig. 1e). Conversely, depletion of the exonuclease XRN2, which, along with SETX, has been linked to positive control of the basal expression of two housekeeping genes20, did not result in a similar greater upregulation of the expression of genes encoding antiviral factors after infection than that of cells left untransfected or treated with siCtrl (Supplementary Fig. 1f–h and Supplementary Table 2). This result further confirmed the specificity of SETX during the cellular response to infection and suggested that its negative regulation of genes encoding antiviral molecules was exerted via an unknown mechanism.

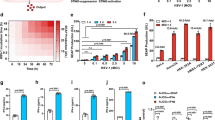
(a) Expression of genes encoding known cytokines and ISGs (left margin) in A549 cells left untransfected (No si) or transfected with siSETX, assessed early (4 h) after infection with PR8NS1, presented relative to that in A549 cells transfected with siCtrl. P < 0.01 (analysis of variance (ANOVA) with post-hoc TUKEY HSD test). (b–d) Gene expression (vertical axes) in A549 cells transfected with siRNA as in a, assessed after mock infection (Mock) or infection with PR8NS1 (Inf) (b) or after no treatment (−) or treatment with (+) exogenous IFN-β (c) or TNF (d); results are presented relative to those of infected cells left untransfected (b) or untransfected cells treated with IFN-β1 (c) or TNF (d). (e) Gene expression (vertical axes) in human STAT1-deficient (STAT1-def) and STAT1-sufficient (WT) human fibroblast cells mock infected or infected with PR8ΔNS1 and transfected with siRNA as in a, assessed 4 h after infection; results are presented relative to those of infected wild-type cells left untransfected. *P < 0.05, **P < 0.005 and ***P < 0.0005 (t-test). Data are from three independent experiments (mean and s.d. in b–e).
Phosphorylation of IRF3, which is a terminal event in the signaling cascade subsequent to sensing of a virus6, was unchanged in infected cells in which the gene encoding SETX was silenced (via siSETX) relative to its phosphorylation in cells left untransfected or treated with siCtrl (Supplementary Fig. 1i); this demonstrated that the increased expression of early genes encoding antiviral molecules in cells depleted of SETX was not caused by dysregulated signaling. The initial levels of viral genomic and messenger RNA in cells depleted of SETX and control cells were also equal (Supplementary Fig. 1j,k), which ruled out the possibility of indirect effects that resulted from differences in infection efficiency. Together these data suggested that upon infection, SETX negatively regulated antiviral factor–encoding genes induced immediately after sensing of a virus, when transcription of these genes was driven mainly by the transcription factor IRF3.
To determine whether the activity of SETX in our system was specific to viral infection, we further characterized the involvement of SETX in response to various stimuli. Our analyses confirmed that depletion of SETX increased the expression of genes encoding antiviral molecules in response to infection with PR8ΔNS1 (Fig. 1b) and showed that neither the addition of exogenous IFN-β (which signals through the Jak-STAT signaling cascade6) nor stimulation with tumor-necrosis factor (TNF) (which acts through other signaling cascades) was able to recapitulate the viral infection–induced gene-expression phenotype (Fig. 1c,d).
To further confirm the specificity of SETX activity downstream of the sensing of virus, we infected immortalized STAT1-deficient human fibroblasts with PR8ΔNS1. Since STAT1 protein is activated downstream of IFN-β and is essential for the IFN-β-dependent gene-expression program, this system allowed us to uncouple the specific arms of signaling linked to the sensing of virus and the IFN-β-dependent response. Depletion of SETX (via siSETX) in STAT1-deficient cells during infection resulted in higher expression of genes encoding antiviral molecules than that in STAT1-deficient control cells treated with siCtrl (Fig. 1e), which further supported the proposal of a model in which SETX acts specifically on early interferon-independent expression of genes encoding antiviral molecules.
SETX inhibits transcription of genes encoding antiviral molecules
We then investigated the effect that depletion of SETX had on viral infection–induced transcription by genome-wide analysis of nascent (elongating) RNA with global run-on sequencing (GRO-seq)18,26 in A549 cells treated with siCtrl or siSETX before and 4 h after infection with PR8ΔNS1 (Fig. 2 and Supplementary Fig. 2). Our GRO-Seq analyses show that depletion of SETX slightly reduced basal transcription in uninfected cells (Fig. 2a and Supplementary Fig. 2a–d), which confirmed published findings of a limited effect of SETX deficiency on the transcription of housekeeping genes20. In contrast, upon infection, SETX functioned as a negative regulator of the transcription of genes encoding antiviral molecules: the average profiles of engaged RNAPII indicated that depletion of SETX specifically increased the transcription of genes induced 4 h after infection. This set of genes displayed substantial enrichment for genes encoding molecules involved in the antiviral response (P = 2.35 × 10−78 (hypergeometric test); Fig. 2a and Supplementary Table 3). We illustrated the increase in inducible gene transcription in cells depleted of SETX for the representative virus-triggered gene IFNB1 (Fig. 2b) and quantified this by run-on quantitative PCR for the representative virus-induced genes IFIT1 and IFI6 (Fig. 2c).

(a) Meta-analysis of the GRO-seq signals of 872 genes in A549 cells treated with siCtrl or siSETX, assessed before (uninfected (UI)) and 4 h after (Inf) infection with PR8ΔNS1, presented as geometric mean coverage per 5 × 106 'reads' in a 30–base pair (bp) sliding window (vertical axis). (b) GRO-seq read coverage in the IFNB1 region (−, antisense strand; +, sense strand) for cells as in a, presented as base coverage per 5 × 106 'reads' (vertical axis); results are presented relative to those of infected cells transfected with siSETX. (c) Quantification of GRO-seq by quantitative PCR analysis of IFI6 and IFIT1 in cells as in a. UTR, untranslated region. (d) Traveling ratios (TR) of active infection-induced genes in cells as in a, calculated as average tags per bp in the promoter-proximal region (+30 to +300 bp around transcriptional start site) divided by that on the gene body (+300 bp to end of gene); numbers in parentheses (key) indicate total active genes. (e) Traveling ratios of active genes in siCtrl- or siSETX-treated A549 cells 4 h after infection with PR8ΔNS1; red indicates known ISGs. Data are from two experiments (error bars (c), s.d.).
To explore the mechanism responsible for the increased transcription in response to depletion of SETX, we compared the RNAPII density at the promoter divided by its density in the gene body in the two experimental conditions. This parameter, called the RNAPII 'traveling ratio'27, defines the propensity of RNAPII to be paused or 'licensed' into elongation. The traveling ratio of infection-induced genes and ISGs was indistinguishable in cells treated with siCtrl versus those treated with siSETX (P = 0.7093 (Kolmogorov–Smirnov test); Fig. 2d,e), which indicated that the increased antiviral gene transcription in siSETX-treated cells did not involve changes in elongation18 or pause-release27 via RNAPII and was therefore probably achieved at (or shortly after) transcription initiation28. Indeed, treatment of cells with flavopiridol, an inhibitor of the transcription-elongation factor p-TEFb that blocks transcription immediately following initiation, diminished the induction of genes encoding antiviral molecules to a similar extent in infected cells treated with siCtrl and those treated with siSETX (Supplementary Fig. 2e). Together our results suggested that during infection, SETX negatively regulated the transcription and, in turn, the expression of virus-induced genes.
SETX promotes premature termination at virus-induced genes
To gain additional insight into the mechanism of action of SETX, we undertook a series of experiments that relied on the transient expression of a constitutively active form of the transcription factor IRF3 (dominant active IRF3 (daIRF3)) that is sufficient to induce the expression of IFN-β and ISGs in uninfected cells6. If the effects of SETX were exerted downstream of IRF3, co-expression of SETX would counteract daIRF3-induced activation, and we did indeed observe this effect (Fig. 3a).

(a) Expression of IFNB and IFIT1 in A549 cells with transient expression of SETX or daIRF3 (−, 0 μg; +, 1 μg; ++, 5 μg) (horizontal axis), left untransfected (UT) or transfected with empty vector (EV) or vector encoding wild-type SETX (WT SETX), the SETX mutant lacking helicase activity (MUT1) or the SETX mutant lacking RNA-binding activity (MUT2) (key); results are presented relative to those of untransfected cells. (b) ChIP–quantitative PCR analysis of the recruitment of RNAPII and Flag-tagged wild-type or mutant SETX to the transcription start sites of IFIT1, IFIT2 and GAPDH (housekeeping gene) in A549 cells with (+) or without (−) transient overexpression of daIRF3 (horizontal axis), transfected as in a (key); results are presented relative to input. IgG, control immunoprecipitation with immunoglobulin G. (c) ChIP analysis of endogenous RNAPII and SETX at the transcriptional start sites of IFIT1, IFIT2 and GAPDH in A549 cells left untransfected or transfected with siCtrl or siSETX, assessed before (0 h) or 4 h after infection with PR8ΔNS1 (horizontal axis). (d) RNA-immunoprecipitation and quantitative PCR analysis of the binding of SETX to RNA derived from the 5′ untranslated region of IFIT1, IFIT2 or GAPDH in uninfected or PR8ΔNS1-infected A549 cells transfected with vector as in a (key); results are presented relative to those obtained with IgG. (e) Quantitative PCR analysis of RNA associated with the transcriptional start site (TSS RNA) of IFIT1, IFIT2 and GAPDH in A549 cells with (+) or without (−) overexpression of daIRF3 (horizontal axis) and transfected with vector as in a (key); results are presented relative to those of cells without overexpression of daIRF3 and transfected with empty vector. *P < 0.05, **P < 0.005 and ***P < 0.0005 (t-test). Data are from three experiments (a; mean and s.d.), one experiment representative of three independent experiments (b,c; mean and s.d. of three technical replicates) or three independent experiments (d,e; mean and s.d.).
The yeast ortholog of SETX, Sen1, induces the termination of transcription at small nucleolar RNAs29 by virtue of its RNA-binding and ATPase activities30. We thus assessed whether RNA-binding and enzymatic activities were similarly required for the function of human SETX. We first confirmed the helicase activity of purified human SETX (Supplementary Fig. 3a,b). In addition, we generated a human SETX mutant devoid of its enzymatic (helicase and translocase) activity by altering the conserved Walker A motif shared by many cellular ATPases (by substitution of glutamine for the lysine at position 1969) and another SETX mutant lacking RNA-binding activity by altering SETX motif IV (via substitution of aspartic acid for the glycine at position 2343)31 (Supplementary Fig. 3a,e). Our results demonstrated that both activities were essential for inhibiting daIRF3-dependent transcription (Fig. 3a). Chromatin-immunoprecipitation (ChIP) experiments with the same experimental conditions further showed that although wild-type and mutant SETX proteins were present in equal amounts at the promoter regions of induced genes encoding antiviral products, only wild-type SETX caused a concomitant decrease in RNAPII at the promoters (Fig. 3b). This effect was thus dependent on the ATPase and RNA-binding activity of SETX. Consistent with those observations, a decrease in endogenous SETX led to an increase in the loading of RNAPII at the promoters of the virus-induced genes IFIT1 and IFIT2 (Fig. 3c). Our data showing that the amount of RNAPII at promoters of antiviral molecule–encoding genes was inversely correlated with SETX activity, not SETX binding, suggested that SETX acted to promote premature termination of the transcription of IRF3-dependent genes by RNAPII.
We thought that our results would fit a simple model whereby SETX, via binding to nascent RNA, would translocate (by means of its ATPase activity) toward RNAPII and induce early termination. This model is reminiscent of how the helicase RhoA controls termination in Escherichia coli32 and is analogous to how Sen1 (the yeast ortholog of human SETX) controls termination at small nucleolar RNAs30. For this model to be true, wild-type SETX, unlike the mutant SETX lacking RNA-binding activity, would bind to 5′ RNA of active IRF3–induced genes encoding antiviral products. Moreover, in the same experimental setting, expression of wild-type SETX, unlike expression of the SETX mutants, would lead to the enrichment of prematurely terminated RNA derived from the 5′ end of the gene. To test this model, we first used RNA immunoprecipitation to investigate the binding of RNA by SETX. Consistent with our hypothesis, we observed that the SETX mutant lacking RNA-binding activity (and, to a similar extent, the SETX mutant lacking ATPase activity) was impaired in binding to RNA derived from the 5′ end of genes encoding antiviral products, relative to such binding by wild-type SETX (Fig. 3d). These results suggested that although both mutants were targeted as efficiently as wild-type SETX to promoters of IRF3-dependent genes (Fig. 3b), they were both unable to bind to such nascent RNAs. These results indicated that the targeting of SETX to gene promoters was independent of its nascent RNA–binding and ATPase activity but that both activities were needed to promote termination of transcription by RNAPII.
We next investigated the effect of SETX on the amount of transcription start site–associated short RNA33,34. We observed that expression of wild-type SETX increased the amount of such RNA at IRF3-dependent genes, but expression of the SETX mutants lacking ATPase or RNA-binding activity did not (Fig. 3e and Supplementary Fig. 3f–h). Together these data suggested that SETX promoted premature termination of transcription at the promoters of virus-induced genes and that depletion of SETX rendered the cells amenable to higher expression of genes encoding antiviral products.
SETX and the cofactor TAF4 coordinate the antiviral response
We next sought to determine which factors were responsible for coordinating the targeting of SETX to loci encoding antiviral molecules. An assay by affinity purification and tandem mass spectrometry revealed that the transcription-initiation factor subunit TAF4 was a specific interacting partner of SETX in influenza virus–infected cells (Supplementary Fig. 4a,b and Supplementary Table 4). In line with that finding, siRNA-mediated depletion of TAF4 in PR8ΔNS1-infected A549 cells resulted in reduced amounts of SETX at the target genes IFIT1 and IFIT2 (Fig. 4a). TAF4 has been shown to function as a transcriptional co-activator in inducible and stimulus-dependent gene-expression programs (for example, Wnt signaling)35,36. Upon infection with PR8ΔNS1, depletion of TAF4 led to less induction of the expression of genes encoding antiviral molecules than that in control cells left untransfected or treated with nontargeting siRNA (Fig. 4b). Conversely, overexpression of TAF4 increased the expression of such genes upon infection, compared with that of cells transfected with empty vector (Fig. 4c). These results suggested that SETX expression was able to inhibit the TAF4-dependent transactivation of IRF3-dependent genes. To prove this directly, we overexpressed daIRF3 together with TAF4 and SETX. Consistent with our hypothesis, our results showed that IRF3-dependent transcription was enhanced by coexpression of TAF4 and was counteracted by SETX (Fig. 4d).

(a) ChIP analysis of RNAPII and SETX the transcriptional start of IFIT1, IFIT2 and GAPDH in uninfected (0 h) or PR8ΔNS1-infected (4 h) A549 cells left untreated (No si) or treated with control siRNA (siCtrl) or TAF4-specific siRNA (siTAF4). (b) Gene expression (vertical axes) in A549 cells treated with siRNA as in a, assessed after mock infection or infection with PR8ΔNS1; results are presented relative those of infected cells without siRNA. (c) Gene expression (vertical axes) in A549 cells treated with siRNA as in a, transfected with empty vector or vector for the overexpression of wild-type TAF4 (TAF4 WT), and mock infected or infected with PR8ΔNS1; results are presented relative to those of infected, siCtrl-treated cells transfected with empty vector. (d) Gene expression (vertical axes) in A549 cells with (+) or without (−) transient expression of SETX or daIRF3 (horizontal axis) and left untransfected (UT) or transfected to overexpress wild-type TAF4 (TAF4 WT) or TAF4 lacking the homology domain required for activator-dependent gene expression (TAF4ΔTAFH) (key); results are presented relative to those of untransfected cells. *P < 0.05, **P < 0.005 and ***P < 0.0005, versus siCtrl-treated cells (a–c) or the corresponding daIRF3-overexpressing cells (d) (t-test). Data are from three independent experiments (mean and s.d.).
TAF4 contains a homology domain (TAFH) shown to be required for activator-dependent gene expression35 via direct recruitment of positive and negative regulators of transcription37. As expected, overexpression of a TAF4 mutant with deletion of this domain abolished TAF4's transactivation of the expression of genes encoding antiviral molecules (Fig. 4d). These data, along with protein-interaction studies showing binding of TAFH and SETX (Supplementary Fig. 4c and Supplementary Table 4), suggested that TAF4 co-activated genes encoding antiviral factors and that its effect was modulated by SETX. Our data showed that a SETX-TAF4 axis might thus represent a pivotal arm of the cellular response to pathogens, whereby loss of function in either protein results in dysregulated innate immune responses to pathogens.
SETX-deficient human cells are hyper-responsive to infection
Finally, to determine the physiological consequence of the loss of SETX function on viral replication, we first profiled viral growth kinetics in human cells depleted of SETX. The growth of both wild-type influenza virus PR8 and PR8ΔNS1 was repressed in the absence of SETX (Supplementary Fig. 5a,b), consistent with a role for SETX in controlling the expression of cytokines and antiviral mediators. Consistent with the role of SETX in controlling the early antiviral response to the sensing of virus, its effect on viral growth depended on the presence of RIG-I, the cellular sensor for influenza virus (Supplementary Fig. 5c). Overall, these results suggested that the increased expression of genes encoding antiviral products in SETX-deficient cells inhibited viral growth, which would directly link SETX to attenuation of the antiviral response and would indicate SETX is key in a cellular buffering system that controls the magnitude of the host response to viruses.
Loss of SETX function has been linked the neurodegenerative diseases ALS4 (ref. 38) and AOA2 (ref. 39). We therefore performed ex vivo experiments to determine whether SETX-deficient cells derived from patients with AOA2 could recapitulate the enhanced antiviral response observed in A549 cells depleted of SETX (Fig. 1c,d) and immortalized glial cells treated with siSETX (Fig. 5a). In line with our initial observations, SETX-deficient lymphoblastoid cells (Fig. 5b) and primary human fibroblasts (Fig. 5c) derived from patients with AOA2 (ref. 25) displayed higher expression of virus-induced genes upon infection than that of cells from healthy donors. We were also able to recapitulate these observations in an in vivo setting by intranasal administration of immunostimulatory defective-interfering RNA from Sendai virus into Setx−/− mice40. We observed higher expression of genes encoding antiviral products in Setx−/− lungs than in their wild-type counterparts (Fig. 5d), which further reinforced the hypothesis of a physiological role for SETX in regulating the expression of such genes.

(a) Expression of IFNB1, IFIT1 and SETX in human glial cells treated with no siRNA, siCtrl or siSETX (key), assessed 4 h after mock infection or infection with PR8ΔNS1 (horizontal axes); results are presented relative to those of infected cells with no siRNA. (b,c) Expression of IFNB1, CXCL9 and IFIT1 in lymphoblastoid (LCL) cells (b) and fibroblasts (c) isolated from healthy donors (WT) or patients with AOA2 (SETX-def), assessed 4 h after infection as in a; results are presented relative to those of infected wild-type cells. (d) Quantitative PCR analysis of Ifnb1, Ddx58 (encoding RIG-I) and Ifit1 in the lungs of wild-type or Setx−/− mice 6 h after intranasal treatment with 4 μg of in vitro–transcribed Sendai virus defective-interfering RNA (SeV DI) (n = 3 mice per group) or PBS (n = 4 mice per group); results are presented relative to those of wild-type lungs treated with Sendai virus defective-interfering RNA. (e) Quantitative PCR analysis of IFNB1, IFIT1 and CXCL9 in wild-type or SETX-deficient human lymphoblastoid cells not reconstituted (−) or reconstituted with empty vector (EV) or vector encoding wild-type (SETX) or mutant SETX lacking ATPase activity (MUT) and mock infected or infected with PR8ΔNS1; results are presented relative to those of infected wild-type cells. (f) Expression of IFNB1 and IFIT1 in wild-type and SETX-deficient human fibroblasts in the absence (−) or presence of vehicle control (DMSO) or flavopiridol (key), mock infected or infected with PR8ΔNS1. (g) Luciferase signal in A549 cells pretreated for 12 h with supernatants of PR8ΔNS1-infected wild-type or SETX-deficient lymphoblastoid cells (high multiplicity of infection; collected at 8 and 12 h after infection), then infected with a PR8-GLuc reporter virus (multiplicity of infection, 1) and assessed 8 h after that infection. *P < 0.05, **P < 0.005 and ***P < 0.0005 (t-test). Data are from three independent experiments (mean and s.d. in a–f; mean and s.e.m. in g).
We also confirmed that the function of SETX was dependent on its ATPase activity in primary human fibroblasts: expression of wild-type SETX in SETX-deficient human fibroblasts, unlike expression of the SETX mutant lacking enzymatic activity, was able to reconstitute a normal host response to infection (Fig. 5e and Supplementary Fig. 5d). Notably, similar to their upregulation in epithelial cells (Supplementary Fig. 2e), the upregulation of genes encoding antiviral molecules in SETX-deficient cells was reduced to wild-type levels by the p-TEFb inhibitor flavopiridol (Fig. 5f).
The consequences of dysregulated expression of genes encoding antiviral molecules in SETX-deficient cells were also apparent at the protein level in infected cells: secretion of cytokines and chemokines was greater in SETX-deficient fibroblasts than in wild-type fibroblasts (Supplementary Fig. 5e). Naive cells pretreated with the supernatants of infected SETX-deficient fibroblasts were also more resistant to infection with a reporter virus than were such cells treated with supernatants derived from infected wild-type cells (Fig. 5g). Similar to siSETX-treated A549 cells, SETX-deficient fibroblasts suppressed the growth of wild-type influenza virus PR8 (Supplementary Fig. 5f). Finally, we also observed higher expression of genes encoding antiviral molecules and concomitant inhibition of viral biogenesis in SETX-deficient lymphoblastoid cells upon infection with the neurotropic Kunjin virus, a subtype of West Nile virus, compared with that in SETX-sufficient lymphoblastoid cells (Supplementary Fig. 5g,h). Although the absence or depletion of SETX rendered the epithelial, fibroblast and glial cells we tested hyper-responsive to viral infection, the overall transcriptional regulation of SETX was different among the three cell types (Fig. 5a); this suggested an additional layer of cell-type and stimulus specificity in the control of SETX-dependent transcription activity. Of note, we found that infection of glial cells with influenza virus resulted in higher SETX expression at the level of RNA and protein than that in uninfected cells (Fig. 5a and Supplementary Fig. 5i). It is possible that upregulation of SETX expression might reflect a cell type–specific built-in system to downregulate genes encoding antiviral products via a negative feedback loop that controls the magnitude of the antiviral response. Overall, these results indicated that SETX-mediated transcriptional control was disrupted in cells isolated from patients with SETX deficiency and confirmed, across a broad range of cell types and viruses, the role of SETX in regulating the initial steps of the transcription of genes encoding antiviral molecules (Supplementary Fig. 5j).
Discussion
The cellular transcriptional response to infection is regulated by various cellular factors that ensure the temporal and dynamic expression of specific genes encoding antiviral factors. We have described here several lines of evidence that identified SETX as a factor that controls the antiviral response. First, depletion of SETX resulted in increased expression of several early infection-induced genes, including those encoding key cytokines (for example, IFN-β) and other antiviral mediators (for example, IFI6 and IFIT1) important for the resolution of infection. Second, overexpression of SETX was able to counteract the IRF3-dependent activation of endogenous loci encoding antiviral products. Third, human cells (lymphoblastoid and fibroblast) derived from patients with loss-of-function mutations in SETX displayed greater upregulation of virus-induced genes upon infection than that of wild-type cells. Fourth, SETX-deficient human cells, like cells depleted of SETX, inhibited viral biogenesis. Finally, we were also able to recapitulate in vivo the effects seen with depletion of SETX, whereby Setx−/− mice had higher expression of inflammatory mediators upon immunostimulation than did wild-type mice.
Our findings indicated that SETX negatively regulated genes encoding antiviral products by promoting early termination by RNAPII. The mechanism used by SETX to inhibit genes encoding antiviral products parallels the mechanism described for Sen1, the yeast homolog of SETX, in controlling basal transcription at noncoding RNA loci. In fact, experiments with mutant yeast strains with altered RNAPII processivity have shown that Sen1 tracks along nascent RNA and induces termination by RNAPII29 in a manner very similar to the Rho-dependent attenuation of transcription in bacteria. In vitro experiments have supported that view and have revealed additional features of Sen1 activity30 that suggest such a mechanism could be in place for regulating promoter activity.
Several of our observations supported the proposal of promoter-proximal termination events during RNAPII-mediated transcription as the mechanism by which SETX limited the transcription of genes encoding antiviral molecules. Our experimental data showed that depletion of SETX caused a specific increase in transcriptionally active RNAPII across virus-induced genes, with a concomitant increase in the level of their mRNA, in a manner that was independent of pause-release, elongation and termination by RNAPII. The ability of SETX to counteract transcriptional activity depended on its ATPase activity. In line with that, wild-type SETX, unlike the SETX mutants lacking enzymatic or RNA-binding activity, bound to the 5′ end of nascent RNA at genes encoding antiviral molecules and increased the level of prematurely terminated RNA at the transcriptional start site of IRF3-dependent genes encoding antiviral factors. Finally, overexpression of SETX modulated IRF3-dependent gene expression in a dose-dependent manner, which suggested that SETX levels can dynamically compete with IRF3-mediated transcription. Overall, our data have revealed a previously unknown function for SETX in negatively regulating the magnitude of an antiviral response at both the cellular level and the organismal level. We thus suggest that the recruitment of SETX to its target genes drives the early termination of the transcription of genes encoding antiviral factors and limits their overall expression. SETX-mediated termination might depend directly on sequence-specific recognition of the target RNA itself through its RNA-binding domain and might require the translocase activity of SETX. Alternatively, SETX-mediated termination might be driven by its interaction with structural elements, such as R loops, formed at nascent transcript. The latter would require the helicase activity of SETX and is a likelier possibility, given that SETX can resolve RNA-DNA hybrids, such structures can be formed at promoters of genes encoding inflammatory molecules41, and free 5′-end RNA at promoters can increase R-loop formation and avoid the requirement for RNA cleavage to promote transcriptional termination.
Mechanistically, depletion of SETX led to diminished transcription of some housekeeping genes in the basal state, while during infection it increased the activation of genes encoding antiviral molecules. Such positive and negative regulation could simply be dependent on the targeting of SETX activity to different gene regions (the 3′ end region for housekeeping genes versus the promoter region for genes encoding antiviral molecules) by different binding partners. This would then result in a different effect on transcription for specific gene subsets. For the housekeeping genes, the interaction of SETX with 3′-end cleavage and termination complexes is supposed to coordinate proper termination16. In contrast, for the genes encoding antiviral products, control of premature termination by SETX at promoters was dependent on IRF3, a master transcription factor activated by the incoming virus, and required the cofactor TAF4. Indeed, upon infection, IRF3-dependent gene expression was potentiated by TAF4 and was subject to negative regulation by SETX. Such multilevel regulation is key during the cell response to viruses due to the detrimental effect of high and/or prolonged expression of genes encoding antiviral molecules (discussed below) and is reminiscent of how signaling via E protein and Wnt coordinates the recruitment of transcription factors and other cofactors to regulate stimulus-dependent gene expression35,36. Strikingly, in those studies35,36, as in ours, TAF4 seemed to serve a pivotal role as a binding platform for the recruitment of positive and negative effectors that coordinate transcriptional activity during a cell response.
In summary, we have revealed a previously unknown mechanism for regulating the expression of genes encoding antiviral molecules by an ATP-dependent helicase that controlled the expression of the gene encoding IFN-β and other key genes encoding antiviral products. Our results suggest that during viral infection, SETX functions as a surveillance factor to ensure that an appropriate physiological response to viruses is elicited. We hypothesize that SETX has evolved a role in controlling transcriptional responses to viruses and thus might be a target amenable to the treatment of infections and inflammatory disorders. In the conditions studied here, the activity and regulation of SETX were specific for signaling downstream of the sensing of a virus; however, we do not discount the possibility that loss of SETX might disrupt other transcriptional networks.
Dysregulation of transcription has been linked to disease development42, as shown for TAF4 deficiency, which confers susceptibility to Huntington disease43. Inborn SETX mutations have been linked to the development of the neurodegenerative diseases ALS4 (ref. 38) and AOA2 (ref. 39). Our data indicated that loss of SETX resulted in an altered temporal activation of the innate immune response to viruses. Unlike congenital diseases with constitutive expression of inflammatory mediators4,44, SETX-driven changes to the innate immune response manifested experimentally only upon infection. We postulate that the inducible phenotypes we observed in the context of SETX deficiency might be triggered environmentally by exposure to viral agents. Transient states of excessive inflammation might, over time, result in the local dysregulation of immunological homeostasis and progressive deterioration of affected tissues. We propose that infection has an important role in the initiation or progression of AOA2 and ALS4.
Methods
Cells and viruses.
A549 cells, 293T human embryonic kidney cells, Madin-Darby canine kidney (MDCK) cells and SVGp12 fetal glial cells were originally obtained from the American Type Culture Collection. Cells were maintained at 37 °C with 5% CO2 in Dulbecco's minimal essential medium (Gibco, Life Technologies) supplemented with 2 mM glutamine, 10% FBS (Hyclone), 100 U/ml penicillin and 100 μg/ml streptomycin (Gibco, Life Technologies). SETX-2RM (SETX-deficient) and C3ABR (wild-type) lymphoblastoid cells were maintained as described above. SETX-1RM human fibroblasts (SETX-deficient fibroblasts) and normal foreskin fibroblasts25 were maintained in Dulbecco's minimal essential medium supplemented with 2 mM glutamine, 20% FBS (Hyclone), 100 U/ml penicillin and 100 μg/ml streptomycin. Influenza virus strain A/Puerto Rico/8/1934 (PR8) was propagated in 8- to 10-day-old embryonated chicken eggs. PR8ΔNS1 was propagated in MDCK cells expressing influenza virus nonstructural protein 1 (NS1). Sendai virus, Cantell strain, was propagated in 10-day-old embryonated chicken eggs. Kunjin virus was provided by J. Lim. RIG-I deficient A549 cells were provided by A. Garcia-Sastre.
Viral growth curves.
A549 cells were treated with 50 nM SETX-specific siRNA or nontargeting siRNA (ON-TARGETplus SMARTpool; Thermo Scientific) or were left untreated for 48 h before infection with either influenza virus strain PR8 at a multiplicity of infection (MOI) of 0.05 plaque-forming units (PFU) per cell or with PR8ΔNS1 at an MOI of 5 PFU/cell. Supernatants of the infected cells were collected at 12 and 24 h after infection. Where RIG-I-deficient A549 cells or SETX-deficient primary fibroblasts were used, cells were plated the day before being infected with influenza virus strain PR8 at a low MOI (0.05 PFU/cell). Supernatants from infection were collected at 12 and 24 h after infection. Viral titers were determined by plaque assay on MDCK cells. For infection with Kunjin virus, wild-type or SETX-deficient fibroblasts were infected with virus at an MOI of 0.05 PFU/cell. Supernatants were collected 32 h after infection, and viral titers were determined by plaque assay on Vero cells. Plaques were visualized by immunostaining with monoclonal antibody to the West Nile Virus envelope protein (E24; BEI Resources), horseradish peroxidase–conjugated goat anti–mouse IgG (sc-2005; Santa Cruz Biotechnology) and True Blue peroxidase substrate (KPL).
Immunofluorescence.
A549 cells were cultured on coverslips overnight and then were infected with the appropriate viral strain. At the appropriate time after infection, cells were fixed for 10 min at room temperature in 3% paraformaldehyde (EMS). Coverslips were washed in 1× PBS and nonspecific binding was blocked by incubation for 1 h at room temperature with blocking solution (1 mg/ml BSA, 3% FBS, 0.1% Triton X-100 and 1 mM EDTA, pH 8.0, in PBS). Cells were then probed for 1 h with custom-made sheep antibody to SETX (diluted 1:400; produced in-house by O.J.B. and M.F.L.) and mouse antibody to tubulin (diluted 1:2,000; 100109-MM05; Sino Biological), followed by detection with Alexa Fluor 488–conjugated (green) donkey anti–sheep IgG (heavy and light chain) (A-11015; Life Technologies) and Alexa Fluor 594–conjugated (red) donkey anti–sheep IgG (heavy and light chain) (A-11005; Life Technologies). DNA was counterstained with DAPI (4,6-diamidino-2-phenylindole).
Enzyme-linked immunosorbent assay.
SETX-1RM cells and normal foreskin fibroblasts were left untreated or were treated with PR8ΔNS1 at an MOI of 100 PFU/cell, and supernatants were collected at 4 and 12 h after infection. Human IFN-α2, IFN-γ and chemokine CXCL10 in culture supernatants were analyzed by Luminex xMAP multiplexing technology (Millipore).
GRO sequencing.
Transcriptionally active nuclei from infected or untreated A549 cells were prepared after the cells were allowed to swell for 5 min in ice-cold swelling buffer (10 mM Tris, pH 7.5, 2 mM MgCl2 and 3 mM CaCl2). Pelleted cells were re-suspended in 1 ml lysis buffer (10 mM Tris, pH 7.5, 2 mM MgCl2, 3 mM CaCl2, 10% glycerol, 0.5% NP40, 2U/ml SUPERaseIN (Ambion)) and were pipetted 20 times with a P1000 tip with the end cut off to reduce shearing. The volume was brought to 10 ml with lysis buffer and nuclei were pelleted at 600g for 5 min. Nuclei were washed in 10 ml lysis buffer and re-pelleted. A small aliquot was taken for staining with Trypan blue to ensure that lysis occurred and nuclei were still intact. Nuclei were re-suspended in 1 ml freezing buffer (50 mM Tris-Cl, pH 8.3, 40% glycerol, 5 mM MgCl2 and 0.1 mM EDTA) with a P1000 tip with the end cut off and were re-pelleted and re-suspended in 500 μl freezing buffer and separated into 100-μl aliquots and frozen in liquid nitrogen. GRO-Seq and library preparation was done as described26. For confirmation of ChIP followed by deep sequencing and GRO-seq, experiments with independent biological replicates were done by quantitative PCR. Samples were sequenced on a HiSeq2000 or a HiSeq 2500 instrument in accordance with the manufacturer's protocols (Illumina).
Analysis of transcription start site–associated RNA.
Transcription start site–associated RNA was quantified by a published method33. Total RNA was extracted and large RNA (>200 nucleotides) and short RNA (<200 nucleotides) were isolated separately with miRNeasy Mini and MinElute kits (Qiagen). Oligo-adenylation and cDNA synthesis of short RNA was performed with Superscript III and an oligo(dT) primer with an additional 5′ adaptor sequence. Quantitative PCR analysis of the short RNA was performed with forward primers specific to the 5′ untranslated region of the gene of interest and a universal reverse primer that matched the 5′ adaptor sequence. 5s rRNA served as a normalization standard. Statistical significance was determined with a two-tailed Student's paired t-test.
Immunological stimulation of Setx−/− and wild-type mice.
Wild type and Setx−/− mice40 were anesthetized before receiving an intranasal dose of either PBS alone or 4 μg of in vitro–transcribed Sendai virus defective-interfering RNA diluted in PBS. 20 μl of PBS was administered in each nostril. Six hours after administration, mice were killed and lungs were extracted. RNA was isolated from lungs with an RNeasy Mini kit according to the manufacturer's recommendations (Qiagen). cDNA was synthesized with Superscript III Reverse Transcriptase according to the manufacturer's recommendations (Life Technologies). All procedures were performed in compliance with procedures approved by the University of Queensland animal use committee.
Nuclear extracts and glycerol-gradient sedimentation.
Nuclear extracts were prepared according to Dignam Roeder methods and were separated by centrifugation for 8 h at 50,000 r.p.m. (in a Beckman SW-Ti55) on a 5–40% glycerol gradient before being fractionated.
Generation and transduction of lentivirus.
Lentivirus expressing wild-type SETX, mutant SETX lacking ATPase activity or GFP were generated by transfection of plasmids encoding the relevant transgene in the pLX304 lentivirus backbone (Addgene) or the pLentiIII-2A-GFP backbone (ABM), VSV-G (pMD2.G, Addgene) and Gag-Pol (a gift from M. Evans), at a ratio of 2.5:1.5:1, into 293T cells. Supernatants were collected 24 and 48 h after infection and viruses were concentrated with a Lenti-X concentrator according to the manufacturer's recommendations (Clontech). Optimal titers for transduction were determined by serial dilution on the appropriate cell type in the presence of 10 μg/ml polybrene (Millipore).
Quantitative PCR.
RNA was extracted with an RNeasy Mini Kit (74106; Qiagen). RNA was treated with an RNase free DNase kit (Qiagen). Cells were homogenized with QIAshredder columns (79656; Qiagen) before RNA extraction. Proteins were also simultaneously recovered from cell lysates by acetone precipitation of the flow-through from RNeasy spin columns, according to manufacturer's instructions (Qiagen). cDNA was synthesized with SuperScript VILO Master Mix (11755050; Life Technologies) or SuperScript III First-Strand Synthesis SuperMix (18080-400; Life Technologies). Quantitative PCR was performed with SYBR Green (Roche) or LightCycler 480 Probes Master mix (Roche).
Primers.
Taqman Probes (4331182; Life Technologies) were as follows (Life Technologies catalog numbers in parentheses): SETX (Hs00209294_m1), IFIT1 (Hs01911452_s1),ISG15 (Hs01921425_s1), IFNB1 (Hs01077958_s1), EGR1 (Hs001152928_m1).
Sequences of primers used for quantitative RT-PCR (human) were as follows: β-actin forward, 5′-ACCTTCTACAATGAGCTGCG-3′, and β-actin reverse, 5′-CCTGGATAGCAACGTACATGG-3′; GAPDH forward, 5′-GCAAATTCCATGGCACCGT-3′, and GAPDH reverse, 5′-GCCCCACTTGATTTTGGAGG-3′; 18S forward, 5′-GTAACCCGTTGAACCCCATT-3′, and 18S reverse, 5′-CCATCCAATCGGTAGTAGCG-3′; PR8 HA forward, 5′-AAAGAAAGCTCATGGCCCAACC-3′, and PR8 HA reverse, 5′-TCCTTCTCCGTCAGCCATAGCA-3′; PR8 PB1 forward, 5′-TCATGAAGGGATTCAAGCCG-3′, and PR8 PB1 reverse, 5′-GGAAGCTCCATGCTGAAATTG-3′; PR8 PB2 forward, 5′-AGAGACGAACAGTCGATTGCCG-3′, and PR8 PB2 reverse, 5′-ATCGCTGATTCGCCCTATTGAC-3′; PR8 M1 forward, 5′-GCAGGGAAGAACACCGATCTTGA-3′, and PR8 M1 reverse, 5′-ACGGTGAGCGTGAACACAATCC-3′; IFIT2 forward, 5′-AGGCTTTGCATGTCTTGG-3′, and IFIT2 reverse, 5′-GAGTCTTCATCTGCTTGTTGC-3′; CXCL10 forward, 5′-CCTTATCTTTCTGACTCTAAGTGGC-3′, and CXCL10 reverse, 5′-ACGTGGACAAAATTGGCTTG-3′; CXCL9 forward, 5′-TGGTGTTCTTTTCCTCTTGGG-3′, and CXCL9 reverse, 5′-AACAGCGACCCTTTCTCAC-3′; TAF4 forward, 5′-CAGCGAATCCTCAAACAGTCA-3′, and TAF4 reverse, 5′-ACCACAGTAGTTCCTATTTGTGC-3′; IFI6 exon forward, AGCAGCGTCGTCATAGGTAA, and IFI6 exon reverse, AGATACTTGTGGGTGGCGTA.
Sequences of primers used for quantitative RT-PCR (mouse) were as follows: Ifit1 forward, GCCTATCGCCAAGATTTAGATGA, and Ifit1 reverse, TTCTGGATTTAACCGGACAGC; Ifnb1 forward, AGCTCCAAGAAAGGACGAACA, and Ifnb1 reverse, GCCCTGTAGGTGAGGTTGAT; Actb forward, TTACGGATGTCAACGTCACAGTTC, and Actb reverse, ACTATTGGCAACGAGCGGTTC; Ddx58 (RIG-I) forward, GAAGAGCCAGAGTGTCAGAATC, and Ddx58 (RIG-I) reverse, AGCTCCAGTTGGTAATTTCTTGG.
Sequences of primers used for ChIP followed by quantitative PCR (human) were as follows: GAPDH 5′ forward, ACAGTCAGCCGCATCTTCTT, and GAPDH 5′ reverse, TTCTCTCCGCCCGTCTTC; IFIT1 5′ forward, AGAGGAGCCTGGCTAAGCA, and IFIT1 5′ reverse, GGTTGCTGTAAATTAGGCAGC; IFIT2 5′ forward, TGCACTGCAACCATGAGG, and IFIT2 5′ reverse, TGACTCAACAGCACTACCGA; and 5s RNA forward, TACGGCCATACCACCCTGAA, and 5s rRNA reverse, GCGGTCTCCCATCCAAGTAC.
Sequences of primers used for transcription start site–associated RNA were as follows: universal primer, GGCCACGCGTCGACTAGTAC; IFIT1 forward, GGACACACCCACAGCTTCA; IFIT2 forward, AAAAGGAACCAGAGCCACTTGT; IFI6 forward, CCTTCAGCCGGAGAACCGT; and GAPDH forward, CAAGACCTTGGGCTGGGACTG.
siRNA transfection.
Cells were transfected through the use of Lipofectamine RNAiMAX transfection reagent according to the manufacturer's instructions (Invitrogen). Cells were transfected with siRNA pools targeted to the gene encoding human SETX (23064; Dharmacon), TAF4 (L-006878-00-0005; Dharmacon) or XRN2 (L-017622-01-0010; Dharmacon) or a control non-targeting pool (D-001810-10-05; Dharmacon) at a final siRNA concentration of 50 nM. Transfected cells were used for further assays at 48 h after transfection, and the efficiency of gene knockdown was determined by quantitative PCR and/or immunoblot analysis.
Microarray analysis.
A549 cells transfected with siRNA targeting the gene encoding SETX or XRN2 or control nontargeting siRNA were infected in triplicate with PR8ΔNS1 (MOI 3). Untransfected cells were also infected, as a control. Total RNA was isolated from infected and uninfected cells with a Qiagen RNeasy kit. 200 ng of total RNA per sample was then used to prepare labeled RNA that was hybridized to Human HT-12 v4 Expression BeadChips (Illumina). Data were analyzed with Genespring software (version 12.5).
Analyses of the effect of treatment with siSETX and siXRN2 were conducted separately. For specific analysis of the effect of depletion of SETX or XRN2 on the magnitude of the cell response during infection, raw signal values obtained with uninfected and infected cells in the various siRNA treatment conditions in were quantile-normalized before being baseline-transformed to the medians of signal values for the corresponding uninfected siRNA-treated samples. For the identification of probe sets with statistically significant differences in response magnitude (P < 0.01), ANOVA followed by a post-hoc (Tukey's honest significant difference) test was conducted. Genes differentially regulated by treatment with siSETX or siXRN2 were identified as genes with a change expression of ≥1.5-fold (P < 0.01) relative to their expression in siCtrl-treated cells. Where indicated, infection-induced genes were identified as genes with a change expression of ≥1.5-fold (P < 0.01) in virus-infected, siCtrl-treated cells relative to their expression in uninfected siCtrl-treated cells. All computations of P values were subjected to multiple-testing correction by the Benjamini-Hochberg method. For purposes of presentation in the heat maps, genes represented by multiple probe sets in the microarray were plotted as the averaged values of those probe sets.
For evaluation of the effect of depletion of SETX or XRN2 under basal conditions, raw signal values of uninfected cells under the siRNA-treatment conditions were quantile-normalized before being baseline-transformed to the median of all samples. Analyses for SETX and XRN2 were done separately. An ANOVA followed by a post-hoc (Tukey's honest significant difference) test was conducted for the identification of siRNA-regulated genes. Genes regulated by siRNA (targeting the gene encoding SETX or XRN2) were defined as genes with a change expression of ≥1.5-fold (P < 0.01) in cells treated with siRNA targeting the gene encoding SETX or XRN2 relative to their expression in siCtrl-treated cells. Normalized signal-intensity values of a list of canonical housekeeping genes were also used to determine the overall effect of the depletion XRN2 or SETX in cells. Full lists of affected genes are in Supplementary Tables 1 (SETX depletion) and 2 (XRN2 depletion).
Functional analyses of differentially regulated genes were conducted through the use of Ingenuity Pathways Analysis (Ingenuity Systems). This system was used for the identification of canonical pathways that showed 'enrichment' among groups of genes with significant changes in expression by microarray analysis. A right-tailed Fisher's exact test was used for calculation of P values determining the probability that each pathway assigned to that data set was due to chance alone. In addition, we also used DAVID gene-ontology analysis45 to identify genes associated with cytokine activity.
Antibodies.
Antibodies used were as follows: anti-RNAPII (MMS-126R; Covance); antibody to the RNAPII carboxy-terminal domain repeat (AB12795; Abcam); anti-RNAPII (A310-190A; Bethyl Laboratories); antibody to phosphorylated IRF3 (AB76493; Abcam); antibody to total IRF3 (SC9082; Santa Cruz Biotech); anti-HA (S190-108; Bethyl Laboratories); anti-LARP7 (A303-724A; Bethyl Laboratories); anti-HEXIM1 (A303-113A; Bethyl Laboratories); anti-DIS3 (A303-764A; Bethyl Laboratories); anti-TAF4 (ab69949; Abcam); custom-made anti-TAF4 (provided by R. Roeder); anti-tubulin (3873; Cell Signaling Technology); custom-made anti-PB2 (provided by P. Palese); and custom-made anti-SETX (provided by M.F. Lavin).
Immunoblot analysis.
Gradient gels were used based on the molecular mass of the proteins to be evaluated, followed by wet-transfer onto PVDF membranes.
ChIP.
ChIP was conducted as described18. For experiments with ChIP followed by quantitative PCR (described below), 10 min of crosslinking was used for both M1 antibody to Flag (overexpression experiments) and anti-SETX. Sonication was performed in a refrigerated Bioruptor (Diagenode), and conditions were optimized to generate DNA fragments of approximately 200–1,000 bp. Lysates were pre-cleared for 3 h with the appropriate isotype-matched control antibody (rabbit IgG (2729; Cell Signaling) or mouse IgG (5415; Cell Signaling)). Specific antibodies were coupled for 6 h with magnetic beads bound to anti–mouse IgG (Dynabeads M-280 Sheep Anti-Mouse IgG; 112-02; Invitrogen) or anti–rabbit IgG (Dynabeads M-280 Sheep Anti-Mouse IgG; 112-04; Invitrogen). Antibody-bound beads and chromatin were then immunoprecipitated overnight at 4 °C with rotation. After the wash steps, reverse crosslinking was carried out overnight at 65 °C. After digestion with RNase and proteinase K, DNA obtained by ChIP was then isolated with a MinElute kit (28004; Qiagen) and was used for downstream applications. The statistical significance of ChIP quantitative PCR analysis was determined with a two-tailed Student's paired t-test.
Next-generation sequencing data analysis.
Following adaptor trimming and quality filtering, reads were mapped to the human reference genome (GRCh37, hg19) with Bowtie software (version 0.12.9)46, for GRO-Seq and short RNA-Seq data, or Tophat software (version 2.0.10)47, for directional RNA-Seq data. Unique best matches were kept and were normalized to a total level of 5 × 106 tags per sample (GRO-Seq) or 30 × 106 tags per sample (directional RNA-Seq).
To identify genes with differences in the level of engaged RNAPII, we first counted the GRO-Seq tags that mapped to the gene body, excluding the first 1-kilobase region following the transcription start site. The resulting counts were then normalized by the 'trimmed mean of M' method48, and genes with a statistically significant difference in expression in infected cells relative to that in uninfected cells (q < 0.05) were identified in a gene-wise log-likelihood ratio test with correction for multiple testing by the Benjamini-Hochberg false-discovery rate, with the Bioconductor EdgeR software package49. A list of differentially expressed genes is in Supplementary Table 3.
For GRO-Seq and directional RNA-Seq, coverage maps relative to gene features and the transcription start site, we calculated base coverage in 200 bins of equal size across each feature and/or 5-kilobase flanking regions. Aggregate plots of GRO-Seq profiles across genes were prepared by calculation of the geometric mean coverage for each bin. GRO-Seq traveling ratios for active genes were calculated as described27.
Affinity purification–tandem mass spectrometry.
A549 cells expressing Flag-SETX or empty vector (2 × 108 cells) were infected for 4 h with PR8ΔNS1 or were left untreated. Cells were collected by scraping and washed with PBS, then were pelleted and re-suspended in 20 ml cold lysis buffer (0.25% NP40, 50 mM Tris-HCl, pH 7.5, 200 mM NaCl, 1 mM EDTA, protease, phosphatase, inhibitors and NEM). Cells were mechanically disrupted with a Dounce homogenizer, and sonication with a Bioruptor was performed at 4 °C for five cycles (30 s on and 30 s off). Insoluble material was then pelleted for 20 min at 15,000 r.p.m., and the supernatant was precleared by incubation for 3 h on an overhead shaker at 4 °C with nonspecific beads. 50 μl of beads immunoprecipitated with Dynabeads coupled to anti-Flag (M2; Sigma) was added to the pre-cleared lysate and immunoprecipitation was performed for 3 h at 4 °C. Beads were then washed four times with cold buffer (50 mM Tris–HCl, pH 7.5, 200 mM NaCl, 0.25% NP40 and 1 mM EDTA) and two successive times with detergent-free buffer to avoid interference with mass spectrometry analysis. Beads were eluted three times with 3xFlag peptide (Sigma) in a buffer of 50 mM Tris–HCl, pH 7.5, and 200 mM NaCl. The eluted material was subjected to acetone precipitation and resuspended proteins were analyzed via mass spectrometry after digestion with trysin.
For the analysis of mass spectrometry 'hits', initial comparisons were first made between proteins precipitated with empty vector and Flag. Proteins with a threefold increase (from duplicate experiment averages) in either peptide score or area score for Flag precipitation versus EV precipitation were then considered. Shared proteins were then selected on the basis of a stringency setting as those with at least three peptides and an area score of greater than 5 × 106.
Helicase assays.
Flag tagged wild-type SETX or mutant SETX lacking ATPase activity was purified from 5 × 108 A549 cells expressing those. Cells were lysed in 30 ml cold lysis buffer (0.25% NP40, 50 mM Tris–HCl, pH 7.5, 200 mM NaCl, 1 mM EDTA, protease, phosphatase, inhibitors and 10 mM NEM). Cells were mechanically disrupted with a Dounce homogenizer, and sonication with a Bioruptor was performed at 4 °C for five cycles (30 s on and 30 s off). The NP40 concentration was raised to 0.5% and NaCl was raised to 300 mM. The extract was rotated for 15 min in a cold room. Insoluble material was then pelleted at 15,000 r.p.m. for 20 min. The supernatant was precleared by incubation for 3 h at 4 °C on an overhead shaker with nonspecific beads. 200 μl immunoprecipitaion beads (anti-Flag (M2; Sigma) coupled to Dynabeads) were added to the precleared lysate, followed immunoprecipitation for 3 h at 4 °C. Beads were then washed four times with cold buffer (50 mM Tris–HCl, pH 7.5, 300 mM NaCl, 0.25% NP40, 1 mM EDTA) and two successive times with helicase buffer (20 mM HEPES-KOH, pH 7.8, 50 mM NaCl, 2 mM ATP, 1 mM MgCl2, 2 mM DTT and 100 μg/mL BSA). Aliquots of beads were quantified by gel electrophoresis followed by Coomassie staining. Immobilized Flag-tagged wild-type SETX, mutant SETX and GFP (control) were used in the unwinding assay with 200 ng of synthetic DNA-RNA hybrid (Integrated DNA Technologies) as described50. After incubation for 1.5 h at 37 °C on a Thermomixer programmed for mixing at 700 r.p.m. with 30 s on and 30 s off, reactions were analyzed with an Agilent 6000 Pico Kit. The RNA and/or DNA oligonucleotide sequences used were as follows: 5-GAAUACACGGAAUUCGAGCUCGCCCAUCCUGGAUU GUAAAUUGUAAUGUUGUUGCGCGCGCGCUAUAGUGAGUCGUAUUAGGG-3; biotin, 5-GAATACACGGAATTCGAGCTCGCCCATCCTGGATTGTAAATTGTAATGTTGTT GCGCGCGCGCTATAGTGAGTCGTATTAGGG-3.
Short RNA sequencing.
RNA from A549 cells induced with daIRF3 and overexpressing wild-type SETX, a SETX mutant with no enzymatic activity (ATPase mutant) or a SETX mutant lacking RNA-binding activity was extracted with Trizol and by ethanol precipitation. 1 μg of total RNA was then treated for 30 min at 37 °C with 1 U of Baseline Zero DNase and was cleaned by ethanol precipitation. To facilitate ligation of adapters to capped short transcription start site–associated RNA, 5′ caps were removed by treatment of each sample for 1.5 h at 37 °C with 10 U of tobacco acid pyrophosphotase, followed by reaction cleanup via ethanol precipitation. Next, small RNA libraries were prepared with the NEBNext Small RNA Library Prep Set for Illumina kit, according to the manufacturer's instructions (NEB), except that the NEB NEBNext 3′ SR Adaptor (5′-adenylated, 3′-blocked oligodeoxynucleotide 5′-rAppAGATCGGAAGAGCACACGTCT-NH2-3′) was ligated overnight (∼16 h) at 16 °C. Ligation of the NEBNext 5′ SR Adaptor (5′-rGrUrUrCrAr GrArGrUrUrCrUrArCrArGrUrCrCrGrArCrGrArUrC-3′) was done according to the manufacturer's instructions (NEB). Final PCR amplification and 'barcoding' of libraries (15 cycles) was performed with LongAmp Taq 2X MasterMix and KAPA HiFi HotStart ReadyMix (2X) and was followed by purification with 1.8X of Agencourt AMPure XP beads. After analysis on an Agilent Bioanalyzer (DNA7500 kit) to verify the presence of canonical microRNA and other short RNA peaks, size-selection was performed with 3% gel cassettes on the BluePippin system (Sage Science), with selection for fragments 135–185 nucleotides in length. Size-selected libraries were assessed on the Agilent Bioanalyzer (High Sensitivity DNA chip), which showed a single peak for each sample corresponding to short RNA fragments ranging from 30 nucleotides to 70 nucleotides . Finally, samples were 'multiplexed' and sequenced on a MiSeq platform (1x 50-nucleotide reads) in accordance with the manufacturer's protocols (Illumina).
RNA immunoprecipitation.
Crosslinking was performed for 10 min. Sonication was performed at 4 °C and conditions were optimized to generate nucleotide fragments of approximately 200–500 base pairs. Extracts were pre-cleared for 3 h before immunoprecipitation overnight with control antibody (5415; Cell Signaling) or anti-Flag (M2; SIGMA). After the washing steps, reverse crosslinking was carried out at 42 °C for 1 h and 65 °C for another hour. Immunoprecipitated RNA was then precipitated for 2 h at −80 °C. RNA was treated with Turbo DNase (Life Technologies), followed by reverse transcription and quantitative PCR.
Statistical methods.
The statistical significance of all pairwise comparisons in quantitative PCR assays (change in cycling threshold (ΔCT) values) was determined with a two-tailed Student's t-test under the assumption of equal variances between groups. We did not find significant differences (false-discovery rate, q < 0.05) between contrast groups in Levene's tests of equality of variances, or departures from normality as assessed by Shapiro-Wilk tests.
Accession codes.
GEO: microarray data, GSE52937.
Accession codes
References
Pichlmair, A. & Sousa, C.R.E. Innate recognition of viruses. Immunity 27, 370–383 (2007).
Ward, P.A. New approaches to the study of sepsis. EMBO Mol. Med. 4, 1234–1243 (2012).
Brandes, M., Klauschen, F., Kuchen, S. & Germain, R.N. A systems analysis identifies a feedforward inflammatory circuit leading to lethal influenza infection. Cell 154, 197–212 (2013).
Crow, Y.J. et al. Mutations in genes encoding ribonuclease H2 subunits cause Aicardi-Goutieres syndrome and mimic congenital viral brain infection. Nat. Genet. 38, 910–916 (2006).
Garcia-Sastre, A. & Biron, C.A. Type 1 interferons and the virus-host relationship: a lesson in detente. Science 312, 879–882 (2006).
Hiscott, J. Triggering the innate antiviral response through IRF-3 activation. J. Biol. Chem. 282, 15325–15329 (2007).
Dixit, E. et al. Peroxisomes are signaling platforms for antiviral innate immunity. Cell 141, 668–681 (2010).
Weake, V.M. & Workman, J.L. Inducible gene expression: diverse regulatory mechanisms. Nat. Rev. Genet. 11, 426–437 (2010).
Ramirez-Carrozzi, V.R. et al. A unifying model for the selective regulation of inducible transcription by CpG islands and nucleosome remodeling. Cell 138, 114–128 (2009).
Kouzine, F. et al. Global regulation of promoter melting in naive lymphocytes. Cell 153, 988–999 (2013).
Badeaux, A.I. & Shi, Y. Emerging roles for chromatin as a signal integration and storage platform. Nat. Rev. Mol. Cell Biol. 14, 211–224 (2013).
Cheung, A.C. & Cramer, P. A movie of RNA polymerase II transcription. Cell 149, 1431–1437 (2012).
Buratowski, S. Progression through the RNA polymerase II CTD cycle. Mol. Cell 36, 541–546 (2009).
Lauberth, S.M. et al. H3K4me3 Interactions with TAF3 regulate preinitiation complex assembly and selective gene activation. Cell 152, 1021–1036 (2013).
Bentley, D.L. Rules of engagement: co-transcriptional recruitment of pre-mRNA processing factors. Curr. Opin. Cell Biol. 17, 251–256 (2005).
Kuehner, J.N., Pearson, E.L. & Moore, C. Unravelling the means to an end: RNA polymerase II transcription termination. Nat. Rev. Mol. Cell Biol. 12, 283–294 (2011).
Horng, T. & Medzhitov, R. Transcriptional control of the inflammatory response. Nat. Rev. Immunol. 9, 692–703 (2009).
Marazzi, I. et al. Suppression of the antiviral response by an influenza histone mimic. Nature 483, 428–433 (2012).
Nicodeme, E. et al. Suppression of inflammation by a synthetic histone mimic. Nature 468, 1119–1123 (2010).
Skourti-Stathaki, K., Proudfoot, N.J. & Gromak, N. Human senataxin resolves RNA/DNA hybrids formed at transcriptional pause sites to promote Xrn2-dependent termination. Mol. Cell 42, 794–805 (2011).
Yüce, Ö. & West, S.C. Senataxin, defective in the neurodegenerative disorder ataxia with oculomotor apraxia 2, lies at the interface of transcription and the DNA damage response. Mol. Cell. Biol. 33, 406–417 (2013).
Steinmetz, E.J. et al. Genome-wide distribution of yeast RNA polymerase II and its control by Sen1 helicase. Mol. Cell 24, 735–746 (2006).
Kim, K.Y. & Levin, D.E. Mpk1 MAPK association with the Paf1 complex blocks Sen1-mediated premature transcription termination. Cell 144, 745–756 (2011).
Downes, C.P., Wolf, C.R. & Lane, D.P. in Cellular Responses to Stress (Princeton University Press, 2014).
Suraweera, A. et al. Senataxin, defective in ataxia oculomotor apraxia type 2, is involved in the defense against oxidative DNA damage. J. Cell Biol. 177, 969–979 (2007).
Core, L.J., Waterfall, J.J. & Lis, J.T. Nascent RNA sequencing reveals widespread pausing and divergent initiation at human promoters. Science 322, 1845–1848 (2008).
Rahl, P.B. et al. c-Myc regulates transcriptional pause release. Cell 141, 432–445 (2010).
Chopra, V.S. et al. The polycomb group mutant esc leads to augmented levels of paused Pol II in the Drosophila embryo. Mol. Cell 42, 837–844 (2011).
Hazelbaker, D.Z., Marquardt, S., Wlotzka, W. & Buratowski, S. Kinetic competition between RNA polymerase II and Sen1-dependent transcription termination. Mol. Cell 49, 55–66 (2013).
Porrua, O. & Libri, D. A bacterial-like mechanism for transcription termination by the Sen1p helicase in budding yeast. Nat. Struct. Mol. Biol. 20, 884–891 (2013).
Bleichert, F. & Baserga, S.J. The long unwinding road of RNA helicases. Mol. Cell 27, 339–352 (2007).
Jin, D.J., Burgess, R.R., Richardson, J.P. & Gross, C.A. Termination efficiency at rho-dependent terminators depends on kinetic coupling between RNA polymerase and rho. Proc. Natl. Acad. Sci. USA 89, 1453–1457 (1992).
Henriques, T. et al. Stable pausing by RNA polymerase II provides an opportunity to target and integrate regulatory signals. Mol. Cell 52, 517–528 (2013).
Valen, E. et al. Biogenic mechanisms and utilization of small RNAs derived from human protein-coding genes. Nat. Struct. Mol. Biol. 18, 1075–1082 (2011).
Chen, W.Y. et al. TAF4 coactivator function for E proteins that involves enhanced TFIID binding. Genes Dev. 27, 1596–1609 (2013).
Wright, K.J. & Tjian, R. Wnt signaling targets ETO coactivation domain of TAF4/TFIID in vivo. Proc. Natl. Acad. Sci. USA 106, 55–60 (2009).
Wei, Y. et al. A TAF4-homology domain from the corepressor ETO is a docking platform for positive and negative regulators of transcription. Nat. Struct. Mol. Biol. 14, 653–661 (2007).
Chen, Y.Z. et al. DNA/RNA helicase gene mutations in a form of juvenile amyotrophic lateral sclerosis (ALS4). Am. J. Hum. Genet. 74, 1128–1135 (2004).
Moreira, M.C. et al. Senataxin, the ortholog of a yeast RNA helicase, is mutant in ataxia-ocular apraxia 2. Nat. Genet. 36, 225–227 (2004).
Becherel, O.J. et al. Senataxin plays an essential role with DNA damage response proteins in meiotic recombination and gene silencing. PLoS Genet. 9, e1003435 (2013).
Ginno, P.A., Lott, P.L., Christensen, H.C., Korf, I. & Chedin, F. R-loop formation is a distinctive characteristic of unmethylated human CpG island promoters. Mol. Cell 45, 814–825 (2012).
Lee, T.I. & Young, R.A. Transcriptional regulation and its misregulation in disease. Cell 152, 1237–1251 (2013).
Dunah, A.W. et al. Sp1 and TAFII130 transcriptional activity disrupted in early Huntington′s disease. Science 296, 2238–2243 (2002).
Rice, G. et al. Clinical and molecular phenotype of Aicardi-Goutieres syndrome. Am. J. Hum. Genet. 81, 713–725 (2007).
Huang, D.W., Sherman, B.T. & Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44–57 (2009).
Langmead, B., Trapnell, C., Pop, M. & Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 10, R25 (2009).
Trapnell, C., Pachter, L. & Salzberg, S.L. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25, 1105–1111 (2009).
Robinson, M.D. & Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 11, R25 (2010).
Robinson, M.D., McCarthy, D.J. & Smyth, G.K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
Rozen, F. et al. Bidirectional RNA helicase activity of eucaryotic translation initiation factors 4A and 4F. Mol. Cell. Biol. 10, 1134–1144 (1990).
Acknowledgements
We thank J. Lim (Icahn School of Medicine at Mount Sinai) for Kunjin virus; M. Evans (Icahn School of Medicine at Mount Sinai) for Gag-Pol; P. Palese (Icahn School of Medicine at Mount Sinai) for comments, anti-PB2 and unpublished observations; A. Telenti for comments; A. García-Sastre (Icahn School of Medicine at Mount Sinai) for comments, RIG-I deficient A549 cells and unpublished observations; R. Roeder (Rockefeller University) for TAF4 reagents; M.F. Lavin (The University of Queensland) for anti-SETX; and the Genomic facility at Icahn and the Proteomics facilities at Rockefeller University. Supported by the computational resources and staff of the Department of Scientific Computing at the Icahn School of Medicine at Mount Sinai; the Canadian Institutes of Health Research (M.S.M.); the US National Institutes of Health (U19AI106754 to I.M.); and the National Institute of Allergy and Infectious Diseases Center of Excellence for Influenza Research and Surveillance (HHSN272201400008C to I.M. and H.v.B.).
Author information
Authors and Affiliations
Contributions
M.S.M., L.M.-G., A.M.M. and A.F.-S. performed experiments with reporters and viruses; M.S.M., P.M., J.M., O.J.B. and M.F.L. performed experiments involving mice; A.R., M.T., N.P.M., Z.P., J.N., C.M. and I.M. performed next-generation sequencing and biochemical experiments; J.S.Y.H. and H.v.B. performed data analysis for next-generation sequencing experiments; C.F.B. and A.R.F. performed fractionation experiments; A.M.M., S.H., C.B., J.A.T.Y., A.F.-S., O.J.B. and M.F.L. provided materials and insights in experimental procedures; and H.v.B. and I.M. supervised the project and wrote the paper with feedback from coauthors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Control experiments for Figure 1.
(a) Localization of SETX in untreated or PR8ΔNS1 virus infected A549 cells (4hours). Nuclear (DAPI) and Tubulin staining are shown. SETX antibody specificity was validated using human fibroblast cells proficient (WT) or deficient for SETX (SETX deficient). Shown are representative images from one of two independent experiments. Scale bars represent 50μm. (b) SETX co-elutes with high molecular weight complexes. Lysates from uninfected or PR8ΔNS1 infected A549 cells were subjected to glycerol gradient sedimentation and the presence of SETX, RNAPII or the Influenza A RNA polymerase PB2 subunit in sequential fractions revealed through immunoblotting. LARP7 is a control for elution in high molecular weight complexes. (c). Co-immunoprecipitation of RNAPII and SETX in the absence or presence of RNaseA. (d). Western blot analysis of SETX depletion in siSETX cells relative to siCtrl and untreated (No Si) condition prior to and after infection with PR8ΔNS1 virus. (e) The expression levels of 12 housekeeping genes were determined by microarray analysis of un-transfected (No Si), siSETX- or siCtrl- transfected cells under basal conditions (without infection). Results for individual probesets are shown for genes that are represented by multiple probesets on the microarray. A full list of genes and their expression in the experimental condition tested can also be found in Supplementary Table 1. (f) Western blot analysis of XRN2 depletion in siXRN2-treated cells relative to siCtrl and untreated (No Si) conditions prior to and after infection with PR8ΔNS1 virus. (g) Heatmap showing differentially expressed genes (Fold change relative to siCtrl >1.5, p < 0.01; ANOVA with post-hoc TUKEY test) in un-transfected cells (No si) or XRN2 siRNA (siXRN2) transfected cells as compared to control non-targeting siRNA transfected cells (siCtrl) at an early (4 hours) time point post-infection. (h). The expression levels of 12 housekeeping genes were determined by microarray analysis of un-transfected (No Si), siXRN2 or siCtrl transfected cells under basal conditions (without infection). Results for individual probesets are shown for genes that are represented by multiple probesets on the microarray. A full list of genes and their expression in the experimental condition tested can also be found in Supplementary Table 2. (i). Protein levels of SETX and phosphorylated IRF3 (pIRF3) were measured in siCtrl and siSETX cells left untreated or infected with PR8∆NS1 virus. Tubulin protein levels were used as a loading control. (j and k) Expression of virus-derived messenger RNA (j), and viral genomic RNA (k) from SETX siRNA transfected (siSETX, white bars), control siRNA transfected (siCtrl, Gray bars) and un-transfected (No si, black bars) cells infected for 4 hours with the PR8∆NS1 virus. Data shown are from three experiments (e-f, g-h and j-k; with the mean and s.d shown for j-k). A representative blot of two independent experiments (b and c) or blots of pooled extracts from two independent experiments are shown (d, f).
Supplementary Figure 2 GRO-seq controls.
(a) Meta-analysis of GRO-seq signals across all RefSeq genes >1kb in A549 cells upon infection with influenza PR8ΔNS1, before (dashed lines) and 4 hours after infection (solid lines) in siCtrl (blue) and siSETX (red) treated cells. Y axis indicates the geometric mean coverage per 5 million reads. (b) Travelling ratios (TR) of all active genes for the conditions indicated in (a). TRs were calculated as the average number of tags per bp in the promoter-proximal region (30 to +300 bp around TSS) divided by that on the gene body (+300 bp to the end of the gene). The number of active genes identified in each condition is indicated between brackets. (c) GRO-seq read coverage for the RPL10 gene, showing no changes in siCtrl over siSETX conditions. Y-axis indicates base coverage per 5 million reads. (d) Overview of the fraction of GRO-seq reads mapping to genic (exons, introns and antisense) and intergenic regions. (e) Expression of IFNB1 and IFIT1 mRNA levels in siSETX and siCtrl A549 cells in the presence of flavopiridol (blue bars), DMSO vehicle control (red bars), or untreated control cells (gray bars). Values represent the mean and standard deviation of three independent experiments; Asterisks (*and **) indicate t-test p-values < 0.05 and < 0.005, respectively.
Supplementary Figure 3 SETX has ATP-dependent helicase activity and affects the induction of transcription start site-associated short RNA at gene promoters.
(a) Agilent bioanalyzer gel-like image of a helicase assay, showing a decrease in relative intensity of the DNA/RNA hybrid band (top) vs. the dissociated simplex band (bottom), as a function of the concentration of SETX. Samples were analyzed using the RNA 6000 Pico Kit. Hybrid-only and RNase treated lanes are shown as controls. (b) Log2 ratio of quantified simplex/hybrid signals (time-corrected peak area) as shown in (a). (c). Same as in a., with NS3 Helicase from hepatitis C virus, wild type SETX (wt), SETX ATPase mutant (ATP MUT) and RNA-binding Mutant (RNA MUT). (d) Log2 ratios of quantified simplex/hybrid signals (time-corrected peak area) as shown in (c). (e) Colloidal Coomassie staining of 3-8% Tris-Acetate gel of wildtype SETX (WT), ATPase (ATP MUT) RNA binding mutant (RNA MUT) purified from HEK 293 cells and used in (a) and (c), as indicated. Co-precipitation of exosome components with SETX (Supplementary. Fig.4 and Supplementary Table 4) does not exclude the possibility that wildtype SETX activity could be modulated by residual exosome activity. (f) Overview of whole-genome short-RNA-Seq data (30-70nt fragments) generated for cells overexpressing wildtype (WT) or mutant SETX (ATPase mutant: MUT1 and RNA binding mutant: MUT2) induced with daIRF3. Raw reads were first mapped against a reference set of sn(o)RNA, tRNA and rRNA transcripts that are typically found in the same size range, and the remainder were mapped against the hg19 human reference genome using Bowtie (v0.12.9). Reads mapping to more than one location in the genome were discarded from analysis. TSS-proximal reads were defined as reads mapping within 300 bp of a transcription start site (GENCODE v18). The table lists the read counts obtained at each stage of data processing. (g) Scatterplots comparing normalized TSS-proximal short RNA-Seq read counts (sense strand only) in cells over-expressing either WT or mutant SETX, after induction with daIRF3. Correlation coefficients (r) are indicated for each comparison. (h) Short RNA-Seq profiles (blue) of antiviral induced genes IFIT1, IFIT2 and IFIT3 (top) and GAPDH and ACTG1 controls (bottom). Short-read track scales correspond to the normalized read coverage per 25,000 uniquely mapped genomic reads in each dataset. Promoter-proximal TSS areas are highlighted in grey. No mapped reads were found at the IFNB1 locus in the SETX mutant or WT profiles (data not shown), presumably due to limitations in read depth. Top tracks (black) indicate the frequency of initiation site usage across each gene, derived from 5’-end RNA-sequencing tags of capped RNAs >300 nt obtained from the same cells. Short-read tags align exactly to the preferred transcription initiation site of each gene.
Supplementary Figure 4 Interaction between SETX and TAF4.
(a) Validation of the IP-MS/MS via immunoprecipitation of SETX and immunoblotting for the indicated endogenous proteins in cells expressing Flag-tagged SETX. Input (Inp) and the IgG control IP are shown. (b) Association of TAF4 with SETX. FLAG tagged wildtype SETX and the SETX ATPase (M1) and RNA binding mutants (M2) were immobilized on FLAG-beads and probed with cell extracts (prepared via Dignam-Roeder method). After 4h binding in BC100 (20 mM Tris–HCl pH 7.9, 20% glycerol, 0.1 mM EDTA, 150 mM KCl), 4 washes in BC150 + 0.1% NP40 in the presence of absence of RNAse, the immunoprecipitated (IP) material was subjected to western blotting with the anti-Flag antibody (to control for equal levels of SETX protein bound to beads) and the TAF4 antibody (detecting endogenous TAF4). UT: untreated control; EV: empty vector control (c) Association of SETX with TAF4. As in (b) but with immobilized wild type TAF4 (WT) and TAF4 ΔTAFH mutant (Δ). After 4h of binding in BC100 followed by 4 washes in a BC150 + 0.1% NP40, the IP material was subjected to western blotting with the anti-Flag antibody (to control for equal level of TAF4 proteins on beads) and SETX antibody (detecting endogenous SETX). For a-c, representative blots of pooled extracts from biological replicates or two independent experiments are shown.
Supplementary Figure 5 SETX deficiency inhibits viral biogenesis.
(a and b). Growth of influenza PR8 (a) and PR8ΔNS1 (b) viruses in A549 cells untreated or treated with control non-targeting siRNA (siCtrl) or SETX-specific siRNA (siSETX). Cells that were not siRNA treated (No si) were also included as controls. Viral titers were determined by plaque assays on MDCK cells (for PR8) or MDCK cells stably expressing NS1 (for PR8ΔNS1). (c) Growth of influenza PR8 virus in siSETX-, siCtrl-treated or untreated (No si) A549 cells that stably express a shRNA targeting DDX58 (RIGI). (d) Western blot showing reconstitution of SETX-deficient cells with FLAG-tagged wildtype SETX and the SETX ATPase mutant (Mut). (e) Quantification of secreted cytokine levels in SETX-deficient and control human fibroblasts 4 hours post infection with PR8ΔNS1. Note that although amounts of IP-10 levels are easily detectable via ELISA, IFNA2 levels were much lower and could result from an IFNB1-dependent induction of IFNA2. (f) Growth of influenza PR8 virus in SETX-deficient and WT human fibroblasts. Viral titers were determined by plaque assay on MDCK cells. Note that final viral titers are similar to A549 cells but the kinetics of viral growth differs between the two cell types. (g) Expression of IFNB1, IFIT1, CXCL9 and SETX in SETX-deficient and wild-type control fibroblasts 4 hours post-infection with West Nile Virus (Kunjin strain; WNV KUNV). **P < 0.005 (t-test). (h) KUNV growth in wildtype and SETX-deficient primary human fibroblast cells. Viral titers were determined by focus-forming assay on Vero cells. (i). Western blot of SETX, RIG-I, IFIT1 and Tubulin proteins isolated from human glial cells after 0, 4, 12 and 24 hours post-infection with PR8∆NS1. Data are from three independent experiments (mean and s.e.m in a-b and f; s.d in c, e, g and h). Representative western blots are shown in d and i. (j) Model of RNAPII- and activator-dependent recruitment of SETX at antiviral loci during infection. Promoter proximal termination is elicited via SETX translocation over nascent transcript as a result of sequence-specific RNA recognition (upper panel) or via helicase activity and R-loop resolution as a result of structure-specific recognition of RNA/DNA hybrids at promoters (lower panel).
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–5 (PDF 3219 kb)
Supplementary Table 1
List of probesets that are affected by siSETX treatment in uninfected cells. (XLS 238 kb)
Supplementary Table 2
List of probesets that are affected by siXRN2 treatment in A549 cells in uninfected cells (XLSX 18 kb)
Supplementary Table 3
List of genes with significantly changed levels of engaged RNAPII in human A549 cells after 4 hours of infection with influenza PR8ΔNS1. (XLS 293 kb)
Supplementary Table 4
Shared Proteins (XLSX 32 kb)
Rights and permissions
About this article

Cite this article
Miller, M., Rialdi, A., Ho, J. et al. Senataxin suppresses the antiviral transcriptional response and controls viral biogenesis. Nat Immunol 16, 485–494 (2015). https://doi.org/10.1038/ni.3132
Received:
Accepted:
Published:
Issue date:
DOI: https://doi.org/10.1038/ni.3132
This article is cited by
-
Senataxin helicase, the causal gene defect in ALS4, is a significant modifier of C9orf72 ALS G4C2 and arginine-containing dipeptide repeat toxicity
Acta Neuropathologica Communications (2023)
-
Hypoxia-induced SETX links replication stress with the unfolded protein response
Nature Communications (2021)
-
Mitochondria-localised ZNFX1 functions as a dsRNA sensor to initiate antiviral responses through MAVS
Nature Cell Biology (2019)
-
Disruption of Spermatogenesis and Infertility in Ataxia with Oculomotor Apraxia Type 2 (AOA2)
The Cerebellum (2019)
-
Chromatin dependencies in cancer and inflammation
Nature Reviews Molecular Cell Biology (2018)
