
Abstract
Tissue-resident immune cells (TRICs) are a highly heterogeneous and plastic subpopulation of immune cells that reside in lymphoid or peripheral tissues without recirculation. These cells are endowed with notably distinct capabilities, setting them apart from their circulating leukocyte counterparts. Many studies demonstrate their complex roles in both health and disease, involving the regulation of homeostasis, protection, and destruction. The advancement of tissue-resolution technologies, such as single-cell sequencing and spatiotemporal omics, provides deeper insights into the cell morphology, characteristic markers, and dynamic transcriptional profiles of TRICs. Currently, the reported TRIC population includes tissue-resident T cells, tissue-resident memory B (BRM) cells, tissue-resident innate lymphocytes, tissue-resident macrophages, tissue-resident neutrophils (TRNs), and tissue-resident mast cells, but unignorably the existence of TRNs is controversial. Previous studies focus on one of them in specific tissues or diseases, however, the origins, developmental trajectories, and intercellular cross-talks of every TRIC type are not fully summarized. In addition, a systemic overview of TRICs in disease progression and the development of parallel therapeutic strategies is lacking. Here, we describe the development and function characteristics of all TRIC types and their major roles in health and diseases. We shed light on how to harness TRICs to offer new therapeutic targets and present burning questions in this field.
Similar content being viewed by others

Introduction
Many diseases are caused by immune dysfunction. The innate and adaptive immune systems coordinate to ensure immune surveillance, clearance, and homeostasis of organisms. Innate immune cells act as the first barrier line and contribute to motivating adaptive immune responses,1 while the adaptive immune cells usually circulate between different organs and perform specific immune responses.2 Interestingly, some immune cell populations from innate or adaptive immunity develop tissue-resident features and hardly perform circulation, and they often undergo specific phenotypic and functional reprogramming.3,4,5 Here, we used the term, tissue-resident immune cells (TRICs), to characterize these cell populations. Studies in the last 20 years have constantly explored each member of TRICs. Inspiringly, under the help of rapidly iterative technologies, their distinct origins, developmental trajectories, maintenance mechanisms, specific phenotypic and transcriptional characteristics, and functions are increasingly clear. Landmark studies on TRICs are shown in Fig. 1 (Fig. 1).

Milestone research of TRICs. Key events in the exploration of TRICs were retrospectively shown from 2009 to the present. The text box with a blue background depicts lymphoid-lineage TRICs, and the text box with a pink background describes myeloid-lineage TRICs. TRM tissue-resident memory T, HSV herpes simplex virus, ILC innate lymphoid cell, BRM resident memory B, MAIT mucosal-associated invariant T, iNKT invariant natural killer T, TRN tissue-resident neutrophil, NSCLC non-small cell lung cancer, PD-L1 programmed cell death-1-ligand-1, CAR chimeric antigen receptor, Blimp B lymphocyte-induced maturation protein, Hobit Homology of Blimp in T cells. This figure is created with BioRender.com
Lymphoid or myeloid-lineage immune cells can develop tissue-resident features and possess specific properties that circulating immune cells are not equipped with.5,6 We define these cells as lymphoid-lineage TRICs (hereafter lymphoid TRICs) and myeloid-lineage TRICs (hereafter myeloid TRICs) respectively. They extensively distribute in second lymphoid organs (SLOs) or peripheral non-lymphoid tissues, such as skin, brain, liver, lung, kidney, and gastrointestinal tract.5 Endogenous regulatory mechanisms- and environmental factors-inducible phenotypic reprogramming co-maintain the long-term persistence of TRICs in peripheral tissues and inhibit their detachment from tissues into circulation (Fig. 2).

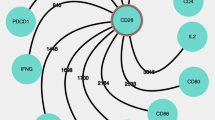
The types, representative markers, and fundamental functions of TRICs. The surface markers used for defining and identifying TRICs are shown. The markers include differentiation markers like CD4, CD8, and CD20, and tissue residency markers include CD103, CD49a, and CD69. Some critical chemokine receptors for tissue location and key TFs responsible for TRIC development are displayed. The transcriptional characteristics of BRM cells are undefined, and myeloid TRICs have distinct transcriptional regulation in different tissues that is difficult to summarize in one plot. The main functions of each TRIC in physiological and pathological conditions are listed. trNK tissue-resident natural killer, tr-ILC tissue-resident ILC, tr-mast cell tissue-resident mast cell, tr-macrophage tissue-resident macrophage, tr-γδT tissue-resident γδT, tr-MAIT tissue-resident MAIT, tr-iNKT tissue-resident iNKT, TCR T cell receptor, BCR B cell receptor, CXCR C-X-C chemokine receptor, CD the cluster of differentiation, HLA human leukocyte antigen, MARCO macrophage receptor with collagenous structure. This figure is created with BioRender.com
TRICs are highly heterogeneous across different tissues and participate in many biological processes. They share some common functions with canonical circulative immune cells, including defense against pathogens and cell collaboration. Notably, long-term tissue residency endows some specific functions of TRICs, such as maintaining durable immune defense and local homeostasis.7 Nonetheless, the lasting tissue retention also has side effects, increasing the vulnerability to autoimmune disease, cancer, and graft rejection.5,7,8 Therefore, how to utilize TRICs to maintain conducive immune functions is an issue worth exploring.
With the growing applications of immunotherapies in many diseases, TRICs are thought to be a potential target for novel therapy development. People have begun to explore the feasibility of targeting disease-specific TRICs as therapeutic strategies.9,10,11,12,13,14 For example, some vaccines can induce the formation of CD8+ tissue-resident memory T (TRM) cells that defend against pathogen reinfection and amplify anti-tumor immunity.15,16,17 Furthermore, neutralizing antibodies blocking proinflammatory cytokines produced by TRICs can alleviate autoimmune disease progression.18,19 Therefore, a thorough understanding of TRICs is very essential for further therapy optimization and development.
In this review, we comprehensively describe the origins, biological characteristics, and functional evolutions of each TRIC type. We summarize their roles in physiological conditions and pathological contexts, covering the major diseases in which TRICs intervene. Moreover, we emphasize the potential of TRICs as promising therapeutic targets and predictive biomarkers for diseases. We hope the understanding of TRICs is helpful for future investigation of immunity in health and diseases.
Definition of tissue-resident immune cells
The origins of TRICs
The precursors and original sites of TRICs are distinct. Some TRICs acquire tissue-resident properties during embryogenesis, such as tissue-resident macrophages and mast cells, tissue-resident natural killer (trNK) cells, tissue-resident innate lymphoid cells (ILCs), and γδT cells, while other TRIC types postnatally establish tissue residency during the effector stages (Fig. 3).20,21 TRICs have different lifetimes under different contexts. In most cases, TRICs achieve long-term tissue residence by self-renewal, but they also depend on the replenishment of circulating immune cells to varying degrees.5,6,22

The developmental trajectories of TRICs. HSCs from bone marrow can differentiate into TRIC precursors. Embryonic hematopoiesis is another progenitor cell source for TRICs. Tissue-resident macrophages and mast cells originate from embryonic EMPs, and circulating monocytes or mast cell precursors can also acquire tissue-resident features after entering tissues. Most lymphoid TRICs develop postnatally and derive from lymphoid progenitors. Tissue-resident ILCs and trNK cells derive from fetal innate lymphoid progenitors during the embryonic period or postnatal lymphoid progenitors. Local progenitors can also differentiate into tissue-resident ILCs and trNK cells in some tissues. After leaving the thymus, tissue-resident MAIT cells, iNKT cells, and γδT cells localize in peripheral tissues, while naïve αβT cells migrate into SLOs. In SLOs, naïve T cells are activated by mature DCs, then T cells proliferate and differentiate into TEFF and memory-like precursors. TEFF will transform into memory T cells stimulated by cognate antigens, including TCM and TEM cells. TRM cells can derive from primary TRM precursors or TEM cells. The interaction with TFH cells promotes GC B cells to differentiate into memory B cells and plasma cells. During the effector stages, circulating memory B cells can become the precursor of BRM cells. BM bone marrow, EMP erythroid myeloid progenitor, HSC hematopoietic stem cell, DC dendritic cell, TEFF effector T, TEM effector memory T, TCM central memory T, TFH follicular helper T, GC germinal center, MBC memory B cell. This figure is created with BioRender.com
Regarding myeloid cells, tissue-resident macrophages and mast cells can originate from both embryogenic precursors and adult hematopoietic stem cells (HSCs)-derived myeloid progenitors.21,23,24,25,26,27,28 They primarily rely on self-renewal for long-term persistence within tissues.23,29,30,31,32,33,34,35,36,37 Exactly, macrophages in different tissues have distinct requirements for the extra supplement of bone marrow-derived progenitors.20,37,38,39,40,41,42 Subsequently, regardless of their origin, they are plastic and develop tissue-specific phenotypes after entering tissues.39,43,44,45,46,47,48 Neutrophils are always short-lived in tissues. They can quickly develop tissue-specific behaviors under the steady state and rely on some factors to prolong their tissue-resident lifespan.49,50,51,52 These neutrophils with a relatively long-tissue livelihood are termed tissue-resident neutrophils (TRNs).
Among lymphoid cells, TRM cells are the most widely studied TRIC type. They derive from circulating memory precursor cells following T cell activation and develop tissue-specific phenotypes across different tissues.53,54 BRM cells may be derived from memory B cells (MBCs) triggered by pathogen infection during adulthood, and the inducible bronchus-associated lymphoid tissue (iBALT) may also specifically contribute to their formation in the lung.55,56,57,58 As for unconventional T cells, tissue-resident mucosal-associated invariant T (MAIT) cells and invariant natural killer T (iNKT) cells derive from postnatal precursors in the thymus, and they adopt tissue-resident characteristics after lineage commitment.59,60,61,62 In contrast, tissue-resident γδT cells derive from thymocytes during embryonic and adult periods, but they are endowed with tissue residency properties before exiting from the thymus and residing in peripheral tissues.63,64,65 Uniquely, the progenitors of resident ILCs distribute into peripheral tissues during the embryonic periods, such as the fetal intestine, liver, and tonsils. ILCs can also derive from progenitors in the bone marrow and local tissues during the adult period.66,67,68,69 Tissue-resident ILCs predominantly maintain themselves by self-renewal, with less role for circulating or locally derived hematopoietic progenitors in supporting ILC maintenance.68,69,70,71 Similarly, trNK cells differentiate from bone-marrow-derived progenitors, differentiated classical NK (cNK) cells in adulthood, or tissue-resident progenitors during embryogenesis.66,67,68,72
Advanced technologies for TRIC finding
The identification of TRICs is subject to experimental technologies. Parabiotic surgery is the direct method to identify TRICs. People combine the circulation of two homogeneous mice, and the circulating cells will establish equilibrium in both mice, and tissue-resident cells do not enter circulation.73,74 TRM cells, ILCs, trNKs, BRM cells, tissue-resident macrophages, tissue-resident iNKT, MAIT cells, and TRNs have been identified via this approach.30,38,49,56,62,68,75,76,77,78,79 Intravascular labeling employs antibodies to distinguish blood-borne and tissue-resident lymphocytes that will not be labeled, like TRM cells and BRM cells.56,80,81 Moreover, flow cytometry and cytometry by time of flight are used to identify TRICs by detecting the specific marker profiles.79,82,83,84,85
High-throughput sequencing technologies help to construct the comprehensive fate and feature map of TRICs.86 Fate-mapping analysis and lineage tracing system contribute to clarifying their origins and developmental trajectories.31,38,49,79,87 T cell receptor (TCR) and B cell receptor sequencing analyses have been used to explore the immunological characteristics of TRM cells, γδT cells, and BRMs.7,57,88 Especially, single-cell RNA-sequencing (scRNA-seq) analysis is effective for exploring the phenotypic or transcriptional profiles of TRM cells, BRM cells, trNK cells, tissue-resident macrophages, TRNs, and ILC2s in physiological states and even diseases.49,55,57,69,83,89,90,91,92,93,94,95,96,97,98,99 In this way, trNK cells are found to express distinct cytokine receptors from cNK cells with heterogeneous phenotypes across the liver, lung, and salivary glands.83,100,101,102,103,104 In addition, three distinct tissue-resident macrophage subsets with different transcription profiles in tumor tissues were also identified by scRNA-seq.40 In disease studies, scRNA-seq analysis of patients’ samples confirmed the existence of TRICs in multiple sclerosis (MS),105 autoimmune nephritis,106 influenza infections,107 and cancers.84
The phenotypic markers and transcriptional profiles of TRICs
The phenotypes of TRICs are deeply shaped by molecules and cellular interaction within the local tissue environment, so the same kinds of TRIC render to acquire high heterogeneity in different organs.22,82 Characteristic phenotype markers and regulatory transcription factors (TFs) are useful in distinguishing them (Fig. 2).
CD69, CD103, and CD49a (very late antigen-1, VLA-1) are canonical tissue-resident markers for lymphoid TRICs, but their expression varies across different tissues. Most TRICs, including TRM cells, tissue-resident ILCs, and γδT cells, predominantly express CD103 in mucosal tissues. However, in non-mucosal tissues, CD103 expression is not common or even absent, while CD69 and/or CD49a are more expressed in these cell populations.108,109 A combined expression of CD40L, CD56, CD161, and CD69 characterizes iNKT cells,110,111 MAIT cells are defined by CD161, C-C motif chemokine receptor (CCR) 5, and CCR6.60 BRM cells generally express CD69 and CD45RB,89 and trNK cells express CD69, CD49a, and C-X-C chemokine receptor (CXCR) 6.112 In addition to those markers, most lymphoid TRICs upregulate genes encoding tissue adhesion- or tissue recruitment-related molecules accompanied by downregulating genes that induce them to detach tissues, such as sphingosine 1 phosphate receptor-1 (S1PR1) and CCR7.
Lymphoid TRICs share similar transcription profiles. Several TFs are essential for the formation and development of lymphoid TRICs. First, Krüppel-like factor 2 (KLF2) regulates the tissue egress of lymphocytes and T cell homeostasis by increasing S1PR1 and CD62L expression.101,113 Low KLF2 is a unique feature in lymphoid TRICs, such as CD8+TRM cells, CD4+TRM cells, trNK cells, and iNKT cells.101,114,115,116,117
B lymphocyte-induced maturation protein (Blimp) and Homology of Blimp in T cells (Hobit) are indispensable for the maintenance of TRM cells, iNKT cells, and liver-resident NK cells.101,118 They repress the expression of circulation-promoting factors KLF2, S1PR1, CCR7, and TCF1 in lymphoid TRICs.74 Notably, they exhibit functional complementarity.101,119 Lacking either Hobit or Blimp partially inhibits the persistence of CD8+TRM cells and iNKT cells in tissues, but the absence of both greatly suppresses the formation of these TRICs.101,118,120,121,122 In contrast to Hobit and Blimp, T-bet and Eomesodermin (Eomes) are negative regulators for TRM cells. The forced expression of either T-bet or Eomes impairs CD8+TRM cell formation.123,124 Other TFs also contribute to the TRIC development, including aryl hydrocarbon receptors for skin CD8+TRM cells and liver-trNK cells,125,126 and Notch for lung CD8+TRM cells.127
However, unlike lymphoid TRICs, tissue-resident macrophages, TRNs, and mast cells do not have identical molecular features, which may be due to myeloid TRICs developing natural tissue-resident properties during embryogenesis.22,82
Cellular interactions involving TRICs
The interactions between TRICs and other cell components in tissues are important for the tissue residency of TRICs and their functions.128 The interaction mechanisms include the direct binding of membrane surface receptors and ligands such as CD103 expressed on TRICs binding to E-cadherin on epithelial cells to ensure the tissue maintenance of TRICs,129 and cell communication mediated by secreted cytokines. For example, CD4+TRM cells can regulate the production of tumor necrosis factor-α (TNF-α) in CD8+TRM cells and promote the maturation of dendritic cells (DCs).70,130 IL-2 from CD8+TRM cells enhances the cytotoxic abilities of NK cells.130 In addition, the multilateral cellular interactions can lead to more complicated immune effects. Alveolar macrophages (AMs) guide CXCR3+lung-BRM cells to secondary infected sites by inducing the expression of CXCL9 and CXCL10 by other immune cells.131 CD69+CD4+lung-resident helper T cells promote the local retention of BRM cells and CD8+TRM cells after primary viral clearance, thus developing protective immune responses against secondary infection.132 Tissue-resident mast cells can induce fibroblast infiltration and interact with neutrophils to mediate wound repair and inflammatory processes.133,134,135,136,137 Furthermore, tissue-resident mast cells even participate in adaptive immune responses by serving as antigen-presenting cells (APCs) and recruiting and activating B cells and T cells.137,138,139
Overview of TRIC functions
TRICs perform essential regulatory effects on local tissues. Under physiological conditions, the fundamental functions of TRICs include maintaining tissue repair and homeostasis, defending against pathogens, and regulating local immune responses.
The canonical tissue homeostasis-related TRICs are cutaneous MAIT cells, tissue-resident macrophages, and mast cells. MAIT cells promote wound healing.140 Microglia, the brain-resident macrophages located in the central nervous system (CNS), contribute to brain homeostasis.141 Tissue-resident mast cells also mediate intestinal mucosal homeostasis and tissue remodeling.142,143,144,145 Liver-resident NK cells suppress liver fibrosis by removing the pathological hepatic stellate cell-derived myofibroblasts.146 Tissue-resident γδT cells secret the growth factor amphiregulin to mediate tissue repair.147,148 Even in the heart, CCR2−heart-resident macrophages express high levels of growth factors and extracellular matrix (ECM) to mediate tissue remodeling.149
Given their long-term tissue residency, TRICs are frequently exposed to pathogen stimulation. So it is rationally thought that TRICs have enduring immune defense capability.7 Studies have proven this issue in many cases, including CD8+TRM cells against herpesvirus (HSV), respiratory syncytial virus, and influenza infection,150,151,152 CD4+TRM cells against HSV-2,116 and trNK cells against mouse cytomegalovirus and influenza infection.153,154 BRM cells also provide durable protective responses against influenza virus infection by producing neutralizing antibodies,55 and tissue-resident mast cells eliminate parasites and bacteria to achieve tissue defense.139,155,156,157,158,159 TRICs mainly secret proinflammatory cytokines, including TNF-α, interferon (IFN)-γ, and interleukin (IL)-2 to kill pathogens and expand tissue inflammation.160,161,162
TRICs keep local immune balance and prevent excessive inflammation mainly through expressing inhibitory factors or immune checkpoint molecules. For example, TRM cells, γδT cells, and trNK cells have been found to express the inhibitory immune checkpoints in many tissues, such as programmed death receptor-1 (PD-1), cytotoxic T lymphocyte-associated antigen-4 (CTLA-4), T cell immunoglobulin and ITIM domain (TIGIT), lymphocyte-activation gene-3 (LAG-3), and T cell immunoglobulin domain and mucin domain-3 (TIM-3).112,163,164,165
In contrast to the tissue-adaptive functions of lymphoid TRICs, myeloid TRICs with compartmentalized distribution always have tissue-specific functions, especially macrophages. For example, human fetal microglia contribute to regulating neuron differentiation.99,166,167 Intestine-resident macrophages and mast cells can positively regulate blood vessel integrity, intestine motility, and the enteric nervous system.168,169,170,171,172 Bone-resident macrophages (Osteoclasts) participate in remodeling bone tissues and the progression of genetic skeletal disease,173,174 and bone-resident mast cells have been found to regulate bone metabolism.175,176 Moreover, Kupffer cells localized in the liver secret hepatocyte growth factors to promote liver regeneration.177 Fat-associated macrophages facilitate fat storage and control energy loss.73,178,179,180
In conclusion, each type of TRIC has both genetic and adaptive tissue-specific functions. Their dysregulation could be the pathogenesis of certain diseases.
Characteristics of lymphoid-lineage tissue-resident immune cells
Lymphoid-lineage TRICs include TRM cells, tissue-resident unconventional T cells, tissue-resident ILCs, trNK cells, and BRM cells. These lymphoid TRICs have effector functions and regulatory mechanisms similar to their circulative counterparts, whereas it is more interesting and significant to investigate the specific characteristics of tissue-resident lymphocytes.
Tissue-resident memory T cells
Based on the expression profiles comprising lymphocytes homing receptor CD62L, chemokine receptor CCR7, memory T cell marker CD45RA and CD45RO,181 and co-stimulatory receptor CD28, memory T cells are divided into CD45RA−CD45RO+CD62L−CCR7−CD28+effector memory T cells (TEM) and CD45RA−CD45RO+CD28+CCR7+CD62L+central memory T cells (TCM).182,183,184,185,186 TRM cells are thought to be differentiated from primary memory precursors or circulating precursor TEM with low killer cell lectin-like receptor subfamily G member 1 (KLRG1) expression, which undergoes activation and localization in peripheral tissues during the effector stage.187,188,189,190,191,192,193,194
TRM cells display distinct phenotypes from TEM cells and TCM cells. They highly express CD103, CD69, and CD49a, the representative markers of tissue residency, instead of expressing CD62L and CCR7.110,195,196,197 In brief, the classic TRM cell marker combination is CD45RA−CD45RO+CD103+/−CD69+/−CD49a+/−CCR7−CD62L−KLRG1−S1PR1−CD4+/CD8+. CD103 is the α subunit of αEβ7-integrin, binding to E-cadherin and helping T cells home to the epithelium and keep residence.195,198 Many studies have found CD103+CD8+TRM cells in epithelial tissues including skin,199,200 lung,201, and intestine.202 They are also in the SLOs, such as the spleen and lymphoid nodes.3,4,5 However, not all TRM cells express CD103, such as some CD4+TRM cells,200,203 so CD103 cannot be used alone to identify TRM cells. C-type lectin CD69 is an agonist of S1PR1 that is important for lymphocyte egress,70 thus CD69 negatively regulates the recirculation of T lymphocytes.204 Most TRM cells have a high level of CD69 and a low level of S1PR1.197,205 CD49a also plays an important role in the retention and survival of memory T cells in peripheral tissues.206,207
Parabiotic surgery shows circulating T cells of two congenic mice that are conjoined by the skin and exchanged blood will reach equilibrium within one week, whereas the resident T cells remain in the original position.73,208 But this approach cannot exclude the influence of local inflammation on circulating memory T cell recruiting.209,210 In vivo labeling by fluorochrome-conjugated antibodies combined with confocal imaging can also be used to distinguish the TRM cells and circulating T cells.211 However, this method is not fully suitable for identifying TRM cells in the red pulp of the spleen and liver sinusoids.80 Tissue transplantation can also be used to identify TRM cells, despite being restricted to surgical technologies.22 Parabiosis or in vivo labeling is not suitable for human TRM cell identification, but high-throughput sequencing technologies are available.91,92,93,94
With the regulatory support of intracellular mechanisms and local tissue environments, TRM cells undergo adaptive differentiation and keep tissue residency.212,213 First, epigenetic mechanisms regulate the expression of resident-related molecules, as the assay for transposase-accessible chromatin using sequencing (ATAC-seq) showed that the transcription start site region of Itage (encoding CD103) was accessible in TRM cells from the small intestine and salivary gland, as did the transcription start site of Ccr9 (encoding CCR9) in intraepithelial lymphocytes TRM cells.214
In addition, many animal studies showed that TF Hobit, Blimp-1, Notch, and Runx3 were essential for the differentiation and maintenance of TRM cells in the lung, small intestine, and salivary gland.101,127,214,215,216 Runx2 and Runx3 also induce the cytotoxic functions of CD8+TRM cells.217 TF Bhlhe40 preferentially regulates the fitness and function of CD8+TRM cells by programming mitochondrial metabolism.218 In contrast, Eomes and T-bet suppress TRM cell differentiation.124,219 Interestingly, TFs involved in regulating TRM cells perform tissue-specific modulation. For example, Blimp deficiency greatly blocks the formation of lung-CD8+TRM cells but hardly affects skin-CD8+TRM cells.49,101 However, most results and conclusions are based on animal models. The transcriptional regulations of human TRM cells require further explicit investigation.101
Transforming growth factor (TGF)-β, IL-33, and TNF are potent external cytokines that enhance tissue residency of TRM cells. These TFs inhibit TRM cell migration by downregulating KLF2 and S1PR1,114,220,221 and TGF-β also induces CD103 expression on TRM cells.179,193,222,223,224,225 IL-15 and IL-7 are also crucial for maintaining skin-TRM cells. Deficiency in either breaks the local immune homeostasis, inhibiting TRM cells and impairing their responses in contact hypersensitivity.226 In addition, TRM cells also secrete cytokines to modulate biological processes which will be described later.130,227
Chemokines/chemokine receptors are also involved in regulating the location and functions of TRM cells, including CCR6, CXCR3, CCR5, CCR8, CCR9, and their ligands.228 As the tissue-homing molecule, CXCR6 is widely expressed in TRM cells. CXCR6 recognizes CXCL16 produced by endothelial cells in the lung and skin mucosa to promote TRM cells to keep retention in these tissue sites.229,230 In addition, the CXCL10/CXCR3 axis is pivotal for HSV-specific CD8+TRM cell differentiation and increment in trigeminal ganglia.150 High CXCR3 is also upregulated on CD8+TRM cells in the liver.162 CCR9 expression promotes CD8+TRM cells to remain in the small intestine epithelium where the ligand chemokine ligand (CCL) 25 is abundant.231 The CCR9/CCL25 axis also contributes to CD103+CD8+TRM cell differentiation.232 CCR8 maintains TRM cells in human skin.233 These findings suggest the importance of chemokines in maintaining the tissue residency of TRM cells, thereby influencing their functional adaptation.
Antigen stimulation is another essential motivator for TRM cell priming,124 and the sites of stimulation greatly determine the location of TRM cells.211 For example, influenza virus vaccines effectively induce the aggregation of CD4+TRM cells and influenza-specific CD8+TRM cells within lung tissues, thereby conferring durable immune protection.234 Other localized viral infections and even house dust mite infections also induced CD103+CD8+TRM cells to differentiate and reside in the brain and skin in mice models.235,236,237 However, some studies indicated that antigen stimulation was not always required for CD8+TRM cell establishment, and even the persistent antigen stimulation inhibited the expression of CD103 in local T cells.70,224 Therefore, the roles of antigen stimulation in regulating TRM cell formation need to be further confirmed.
TRM cells express inhibitory exhaustion molecules to maintain immune homeostasis, including PD-1, CTLA-4, TIM-3, and LAG-3.238 These negative regulators can attenuate the immune responses induced by TRM cells to avoid overreactions. These checkpoint molecules are targets of immune checkpoint blockades (ICBs), suggesting that TRM cells can respond to ICB therapies.239,240,241
Tissue-resident unconventional T cells
Unconventional T cells (also termed innate-like T cells, ILTCs) are developed in the thymus.59 They are mainly involved in immune responses in non-lymphoid tissues, including iNKT cells, γδT cells, and MAIT cells.59,242 These ILTCs have TCRs with less diversity and recognize antigens without limitation of major histocompatibility complex (MHC)-related molecules,242 so they can make quick responses early when encountering antigen stimulation.243
Most γδT cells exist in the epithelium and mucosal tissues, and only a small number of γδT cells are present in circulation.244 Unlike TRM cells, tissue-resident γδT cells derive from double negative thymocytes during the embryonic period or postnatal thymus and directly seed into barrier tissues after maturing and exporting from the thymus,63,245 including skin,246 liver,247 and intestine epithelium.248 The known functions of tissue-resident γδT cells include defending against infection, maintaining homeostasis, and repairing tissues.248
iNKT cells develop in the thymus after birth. TCR coordinating with CD1d molecules expressed by CD4+CD8+double-positive thymocytes promotes the development of iNKT cells.249,250,251 iNKT cells express lymphocyte function-associated antigen-1, intercellular adhesion molecules-1, and tissue-resident molecule CD69.74,252 Given the different TF profiles and cytokine secretion, iNKT cells can be classified into three subtypes, iNKT1, iNKT2, and iNKT17 cells.111,253 T-bet regulates the development of IFN-γ-producing iNKT1 cells, while iNKT2 cells are regulated by GATA3 and mainly produce IL-4 or IL-13, independently of RORγt regulation. iNKT17 cells that produce IL-17 are regulated by RORγt.254,255 Additionally, BTB-zine finger TF, PLZF is also important for iNKT cell differentiation, and PLZF-deficient iNKT cells downregulate the secretion of granzyme B (GZMB), IL-4, IFN-γ, and TNF-α upon stimulation.256,257,258
Following the lineage commitment of iNKT cells, Hobit and Blimp-1 drive their tissue-resident transcriptional programs.122 iNKT cells show highly heterogeneous in peripheral tissues such as the skin, liver, lung, and adipose tissues without recirculating in the blood or lymphoid tissues.62,259,260,261,262,263,264,265,266 They can initiate a rapid immune response at the early stage of pathogenic lipid stimulation presented on CD1d and produce cytotoxic molecules or cytokines like IFN-γ, IL-17, and TNF-α. They also activate other immune cells, such as DCs and CD8+T cells, and further amplify the immune response.110,267 Therefore, they are one of the potent contributors to support barrier immunity.243,259,260
MAIT cells develop in the thymus after birth.60,61,268 Notably, iNKT cells and MAIT cells acquire tissue-resident transcriptional profiles before leaving the thymus.62 MAIT cells express CD161 and tissue-homing receptors, including CCR5 and CCR6, and they always aggregate in mucosal tissues to exert innate-like immune functions via non-TCR signals.269 However, the tissue-specific regulation of MAIT cells and γδT cells needs to be further explored, and it is still unknown how tissue-resident unconventional T cells renew in tissues.
Tissue-resident innate lymphoid cells
The development of ILCs and NK cells shares some conserved processes between mice and humans. However, human ILCs exhibit greater heterogeneity and NK cells are highly specialized with distinct transcriptional regulation and functions.67,270 In both species, innate lymphoid progenitors generally differentiate into NK cell precursors and common helper innate lymphoid progenitors. Then, NK cell precursors give rise to cNK cells that usually exist in circulation.270,271 However, human NK cells display more functional and tissue-specific diversity, including the CD56bright and CD56dim NK cell subsets with distinct cytotoxic functions, as well as trNK cells like decidual and liver NK cells, which are less prominent in mice.270 Common helper innate lymphoid progenitors further differentiate into innate lymphoid cell precursors that generate helper-like ILCs, including ILC1s, ILC2s, and ILC3s, and another progenitor that generates lymphoid tissue inducer.70,71,271,272,273 In humans, ILC subsets are more phenotypically and functionally diverse than in mice. For instance, ILC3 subsets show great heterogeneity, including NKp44+ and NKp44−populations, which are less defined in mice.270,274 ILCs do not express antigen receptors but regulate innate immunity and tissue remodeling.243,275,276
Helper-like ILCs have specific cytokine-sensitive pathways and intrinsic regulatory TF profiles, which serve to modulate their differentiation processes.275 ILC1s are induced by IL-12 and IL-18, regulated by the T-bet, and mainly produce IFN-γ and TNF-α to mediate immune defense against intracellular pathogens.277 ILC2s develop in response to IL-25 and IL-33, and their dominant TF is GATA3. ILC2s secrete cytokines associated with the Th2 population, including IL-4, IL-5, IL-9, and IL-13, to promote tissue repair and maintain tissue homeostasis.278 RORγt regulates ILC3s development and ILC3s produce IL-17, IL-22, TNF, and granulocyte-macrophage-colony-stimulating factor.70,279,280,281,282 ILC3s are important for defending extracellular bacteria and are associated with autoimmune diseases.67,283,284,285
According to a parabiosis investigation, the majority of helper-like ILCs are found in host SLOs and peripheral tissues, indicating their tissue-resident characteristics.68 Most helper-like ILCs can maintain themselves by self-renewal in the physiological states but depend on the supplement of adult bone marrow-derived or local precursors in a few cases.68,69,70,71 For example, upon acute infection, tissue-resident ILC2s proliferate locally and still keep self-maintenance. The hematopoietic progenitors partially supplement the tissue-resident ILC2s pool under inflammation resolution and tissue repair.68 Therefore, the constitution of the tissue-resident ILCs pool is not identical under different conditions.
Tissue-resident NK cells
NK cells are the core effector of innate immunity.286 The typical phenotypic markers of human NK cells are CD56 and CD16, and murine NK cells express NK1.1, NKp46, and CD49b.287 There are two NK cell subtypes, cNK cells, and trNK cells, entirely different in developmental trajectories, phenotypes, locations, and functions.83
Splenic cNK cells originate from bone marrow-derived progenitors in a T-bet and Eomes-dependent manner.75,76 trNK cells derive from various progenitors. They can originate from tissue-resident progenitors during the embryonic and adult stages, bone marrow-derived progenitors during adulthood, or even differentiated cNK cells.66,67,68,72 For example, liver-trNK cells originate from hepatic hematopoietic progenitor cells or hepatic stem cells.70,75 Lin−Sca-1+Mac-1+fetal-liver-derived hematopoietic precursor cells tend to develop into liver-trNK cells in the adult liver.72,288 Moreover, a study found that CD62L+Eomes+cNK cells could enter infectious sites and differentiate into Tcf1+CD69+trNK cells upon acute infection in the skin. Therefore, liver-trNK cells can maintain themselves in a bone-marrow-independent manner during adulthood. The mechanisms of renewal and maintenance of trNK cells in other organs are poorly understood.
trNK cells are widely distributed across many non-lymphoid tissues, such as the uterus, lung, intestine, skin, liver, adipose tissue, and salivary gland.76,153,273,289,290,291,292 Most of them express canonical markers of tissue residency, including CD69, CXCR6, and CD49a, whereas CD103 expression is not constitutive.76,293,294 The phenotypic and transcriptional profiles of trNK cells are highly heterogeneous within different tissues.85,96 Human liver-trNK cells can express CD56, CD69, CD49a, and CXCR6. Hobit and Blimp are the main TFs that regulate liver-trNK cell development.100,101,102 Lung-trNK cells express CD69 but selectively express CD49a or CD103.177,295 trNK cells located in the salivary gland upregulate both CD49a and CD49b, and T-bet and TGF-β signaling are indispensable for their differentiation.103,104 trNK cells in the uterine express CD94, NKG2A, CD49a, and CD103, and Eomes regulates their activation.296,297 Moreover, trNK cells downregulate the expression of lymphocyte egression markers, such as KLF2, S1PR1, and SELL. Some trNK cells also express immunosuppressive molecules PD-1, CTLA-4, and TIGIT.83
Therefore, the categories, localized development, and functions of trNK cells provide us with a better understanding of NK cell-involved immunity. However, their immune responses to stimulation especially those different from cNK cells need to be further studied.
Tissue-resident memory B cells
Naïve B cells migrate to SLOs from bone marrow and mature after interacting with CD4+follicular helper T cells and DCs, and then mature B cells differentiate into plasma and MBCs upon antigen stimulation. MBCs will differentiate into effective cells and offer humoral immune responses to antigen stimulation.298 The long-tissue residency characteristics of MBCs are reported in many studies. Influenza-specific B cells that highly express CD69 and CXCR3 in the lung can persist in the same sites for a long time after the primary infection and immediately differentiate and produce antibodies against reinfection.55,299,300 Researchers also found a novel subset of MBCs that express CD69 and CD45RB were predominant in the gut and spleen but did not exist in the blood.89 CD45RB is associated with MBC differentiation, and CD45RB+B cells can localize outside germinal centers.301 Therefore, CD69+CD45RB+MBCs are recognized as a unique subset of MBC cells, defined as BRM cells.89,302,303 BRM cells have distinct features from non-resident MBCs. Similar to TRM cells, BRM cells downregulate the expression of CD62L and S1PR1, whereas they upregulate CD69 and CXCR3.55,57,89,304
The origins of BRM cells in tissues are still unclear. A study revealed that GC in lymphoid organs, instead of iBALT, is the source of most lung-BRM cells,55,57 and the small percentage of MBCs in the lung originating from iBALT had a distinct phenotype from other BRM cells.55,305 Studies demonstrated that BRM cells form where B cells mediate immune responses to infection.56 Intranasal administration of the influenza virus can induce the recruitment of CXCR3+circulating MBCs by CXCL9 and CXCL10 to infected lung tissues, and these MBCs will develop into immunoglobulin A-producing BRM cells in the respiratory mucosa.90 Pneumococcus infection also induces CD69+BRM cells in the lung.58 Therefore, BRM cells may born in B cell-induced immune responses.
According to a recent study, BRM cells can be classified into two categories based on their antigen specificity and B cell receptor repertoire, termed “bona fide” BRMs and “bystanders” BRMs. The former cells have a special affinity for viruses that activates them to differentiate and function. In contrast, bystander cells cannot recognize viruses directly.304 Intriguingly, a study found that BRM cells can transiently migrate after the secondary infection in the lung. The chemokines secreted by other infiltrating immune cells recruit BRM cells to infected regions and amplify localized immune responses.131 However, the roles of BRM cells in other tissues or pathological status are unclear.
Characteristics of myeloid tissue-resident immune cells
Myeloid-lineage leukocytes perform innate immunity and regulate adaptive immune activity. Myeloid TRICs, including tissue-resident macrophages, TRNs, and tissue-resident mast cells, have been found in many tissues and organs.20,49,79 Here, we systematically summarize their development, maintenance, phenotypes, and functions.
Tissue-resident macrophages
Tissue-resident macrophages are a highly heterogeneous subgroup. Macroglia in the brain, Kupffer cells in the liver, AMs in the lung, nerve- and airway-associated macrophages in the lung mesenchyme, Langerhans cells in the skin, fat-associated macrophages in the adipose tissues, red pulp macrophages in the spleen, large peritoneal macrophage in the peritoneum, and osteoclasts in the bone are widely studied.169,306 The canonical markers of human classical macrophages are CD11b and HLA-DR.307 Different tissue-resident macrophages have specific phenotypic profiles, such as AMs express CD11c, CD14, CD206, and CD169,308 Kupffer cells express resident markers CD163, CD32, and macrophage receptors with collagenous structure but do not express CD169,309,310 as well as interstitial macrophages express HLA-DR and CD14.311,312 The functions of tissue-specific resident macrophages are also highly tissue-adaptive. In brief, they are responsible for tissue homeostasis, tissue repair, tissue remodeling, and regeneration.313
Some macrophages originate from yolk-sac or fetal-liver embryonic progenitors before HSC formally appears, and others originate from circulating monocytes derived from HSC in adults.21,23,24,25,314 Embryonic progenitors-derived macrophages are a distinct cell population from canonical macrophages in the mononuclear phagocyte system with specific developmental trajectories. There are three main hematopoiesis waves during the embryonic period of mice.25,315 The primary hematopoiesis was around the embryonic day (E) 7.5, which produces early erythroid myeloid precursors (EMPs). Early EMPs can further differentiate into C-Myb−CD45+F4/80brightCD11blow macrophages, which can migrate to the brain to form microglia.316,317,318 Brain microglia have been thought to originate exclusively from early yolk-sac-derived progenitors.31,98 Around E8.5, the second wave produces late EMPs. Late EMPs will colonize the fetal liver. Fetal-liver progenitors give rise to many kinds of tissue-resident macrophages, including Kupffer cells, Langerhans cells, AMs, etc.319,320 The third wave originated from HSCs produced in the aorta-gonad-mesonephros region at about E10.5, and these HSCs were transplanted into the fetal liver and differentiated into a variety of lineage blood cells, including CD45+F4/80lowCD11bhighmacrophages/monocytes.321,322 Furthermore, cardiac-resident macrophages can derive from progenitors in both primitive and definitive hematopoiesis during embryogenesis.323
Many studies indicated that the postnatal tissue maintenance of yolk-sac or fetal-liver-derived macrophages in the brain, liver, spleen, lung, peritoneal, pancreas, and BM are not or minimal dependent on the replenishment of bone marrow-derived HSC and circulating monocytes.23,29,30,31,32,78,79,324 However, sometimes HSC-derived monocytes do constitute significant parts in resident macrophage pools.38,39 Circulating monocyte-derived macrophages will acquire identical tissue-specific characteristics, functions, and the ability of long-term self-renew as embryonic progenitors-derived macrophages, such as in the lung and liver,31,39,43,44,45,46 which fully manifest the plasticity of monocytes. For example, a study found that the Kupffer cell niches would be replaced by monocyte-derived macrophages that were phenotypically and transcriptionally similar to embryonic progenitors-derived Kupffer cells after depleting embryonic progenitors-derived Kupffer cells by diphtheria toxin (DT) in the mice model.39 In the lung and bone, circulating monocyte-derived macrophages are also found.169,313 Furthermore, a study using scRNA-seq analysis, parabiotic experiment, and fate-mapping analysis defined three tissue-resident macrophage subpopulations in the liver, kidney, heart, brain, and lung, based on their representative gene markers and the way they maintain themselves in tissues.40 One macrophage subtype TLF+TIMD4+/−LYVE1+/−FOLR2+/− macrophages hardly depend on monocytes for maintenance, whereas another two macrophage types MHCIIhiTIMD4−LYVE1−FOLR2−CCR2−macrophages and TIMD4−LYVE1−FOLR2− CCR2+macrophages depend on monocyte-derived macrophage for renewing to varying degrees.40,41 In cancer, infiltrating monocytes and tumor-associated macrophages (TAMs) also express tissue-resident markers.311,325,326 Initially, the yolk-sac-derived macrophages were thought to be replaced by Ly6Chi monocyte-derived macrophages and were not present in the adult intestine.41 However, a study found the existence of self-maintaining gut-resident macrophages during the embryonic period, and they were not completely replaced by monocyte-derived macrophages after birth. This sample further confirms the complementarity between two macrophages with different origins.327 Hence, at least a partial population of monocyte-derived macrophages can be categorized as tissue-resident macrophages.
We think tissue-resident macrophages are composed of different ratios of embryonic and monocyte-derived macrophages, which is associated with the accessibility and availability of corresponding tissues. Take microglia as an example, which solely originates from yolk-sac-derived progenitors due to macrophages derived from the fetal liver and monocytes being impeded by the blood-brain barrier.31,328
The transcriptional profiles of tissue-resident macrophages in different tissues are determined by their lineage-determining factors and environmental signals.313 SALL1, SALL3, and MEIS3 mainly regulate microglia, and in Langerhans cells, it is RUNX3 and ID2 that do. Both of the two tissue-resident macrophages are regulated by TGF-β produced by epithelial cells and neurons.329,330,331,332 In AMs, colony-stimulating factor (CSF) 2 and TGF-β control PPARγ expression. In bone, the expression of NFATC1 in osteoclasts is regulated by RANKL, OPG, and TGF-β.329,330,331,332,333,334
The study of human macrophages poses significant challenges, primarily due to the lack of macrophages in early embryos and the exceeding difficulty in isolating specific macrophages. The study findings based on animal models cannot be confirmed in humans. Encouragingly, transcriptomics is utilized as a research method in this field. Many studies using scRNA-seq have disclosed the origin and lineage of macrophages, including the detailed single-cell profile of human embryonic macrophages, which clarified the origin of resident macrophages in many tissues.98 scRNA-seq also assists people in exploring novel macrophage populations.98,99,335 In addition, spatial transcriptomics, proteomics, and organoid cultures are useful in deciphering the functions and crosstalk of resident macrophages in tissues.336,337,338,339
Tissue-resident neutrophils
Embryonic neutrophils derive from definitive hematopoiesis,49,79, and adult neutrophils originate from HSCs in the bone marrow and migrate into tissues to provide immediate immune responses.340 The existence time of neutrophils in tissues is relatively short, which may explain why people pay less attention to how neutrophils acquire tissue-specific properties and their efforts for tissue residency.341,342
However, short-lived neutrophils depend on different mechanisms to reside in tissues and show distinct functional phenotypes.343 The analyses using mass cytometry and high-throughput sequencing have identified CD11b+Ly6G+neutrophil clusters that acquired tissue-specific phenotypes in different organs, including bone marrow, spleen, blood, skin, intestine, and lung. Regardless of the short time (about one day) that these neutrophils exist in tissues,49 neutrophils entering tissues also show plasticity in adapting to local environments. The results of parabiosis and transfer experiments indicated that neutrophils would quickly adopt similar characteristics as the cells that were initially present in this tissue after migrating into tissues.49 Therefore, we named those neutrophils with similar tissue-resident phenotypes to other TRICs as TRNs.
Neutrophils in the lung are the representative example of TRNs. Lung-resident neutrophils possess distinct transcription profiles and surface markers that help distinguish them from bone marrow and blood-circulating neutrophils.51 For example, neutrophils isolated from the lung express higher CD11b and CXCR4 but lower CD62L than neutrophils derived from the blood and bone marrow. They also upregulate the mRNA expression levels of Cd14, Plet1, and Sca1/Ly6a, which demonstrate how they differ from other origins-derived neutrophils.51 In addition, many factors within lung tissues promote neutrophil development, such as prostaglandin E2 (PGE2).51 Moreover, PGE2 prolongs neutrophil persistence by increasing the expression of antiapoptotic factor Nr4a2 and Nr4a3, and strengthens ROS production.50,51 The CXCL12-CXCR4 axis also promotes the retention of neutrophils in the lungs and inhibits their circulation.52 However, the existence of TRNs in other tissues is less defined.
In the tumor microenvironment (TME), the scRNA-seq of NSCLC tumor tissues identified two kinds of TRNs, including tumor-associated neutrophils (TANs) and normal adjacent tissue-associated neutrophils.84 TANs downregulate CXCR2 and SELL, the markers of mature neutrophils, whereas normal adjacent tissue-associated neutrophils highly express them.84 The common markers of TANs include OLR1, VEGFA, CXCR4, and CD83.84 These TANs are deeply involved in TME reshaping and regulation of cancer immunity.
Although studies focus on the regulatory mechanisms and biological significance of tissue-resident neutrophils are less, we think that investigating this neutrophil subpopulation will help us understand the role of neutrophils in diseases, especially infection and cancer.
Tissue-resident mast cells
Mast cells are distributed in many organs, especially the respiratory tract and digestive tract where they are directly exposed to external environments.344 In rodent animal studies, tissue-resident mast cells can be divided into two subpopulations, connective tissue mast cells (CTMCs) and mucosal mast cells (MMCs).345
Embryonic CTMCs derive from three different hematopoietic progenitors in succession distributing into many tissues for maturation before birth.26,27,28 Postnatal CTMCs in most connective tissues mainly derive from late EMPs, whereas early EMPs specifically give rise to CTMCs in adipose tissue and pleural cavities.20 MMCs are mainly distributed in the epithelium of the respiratory tract and gastrointestinal tract. They mostly derive from definitive hematopoietic progenitors and adult cecum and utero MMCs can be replaced by bone marrow-derived HSCs.20,37
The adult skin-resident mast cells and utero-resident CTMCs minimally depend on bone marrow HSCs-derived cells for renewal.33,34,35,36,37 In contrast, bone marrow-derived mast cell progenitors partially get involved in the skin-resident mast cell pool and adaptively acquire CTMC phenotypes in the atopic dermatitis-like skin model.42 In addition, blood mast cell progenitors can also be recruited into the lung and gut to reside.35,346,347 However, the specific constitutions and the regulatory mechanisms of adult mast cells in different tissues are not confirmed. MMCs persistently reside in tissues for a relatively shorter life than CTMCs.348,349,350 A study explained that the Fcγ-induced cell apoptosis mechanism made MMCs more prone to apoptosis, seemingly resulting in the different tissue persistence of two mast cell populations.351 Especially, MMCs can migrate to infectious niches and expand.352,353,354,355 Therefore, we speculate that MMCs usually exist in tissues that are frequently exposed to allergens and pathogens, which also promote the short persistence of MMCs. However, the origin and development of human tissue-resident mast cells are unclear.
scRNA-seq analyses help people find more tissue-resident mast cell subtypes with high heterogeneity and plasticity in different tissues, such as skin, esophagus, trachea, and peritoneum.87,135,144,345,356,357,358,359,360,361 For example, skin-resident mast cells mainly upregulate genes involved in the development signaling pathway, including Lgr4, Smo, and Igf2r, and also TF SOX7.87,349 Mcpt1 and Mcpt2 are expressed in esophagus-resident mast cells.349 Lipf is enriched in tracheal-resident mast cells, and Itgb2 and Bmp2 are upregulated in peritoneal-resident mast cells.349 In addition, mast cells distributed in different tissues can produce different factors even during embryogenesis, such as granulocyte-macrophage-colony-stimulating factor (GM-CSF), vascular endothelial growth factor (VEGF), and platelet-derived growth factor (PDGF), which may contribute to specific tissue development.87
Tissue-resident mast cells can secrete cytokines, chemokines, and growth factors to regulate physiological or pathological processes, such as regulating angiogenesis by secreting VEGF, TGF-β, and CXCL18, and promoting vascular and blood-brain barrier permeability and facilitating fibrosis of chronic diseases dependent on TGFs and fibroblast growth factors.362 In addition, mast cells also mediate wound healing and nerve regulation.363,364,365 Although the role of mast cells in allergic inflammation is familiar, whether mast cells dominate other immune events such as its effects on cancer immunity is unclear.363,366,367,368
Roles of TRICs in diseases
Given the wide existence and crucial functions of TRICs in tissues, the investigation of TRICs in pathological processes is important for deepening our understanding of pathogenesis and disease therapies. Here, we delineate the roles of TRICs in tissue homeostasis and repair, autoimmune diseases, infectious diseases, cancer, and others (Table 1).369,370,371,372,373
Tissue homeostasis and repair
Tissue-resident myeloid TRICs are the dominant regulators in tissue homeostasis, covering tissue repair, inflammatory damage inhibition, and stability of tissue architecture.
Tissue-specific macrophages have adaptive functions linked to their environmental backgrounds.374 They can resemble sensors and react to many environmental signals, such as pH, temperature, hypoxia, and stress, thus maintaining tissue homeostasis.313 In the process of tissue repair, macrophages can suppress neutrophil activation and inflammatory damage, and recruit myofibroblasts that contribute to tissue remodeling.375,376,377 For example, Kupffer cells participate in immune tolerance by suppressing CD4+ and CD8+T cell activation,378,379 triggering antigen-specific CD8+T cell apoptosis,380 and promoting the expansion of regulatory T cells (Tregs).379 In the gut, resident macrophages maintain intestinal functions and homeostasis. The depletion of this cell population will result in neuronal apoptosis and blood vessel leakage, decrease neuron secretion, and impair intestine motility.168,327 In addition, tissue-resident macrophages also participate in regulating homeostatic energy expenditure and heat generation by controlling sympathetic innervation in brown adipose tissues. Mecp2 deletion in adipose-resident macrophages will disrupt their cross-talks with neurons, resulting in decreased production of thermogenic factors by adipocytes, which are controlled by sympathetic nerve axons.178 Therefore, tissue-resident macrophages are multifunctional across different tissues.
Studies showed that tissue-resident mast cells regulated hair follicle growth, bone metabolism, and mucosal integrity in the intestine.142,143 The bone quality and strength of the mice with mast cell elimination were impaired.175,176 Intestine-resident mast cells contribute to maintaining mucosal barrier homeostasis by regulating epithelial cell activity and mucosal integrity.144,145 They also reverse tissue injury and promote wound healing.381 In wound sites, mast cells produce IL-4 and nerve growth factors to induce the proliferation and migration of fibroblasts to facilitate tissue healing.133,134,135
As for TRNs, some studies reported that they function in regulating vascular growth and repair in the lung and intestine, as well as hematopoiesis.49,342,382,383
Lymphoid TRICs are also involved in regulating tissue homeostasis, although few studies focus on this issue. An example is the regulatory feedback loop between ILC2s and stromal cells in white adipose tissues. ILC2s induce tissue-resident multipotent stromal cells to produce eotaxin and to recruit eosinophils for improving tissue homeostasis.384 Concurrently, white adipose tissue-resident mesenchymal stromal cells promote ILC2 to produce type 2 cytokines and similarly facilitate eosinophil recruiting.385 In addition, tissue-resident iNKT cells maintain adipose tissue homeostasis by inducing anti-inflammatory phenotypes of macrophages and suppressing Treg cell activation.258
Autoimmune disease
The overactivation of immune systems will cause exacerbated inflammation and extensive organ damage, finally leading to the initiation of autoimmune diseases.386,387 However, the pathogenic mechanisms of autoimmune diseases are not comprehensively studied. Many studies have investigated that TRICs are closely associated with the development of autoimmune diseases, and some of them will become potential therapeutic targets (Fig. 4).8,388,389

The regulatory roles of TRICs in autoimmune diseases. The red “+” means that the TRIC type mainly plays an inflammation-promoting role in corresponding autoimmune diseases, which will exacerbate disease progression. The green “-” means that the TRIC type can inhibit immune activity to attenuate diseases. RA rheumatoid arthritis, PsA psoriasis arthritis, MS multiple sclerosis, LN lupus nephritis, ANCN-associated GN anti-neutrophil cytoplasmic antibody-associated glomerulonephritis, AIC autoimmune cholangitis, AIH autoimmune hepatitis, IBD immune bowel disease, pSS primary Sjögren’s syndrome. This figure is created with BioRender.com
TRICs in autoimmune arthritis
Rheumatoid arthritis (RA) is an autoimmune disease with high incidences. The most common drug is disease-modifying antirheumatic drugs (DMARDs),390,391 though not all patients respond well to treatment. Therefore, it is important to explore new therapeutic targets.392 A better understanding of the pathological mechanisms of RA initiation and recurrence may help break through the barrier. Notably, the long-term tissue-resident properties of TRICs contribute to the pathology of RA.
CD8+T cells have been found to exist in the synovial fluid of RA patients for a long time. They exhibit canonical phenotypic of TRM cells, such as upregulating the expression of CD69, CXCR6, and CD49a, and downregulating S1PR1 and KLF2.393,394 In addition, These CD8+TRM cells prefer to uptake fatty acid and display enhanced activities in response to stimulation.393 The long-term resident synovial CD8+TRM cells can recruit circulating effector T cells by releasing CCL5 to aggravate joint injury and RA recurrence.393,395,396 Some studies also indicated that TRM cells may promote the recurrence of RA by perforin-mediated histone citrullination and the production of anti-citrullinated protein antibodies, which are the main characteristics of RA pathogenesis.394,397,398,399 Therefore, TRM cells may be the key regulator of RA recurrence, and their features are reversely influenced by the tissue environments of RA. CD8+T cells with similar phenotypes of TRM cells are also found in the tissues of psoriatic arthritis (PsA) patients, such as synovial fluid and the skin.19,199,400,401 However, their roles in regulating PsA are not confirmed.
The local composition of CD8+TRM cells in PsA and RA are distinct. The proinflammatory type 17-like CD103+CD69+CD8+TRM cells are specifically aggregated in the synovial fluid of PsA, whereas cytotoxic and Treg-like CD8+TRM cells are found in both RA and PsA.19 Whether the different phenotypes of TRM cells in the two arthritis attributed to the plasticity of TRM cells in different environments and the specific environmental stimulators needs further validation.
The roles of tissue-resident macrophages in RA have also been studied. In contrast to circulating monocyte-derived macrophages that mainly promote inflammation in RA,402,403,404 a recent study demonstrated that CX3CR1+ synovial lining-resident macrophages exhibited characteristics of epithelial cells. These tissue-resident macrophages limit joint inflammation by constructing a membrane-like immunological barrier in the synovial lining.405,406,407,408 Furthermore, after accessing the synovial niche, circulating monocytes can differentiate into tissue-resident macrophages, which mitigates the chronic inflammation of RA.409 These studies suggest the effector roles of tissue-resident macrophages and TRM cells in RA, but further studies on manipulating these TRICs to treat autoimmune arthritis are needed.
TRICs in skin-autoimmune diseases
Autoimmune skin diseases include psoriasis, vitiligo, et al. Many studies also revealed that TRIC subpopulations participate in the pathogenetic process of these diseases.410
Psoriasis is a chronic, relapsing autoimmune skin disease induced by IL-17A.411,412,413 The existence of TRM cells usually aggravates psoriasis. CD8+TRM cells have been identified to exacerbate skin inflammation in the psoriatic skin by producing IL-17A.414,415,416 Moreover, IL-22 produced by CD69+CD4+TRM cells also enhanced psoriatic inflammation by targeting epidermal keratinocytes, a predictive feature of psoriatic skin.417,418 Therefore, strategies for eliminating TRM cells may be effective in treating psoriasis.
Vitiligo is caused by the loss of melanocytes, and the main clinical manifestation includes recurrent depigmented patches.419 CD103+CD69+CD8+TRM cells with high levels of IFN-γ and TNF-α production have been identified in the skin lesions.420,421 These TRM cells synergize with circulating memory T cells to exacerbate the disease.422 Another study indicated that melanocyte-specific CD103+CD69+CD49a+CD8+TRM cells produced IFN-γ after recognizing Melan-A to perform cytotoxic killing.415 Interestingly, these CD8+TRM cells express CD122, a subunit of the IL-15 receptor, and anti-CD122 antibody treatment ameliorates the vitiligo lesion in the mice model.18 In addition, JAK inhibitors can reverse the depigmentation of vitiligo lesions due to the JAK/STAT signaling pathway being the driving pathway for cytokine production in TRM cells, which stresses that TRM cells are one of the feasible therapeutic targets of vitiligo.421
TRICs in nervous system-autoimmune diseases
Autoimmune diseases involving the nervous system are more harmful. The complicated pathogenesis and limited treatment strategies lead to unfavorable treatment outcomes and poor prognoses of patients.
The scRNA-seq studies showed that CD8+TRM cells, CD4+TRM cells, and trNK cells existed in the cerebrospinal fluid of MS patients.423,424,425 CD8+T cells infiltrating in the recurrent MS lesions display tissue-resident phenotypes, contributing to the compartmentalized inflammation in the brain.426 Mechanistically, TRM cells interact with other immune cells to develop their tissue-resident characteristics in the CNS. TGF-β released by Treg cells promotes the CD103 expression in the brain-infiltrated T cells.427 Moreover, PD-L1+glial cells combine with brain-CD103+CD69+CD8+TRM cells expressing PD-1, and promote their development.428 CD8+TRM cells can trigger pathological neuroinflammation and impair neurons with the assistance of CD4+T cells.429,430 Moreover, CD103+CD69+CD49a+CD8+TRM cells located in the white matter lesions of progressive MS have been demonstrated to promote inflammatory activity.431 Therefore, CD8+TRM cells are likely to be the pathogenic factors for MS progression.
T-bet-dependent NKp46+ILCs also get involved in driving CNS autoimmune disorders. They establish a proinflammatory environment for T helper type (Th)17 cells by secreting cytokines and matrix metalloproteinases, and all of these biological factors promote the migration of Th17 cells into CNS parenchyma.432 In contrast, tissue-resident ILC3s can suppress neuroinflammation by forming immune tolerance and eliminating autoimmune T cells.433 MAIT cells also promote MS progression by producing IL-17.434 The non-myeloablative autologous hematopoietic stem cell transplantation limited the proinflammatory process in MS by depleting IL-17-producing MAIT cells.435 These results suggest that IL-17-related lymphocytes are one of the drivers for CNS autoimmune diseases.
Microglia plays a dual role in nervous system-related autoimmune diseases.436,437 Regarding disease suppression, they can generate anti-inflammatory cytokines in favor of the differentiation of immunosuppressive cell populations such as Treg and Th2 cells, and display protective roles in neurological disorders.438,439 However, activated microglia will damage neurons in some cases. Microglia release harmful molecules such as reactive oxygen and nitrogen species and glutaminase,440,441, or secrete inflammatory molecules such as TNF-α.442 Moreover, they also promote the differentiation of pathogenic CD4+T cells, Th1, and Th17 cells which facilitate the progression of experimental autoimmune encephalomyelitis, a general model for studying MS.443,444 Blocking microglia alleviated the experimental autoimmune encephalomyelitis.445,446 Microglia also serve as APCs to present myelin antigens to activate T cells and then motivate the intense inflammatory responses.447,448 Therefore, microglia is recognized as the pathogenic participant in MS.
TRICs in digestive system-autoimmune diseases
TRICs also mediate the initiation and progression of several kinds of inflammatory bowel disease (IBD). CD103+CD69+CD4+TRM cells were found in the mucosa of patients, and the density of them was positively correlated with disease severity.449 Intriguingly, the deletion of Hobit and Blimp, the important TFs for the development of TRM cells, indeed induced IBD remission.449 Activated MAIT cells derived from IBD patients also exhibit high IL-17-secreting phenotypes, related to aggravating inflammation.450
Liver-CD8+TRM cells are found to induce the apoptosis of intrahepatic small bile duct epithelial cells and contribute to autoimmune cholangitis. Interestingly, they can express PD-1, and PD-1-targeted chimeric antigen receptor (CAR)-T therapy eliminated these CD8+TRM cells and achieved cholangitis remission.95 Scientists found the enrichment of TGF-β, IL-15, and E-cadherin in the liver of autoimmune hepatitis (AIH) patients, which orchestrated tissue residency of CD8+TRM cells.451 The infiltration levels of CD8+TRM cells are positively related to disease severity. Long-term tissue inflammation will lead to chronic organ fibrosis. MAIT cells have been found to promote liver fibrosis by producing IL-17A to activate hepatic stellate cells, an important component of liver fibrosis.452
TRICs in autoimmune nephritis
Lupus nephritis (LN) is a common complication of systemic lupus erythematosus.453 CD103+CD69+CD8+TRM cells are found to infiltrate the kidneys of LN patients or MRL/MpJ-Faslpr/J (MRL/lpr) mice. These cells proliferate in situ to maintain themselves and elicit renal inflammation and flare.454,455 However, their disease-specific phenotypes are unclear. It was found that JAK/STAT signaling was important for the maintenance of renal TRM cells in MRL/lpr mice, and the application of tofacitinib, an inhibitor of JAK/STAT signaling, attenuated kidney inflammation.455 In addition, the depletion of kidney tissue-resident macrophages also inhibited inflammation by reducing the recruitment of monocytes and factors that encouraged localized B cells to produce autoantibodies.456
Moreover, the scRNA-seq profile of anti-neutrophil cytoplasmic antibody (ANCA)-associated glomerulonephritis (GN) showed higher infiltration levels of TRM17 cells than health.106 Genes associated with T cell activation, proliferation, and cytokine production were upregulated in the TRM cells derived from ANCA-associated GN.106 Therefore, CD8+TRM cells at least partially contribute to kidney autoimmune diseases.
TRICs in other autoimmune diseases
A study on primary Sjögren’s syndrome (pSS) indicated a cellular crosstalk dominated by TRICs to mediate autoimmune damage. CD11bhigh tissue-resident macrophages secreted CCL22 to chemotaxis CD4+T cells with upregulated IFN-γ production, which aggravated local inflammation. Anti-CCL22 antibodies alleviated the progression of pSS.457 Similarly, the depletion of CD103+CD8+TRM cells by blocking CD103 is also helpful for SS inhibition.97,458 In contrast, salivary gland-trNK cells participate in attenuating SS progression. They may play protective roles in SS, fighting against the immune responses mediated by T cells.459
Cardiac tissue-resident macrophages are reported to promote autoimmune vasculitis by activating the CCL2/CCR2 axis, which can start the cascade of proinflammatory events as well as recruit circulating CCR2+monocytes and neutrophils.460
There remain some undetermined effects of TRICs in autoimmune diseases. For example, CD103+CD69+CD8+TRM cells have been identified as the predominant T cell population in the insulitic lesion of type 1 diabetes mellitus rather than classical cytotoxic CD8+T cells, which indicates the potentially important roles of TRM cells in diabetes mellitus.461 Studies on uterus-related autoimmune diseases showed that uterus-γδT cells upregulate IL-17 production after being activated, which may cause disease progression.105,462,463
Taken together, TRICs play inhibiting or promoting roles in different autoimmune diseases. However, limited to the research technologies and sample access, the available elucidation of TRICs in autoimmune diseases is still not enough to support clinical transformation.
Therapies targeting TRICs in autoimmune diseases
Repressing the excessive immune response is the most critical strategy for autoimmune disease treatment. We propose that the combination of routine immunosuppressive therapies with TRIC-targeting therapies may help amplify therapeutic efficacy.
First, blocking the critical molecules that contribute to the formation and maintenance of TRICs is feasible. For example, CD103 is the representative marker for most lymphoid TRICs, and a study found that CD103 inhibition remarkably alleviated the progression of SS.97 Fatty-acid-binding proteins 4 and 5 (FABP4/5) are indispensable for CD8+TRM cell survival, and their deletion also impair the formation of CD8+TRM cells.464
Second, therapeutic antibodies or small molecule inhibitors targeting effector cytokines or receptors of TRICs are implementable. Such as the anti-CD122 antibody achieves vitiligo remission.18 In the Lck-iDTR-engineered murine model with all T cells expressing DT receptors, synovial TRM cells are susceptible to DT-mediated depletion. The intra-articular injection of DT selectively depletes synovial TRM cells, resulting in attenuated joint flares in RA.393 In addition, IL-17A produced by CD8+TRM cells contributes to local inflammatory injury, and patients with PsA, autoimmune kidney disease, and psoriasis may benefit from IL-17-blockade therapy.19,106,411,412
Taken together, TRICs are potential candidates as disease-predictive biomarkers or therapeutic targets.
Infectious disease
TRICs located in peripheral tissues are one of the major components of infection defense barriers. TRM cells, BRM cells, tissue-resident unconventional T cells, and trNK cells are induced after infection, and they keep long-term immune memory and generate potent immune responses against the reinfected pathogens. Mechanistically, activated TRICs will produce cytotoxic molecules to kill pathogens directly, or secrete cytokines and chemokines to recruit circulated immune cells to infected sites. Moreover, the primary localized myeloid TRICs also construct the defensive line at the environmental interface. Here, we provide an overview of TRICs in infectious diseases (Fig. 5).

The anti-pathogen mechanisms of TRICs in infectious diseases. Upon pathogen reinfection, TRICs cooperatively perform immune defense functions. a After being stimulated by pathogens, CD4+TRM cells will rapidly produce IFN-γ, CCL2, and CCL7 to recruit circulating neutrophils and monocytes, which produce NO and ROS to defend against pathogens. CD8+TRM cells perform potent immune responses by producing IFN-γ and TNF-α. They also directly kill pathogens with perforin and GZMB. The high level of IFN-γ-inducible CXCL9 and CXCL10 in infected sites recruit circulating memory T cells to amplify local immune responses. b Upon bacterial reinfection, AMs and tissue-resident ILC3s quickly initiate the recruitment of circulating neutrophils and monocytes into the infected sites, while tissue-resident ILC3s further promote AM accumulation. Moreover, AMs enhance phagocytotic function and the autophagic clearance capacity by producing ROS, and they generate NO to directly kill bacteria. In addition, tissue-resident γδT cells, CD4+TRM cells, CD8+TRM cells, and MAIT cells also play important anti-bacterial roles by secreting regulatory cytokines or cytotoxic molecules. BRM cells can make rapid humoral immunity to resist bacteria reinfection. c Upon hepatitis virus infection, CXCL16 produced by hepatocytes and hepatic sinusoid epithelial cells promotes liver-resident CD8+TRM cells and γδT cells to retain in the liver. The two lymphoid TRICs exhibit potent immune responses against the HBV (Left). In the case of lung viral infections, multiple lung-TRICs cooperate to perform immune defenses. Lung-resident NK cells and iNKT cells highly produce IFN-γ, TNF-α, and cytotoxic molecules perforin and GZMB. Moreover, AMs produce chemokines to recruit circulating T cells and NK cells from the blood to enhance local immunity. AMs phagocytose virus-infected cells and defend against viruses via type I IFN. They also promote the recruitment of CXCR3+BRM cells by cooperating with neutrophils and monocyte-derived macrophages. After aggregating in infected sites, CXCR3+BRM cells differentiate into plasma cells to produce specific antibodies in response to virus infection. Moreover, TRM cells partially control the activation of innate cells in anti-viral protective immunity by producing IFN-γ (Middle). Upon HIV infection, CD8+TRM cells defend against HIV-infected CD4+T cells by producing IFN-γ and CD107a. In contrast, tissue-resident macrophages and CD4+TRM cells promote HIV survival and induce long-term infection (Right). AM alveolar macrophage, HBV hepatitis infection, HIV human immunodeficiency virus, IFN-γ interferon-γ, TNF-α tumor necrosis factor-α, GZMB granzyme B, IL interleukin, CCL C-C motif chemokine ligand, CXCL C-X-C motif chemokine ligand, NO nitrogen oxide, ROS reactive oxygen species, iNOS inducible nitric oxide sythase. This figure is created with BioRender.com
Viral infection
Viruses are the most common pathogens that TRICs encounter. In this subsection, we introduce the roles of TRICs in several common or serious viral infections.
Influenza infection
Influenza infects the host through the respiratory tract where AMs are involved in establishing the first protective line. AMs initiate the infectious defense against the influenza virus by producing type I IFNs and recruiting circulating monocytes into infected sites.465,466 RNA-seq analysis also validated that AMs developed proinflammatory phenotypes during influenza infection.107 During the viral pneumonia resolution phase, the placenta-expressed transcript 1 generated by AMs aids in healing lung damage by promoting the regeneration of the lung epithelial barrier.467 However, chronic influenza infection also induces immune tolerance by damaging the migratory and phagocytotic capacities of AMs towards invasive bacteria, ultimately increasing the possibility of secondary bacterial infection.107,468,469,470 Researchers also detected a unique monocyte-derived AM population in mice with influenza infection. Although this cell population is distinct from resident AMs, they can offer prolonged protective immunity against bacterial infection by increasing IL-6 production.471 Therefore, the complicated lung-AMs pool significantly inhibits influenza infections.
The existence of virus-specific TRM cells in the lung after influenza infection has been validated in patient samples and animal models.211,472,473,474 TRM cells quickly expand and perform immune responses against invasive influenza by producing IFN-γ, TNF-α, and GZMB.151,473,475 Notably, influenza-specific CD8+TRM cells can offer cross-protection defense against various influenza virus strains, including influenza A, B, and C, which provides the basis for developing a broad-spectrum influenza virus vaccine.475,476,477,478 However, a study found that CD8+TRM cells aggregating in the aged lungs were dysfunctional and pathogenic. These CD8+TRM cells led to excessive inflammation and lung fibrosis.479 This finding suggests that CD8+TRM cells play bi-directional roles at different life stages.480,481 In addition, BRM cells are also induced by influenza stimulation in the lung. They differentiate into plasma cells and further perform humoral immunity upon virus rechallenge.55,56,90 Interestingly, innate and lymphoid TRICs can coordinate with each other to protect the host from influenza infection. For example, AMs promote BRM cells to migrate into secondary influenza-infected sites to enhance anti-virus immunity.131 Moreover, IL-18 produced by AMs contributes to the establishment of CD103+CD8+TRM cells and the expansion of CD8+TRM cells upon influenza reinfection.482 Conclusively, the development of vaccines that can induce TRM and BRM cells to amplify host defense against virus infection is a promising strategy.
Furthermore, other lymphoid TRICs also play important roles in protective immunity against influenza infection. First, lung-resident ILCs maintain lung homeostasis after influenza infection.243,483 The loss of lung-resident ILCs compromises the integrity of the lung epithelium barrier, but this can be reversed by replenishing the amphiregulin generated by lung-resident ILCs, a ligand for the epidermal growth factor receptor.483 Lung-ILC1s make rapid response upon influenza infection by producing IFN-γ at the early stage.484 Lung-ILC2s prolong the survival of influenza-infected mice by secreting IL-5 and further recruiting eosinophils at the late infectious stage.485,486,487 In addition, CD49+CD103+CD69+lung-trNK cells induced by the influenza A infection also enhance cytokine production targeting virus-infected cells in the lung.154,289,488 Tissue-resident γδT cells also eliminate influenza infection by producing IFN-γ.489 Therefore, lung-specific TRICs perform critical immune defense functions in controlling influenza infection. They are likely to be novel targets for influenza prevention and control.
HSV infection
HSV is the neurotropic herpesvirus that mainly invades the skin and mucosal tissues of the mouth, eyes, and reproductive tracts. HSV has two types, HSV-1 and HSV-2. The most common manifestation of HSV infection is mucocutaneous injuries, and some patients also have encephalitis.490 HSV-1 is the general leading cause of gingivostomatitis and keratoconjunctivitis, whereas HSV-2 preferentially leads to genital herpes.491 Mucosal immunity is the most critical defense against HSV infection. Given the wide distribution of TRICs in mucosal tissue sites, many studies focused on the protective roles of TRICs in HSV infection.
Microglia are involved in fighting HSV-1 infection.492 They resist HSV infections by producing proinflammatory cytokines like type I IFN and recruiting circulating monocytes into infected sites.493,494 Microglia deletion will aggravate herpes simplex encephalitis.495 However, microglia overactivation also impairs surrounding neurons and causes severe herpes simplex encephalitis.496,497,498,499 Hence, the moderate activation of microglia will have maximum inhibitory effects.495
TRM cells also respond to HSV infection rapidly.500,501 For example, CD4+T cells and CD8+T cells isolated from endocervical biopsy tissues of HSV-2 infected patients show tissue-resident phenotypes, and they defend against HSV by producing IL-4 and IFN-γ.502 In addition, people found CXCR3+CD8+TRM cells and CD69+CD14+CD8+TRM cells in recurrent ocular HSV-1 infection and vagina HSV-2 infection respectively.116 In the genital mucosa, CD69+CD103−CD4+TRM cells produce IFN-γ against HSV-2 infection.116 Therefore, virus vaccines promoting the formation of TRM cells in mucosal sites are significant for controlling HSV infection.
Human immunodeficiency virus (HIV) infection
HIV is the pathogen of acquired immunodeficiency syndrome (AIDS), which causes a huge burden of disease worldwide.503 Mucosal tissues are the primary barrier to preventing HIV invasion.504 HIV replicates in mucosal sites, and HIV-specific CD8+T cells are the main effector for controlling HIV infection.505,506 Many studies have validated the presence of HIV-specific CD8+TRM cells in the female genital tract,507,508 lymphoid tissues,509 gastrointestinal tract,510,511 and these cells can fight against HIV by performing polyfunctional responses, including producing IFN-γ, TNF-α, and other cytokines or chemokines, rather than enhancing cytotoxic capacity.510,512,513 In the mice model, the reactivated TRM cells can upregulate the local concentration of passively transferred anti-HIV antibodies in the genital mucosa.514 Except for TRM cells, in neurocognitive disorders caused by HIV infection, microglia potentiate neurotoxicity by raising glutaminase production, which in turn causes direct neuronal dysfunction and death.440
In addition, some subsets of TRICs are associated with persistent HIV infection. CD4+TRM cells and tissue-resident macrophages are the main reservoirs of persistent HIV infection, which support HIV replication and the progression toward AIDS.515,516,517,518,519 Adipose tissue has been recognized as the latent location for HIV infection.520 Staying in HIV-infected subcutaneous adipose tissues, Treg cells and PD-1+CD4+TRM cells construct an immunomodulatory milieu that protects HIV persistence.521
Hepatitis virus infection
Hepatitis virus is the pathogen of viral hepatitis, and it has six subtypes. The most common hepatitis virus subtypes include hepatitis A virus (HAV), HBV, and HCV.522 Different from HAV triggering acute liver inflammation, HBV and HCV can cause chronic liver inflammation which may progress to liver cirrhosis or even malignancies.523 Studies have found that liver-resident TRICs extensively participate in the pathogenic processes of chronic liver inflammation caused by hepatitis virus infection. The intrahepatic CD103+CD69+CD8+TRM cells can rapidly expand and produce a large amount of IFN-γ and TNF-α in response to HBV and HCV infection,162,524,525 Liver-resident γδT cells are also involved in HBV cleaning.526 However, whether their anti-virus activities harm normal liver parenchyma is yet to be determined.
Persistent HBV infection is an important contributor to hepatocellular carcinoma (HCC). Notably, HBV-specific CD8+TRM cells congregate in the livers of patients with HCC. These cells play cancer-inhibiting roles and are positively correlated with the patients’ relapse-free survival.527,528 Therefore, virus-specific CD8+TRM cells may serve as a predictive marker for HBV-induced HCC.
A recent study indicates that Kupffer cells can coordinate with liver-trNK cells to defend against pathogen infections.529 The two kinds of cells produce more potent immune responses to virus infection than bacterium infection, which is partially attributed to the different activation of toll-like receptor (TLR)-dependent signaling pathways in Kupffer cells and their regulatory effects on trNK cell cytotoxicity. Bacterial antigens combine with TLR2 or TLR4 expressed on Kupffer cells and transduced negative signals for trNK cells in an intracellular MyD88-dependent manner. In contrast, virus infection induces trNK cell activation through the MyD88-independent way.529 The roles of Kupffer cells and trNK cells in hepatitis virus infection are worth further exploring.
In addition to the above-mentioned types of infections, some studies also reported the role of TRICs in other viral infections. For example, CD8+TRM cells located in lung tissues rapidly secrete protective molecules such as IFN-γ and IL-2 after confronting with RSV, SARS-CoV-1, and SARS-CoV-2.151,530 In mice model with virus infection, brain-CD8+TRM cells provide immune defense against the reinfection of mouse polyomavirus,531,532 and tissue-resident ILC1s can make an early robust immune response against mouse cytomegalovirus in the liver.484 Generally, most TRICs contribute to protective immunity in response to various virus infections.
Bacterial infection
The genital tract, skin, and respiratory tract where TRICs are known to exist are the main entries for the exogenous bacterium to invade. Mucosal immunity serves as a crucial defense against bacterial infection.533 Especially, the pivotal roles that TRICs play in controlling bacterial infections within the respiratory tract serve as a representative example of TRICs’ function in bacterial infections, as outlined below.
Mycobacterium tuberculosis infection
Mycobacterium tuberculosis (MTB) is the pathogen of TB.534 AMs are the predominant immune cells to encounter MTB invasion, and they can phagocyte and kill intracellular pathogens in the early infectious stage by producing inducible nitric oxide synthase.535,536,537,538 However, MTB can escape the attack from innate immunity and reside in the niches enriched AMs which serve as a reservoir for MTB growth and help them to disseminate into the lung parenchyma.537,539 AM deletion has been identified to help inhibit MTB and enhance protective immunity in the lung.540
Numerous investigations have demonstrated the significant functions of antigen-specific lung-TRM cells in controlling MTB infection.541,542 Mycobacterium bovis Bacille-Calmette-Guérin (BCG) is the only prophylactic vaccine approved for treating TB.543 The administration of BCG can induce the formation of TRM cells that strengthen local anti-bacterial responses in the lung.544,545 CD4+TRM cells and CD8+TRM cells suppress MTB by secreting IFN-γ, TNF-α, and IL-2, and CD4+TRM cells also selectively generate IL-17 to further enhance the immune control.546,547,548 However, it is not clear whether long-term resident TRM cells contribute to excessive inflammation in the lung and their roles in the progression of chronic TB.
Other TRICs in the lung also contribute to the immune response against the MTB infection. For example, γδT cells provide a protective role against MTB infection by producing IL-17, which is important for the maturation of granulomas.549,550 Moreover, IL-17 and IL-22 produced by lung-resident ILC3s initiate early innate immune responses.551 CD103+CD69+MAIT cells aggregating in the tuberculous pleural effusions highly secrete IFN-γ and GZMB for MTB clearing.552 In summary, various lung-TRICs participate in regulating local immunity upon MTB infection.
Streptococcus pneumoniae (S. pneumoniae) infection
S. pneumoniae infection will cause pneumococcal pneumonia.553 The primary immunological line of defense against bacterial infection is innate immunity mediated by AMs. AMs can eliminate ingested S. pneumoniae by promoting phagolysosomal fusion and the production of proinflammatory cytokines, nitric oxide, and reactive oxygen species.554,555,556,557 Furthermore, AMs restrain local inflammation by eradicating apoptotic polymorphonuclear cells.558 During the resolution phase of infection, AMs also promote tissue repair, and lung-resident CD69+γδT cells prevent hyperinflammation by maintaining normal numbers of AMs and DCs.559,560,561
Lymphoid TRICs also play essential roles in resisting S. pneumoniae infection. The depletion of lung-BRM cells in mice suffering from pneumococcal pneumonia will enhance bacterial activities along with less antibody production.58 Additionally, ILC3s will undergo rapid expansion and produce IL-17 and IL-22 upon S. pneumoniae infection.562,563,564 As for TRM cells, studies found that intranasal administration of pneumococci induces CD4+TRM cells in local mucosa. These cells provide long-term protection against cognate antigen challenge.565,566,567,568 CD4+TRM cells and CD8+TRM cells upregulate cytotoxic factors and promote neutrophil infiltration, which positively amplifies the immune responses against pneumococci.569,570
Staphylococcus aureus (S. aureus) infection
AMs are the primary immune cells that defend against S. aureus.554 However, the impaired chemotaxis of AMs will lead to increasing infiltration of neutrophils upon S. aureus infection, thus breaking down the lung immune homeostasis and resulting in excessive inflammation.107 As to this problem, AMs can conceal the inhaled bacteria, which is useful for preventing the hyperinflammatory response induced by recruited neutrophils. In addition, lung-CD4+TRM cells inhibit S. aureus infection by producing IL-17 and IFN-γ.571,572,573 Consequently, vaccinations that stimulate CD4+TRM cells in the lung may help avoid S. aureus invasion.
Parasite infection
Given the complex symptoms and serious injury, parasite infections pose a significant threat to human life. Few investigations are examining the association between TRICs and parasite infection. Here we review the current understanding of TRICs in anti-parasitic immunity.
Malaria
Malaria is a dangerous parasite infection disease that greatly threatens human life, caused by plasmodium infection.574 During the primary infection, plasmodium will reproduce in the host liver. Then they enter into the blood and invade red blood cells.575 Therefore, eliminating malaria parasites in the liver is essential to halt malaria infection.
Liver-CD8+TRM cells elicited by the immunization with radiation-attenuated sporozoites will possess specific transcriptional profiles.576 Pathogen-specific CD8+TRM cells mediate the malaria-killing immune responses by generating IFN-γ, TNF-α, and IL-2 to destroy parasite-infected hepatocytes.227,577 Many studies are employing vaccines to treat malaria with the primary objective of eliciting malaria-specific liver-resident CD8+TRM cells, and the treatment outcomes are favorable.578,579,580,581
Leishmaniasis
Leishmaniasis is a zoonotic disease caused by the Leishmania spp. The common clinical manifestations include cutaneous, visceral, and mucocutaneous lesions.582 There are no specific vaccines for providing long-term protection from Leishmaniasis, so it is necessary to develop novel precautionary approaches.
Skin is the natural barrier for defending against parasite invasion. A study first demonstrates the presence of Leishmania-specific CD4+TRM cells in the infected skin. At the early infection stage, skin-CD4+TRM cells make rapid responses to the Leishmania parasite. This response is dependent on the recruitment of inflammatory monocytes rather than circulating memory T cells.583,584 CD4+TRM cells also produce IFN-γ and recruit circulating T cells into the skin lesion to carry out anti-parasite immune responses.585,586 Therefore, CD4+TRM cells are the candidate for developing vaccines against Leishmania.587
In contrast to parasite-specific CD4+TRM cells, skin-resident macrophages provide niches for parasite growth.588 After being stimulated by IL-4 secreted by local eosinophils, skin-resident macrophages produce CCL24 to recruit more eosinophils, which construct a positive feedback loop to promote cutaneous Leishmania spp infection.589,590,591
In addition to TRM cells and tissue-resident macrophages, tissue-resident mast cells also participate in regulating other parasite infections. For example, mast cells repel parasites in the intestine, such as worms and Toxoplasma gondii.155,156,157,158 The tryptase mouse mast cell protease-6 secreted by intestinal CTMCs attracts eosinophils to the chronic infection sites and significantly inhibits Trichinella spiralis invasion.159 The roles of other TRICs in regulating parasite infection are less reported.
Conclusively, certain TRICs are crucial for defending against pathogen infections, including viruses, bacteria, and some kinds of parasites. They become candidate targets for the prevention or treatment of these infectious diseases. It is waiting to explore whether TRICs could sustain potent and durable anti-infection activities throughout the infection cycle, and how to prevent them from over-activating.
Therapies targeting TRICs in infectious diseases
Based on the capability of TRM cells to provide long-term immune protection and their immune memory features, many researchers have developed vaccines targeting TRM cell activation.592 Studies found that vaccines can effectively induce TRM cells to defend against pathogens in infectious diseases.234,581,593,594,595,596 For example, mucosal immunization elicit HIV-specific CD8+TRM cells in the genital and respiratory tract.593,597,598 These cells perform potent anti-virus immune responses upon the HIV infection.597 In the lung, IL-17-producing CD4+TRM cells can be induced by vaccines to provide potent immune protection against MTB infection.596 Activated TRM cells also amplify the local immunity by activating and recruiting other immune cells into the infected mucosa.593
Notably, “Prime and pull” refers to a representative immunization strategy employing TRM cells.596 Firstly, a canonical parenteral vaccination is required to induce T cell responses, resulting in the existence of effector T cells in the circulation or infected sites. Subsequently, the “pull” step is the administration of chemokines or other stimulators to recruit activated T cells into organ-specific niches and develop tissue-resident features.595 This strategy has been successfully used in controlling HSV,595,599,600 influenza virus,15, and poxvirus infection.16 Of note, the delivery route of vaccines also influences the establishment of protective immunity, including T cell-mediated immune responses and antibody production.592,596,601 In contrast to intranasal and epidermal administration, intra-muscular vaccination cannot trigger robust T cell responses, possibly due to few dendritic cells in the muscles.592,602 Thus, the vaccination strategies need to be formulated according to the biological features of invasive pathogens.
Cancer
Overview of TRICs in cancer
TME is a highly complex ecosystem where soluble mediators, ECM, and multiple cellular components assemble to establish the living environments for solid tumors. The cell components include many types of immune cells, stromal cells, and blood vessel-associated cells.603,604 As the first step of the cancer-immune cycle, tumor-derived neoantigens stimulate APCs to initiate the downstream immune responses. Different immune cells coordinate with each other to make cancer-killing or cancer-protective effects. In addition, the intercellular crosstalk not limited to immune cells also modulates cancer development, metastasis, therapy responses, and drug resistance.605,606,607 The long-term interaction between cancer cells and immune-killing effector components encourages cancer cells to acquire the ability to immune escape.603 Therefore, the persistent and sufficient cancer-killing abilities of immune cells are the decisive factor for cancer immunity. The long-term tissue residency properties render TRICs involved in the whole process of cancer immunity evolution.608
As a non-negligible constituent of TIME, many studies have elucidated the role of TRICs in cancer immunity (Fig. 6, Table 2).239,609,610,611 Their phenotypes and their potential pro- or anti-tumor effects are regulated by the intrinsic mechanisms and external TME.22

The roles of TRICs in cancer. Each TRIC shows opposite roles in tumor immunity. The red arrows indicate the pro-tumor effects, while the green arrows indicate the anti-tumor effects. VEGFA vascular endothelial growth factor-A, FGF-2 fibroblast growth factor 2. This figure is created with BioRender.com
The lymphoid TRICs are the primary players involved in anti-tumor immunity. CD8+TRM cells have been reported to enhance cancer-killing responses in BC,241,612 melanoma,239,613 NSCLC,197,240 cervical cancer,9 high-grade serous ovarian cancer (HGSC),163 HCC,614 and intrahepatic cholangiocarcinoma.614,615 The levels of CD8+TRM cells have positive clinical prognostic value for these cancer patients. Mechanistically, CD8+TRM cells release TNF-α, IFN-γ, perforin, and GZMB to perform cancer-inhibiting functions.239,613,614,615 Interestingly, these tissue-resident molecules such as CD103 extensively expressed by TRICs also regulate cancer immunity, not limited to the residency-related effects. CD103 serves as a co-stimulator for TCR-mediated cancer-killing signals.616 In addition, TGF-β, IL-33, and IFN-γ could induce the formation of CD8+TRM cells.219 TGF-β promotes CD8+TRM cells to express CD103 and enhances their adhesion to E-cadherin on epithelial cancer cells.193,617,618 Mechanically, the recruitment and phosphorylation of ILK are induced by TGF-β binding to TFGBR. ILK then interacts with the CD103 intracellular domain and active PKB/AKT to transduce transmembrane integrin signals. This process strengthens the recruitment of CD103+CD8+tumor-infiltrating lymphocytes (TILs) into epithelial tumor sites and amplifies their anti-tumor responses.619
Innate-like lymphocytes are not restricted to specific antigen-receptor recognition and activation, so they can exert rapid responses. Especially, most of them have tissue residency characteristics.120 Some studies have reported that MHC-independent γδT cells secrete high levels of cytotoxic factors such as IFN-γ, GZMB, and TNF-α in TME to motivate anti-cancer activity in triple-negative breast cancer (TNBC),620,621 NSCLC,109 colorectal cancer (CRC),622 and HCC.621 The presence of γδT cells has been identified as a favorable prognostic marker.109,620,622 Similarly, iNKT cells display robust killing effects on CRC tumor cells, and MAIT cells also play a cancer-repressive role in cholangiocarcinoma (CCA).623,624 In studies of lung cancer and ovarian cancer, trNK cells are the main anti-tumor force. They can suppress tumor growth and metastasis by secreting proinflammatory cytokines and cytotoxic molecules.625,626
Moreover, myeloid TRICs also participate in regulating tumor immunity. A study showed that tissue-resident macrophages got involved in magnifying anti-tumor immune response through interacting with CD8+T cells, and their high infiltration correlated with improved overall survival in BC patients.372
However, TRICs play dichotomous roles in different cancer types, regarded as cancer-killing effector cells in some studies, yet showing cancer-protective activity in others. For example, in cSCC and clear cell renal carcinoma, CD8+TRM cells repress cancer immunity through upregulating inhibitory receptors such as PD-1 and CTLA-4, or secreting inhibitory cytokines, and their numbers are negatively correlated to patient prognosis and clinical survival.205,627 trNK cells suppress tumor growth and metastasis in lung cancer,625 but in a study of HCC, CD49a+liver-trNK cells expressing inhibitory molecules PD-1 and LAG-3 were associated with tumor progression.112 Moreover, MAIT cells inhibit anti-cancer immunity and promote tumor progression in HCC and melanoma, correlated to the shorter overall survival and recurrence-free survival of patients.628,629
Taken together, the regulatory effects of TRICs provide new insight into cancer immunity. However, the current exploration of TRICs is mainly based on animal studies. Breaking through the technical limitations of human research and verifying the reliability of animal research conclusions in humans is critical for future TRIC investigations.
Roles of lymphoid TRICs in regulating cancer immunity
T cells are the main force of anti-tumor immunity. Circulating CD8+T cells migrate into TME and perform cancer-killing effects upon stimulation by tumor antigens presented on DCs. However, it is hard for effector T cells to maintain long-term immune protection due to functional exhaustion or cancer-immune escape.630 Many studies found CD8+TILs aggregating in the TME acquired the classic phenotypic and transcriptional markers of TRM cells, including CD103 and CD69, which were indispensable for the long-term tissue residency of CD8+T cells in tumors.631 Anusha-Preethi Ganesan et al. discovered the isolated tumor-infiltrating CTLs from lung cancer patients having transcriptional profiles of CD8+TRM cells, and CD103+CD8+TRM cells were a favorable prognostic maker of lung cancer patients.197 Moreover, CD103+CD8+TRM cells have been identified in other epithelial tumors, including high-grade ovarian cancer and CRC.632,633
Mechanistically, TRM cells eliminate cancer cells through TCR-mediated cytotoxic effects. The functional initiation of TRM cells requires the activation of co-stimulating TCR signals, including E-cadherin-CD103, extracellular signal-regulated kinase1/2-phospholipaseCγ1, phosphoinositide-3-kinase-AKT, proline-rich tyrosine kinase 2, and paxillin (Pxn).228,634,635 Activated TRM cells produce cytotoxic molecules, including GZMB, perforin, and IFN-γ, and upregulate 4-1BB, CD27, and Ki67 expression, sustaining anti-tumor activities.197,636 Many cytokines engage in regulating the immune abilities of TRM cells. IL-15 promotes the establishment of TRM cells and ILCs in BC.637,638 In lung cancer, the interaction between CD103 and E-cadherin induces CCR5 recruitment at the immunologic synapse to further retain CD103+TILs.639
Lymphoid TRICs also directly regulate tumor progression in a cytokine-dependent manner. For example, IL-22 produced by tissue-resident iNKT cells or CD4+TRM cells promotes cancer metastasis at different stages. At the early stage of cancer metastasis, IL-22 produced by tissue-resident iNKT17 cells facilitates the extravasation of cancer cells into the liver, and CD4+TRM cell-derived IL-22 promotes tumor metastasis at the late stage. IL-22 neutralization can halt this process.609,640
In contrast, many studies have demonstrated that TRM cells play opposite roles in regulating anti-tumor immunity. TRM cells can accelerate cancer progression by secreting immune-repressive molecules or aiding tumor immune escape processes. In cSCC, the CD103+CD39+CD8+TRM cells promote tumor metastasis and inhibit anti-tumor response by increasing IL-10 production and upregulating the expression of exhaustion markers like PD-1 and CTLA-4.205 A study revealed that the pre-existing TRM-like cells in lung tissues strengthened the recruitment and activation of host T cells within TME, and they also expressed high PD-1 and IFN-γ.641 In addition, the pre-existing CD8+TRM cells induced by cancer vaccines facilitate anti-tumor immunity in head and neck tumors.229
The tertiary lymphoid structures (TLSs) are the aggregation of immune cells resembling SLOs anatomically present in non-lymphoid tissues upon chronic inflammation, including autoimmune diseases, infection, and cancer.642 TLS is mainly composed of a T cell-rich zone adjacent to a B cell follicle having germinal center characteristics and is surrounded by mature DCs. The existence of TLSs in cancer loci is always positively associated with the prognosis and clinical outcomes of patients.643 Many studies suggested that TLS formation is closely related to TRM cell evolution.643,644 In GC patients, CD103+CD8+TRM cells were located around TLSs in the cancerous areas, and the high enrichment of CD103+CD8+TRM cells in TLSs was correlated with favorable patients’ responses to anti-PD-1 therapy and prognosis.645 CD103+TRM cells were also set within TLSs in lung adenocarcinoma neoplastic tissues, and their high density was positively associated with TLS maturity.646 CXCL13 produced by CD103+TRM cells promotes the establishment of TLS.647,648,649 Therefore, we speculate whether some driver factors contribute to the coordinative formation of TLS and TRM cells to maintain long-term anti-tumor function.
Unconventional T cells also play important roles in regulating tumor progression. In TME, iNKT cells recognize cancer cells or others expressing CD1d-lipid antigen and clean them by secreting cytotoxic molecules.259 They also activate other effector immune cells, such as DCs and CD8+T cells, and simultaneously repress immunosuppressive cells to amplify the immune response.110,267 In pancreatic cancer liver metastasis and chronic lymphocytic leukemia model, iNKT cells reverse immunosuppressive TME by promoting the infiltration of effector T cells or restraining TAMs.650,651 Nonetheless, iNKT cells showed cancer-protective roles in some cancers.609
γδT cells can directly kill cancer cells by antibody-dependent cell-mediated cytotoxicity or induce other immune cells to amplify anti-tumor responses cooperatively in TME.621,652 Studies also found that CD103+ or CD69+tissue-resident Vδ1+γδT cells were significantly related to the improved clinical outcome of cancer patients.109,620,622 Especially due to the non-MHC restriction and powerful cytotoxic functions of γδT cells, a substantial number of cancer therapy developments focus on allogeneic γδT cells, including endogenous γδT cell activation.653,654 Tissue-resident γδT cells are also potential therapeutic targets to offer new avenues for cancer immunotherapy.
MAIT cells exist in primary and metastatic tumors and mainly perform immunosuppressive functions in cancers, including promoting angiogenesis, suppressing T cells and/or NK cells, producing immunosuppressive cytokines, and recruiting cancer-protective immune cells into TME, consequently promoting tumor progression.624,628,655,656,657,658 For example, MAIT cells accelerate HCC progression by reducing Th1 cytokines, producing immunosuppressive cytokines, and upregulating immune checkpoint PD-1 and CTLA-4.628,629 On the contrary, MAIT cells also strengthen immune surveillance by producing cytotoxic molecules in some tumors.659
ILCs also get involved in cancer immunity, and their phenotypes will be reshaped within TME.660,661 In the MMTV-PyMT mammary tumor model, IL-15 promotes the establishment of a tissue-resident ILC1s subgroup termed type 1-like ILCs, which exhibit cytotoxic functions against cancer cells by producing GZMB, IFN-γ, and TNF-α.638 In contrast, IL-13 produced by ILC2s is correlated with poor prognosis of cancer patients.278 ILC3s reshape the TIME of intestine cancers, and the IL-22 secreted by them promotes tumor development,662 suggesting that ILC2 and ILC3 cells play immunosuppressive roles.
Notably, NK cells would transform a lot after entering solid tumor sites. They upregulate the expression of CD103, CD49a, and some inhibitory receptors, along with impaired cancer-killing effects.663 The cancer-specific phenotypes of NK cells are caused by cytokines within TME, like TGF-β, PGE-2, and IL-15.663 The roles of trNK cells in cancer immunity are seemingly dichotomous, but the specific mechanisms are unclear. In liver cancer, NK cells have been found to accelerate cancer progression by upregulating immunosuppressive molecules, including CD96, PD-1, and TIGIT, and are identified as a poor prognostic factor.112 In contrast, trNK cells have higher degranulation levels for cytotoxic molecules in ovarian cancer and NSCLC to promote anti-cancer ability.626,664 Considering the advances in NK cell therapies such as CAR-NK therapy, it is essential to further elucidate the role of trNK cells in cancer.
Roles of myeloid TRICs in regulating cancer immunity
TAMs infiltrated in the TME are one of the important components in regulating tumor immunity, and they can interact with TRM cells to regulate their maintenance.665,666 In terms of TME, a study found that in early lung cancer, M1 type-TAMs enhanced the infiltration and maintenance of CD8+TRM cells in tumor sites by producing CXCL9 and fatty acids, which CD8+TRM cells depend on for survival and functions.667 In addition, another study also indicated that IL-15 secreted by TAMs contributed to the persistent residency of TRM cells in BC.637
Other tissue-resident macrophages regulate cancer processes by specific mechanisms. Kupffer cells play important roles in preventing tumor cell engraftment and restricting metastatic tumor outgrowth in the liver.44,668,669,670 They exert anti-tumor immune responses by phagocyting tumor cells and releasing various chemokines and cytokines to recruit effector T cells and NK cells into the tumor border.44,45 Thus, Kupffer cells can serve as promising targets for treating liver metastatic malignancies. Furthermore, FOLR2+tissue-resident macrophages have been identified in BC for their promoting effects on CD8+T cell infiltration.372 In CRC, scientists identified that FCN1+monocyte-like cells enriched in tumor tissues are the precursors of many tissue-resident macrophages, which finally develop into C1QC+ and SPP1+TAMs.326 C1QC+TAMs are highly related to IL1B+tissue-resident macrophage, and they perform cellular phagocytosis and antigen presentation. SPP1+TAMs mainly connect to NLRP3+tissue-resident macrophages, which interact with tumor-associated fibroblasts to promote tumor angiogenesis and tumor metastasis.326 Moreover, the LYVE1+macrophages and NLRP3+ macrophages are found in adjacent non-cancer tissues across many cancer types and have tissue-resident characteristics.325 LYVE1+tissue-resident macrophages contribute to constraining tissue inflammation in CRC, while NLPR3+macrophages appear to have proinflammatory effects.311,326
Notably, tissue-resident macrophages are also involved in the epithelial-mesenchymal transition (EMT) of cancer cells. EMT is a prerequisite process of epithelial tumor metastasis, describing the process of epithelial cells gradually changing into mesenchymal-like cells while also losing their original characteristics.671,672,673 Tissue-resident macrophages can accelerate tumor progression by regulating the EMT, such as AMs accelerating NSCLC progression by activating the EMT process.610,674 Similarly, CD163+TIM4+ omental macrophages promote ovarian cancer metastasis by supporting the EMT and the formation of cancer stem cells.675 In pancreatic ductal adenocarcinoma, tissue-resident macrophages evolved a profibrotic transcriptional profile. Eliminating these cells reduces tumor burden.676 However, the roles of TRNs and tissue-resident mast cells in cancer processes have not been reported.
Taken together, the tissue-residence features facilitate their long-term exposure to cancer cells and functions of regulating cancer immunity, which suggests the strategies motivating their immune activities are likely to achieve long-acting therapeutic efficacy. Further explorations are warranted to elucidate the aspects of cancer immunity dominated by TRICs.
Therapies targeting TRICs in cancer
Cancer immunotherapy has become one of the treatment pillars in tumors,677 including ICB therapies, oncolytic virus therapies, immune modulators, cancer vaccines, and adoptive transfer therapy (ACT), directly or indirectly enhancing immune-derived cancer-killing capacities.197,678 However, many patients do not respond well to immunotherapies, which may be ascribed to the highly heterogeneous and complex cancer-immune interactions among individuals.679,680 Therefore, it is important to expand our understanding of TME further and discover new targets for immunotherapies. More and more studies focus on TRICs-targeted therapy development. Here, we have summarized the feasible utilization of TRICs in cancer immunotherapies (Fig. 7).

Available strategies targeting CD8+TRM cells in cancer immunotherapies. a CAR-loading renders greater cancer-killing abilities of CD8+TRM cells. b The injection of tumor antigen promotes the differentiation of memory CD8+T cells into TRM cells which exert cancer-killing functions. c The blockade of immunosuppressive CD8+TRM cells by applying specific antibodies can suppress their cancer-protective activities and restore anti-cancer immunity. d ICBs target the immune checkpoint molecules expressed on CD8+TRM cells to reactivate their cytotoxic effects. e The injection of oncolytic viruses motivates CD8+TRM cells to enhance specific anti-cancer immunity. f Cytokine therapies regulate anti-cancer-immune responses. TAA tumor-associated antigen, PD-1 programmed cell death-1, CTLA-4 cytotoxic T lymphocyte-associated antigen-4, Ab antibody, LAG-3 lymphocyte-activation gene-3, TIM-3 T cell immunoglobulin domain and mucin domain-3, ICB immune checkpoint blockade, MHC major histocompatibility complex. This figure is created with BioRender.com
Immune checkpoint blockade therapies
Immune checkpoint molecules are negative regulators of immune activation, facilitating immune homeostasis and immune escape.681 The roles of TRM cells in ICB immunotherapies have been investigated in melanoma, lung cancer, BC, and intrahepatic cholangiocarcinoma.239,240,241 Since many kinds of TRICs express these negative factors, they are potential biomarkers of patients’ response to ICB therapies. Especially, the number of CD8+TRM cells is identified as a prognostic factor for evaluating the efficacy of anti-PD-1 therapy in melanoma, TNBC, and NSCLC.11,239,240,682 Studies of BC explained that CD8+ TRM cells strengthened the local anti-tumor responses and cytotoxic capacity and restored the production of IFN-γ and TNF upon anti-CTLA-4 therapy.241 The interaction between IFN-γ-producing T cells and IL-12-producing DCs plays an important role in the effective response to anti-PD-1 immunotherapy,683 and CD49a+CXCR6+trNK cells-derived CCL5 can attract IL-12-secreted DCs and enhance intra-tumoral DC-CD8+T cells interaction.14
Apart from TRMs, other kinds of TRICs also influence the efficacy of ICB therapies. Tumor-residing MAIT cells have been found to upregulate IFN-γ production in squamous cell carcinoma following PD-1 blockade treatment, and the higher proportions of MAIT cells were positively associated with favorable responses to anti-PD-1 immunotherapy.684,685 PD-1+CD8+TRM cells and PD-1+MAIT cells interact with PD-L1+tissue-resident macrophages to repress immune activities,165 and anti-PD-1/anti-PD-L1 therapies block the interaction to reactive anti-cancer immunity.10 Interestingly, the combination of PD-1 blockade therapy with chemotherapy can lead to an increase of tissue-resident macrophages and TRMs infiltration in NSCLC, which provides novel biomarkers to predict immunotherapeutic efficacy.686,687
Cancer vaccine therapies
The basic principle of cancer vaccines is to improve the immune system’s ability to recognize and kill tumor cells by targeting tumor-associated antigens or tumor-specific antigens.688 Many studies have validated that cancer vaccine treatments can inhibit cancer progression partially by activating TRM cells. For example, intra-cervicovaginal mucosal immunization of female mouse models with human HPV vectors induces CD103+CD8+TRM cells that upregulate cytokines and perform cytotoxicity.17,689 And in the HPV E6/E7 oncoproteins-transformed TC-1 mouse model, the cervicovaginal vaccination with HPV E6/E7 recombinant vaccinia vaccine induces robust anti-tumor response harnessing CD103+CD8+TRM cells.690
Cancer vaccines also induce the generation of TRM cells in cancer.9,691 For example, after delivering a cancer vaccine by the intranasal mucosal route, the inducible mucosal-specific CD49a+CTLs suppress mucosal tumor growth.692 Additionally, the intranasal administration of the B subunit of Shiga toxin (STxB)-E7 vaccine elicits CD103+CD8+TRM cells in lung tissues, and these cells can strengthen the local anti-tumor response.691 Notably, the location of TRM cells is similarly established upon different antigen stimulations such as influenza vaccines or direct pathogen infections,234,237 but whether tumor antigens directly influence the development of TRM cells within TME is unclear.
Vaccine formulations, injection routes, and tissue-derived factors all influence the inducible efficacy of TRICs. Each tissue origin of cancers requires the appropriate vaccination methods to stimulate the production of TRM cells. For example, mucosal immunization is much better than intra-muscular or subcutaneous ways to elicit the formation of CD8+TRM cells in mucosal tumor sites.692 The superior effect of CD8+TRM cells over effector CD8+T cells in the same vaccination route may be due to the persistent mucosal location by which TRM cells provide rapid and long-term protection against tumors. In lung cancer, CD8+TRM cells induced by intranasal routes of tumor vaccines also play an indispensable and persistent role against tumor growth.691 Similarly, subcutaneous treatment of vaccines is effective to induce the formation of TRM cells in melanoma. In murine B16F10 melanoma models with the deletion of circulating CD8+T cells, intradermal vaccination motivates endogenous T cell repertoire and induces CD103+CD69+CD8+TRM cells that actively respond to tumor-specific (OVA) and self-antigens (GP100), and these cells also suppress tumor growth in vaccination sites and distant sites of skin, whereas the intraperitoneal route greatly reduces the anti-tumor efficacy.693 In the OVA-OT-1 mouse model, TILs that express VLA-1 and/or CD103 produce more IFN-γ and GZMB than integrin-negative OT-1 TILs, and the caner-repressive effects of host immunity are impaired after blocking CD103 or VLA-1.694 The preclinical orthotopic HCC mouse model shows that the anti-tumor effects of neoantigen vaccine are strengthened by combining with anti-PD-1 therapy, the synergy mainly mediated by CD8+TRM cells.695 Since the activation of TRM and BRM cells requires the stimulation of antigens,56 it is rational to prioritize the development strategies for novel cancer vaccines, with a focus on activating and inducing TRICs.
Adoptive cell therapies
ACT refers to transfusing specific and nonspecific immune cells with anti-tumor activity into patients’ circulation to kill tumor cells directly or motivate the anti-tumor immune responses of hosts. Many TRIC types are the candidate cell types in ACT.12
The technology of CAR is the most successful advance in the ACT field. General CAR-T products are made by engineering circulating T cells and play effective roles in treating hematopoietic malignancies. However, their application in solid tumors is still limited.696 The physical barriers and immunosuppressive cells and cytokines within TME greatly obstruct CAR-T cells from entering solid tumors and persistent functioning.697,698 To address this problem and improve CAR-T therapy for solid tumors, scientists fused ex vivo blood T cells with CAR engineering to generate CAR-TRM cells. Compared to CAR-T cells, the transcriptional and epigenetic profiles of CAR-TRM cells exhibit stem-like resident memory T cell characteristics. Functionally, they produce higher levels of effector cytokines and show superior resistance to exhaustion, which renders them with potent anti-tumor efficacy in both solid and hematopoietic tumor models.699 This innovative ACT strategy sheds light on the prospect of TRM cells in developing cancer immunotherapies.
Therapeutic antibodies
Targeting cancer-associated antibodies can be used to control TRIC subsistence and block their immunosuppressive effects to inhibit cancer. Here, we briefly summarize several studies that report the promising roles of therapeutic antibodies in regulating tumor immunity, including antibodies targeting receptors in tissue-resident macrophages, or basic phenotypic markers of TRICs.
In HCC, signal regulatory protein α (SIPRα) is an inhibitory receptor expressed on Kupffer cells, and its ligand CD47 is expressed on cancer cells. Inhibiting the interaction between Kupffer cells and tumor cells through blocking SIPRα-CD47 enhances phagocytosis of Kupffer cells on tumor cells.700
CSF plays an indispensable role in controlling the development of tissue-resident macrophages. CSF/CSF1R can trigger a cascade of intracellular tyrosine kinase activation, which promotes the survival of macrophages. Therefore, the inhibitory antibodies targeting CSF1R can alleviate the immunosuppressive effects of tissue-resident macrophages in some cancers.701
Many molecules participate in inducing TRM cell establishment and maintenance. For example, ICOS is a co-stimulatory receptor that plays a crucial role in inducing the formation of CD8+TRM cells, which will be inhibited by ICOS blockade.702 Moreover, the application of anti-CD45 antibodies kills tissue-resident myeloid cells in many tissues.703 Neutralized antibodies of CD103 and CD69 also effectively suppress the formation of immune-suppressive TRICs in some tumors.205,627 In short, the efficacy and kinds of therapeutic antibodies against TRICs in different diseases need further exploration.
Other therapies
Oncolytic viruses are genetically modified in vitro to inactivate their cytotoxicity but retain replication capacity. Targeted delivery of them to cancer cells can efficiently amplify systemic anti-tumor immunity by reversing immunosuppressive TME and triggering a cascade of proinflammatory events.704,705 However, the studies focusing on the application of oncolytic virotherapies functioning in a TRIC-activating/inactivating manner for cancer treatment are few. A study of pancreatic cancer showed that the administration of a variant of suratadenoturev armed with p53 (OBP-702) oncolytic virotherapy facilitated the activation and expansion of CD8+TRM cells and enhanced anti-tumor immunity.706
Some nonspecific immunotherapies also have favorable efficacy in cancer treatment such as cytokine therapy and vaccine adjuvant.707,708,709 IL-15 has been confirmed to be important for the formation of TRM cells in many tissues, and the application of IL-15 to the TME can enhance TRM cell establishment and thus promote tumor immunity.710,711 Adjuvants refer to substances that are usually added to vaccines to modulate immune responses, such as CpG oligodeoxynucleotide (ODN) is useful in enhancing the formation of skin-CD8+TRM cells and improving cancer-killing in mice B16-OVA melanoma.712,713 In conclusion, TRICs are promising targets for developing novel cancer immunotherapies.
Transplantation
Organ or tissue transplantation is an important treatment strategy for some diseases. However, the allograft-induced transplant rejection greatly threatens the clinical outcomes of patients following surgery.714 It is required to find more ways to increase the survival of patients who have received organ transplants. TRICs persistently reside in local tissues, as well as in some transplanted solid organs.715,716 It is worth exploring whether those TRICs will affect organ transplant rejection.
First, TRM cells have been identified to regulate transplant rejection immunity of different organs.715,717,718 Donor-derived CD103+CD69+CD49a+TRM cells have been found to persist in lung transplants, and their numbers are positively correlated with the clinical prognosis of recipients.719 A study of small intestine transplantation found that CD8+TRM cells in transplants could reside for more than one year, and their capacity to produce cytokines and cytotoxic molecules helped defend against intestinal infections.720
However, TRM cells are closely related to transplant rejection. Studies of facial transplant rejection found that lymphocytes aggregated around recipients’ injured cells were donor-derived and expressed CD103 and CD69. These donor cells could attack the host skin microstructures and aggravate skin allograft rejection.721 A previous study indicated that the CD103+CD8+effector T cells are essential for promoting graft damage following renal transplantation.722 In a kidney transplant mouse model with chronic rejection features, recipient CD8+T cells infiltrated in the graft did not recirculate and also acquired similar phenotypic and transcriptional characteristics to TRM cells. Moreover, the depletion of graft-CD8+TRM cells ameliorates inflammatory damage to grafts.723 The unclear issue is whether the TRM cells from the recipient or the host all contribute to transplant rejection.
Except for TRM cells, tissue-resident macrophages also regulate lung transplantation.716 Mismatched with de novo donor-specific antibodies, donor-AMs can initiate adaptive immune responses against the transplanted lung tissues by secreting proinflammatory cytokines, resulting in obstructive airway and lung fibrosis.724,725,726
Conventional anti-rejection therapies for organ transplant patients include systemic immunosuppressants.727 However, the poor therapeutic effects of these drugs may be attributed to mainly targeting circulation-effector T cells, while proinflammatory TRM cells are not mentioned. Therefore, these TRM cells may serve as therapeutic targets to prolong the survival of allografts to improve clinical outcomes in the future.
Conclusion and perspective
With the high-speed development of sequencing analysis and omics techniques, the developmental trajectories and heterogeneities of TRICs are comprehensively investigated. TRICs have been demonstrated to derive from embryonic organs or adult bone marrow, and they develop tissue-resident features at different developmental phases.2,5,11,32,56,63,64,65,728 Most TRICs can self-renew to maintain their residency in tissues, but they also depend on the supplement of blood-derived progenitors in different tissues. Tissue-resident mast cells and macrophages, trNK cells, and ILCs can derive from extramedullary hematopoiesis during adulthood, which highlights the significance of extramedullary hematopoiesis in maintaining immune cells in different tissues during adulthood, and provide us with a better understanding of the local TRIC pool composition.68,69,70,71 However, few studies are focusing on TRNs, and their tissue-resident mechanisms are still unclear.
Although most tissue-resident counterparts of circulating immune cells have been identified, there is no definition of whether DCs have tissue residency characteristics.729 Pre-DCs generate lymphoid “tissue-resident” CD8+DCs and CD11b+DCs, especially in SLOs.730 However, some studies indicated that DCs residing in peripheral tissues would migrate to lymphoid nodes under physiological and homeostatic conditions,731 so it is difficult to define the long-term “tissue residency” for DCs in different tissues.
Many factors within tissue niches participate in reshaping TRIC phenotypes and functions.732 Hence, the formation processes of TRICs are remarkably heterogeneous. Understanding the evolution of TRICs is important for people to develop and handle strategies of them. Interestingly, it has been found that the steady-state niches partially limit the plasticity of localized TRICs, except for endowing resident properties to preserve tissue homeostasis.733 Therefore, the reprogramming effects of tissue environments on TRICs are more complicated than the general imagination.
One of the major challenging tasks in TRIC studies is the identification of characteristic markers of TRICs in different tissues. Encouragingly, scRNA-seq analysis is a powerful way to find and identify the subpopulation and molecular characteristics of different TRICs. Spatial transcriptomics can help understand the compartmentalized locations of TRICs and their potential roles in controlling local immunity.
TRICs play regulator roles in different diseases. Their presence in infectious diseases is frequently correlated with favorable disease progression, suggesting their potency ability of immune defense. However, their activated immune-killing also exacerbates autoimmune diseases. Of note, the presence of most lymphoid TRICs in autoimmune diseases is induced by the previous infection. For example, the early transient viral infection results in the development of CCL5-producing TRM cells in the brain lesions of mice models with autoimmune disease. These CCL5+TRM cells preserve a long-term proinflammatory environment by recruiting circulating CCR5+autoreactive T cells and macrophages into the CNS.734 Lymphocytic choriomeningitis virus infection induces the formation of CNS-CD8+TRM cells, which can reside behind the blood-brain barrier for a long time. However, these TRM cells will attack glial cells that express the lymphocytic choriomeningitis virus glycoprotein after being infected for a while.429 Similarly, bacterium infection-induced TRM17 cells aggravate local renal inflammation by producing IL-17A in the experimental glomerulonephritis models.106 Therefore, although activated TRICs play beneficial roles in defending against infection, their long-term existence may disturb the host immune balance. This provides us with an updated understanding of TRIC functions in regulating immunity. Furthermore, monitoring the establishment and functional activities of TRICs after infections may help prevent autoimmune diseases.
Recent studies found that the infection-inducible TRICs also play a role in cancer immunity. Following influenza infection, “trained” AMs exhibit improved phagocytotic and cytotoxic functions and thus contribute to long-lasting anti-tumor immunity in a lung metastatic cancer mice model.735 This study provides novel insights into improving local cancer immunotherapy by trained immunity. In addition, some viral vaccines can induce TRM cells in tissues and thus enhance local anti-infection immunity.234,581,593,594,595,596 These vaccines may also restrain cancer development.
Many critical questions about TRICs in cancer immunity are waiting for answers: Do TRICs residing in distinct TMEs share common regulatory mechanisms of phenotypes that can be utilized for therapy development? How to activate the cancer-killing abilities of TRICs along with inactivating their immunosuppressive effects? How to evaluate the feasibility and value of TRICs as therapy and prognosis predicters? That’s not all.
Nowadays, animal models are the main methods to explore TRICs. Studies using patient samples are very few.736 Formulating research paradigms of TRICs based on high-throughput sequencing, flow cytometry, and tissue pathology technologies, or developing humanized mouse models may be feasible.
In conclusion, we comprehensively summarize the characteristics and functions of each TRIC type. We review their roles in physiological and pathological processes, especially their functions in regulating disease immunity. Therapies targeting TRIC regulation also show favorable outcomes in some disease treatments. Importantly, the knowledge of TRICs further expands people’s understanding of the immune system. We hope that this review will provide fundamental and comprehensive references for future research.
References
Vesely, M. D., Kershaw, M. H., Schreiber, R. D. & Smyth, M. J. Natural innate and adaptive immunity to cancer. Annu. Rev. Immunol. 29, 235–271, (2011).
Klose, C. S. N. & Artis, D. Innate lymphoid cells control signaling circuits to regulate tissue-specific immunity. Cell Res. 30, 475–491 (2020).
Miron, M. et al. Maintenance of the human memory T cell repertoire by subset and tissue site. Genome Med. 13, 100 (2021).
Kumar, B. V. et al. Human tissue-resident memory T cells are defined by core transcriptional and functional signatures in lymphoid and mucosal sites. Cell Rep. 20, 2921–2934 (2017).
Gray, J. I. & Farber, D. L. Tissue-resident immune cells in humans. Annu. Rev. Immunol. 40, 195–220 (2022).
Szabo, P. A., Miron, M. & Farber, D. L. Location, location, location: Tissue resident memory T cells in mice and humans. Sci. Immunol. 4, eaas9673 (2019).
Masopust, D. & Soerens, A. G. Tissue-resident T cells and other resident leukocytes. Annu. Rev. Immunol. 37, 521–546 (2019).
Ryan, G. E., Harris, J. E. & Richmond, J. M. Resident memory T cells in autoimmune skin diseases. Front. Immunol. 12, 652191 (2021).
Komdeur, F. L. et al. CD103+ tumor-infiltrating lymphocytes are tumor-reactive intraepithelial CD8+ T cells associated with prognostic benefit and therapy response in cervical cancer. Oncoimmunology 6, e1338230 (2017).
Ruf, B. et al. Tumor-associated macrophages trigger MAIT cell dysfunction at the HCC invasive margin. Cell 186, 3686–3705.e3632 (2023).
Adams, S. et al. Pembrolizumab monotherapy for previously treated metastatic triple-negative breast cancer: cohort A of the phase II KEYNOTE-086 study. Ann. Oncol. 30, 397–404 (2019).
Scheper, W., Gründer, C. & Kuball, J. Multifunctional γδ T cells and their receptors for targeted anticancer immunotherapy. Oncoimmunology 2, e23974 (2013).
Motohashi, S. et al. A phase I study of in vitro expanded natural killer T cells in patients with advanced and recurrent non-small cell lung cancer. Clin. Cancer Res. 12, 6079–6086 (2006).
Kirchhammer, N. et al. NK cells with tissue-resident traits shape response to immunotherapy by inducing adaptive antitumor immunity. Sci. Transl. Med. 14, eabm9043 (2022).
Wang, Z. et al. A novel “prime and pull” strategy mediated by the combination of two dendritic cell-targeting designs induced protective lung tissue-resident memory T cells against H1N1 influenza virus challenge. J. Nanobiotechnol. 21, 479 (2023).
Liu, L. et al. Epidermal injury and infection during poxvirus immunization is crucial for the generation of highly protective T cell-mediated immunity. Nat. Med. 16, 224–227 (2010).
Çuburu, N. et al. Adenovirus vector-based prime-boost vaccination via heterologous routes induces cervicovaginal CD8(+) T cell responses against HPV16 oncoproteins. Int. J. Cancer 142, 1467–1479 (2018).
Richmond, J. M. et al. Antibody blockade of IL-15 signaling has the potential to durably reverse vitiligo. Sci. Transl. Med. 10, (2018).
Povoleri, G. A. M. et al. Psoriatic and rheumatoid arthritis joints differ in the composition of CD8+ tissue-resident memory T cell subsets. Cell Rep. 42, 112514 (2023).
Li, Z. et al. Adult connective tissue-resident mast cells originate from late erythro-myeloid progenitors. Immunity 49, 640–653.e645 (2018).
Ginhoux, F. et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science 330, 841–845 (2010).
Sun, H., Sun, C., Xiao, W. & Sun, R. Tissue-resident lymphocytes: from adaptive to innate immunity. Cell Mol. Immunol. 16, 205–215 (2019).
Schulz, C. et al. A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science 336, 86–90 (2012).
McGrath, K. E. et al. Distinct sources of hematopoietic progenitors emerge before HSCs and provide functional blood cells in the mammalian embryo. Cell Rep. 11, 1892–1904 (2015).
Hoeffel, G. et al. C-Myb(+) erythro-myeloid progenitor-derived fetal monocytes give rise to adult tissue-resident macrophages. Immunity 42, 665–678 (2015).
Hayashi, C. et al. Mast-cell precursors in the skin of mouse embryos and their deficiency in embryos of Sl/Sld genotype. Dev. Biol. 109, 234–241 (1985).
Kitamura, Y., Shimada, M. & Go, S. Presence of mast cell precursors in fetal liver of mice. Dev. Biol. 70, 510–514, (1979).
Sonoda, T., Hayashi, C. & Kitamura, Y. Presence of mast cell precursors in the yolk sac of mice. Dev. Biol. 97, 89–94 (1983).
Yona, S. et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity 38, 79–91 (2013).
Kuziel, W. A. et al. Severe reduction in leukocyte adhesion and monocyte extravasation in mice deficient in CC chemokine receptor 2. Proc. Natl Acad. Sci. USA 94, 12053–12058 (1997).
Sheng, J., Ruedl, C. & Karjalainen, K. Most tissue-resident macrophages except microglia are derived from fetal hematopoietic stem cells. Immunity 43, 382–393, (2015).
Sawyer, R. T., Strausbauch, P. H. & Volkman, A. Resident macrophage proliferation in mice depleted of blood monocytes by strontium-89. Lab. Invest. 46, 165–170, (1982).
Kitamura, Y., Go, S. & Hatanaka, K. Decrease of mast cells in W/Wv mice and their increase by bone marrow transplantation. Blood 52, 447–452 (1978).
Kitamura, Y., Shimada, M., Hatanaka, K. & Miyano, Y. Development of mast cells from grafted bone marrow cells in irradiated mice. Nature 268, 442–443, (1977).
Kitamura, Y., Hatanaka, K., Murakami, M. & Shibata, H. Presence of mast cell precursors in peripheral blood of mice demonstrated by parabiosis. Blood 53, 1085–1088, (1979).
Matsuda, H., Kitamura, Y., Sonoda, T. & Imori, T. Precursor of mast cells fixed in the skin of mice. J. Cell Physiol. 108, 409–415, (1981).
Tauber, M. et al. Landscape of mast cell populations across organs in mice and humans. J. Exp. Med. 220, (2023).
McGovern, N. et al. Human dermal CD14+ cells are a transient population of monocyte-derived macrophages. Immunity 41, 465–477 (2014).
Scott, C. L. et al. Bone marrow-derived monocytes give rise to self-renewing and fully differentiated Kupffer cells. Nat. Commun. 7, 10321 (2016).
Dick, S. A. et al. Three tissue resident macrophage subsets coexist across organs with conserved origins and life cycles. Sci. Immunol. 7, eabf7777 (2022).
Bain, C. C. et al. Constant replenishment from circulating monocytes maintains the macrophage pool in the intestine of adult mice. Nat. Immunol. 15, 929–937 (2014).
Keith, Y. H. et al. Infiltration and local differentiation of bone marrow-derived integrinβ7-positive mast cell progenitors in atopic dermatitis-like skin. J. Allergy Clin. Immunol. 151, 159–171.e158 (2023).
van de Laar, L. et al. Yolk sac macrophages, fetal liver, and adult monocytes can colonize an empty niche and develop into functional tissue-resident macrophages. Immunity 44, 755–768 (2016).
Deng, Z. et al. The nuclear factor ID3 endows macrophages with a potent anti-tumour activity. Nature 626, 864–873 (2024).
Thomas, S. K. et al. Kupffer cells prevent pancreatic ductal adenocarcinoma metastasis to the liver in mice. Nat. Commun. 14, 6330 (2023).
Guilliams, M. & Svedberg, F. R. Does tissue imprinting restrict macrophage plasticity? Nat. Immunol. 22, 118–127 (2021).
Traina, F. et al. Single nucleotide polymorphism array lesions, TET2, DNMT3A, ASXL1 and CBL mutations are present in systemic mastocytosis. PLoS ONE 7, e43090 (2012).
Yang, I. V., Lozupone, C. A. & Schwartz, D. A. The environment, epigenome, and asthma. J. Allergy Clin. Immunol. 140, 14–23 (2017).
Ballesteros, I. et al. Co-option of neutrophil fates by tissue environments. Cell 183, 1282–1297.e1218 (2020).
Prince, L. R. et al. NR4A orphan nuclear receptor family members, NR4A2 and NR4A3, regulate neutrophil number and survival. Blood 130, 1014–1025 (2017).
Bae, G. H. et al. Unique characteristics of lung-resident neutrophils are maintained by PGE2/PKA/Tgm2-mediated signaling. Blood 140, 889–899 (2022).
Devi, S. et al. Neutrophil mobilization via plerixafor-mediated CXCR4 inhibition arises from lung demargination and blockade of neutrophil homing to the bone marrow. J. Exp. Med. 210, 2321–2336 (2013).
Ely, K. H., Cookenham, T., Roberts, A. D. & Woodland, D. L. Memory T cell populations in the lung airways are maintained by continual recruitment. J. Immunol. 176, 537–543, (2006).
Buggert, M., Price, D. A., Mackay, L. K. & Betts, M. R. Human circulating and tissue-resident memory CD8(+) T cells. Nat. Immunol. 24, 1076–1086 (2023).
Tan, H. X. et al. Lung-resident memory B cells established after pulmonary influenza infection display distinct transcriptional and phenotypic profiles. Sci. Immunol. 7, eabf5314 (2022).
Allie, S. R. et al. The establishment of resident memory B cells in the lung requires local antigen encounter. Nat. Immunol. 20, 97–108 (2019).
Mathew, N. R. et al. Single-cell BCR and transcriptome analysis after influenza infection reveals spatiotemporal dynamics of antigen-specific B cells. Cell Rep. 35, 109286 (2021).
Barker, K. A. et al. Lung-resident memory B cells protect against bacterial pneumonia. J. Clin. Invest. 131, (2021).
Pellicci, D. G., Koay, H. F. & Berzins, S. P. Thymic development of unconventional T cells: how NKT cells, MAIT cells and γδ T cells emerge. Nat. Rev. Immunol. 20, 756–770 (2020).
Treiner, E. et al. Selection of evolutionarily conserved mucosal-associated invariant T cells by MR1. Nature 422, 164–169 (2003).
Seach, N. et al. Double-positive thymocytes select mucosal-associated invariant T cells. J. Immunol. 191, 6002–6009 (2013).
Salou, M. et al. A common transcriptomic program acquired in the thymus defines tissue residency of MAIT and NKT subsets. J. Exp. Med. 216, 133–151 (2019).
Buus, T. B., Ødum, N., Geisler, C. & Lauritsen, J. P. H. Three distinct developmental pathways for adaptive and two IFN-γ-producing γδ T subsets in adult thymus. Nat. Commun. 8, 1911 (2017).
Prinz, I., Silva-Santos, B. & Pennington, D. J. Functional development of γδ T cells. Eur. J. Immunol. 43, 1988–1994, (2013).
Lombes, A. et al. Adaptive immune-like γ/δ T lymphocytes share many common features with their α/β T cell counterparts. J. Immunol. 195, 1449–1458 (2015).
Bando, J. K., Liang, H. E. & Locksley, R. M. Identification and distribution of developing innate lymphoid cells in the fetal mouse intestine. Nat. Immunol. 16, 153–160, (2015).
Liu, C. et al. Delineating spatiotemporal and hierarchical development of human fetal innate lymphoid cells. Cell Res. 31, 1106–1122 (2021).
Gasteiger, G. et al. Tissue residency of innate lymphoid cells in lymphoid and nonlymphoid organs. Science 350, 981–985 (2015).
Zeis, P. et al. In situ maturation and tissue adaptation of type 2 innate lymphoid cell progenitors. Immunity 53, 775–792.e779 (2020).
Klose, C. S. N. et al. Differentiation of type 1 ILCs from a common progenitor to all helper-like innate lymphoid cell lineages. Cell 157, 340–356 (2014).
Constantinides, M. G., McDonald, B. D., Verhoef, P. A. & Bendelac, A. A committed precursor to innate lymphoid cells. Nature 508, 397–401 (2014).
Bai, L. et al. Liver type 1 innate lymphoid cells develop locally via an interferon-γ-dependent loop. Science. 371, (2021).
Jiang, X. et al. Skin infection generates non-migratory memory CD8+ T(RM) cells providing global skin immunity. Nature 483, 227–231 (2012).
Mackay, L. K. & Kallies, A. Transcriptional regulation of tissue-resident lymphocytes. Trends Immunol. 38, 94–103 (2017).
Peng, H. et al. Liver-resident NK cells confer adaptive immunity in skin-contact inflammation. J. Clin. Invest. 123, 1444–1456 (2013).
Sojka, D. K. et al. Tissue-resident natural killer (NK) cells are cell lineages distinct from thymic and conventional splenic NK cells. Elife 3, e01659 (2014).
Tieu, R. et al. Tissue-resident memory T cell maintenance during antigen persistence requires both cognate antigen and interleukin-15. Sci. Immunol. 8, eadd8454 (2023).
Hashimoto, D. et al. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity 38, 792–804 (2013).
Gomez Perdiguero, E. et al. Tissue-resident macrophages originate from yolk-sac-derived erythro-myeloid progenitors. Nature 518, 547–551 (2015).
Anderson, K. G. et al. Intravascular staining for discrimination of vascular and tissue leukocytes. Nat. Protoc. 9, 209–222 (2014).
Teijaro, J. R. et al. Cutting edge: tissue-retentive lung memory CD4 T cells mediate optimal protection to respiratory virus infection. J. Immunol. 187, 5510–5514 (2011).
Poon, M. M. L. et al. Tissue adaptation and clonal segregation of human memory T cells in barrier sites. Nat. Immunol. 24, 309–319 (2023).
Hu, L. et al. Genetic distinction between functional tissue-resident and conventional natural killer cells. iScience 26, 107187 (2023).
Salcher, S. et al. High-resolution single-cell atlas reveals diversity and plasticity of tissue-resident neutrophils in non-small cell lung cancer. Cancer Cell 40, 1503–1520.e1508 (2022).
Peng, H. & Tian, Z. Diversity of tissue-resident NK cells. Semin. Immunol. 31, 3–10 (2017).
VanHorn, S. & Morris, S. A. Next-generation lineage tracing and fate mapping to interrogate development. Dev. Cell 56, 7–21 (2021).
Gentek, R. et al. Hemogenic endothelial fate mapping reveals dual developmental origin of mast cells. Immunity 48, 1160–1171.e1165 (2018).
Anadon, C. M. et al. Ovarian cancer immunogenicity is governed by a narrow subset of progenitor tissue-resident memory T cells. Cancer Cell 40, 545–557.e513 (2022).
Weisel, N. M. et al. Comprehensive analyses of B-cell compartments across the human body reveal novel subsets and a gut-resident memory phenotype. Blood 136, 2774–2785 (2020).
Oh, J. E. et al. Intranasal priming induces local lung-resident B cell populations that secrete protective mucosal antiviral IgA. Sci. Immunol. 6, eabj5129 (2021).
Chen, Y. et al. Single-cell transcriptomics reveals core regulatory programs that determine the heterogeneity of circulating and tissue-resident memory CD8(+) T Cells. Cells 10, (2021).
Noé, A. et al. Deep immune phenotyping and single-cell transcriptomics allow identification of circulating TRM-like cells which correlate with liver-stage immunity and vaccine-induced protection from malaria. Front. Immunol. 13, 795463 (2022).
Molodtsov, A. K. et al. Resident memory CD8(+) T cells in regional lymph nodes mediate immunity to metastatic melanoma. Immunity 54, 2117–2132.e2117 (2021).
Fang, J. et al. Decoding the transcriptional heterogeneity, differentiation lineage, clinical significance in tissue-resident memory CD8 T cell of the small intestine by single-cell analysis. J. Transl. Med. 22, 203 (2024).
Zhu, H. X. et al. Targeting pathogenic CD8(+) tissue-resident T cells with chimeric antigen receptor therapy in murine autoimmune cholangitis. Nat. Commun. 15, 2936 (2024).
Dogra, P. et al. Tissue determinants of human NK cell development, function, and residence. Cell 180, 749–763.e713 (2020).
Mauro, D. et al. CD8(+) tissue-resident memory T cells are expanded in primary Sjögren’s disease and can be therapeutically targeted by CD103 blockade. Ann. Rheum. Dis. (2024).
Bian, Z. et al. Deciphering human macrophage development at single-cell resolution. Nature 582, 571–576 (2020).
Wang, Z. et al. An immune cell atlas reveals the dynamics of human macrophage specification during prenatal development. Cell 186, 4454–4471.e4419 (2023).
Lunemann, S. et al. Hobit expression by a subset of human liver-resident CD56(bright) Natural Killer cells. Sci. Rep. 7, 6676 (2017).
Mackay, L. K. et al. Hobit and Blimp1 instruct a universal transcriptional program of tissue residency in lymphocytes. Science 352, 459–463 (2016).
Hudspeth, K. et al. Human liver-resident CD56(bright)/CD16(neg) NK cells are retained within hepatic sinusoids via the engagement of CCR5 and CXCR6 pathways. J. Autoimmun. 66, 40–50 (2016).
Cortez, V. S. et al. Cutting edge: salivary gland NK cells develop independently of Nfil3 in steady-state. J. Immunol. 192, 4487–4491 (2014).
Cortez, V. S. et al. Transforming growth factor-β signaling guides the differentiation of innate lymphoid cells in salivary glands. Immunity 44, 1127–1139 (2016).
Kang, S. et al. Tissue resident memory γδT cells in murine uterus expressed high levels of IL-17 promoting the invasion of trophocytes. Front. Immunol. 11, 588227 (2020).
Krebs, C. F. et al. Pathogen-induced tissue-resident memory T(H)17 (T(RM)17) cells amplify autoimmune kidney disease. Sci. Immunol. 5, (2020).
Neupane, A. S. et al. Patrolling alveolar macrophages conceal bacteria from the immune system to maintain homeostasis. Cell 183, 110–125.e111 (2020).
Guia, S. & Narni-Mancinelli, E. Helper-like innate lymphoid cells in humans and mice. Trends Immunol. 41, 436–452 (2020).
Wu, Y. et al. A local human Vδ1 T cell population is associated with survival in nonsmall-cell lung cancer. Nat. Cancer 3, 696–709 (2022).
Fan, X. & Rudensky, A. Y. Hallmarks of tissue-resident lymphocytes. Cell 164, 1198–1211 (2016).
Liechti, T. & Roederer, M. OMIP-058: 30-parameter flow cytometry panel to characterize iNKT, NK, unconventional and conventional T cells. Cytom. A 95, 946–951 (2019).
Sun, H. et al. Accumulation of tumor-infiltrating CD49a(+) NK cells correlates with poor prognosis for human hepatocellular carcinoma. Cancer Immunol. Res. 7, 1535–1546 (2019).
Carlson, C. M. et al. Kruppel-like factor 2 regulates thymocyte and T-cell migration. Nature 442, 299–302 (2006).
Skon, C. N. et al. Transcriptional downregulation of S1pr1 is required for the establishment of resident memory CD8+ T cells. Nat. Immunol. 14, 1285–1293 (2013).
Woon, H. G. et al. Compartmentalization of total and virus-specific tissue-resident memory CD8+ T cells in human lymphoid organs. PLoS Pathog. 12, e1005799 (2016).
Iijima, N. & Iwasaki, A. T cell memory. A local macrophage chemokine network sustains protective tissue-resident memory CD4 T cells. Science 346, 93–98, (2014).
Daussy, C. et al. T-bet and Eomes instruct the development of two distinct natural killer cell lineages in the liver and in the bone marrow. J. Exp. Med. 211, 563–577 (2014).
Kallies, A., Xin, A., Belz, G. T. & Nutt, S. L. Blimp-1 transcription factor is required for the differentiation of effector CD8(+) T cells and memory responses. Immunity 31, 283–295, (2009).
Kallies, A. et al. A role for Blimp1 in the transcriptional network controlling natural killer cell maturation. Blood 117, 1869–1879 (2011).
Xin, A. et al. A molecular threshold for effector CD8(+) T cell differentiation controlled by transcription factors Blimp-1 and T-bet. Nat. Immunol. 17, 422–432 (2016).
van Gisbergen, K. P. et al. Mouse Hobit is a homolog of the transcriptional repressor Blimp-1 that regulates NKT cell effector differentiation. Nat. Immunol. 13, 864–871 (2012).
Kragten, N. A. et al. Hobit and Blimp-1 instruct the differentiation of iNKT cells into resident-phenotype lymphocytes after lineage commitment. Eur. J. Immunol. 52, 389–403 (2022).
Laidlaw, B. J. et al. CD4+ T cell help guides formation of CD103+ lung-resident memory CD8+ T cells during influenza viral infection. Immunity 41, 633–645 (2014).
Mackay, L. K. et al. T-box transcription factors combine with the cytokines TGF-β and IL-15 to control tissue-resident memory T cell fate. Immunity 43, 1101–1111 (2015).
Zaid, A. et al. Persistence of skin-resident memory T cells within an epidermal niche. Proc. Natl Acad. Sci. USA 111, 5307–5312 (2014).
Zhang, L. H., Shin, J. H., Haggadone, M. D. & Sunwoo, J. B. The aryl hydrocarbon receptor is required for the maintenance of liver-resident natural killer cells. J. Exp. Med 213, 2249–2257 (2016).
Hombrink, P. et al. Programs for the persistence, vigilance and control of human CD8(+) lung-resident memory T cells. Nat. Immunol. 17, 1467–1478 (2016).
Hasan, M. H. & Beura, L. K. Cellular interactions in resident memory T cell establishment and function. Curr. Opin. Immunol. 74, 68–75 (2022).
Xu, W., Bergsbaken, T. & Edelblum, K. L. The multifunctional nature of CD103 (αEβ7 integrin) signaling in tissue-resident lymphocytes. Am. J. Physiol. Cell Physiol. 323, C1161–c1167 (2022).
Schenkel, J. M. et al. T cell memory. Resident memory CD8 T cells trigger protective innate and adaptive immune responses. Science 346, 98–101 (2014).
MacLean, A. J. et al. Secondary influenza challenge triggers resident memory B cell migration and rapid relocation to boost antibody secretion at infected sites. Immunity 55, 718–733.e718 (2022).
Son, Y. M. et al. Tissue-resident CD4(+) T helper cells assist the development of protective respiratory B and CD8(+) T cellmemory responses. Sci. Immunol. 6, eabb6852 (2021).
Levi-Schaffer, F. & Kupietzky, A. Mast cells enhance migration and proliferation of fibroblasts into an in vitro wound. Exp. Cell Res. 188, 42–49 (1990).
Micera, A. et al. Nerve growth factor displays stimulatory effects on human skin and lung fibroblasts, demonstrating a direct role for this factor in tissue repair. Proc. Natl Acad. Sci. USA 98, 6162–6167 (2001).
Maurer, M. et al. What is the physiological function of mast cells? Exp. Dermatol. 12, 886–910, (2003).
Dudeck, J. et al. Directional mast cell degranulation of tumor necrosis factor into blood vessels primes neutrophil extravasation. Immunity 54, 468–483.e465 (2021).
Malaviya, R. et al. Mast cells process bacterial Ags through a phagocytic route for class I MHC presentation to T cells. J. Immunol. 156, 1490–1496 (1996).
Henz, B. M. et al. Mast cells as initiators of immunity and host defense. Exp. Dermatol. 10, 1–10 (2001).
Malaviya, R., Ikeda, T., Ross, E. & Abraham, S. N. Mast cell modulation of neutrophil influx and bacterial clearance at sites of infection through TNF-alpha. Nature 381, 77–80 (1996).
Constantinides, M. G. et al. MAIT cells are imprinted by the microbiota in early life and promote tissue repair. Science 366, eaax6624 (2019).
Zhang, W. et al. Microglia-containing human brain organoids for the study of brain development and pathology. Mol. Psychiatry 28, 96–107 (2023).
Maurer, M. et al. Activated skin mast cells are involved in murine hair follicle regression (catagen). Lab Invest. 77, 319–332 (1997).
Maurer, D. & Stingl, G. Immunoglobulin E-binding structures on antigen-presenting cells present in skin and blood. J. Invest. Dermatol. 104, 707–710 (1995).
Albert-Bayo, M. et al. Intestinal mucosal mast cells: key modulators of barrier function and homeostasis. Cells. 8, 135 (2019).
Groschwitz, K. R. et al. Mast cells regulate homeostatic intestinal epithelial migration and barrier function by a chymase/Mcpt4-dependent mechanism. Proc. Natl Acad. Sci. USA 106, 22381–22386 (2009).
Fasbender, F., Widera, A., Hengstler, J. G. & Watzl, C. Natural killer cells and liver fibrosis. Front. Immunol. 7, 19 (2016).
Lam, A. J. et al. Innate control of tissue-reparative human regulatory T cells. J. Immunol. 202, 2195–2209 (2019).
Krishnan, S. et al. Amphiregulin-producing γδ T cells are vital for safeguarding oral barrier immune homeostasis. Proc. Natl Acad. Sci. USA 115, 10738–10743 (2018).
Bajpai, G. et al. The human heart contains distinct macrophage subsets with divergent origins and functions. Nat. Med 24, 1234–1245 (2018).
Srivastava, R. et al. CXCL10/CXCR3-dependent mobilization of herpes simplex virus-specific CD8(+) T(EM) and CD8(+) T(RM) cells within infected tissues allows efficient protection against recurrent herpesvirus infection and disease. J Virol. 91, e00278-17 (2017).
Piet, B. et al. CD8+ T cells with an intraepithelial phenotype upregulate cytotoxic function upon influenza infection in human lung. J. Clin. Invest. 121, 2254–2263 (2011).
Jozwik, A. et al. RSV-specific airway resident memory CD8+ T cells and differential disease severity after experimental human infection. Nat. Commun. 6, 10224 (2015).
Tessmer, M. S., Reilly, E. C. & Brossay, L. Salivary gland NK cells are phenotypically and functionally unique. PLoS Pathog. 7, e1001254 (2011).
Scharenberg, M. et al. Influenza A virus infection induces hyperresponsiveness in human lung tissue-resident and peripheral blood NK cells. Front. Immunol. 10, 1116 (2019).
Nawa, Y., Kiyota, M., Korenaga, M. & Kotani, M. Defective protective capacity of W/Wv mice against Strongyloides ratti infection and its reconstitution with bone marrow cells. Parasite Immunol. 7, 429–438, (1985).
Khan, A. I. et al. Mucosal mast cells and the expulsive mechanisms of mice against Strongyloides venezuelensis. Int J. Parasitol. 23, 551–555 (1993).
Watanabe, N., Nawa, Y., Okamoto, K. & Kobayashi, A. Expulsion of Hymenolepis nana from mice with congenital deficiencies of IgE production or of mast cell development. Parasite Immunol. 16, 137–144, (1994).
Henderson, W. R. Jr & Chi, E. Y. The importance of leukotrienes in mast cell-mediated Toxoplasma gondii cytotoxicity. J. Infect. Dis. 177, 1437–1443 (1998).
Shin, K. et al. Mouse mast cell tryptase mMCP-6 is a critical link between adaptive and innate immunity in the chronic phase of Trichinella spiralis infection. J. Immunol. 180, 4885–4891 (2008).
Oja, A. E. et al. Trigger-happy resident memory CD4(+) T cells inhabit the human lungs. Mucosal Immunol. 11, 654–667 (2018).
FitzPatrick, M. E. B. et al. Human intestinal tissue-resident memory T cells comprise transcriptionally and functionally distinct subsets. Cell Rep. 34, 108661 (2021).
Pallett, L. J. et al. IL-2(high) tissue-resident T cells in the human liver: Sentinels for hepatotropic infection. J. Exp. Med. 214, 1567–1580 (2017).
Webb, J. R., Milne, K. & Nelson, B. H. PD-1 and CD103 are widely coexpressed on prognostically favorable intraepithelial CD8 T cells in human ovarian cancer. Cancer Immunol. Res. 3, 926–935, (2015).
Park, S. L. et al. Local proliferation maintains a stable pool of tissue-resident memory T cells after antiviral recall responses. Nat. Immunol. 19, 183–191 (2018).
Weisberg, S. P. et al. Tissue-resident memory T cells mediate immune homeostasis in the human pancreas through the PD-1/PD-L1 pathway. Cell Rep. 29, 3916–3932.e3915 (2019).
Pollard, J. W. Trophic macrophages in development and disease. Nat. Rev. Immunol. 9, 259–270 (2009).
Kondo, T. et al. Heterogeneity of presenile dementia with bone cysts (Nasu-Hakola disease): three genetic forms. Neurology 59, 1105–1107 (2002).
De Schepper, S. et al. Self-maintaining gut macrophages are essential for intestinal homeostasis. Cell 175, 400–415.e413 (2018).
Mass, E. Specification of tissue-resident macrophages during organogenesis. Science. 353, aaf4238 (2016).
Furness, J. B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 9, 286–294 (2012).
Bischoff, S. C. Role of mast cells in allergic and non-allergic immune responses: comparison of human and murine data. Nat. Rev. Immunol. 7, 93–104 (2007).
Forsythe, P. Mast cells in neuroimmune interactions. Trends Neurosci. 42, 43–55 (2019).
Jacome-Galarza, C. E. et al. Developmental origin, functional maintenance and genetic rescue of osteoclasts. Nature 568, 541–545 (2019).
Veis, D. J. & O’Brien, C. A. Osteoclasts, master sculptors of bone. Annu Rev. Pathol. 18, 257–281 (2023).
Cindik, E. D. et al. Phenotypical characterization of c-kit receptor deficient mouse femora using non-destructive high-resolution imaging techniques and biomechanical testing. Technol. Health Care 8, 267–275 (2000).
Silberstein, R., Melnick, M., Greenberg, G. & Minkin, C. Bone remodeling in W/Wv mast cell deficient mice. Bone 12, 227–236 (1991).
Marquardt, N. et al. Unique transcriptional and protein-expression signature in human lung tissue-resident NK cells. Nat. Commun. 10, 3841 (2019).
Wolf, Y. et al. Brown-adipose-tissue macrophages control tissue innervation and homeostatic energy expenditure. Nat. Immunol. 18, 665–674 (2017).
Sheridan, B. S. et al. Oral infection drives a distinct population of intestinal resident memory CD8(+) T cells with enhanced protective function. Immunity 40, 747–757 (2014).
Osborn, J. F. et al. Central memory CD8+ T cells become CD69+ tissue-residents during viral skin infection independent of CD62L-mediated lymph node surveillance. PLoS Pathog. 15, e1007633 (2019).
Benichou, G. et al. Role of memory T cells in allograft rejection and tolerance. Front. Immunol. 8, 170 (2017).
Sallusto, F. et al. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 401, 708–712 (1999).
Watanabe, R. et al. Human skin is protected by four functionally and phenotypically discrete populations of resident and recirculating memory T cells. Sci. Transl. Med. 7, 279ra239 (2015).
Gerlach, C. et al. The Chemokine receptor CX3CR1 defines three antigen-experienced CD8 T cell subsets with distinct roles in immune surveillance and homeostasis. Immunity 45, 1270–1284 (2016).
Gattinoni, L. et al. A human memory T cell subset with stem cell-like properties. Nat. Med. 17, 1290–1297 (2011).
Olson, J. A., McDonald-Hyman, C., Jameson, S. C. & Hamilton, S. E. Effector-like CD8+ T cells in the memory population mediate potent protective immunity. Immunity 38, 1250–1260, (2013).
Xu, H., Zhou, R. & Chen, Z. Tissue-resident memory T cell: ontogenetic cellular mechanism and clinical translation. Clin. Exp. Immunol. 214, 249–259 (2023).
Chen, L. & Shen, Z. Tissue-resident memory T cells and their biological characteristics in the recurrence of inflammatory skin disorders. Cell Mol. Immunol. 17, 64–75 (2020).
Kok, L., Masopust, D. & Schumacher, T. N. The precursors of CD8(+) tissue resident memory T cells: from lymphoid organs to infected tissues. Nat. Rev. Immunol. 22, 283–293 (2022).
Joshi, N. S. et al. Inflammation directs memory precursor and short-lived effector CD8(+) T cell fates via the graded expression of T-bet transcription factor. Immunity 27, 281–295 (2007).
Chang, J. T., Wherry, E. J. & Goldrath, A. W. Molecular regulation of effector and memory T cell differentiation. Nat. Immunol. 15, 1104–1115, (2014).
Masopust, D. et al. Dynamic T cell migration program provides resident memory within intestinal epithelium. J. Exp. Med. 207, 553–564 (2010).
Mackay, L. K. et al. The developmental pathway for CD103(+)CD8+ tissue-resident memory T cells of skin. Nat. Immunol. 14, 1294–1301 (2013).
Mackay, L. K. et al. Long-lived epithelial immunity by tissue-resident memory T (TRM) cells in the absence of persisting local antigen presentation. Proc. Natl Acad. Sci. USA 109, 7037–7042 (2012).
Mueller, S. N., Gebhardt, T., Carbone, F. R. & Heath, W. R. Memory T cell subsets, migration patterns, and tissue residence. Annu Rev. Immunol. 31, 137–161, (2013).
Evrard, M. et al. Single-cell protein expression profiling resolves circulating and resident memory T cell diversity across tissues and infection contexts. Immunity 56, 1664–1680.e1669 (2023).
Ganesan, A. P. et al. Tissue-resident memory features are linked to the magnitude of cytotoxic T cell responses in human lung cancer. Nat. Immunol. 18, 940–950 (2017).
Cepek, K. L. et al. Adhesion between epithelial cells and T lymphocytes mediated by E-cadherin and the alpha E beta 7 integrin. Nature 372, 190–193 (1994).
Leijten, E. F. et al. Tissue-resident memory CD8+ T cells from skin differentiate psoriatic arthritis from psoriasis. Arthritis Rheumatol. 73, 1220–1232 (2021).
Bromley, S. K. et al. Recirculating memory T cells are a unique subset of CD4+ T cells with a distinct phenotype and migratory pattern. J. Immunol. 190, 970–976 (2013).
Cheon, I. S., Son, Y. M. & Sun, J. Tissue-resident memory T cells and lung immunopathology. Immunol. Rev. 316, 63–83 (2023).
Lin, Y. H. et al. Small intestine and colon tissue-resident memory CD8(+) T cells exhibit molecular heterogeneity and differential dependence on Eomes. Immunity 56, 207–223.e208 (2023).
Romagnoli, P. A. et al. Differentiation of distinct long-lived memory CD4 T cells in intestinal tissues after oral Listeria monocytogenes infection. Mucosal Immunol. 10, 520–530 (2017).
Shiow, L. R. et al. CD69 acts downstream of interferon-alpha/beta to inhibit S1P1 and lymphocyte egress from lymphoid organs. Nature 440, 540–544 (2006).
Lai, C. et al. CD8+CD103+ tissue-resident memory T cells convey reduced protective immunity in cutaneous squamous cellcarcinoma. J. Immunother. Cancer 9, e001807 (2021).
Takamura, S. et al. Specific niches for lung-resident memory CD8+ T cells at the site of tissue regeneration enable CD69-independent maintenance. J. Exp. Med. 213, 3057–3073 (2016).
Ray, S. J. et al. The collagen binding alpha1beta1 integrin VLA-1 regulates CD8 T cell-mediated immune protection against heterologous influenza infection. Immunity 20, 167–179 (2004).
Klonowski, K. D. et al. Dynamics of blood-borne CD8 memory T cell migration in vivo. Immunity 20, 551–562 (2004).
Collins, N. et al. Skin CD4(+) memory T cells exhibit combined cluster-mediated retention and equilibration with the circulation. Nat. Commun. 7, 11514 (2016).
Carbone, F. R. & Gebhardt, T. Should I stay or should I go-reconciling clashing perspectives on CD4(+) tissue-resident memory T cells. Sci. Immunol. 4, eaax5595 (2019).
Turner, D. L. et al. Lung niches for the generation and maintenance of tissue-resident memory T cells. Mucosal Immunol. 7, 501–510 (2014).
Yang, K. & Kallies, A. Tissue-specific differentiation of CD8(+) resident memory T cells. Trends Immunol. 42, 876–890 (2021).
Masopust, D. et al. Cutting edge: gut microenvironment promotes differentiation of a unique memory CD8 T cell population. J. Immunol. 176, 2079–2083 (2006).
Crowl, J. T. et al. Tissue-resident memory CD8(+) T cells possess unique transcriptional, epigenetic and functional adaptations to different tissue environments. Nat. Immunol. 23, 1121–1131 (2022).
Milner, J. J. et al. Runx3 programs CD8(+) T cell residency in non-lymphoid tissues and tumours. Nature 552, 253–257 (2017).
Behr, F. M. et al. Blimp-1 rather than Hobit drives the formation of tissue-resident memory CD8(+) T cells in the lungs. Front. Immunol. 10, 400 (2019).
Zitti, B. et al. Human skin-resident CD8(+) T cells require RUNX2 and RUNX3 for induction of cytotoxicity and expression of the integrin CD49a. Immunity 56, 1285–1302.e1287 (2023).
Li, C. et al. The transcription factor Bhlhe40 programs mitochondrial regulation of resident CD8(+) T cell fitness and functionality. Immunity 51, 491–507.e497 (2019).
Schenkel, J. M. & Pauken, K. E. Localization, tissue biology and T cell state - implications for cancer immunotherapy. Nat. Rev. Immunol. 23, 807–823 (2023).
Alfonso, C., McHeyzer-Williams, M. G. & Rosen, H. CD69 down-modulation and inhibition of thymic egress by short- and long-term selective chemical agonism of sphingosine 1-phosphate receptors. Eur. J. Immunol. 36, 149–159 (2006).
Hofmann, M. & Pircher, H. E-cadherin promotes accumulation of a unique memory CD8 T-cell population in murine salivary glands. Proc. Natl Acad. Sci. USA 108, 16741–16746, (2011).
El-Asady, R. et al. TGF-{beta}-dependent CD103 expression by CD8(+) T cells promotes selective destruction of the host intestinal epithelium during graft-versus-host disease. J. Exp. Med. 201, 1647–1657 (2005).
Casey, K. A. et al. Antigen-independent differentiation and maintenance of effector-like resident memory T cells in tissues. J. Immunol. 188, 4866–4875 (2012).
Zhang, N. & Bevan, M. J. Transforming growth factor-β signaling controls the formation and maintenance of gut-resident memory T cells by regulating migration and retention. Immunity 39, 687–696, (2013).
Mohammed, J. et al. Stromal cells control the epithelial residence of DCs and memory T cells by regulated activation of TGF-β. Nat. Immunol. 17, 414–421 (2016).
Adachi, T. et al. Hair follicle-derived IL-7 and IL-15 mediate skin-resident memory T cell homeostasis and lymphoma. Nat. Med. 21, 1272–1279 (2015).
Schenkel, J. M., Fraser, K. A., Vezys, V. & Masopust, D. Sensing and alarm function of resident memory CD8+ T cells. Nat. Immunol. 14, 509–513, (2013).
Mami-Chouaib, F. et al. Resident memory T cells, critical components in tumor immunology. J. Immunother. Cancer 6, 87 (2018).
Karaki, S. et al. CXCR6 deficiency impairs cancer vaccine efficacy and CD8(+) resident memory T-cell recruitment in headand neck and lung tumors. J. Immunother. Cancer 9, e001948 (2021).
Zaid, A. et al. Chemokine receptor-dependent control of skin tissue-resident memory T cell formation. J. Immunol. 199, 2451–2459 (2017).
Ericsson, A., Svensson, M., Arya, A. & Agace, W. W. CCL25/CCR9 promotes the induction and function of CD103 on intestinal intraepithelial lymphocytes. Eur. J. Immunol. 34, 2720–2729, (2004).
Qiu, Z. et al. Retinoic acid signaling during priming licenses intestinal CD103+ CD8 TRM cell differentiation. J. Exp. Med. 220, e20210923 (2023).
McCully, M. L. et al. CCR8 expression defines tissue-resident memory T cells in human skin. J. Immunol. 200, 1639–1650 (2018).
Zens, K. D., Chen, J. K. & Farber, D. L. Vaccine-generated lung tissue-resident memory T cells provide heterosubtypicprotection to influenza infection. JCI Insight 1, e85832 (2016).
Wakim, L. M., Woodward-Davis, A. & Bevan, M. J. Memory T cells persisting within the brain after local infection show functional adaptations to their tissue of residence. Proc. Natl Acad. Sci. USA 107, 17872–17879, (2010).
Khan, T. N. et al. Local antigen in nonlymphoid tissue promotes resident memory CD8+ T cell formation during viral infection. J. Exp. Med. 213, 951–966 (2016).
Hondowicz, B. D. et al. Interleukin-2-dependent allergen-specific tissue-resident memory cells drive asthma. Immunity 44, 155–166 (2016).
Sasson, S. C. et al. Interferon-gamma-producing CD8(+) tissue resident memory T cells are a targetable hallmark of immune checkpoint inhibitor-colitis. Gastroenterology 161, 1229–1244.e1229 (2021).
Enamorado, M. et al. Enhanced anti-tumour immunity requires the interplay between resident and circulating memory CD8(+) T cells. Nat. Commun. 8, 16073 (2017).
Djenidi, F. et al. CD8+CD103+ tumor-infiltrating lymphocytes are tumor-specific tissue-resident memory T cells and a prognostic factor for survival in lung cancer patients. J. Immunol. 194, 3475–3486 (2015).
Virassamy, B. et al. Intratumoral CD8(+) T cells with a tissue-resident memory phenotype mediate local immunity and immune checkpoint responses in breast cancer. Cancer Cell 41, 585–601.e588 (2023).
Godfrey, D. I. et al. The burgeoning family of unconventional T cells. Nat. Immunol. 16, 1114–1123 (2015).
Hildreth, A. D. & O’Sullivan, T. E. Tissue-resident innate and innate-like lymphocyte responses to viral infection. Viruses 11, 272 (2019).
Wilharm, A. et al. Mutual interplay between IL-17-producing γδT cells and microbiota orchestrates oral mucosal homeostasis. Proc. Natl Acad. Sci. USA 116, 2652–2661 (2019).
Goodman, T. & Lefrancois, L. Intraepithelial lymphocytes. Anatomical site, not T cell receptor form, dictates phenotype and function. J. Exp. Med. 170, 1569–1581 (1989).
Holtmeier, W. et al. The TCR-delta repertoire in normal human skin is restricted and distinct from the TCR-delta repertoire in the peripheral blood. J. Invest. Dermatol. 116, 275–280 (2001).
Hunter, S. et al. Human liver infiltrating γδ T cells are composed of clonally expanded circulating and tissue-resident populations. J. Hepatol. 69, 654–665 (2018).
Li, G. Q. et al. The intestinal γδ T cells: functions in the gut and in the distant organs. Front. Immunol. 14, 1206299 (2023).
Coles, M. C. & Raulet, D. H. NK1.1+ T cells in the liver arise in the thymus and are selected by interactions with class I molecules on CD4+CD8+ cells. J. Immunol. 164, 2412–2418 (2000).
Bendelac, A. Positive selection of mouse NK1+ T cells by CD1-expressing cortical thymocytes. J. Exp. Med. 182, 2091–2096 (1995).
Wei, D. G. et al. Expansion and long-range differentiation of the NKT cell lineage in mice expressing CD1d exclusively on cortical thymocytes. J. Exp. Med. 202, 239–248 (2005).
Sharma, A. et al. LFA-1 ligation by high-density ICAM-1 is sufficient to activate IFN-γ release by innate T lymphocytes. J. Immunol. 201, 2452–2461 (2018).
Lee, Y. J. et al. Tissue-specific distribution of iNKT cells impacts their cytokine response. Immunity 43, 566–578 (2015).
Krovi, S. H. & Gapin, L. Invariant natural killer T cell subsets-more than just developmental intermediates. Front. Immunol. 9, 1393 (2018).
Baranek, T., de Amat Herbozo, C., Mallevaey, T. & Paget, C. Deconstructing iNKT cell development at single-cell resolution. Trends Immunol. 43, 503–512 (2022).
Kovalovsky, D. et al. The BTB-zinc finger transcriptional regulator PLZF controls the development of invariant natural killer T cell effector functions. Nat. Immunol. 9, 1055–1064 (2008).
Constantinides, M. G. & Bendelac, A. Transcriptional regulation of the NKT cell lineage. Curr. Opin. Immunol. 25, 161–167, (2013).
Lynch, L. et al. Regulatory iNKT cells lack expression of the transcription factor PLZF and control the homeostasis of T(reg) cells and macrophages in adipose tissue. Nat. Immunol. 16, 85–95 (2015).
Wolf, B. J., Choi, J. E. & Exley, M. A. Novel approaches to exploiting invariant NKT cells in cancer immunotherapy. Front. Immunol. 9, 384 (2018).
Crosby, C. M. & Kronenberg, M. Tissue-specific functions of invariant natural killer T cells. Nat. Rev. Immunol. 18, 559–574 (2018).
Scanlon, S. T. et al. Airborne lipid antigens mobilize resident intravascular NKT cells to induce allergic airway inflammation. J. Exp. Med. 208, 2113–2124 (2011).
Geissmann, F. et al. Intravascular immune surveillance by CXCR6+ NKT cells patrolling liver sinusoids. PLoS Biol. 3, e113 (2005).
Thomas, S. Y. et al. PLZF induces an intravascular surveillance program mediated by long-lived LFA-1-ICAM-1 interactions. J. Exp. Med. 208, 1179–1188 (2011).
Berzins, S. P. et al. Long-term retention of mature NK1.1+ NKT cells in the thymus. J. Immunol. 176, 4059–4065 (2006).
Wang, W. B. et al. Developmentally programmed early-age skin localization of iNKT cells supports local tissue development and homeostasis. Nat. Immunol. 24, 225–238 (2023).
Yip, K. H., Papadopoulos, M., Pant, H. & Tumes, D. J. The role of invariant T cells in inflammation of the skin and airways. Semin. Immunopathol. 41, 401–410 (2019).
Qin, Y. et al. Invariant NKT cells facilitate cytotoxic T-cell activation via direct recognition of CD1d on T cells. Exp. Mol. Med. 51, 1–9 (2019).
Franciszkiewicz, K. et al. MHC class I-related molecule, MR1, and mucosal-associated invariant T cells. Immunol. Rev. 272, 120–138 (2016).
Dusseaux, M. et al. Human MAIT cells are xenobiotic-resistant, tissue-targeted, CD161hi IL-17-secreting T cells. Blood 117, 1250–1259 (2011).
Montaldo, E. et al. Human innate lymphoid cells. Immunol. Lett. 179, 2–8 (2016).
Serafini, N., Vosshenrich, C. A. & Di Santo, J. P. Transcriptional regulation of innate lymphoid cell fate. Nat. Rev. Immunol. 15, 415–428, (2015).
Seillet, C. et al. Deciphering the innate lymphoid cell transcriptional program. Cell Rep. 17, 436–447 (2016).
Zhou, J., Tian, Z. & Peng, H. Tissue-resident NK cells and other innate lymphoid cells. Adv. Immunol. 145, 37–53 (2020).
Simoni, Y. & Newell, E. W. Dissecting human ILC heterogeneity: more than just three subsets. Immunology 153, 297–303 (2018).
Vivier, E. et al. Innate lymphoid cells: 10 years on. Cell 174, 1054–1066 (2018).
Ishizuka, I. E., Constantinides, M. G., Gudjonson, H. & Bendelac, A. The innate lymphoid cell precursor. Annu. Rev. Immunol. 34, 299–316 (2016).
Shannon, J. P. et al. Group 1 innate lymphoid-cell-derived interferon-γ maintains anti-viral vigilance in the mucosal epithelium. Immunity 54, 276–290.e275 (2021).
Zhu, J. Mysterious ILC2 tissue adaptation. Nat. Immunol. 19, 1042–1044 (2018).
Bar-Ephraim, Y. E. et al. Cross-tissue transcriptomic analysis of human secondary lymphoid organ-residing ILC3s reveals a quiescent state in the absence of inflammation. Cell Rep. 21, 823–833 (2017).
Simoni, Y. et al. Human innate lymphoid cell subsets possess tissue-type based heterogeneity in phenotype and frequency. Immunity 46, 148–161 (2017).
Hoorweg, K. et al. Functional differences between human NKp44(-) and NKp44(+) RORC(+) innate lymphoid cells. Front. Immunol. 3, 72 (2012).
Artis, D. & Spits, H. The biology of innate lymphoid cells. Nature 517, 293–301 (2015).
Buonocore, S. et al. Innate lymphoid cells drive interleukin-23-dependent innate intestinal pathology. Nature 464, 1371–1375 (2010).
Song, C. et al. Unique and redundant functions of NKp46+ ILC3s in models of intestinal inflammation. J. Exp. Med. 212, 1869–1882 (2015).
Panda, S. K. & Colonna, M. Innate lymphoid cells in mucosal immunity. Front. Immunol. 10, 861 (2019).
Yokoyama, W. M. Natural killer cell receptors. Curr. Opin. Immunol. 7, 110–120 (1995).
Wu, S. Y., Fu, T., Jiang, Y. Z. & Shao, Z. M. Natural killer cells in cancer biology and therapy. Mol. Cancer 19, 120 (2020).
Taniguchi, H., Toyoshima, T., Fukao, K. & Nakauchi, H. Presence of hematopoietic stem cells in the adult liver. Nat. Med. 2, 198–203 (1996).
Brownlie, D. et al. Expansions of adaptive-like NK cells with a tissue-resident phenotype in human lung and blood. Proc. Natl Acad. Sci. USA 118, e2016580118 (2021).
Kalkunte, S. et al. Evolution of non-cytotoxic uterine natural killer cells. Am. J. Reprod. Immunol. 59, 425–432 (2008).
Sharkey, A. M. et al. Tissue-specific education of decidual NK cells. J. Immunol. 195, 3026–3032 (2015).
Keskin, D. B. et al. TGFbeta promotes conversion of CD16+ peripheral blood NK cells into CD16- NK cells with similarities to decidual NK cells. Proc. Natl Acad. Sci. USA 104, 3378–3383 (2007).
Peng, H. & Tian, Z. Tissue-resident natural killer cells in the livers. Sci. China Life Sci. 59, 1218–1223 (2016).
Sojka, D. K., Tian, Z. & Yokoyama, W. M. Tissue-resident natural killer cells and their potential diversity. Semin. Immunol. 26, 127–131, (2014).
Marquardt, N. et al. Human lung natural killer cells are predominantly comprised of highly differentiated hypofunctional CD69(-)CD56(dim) cells. J. Allergy Clin. Immunol. 139, 1321–1330.e1324 (2017).
Redhead, M. L. et al. The transcription factor NFIL3 is essential for normal placental and embryonic development but not for uterine natural killer (UNK) cell differentiation in mice. Biol. Reprod. 94, 101 (2016).
Tayade, C. et al. Differential transcription of Eomes and T-bet during maturation of mouse uterine natural killer cells. J. Leukoc. Biol. 78, 1347–1355 (2005).
Kurosaki, T., Kometani, K. & Ise, W. Memory B cells. Nat. Rev. Immunol. 15, 149–159, (2015).
Joo, H. M., He, Y. & Sangster, M. Y. Broad dispersion and lung localization of virus-specific memory B cells induced by influenza pneumonia. Proc. Natl Acad. Sci. USA 105, 3485–3490, (2008).
Onodera, T. et al. Memory B cells in the lung participate in protective humoral immune responses to pulmonary influenza virus reinfection. Proc. Natl Acad. Sci. USA 109, 2485–2490 (2012).
Koethe, S. et al. Pivotal advance: CD45RB glycosylation is specifically regulated during human peripheral B cell differentiation. J. Leukoc. Biol. 90, 5–19 (2011).
Savelyeva, N. & Ottensmeier, C. H. Novel players: tissue-resident memory B cells. Blood 136, 2722–2723 (2020).
Chen, C. & Laidlaw, B. J. Development and function of tissue-resident memory B cells. Adv. Immunol. 155, 1–38 (2022).
Gregoire, C. et al. Viral infection engenders bona fide and bystander subsets of lung-resident memory B cells through a permissive mechanism. Immunity 55, 1216–1233.e1219 (2022).
Reusch, L. & Angeletti, D. Memory B-cell diversity: From early generation to tissue residency and reactivation. Eur. J. Immunol. 53, e2250085 (2023).
Ural, B. B. et al. Identification of a nerve-associated, lung-resident interstitial macrophage subset with distinct localization and immunoregulatory properties. Sci. Immunol. 5, eaax8756 (2020).
Bharat, A. et al. Flow cytometry reveals similarities between lung macrophages in humans and mice. Am. J. Respir. Cell Mol. Biol. 54, 147–149 (2016).
Yu, Y. R. et al. Flow cytometric analysis of myeloid cells in human blood, bronchoalveolar lavage, and lung tissues. Am. J. Respir. Cell Mol. Biol. 54, 13–24 (2016).
Wu, X. et al. Human liver macrophage subsets defined by CD32. Front. Immunol. 11, 2108 (2020).
MacParland, S. A. et al. Single cell RNA sequencing of human liver reveals distinct intrahepatic macrophage populations. Nat. Commun. 9, 4383 (2018).
Chakarov, S. et al. Two distinct interstitial macrophage populations coexist across tissues in specific subtissular niches. Science 363, eaau0964 (2019).
Bujko, A. et al. Transcriptional and functional profiling defines human small intestinal macrophage subsets. J. Exp. Med. 215, 441–458 (2018).
Lazarov, T., Juarez-Carreño, S., Cox, N. & Geissmann, F. Physiology and diseases of tissue-resident macrophages. Nature 618, 698–707 (2023).
Davies, L. C., Jenkins, S. J., Allen, J. E. & Taylor, P. R. Tissue-resident macrophages. Nat. Immunol. 14, 986–995, (2013).
Hoeffel, G. & Ginhoux, F. Ontogeny of tissue-resident macrophages. Front. Immunol. 6, 486 (2015).
Masuda, T. et al. Specification of CNS macrophage subsets occurs postnatally in defined niches. Nature 604, 740–748 (2022).
Viola, M. F. et al. Dedicated macrophages organize and maintain the enteric nervous system. Nature 618, 818–826 (2023).
Zhao, Y. X. et al. Single-cell RNA sequencing-guided fate-mapping toolkit delineates the contribution of yolk sac erythro-myeloid progenitors. Cell Rep. 42, 113364 (2023).
Mass, E., Nimmerjahn, F., Kierdorf, K. & Schlitzer, A. Tissue-specific macrophages: how they develop and choreograph tissue biology. Nat. Rev. Immunol. 23, 563–579 (2023).
Ginhoux, F. & Guilliams, M. Tissue-resident macrophage ontogeny and homeostasis. Immunity 44, 439–449 (2016).
Epelman, S., Lavine, K. J. & Randolph, G. J. Origin and functions of tissue macrophages. Immunity 41, 21–35 (2014).
Medvinsky, A. & Dzierzak, E. Definitive hematopoiesis is autonomously initiated by the AGM region. Cell 86, 897–906 (1996).
Liu, K. et al. Lineage tracing clarifies the cellular origin of tissue-resident macrophages in the developing heart. J. Cell Biol. 221, e202108093 (2022).
Bigley, V. et al. The human syndrome of dendritic cell, monocyte, B and NK lymphoid deficiency. J. Exp. Med. 208, 227–234 (2011).
Cheng, S. et al. A pan-cancer single-cell transcriptional atlas of tumor infiltrating myeloid cells. Cell 184, 792–809.e723 (2021).
Zhang, L. et al. Single-cell analyses inform mechanisms of myeloid-targeted therapies in colon cancer. Cell 181, 442–459.e429 (2020).
Viola, M. F. & Boeckxstaens, G. Niche-specific functional heterogeneity of intestinal resident macrophages. Gut 70, 1383–1395 (2021).
Haruwaka, K. et al. Dual microglia effects on blood brain barrier permeability induced by systemic inflammation. Nat. Commun. 10, 5816 (2019).
Buttgereit, A. et al. Sall1 is a transcriptional regulator defining microglia identity and function. Nat. Immunol. 17, 1397–1406 (2016).
Butovsky, O. et al. Identification of a unique TGF-β-dependent molecular and functional signature in microglia. Nat. Neurosci. 17, 131–143 (2014).
Merad, M. et al. Langerhans cells renew in the skin throughout life under steady-state conditions. Nat. Immunol. 3, 1135–1141 (2002).
Chorro, L. et al. Langerhans cell (LC) proliferation mediates neonatal development, homeostasis, and inflammation-associated expansion of the epidermal LC network. J. Exp. Med. 206, 3089–3100 (2009).
Baker, A. D. et al. Targeted PPAR{gamma} deficiency in alveolar macrophages disrupts surfactant catabolism. J. Lipid Res. 51, 1325–1331 (2010).
Okabe, Y. & Medzhitov, R. Tissue-specific signals control reversible program of localization and functional polarization of macrophages. Cell 157, 832–844, (2014).
Petegrosso, R., Li, Z. & Kuang, R. Machine learning and statistical methods for clustering single-cell RNA-sequencing data. Brief. Bioinform. 21, 1209–1223 (2020).
Guilliams, M. et al. Spatial proteogenomics reveals distinct and evolutionarily conserved hepatic macrophage niches. Cell 185, 379–396.e338 (2022).
Park, D. S. et al. iPS-cell-derived microglia promote brain organoid maturation via cholesterol transfer. Nature 623, 397–405 (2023).
Schafer, S. T. et al. An in vivo neuroimmune organoid model to study human microglia phenotypes. Cell 186, 2111–2126.e2120 (2023).
Li, C. et al. Single-cell brain organoid screening identifies developmental defects in autism. Nature 621, 373–380 (2023).
Liew, P. X. & Kubes, P. The neutrophil’s role during health and disease. Physiol. Rev. 99, 1223–1248 (2019).
Hidalgo, A., Chilvers, E. R., Summers, C. & Koenderman, L. The neutrophil life cycle. Trends Immunol. 40, 584–597 (2019).
Silvestre-Roig, C., Hidalgo, A. & Soehnlein, O. Neutrophil heterogeneity: implications for homeostasis and pathogenesis. Blood 127, 2173–2181, (2016).
Hedrick, C. C. & Malanchi, I. Neutrophils in cancer: heterogeneous and multifaceted. Nat. Rev. Immunol. 22, 173–187 (2022).
Metcalfe, D. D., Baram, D. & Mekori, Y. A. Mast cells. Physiol. Rev. 77, 1033–1079, (1997).
Vliagoftis, H. & Befus, A. D. Rapidly changing perspectives about mast cells at mucosal surfaces. Immunol. Rev. 206, 190–203 (2005).
Abonia, J. P. et al. Alpha-4 integrins and VCAM-1, but not MAdCAM-1, are essential for recruitment of mast cell progenitors to the inflamed lung. Blood 108, 1588–1594 (2006).
Gurish, M. F. et al. Intestinal mast cell progenitors require CD49dbeta7 (alpha4beta7 integrin) for tissue-specific homing. J. Exp. Med. 194, 1243–1252 (2001).
Enerbäck, L. Mucosal mast cells in the rat and in man. Int. Arch. Allergy Appl. Immunol. 82, 249–255 (1987).
Dwyer, D. F., Barrett, N. A. & Austen, K. F. Expression profiling of constitutive mast cells reveals a unique identity within the immune system. Nat. Immunol. 17, 878–887, (2016).
Gurish, M. F. & Austen, K. F. Developmental origin and functional specialization of mast cell subsets. Immunity 37, 25–33 (2012).
Wu, T. et al. FcγR-dependent apoptosis regulates tissue persistence of mucosal and connective tissue mast cells. Eur. J. Immunol. 53, e2250221 (2023).
Friend, D. S. et al. Mast cells that reside at different locations in the jejunum of mice infected with Trichinella spiralis exhibit sequential changes in their granule ultrastructure and chymase phenotype. J. Cell Biol. 135, 279–290 (1996).
Galli, S. J. New insights into “the riddle of the mast cells”: microenvironmental regulation of mast cell development and phenotypic heterogeneity. Lab Invest. 62, 5–33 (1990).
Tsai, M. et al. The rat c-kit ligand, stem cell factor, induces the development of connective tissue-type and mucosal mast cells in vivo. Analysis by anatomical distribution, histochemistry, and protease phenotype. J. Exp. Med. 174, 125–131 (1991).
Galli, S. J. et al. Reversible expansion of primate mast cell populations in vivo by stem cell factor. J. Clin. Invest. 91, 148–152 (1993).
Valent, P. et al. Mast cells as a unique hematopoietic lineage and cell system: from Paul Ehrlich’s visions to precision medicine concepts. Theranostics 10, 10743–10768 (2020).
Cildir, G. et al. Understanding mast cell heterogeneity at single cell resolution. Trends Immunol. 42, 523–535 (2021).
Martini, E. et al. Single-cell sequencing of mouse heart immune infiltrate in pressure overload-driven heart failure reveals extent of immune activation. Circulation 140, 2089–2107 (2019).
Lehmann, G. L. et al. Single-cell profiling reveals an endothelium-mediated immunomodulatory pathway in the eye choroid. J. Exp. Med. 217, e20190730 (2020).
Huang, H., Li, Y. & Liu, B. Transcriptional regulation of mast cell and basophil lineage commitment. Semin. Immunopathol. 38, 539–548 (2016).
Putro, E. et al. New insight into intestinal mast cells revealed by single-cell RNA sequencing. Int. J. Mol. Sci. 25, 5594 (2024).
Inoue, Y. et al. Human mast cell basic fibroblast growth factor in pulmonary fibrotic disorders. Am. J. Pathol. 149, 2037–2054 (1996).
Mukai, K., Tsai, M., Saito, H. & Galli, S. J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 282, 121–150 (2018).
Sobiepanek, A. et al. The multifaceted roles of mast cells in immune homeostasis, infections and cancers. Int. J. Mol. Sci. 23, 2249 (2022).
Norrby, K. Mast cells and angiogenesis. Apmis 110, 355–371 (2002).
Galli, S. J. & Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 18, 693–704 (2012).
Nadler, M. J., Matthews, S. A., Turner, H. & Kinet, J. P. Signal transduction by the high-affinity immunoglobulin E receptor Fc epsilon RI: coupling form to function. Adv. Immunol. 76, 325–355, (2000).
Theoharides, T. C., Valent, P. & Akin, C. Mast cells, mastocytosis, and related disorders. N. Engl. J. Med. 373, 163–172, (2015).
Sasson, S. C. et al. Local heroes or villains: tissue-resident memory T cells in human health and disease. Cell Mol. Immunol. 17, 113–122 (2020).
Asada, N. et al. Tissue-resident memory T cells in the kidney. Semin. Immunopathol. 44, 801–811 (2022).
Li, Y., You, Z., Tang, R. & Ma, X. Tissue-resident memory T cells in chronic liver diseases: Phenotype, development and function. Front. Immunol. 13, 967055 (2022).
Nalio Ramos, R. et al. Tissue-resident FOLR2(+) macrophages associate with CD8(+) T cell infiltration in human breast cancer. Cell 185, 1189–1207.e1125 (2022).
Kveštak, D., Mihalić, A., Jonjić, S. & Brizić, I. Innate lymphoid cells in neuroinflammation. Front. Cell Neurosci. 18, 1364485 (2024).
Lavin, Y., Mortha, A., Rahman, A. & Merad, M. Regulation of macrophage development and function in peripheral tissues. Nat. Rev. Immunol. 15, 731–744, (2015).
Uderhardt, S. et al. Resident macrophages cloak tissue microlesions to prevent neutrophil-driven inflammatory damage. Cell 177, 541–555.e517 (2019).
Shook, B. A. et al. Myofibroblast proliferation and heterogeneity are supported by macrophages during skin repair. Science 362, eaar2971 (2018).
Mirza, R., DiPietro, L. A. & Koh, T. J. Selective and specific macrophage ablation is detrimental to wound healing in mice. Am. J. Pathol. 175, 2454–2462, (2009).
You, Q., Cheng, L., Kedl, R. M. & Ju, C. Mechanism of T cell tolerance induction by murine hepatic Kupffer cells. Hepatology 48, 978–990, (2008).
Heymann, F. et al. Liver inflammation abrogates immunological tolerance induced by Kupffer cells. Hepatology 62, 279–291 (2015).
Yu, J. et al. Liver metastasis restrains immunotherapy efficacy via macrophage-mediated T cell elimination. Nat. Med. 27, 152–164 (2021).
Bernard, J. K., Marakovits, C., Smith, L. G. & Francis, H. Mast cell and innate immune cell communication in cholestatic liver disease. Semin Liver Dis. 43, 226–233 (2023).
Nicolás-Ávila, J., Adrover, J. M. & Hidalgo, A. Neutrophils in homeostasis, immunity, and cancer. Immunity 46, 15–28 (2017).
Casanova-Acebes, M. et al. Neutrophils instruct homeostatic and pathological states in naive tissues. J. Exp. Med. 215, 2778–2795 (2018).
Rana, B. M. J. et al. A stromal cell niche sustains ILC2-mediated type-2 conditioning in adipose tissue. J. Exp. Med. 216, 1999–2009 (2019).
Mahlakõiv, T. et al. Stromal cells maintain immune cell homeostasis in adipose tissue via production of interleukin-33. Sci. Immunol 4, eaax0416 (2019).
Vargas-Uricoechea, H. Molecular mechanisms in autoimmune thyroid disease. Cells 12, 918 (2023).
Yasunaga, M. Antibody therapeutics and immunoregulation in cancer and autoimmune disease. Semin. Cancer Biol. 64, 1–12 (2020).
Wu, H. et al. Pathogenic role of tissue-resident memory T cells in autoimmune diseases. Autoimmun. Rev. 17, 906–911 (2018).
Schulz-Kuhnt, A., Wirtz, S., Neurath, M. F. & Atreya, I. Regulation of human innate lymphoid cells in the context of mucosal inflammation. Front. Immunol. 11, 1062 (2020).
Smolen, J. S., Aletaha, D. & McInnes, I. B. Rheumatoid arthritis. Lancet 388, 2023–2038 (2016).
Huang, J. et al. Promising therapeutic targets for treatment of rheumatoid arthritis. Front. Immunol. 12, 686155 (2021).
McInnes, I. B. & Schett, G. The pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 365, 2205–2219, (2011).
Chang, M. H. et al. Arthritis flares mediated by tissue-resident memory T cells in the joint. Cell Rep. 37, 109902 (2021).
Jung, J. et al. Synovial fluid CD69(+)CD8(+) T cells with tissue-resident phenotype mediate perforin-dependent citrullination in rheumatoid arthritis. Clin. Transl. Immunol. 9, e1140 (2020).
Gao, A. et al. Tissue-resident memory T cells: the key frontier in local synovitis memory of rheumatoid arthritis. J. Autoimmun. 133, 102950 (2022).
Onuora, S. Erasing the memory of arthritis in joints. Nat. Rev. Rheumatol. 18, 5 (2022).
Scherer, H. U., Häupl, T. & Burmester, G. R. The etiology of rheumatoid arthritis. J. Autoimmun. 110, 102400 (2020).
Fert-Bober, J., Darrah, E. & Andrade, F. Insights into the study and origin of the citrullinome in rheumatoid arthritis. Immunol. Rev. 294, 133–147 (2020).
Corsiero, E. et al. NETosis as source of autoantigens in rheumatoid arthritis. Front. Immunol. 7, 485 (2016).
Menon, B. et al. Interleukin-17+CD8+ T cells are enriched in the joints of patients with psoriatic arthritis and correlate with disease activity and joint damage progression. Arthritis Rheumatol. 66, 1272–1281 (2014).
Steel, K. J. A. et al. Polyfunctional, proinflammatory, tissue-resident memory phenotype and function of synovial interleukin-17A+CD8+ T Cells in psoriatic arthritis. Arthritis Rheumatol. 72, 435–447 (2020).
Haringman, J. J. et al. Synovial tissue macrophages: a sensitive biomarker for response to treatment in patients with rheumatoid arthritis. Ann. Rheum. Dis. 64, 834–838 (2005).
Udalova, I. A., Mantovani, A. & Feldmann, M. Macrophage heterogeneity in the context of rheumatoid arthritis. Nat. Rev. Rheumatol. 12, 472–485, (2016).
Mulherin, D., Fitzgerald, O. & Bresnihan, B. Synovial tissue macrophage populations and articular damage in rheumatoid arthritis. Arthritis Rheum. 39, 115–124, (1996).
Buckley, C. D. Macrophages form a protective cellular barrier in joints. Nature 572, 590–592 (2019).
Collison, J. Synovial macrophages shield the joints. Nat. Rev. Rheumatol. 15, 573 (2019).
Culemann, S. et al. Locally renewing resident synovial macrophages provide a protective barrier for the joint. Nature 572, 670–675 (2019).
Kemble, S. & Croft, A. P. Critical role of synovial tissue-resident macrophage and fibroblast subsets in the persistence of joint inflammation. Front. Immunol. 12, 715894 (2021).
Huang, Q. Q. et al. Critical role of synovial tissue-resident macrophage niche in joint homeostasis and suppression ofchronic inflammation. Sci. Adv. 7, eabd0515 (2021).
Sontheimer, R. D. Skin manifestations of systemic autoimmune connective tissue disease: diagnostics and therapeutics. Best. Pract. Res. Clin. Rheumatol. 18, 429–462, (2004).
Yamanaka, K., Yamamoto, O. & Honda, T. Pathophysiology of psoriasis: a review. J. Dermatol. 48, 722–731 (2021).
Suárez-Fariñas, M., Fuentes-Duculan, J., Lowes, M. A. & Krueger, J. G. Resolved psoriasis lesions retain expression of a subset of disease-related genes. J. Invest. Dermatol. 131, 391–400 (2011).
Mak, R. K., Hundhausen, C. & Nestle, F. O. Progress in understanding the immunopathogenesis of psoriasis. Actas Dermosifiliogr. 100 (Suppl. 2), 2–13 (2009).
Boyman, O. et al. Spontaneous development of psoriasis in a new animal model shows an essential role for resident T cells and tumor necrosis factor-alpha. J. Exp. Med. 199, 731–736 (2004).
Cheuk, S. et al. CD49a expression defines tissue-resident CD8(+) T cells poised for cytotoxic function in human skin. Immunity 46, 287–300 (2017).
Kurihara, K. et al. Significance of IL-17A-producing CD8(+)CD103(+) skin resident memory T cells in psoriasis lesion and their possible relationship to clinical course. J. Dermatol Sci. 95, 21–27 (2019).
Owczarczyk Saczonek, A., Krajewska-Włodarczyk, M., Kasprowicz-Furmańczyk, M. & Placek, W. Immunological memory of psoriatic lesions. Int. J. Mol. Sci. 21, 625 (2020).
Tohyama, M. et al. Bcl-3 induced by IL-22 via STAT3 activation acts as a potentiator of psoriasis-related gene expression in epidermal keratinocytes. Eur. J. Immunol. 48, 168–179 (2018).
Frisoli, M. L., Essien, K. & Harris, J. E. Vitiligo: mechanisms of pathogenesis and treatment. Annu. Rev. Immunol. 38, 621–648 (2020).
Boniface, K. et al. Vitiligo skin is imprinted with resident memory CD8 T cells expressing CXCR3. J. Invest. Dermatol. 138, 355–364 (2018).
Harris, J. E. et al. Rapid skin repigmentation on oral ruxolitinib in a patient with coexistent vitiligo and alopecia areata (AA). J. Am. Acad. Dermatol. 74, 370–371 (2016).
Richmond, J. M. et al. Resident memory and recirculating memory T cells cooperate to maintain disease in a mouse model of vitiligo. J. Invest. Dermatol. 139, 769–778 (2019).
Ostkamp, P. et al. A single-cell analysis framework allows for characterization of CSF leukocytes and their tissue of origin in multiple sclerosis. Sci. Transl. Med. 14, eadc9778 (2022).
Beltrán, E. et al. Early adaptive immune activation detected in monozygotic twins with prodromal multiple sclerosis. J. Clin. Invest. 129, 4758–4768 (2019).
Tian, J. et al. Tissue-resident immune cells in the pathogenesis of multiple sclerosis. Inflamm. Res. 72, 363–372 (2023).
Machado-Santos, J. et al. The compartmentalized inflammatory response in the multiple sclerosis brain is composed of tissue-resident CD8+ T lymphocytes and B cells. Brain 141, 2066–2082 (2018).
Graham, J. B., Da Costa, A. & Lund, J. M. Regulatory T cells shape the resident memory T cell response to virus infection in the tissues. J. Immunol. 192, 683–690, (2014).
Prasad, S. et al. Reactive glia promote development of CD103(+) CD69(+) CD8(+) T-cells through programmed cell death-ligand 1 (PD-L1). Immun. Inflamm. Dis. 6, 332–344 (2018).
Vincenti, I. et al. Tissue-resident memory CD8(+) T cells cooperate with CD4(+) T cells to drive compartmentalized immunopathology in the CNS. Sci. Transl. Med. 14, eabl6058 (2022).
Frieser, D. et al. Tissue-resident CD8(+) T cells drive compartmentalized and chronic autoimmune damage against CNS neurons. Sci. Transl. Med. 14, eabl6157 (2022).
Fransen, N. L. et al. Tissue-resident memory T cells invade the brain parenchyma in multiple sclerosis white matter lesions. Brain 143, 1714–1730 (2020).
Kwong, B. et al. T-bet-dependent NKp46(+) innate lymphoid cells regulate the onset of T(H)17-induced neuroinflammation. Nat. Immunol. 18, 1117–1127 (2017).
Grigg, J. B. et al. Antigen-presenting innate lymphoid cells orchestrate neuroinflammation. Nature 600, 707–712 (2021).
Willing, A. et al. Production of IL-17 by MAIT cells is increased in multiple sclerosis and is associated with IL-7 receptor expression. J. Immunol. 200, 974–982 (2018).
Abrahamsson, S. V. et al. Non-myeloablative autologous haematopoietic stem cell transplantation expands regulatory cells and depletes IL-17 producing mucosal-associated invariant T cells in multiple sclerosis. Brain 136, 2888–2903 (2013).
Yamasaki, R. et al. Differential roles of microglia and monocytes in the inflamed central nervous system. J. Exp. Med. 211, 1533–1549 (2014).
Miron, V. E. & Franklin, R. J. Macrophages and CNS remyelination. J. Neurochem 130, 165–171, (2014).
González, H., Elgueta, D., Montoya, A. & Pacheco, R. Neuroimmune regulation of microglial activity involved in neuroinflammation and neurodegenerative diseases. J. Neuroimmunol. 274, 1–13 (2014).
Ponomarev, E. D., Maresz, K., Tan, Y. & Dittel, B. N. CNS-derived interleukin-4 is essential for the regulation of autoimmune inflammation and induces a state of alternative activation in microglial cells. J. Neurosci. 27, 10714–10721, (2007).
Huang, Y. et al. Glutaminase dysregulation in HIV-1-infected human microglia mediates neurotoxicity: relevant to HIV-1-associated neurocognitive disorders. J. Neurosci. 31, 15195–15204 (2011).
Nikić, I. et al. A reversible form of axon damage in experimental autoimmune encephalomyelitis and multiple sclerosis. Nat. Med. 17, 495–499 (2011).
Chabot, S., Williams, G. & Yong, V. W. Microglial production of TNF-alpha is induced by activated T lymphocytes. Involvement of VLA-4 and inhibition by interferonbeta-1b. J. Clin. Invest. 100, 604–612, (1997).
Ben-Sasson, S. Z. et al. IL-1 acts directly on CD4 T cells to enhance their antigen-driven expansion and differentiation. Proc. Natl Acad. Sci. USA 106, 7119–7124 (2009).
Gao, Z. & Tsirka, S. E. Animal models of MS reveal multiple roles of microglia in disease pathogenesis. Neurol. Res. Int. 2011, 383087 (2011).
Heppner, F. L. et al. Experimental autoimmune encephalomyelitis repressed by microglial paralysis. Nat. Med. 11, 146–152 (2005).
Bhasin, M., Wu, M. & Tsirka, S. E. Modulation of microglial/macrophage activation by macrophage inhibitory factor (TKP) or tuftsin (TKPR) attenuates the disease course of experimental autoimmune encephalomyelitis. BMC Immunol. 8, 10 (2007).
Windhagen, A. et al. Expression of costimulatory molecules B7-1 (CD80), B7-2 (CD86), and interleukin 12 cytokine in multiple sclerosis lesions. J. Exp. Med. 182, 1985–1996 (1995).
Dong, Y. & Yong, V. W. When encephalitogenic T cells collaborate with microglia in multiple sclerosis. Nat. Rev. Neurol. 15, 704–717 (2019).
Zundler, S. et al. Hobit- and Blimp-1-driven CD4(+) tissue-resident memory T cells control chronic intestinal inflammation. Nat. Immunol. 20, 288–300 (2019).
Serriari, N. E. et al. Innate mucosal-associated invariant T (MAIT) cells are activated in inflammatory bowel diseases. Clin. Exp. Immunol. 176, 266–274 (2014).
You, Z. et al. The clinical significance of hepatic CD69(+) CD103(+) CD8(+) resident-memory T cells in autoimmune hepatitis. Hepatology 74, 847–863 (2021).
Böttcher, K. et al. MAIT cells are chronically activated in patients with autoimmune liver disease and promote profibrogenic hepatic stellate cell activation. Hepatology 68, 172–186 (2018).
Parikh, S. V., Almaani, S., Brodsky, S. & Rovin, B. H. Update on lupus nephritis: core curriculum 2020. Am. J. Kidney Dis. 76, 265–281 (2020).
Ginsberg, P., Panzer, U. & Asada, N. Tissue-resident memory T cells in renal autoimmune diseases. Front. Immunol. 14, 1111521 (2023).
Zhou, M. et al. JAK/STAT signaling controls the fate of CD8(+)CD103(+) tissue-resident memory T cell in lupus nephritis. J. Autoimmun. 109, 102424 (2020).
Richoz, N. et al. Distinct pathogenic roles for resident and monocyte-derived macrophages in lupus nephritis. JCI Insight 7, e159751 (2022).
Ushio, A. et al. CCL22-producing resident macrophages enhance T cell response in Sjögren’s syndrome. Front. Immunol. 9, 2594 (2018).
Gao, C. Y. et al. Tissue-resident memory CD8+ T cells acting as mediators of salivary gland damage in a murine model of Sjögren’s syndrome. Arthritis Rheumatol. 71, 121–132 (2019).
Sato, M. et al. Disturbed natural killer cell homeostasis in the salivary gland enhances autoimmune pathology via IFN-γ in a mouse model of primary Sjögren’s syndrome. Front. Med. 9, 1036787 (2022).
Miyabe, C. et al. Dectin-2-induced CCL2 production in tissue-resident macrophages ignites cardiac arteritis. J. Clin. Invest. 129, 3610–3624 (2019).
Kuric, E. et al. Demonstration of tissue resident memory CD8 T cells in insulitic lesions in adult patients with recent-onset type 1 diabetes. Am. J. Pathol. 187, 581–588 (2017).
Kang, S., Wu, Q., Yang, B. & Wu, C. Estrogen enhanced the expression of IL-17 by tissue-resident memory γδT cells from uterus via interferon regulatory factor 4. FASEB J. 36, e22166 (2022).
Cuthbert, R. J. et al. Evidence that tissue resident human enthesis γδT-cells can produce IL-17A independently of IL-23R transcript expression. Ann. Rheum. Dis. 78, 1559–1565 (2019).
Pan, Y. et al. Survival of tissue-resident memory T cells requires exogenous lipid uptake and metabolism. Nature 543, 252–256 (2017).
Purnama, C. et al. Transient ablation of alveolar macrophages leads to massive pathology of influenza infection without affecting cellular adaptive immunity. Eur. J. Immunol. 44, 2003–2012 (2014).
Kumagai, Y. et al. Alveolar macrophages are the primary interferon-alpha producer in pulmonary infection with RNA viruses. Immunity 27, 240–252 (2007).
Pervizaj-Oruqaj, L. et al. Alveolar macrophage-expressed Plet1 is a driver of lung epithelial repair after viral pneumonia. Nat. Commun. 15, 87 (2024).
Yao, Y. et al. Induction of autonomous memory alveolar macrophages requires T cell help and is critical to trained immunity. Cell 175, 1634–1650.e1617 (2018).
Didierlaurent, A. et al. Sustained desensitization to bacterial Toll-like receptor ligands after resolution of respiratory influenza infection. J. Exp. Med. 205, 323–329 (2008).
Verma, A. K. et al. Influenza infection induces alveolar macrophage dysfunction and thereby enables noninvasive streptococcus pneumoniae to cause deadly pneumonia. J. Immunol. 205, 1601–1607 (2020).
Aegerter, H. et al. Influenza-induced monocyte-derived alveolar macrophages confer prolonged antibacterial protection. Nat. Immunol. 21, 145–157 (2020).
Lee, S., Yeung, K. K. & Watts, T. H. Tissue-resident memory T cells in protective immunity to influenza virus. Curr. Opin. Virol. 65, 101397 (2024).
de Bree, G. J. et al. Characterization of CD4+ memory T cell responses directed against common respiratory pathogens in peripheral blood and lung. J. Infect. Dis. 195, 1718–1725 (2007).
de Bree, G. J. et al. Selective accumulation of differentiated CD8+ T cells specific for respiratory viruses in the human lung. J. Exp. Med. 202, 1433–1442 (2005).
Pizzolla, A. et al. Influenza-specific lung-resident memory T cells are proliferative and polyfunctional and maintain diverse TCR profiles. J. Clin. Invest. 128, 721–733 (2018).
Koutsakos, M. et al. Human CD8(+) T cell cross-reactivity across influenza A, B and C viruses. Nat. Immunol. 20, 613–625 (2019).
Zheng, M. Z. M. et al. Single-cycle influenza virus vaccine generates lung CD8(+) Trm that cross-react against viral variants and subvert virus escape mutants. Sci. Adv. 9, eadg3469 (2023).
Paik, D. H. & Farber, D. L. Influenza infection fortifies local lymph nodes to promote lung-resident heterosubtypicimmunity. J. Exp. Med. 218, e20200218 (2021).
Goplen, N. P. et al. Tissue-resident CD8(+) T cells drive age-associated chronic lung sequelae after viral pneumonia. Sci. Immunol 5, eabc4557 (2020).
Quinn, K. M. et al. Heightened self-reactivity associated with selective survival, but not expansion, of naïve virus-specific CD8+ T cells in aged mice. Proc. Natl Acad. Sci. USA 113, 1333–1338 (2016).
Quinn, K. M. et al. Age-related decline in primary CD8(+) T cell responses is associated with the development of senescence in virtual memory CD8(+) T cells. Cell Rep. 23, 3512–3524 (2018).
Kawasaki, T. et al. Alveolar macrophages instruct CD8(+) T cell expansion by antigen cross-presentation in lung. Cell Rep. 41, 111828 (2022).
Monticelli, L. A. et al. Innate lymphoid cells promote lung-tissue homeostasis after infection with influenza virus. Nat. Immunol. 12, 1045–1054 (2011).
Weizman, O. E. et al. ILC1 confer early host protection at initial sites of viral infection. Cell 171, 795–808.e712 (2017).
Gorski, S. A., Hahn, Y. S. & Braciale, T. J. Group 2 innate lymphoid cell production of IL-5 is regulated by NKT cells during influenza virus infection. PLoS Pathog. 9, e1003615 (2013).
Li, B. W. S. et al. T cells and ILC2s are major effector cells in influenza-induced exacerbation of allergic airway inflammation in mice. Eur. J. Immunol. 49, 144–156 (2019).
Califano, D. et al. IFN-γ increases susceptibility to influenza A infection through suppression of group II innate lymphoid cells. Mucosal Immunol. 11, 209–219 (2018).
Cooper, G. E. et al. Human CD49a(+) lung natural killer cell cytotoxicity in response to influenza A virus. Front. Immunol. 9, 1671 (2018).
Qin, G. et al. Type 1 responses of human Vγ9Vδ2 T cells to influenza A viruses. J. Virol. 85, 10109–10116 (2011).
Fatahzadeh, M. & Schwartz, R. A. Human herpes simplex virus infections: epidemiology, pathogenesis, symptomatology, diagnosis, and management. J. Am. Acad. Dermatol. 57, 737–763 (2007). quiz 764-736.
Zhu, S. & Viejo-Borbolla, A. Pathogenesis and virulence of herpes simplex virus. Virulence 12, 2670–2702 (2021).
Filgueira, L., Larionov, A. & Lannes, N. The influence of virus infection on microglia and accelerated brain aging. Cells 10, 1836 (2021).
Fekete, R. et al. Microglia control the spread of neurotropic virus infection via P2Y12 signalling and recruit monocytes through P2Y12-independent mechanisms. Acta Neuropathol. 136, 461–482 (2018).
Katzilieris-Petras, G. et al. Microglia activate early antiviral responses upon herpes simplex virus 1 entry into the brain to counteract development of encephalitis-like disease in mice. J. Virol. 96, e0131121 (2022).
Uyar, O. et al. An early microglial response is needed to efficiently control herpes simplex virus encephalitis. J Virol 94, e01428-20 (2020).
Song, X. et al. Nicotinamide n-Oxide Attenuates HSV-1-Induced Microglial Inflammation through Sirtuin-1/NF-κB signaling. Int. J. Mol. Sci. 23, 16085 (2022).
Marcocci, M. E. et al. Herpes simplex virus-1 in the brain: the dark side of a sneaky infection. Trends Microbiol. 28, 808–820 (2020).
McMillan, R. E., Wang, E., Carlin, A. F. & Coufal, N. G. Human microglial models to study host-virus interactions. Exp. Neurol. 363, 114375 (2023).
Wang, Y. et al. Roles of HSV-1 infection-induced microglial immune responses in CNS diseases: friends or foes? Crit. Rev. Microbiol. 45, 581–594 (2019).
Roychoudhury, P. et al. Tissue-resident T cell-derived cytokines eliminate herpes simplex virus-2-infected cells. J. Clin. Invest. 130, 2903–2919 (2020).
Unger, P. A. et al. T-cells in human trigeminal ganglia express canonical tissue-resident memory T-cell markers. J. Neuroinflamm. 19, 249 (2022).
Posavad, C. M. et al. Enrichment of herpes simplex virus type 2 (HSV-2) reactive mucosal T cells in the human female genital tract. Mucosal Immunol. 10, 1259–1269 (2017).
Feinberg, J. & Keeshin, S. Prevention and Initial Management of HIV Infection. Ann. Intern. Med. 175, Itc81–itc96 (2022).
Shacklett, B. L. & Ferre, A. L. Mucosal immunity in HIV controllers: the right place at the right time. Curr. Opin. HIV AIDS 6, 202–207 (2011).
Shacklett, B. L. et al. Trafficking of human immunodeficiency virus type 1-specific CD8+ T cells to gut-associated lymphoid tissue during chronic infection. J. Virol. 77, 5621–5631 (2003).
Ibarrondo, F. J. et al. Parallel human immunodeficiency virus type 1-specific CD8+ T-lymphocyte responses in blood and mucosa during chronic infection. J. Virol. 79, 4289–4297 (2005).
Gibbs, A. et al. Human immunodeficiency virus-infected women have high numbers of CD103-CD8+ T cells residing close to the basal membrane of the ectocervical epithelium. J. Infect. Dis. 218, 453–465 (2018).
Moylan, D. C. et al. Diminished CD103 (αEβ7) expression on resident T cells from the female genital tract of HIV-positive women. Pathog. Immun. 1, 371–387, (2016).
Buggert, M. et al. Identification and characterization of HIV-specific resident memory CD8(+) T cells in human lymphoidtissue. Sci. Immunol. 3, eaar4526 (2018).
Kiniry, B. E. et al. Detection of HIV-1-specific gastrointestinal tissue resident CD8(+) T-cells in chronic infection. Mucosal Immunol. 11, 909–920 (2018).
Kiniry, B. E. et al. Predominance of weakly cytotoxic, T-bet(Low)Eomes(Neg) CD8(+) T-cells in human gastrointestinal mucosa: implications for HIV infection. Mucosal Immunol. 10, 1008–1020 (2017).
Roan, N. R. Resident T cells stand up to HIV. Sci. Immunol. 3, eaat6121 (2018).
Mvaya, L. et al. Differential localization and limited cytotoxic potential of duodenal CD8+ T cells. JCI Insight 7, e154195 (2022).
Rosato, P. C. et al. Tissue-resident memory T cells trigger rapid exudation and local antibody accumulation. Mucosal Immunol. 16, 17–26 (2023).
Chitrakar, A., Sanz, M., Maggirwar, S. B. & Soriano-Sarabia, N. HIV latency in myeloid cells: challenges for a cure. Pathogens 11, 611 (2022).
Hendricks, C. M., Cordeiro, T., Gomes, A. P. & Stevenson, M. The interplay of HIV-1 and macrophages in viral persistence. Front. Microbiol. 12, 646447 (2021).
Real, F. et al. S100A8-mediated metabolic adaptation controls HIV-1 persistence in macrophages in vivo. Nat. Commun. 13, 5956 (2022).
O'Neil, T. R. et al. The role of tissue resident memory CD4 T cells in herpes simplex viral and HIV infection. Viruses 13, 359 (2021).
Wong, M. E., Jaworowski, A. & Hearps, A. C. The HIV reservoir in monocytes and macrophages. Front. Immunol. 10, 1435 (2019).
Couturier, J. & Lewis, D. E. HIV persistence in adipose tissue reservoirs. Curr. HIV/AIDS Rep. 15, 60–71 (2018).
Damouche, A. et al. High proportion of PD-1-expressing CD4(+) T cells in adipose tissue constitutes an immunomodulatory microenvironment that may support HIV persistence. Eur. J. Immunol. 47, 2113–2123 (2017).
Odenwald, M. A. & Paul, S. Viral hepatitis: past, present, and future. World J. Gastroenterol. 28, 1405–1429 (2022).
Lanini, S. et al. Viral hepatitis: etiology, epidemiology, transmission, diagnostics, treatment, and prevention. Infect. Dis. Clin. North Am. 33, 1045–1062 (2019).
Stelma, F. et al. Human intrahepatic CD69 + CD8+ T cells have a tissue resident memory T cell phenotype with reduced cytolytic capacity. Sci. Rep. 7, 6172 (2017).
Sung, C. C. et al. Asialo GM1-positive liver-resident CD8 T cells that express CD44 and LFA-1 are essential for immune clearance of hepatitis B virus. Cell Mol. Immunol. 18, 1772–1782 (2021).
Wang, Y. et al. Murine CXCR3(+)CXCR6(+)γδT cells reside in the liver and provide protection against HBV infection. Front. Immunol. 12, 757379 (2021).
Cheng, Y. et al. Non-terminally exhausted tumor-resident memory HBV-specific T cell responses correlate with relapse-free survival in hepatocellular carcinoma. Immunity 54, 1825–1840.e1827 (2021).
Wei, J. & Ishizuka, J. J. Going viral: HBV-specific CD8(+) tissue-resident memory T cells propagate anti-tumor immunity. Immunity 54, 1630–1632 (2021).
Tu, Z. et al. TLR-dependent cross talk between human Kupffer cells and NK cells. J. Exp. Med. 205, 233–244 (2008).
Zhang, M. et al. Pulmonary resident memory T cells in respiratory virus infection and their inspiration on therapeutic strategies. Front. Immunol. 13, 943331 (2022).
Shwetank et al. PD-1 dynamically regulates inflammation and development of brain-resident memory CD8 T cells during persistent viral encephalitis. Front. Immunol. 10, 783 (2019).
Mockus, T. E. et al. CD4 T cells control development and maintenance of brain-resident CD8 T cells during polyomavirus infection. PLoS Pathog. 14, e1007365 (2018).
Cuthbertson, L. et al. Genomic attributes of airway commensal bacteria and mucosa. Commun. Biol. 7, 171 (2024).
Koch, A. & Mizrahi, V. Mycobacterium tuberculosis. Trends Microbiol. 26, 555–556 (2018).
Bo, H. et al. Mycobacterium tuberculosis-macrophage interaction: molecular updates. Front. Cell Infect. Microbiol. 13, 1062963 (2023).
Thuong, N. T. et al. MARCO variants are associated with phagocytosis, pulmonary tuberculosis susceptibility and Beijing lineage. Genes Immun. 17, 419–425 (2016).
Ahmad, F. et al. Macrophage: a cell with many faces and functions in tuberculosis. Front. Immunol. 13, 747799 (2022).
Nicholson, S. et al. Inducible nitric oxide synthase in pulmonary alveolar macrophages from patients with tuberculosis. J. Exp. Med. 183, 2293–2302 (1996).
Cohen, S. B. et al. Alveolar macrophages provide an early mycobacterium tuberculosis niche and initiate dissemination. Cell Host Microbe 24, 439–446.e434 (2018).
Leemans, J. C. et al. Depletion of alveolar macrophages exerts protective effects in pulmonary tuberculosis in mice. J. Immunol. 166, 4604–4611 (2001).
Ogongo, P., Porterfield, J. Z. & Leslie, A. Lung tissue resident memory T-cells in the immune response to Mycobacterium tuberculosis. Front. Immunol. 10, 992 (2019).
Yu, S., Lao, S., Yang, B. & Wu, C. Tissue-resident memory-like CD8(+) T cells exhibit heterogeneous characteristics in tuberculous pleural effusion. J. Immunol. Res. 2021, 6643808 (2021).
Mangtani, P. et al. Protection by BCG vaccine against tuberculosis: a systematic review of randomized controlled trials. Clin. Infect. Dis. 58, 470–480 (2014).
Bull, N. C. et al. Enhanced protection conferred by mucosal BCG vaccination associates with presence of antigen-specific lung tissue-resident PD-1(+) KLRG1(-) CD4(+) T cells. Mucosal Immunol. 12, 555–564 (2019).
Hu, Z. et al. Sendai virus mucosal vaccination establishes lung-resident memory CD8 T cell immunity and boosts BCG-primed protection against TB in mice. Mol. Ther. 25, 1222–1233 (2017).
Yang, Q. et al. Cutting edge: characterization of human tissue-resident memory T cells at different infection sites in patients with tuberculosis. J. Immunol. 204, 2331–2336 (2020).
Ogongo, P. et al. Tissue-resident-like CD4+ T cells secreting IL-17 control Mycobacterium tuberculosis in the human lung. J. Clin. Invest. 131, e142014 (2021).
Pahuja, I. et al. Berberine governs NOTCH3/AKT signaling to enrich lung-resident memory T cells during tuberculosis. PLoS Pathog. 19, e1011165 (2023).
Lockhart, E., Green, A. M. & Flynn, J. L. IL-17 production is dominated by gammadelta T cells rather than CD4 T cells during Mycobacterium tuberculosis infection. J. Immunol. 177, 4662–4669, (2006).
Okamoto Yoshida, Y. et al. Essential role of IL-17A in the formation of a mycobacterial infection-induced granuloma in the lung. J. Immunol. 184, 4414–4422 (2010).
Ardain, A. et al. Group 3 innate lymphoid cells mediate early protective immunity against tuberculosis. Nature 570, 528–532 (2019).
Jiang, J. et al. Tim-3 expression is induced by mycobacterial antigens and identifies tissue-resident subsets of MAIT cells from patients with tuberculosis. Microbes Infect. 25, 105021 (2023).
Weiser, J. N., Ferreira, D. M. & Paton, J. C. Streptococcus pneumoniae: transmission, colonization and invasion. Nat. Rev. Microbiol. 16, 355–367 (2018).
Cole, J., Aberdein, J., Jubrail, J. & Dockrell, D. H. The role of macrophages in the innate immune response to Streptococcus pneumoniae and Staphylococcus aureus: mechanisms and contrasts. Adv. Micro. Physiol. 65, 125–202 (2014).
Marriott, H. M., Hellewell, P. G., Whyte, M. K. & Dockrell, D. H. Contrasting roles for reactive oxygen species and nitric oxide in the innate response to pulmonary infection with Streptococcus pneumoniae. Vaccine 25, 2485–2490, (2007).
Machado, M. G. et al. Acetate improves the killing of Streptococcus pneumoniae by alveolar macrophages via NLRP3 inflammasome and glycolysis-HIF-1α axis. Front. Immunol. 13, 773261 (2022).
Aberdein, J. D. et al. Alveolar macrophages in pulmonary host defence the unrecognized role of apoptosis as a mechanism of intracellular bacterial killing. Clin. Exp. Immunol. 174, 193–202 (2013).
Knapp, S. et al. Alveolar macrophages have a protective antiinflammatory role during murine pneumococcal pneumonia. Am. J. Respir. Crit. Care Med. 167, 171–179 (2003).
Aggarwal, N. R., King, L. S. & D’Alessio, F. R. Diverse macrophage populations mediate acute lung inflammation and resolution. Am. J. Physiol. Lung Cell Mol. Physiol. 306, L709–L725 (2014).
Kirby, A. C., Newton, D. J., Carding, S. R. & Kaye, P. M. Pulmonary dendritic cells and alveolar macrophages are regulated by gammadelta T cells during the resolution of S. pneumoniae-induced inflammation. J. Pathol. 212, 29–37 (2007).
Kirby, A. C., Newton, D. J., Carding, S. R. & Kaye, P. M. Evidence for the involvement of lung-specific gammadelta T cell subsets in local responses to Streptococcus pneumoniae infection. Eur. J. Immunol. 37, 3404–3413, (2007).
Van Maele, L. et al. Activation of type 3 innate lymphoid cells and interleukin 22 secretion in the lungs during Streptococcus pneumoniae infection. J. Infect. Dis. 210, 493–503 (2014).
Ardain, A., Porterfield, J. Z., Kløverpris, H. N. & Leslie, A. Type 3 ILCs in lung disease. Front. Immunol. 10, 92 (2019).
Hoffmann, J. P., Kolls, J. K. & McCombs, J. E. Regulation and function of ILC3s in pulmonary infections. Front. Immunol. 12, 672523 (2021).
O’Hara, J. M. et al. Generation of protective pneumococcal-specific nasal resident memory CD4(+) T cells via parenteral immunization. Mucosal Immunol. 13, 172–182 (2020).
Smith, N. M. et al. Regionally compartmentalized resident memory T cells mediate naturally acquired protection against pneumococcal pneumonia. Mucosal Immunol. 11, 220–235 (2018).
Lyon De Ana, C. et al. GL7 ligand expression defines a novel subset of CD4(+) T(RM) cells in lungs recovered from pneumococcus. Mucosal Immunol. 16, 699–710 (2023).
Lindenberg, M. et al. Clarithromycin impairs tissue-resident memory and Th17 responses to macrolide-resistant Streptococcus pneumoniae infections. J. Mol. Med. 99, 817–829 (2021).
Shenoy, A. T. et al. Lung CD4(+) resident memory T cells remodel epithelial responses to accelerate neutrophil recruitment during pneumonia. Mucosal Immunol. 13, 334–343 (2020).
Ge, C. et al. Bystander activation of pulmonary Trm cells attenuates the severity of bacterial pneumonia by enhancing neutrophil recruitment. Cell Rep. 29, 4236–4244.e4233 (2019).
Hendriks, A. et al. Staphylococcus aureus-specific tissue-resident memory CD4(+) T cells are abundant in healthy human skin. Front. Immunol. 12, 642711 (2021).
Brown, A. F. et al. Memory Th1 cells are protective in invasive Staphylococcus aureus Infection. PLoS Pathog. 11, e1005226 (2015).
Braverman, J. et al. Staphylococcus aureus specific lung resident memory CD4(+) Th1 cells attenuate the severity of influenza virus induced secondary bacterial pneumonia. Mucosal Immunol. 15, 783–796 (2022).
Milner, D. A., Jr. Malaria pathogenesis. Cold Spring Harb. Perspect. Med. 8, a025569 (2018).
Basu, S. & Sahi, P. K. Malaria: an update. Indian J. Pediatr. 84, 521–528 (2017).
Tse, S. W. et al. Unique transcriptional profile of liver-resident memory CD8+ T cells induced by immunization with malaria sporozoites. Genes Immun. 14, 302–309 (2013).
Fernandez-Ruiz, D. et al. Liver-resident memory CD8(+) T cells form a front-line defense against malaria liver-stage infection. Immunity 45, 889–902 (2016).
Walk, J., Stok, J. E. & Sauerwein, R. W. Can patrolling liver-resident T cells control human malaria parasite development? Trends Immunol. 40, 186–196 (2019).
Olsen, T. M., Stone, B. C., Chuenchob, V. & Murphy, S. C. Prime-and-trap malaria vaccination to generate protective CD8(+) liver-resident memory t cells. J. Immunol. 201, 1984–1993 (2018).
Valencia-Hernandez, A. M. et al. A natural peptide antigen within the plasmodium ribosomal protein RPL6 confers liver T(RM) cell-mediated immunity against malaria in mice. Cell Host Microbe 27, 950–962.e957 (2020).
Ganley, M. et al. mRNA vaccine against malaria tailored for liver-resident memory T cells. Nat. Immunol. 24, 1487–1498 (2023).
Pace, D. Leishmaniasis. J. Infect. 69 S10–S18 (2014).
Glennie, N. D., Volk, S. W. & Scott, P. Skin-resident CD4+ T cells protect against Leishmania major by recruiting and activating inflammatory monocytes. PLoS Pathog. 13, e1006349 (2017).
Nateghi-Rostami, M. & Sohrabi, Y. Memory T cells: promising biomarkers for evaluating protection and vaccine efficacy against leishmaniasis. Front. Immunol. 15, 1304696 (2024).
Glennie, N. D. et al. Skin-resident memory CD4+ T cells enhance protection against Leishmania major infection. J. Exp. Med. 212, 1405–1414 (2015).
Scott, P. Long-lived skin-resident memory T cells contribute to concomitant immunity in cutaneous leishmaniasis. Cold Spring Harb. Perspect. Biol. 12, a038059 (2020).
Louis, L. et al. Intradermal synthetic DNA vaccination generates Leishmania-specific T cells in the skin and protectionagainst Leishmania major. Infect. Immun. 87, e00227-19 (2019).
Lee, S. H. et al. Mannose receptor high, M2 dermal macrophages mediate nonhealing Leishmania major infection in a Th1 immune environment. J. Exp. Med. 215, 357–375 (2018).
Lee, S. H. et al. M2-like, dermal macrophages are maintained via IL-4/CCL24-mediated cooperative interaction with eosinophils in cutaneous leishmaniasis. Sci. Immunol. 5, eaaz4415 (2020).
Lee, S. H. et al. Dermis resident macrophages orchestrate localized ILC2 eosinophil circuitries to promote non-healing cutaneous leishmaniasis. Nat. Commun. 14, 7852 (2023).
Vellozo, N. S., Rigoni, T. S. & Lopes, M. F. New therapeutic tools to shape monocyte functional phenotypes in leishmaniasis. Front. Immunol. 12, 704429 (2021).
Rotrosen, E. & Kupper, T. S. Assessing the generation of tissue resident memory T cells by vaccines. Nat. Rev. Immunol. 23, 655–665 (2023).
Tan, H. X. et al. Induction of vaginal-resident HIV-specific CD8 T cells with mucosal prime-boost immunization. Mucosal Immunol. 11, 994–1007 (2018).
Mekonnen, Z. A. et al. Toward DNA-based T-cell mediated vaccines to target HIV-1 and hepatitis C virus: approaches to elicit localized immunity for protection. Front. Cell Infect. Microbiol. 9, 91 (2019).
Shin, H. & Iwasaki, A. A vaccine strategy that protects against genital herpes by establishing local memory T cells. Nature 491, 463–467, (2012).
Counoupas, C. et al. Mucosal delivery of a multistage subunit vaccine promotes development of lung-resident memory T cells and affords interleukin-17-dependent protection against pulmonary tuberculosis. NPJ Vaccines 5, 105 (2020).
Zaric, M. et al. Long-lived tissue resident HIV-1 specific memory CD8(+) T cells are generated by skin immunization with live virus vectored microneedle arrays. J. Control Release 268, 166–175 (2017).
Adnan, S. et al. Persistent low-level replication of SIVΔnef drives maturation of antibody and CD8 T cell responses to induce protective immunity against vaginal SIV infection. PLoS Pathog. 12, e1006104 (2016).
Bernstein, D. I. et al. Successful application of prime and pull strategy for a therapeutic HSV vaccine. NPJ Vaccines 4, 33 (2019).
Gopinath, S., Lu, P. & Iwasaki, A. Cutting edge: the use of topical aminoglycosides as an effective pull in “prime and pull” vaccine strategy. J. Immunol. 204, 1703–1707 (2020).
Künzli, M. et al. Route of self-amplifying mRNA vaccination modulates the establishment of pulmonary resident memory CD8 and CD4 T cells. Sci. Immunol. 7, eadd3075 (2022).
Pan, Y. et al. Epicutaneous immunization with modified vaccinia Ankara viral vectors generates superior T cell immunity against a respiratory viral challenge. NPJ Vaccines 6, 1 (2021).
de Visser, K. E. & Joyce, J. A. The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth. Cancer Cell 41, 374–403 (2023).
Elhanani, O., Ben-Uri, R. & Keren, L. Spatial profiling technologies illuminate the tumor microenvironment. Cancer Cell 41, 404–420 (2023).
Hinshaw, D. C. & Shevde, L. A. The tumor microenvironment innately modulates cancer progression. Cancer Res. 79, 4557–4566 (2019).
Xiao, Y. & Yu, D. Tumor microenvironment as a therapeutic target in cancer. Pharm. Ther. 221, 107753 (2021).
Pitt, J. M. et al. Targeting the tumor microenvironment: removing obstruction to anticancer immune responses and immunotherapy. Ann. Oncol. 27, 1482–1492 (2016).
Nakamura, K. & Smyth, M. J. Myeloid immunosuppression and immune checkpoints in the tumor microenvironment. Cell Mol. Immunol. 17, 1–12 (2020).
Giannou, A. D. et al. Tissue resident iNKT17 cells facilitate cancer cell extravasation in liver metastasis via interleukin-22. Immunity 56, 125–142.e112 (2023).
Casanova-Acebes, M. et al. Tissue-resident macrophages provide a pro-tumorigenic niche to early NSCLC cells. Nature 595, 578–584 (2021).
Sivori, S. et al. NK cells and ILCs in tumor immunotherapy. Mol. Asp. Med 80, 100870 (2021).
Simoni, Y. et al. Bystander CD8(+) T cells are abundant and phenotypically distinct in human tumour infiltrates. Nature 557, 575–579 (2018).
Park, S. L. et al. Tissue-resident memory CD8(+) T cells promote melanoma-immune equilibrium in skin. Nature 565, 366–371 (2019).
Chen, L. et al. Prognostic values of tissue-resident CD8(+)T cells in human hepatocellular carcinoma and intrahepatic cholangiocarcinoma. World J. Surg. Oncol. 21, 124 (2023).
Kim, H. D. et al. Implication of CD69(+) CD103(+) tissue-resident-like CD8(+) T cells as a potential immunotherapeutic target for cholangiocarcinoma. Liver Int. 41, 764–776 (2021).
Le Floc’h, A. et al. Minimal engagement of CD103 on cytotoxic T lymphocytes with an E-cadherin-Fc molecule triggers lytic granule polarization via a phospholipase Cgamma-dependent pathway. Cancer Res. 71, 328–338 (2011).
Qiu, Z., Chu, T. H. & Sheridan, B. S. TGF-β: many paths to CD103(.) CD8 T cell residency. Cells 10, 989 (2021).
Hasan, F. et al. Hypoxia acts as an environmental cue for the human tissue-resident memory T cell differentiation program. JCI Insight 6, e138970 (2021).
Boutet, M. et al. TGFβ signaling intersects with CD103 integrin signaling to promote T-lymphocyte accumulation and antitumor activity in the lung tumor microenvironment. Cancer Res. 76, 1757–1769 (2016).
Wu, Y. et al. An innate-like Vδ1(+) γδ T cell compartment in the human breast is associated with remission in triplenegativebreast cancer. Sci. Transl. Med. 11, eaax9364 (2019).
Zakeri, N. et al. Characterisation and induction of tissue-resident gamma delta T-cells to target hepatocellular carcinoma. Nat. Commun. 13, 1372 (2022).
Bruni, E. et al. Intrahepatic CD69(+)Vδ1 T cells re-circulate in the blood of patients with metastatic colorectal cancer andlimit tumor progression. J. Immunother. Cancer 10, e004579 (2022).
Burks, J., Olkhanud, P. B. & Berzofsky, J. A. The role of NKT cells in gastrointestinal cancers. Oncoimmunology 11, 2009666 (2022).
Zimmer, C. L. et al. Mucosal-associated invariant T-cell tumor infiltration predicts long-term survival in cholangiocarcinoma. Hepatology 75, 1154–1168 (2022).
Yamamoto, Y. et al. Lung-resident natural killer cells control pulmonary tumor growth in mice. Cancer Sci. 109, 2670–2676 (2018).
Bernson, E. et al. Identification of tissue-resident natural killer and T lymphocytes with anti-tumor properties in ascites of ovarian cancer patients. Cancers 15, 3362 (2023).
Sanders, C. et al. CD103+ tissue resident T-lymphocytes accumulate in lung metastases and are correlated with poor prognosis in ccRCC. Cancers 14, 1541 (2022).
Fu, S. et al. Regulatory mucosa-associated invariant T cells controlled by β1 adrenergic receptor signaling contribute to hepatocellular carcinoma progression. Hepatology 78, 72–87 (2023).
Duan, M. et al. Activated and exhausted MAIT cells foster disease progression and indicate poor outcome in hepatocellular carcinoma. Clin. Cancer Res. 25, 3304–3316 (2019).
Chen, D. S. & Mellman, I. Elements of cancer immunity and the cancer-immune set point. Nature 541, 321–330 (2017).
Damei, I., Trickovic, T., Mami-Chouaib, F. & Corgnac, S. Tumor-resident memory T cells as a biomarker of the response to cancer immunotherapy. Front. Immunol. 14, 1205984 (2023).
Webb, J. R. et al. Tumor-infiltrating lymphocytes expressing the tissue resident memory marker CD103 are associated with increased survival in high-grade serous ovarian cancer. Clin. Cancer Res. 20, 434–444 (2014).
Duhen, T. et al. Co-expression of CD39 and CD103 identifies tumor-reactive CD8 T cells in human solid tumors. Nat. Commun. 9, 2724 (2018).
Schaller, M. D. Paxillin: a focal adhesion-associated adaptor protein. Oncogene 20, 6459–6472, (2001).
Feng, S. et al. Regulatory role of RGMb in lung injury promoted by the combination of carbon ion irradiation and anti-PD-1 antibody through Erk1/2 and p38 MAPK pathways. Biochem. Biophys. Res. Commun. 691, 149334 (2024).
Best, J. A. et al. Transcriptional insights into the CD8(+) T cell response to infection and memory T cell formation. Nat. Immunol. 14, 404–412 (2013).
Boldajipour, B., Nelson, A. & Krummel, M. F. Tumor-infiltrating lymphocytes are dynamically desensitized to antigen but are maintained by homeostatic cytokine. JCI Insight 1, e89289 (2016).
Dadi, S. et al. Cancer immunosurveillance by tissue-resident innate lymphoid cells and innate-like T cells. Cell 164, 365–377 (2016).
Franciszkiewicz, K. et al. Intratumoral induction of CD103 triggers tumor-specific CTL function and CCR5-dependent T-cell retention. Cancer Res. 69, 6249–6255 (2009).
Zhang, T. et al. CD4+ T cell-derived IL-22 enhances liver metastasis by promoting angiogenesis. Oncoimmunology 12, 2269634 (2023).
Weeden, C. E. et al. Early immune pressure initiated by tissue-resident memory T cells sculpts tumor evolution in non-small cell lung cancer. Cancer Cell 41, 837–852.e836 (2023).
Cabrita, R. et al. Tertiary lymphoid structures improve immunotherapy and survival in melanoma. Nature 577, 561–565 (2020).
Sautès-Fridman, C., Petitprez, F., Calderaro, J. & Fridman, W. H. Tertiary lymphoid structures in the era of cancer immunotherapy. Nat. Rev. Cancer 19, 307–325 (2019).
Aloisi, F. & Pujol-Borrell, R. Lymphoid neogenesis in chronic inflammatory diseases. Nat. Rev. Immunol. 6, 205–217, (2006).
Mori, T. et al. Tertiary lymphoid structures show infiltration of effective tumor-resident T cells in gastric cancer. Cancer Sci. 112, 1746–1757 (2021).
Zhao, H. et al. Tumor-resident T cells, associated with tertiary lymphoid structure maturity, improve survival in patients with stage III lung adenocarcinoma. Front. Immunol. 13, 877689 (2022).
Denton, A. E. et al. Type I interferon induces CXCL13 to support ectopic germinal center formation. J. Exp. Med. 216, 621–637 (2019).
Luther, S. A. et al. Differing activities of homeostatic chemokines CCL19, CCL21, and CXCL12 in lymphocyte and dendritic cell recruitment and lymphoid neogenesis. J. Immunol. 169, 424–433 (2002).
Ng, K. W. et al. Antibodies against endogenous retroviruses promote lung cancer immunotherapy. Nature 616, 563–573 (2023).
Yi, Q. et al. scRNA-Seq and imaging mass cytometry analyses unveil iNKT cells-mediated anti-tumor immunity in pancreatic cancer liver metastasis. Cancer Lett. 561, 216149 (2023).
Gorini, F. et al. Invariant NKT cells contribute to chronic lymphocytic leukemia surveillance and prognosis. Blood 129, 3440–3451 (2017).
Silva-Santos, B., Mensurado, S. & Coffelt, S. B. T cells: pleiotropic immune effectors with therapeutic potential in cancer. Nat. Rev. Cancer 19, 392–404 (2019).
Khairallah, C., Chu, T. H. & Sheridan, B. S. Tissue adaptations of memory and tissue-resident gamma delta T cells. Front. Immunol. 9, 2636 (2018).
Legut, M., Cole, D. K. & Sewell, A. K. The promise of γδ T cells and the γδ T cell receptor for cancer immunotherapy. Cell Mol. Immunol. 12, 656–668, (2015).
Yan, J. et al. MAIT cells promote tumor initiation, growth, and metastases via tumor MR1. Cancer Discov. 10, 124–141 (2020).
Cogswell, D. T. et al. MAIT cells: partners or enemies in cancer immunotherapy?. Cancers 13, 1502 (2021).
O'Neill, C., Cassidy, F. C., O'Shea, D. & Hogan, A. E. Mucosal associated invariant T cells in cancer-friend or foe?. Cancers 13, 1582 (2021).
Ouyang, L. et al. Mucosal-associated invariant T cells reduce and display tissue-resident phenotype with elevated IL-17 producing capacity in non-small cell lung cancer. Int. Immunopharmacol. 113, 109461 (2022).
Haeryfar, S. M. M., Shaler, C. R. & Rudak, P. T. Mucosa-associated invariant T cells in malignancies: a faithful friend or formidable foe? Cancer Immunol. Immunother. 67, 1885–1896 (2018).
Heinrich, B. et al. The tumour microenvironment shapes innate lymphoid cells in patients with hepatocellular carcinoma. Gut 71, 1161–1175 (2022).
Stamatiades, E. G. & Li, M. O. Tissue-resident cytotoxic innate lymphoid cells in tumor immunosurveillance. Semin. Immunol. 41, 101269 (2019).
Zhu, Y. et al. Fungal-induced glycolysis in macrophages promotes colon cancer by enhancing innate lymphoid cell secretion of IL-22. EMBO J. 40, e105320 (2021).
Dean, I. et al. Rapid functional impairment of natural killer cells following tumor entry limits anti-tumor immunity. Nat. Commun. 15, 683 (2024).
Brownlie, D. et al. Accumulation of tissue-resident natural killer cells, innate lymphoid cells, and CD8(+) T cells towards the center of human lung tumors. Oncoimmunology 12, 2233402 (2023).
Xiang, X., Wang, J., Lu, D. & Xu, X. Targeting tumor-associated macrophages to synergize tumor immunotherapy. Signal. Transduct. Target. Ther. 6, 75 (2021).
Zhang, H. et al. Roles of tumor-associated macrophages in anti-PD-1/PD-L1 immunotherapy for solid cancers. Mol. Cancer 22, 58 (2023).
Garrido-Martin, E. M. et al. M1(hot) tumor-associated macrophages boost tissue-resident memory T cells infiltration andsurvival in human lung cancer. J. Immunother. Cancer 8, e000778 (2020).
Bayón, L. G. et al. Role of Kupffer cells in arresting circulating tumor cells and controlling metastatic growth in the liver. Hepatology 23, 1224–1231 (1996).
Kimura, Y. et al. The innate immune receptor Dectin-2 mediates the phagocytosis of cancer cells by Kupffer cells for the suppression of liver metastasis. Proc. Natl Acad. Sci. USA 113, 14097–14102 (2016).
van der Bij, G. J. et al. Therapeutic potential of Kupffer cells in prevention of liver metastases outgrowth. Immunobiology 210, 259–265 (2005).
Babaei, G., Aziz, S. G. & Jaghi, N. Z. Z. EMT, cancer stem cells and autophagy; the three main axes of metastasis. Biomed. Pharmacother. 133, 110909 (2021).
Jiang, Y. & Zhan, H. Communication between EMT and PD-L1 signaling: new insights into tumor immune evasion. Cancer Lett. 468, 72–81 (2020).
Huang, Y., Hong, W. & Wei, X. The molecular mechanisms and therapeutic strategies of EMT in tumor progression and metastasis. J. Hematol. Oncol. 15, 129 (2022).
Lavin, Y. et al. Innate immune landscape in early lung adenocarcinoma by paired single-cell analyses. Cell 169, 750–765.e717 (2017).
Etzerodt, A. et al. Tissue-resident macrophages in omentum promote metastatic spread of ovarian cancer. J. Exp. Med. 217, e20191869 (2020).
Zhu, Y. et al. Tissue-resident macrophages in pancreatic ductal adenocarcinoma originate from embryonic hematopoiesis and promote tumor progression. Immunity 47, 597 (2017).
Gotwals, P. et al. Prospects for combining targeted and conventional cancer therapy with immunotherapy. Nat. Rev. Cancer 17, 286–301 (2017).
Wang, T. et al. Tissue-resident memory CD8(+) T cells in cancer immunology and immunotherapy. Pharmacol. Res. 159, 104876 (2020).
Vesely, M. D., Zhang, T. & Chen, L. Resistance mechanisms to anti-PD cancer immunotherapy. Annu. Rev. Immunol. 40, 45–74 (2022).
O’Donnell, J. S., Teng, M. W. L. & Smyth, M. J. Cancer immunoediting and resistance to T cell-based immunotherapy. Nat. Rev. Clin. Oncol. 16, 151–167 (2019).
Darvin, P., Toor, S. M., Sasidharan Nair, V. & Elkord, E. Immune checkpoint inhibitors: recent progress and potential biomarkers. Exp. Mol. Med. 50, 1–11 (2018).
Edwards, J. et al. CD103(+) tumor-resident CD8(+) T cells are associated with improved survival in immunotherapy-naïve melanoma patients and expand significantly during anti-PD-1 treatment. Clin. Cancer Res. 24, 3036–3045 (2018).
Garris, C. S. et al. Successful anti-PD-1 cancer immunotherapy requires T cell-dendritic cell crosstalk involving the cytokines IFN-γ and IL-12. Immunity 55, 1749 (2022).
Yao, T., Shooshtari, P. & Haeryfar, S. M. M. Leveraging public single-cell and bulk transcriptomic datasets to delineate MAIT cell roles and phenotypic characteristics in human malignancies. Front. Immunol. 11, 1691 (2020).
De Biasi, S. et al. Circulating mucosal-associated invariant T cells identify patients responding to anti-PD-1 therapy. Nat. Commun. 12, 1669 (2021).
Hu, J. et al. Tumor microenvironment remodeling after neoadjuvant immunotherapy in non-small cell lung cancer revealed by single-cell RNA sequencing. Genome Med. 15, 14 (2023).
Gaudreau, P. O. et al. Neoadjuvant chemotherapy increases cytotoxic T cell, tissue resident memory T cell, and B cell infiltration in resectable NSCLC. J. Thorac. Oncol. 16, 127–139 (2021).
Kaczmarek, M. et al. Cancer vaccine therapeutics: limitations and effectiveness-a literature review. Cells 12, 2159 (2023).
Çuburu, N. et al. Intravaginal immunization with HPV vectors induces tissue-resident CD8+ T cell responses. J. Clin. Invest. 122, 4606–4620 (2012).
Sun, Y. Y. et al. Local HPV recombinant vaccinia boost following priming with an HPV DNA vaccine enhances local HPV-specific CD8+ T-cell-mediated tumor control in the genital tract. Clin. Cancer Res. 22, 657–669 (2016).
Nizard, M. et al. Induction of resident memory T cells enhances the efficacy of cancer vaccine. Nat. Commun. 8, 15221 (2017).
Sandoval, F. et al. Mucosal imprinting of vaccine-induced CD8+ T cells is crucial to inhibit the growth of mucosal tumors. Sci. Transl. Med. 5, 172ra120 (2013).
Gálvez-Cancino, F. et al. Vaccination-induced skin-resident memory CD8(+) T cells mediate strong protection against cutaneous melanoma. Oncoimmunology 7, e1442163 (2018).
Murray, T. et al. Very late antigen-1 marks functional tumor-resident CD8 T cells and correlates with survival of melanoma patients. Front. Immunol. 7, 573 (2016).
Chen, H. et al. Personalized neoantigen vaccine combined with PD-1 blockade increases CD8(+) tissue-resident memory Tcellinfiltration in preclinical hepatocellular carcinoma models. J. Immunother. Cancer 10, e004389 (2022).
Sterner, R. C. & Sterner, R. M. CAR-T cell therapy: current limitations and potential strategies. Blood Cancer J. 11, 69 (2021).
Craddock, J. A. et al. Enhanced tumor trafficking of GD2 chimeric antigen receptor T cells by expression of the chemokine receptor CCR2b. J. Immunother. 33, 780–788 (2010).
O'Rourke, D. M. et al. A single dose of peripherally infused EGFRvIII-directed CAR T cells mediates antigen loss andinduces adaptive resistance in patients with recurrent glioblastoma. Sci. Transl. Med. 9, eaaa0984 (2017).
Jung, I. Y. et al. Tissue-resident memory CAR T cells with stem-like characteristics display enhanced efficacy against solid and liquid tumors. Cell Rep. Med. 4, 101053 (2023).
Matlung, H. L., Szilagyi, K., Barclay, N. A. & van den Berg, T. K. The CD47-SIRPα signaling axis as an innate immune checkpoint in cancer. Immunol. Rev. 276, 145–164 (2017).
Wen, J., Wang, S., Guo, R. & Liu, D. CSF1R inhibitors are emerging immunotherapeutic drugs for cancer treatment. Eur. J. Med. Chem. 245, 114884 (2023).
Peng, C. et al. Engagement of the costimulatory molecule ICOS in tissues promotes establishment of CD8(+) tissue-resident memory T cells. Immunity 55, 98–114.e115 (2022).
Gustafsson, K. et al. Clearing and replacing tissue-resident myeloid cells with an anti-CD45 antibody-drug conjugate. Blood Adv. 7, 6964–6973 (2023).
Mondal, M., Guo, J., He, P. & Zhou, D. Recent advances of oncolytic virus in cancer therapy. Hum. Vaccines Immunother. 16, 2389–2402 (2020).
Lin, D., Shen, Y. & Liang, T. Oncolytic virotherapy: basic principles, recent advances and future directions. Signal. Transduct. Target. Ther. 8, 156 (2023).
Hashimoto, M. et al. Long-term activation of anti-tumor immunity in pancreatic cancer by a p53-expressing telomerase-specific oncolytic adenovirus. Br. J. Cancer 130, 1187–1195 (2024).
Zhang, Y. & Zhang, Z. The history and advances in cancer immunotherapy: understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell Mol. Immunol. 17, 807–821 (2020).
Waldmann, T. A. Cytokines in cancer immunotherapy. Cold Spring Harb. Perspect. Biol. 10, a028472 (2018).
Xie, Y. Q. et al. Redox-responsive interleukin-2 nanogel specifically and safely promotes the proliferation and memory precursor differentiation of tumor-reactive T-cells. Biomater. Sci. 7, 1345–1357 (2019).
Sowell, R. T. et al. IL-15 complexes induce migration of resting memory CD8 T cells into mucosal tissues. J. Immunol. 199, 2536–2546 (2017).
Tang, L. et al. Enhancing T cell therapy through TCR-signaling-responsive nanoparticle drug delivery. Nat. Biotechnol. 36, 707–716 (2018).
Zhao, T. et al. Vaccine adjuvants: mechanisms and platforms. Signal. Transduct. Target. Ther. 8, 283 (2023).
Lai, J. C. Y. et al. Topical adjuvant application during subcutaneous vaccination promotes resident memory T cell generation. J. Immunol. 203, 2443–2450 (2019).
Waldmann, H. & Cobbold, S. Exploiting tolerance processes in transplantation. Science 305, 209–212 (2004).
Prosser, A. C., Kallies, A. & Lucas, M. Tissue-resident lymphocytes in solid organ transplantation: innocent passengers or the key to organ transplant survival? Transplantation 102, 378–386 (2018).
Aiello, S., Benigni, A. & Remuzzi, G. Tissue-resident macrophages in solid organ transplantation: harmful or protective? J. Immunol. 212, 1051–1061 (2024).
Turner, D. L., Gordon, C. L. & Farber, D. L. Tissue-resident T cells, in situ immunity and transplantation. Immunol. Rev. 258, 150–166, (2014).
Fu, J. & Sykes, M. Emerging concepts of tissue-resident memory T cells in transplantation. Transplantation 106, 1132–1142 (2022).
Snyder, M. E. et al. Generation and persistence of human tissue-resident memory T cells in lung transplantation. Sci. Immunol 4, eaav5581 (2019).
Bartolomé-Casado, R. et al. Resident memory CD8 T cells persist for years in human small intestine. J. Exp. Med. 216, 2412–2426 (2019).
Lian, C. G. et al. Biomarker evaluation of face transplant rejection: association of donor T cells with target cell injury. Mod. Pathol. 27, 788–799 (2014).
Yuan, R. et al. Critical role for CD103+CD8+ effectors in promoting tubular injury following allogeneic renal transplantation. J. Immunol. 175, 2868–2879 (2005).
Abou-Daya, K. I. et al. Resident memory T cells form during persistent antigen exposure leading to allograft rejection. Sci. Immunol 6, eabc8122 (2021).
Nayak, D. K. et al. Long-Term Persistence of Donor Alveolar Macrophages in Human Lung Transplant Recipients That Influences Donor-Specific Immune Responses. Am. J. Transpl. 16, 2300–2311 (2016).
Nayak, D. K. et al. Zbtb7a induction in alveolar macrophages is implicated in anti-HLA-mediated lung allograft rejection. Sci. Transl. Med. 9, eaal1243 (2017).
Liu, Z. et al. Reprogramming alveolar macrophage responses to TGF-β reveals CCR2+ monocyte activity that promotesbronchiolitis obliterans syndrome. J. Clin. Invest. 132, e159229 (2022).
Parlakpinar, H. & Gunata, M. Transplantation and immunosuppression: a review of novel transplant-related immunosuppressant drugs. Immunopharmacol. Immunotoxicol. 43, 651–665 (2021).
Adachi, Y. et al. Distinct germinal center selection at local sites shapes memory B cell response to viral escape. J. Exp. Med. 212, 1709–1723 (2015).
Macri, C., Pang, E. S., Patton, T. & O’Keeffe, M. Dendritic cell subsets. Semin. Cell Dev. Biol. 84, 11–21 (2018).
Naik, S. H. et al. Intrasplenic steady-state dendritic cell precursors that are distinct from monocytes. Nat. Immunol. 7, 663–671 (2006).
Chen, K. et al. Tissue-resident dendritic cells and diseases involving dendritic cell malfunction. Int. Immunopharmacol. 34, 1–15 (2016).
Lavin, Y. et al. Tissue-resident macrophage enhancer landscapes are shaped by the local microenvironment. Cell 159, 1312–1326 (2014).
Sajti, E. et al. Transcriptomic and epigenetic mechanisms underlying myeloid diversity in the lung. Nat. Immunol. 21, 221–231 (2020).
Steinbach, K. et al. Brain-resident memory T cells generated early in life predispose to autoimmune disease in mice. Sci. Transl. Med. 11, eaav5519 (2019).
Wang, T. et al. Influenza-trained mucosal-resident alveolar macrophages confer long-term antitumor immunity in the lungs. Nat. Immunol. 24, 423–438 (2023).
Farber, D. L. Tissues, not blood, are where immune cells function. Nature 593, 506–509 (2021).
Fernandez-Ruiz, D. et al. Liver-resident memory CD8(+) T cells form a front-line defense against malaria liver-stage infection. Immunity 51, 780 (2019).
Savas, P. et al. Single-cell profiling of breast cancer T cells reveals a tissue-resident memory subset associated with improved prognosis. Nat. Med. 24, 986–993 (2018).
Losurdo, A. et al. Single-cell profiling defines the prognostic benefit of CD39(high) tissue resident memory CD8+ T cells in luminal-like breast cancer. Commun. Biol. 4, 1117 (2021).
Petley, E. V. et al. MAIT cells regulate NK cell-mediated tumor immunity. Nat. Commun. 12, 4746 (2021).
Zhu, Y. et al. Tissue-resident macrophages in pancreatic ductal adenocarcinoma originate from embryonic hematopoiesis and promote tumor progression. Immunity 47, 323–338.e326 (2017).
Acknowledgements
This work was supported by the National Key R&D Program of China (2021YFF1201303), the CAMS Innovation Fund for Medical Sciences (CIFMS) (2021-1-I2M-012), and the Beijing Natural Science Foundation (L248050).
Author information
Authors and Affiliations
Contributions
J.H. and C.L. conceived the work. J.L. and C.X. drafted the manuscript. J.L. and C.X. performed visualization. C.L. and J.H. revised the manuscript. All authors have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Li, J., Xiao, C., Li, C. et al. Tissue-resident immune cells: from defining characteristics to roles in diseases. Sig Transduct Target Ther 10, 12 (2025). https://doi.org/10.1038/s41392-024-02050-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41392-024-02050-5
This article is cited by
-
The dynamics of brain T cell populations during the course of rasmussen encephalitis: from expansion to exhaustion
Journal of Neuroinflammation (2025)
-
Impact of exercise on immune cell infiltration in muscle tissue: implications for muscle repair and chronic disease
Clinical and Experimental Medicine (2025)
