
Abstract
The menopausal transition (MT) is associated with an increased risk for many disorders including neurological and mental disorders. Brain imaging studies in living humans show changes in brain metabolism and structure that may contribute to the MT-associated brain disease risk. Although deficits in ovarian hormones have been implicated, cellular and molecular studies of the brain undergoing MT are currently lacking, mostly due to a difficulty in studying MT in postmortem human brain. To enable this research, we explored 40 candidate biomarkers for menopausal status in 42 pre-, peri-, and post-menopausal subjects across three postmortem tissues: blood, the hypothalamus, and pituitary gland. We identified fourteen significant and seven strongest menopausal biomarkers across the three tissues. Using these biomarkers, we generated multi-tissue and tissue-specific composite measures that allow the postmortem identification of the menopausal status across different age ranges, including the “perimenopausal”, 45–55-year-old group. Our findings enable the study of cellular and molecular mechanisms underlying increased neuropsychiatric risk during the MT, opening the path for hormone status-informed, precision medicine approach in women’s mental health.
Similar content being viewed by others
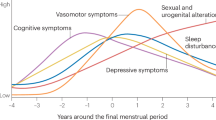
The menopausal transition (MT) is the midlife transition period, typically lasting 2–8 years [1], which extends from the point when menstrual periods become irregular (early MT) until the final menstrual period (FMP) [2] (Fig. 1A). Menstrual changes reflect changes in the function of the hypothalamic-pituitary-gonadal (HPG) axis (Fig. 1B) due to reproductive ovarian aging and declining ovarian follicle reserve. The entire MT is marked by the low antral follicle counts and low levels of the ovarian reserve marker anti-Müllerian hormone (AMH) [3, 4] (Fig. 1B). The levels of ovarian hormones estradiol and progesterone decrease, although this is also a period with the most extreme hormone fluctuations, particularly during the late MT [2]. To compensate for decreased ovarian function, the secretion of follicle stimulating hormone (FSH) from the pituitary (Fig. 1B) is typically elevated and stays stably elevated postmenopause together with low ovarian hormone levels [5]. The hallmarks of MT and menopause are thus low/non-detectable AMH, low estradiol, and elevated FSH, and while cutoff values for blood AMH and FSH levels have been suggested [2, 3], these measures are not standardized as clinical menopausal biomarkers [2].

A The menopausal transition is defined as the time between the beginning of cycle irregularity and the final menstrual period (FMP). One year after the FMP marks menopause, and following this point an individual is considered postmenopausal. Perimenopause includes the entire menopausal transition and one year post-FMP. B Release of reproductive hormones is controlled by the HPG axis. GnRH is released from the hypothalamus into the hypophyseal portal system, signaling release of LH and FSH from the pituitary gland. From the pituitary gland, LH and FSH travel through the periphery until they reach the ovaries, which stimulates the release of estradiol and progesterone. Estradiol and progesterone provide feedback inhibition to the hypothalamus and pituitary gland to prevent release of GnRH, LH, and FSH. Within the ovary, AMH is produced by growing follicles, and serum AMH levels are an indicator of functional ovarian reserve. C Fourteen of the seventeen key steroids involved in sex steroid synthesis were analyzed in the present study (blue; primary ovarian steroids in pink). AMH Anti-Müllerian hormone, DHEA Dehydroepiandrosterone, DHT Dihydrotestosterone, DOC Deoxycorticosterone, FSH Follicle-stimulating hormone, GnRH Gonadotropin-releasing hormone, LH Luteinizing hormone.
Clinically, spontaneous menopause is diagnosed 12 months after the FMP, at the mean age of approximately 51 years [6], ranging from 49.9–52.7 years, depending on the country and ethnicity [7,8,9,10]. The period of MT together with 12 months following the FMP is also known as perimenopause (Fig. 1A). Globally, >850 million women are aged 40–60 years and ~90% of these women will transition through perimenopause within the age range of 40–58 years [6]. In addition, a subset of people with ovaries experience menopause earlier than the normative age range, and this includes premature menopause (younger than 40 years) or induced menopause that can result from medical treatments like chemotherapy or surgical removal of the ovaries in which case the MT is more abrupt [11]. In general, the lack of physiologically meaningful, reliable, and objective criteria for classifying women into menopausal status categories has been impeding progress in understanding the effect of reproductive aging and MT on health and disease [12], particularly with regards to examining postmortem human brain tissue.
While MT represents a natural life transition, it is associated with symptoms that can significantly affect quality of life in up to 90% of women and other people with ovaries, including hot flashes, sleep disturbances, depressed mood, increased anxiety, and trouble concentrating [10]. Specifically, the MT is an important window of vulnerability for the development of mental disorders including mood and psychotic disorders. Studies showed a 2–5 fold increased risk for major depressive disorder (MDD) during the MT, compared with both late premenopause and several years postmenopause [13]. Similarly, women undergoing the MT exhibit a heightened risk for first-onset psychosis and reemergence or exacerbation of symptoms of schizophrenia and related disorders, which are not observed in men of similar age [14].
Although evidence suggests that hormonal changes play a critical role in the manifestation of mood and psychotic symptoms during the MT, the biological mechanisms are poorly understood [13,14,15,16]. Studies in living humans show changes in brain metabolism and brain structure that may contribute to MT-associated psychiatric risk [17]. However, studies on cellular and molecular changes in the human brain during this period are currently lacking. Specifically, working with postmortem human brain tissue represents a particular challenge, as the major US brain banks do not typically have information regarding an individual’s reproductive status. While chronological age at death is informative, it does not represent a good proxy for the menopausal status, which can obscure the results of the investigations focused on the MT’s effects on the brain.
To enable cellular and molecular studies of MT in humans, here we characterize 40 candidate biomarkers in our cohort of 42 individuals across three postmortem tissues – blood, the hypothalamus, and pituitary gland. Our study identifies the strongest menopausal biomarkers across the three tissues and establishes a menopausal component score that allows for the postmortem determination of the menopausal status of any given individual, across a wide age range including the most challenging “perimenopausal” group, within 45–55 years of age. To make this analysis more cost-efficient, feasible, and adaptable for other researchers, we also offer alternative tissue-specific component scores for the reproductive status determination.
Results
Here, we analyzed 40 candidate menopausal biomarkers including: i) a panel of 14 different steroid hormones (Fig. 1C) in blood and the hypothalamus; ii) AMH in blood; iii) FSH protein levels in blood and pituitary gland; iv) gene expression of reproduction-relevant genes in the hypothalamus (CYP19A1, ESR1, ESR2, GPER1, PGR, KISS1) and pituitary gland (FSH, ESR1, GNRHR). Our cohort included 42 individuals whose postmortem tissue samples (blood, the hypothalamus, and pituitary gland) were received from the Human Brain Collection Core (HBCC) of the National Institute of Mental Health - Intramural Research Program. We originally classified the subjects based on their “expected menopausal status”, simply using their chronological age as follows: <40 years (“pre-menopausal”, N = 10), 45–55 years (“peri-menopausal”, N = 21), and > 55 years (“post-menopausal”, N = 11) (Table 1). While our target was the “perimenopausal” group, which is likely to show variability in the menopausal age of onset, we included younger (<40 years) and older (>55 years) groups to define pre- and post-menopausal reference values, respectively, for biomarkers. For the majority of the subjects older than 55 years (9 out of 11), beyond the likelihood based on age, we had reasonable evidence that these subjects were indeed postmenopausal (see Methods). Notably, our cohort also included individuals with MDD or depressive symptoms (N = 18), in addition to individuals with anxiety symptoms or substance use (Table 1). Education level, body mass index (BMI), post-mortem interval (PMI), and RNA integrity number (RIN) for the hypothalamus and pituitary gland were comparable across all three groups (Table 1).
Biological measurements
A total of 40 analytes related to the HPG axis and steroidogenesis were measured in the blood, hypothalamus, and pituitary glands of the 42 samples (Table 2). To identify strongest menopausal biomarker candidates, we initially did statistical analysis on the three groups based on age: pre-, peri, and post-menopausal groups (Fig. 1A). Of the 40 markers, 14 had significantly different measurements between groups determined by non-parametric Kruskal-Wallis test. All 14 of these significant effects were driven by differences between the pre- and post-menopausal groups as determined by posthoc Dunn’s test (Table 2). Perhaps not surprisingly, in blood, we found significant differences between pre- and post-menopausal groups in AMH (p < 0.001), FSH (p < 0.001), estrone (p = 0.009), estradiol (p < 0.001), progesterone (p = 0.011), and DHT (p = 0.021). In the pituitary gland, three markers showed significant changes from pre- to post-menopausal groups including FSH protein levels (p = 0.002) and gene expression levels of FSH (p < 0.001) and GNRHR (p = 0.049). Finally, we found differences in the levels of four hypothalamic steroids between pre- and post-menopausal groups including DHEA (p = 0.005), estrone (p = 0.003), estradiol (p = 0.023), and progesterone (p = 0.042), and in the hypothalamic expression of the gene for the enzyme aromatase, CYP19A1 (p = 0.038), responsible for estradiol synthesis.
Steroid hormones
All three of the major ovarian hormones – estrone (Fig. 2A), estradiol (Fig. 2B), and progesterone (Fig. 2C) – were significantly lower in the postmenopausal group than the premenopausal group in both the blood and hypothalamic tissue. Because of potential issues with accessing blood from different postmortem tissue/brain repositories, we additionally wished to gauge whether the levels of these steroids within the hypothalamus correlated highly with those within the blood. Estrone levels had a very high correlation between the two tissue types (Fig. 2A-iii; r = 0.95, p < 0.001), and a moderate correlation was also observed for both estradiol (Fig. 2B-iii; r = 0.44, p = 0.007) and progesterone (Fig. 2C-iii; r = 0.44, p = 0.006). Moderate to very strong correlations were also observed between blood and hypothalamus measurements of 9 of the other 11 steroids analyzed (Supplementary Fig. 1). The only two steroids that did not have a strong positive correlation were DHT and testosterone, which are likely explained by technical limitations; DHT levels within the hypothalamus were below the limit of quantitation for every sample, and 6 samples were excluded from testosterone measurement due to technical issues with sample processing. Therefore, quantifying steroid hormones extracted from hypothalamic brain tissue could represent a proxy for steroid hormone levels in the absence of access to blood.

A Estrone, B estradiol, and C progesterone measurements are all lower in both i) blood and ii) hypothalamus tissue extracts following menopause. iii) Steroid measurements in the two tissue types had a high correlation for estrone and a moderate correlation for both estradiol and progesterone. Y-axis breaks indicate an adjustment in scale for display of individual value extremes. p values above box plots indicate significance of Dunn’s test posthoc comparisons following non-parametric Kruskal-Wallis test. Box plots (box, 1st–3rd quartile; horizontal line, median; whiskers, 1.5xIQR).
In addition to the steroid levels, hypothalamic CYP19A1 gene expression was also significantly lower in the postmenopausal group compared to the premenopausal group (p = 0.038, Supplementary Fig. 2A). Notably, hypothalamic CYP19A1 expression showed no correlation with blood estradiol levels (R = −0.02, p = 0.896; Supplementary Fig. 2B) yet had a strong positive correlation with hypothalamic estradiol levels (R = 0.5, p < 0.001; Supplementary Fig. 2C). This result indicates that the decrease in hypothalamic estradiol observed from pre- to post-menopause (Fig. 2B) is not only a proxy observation of decreased peripheral estradiol levels but also reflects reduced levels of locally synthesized estradiol.
Glycoproteins
AMH and FSH are often measured in clinical settings to assist with determining reproductive status [2]. Within our cohort of postmortem samples, there was a significant decrease in AMH (Fig. 3A) from the premenopausal group to the postmenopausal group (p < 0.001). Notably, although we measured AMH in frozen postmortem blood samples, our results were comparable to the levels typically reported from the serum of living subjects [3]. In living humans, it was estimated that an individual with serum AMH < 10 pg/ml would likely undergo the FMP within the next 12 months, with probability ranging with age, from 51% at < 48 years old and 79% at ≥ 51 years [3]. On the contrary, the likelihood of a person with AMH > 100 pg/ml not reaching the FMP within the next 12 months was between 97% in those < 48 years old and 90% in those ≥ 51 years old [3].

A AMH in the blood is significantly reduced from premenopause to perimenopause and postmenopause, and is nearly undetectable in the postmenopause group. AMH measurements are displayed on a logarithmic scale. B FSH protein levels in the blood are significantly higher in the perimenopause and postmenopause groups when compared to premenopausal levels. p values above box plots indicate significance of Dunn’s test posthoc comparisons following non-parametric Kruskal-Wallis test. Box plots (box, 1st–3rd quartile; horizontal line, median; whiskers, 1.5xIQR); AMH Anti- Müllerian hormone, FSH Follicle-stimulating hormone.
In our cohort, AMH was below 10 pg/mL in 8 of the 11 postmenopausal samples (>55 years old), indicative of a high likelihood of having reached the FMP [3], with 5 samples having AMH levels below the limit of quantitation indicating a completely depleted ovarian reserve (Supplementary Table 1). 3 postmenopausal samples had AMH levels slightly higher than expected (10–20 pg/ml). Conversely, every premenopausal sample (<40 years) had AMH levels > 100 pg/mL, indicating a very low chance that the FMP would occur within 12 months of the time of measurement [3] (Supplementary Table 1). Thus, our results in premenopausal (<40 years) and postmenopausal (>55 years) groups align with expectations based on clinical observations in living humans of comparable age [3] and based on our medical records, with the exception of 3 postmenopausal samples. Within the perimenopausal group, 11 of the 20 samples had AMH levels below 10 pg/mL (three unquantifiable), indicating that the FMP was likely reached (Supplementary Table 1). The remaining nine measurements were all within the range of 10–100 pg/mL (Supplementary Table 1); these levels provide a low confidence that the FMP has occurred, particularly within individuals younger than 51 years of age [3], but are not well-defined based on the current clinical standards. Thus, while typically providing a good indicator of the reproductive status, AMH measurements alone are not sufficient to characterize perimenopausal samples as pre- or post-menopausal like within our dataset.
There was an inverse effect of MT on blood FSH levels, with a significant increase being observed between pre- and post-menopausal groups (p < 0.001, Fig. 3B). Our measurements once again align with clinical observations [2]. Based on current standards in the field, FSH levels greater than 25 IU/L in a random blood draw are characteristic of being in late transition. FSH levels typically continue to increase approximately 2 years after the FMP, after which they are expected to stabilize. In our study, all premenopausal samples had FSH measurements < 15 mIU/mL and 9 out of 11 postmenopausal samples measured > 25 mIU/mL (Supplementary Table 1). In our “perimenopausal”, 45–55-year-old group, 13 subjects had FSH levels < 25 mIU/mL and 7 had levels > 25 mIU/mL (Supplementary Table 1). Although FSH is often monitored in a clinical setting to help determine menopausal status, it alone is insufficient to diagnose menopause due to extreme variability during the MT, as well as susceptibility to factors such as hormonal contraceptives, hormone replacement therapy, and obesity [5, 18]. Therefore, while aligning well with clinical observations, similar to AMH, FSH levels are insufficient to characterize the reproductive status of perimenopausal samples as pre- or post-menopusal-like within our dataset.
Pituitary gland
As a central component of the HPG axis, FSH production within the pituitary gland is likely significantly impacted across the menopausal transition, although this has not been addressed previously. Here we show that both FSH protein levels (p = 0.002, Fig. 4A) and FSH gene expression (p < 0.001, Fig. 4B) within the pituitary gland were significantly higher in the postmenopausal group compared to the premenopausal group, which aligns with the overall expected increase in FSH production and observed increase in blood FSH in the postmenopausal group (Fig. 3B). Additionally, expression of the gene for the GnRH receptor (GNRHR), which would trigger FSH production and release in response to GnRH released from the hypothalamus (Fig. 1B), was also higher in the postmenopausal group (p = 0.049, Fig. 4C). Once again, in the event that blood is not available to measure FSH, we wished to determine whether the pituitary gland could be an appropriate proxy for measuring FSH. Similar to the steroids, there was a significant moderate correlation between blood FSH and pituitary FSH (Fig. 4D; r = 0.42, p = 0.007). Finally, we also confirmed that FSH protein and FSH gene expression within the pituitary had a significant moderate correlation (Fig. 4E; r = 0.57, p < 0.001). This data shows that the pituitary gland can be used as an alternative tissue source for determining menopausal status postmortem.

A FSH protein levels and B FSH gene expression within the pituitary gland are both significantly increased in the postmenopause group compared to the premenopause group. C GNRHR gene expression within the pituitary gland is higher in the postmenopause group than the premenopause and perimenopause groups. There is a moderate correlation between D blood FSH protein and pituitary gland FSH protein measurements, and E FSH protein measurements and FSH gene expression within the pituitary gland. p values above box plots indicate significance of Dunn’s test posthoc comparisons following non-parametric Kruskal-Wallis test. Box plots (box, 1st–3rd quartile; horizontal line, median; whiskers, 1.5xIQR); FSH Follicle-stimulating hormone (protein), FSH Follicle-stimulating hormone (gene), GNRHR Gonadotropin-releasing hormone receptor (gene).
Composite measurement
Although we discovered 14 biological measurements that significantly differed between the premenopausal and postmenopausal groups (Table 2), with the possible exception of AMH, each measure showed some overlap between pre- and post-menopausal individuals (Figs. 2–4). Therefore, none of the candidate biomarkers per se would be sufficient to determine menopausal status and our next goal was to incorporate these markers into a single composite measure which we predict to be more informative than any of the single measurements. In this way, “perimenopausal samples” in the 45–55-year old group, which are the most challenging to categorize based on age alone (no information whether the FMP had been reached was available), could have the composite score used to be characterized as statistically and biologically more similar to a “premenopause-like” or “postmenopause-like” group. In order to achieve this, principal component analysis (PCA) was performed to integrate biological measurements of interest into a single component score. PCA is a statistical procedure that identifies groups of variables according to degrees of association, and reduces each associated group into a new variable – so called “component.” Thus, this approach can be used to reduce correlating variables into a single component score to be used as our composite measure.
Principal component analysis (PCA)
The first step of PCA was to examine the linearity and correlation between each of the 14 individual markers (Supplementary Fig. 3). Of the 91 comparisons drawn between each of the 14 markers, 37 comparisons had a significant correlation (p < 0.05), and each of the 14 markers correlated with at least one other marker. Parallel analysis was conducted to determine the maximum number of dimensions to be extracted from the model (Supplementary Fig. 4A). Parallel analysis compares the sample eigenvalues (the amount of variance carried in each component) of the markers with the 95th percentile of eigenvalues across multiple randomly generated datasets with the same number of uncorrelated factors. Components with sample eigenvalues being higher than those of the randomly generated data can be extracted. Parallel analysis results revealed that the 14 markers were best reduced to three components, as determined by the sample eigenvalue being higher than that of the randomly generated data at up to three dimensions (Supplementary Fig. 4A). This model would account for approximately 67% of the variance in the data (Supplementary Fig. 4A). Most of the biological measures were appropriately categorized into at least one component, as demonstrated by a factor loading of > 0.3 [19, 20] (Supplementary Fig. 4B).
Notably, this initial model using all 14 significant biological measures included four markers that did not have a strong loading into any of the three components (FSH in the blood and pituitary, and progesterone and DHEA in the hypothalamus). This may indicate that the dimension reduction by PCA may be being muddied by extraneous variables that are not immediately relevant to the model. Additionally, the grouping of biological measures into three components did not meet our needs in order to assign each sample a single composite score. Ignoring all but the first component would not have been appropriate, as the first dimension only accounted for approximately 37% of the variance within the dataset, and many biological measures with strong relevance to menopausal status had a low factor loading with the first component (e.g. blood AMH, FSH, estradiol, and progesterone).
Thus, we sought to perform a new analysis including only the “strongest” markers that were selected based on the following criteria: i) they had established relevance to menopausal status (AMH, FSH, estradiol, and progesterone within the blood), and ii) they were correlated with the aforementioned established biologically relevant measures (FSH and FSH gene expression in the pituitary, and estradiol within the hypothalamus) (Figs. 2 and 4; Supplementary Fig. 3). In addition, as a follow up to non-parametric Kruskal-Wallis test, we performed a robust version of ANOVA and pairwise comparisons to insulate against effects of nonnormality and heterogeneity of variance [21], so passing significance in this test was an additional requirement for the inclusion in the model (Supplementary Table 2). This led to a final model with seven biological markers for characterizing menopausal status (Figs. 5–6). Of the 21 comparisons made between these seven markers, 15 comparisons had a significant correlation (p < 0.05), and every individual marker correlated with at least three others (Fig. 5). This suggested a strong model with highly correlated factors. This suggestion was supported by parallel analysis, which revealed that the data was best reduced to a single dimension that accounted for approximately 47% of the variance within the dataset (Fig. 6A). All markers had factor loading > 0.3 in magnitude, indicating that each factor had a moderate correlation with the final component score (Fig. 6B).

The top-left/bottom-right diagonal represents distribution of each measurement, demonstrating a non-normal distribution for all seven markers. Below the diagonal are scatterplots of each pair of markers with the linear regression lines imposed. Above the diagonal are Pearson’s correlation coefficients for each marker with each other. *p < 0.05, **p < 0.01, ***p < 0.001 AMH Anti- Müllerian hormone, FSH Follicle-stimulating hormone (protein), FSH Follicle-stimulating hormone (gene).

A The seven selected biological markers are most appropriately combined into a single component, as represented by the observed Eigenvalue (light blue line) being higher than the simulated Eigenvalue (dark blue line) only at the first component. This single component accounts for approximately 47% of variance in the data (dashed red line). B All seven biological measures had at least moderate correlation with the final component score, as demonstrated by the absolute value of factor loadings falling between 0.3–0.5. A positive factor loading indicates that as this measure increases the component score increases, and a negative factor loading indicates that as this measure decreases the component score increases. C Plot of chronological age vs. component score. All samples in the premenopause group had a component score of < −0.6, and all samples within the postmenopause group had a component score of > −0.4. A cutoff value to classify samples in the perimenopause group was set halfway between these two limits at −0.5 (dashed line). AMH Anti- Müllerian hormone, E2 Estradiol, FSH Follicle-stimulating hormone (protein), FSH Follicle-stimulating hormone (gene), P4 Progesterone. Red dots represent individuals that were on exogenous hormone treatment at some point of time, as per medical records (See Supplementary Table 1 for more information); note that this information is not available for all subjects.
Finally, the component score was calculated for each sample in order to categorize them into one of two groups – “pre-menopausal like” or “post-menopausal like” groups. Calculation of the component score revealed that the premenopausal and postmenopausal samples created two distinct groups, with every premenopausal sample having a component score < −0.6 and every postmenopausal sample having a component score > −0.4 (Fig. 6C). Thus, based on the measurements obtained in our current cohort, the cutoff value of −0.5 can be used to categorize the 18 perimenopausal samples into two groups – pre- or post-menopausal like groups (Supplementary Table 1). Three of the 21 perimenopausal samples could not have a component score calculated due to missing at least one of the seven biological measures included in the model.
Importantly, two of the samples assigned to the “postmenopausal” group based on age, MT-39 (63.3 years) and MT-33 (56.6 years) had component scores −0.4 and + 0.68, respectively, closest to the “premenopausal” group (Supplementary Table 1, Fig. 6C). In order to better separate post- from pre-menopausal cases, we added hypothalamic CYP19A1 gene expression to the model because it was highly correlated with all but blood estradiol and progesterone markers (Supplementary Fig. 5). This eight-marker model was best reduced to a single component which accounted for approximately 45% of the variance in the dataset, and all markers had a factor loading > 0.3 in magnitude (Supplementary Fig. 6A, B). This model achieved a more confident classification of the two individuals (MT-39 and MT-33) as postmenopausal while preserving classification of perimenopausal cases (Supplementary Fig. 6C).
When we checked the classification of the perimenopausal samples, there was almost perfect agreement between the seven- and eight-marker models, with the exception of the sample MT-23 which was classified as premenopausal-like in the seven and postmenopausal-like in the eight marker model (Supplementary Table 3). This sample is an interesting example of being a borderline MT case in basically all the parameters – the individual was 50 years old with both AMH (12.84 pg/mL) and FSH (21.53 mIU/mL) levels hardly falling into the premenopausal values. Thus, this is likely an individual in the very late perimenopause that exemplifies the complexity of the hormonal profile of the MT but also the strength of having multiple biological measures to identify the reproductive state. We decided to keep our seven-marker model because it appears to be the most precise model of the candidate MT biomarkers that we analyzed. The inclusion of CYP19A1 is unnecessary, particularly if hypothalamic tissue is in short supply and steroid quantification should be prioritized. However, quantification of hypothalamic CYP19A1 levels may assist in classification of the baseline premenopausal vs. postmenopausal cutoff score in the case of edge cases in future studies. In addition, hypothalamic CYP19A1 expression was in agreement with the seven-marker component score in 28 out of 37 (or 75.7%) of cases (Supplementary Table 3), so this marker could even be informative in the case when hypothalamic gene expression is the only available dataset for the cohort under the study.
Since our cohort included individuals with psychiatric diagnoses (Table 1), we also wanted to examine whether having a psychiatric diagnosis impacts the final composite measure. Psychiatric diagnoses were characterized as binary variables (history of psychiatric diagnosis is either present or not present), and multiple factor analysis [22], which extends PCA to handle binary variables, was performed to determine if psychiatric diagnosis would significantly impact categorization of menopausal status. Two sets of multiple factor analyses were performed: one collapsing all diagnoses into a single binary variable (using Axis 1 only), and one separating each diagnosis out into three separate binary variables. None of the psychiatric diagnosis groupings had strong associations with the component of the seven biological measurements. Thus, all psychiatric diagnoses were excluded from the final composite score calculation.
We also repeated the multiple factor analyses by adding participant’s race, BMI and anti-depressants history (positive, negative). Results showed that race, BMI and anti-depressant treatment did not have strong associations with the component of the seven biological measurements. Thus, they were excluded from the final composite score calculation.
After calculating the seven-marker composite measure, we further wished to determine whether this model could more reliably characterize each sample than manual characterization based on measures currently being used in clinical settings. An experimenter blinded to each sample’s age and composite measure examined AMH and FSH levels within each of the 42 samples and characterized each measurement as “premenopausal levels” or “postmenopausal levels” based on proposed clinical standards (postmenopausal levels defined as AMH < 10 pg/mL, FSH > 25 mIU/mL) [2, 3] (Supplementary Table 1). Two samples – one premenopausal (MT-3) and one perimenopausal (MT-21) – lack all blood measures and were excluded from this classification. For the 9 premenopausal samples, grouping by chronological age, manual characterization based on AMH and FSH, and composite score all agreed on group assignment (Supplementary Table 1).
For the remaining 31 perimenopausal and postmenopausal samples, 9 samples had conflicting AMH and FSH measurements that would make it difficult to characterize menopausal status (Supplementary Table 1). Of these 9 samples, three were older than 58 years old, indicating that these two biological measures alone may be unreliable even for individuals with a high confidence of postmenopausal status due to chronological age. All three samples were notably correctly characterized as postmenopausal by the composite score despite conflicting measurements in AMH and FSH. In particular, the sample MT-38 (63.1 yo) has the highest component score (2.97) and the highest FSH levels (162.3 mIU/mL), as well as the medical record consistent with postmenopause, yet AMH levels are still above the levels likely to be postmenopausal based on current recommendation (Supplementary Table 1).
Of the six perimenopausal samples with conflicting AMH and FSH measurements, five had composite scores well outside the cutoff range of −0.6−0.4 (Supplementary Table 1), indicating that the other five measures that contributed to the model provided stronger confidence in the characterization of these samples. One sample had a composite score of −0.59, which falls just within the cutoff range. However, this sample came from an individual that was on birth control pill (sample MT-17; 47.3 yo), which likely resulted in their hormonal profile aligning with premenopausal status despite having low, postmenopausal-like (<10 pg/mL) AMH levels (Supplementary Table 1). For the purpose of studying the effect of ovarian hormone shifts on the brain, it is indeed more useful to consider this individual’s hormonal profile as premenopausal-like, although it is important to take into account that, in this case, the origin of hormones was exogenous (Supplementary Table 1, Fig. 6 – red color point). Interestingly, the additional three “perimenopausal” samples that were on hormonal treatment (MT-15, MT-24, MT-26), all were classified as postmenopausal based on our score, consistent with either both or at least one clinical measure (AMH or FSH), likely either due to progesterone-only or discontinued treatment at the time of death (Supplementary Table 1, Fig. 6 – red color point).
All this together supports our seven-marker composite measure as a more reliable way to characterize individuals’ menopausal status as pre- or post-menopausal-like when proposed clinical measures may conflict with each other.
Alternative, tissue-specific component scores
Finally, considering that many brain banks do not have blood available and that working with multiple tissues may not be feasible or cost effective for researchers, we calculated tissue-specific component scores for hypothalamic markers, pituitary markers, and blood markers. We selected tissue-specific markers that were among the 14 significant markers we initially revealed (Table 2). For the hypothalamus, we included steroids markers only: estrone, estradiol, progesterone and DHEA (Supplementary Fig. 7A, 8A, B). Parallel analysis confirmed that the four hypothalamic steroids are well correlated and best reduced to a single component (Supplementary Fig. 7A, 8A). However, unlike the seven-marker model, the hypothalamus-only PCA did not generate a model where all markers had a factor loading of > 0.3 (Supplementary Fig. 8B) and there was a lack of clear cutoff between premenopausal and postmenopausal groups, making characterization of the “perimenopausal samples” less definitive. All but one premenopausal sample had hypothalamus composite measures of > −0.3, and all but two postmenopausal samples had hypothalamus composite measures of < −0.7, meaning that the most appropriate cutoff score would be between these two values (Fig. 7A). With a cutoff of −0.5, 32 out of 37 samples (86.5%) agreed between the hypothalamus component score and the seven-marker component score (Supplementary Table 3). There was a moderate, negative correlation between these two scores (r = −0.54, p < 0.001; Fig. 7B).

A Plot of chronological age vs. hypothalamus component score. 9/10 premenopausal samples had a component score of > −0.3, and 10/11 postmenopausal samples had a component score of < −0.7. A cutoff value to classify samples in the perimenopause group was set halfway between these two limits at −0.5 (dashed line) B The hypothalamus component score moderately correlated with the seven-marker component score. A total of 5 samples disagree between hypothalamus and seven-marker scores. C Plot of chronological age vs. pituitary component score. All samples in the premenopause group had a component score of < 0, and all samples (but MT-39) within the postmenopause group had a component score of > 0. D The pituitary component score strongly correlated with the seven-marker component score. A total of 6 samples disagree between pituitary and seven-marker scores. E Plot of chronological age vs. blood component score. All samples in the premenopause group had a component score of < 0, and all samples within the postmenopause group had a component score of > 0. F The blood component score strongly correlated with the seven-marker component score. Only one sample disagreed between blood and seven-marker scores. On correlation plots B, D, F, blue areas indicate where respective tissue-specific score classification aligned with seven-marker composite measure classification, and red dots indicate samples with disagreement between the two classifications.
For the pituitary component score, we combined pituitary FSH protein levels and gene expression for FSH and GNRHR and were able to show highly correlated markers (Supplementary Fig. 7B), fitted into one component PCA with excellent > 0.5 factor loading (Supplementary Fig. 8C, D). Importantly, the pituitary score had a clear distinction between the premenopausal and postmenopausal groups in their component score, with all premenopausal samples having a component score < 0 and all postmenopausal samples (but MT-39) having a component score > 0 (Fig. 7C). We note that the sample MT-39 (63.3 years old) would be considered premenopausal-like based on this score, although other parameters indicate that menopause has been reached. In addition, with the proposed cutoff, 31 out of 37 (83.8%) samples agreed between the pituitary component score and the seven-marker component score. There was a strong positive correlation between these two scores (r = 0.80, p < 0.001) (Fig. 7D).
Lastly, we calculated two blood component scores: one with all six markers found to have significant differences between the premenopausal and postmenopausal groups (AMH, FSH, estrone, estradiol, progesterone, and DHT) (Supplementary Fig. 7C, 8E, F), and one only containing the four steroid hormones (Supplementary Fig. 7D, 8G, H). Both models were consistent with one component PCA and factor loading > 0.3. Similar to the pituitary component score, the full blood component scores had a distinct separation between the premenopausal and postmenopausal groups, with all premenopausal samples having scores < 0 and all postmenopausal samples having scores > 0 (Fig. 7E). With this cutoff, 36 out of 37 samples (97.3%) agreed between the blood component score and the seven-marker component score. There was a strong positive correlation between these two scores (r = 0.86, p < 0.001) (Fig. 7F). Conversely, the removal of AMH and FSH measures in the steroids-only score removed the clear distinction between premenopausal and postmenopausal groups, with some overlap of component scores. All but one premenopausal sample had a steroids-only component score of > −0.2, and all postmenopausal samples had component scores < −0.2 (Supplementary Fig. 9A). With a cutoff of −0.2, 35 out of 37 samples (94.6%) agreed between the steroids-only component score and the seven-marker component score. Unlike the full blood component score, the steroids-only score had a strong negative correlation with the seven-marker score (r = −0.73, p < 0.001) (Supplementary Fig. 9B). These analyses reveal that future application of this approach in classifying premenopausal-like and postmenopausal-like cases is feasible in cases of limited tissue availability, as all four tissue-specific models generally have high agreement with the full composite measure (Supplementary Table 3). Finally, to examine the robustness of the PCA results, we performed an additional PCA using the Spearman’s rank correlations of the markers, as well as the partial Pearson’s correlations of the markers controlling for PMI, hypothalamus RNA integrity number (RIN), and pituitary RIN. The eigenvalues and the factor loadings of these results were consistent with those presented previously (Supplementary Fig. 10).
Discussion
With the increasing life expectancy and the average age of menopause being ~51 years, it is expected that women and other people with ovaries will spend nearly half of their lives in postmenopause. Importantly, although the MT is associated with the increased risk for many brain disorders [6, 10, 14, 15], the underlying cellular and molecular mechanisms remain poorly understood. To enable postmortem investigations of the human brain across the MT, here we reveal the biomarkers and composite measures across three different tissues that can be used to characterize an individual’s menopausal status postmortem.
Twelve months of amenorrhea are currently required for a formal diagnosis of menopause in living humans, but this is an arbitrary, agreed-upon clinical requirement that is not clearly defined by any biological marker. Revealing the endocrinology underlying the uterine bleeding that characterizes the MT has been challenging because of the marked variability in hormones during this period. Previous studies have proposed using serum levels of FSH [5, 18] or AMH [3, 23] to either diagnose or predict menopause, although no standardized measures have been established so far. While studies in living humans at least provided some guidelines for reference biological measures and the menstrual cycling pattern can be directly inquired from patients and study participants, there is practically no information on how to determine reproductive status from postmortem human tissues as this individual or clinical information is typically lacking in tissue repositories.
We chose three tissues that synthesize or carry HPG-related hormones of direct relevance to reproductive function: blood, the hypothalamus, and pituitary gland. While plasma or serum are typically tissues of choice for exploring the reproductive status in living humans, it is important to note that brain tissue repositories either do not carry blood/serum or may only provide frozen whole blood samples. Importantly, here we show that AMH, FSH, and steroid hormones can be reliably detected in frozen postmortem blood samples, corresponding well to the values observed in clinical serum samples. As expected, blood AMH, FSH, estradiol, and progesterone are among the strongest menopausal markers. However, similar to findings from clinical populations, AMH or FSH by themselves are not sufficient to clearly determine reproductive state postmortem. Rather, our findings show that using the postmortem blood composite measure of the six strongest blood biomarkers (AMH, FSH, estrone, estradiol, progesterone, and DHT) is allowing for high confidence prediction of whether the sample comes from a “pre-menopausal like” or “post-menopausal like” individual. If measuring all six markers is cost prohibitive since they require three separate tests, the composite measure of four steroid hormones in blood (DHT, estrone, estradiol, and progesterone) also allows for a fairly good prediction of the menopausal status.
Another tissue of interest was the hypothalamus, because this region is both involved in the HPG axis and has high density of ovarian hormone receptors [24]. In addition, for brain banks that do not provide access to blood, the hypothalamus can provide an alternative tissue source to determine steroid hormone levels. Previous studies showed that circulating estradiol [25] and progesterone [26] levels are reflected in the hypothalamus and that they differ between postmenopausal and premenopausal women. However, in a small sample size study (N = 11), it was shown that estradiol levels are significantly reduced during postmenopause in the preoptic area but not in the medial and basal hypothalamus [25] as compared to premenopause. In addition, this study did not find a significant correlation between serum and brain concentration of estradiol. However, here we show a significant correlation between steroid hormone levels in blood and in (an unspecified area of) the hypothalamus, which may reflect a higher sensitivity of the method that we used compared to the previous study (HPLC-Mass Spectrometry vs. Radioimmunoassay).
One important consideration regarding the hypothalamus is that this is a highly heterogeneous region and lack of consistent brain dissections may lead to difficulty in finding appropriate biomarkers from this brain region. Indeed, with the acquired material from the hypothalamus, we were not able to detect any MT-related changes in hypothalamic gene expression with the exception of the gene encoding enzyme aromatase (CYP19A1). Importantly, previous studies of the hypothalamus in humans [27] and mice [28] showed changes in estrogen receptor expression following menopause and ovariectomy, respectively. We failed to detect changes in the expression of genes encoding estrogen and progesterone receptors (ESR1, ESR2, GPER1, and PGR) in our study, likely due to our inability to acquire a more specific part of the hypothalamus. As an example, previous studies have found either increased [29] or decreased [30] expression of the kisspeptin (KISS1) gene in the human hypothalamus following menopause, depending on a specific hypothalamic region. Here, we did not observe a difference in KISS1 expression across the groups and individual values demonstrated extreme variability consistent with its region-specific expression.
For this reason, our finding that steroid hormone levels are correlated with their blood levels reveal the possible utility of hypothalamic steroid measurements in the investigations of the menopausal effects on the brain, as this effect may be less region-specific than gene expression. In addition, there is a modest correlation between multi-tissue and hypothalamic-specific composite measures for the menopausal status implying that the hypothalamus can represent an alternative tissue source for studies of the MT and menopause. Importantly, the hypothalamic aromatase (CYP19A1) gene expression not only correlates with estradiol levels in a hypothalamic region-independent manner, but also provides an alternative biomarker for determination of menopausal status in case when the improvement of the PCA model is needed or when hypothalamic gene expression is the only dataset available for the cohort under the study.
Among the most remarkable and less expected findings was the utility of the pituitary gland as an alternative tissue source for determining the menopausal status. While FSH is produced in the pituitary, the FSH assay is typically performed using serum, not the pituitary. Here, however, we show that pituitary FSH represents an excellent menopausal marker, at both protein and gene expression levels. Our data are consistent with previous findings that FSH levels in serum are primarily regulated at the level of FSH production rather than through regulation of FSH release [31]. But, importantly, combining pituitary FSH protein and gene expression with the pituitary GNRHR gene expression gives almost a perfect composite score that resembles the results of applying multi-tissue composite measure.
What is clear from this study is that having multiple, highly-correlated biomarkers increases the ability to predict an individual’s menopausal status. While combining all three tissue indices together gives the most precise information, having markers from blood or pituitary only provide highly reliable results as well. Finally, while not ideal, the hypothalamus can also provide a solid, alternative tissue source if blood or pituitary are not available to researchers. The results of our study provide a long missing method to determine the reproductive status postmortem, allowing the molecular and cellular studies of the MT effect on the human brain. By enabling the study of mechanisms underlying increased neuropsychiatric risk during the MT, we open the path for the hormone status-informed, precision medicine approach in women’s mental health.
Methods
Subjects
The HBCC collected brains, surrounding tissues and blood primarily from medical examiners in Virginia and Washington, D.C. with the permission from the next-of-kin. A telephone interview was conducted with the next-of-kin to gather demographic information and medical history. This was complemented by collection of medical records where available. Medical history was reviewed, and a consensus clinical diagnosis was reached by two psychiatrists based on DSM-IV or DSM-5 criteria. Postmortem brains were obtained from both individuals with no neuropsychiatric illness in their lifetime and individuals with psychiatric disorders (including mood, anxiety, and substance use disorders; Table 1). All cases were assessed by a neuropathologist and found to be free of neurodegenerative disease.
Samples from a total of 42 individuals were received from the HBCC. We requested tissue from women and other people with ovaries that can undergo the menopausal transition. All subjects were assigned female sex at birth. In terms of gender, 11 subjects were identified as “female’, 1 subjects as “non-binary”, and the remaining subjects had no gender reported. Each sample was categorized based on age at the time of death as either “premenopausal” (<40 years old), “perimenopausal” (45–55 years old), or “postmenopausal” (>55 years old). For the majority of the > 55 years old subjects (9 out of 11), we had reasonable evidence that these subjects were postmenopausal, based on their medical records (atrophic or missing ovaries) or age > 62 years (Supplementary Table 1). We avoided the inclusion of individuals over 68 to minimize chronological age-related biases in our data. For every sample, we received ~200 mg of tissue from the hypothalamus, ~500 mg of tissue from the pituitary gland including both anterior and posterior portions, and 500–1000 μL of whole blood.
Sample preparation
Brain collection and processing was performed as previously described [32]. Briefly, brains were sectioned into ~1 cm coronal slabs, and slabs were flash frozen in a slurry of isopentane and dry ice prior to storage at −80 °C. Upon receipt of tissue request, slabs were removed from storage and blocks of tissue were dissected, maintaining frozen temperature throughout processing. The HBCC provided tissue from the hypothalamus and pituitary as well as aliquots of whole blood for analysis.
Tissue blocks received from the HBCC were pulverized to a coarse powder over dry ice. Pulverized tissue samples were separated into aliquots for separate analyses. Approximately 100 mg of hypothalamic tissue was set aside for steroid analysis, and 50–100 mg of hypothalamic tissue was used for RNA isolation. Between 25–75 mg of pulverized pituitary tissue was used each for RNA isolation and protein extraction. Pulverized tissue was stored at −80 °C until further processing.
Whole blood was thawed and centrifuged for 20 min at 1500xg at 4 °C to separate coagulation before aliquoting. 200 µL of whole blood was set aside for steroid analysis and two 100 μL aliquots were set aside for ELISA assays. All aliquots were stored at −80 °C until analysis.
Three premenopausal samples also had serum provided, which were used during assay optimization to confirm whether each assay was compatible with whole blood and determine the ratio of whole blood measures to plasma measures of AMH and the 14 steroids. All measures had a similar pattern of whole blood measuring at around 50% of the values for plasma (Supplementary Table 4).
Steroid panel
Steroid panel concentration was quantified with high-performance liquid chromatography with tandem-mass spectrometry (HPLC-MS/MS) by OpAns, LLC (Durham, NC). The reference standards for aldosterone, androstenedione, corticosterone, cortisol, cortisone, dehydroepiandrosterone (DHEA), deoxycorticosterone (DOC), 11-deoxycortisol, dihydrotestosterone (DHT), estradiol, estrone, 17OH-progesterone, progesterone, and testosterone were prepared as individual stock solutions at 1 mg/mL in acetonitrile (ACN) and dimethyl sulfoxide (DMSO) (50:50), then further diluted in ACN and DMSO (50:50) to create calibration and quality control standards. Stable isotropic labeled internal standards for each analyte were added to calibration standards, quality control, and matrix samples.
Analytes and their internal standards were extracted from whole blood by liquid-liquid extraction using methyl tert-butyl ether (MTBE). Following room temperature centrifugation at 14,000 RPM, the supernatant was evaporated to dryness and reconstituted in MeOH:water. Hypothalamus samples were homogenized in MeOH and EtOAc (50:50). The resulting homogenate was centrifuged and fortified with internal standards. Following evaporation, the extracts were reconstituted in MeOH:water and then subjected to solid phase extraction using Biotage Evolute Express ABN sorbent. After washing, wells were eluted using MeOH, evaporated to dryness, and reconstituted with MeOH:water. The reconstituted blood and hypothalamus extracts were then analyzed with HPLC-MS/MS using a 1290 series HPLC system (Agilent Technologies) and Agilent 6495 series triple quadrupole tandem mass spectrometer. HPLC-MS/MS data were acquired and quantified using the software application MassHunter Workstation Data Acquisition for Triple Quad version 10.1/ build 10.1.67 (Agilent Technologies).
Pituitary protein extraction
An extraction buffer of phosphate buffered saline (pH 7.4) (PBS) containing 1X protease/phosphatase inhibitor cocktail was prepared and kept on ice during use. Pulverized pituitary was added to 1 mL of chilled extraction buffer and homogenized, then centrifuged for 5 min at 5000xg at 4 °C. The supernatant was then removed and placed into a new chilled tube on ice and the tissue pellet was discarded. Total protein concentration was quantified with a Pierce BCA protein assay kit (Thermo Scientific) according to manufacturer instructions. Protein extract was stored at −80 °C until analysis.
FSH ELISA
Whole blood aliquots and pituitary protein extracts were thawed over ice, and FSH concentration was quantified using an ELISA kit (Novus Biologicals) according to manufacturer instructions. Whole blood was diluted to a 1:10 concentration in PBS, and pituitary protein extracts were sequentially diluted to 1:100 concentration twice in PBS for a final concentration of 1:10,000. Diluted samples and prepared standards were loaded in duplicate into the pre-coated plates, then antibody solution was added. Plates were incubated at 37 °C for 1 h, then washed three times with wash buffer. Following washing, HRP-conjugate was added and the plate was incubated at 37 °C for 30 min. The plates were washed once more, and development substrates were added before one final incubation at 37 °C for 15 min. Finally, stop solution was added and the plates were read with a microplate reader (BMG Labtech) at 450 nm. Optical densities of duplicates were averaged and the optical density of the 0 standard (blank) was subtracted from every value. Sample concentrations were calculated by plotting the log of sample optical density along the standard curve generated by the log of standard optical density against the log of standard concentrations, then appropriately multiplied by the dilution factor to determine the total concentration. Pituitary FSH concentration was then divided by total protein concentration to determine the concentration of FSH per mg of extracted protein.
Blood intra- and inter-assay CV’s were 5.6 and 2.2%, respectively. Pituitary intra- and inter-assay CV’s were 4.2 and 3.5%, respectively.
AMH ELISA
Whole blood aliquots were thawed over ice, and AMH concentration was quantified using the picoAMH ELISA kit (Ansh Labs) according to manufacturer instructions. Microplates were loaded with assay buffer, calibrators, and controls. 20 μL of each whole blood sample was loaded in duplicate, then 80 μL of sample diluent was added to each sample well to bring each sample well to a final 1:5 concentration. Plates were then incubated at room temperature for three hours while on a shaker at ~600 rpm. Plates were then washed five times with wash buffer, and antibody-biotin conjugate was added before another incubation at room temperature for one hour while shaking. Plates were washed again, then loaded with streptavidin-enzyme conjugate before incubating again at room temperature for 30 min while shaking. Plates were washed one final time, then loaded with TMB solution and incubated at room temperature for 10 min while shaking. Stop solution was then added and the plates were read using a microplate reader at 450 nm. Sample concentrations were calculated by plotting the log of sample optical density along the standard curve generated by the log of calibrator optical density against the log of calibrator concentrations, then appropriately multiplied by the dilution factor to determine the total concentration.
Intra- and inter-assay CV’s were 4.1 and 2.6%, respectively. Calculated concentrations for Controls I and II provided within each picoAMH kit were within stated concentration ranges.
RNA isolation and RT-qPCR
Pituitary and hypothalamus tissue was first processed with Trizol reagent (Thermo Scientific) and RNA was isolated from the aqueous layer using the Allprep DNA/RNA Mini kit (Qiagen). Pulverized tissue was homogenized in 1 mL Trizol reagent, 200 uL of chloroform was added and mixed by inversion, then samples incubated at room temperature for 10 min. Samples were then centrifuged at 4 °C for 15 min at 12,000xg. Following centrifugation, the aqueous phase was removed and loaded into the Allprep DNA spin column. From this point samples were processed according to manufacturer instructions, with the RW1 wash step separated into two 350 μL washes before and after DNase (Qiagen) added to the spin column to ensure elimination of gDNA within the sample. Following RNA elution, total RNA concentration was determined using a Qubit RNA High Sensitivity kit and Qubit fluorometer (Invitrogen). RIN was determined with Bioanalyzer using an RNA 6000 Pico kit (Agilent). Isolated RNA was stored at −20 °C until conversion. 500 ng of RNA was reverse transcribed to cDNA using SuperScript III First-Strand Synthesis System (Invitrogen), and stored at −20 °C until analysis. cDNA was analyzed for gene expression with quantitative PCR using a Quant Studio 3 real-time PCR system (Applied Biosystems). Forward and reverse primers for each gene analyzed are listed in Supplementary Table 5. Both hypothalamus and pituitary genes of interest were normalized against three housekeeping genes each using the previously described method [33]. The housekeeping genes used for the hypothalamus were CYC1, UBE2D2, and EIF4A2 [34, 35]. The housekeeping genes used for the pituitary were GAPDH, TBP, and PSMC4 [36, 37]. Gene expression was calculated relative to the premenopausal group.
Statistics
Individual markers
Demographic information (Table 1) including age, BMI, education level, PMI, and RIN values were analyzed with one-way ANOVA with Bonferroni-Hochberg corrections for multiple comparisons. A total of 40 biological measurements were taken for each of the 42 samples, including steroid hormones, glycoproteins, and gene expression (Table 2). For steroid measurements that were detectable but below the limit of quantitation, the value was indicated to be half of the lower limit of quantitation for the purposes of performing statistical analysis. None of the markers met the assumption for normal distribution (Shapiro-Wilk p < 0.05), so non-parametric tests were used for group comparisons. All 40 analytes were first compared with Kruskal-Wallis test, with Dunn’s test for pairwise comparison with Benjamini-Hochberg correction for multiple comparisons. Measurements that had a significant difference between premenopausal and postmenopausal groups as determined by Dunn’s test posthoc were used for subsequent analyses. As a follow up to non-parametric Kruskal-Wallis test, we performed an additional analysis of the individual markers using the robust ANOVA test by Wilcox and pairwise comparisons that are robust to nonnormality and heterogeneity of variance [21] (Supplementary Table 2). In this test, we used the 5% trimmed mean to discard the top and bottom 5% of the outcomes from the ANOVA tests. Results are reported as Median (First quartile – Third quartile) by group.
Composite measure
The composite measure was determined using principal component analysis (PCA), which is a statistical method of dimension reduction used to group variables that highly associate with each other into a single variable known as a “component.” Markers’ distribution was first examined using kernel density plots, demonstrating the non-normal distribution of each of the markers. Scatterplots of each pair of markers with linear regression lines imposed showed that most markers were linearly associated. We repeated the kernel density plots and scatterplots with linear regression lines after the natural-log transformation of the markers. Results were consistent and it was decided that no data transformation was needed.
Parallel analysis [19, 38] was used to determine the dimensionality of the markers. Observed eigenvalues were compared to the 95th percentile of eigenvalues across multiple randomly generated datasets with same number of random, uncorrelated markers (simulated eigenvalues). Components with sample eigenvalues being higher than those of the randomly generated data can be extracted. The number of dimensions with an observed eigenvalue greater than that of the simulated eigenvalue was used for PCA. For PCA, all markers were standardized (mean = 0, standard deviation = 1). After determining the principal component model, multiple factor analysis [22] was performed with confident Axis I diagnoses (including depression, anxiety, and substance use disorders) together with the selected biological measurements. Multiple factor analysis extends PCA to handle binary variables. We compared the degrees of associations between the diagnoses and the component with those between the biological measurements and the component. We also repeated the multiple factor analyses by adding participant’s race (26 White, 14 Black, 1 Asian which was treated as missing due to small size), BMI and anti-depressants history (positive, negative). Notably a component score could not be calculated for any samples missing at least one measurement; this led to exclusion of one premenopausal sample, three perimenopausal samples, and one postmenopausal sample from the seven-marker composite measure. To explore the robustness of the PCA results, we performed an additional PCA using the Spearman’s rank correlations of the markers, as well as the partial Pearson’s correlations of the markers controlling for PMI, hypothalamus RNA integrity number (RIN), and pituitary RIN.
All analyses were conducted in R version 4.3.2 (https://www.R-project.org) [39]. Kruskal-Wallis comparison across groups was performed with kruskal.test function in the stats package. Dunn’s test for pairwise comparison was performed with the dunnTest function in the FSA package [40] (https://CRAN.R-project.org/package=FSA). The fa.parallel function in package psych was used for parallel analysis [40]. The prcomp function in package stats was used for principal component analysis. The MFA function in package FactoMineR was used for multiple factor analysis (CRAN - Package FactoMineR (r-project.org)). The partial.r function in package psych was used for partial Pearson’s correlations.
Data availability
All data supporting the findings of this study are available within the paper and its Supplementary Information, or from the corresponding author upon reasonable request.
References
Mehta JM, Manson JE. The menopausal transition period and cardiovascular risk. Nat Rev Cardiol. 2024;21:203–11.
Harlow SD, Gass M, Hall JE, Lobo R, Maki P, Rebar RW, et al. Executive summary of the stages of reproductive aging workshop + 10: addressing the unfinished agenda of staging reproductive aging. Menopause. 2012;19:387–95.
Finkelstein JS, Lee H, Karlamangla A, Neer RM, Sluss PM, Burnett-Bowie SM, et al. Antimullerian hormone and impending menopause in late reproductive age: the study of Women’s health across the nation. J Clin Endocrinol Metab. 2020;105:e1862–1871.
Moolhuijsen LME, Visser JA. Anti-müllerian hormone and ovarian reserve: update on assessing ovarian function. J Clin Endocrinol Metab. 2020;105:3361–73.
Randolph JF Jr., Zheng H, Sowers MR, Crandall C, Crawford S, Gold EB, et al. Change in follicle-stimulating hormone and estradiol across the menopausal transition: effect of age at the final menstrual period. J Clin Endocrinol Metab. 2011;96:746–54.
Brinton RD, Yao J, Yin F, Mack WJ, Cadenas E. Perimenopause as a neurological transition state. Nat Rev Endocrinol. 2015;11:393–405.
Appiah D, Nwabuo CC, Ebong IA, Wellons MF, Winters SJ. Trends in age at natural menopause and reproductive life span among US women, 1959-2018. JAMA. 2021;325:1328–30.
Chan S, Gomes A, Singh RS. Is menopause still evolving? Evidence from a longitudinal study of multiethnic populations and its relevance to women’s health. BMC Womens Health. 2020;20:74.
Gottschalk MS, Eskild A, Hofvind S, Gran JM, Bjelland EK. Temporal trends in age at menarche and age at menopause: a population study of 312 656 women in Norway. Hum Reprod. 2020;35:464–71.
El Khoudary SR, Greendale G, Crawford SL, Avis NE, Brooks MM, Thurston RC, et al. The menopause transition and women’s health at midlife: a progress report from the study of Women’s health across the nation (SWAN). Menopause. 2019;26:1213–27.
Edwards H, Duchesne A, Au AS, Einstein G. The many menopauses: searching the cognitive research literature for menopause types. Menopause. 2019;26:45–65.
Sherman S. Defining the menopausal transition. Am J Med. 2005;118:3–7. Suppl 12B
Kundakovic M, Rocks D. Sex hormone fluctuation and increased female risk for depression and anxiety disorders: from clinical evidence to molecular mechanisms. Front Neuroendocrinol. 2022;66:101010.
Culbert KM, Thakkar KN, Klump KL. Risk for midlife psychosis in women: critical gaps and opportunities in exploring perimenopause and ovarian hormones as mechanisms of risk. Psychol Med. 2022;52:1612–20.
Deecher D, Andree TH, Sloan D, Schechter LE. From menarche to menopause: exploring the underlying biology of depression in women experiencing hormonal changes. Psychoneuroendocrinology. 2008;33:3–17.
Rubinow DR, Schmidt PJ. Is there a role for reproductive steroids in the etiology and treatment of affective disorders? Dialogues Clin Neurosci. 2018;20:187–96.
Mosconi L, Berti V, Dyke J, Schelbaum E, Jett S, Loughlin L, et al. Menopause impacts human brain structure, connectivity, energy metabolism, and amyloid-beta deposition. Sci Rep. 2021;11:10867.
Burger HG. Diagnostic role of follicle-stimulating hormone (FSH) measurements during the menopausal transition–an analysis of FSH, oestradiol and inhibin. Eur J Endocrinol. 1994;130:38–42.
Fabrigar LR, Wegener DT, MacCallum RC, Strahan EJ. Evaluating the use of exploratory factor analysis in psychological research. Psychol Methods. 1999;4:272.
Tavakol M, Wetzel A. Factor analysis: a means for theory and instrument development in support of construct validity. Int J Med Educ. 2020;11:245–7.
Mair P, Wilcox R. Robust statistical methods in R using the WRS2 package. Behav Res Methods. 2020;52:464–88.
Pagès J Multiple factor analysis by example using R. CRC Press, New York, 2014.
Nelson SM, Davis SR, Kalantaridou S, Lumsden MA, Panay N, Anderson RA. Anti-müllerian hormone for the diagnosis and prediction of menopause: a systematic review. Hum Reprod Update. 2023;29:327–46.
Marrocco J, McEwen BS. Sex in the brain: hormones and sex differences. Dialogues Clin Neurosci. 2016;18:373–83.
Bixo M, Bäckström T, Winblad B, Andersson A. Estradiol and testosterone in specific regions of the human female brain in different endocrine states. J Steroid Biochem Mol Biol. 1995;55:297–303.
Bixo M, Andersson A, Winblad B, Purdy RH, Bäckström T. Progesterone, 5alpha-pregnane-3,20-dione and 3alpha-hydroxy-5alpha-pregnane-20-one in specific regions of the human female brain in different endocrine states. Brain Res. 1997;764:173–8.
Mosconi L, Nerattini M, Matthews DC, Jett S, Andy C, Williams S, et al. In vivo brain estrogen receptor density by neuroendocrine aging and relationships with cognition and symptomatology. Sci Rep. 2024;14:12680.
Ouldibbat L, Rocks D, Sampson B, Kundakovic M. The role of ovarian hormone dynamics in metabolic phenotype and gene expression in female mice. Horm Behav. 2025;169:105693.
Rometo AM, Krajewski SJ, Voytko ML, Rance NE. Hypertrophy and increased kisspeptin gene expression in the hypothalamic infundibular nucleus of postmenopausal women and ovariectomized monkeys. J Clin Endocrinol Metab. 2007;92:2744–50.
Rumpler É, Skrapits K, Takács S, Göcz B, Trinh SH, Rácz G, et al. Characterization of kisspeptin neurons in the human rostral hypothalamus. Neuroendocrinology. 2021;111:249–62.
McNeilly AS, Crawford JL, Taragnat C, Nicol L, McNeilly JR. The differential secretion of FSH and LH: regulation through genes, feedback and packaging. Reprod Suppl. 2003;61:463–76.
Lipska BK, Deep-Soboslay A, Weickert CS, Hyde TM, Martin CE, Herman MM, et al. Critical factors in gene expression in postmortem human brain: focus on studies in schizophrenia. Biol Psychiatry. 2006;60:650–8.
Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3:Research0034.
Rydbirk R, Folke J, Winge K, Aznar S, Pakkenberg B, Brudek T. Assessment of brain reference genes for RT-qPCR studies in neurodegenerative diseases. Sci Rep. 2016;6:37116.
Penna I, Vella S, Gigoni A, Russo C, Cancedda R, Pagano A. Selection of candidate housekeeping genes for normalization in human postmortem brain samples. Int J Mol Sci. 2011;12:5461–70.
van Rijn SJ, Riemers FM, van den Heuvel D, Wolfswinkel J, Hofland L, Meij BP, et al. Expression stability of reference genes for quantitative RT-PCR of healthy and diseased pituitary tissue samples varies between humans, mice, and dogs. Mol Neurobiol. 2014;49:893–9.
Normann KR, Øystese KAB, Berg JP, Lekva T, Berg-Johnsen J, Bollerslev J, et al. Selection and validation of reliable reference genes for RT-qPCR analysis in a large cohort of pituitary adenomas. Mol Cell Endocrinol. 2016;437:183–9.
Horn JL. A rationale and test for the number of factors in factor analysis. Psychometrika. 1965;30:179–85.
Team RC. R: A language and environment for statistical computing. R Foundation for Statistical Computing; 2013 (No Title).
Ogle DH, Doll JC, Wheeler AP, Dinno A. FSA: Simple Fisheries Stock Assessment Methods. R package version 0.10.0. https://fishr-core-team.github.io/FSA/.
Acknowledgements
This work was supported by the National Institutes of Mental Health (NIMH) under Award Number R01MH138370 and the Fordham University’s Faculty Research Grant to MK. The Human Brain Collection Core (HBCC) is supported by the NIMH-IRP project ZIC MH002903. This research was supported in part by the Intramural Research Program of the National Institutes of Health (NIH). The contributions of the NIH author(s) were made as part of their official duties as NIH federal employees, are in compliance with agency policy requirements, and are considered Works of the United States Government. However, the findings and conclusions presented in this paper are those of the author(s) and do not necessarily reflect the views of the NIH or the U.S. Department of Health and Human Services. We would like to thank Brian Nofsinger for his assistance with steroid hormone quantification.
Author information
Authors and Affiliations
Contributions
MK conceived and directed the project and provided funding; MT, HC, PA, SM, and MK designed the study; MT prepared samples for the analyses and performed ELISA and gene expression experiments; LO helped with gene expression analysis; MT and HC performed data analysis; AG performed manual characterization of the samples; SB performed a literature search; PA and SM provided postmortem tissue specimens; MT, HC, and MK interpreted the data and constructed the figures; MT and MK wrote the article with input from all co-authors; HC, PJS, and SM revised the paper. All authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Human postmortem tissue samples were deidentified before being transferred to Fordham University and were, thus, exempt from Institutional Review Board review. All methods in this study were performed in accordance with the relevant guidelines and regulations.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tickerhoof, M., Cham, H., Ouldibbat, L. et al. Postmortem tissue biomarkers of menopausal transition. Mol Psychiatry (2025). https://doi.org/10.1038/s41380-025-03177-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41380-025-03177-9
