
Abstract
Pseudomyxoma peritonei (PMP) is a rare neoplasm coursing with uncontrollable mucus accumulation, with a high relapse rate. RNA biology processes have emerged as new players in cancer development and progression, nevertheless their role in PMP remains unknown. In this study, we aimed to examine RNA-regulatory machineries in PMP and their potential contribution to this disease progression. We analyzed 62 splicing-related genes, 27 RNA exosome and 21 nonsense-mediated decay genes, in a cohort of 29 patients using a microfluidic array, comparing tumor and control/reference tissues, together with external RNA-seq and proteomic data. Our results revealed a profound dysregulation of key components, which correlated to relevant clinical parameters and enabled to distinguish between tumor and control tissues. In vitro splicing inhibition using Pladienolide-B, as well as the modulation of specific splicing factors, reduced aggressiveness parameters, enhanced the effect of clinically used drugs, and revealed a strong correlation between dysregulated genes and key cancer-related genes. This inhibition also affected mucin secretion and mucin variants production. Collectively, our findings provide the first evidence for dysregulation of the genes of pivotal RNA-regulatory processes in PMP, implying that these targetable mechanisms may be functionally altered and play a role in the disease. Hence, a thorough understanding of its RNA biology could aid in the discovery of new clinically actionable vulnerabilities in this rare disease.

Similar content being viewed by others

Introduction
Pseudomyxoma peritonei (PMP) is a rare, progressive, slow-proliferating tumor, with an estimated incidence of 3.2 cases per million per year [1]. PMP is typically caused by the spread and metastasis of appendiceal mucinous neoplasms throughout the peritoneal cavity. The most efficient treatment for patients with PMP is cytoreductive surgery followed by hyperthermic intraperitoneal chemotherapy (HIPEC), with mitomycin-C and cisplatin [2, 3]. These therapeutic options have improved PMP patients’ overall survival (OS); however, the low relapse-free progression (RFP) rate (48%) despite the treatment remains a major issue [4]. In addition, classification of PMP has been controversial. The main approach currently accepted relies on histological evaluation, which allows to divide it into three well-separated categories: low-grade (LG), high-grade (HG), and high-grade with signet ring cells (SRC) tumors [5]. In this line, the use of Ki67 index has recently proven to be a valuable prognostic factor for HG PMP, dividing it into two subcategories with different prognoses [6].
PMP molecular profile is still poorly characterized. Multiomics studies have successfully guided treatment in other cancers; however, although initial studies have described the genetic profile of PMP, there are currently no specific genetic targets that could facilitate treatments with fewer adverse side effects [6]. Knowledge of PMP transcriptomic landscape is more limited, although some studies have classified PMP into different transcriptomics groups: immune-enriched, oncogenes-enriched, or mixed [7]; or based on prognosis. At this point, it is worth noting that, to better understand a disease, recent studies do not solely focused on the transcriptome, but in the processes that regulate the final set of transcripts, as we recently reviewed regarding the different dimensions of RNA processing in neuroendocrine tumors, another type of infrequent neoplasms [8]. As illustrated therein, RNA molecules are finely surveyed and refined at various stages by distinct processes, including 5’ capping, splicing, 3’ end formation, and nuclear export, while competing kinetically with decay-targeting machinery such as nonsense, nonstop or no-go mediated decays, or RNA exosome [8]. Proper RNA-protein and protein-protein interactions must be assured in these macromolecular-guided processes, and fine regulation is essential, since disruptions in these mechanisms have been linked to specific cell dysfunctions and several diseases [9,10,11,12]. Among the mentioned processes, alternative splicing—the excision of the intronic regions in the pre-RNA and subsequent binding of the exons to generate the mature RNA—disruption is currently considered a hallmark of cancer [13,14,15,16]. In addition, the main RNA surveillance processes, Nonsense-mediated decay (NMD) and RNA exosome are responsible for identifying and degrading mispliced or aberrant RNA transcripts, thereby maintaining the vital integrity of gene expression and preventing the accumulation of potentially harmful proteins [17, 18]. In sum, current evidence points to dysregulation of the splicing machinery and RNA surveillance mechanisms as critical players contributing to cancer by the generation and accumulation of aberrant splicing variants with oncogenic potential, which may contribute to tumor development and progression. To date, however, this hypothesis remains unexplored in PMP. Therefore, this study aims to characterize the splicing and RNA surveillance landscape in PMP, evaluate their potential diagnostic and prognostic value, and assess the relationship between putative alterations in the splicing process and clinical parameters, to identify vulnerabilities in this rare and clinically challenging tumor.
Material and Methods
Patient samples
A cohort of 29 fresh samples (discovery cohort) collected and stored in the Andalusian Biobank (Cordoba Node) was used to perform the present study. Specifically, we selected 11 control samples (5 appendix, 4 peritoneum, 1 colon and 1 mesentery samples derived from non-tumor tissue) and 18 PMP samples (10 LG and 8 HG tumors). Histological studies were performed by experienced pathologists to identify tumor and healthy tissue. Clinicopathological data of the cohort are described in Table 1. All experiments were conducted in accordance with the ethical standards of the Helsinki Declaration, of the World Medical Association and with the approval of the University of Cordoba/IMIBIC and Ethics Committees from the Hospital involved in the study (protocol code PI19/01603, v1 – 13/03/2019). Informed consent from each patient was obtained through the Andalusian Biobank.
Two different RNA-seq datasets were obtained from GEO database. RNA-seq (GSE228376), generated from fresh frozen tissue, was composed of 19 samples: 7 controls and 12 PMP samples (12 LG). The second one, RNA-seq (GSE228375), was derived from formalin-fixed, paraffin-embedded tissue (FFPE), and includes 34 samples: 9 controls and 25 PMP samples (19 LG, 5 HG and 1 SRC). Both RNA-seq datasets were performed by Illumina NovaSeq 6000.
Proteomic data was downloaded from ProteomeXchange (PXD037364, PRIDE repository) and included protein quantification results from 25 samples: 10 controls and 15 tumors (8 LG PMP and 7 HG PMP). Identification and quantification of proteins was performed using the TripleTOF 6600 instrument [19].
Cell models and culture conditions
For functional assays, we used a PMP-derived cell line, N14A (ECACC 16110902), kindly provided by Dr. Andrew G. Renehan (Division of Cancer Sciences, School of Medical Sciences, University of Manchester, Manchester, UK) and Dr. Patrick T. Caswell (Wellcome Trust Centre for Cell-Matrix Research, School of Biological Sciences, Faculty of Biology Medicine and Health, Manchester Academic Health Science Centre, University of Manchester, Manchester, UK) [20]. This cell model was established from PMP primary tissue, derived from a female patient. The STR profile was performed when the cell line was developed and matched the genomic DNA isolated from FFPE samples of the same patients, confirming the same origin. Also, the STR profile did not match any cell lines from ATCC (American Type Culture Collection) [20], and to ensure the authenticity of our N14A cell line, we have repeated the STR profiling and confirmed that matches its original description. The cells were cultured in a humidified environment containing 5% CO2 and were regularly examined for mycoplasma contamination [18]. Briefly, cells were cultured in Dulbecco’s Modified Eagle’s Medium (Sigma-Aldrich, Madrid, Spain) with 4.5 g/L of glucose supplemented with 10% fetal bovine serum (FBS; Sigma-Aldrich), 10 mM L-glutamine (Sigma-Aldrich), 25 mM HEPES (Gibco, Madrid, Spain), 5 µg/mL insulin (Gibco) and 0.2% antibiotic/antimycotic (Gentamicin/Amphotericin B; ThermoFisher Scientific, Spain).
In vitro experiments
To evaluate splicing inhibition effect over different functional parameters in vitro, we used Pladienolide-B (Pd; Santa Cruz Biotechnology, Heidelberg, Germany), resuspended in dimethyl sulfoxide (DMSO) and used in the 10-8–10-10 M range. Pladienolide-B directly binds to and impairs the activity of SF3B1, a core protein of the spliceosome [21], hindering spliceosome assembly and, in consequence, changing splicing pattern. For each experiment, this treatment was refreshed every day. To check whether the splicing inhibition in vitro could improve the effect of some of the most used treatments for PMP in the clinics, we combined Pladienolide-B (10-9 M) with one unique exposition during 24 h to mitomycin-C (4 µg/mL; Sigma-Aldrich) and cisplatin (0.5×10-3 M; Sigma-Aldrich), both chemotherapeutic agents widely used in PMP [2]. The doses and time of exposition were selected based on bibliography [18, 20,21,22,23].
Gene expression modulation
Pre-designed and validated specific small interfering RNAs (siRNA) and overexpression plasmids were used to change the expression of selected splicing factors discovered to be altered at the RNA-level. For endogenous PTBP1 silencing 75 nM of the specific siRNA (1:1 from B-C sequences; SR303857; Trilencer-27 siRNA kit; Origene, Rockville, MD, USA) and Lipofectamine RNAiMax® (ThermoFisher Scientific) were used according to the manufacturer’s instructions. Silencer® Select Negative Control siRNA (ThermoFisher Scientific) was used as a scramble control. Two different expression plasmids were utilized to overexpress HNRNPK (Origene, #RC201843) and MBNL1 (Origene, #RC208112) using 1 µg/mL of each one and Lipofectamine 3000 Reagent (ThermoFisher Scientific) following the manufacturer’s instructions. Empty pCMV6‐Entry plasmid served as negative control. 500000 cells per well were seeded for each transfection. After 24 h, cells were collected for validation of the transfection (qPCR) and seeded for different functional assays.
Cell viability
To measure Pladienolide-B effect, alone and in combination with the chemotherapeutics, as well as the effect of the silencing and overexpression of the selected splicing factors over cell proliferation, resazurin (Canvax Biotech S.L., Cordoba, Spain) assay, an indirect method that allow to estimate cell proliferation by measuring cell viability, was used as previously described [24]. Briefly, 6000 cells/well were seeded in 96-well plates and serum-starved for 12 h. Resazurin (10%) was then added, and fluorescence measured after 3 h of incubation with FlexStation III (Molecular Devices, San José, CA, USA) at 0, 24, 48 and 72 h of culture.
Wound‑healing assay
N14A migration capacity was evaluated after 24 h of Pladienolide-B treatment in a wound-healing assay, as previously reported [24]. 40000 cells/well were seeded in 96-well plates and serum-starved for 2 h when confluence was reached. Then, a wound was made in the center of each well using a 100 µL pipette tip. Specifically, images of the scratch were taken after 0 and 16 h and the area covered by the cells was registered. ImageJ software v2.1.2 (NIH, Bethesda, Maryland, USA) was used to calculate the area of the scratch in each case.
Cell medium turbidity
N14A cells produce a layer of mucin in the flask, equivalent to the clinical PMP phenotype [20]. We assessed mucin production in response to treatment by measuring optical density (OD) of the cell culture medium at 900 nm, ensuring no interference from the colorant and other components of the media. Summarizing, 250000 cells/well were seeded in 6-well plates, serum-starved for 24 h and treated for 3 days. Turbidity was measured using the SPECTROstar Nano (BMG LABTECH, Ortenberg, Germany) spectrophotometer. Given the structural complexity of MUC2 three isoforms, that do not share any common sequence, and the lack of antibodies detecting all of them, medium turbidity was used as an indirect measure of mucin secretion, providing a functional readout of the mucinous phenotype in these cells.
RNA isolation and retrotranscription
Total RNA was extracted from 30 mg of frozen tissue samples kept at -80 °C immediately after surgery, using the commercial kit for RNA purification AllPrep RNA/Protein Kit (Qiagen, Gilden, Germany) according to the manufacturer’s recommendations. In the case of RNA extracted from N14A cells, 250000 cells/well were seeded in 6-well plates, and collected using TRIzol Reagent (Invitrogen, Barcelona, Spain) following the manufacturer’s instructions. RNA concentration and purity was always evaluated by Nanodrop One Spectrophotometer (ThermoFisher Scientific) and treated with DNase (Promega, Madison, USA). Total RNA was retrotranscribed using the RevertAid RT Reverse Transcription Kit (ThermoFisher Scientific), following the manufacturer’s instructions.
Quantitative real-time PCR (qPCR)
To measure gene expression, qPCR was performed using the Brilliant III SYBR Green-qPCR MasterMix (Stratagene, La Jolla, CA) in the Stratagene Mx3000p system, as previously described [24]. Specific primers for human transcripts were designed with Primer3 (Applied Biosystems, Foster City, USA) and Primer Blast software as previously reported [24]. Sequences of all primers are shown in Supplemental Table 1. For RNA expression normalization, three different housekeeping genes were tested: ACTB (actin beta), GAPDH (glyceraldehyde 3-phosphate dehydrogenase) and HPRT1 (hypoxanthine phosphoribosyltransferase 1); in the case of N14A cell line, only GAPDH resulted a feasible housekeeping according to geNorm 3.5 analysis [25].
qPCR microfluidic dynamic array
RNA expression level of a set of 110 RNA biology-related genes (Supplemental Table 2) were simultaneously determined in the 29 collected samples by a qPCR dynamic array based on microfluidic technology, as previously reported by our group [24]. Particularly, we evaluated: 62 genes of the splicing machinery, 27 of the RNA exosome complex, 21 of the NMD system, a set of genes involved in cancer altered pathways, as cell cycle, proliferation, immunomodulation and inflammation, and MUC2 isoforms (MUC2-206, canonical isoform; MUC2-204 and MUC2-205). Primers for specific human transcripts were designed with Primer3 and Primer Blast software (see Supplemental Table 1). Biomark System and the Fluidigm Real-Time PCR Analysis Software v3.0.2 and Data Collection Software v3.1.2 (Fluidigm, San Francisco, USA) were used to acquire RNA expression levels. Data were normalized by the normalization factor (NF) of the three housekeeping genes obtained using geNorm 3.5.
Western blot
N14A cells transfected with the specific siRNA (siR PTBP1) or plasmids (overexpression plasmid for MBNL1 and HNRNPK) were seeded (500000 cells/well in a 6-well plate) and lysed using SDS-DTT (Sigma-Aldrich). Protein was denaturalized as previously reported [24] and separated in 4–20% Criterion™ TGX™ Precast Midi Protein Gel (Bio-Rad Laboratories, Madrid, Spain) by SDS-PAGE, transferred to a nitrocellulose membrane (Ref. 1,704,270, Millipore) and blocked with 5% non-fat dry milk in Tris-buffered saline with 0.05% Tween-20 (Ref. 93,773, Sigma-Aldrich). Membranes were then incubated with the following primary antibodies: PTBP1 (1:1000, PA5-81297, ThermoFisher Scientific), MBNL1 (1:500, PA5-96911, ThermoFisher Scientific), HNRNPK (1:500, PA5-95866, ThermoFisher Scientific), phospho-ERK1/2 (1:1000, 4370S, Cell Signaling Technology; Danvers, MA, USA), ERK1/2 (1:1000, SC-154, Santa Cruz Biotechnology; Santa Cruz, CA, USA), phospho-AKT (1:1000, 9271S, Cell Signaling Technology), AKT (1:1000, 9272S, Cell Signaling Technology) and GAPDH, used as housekeeping (1:1000, AM4300, ThermoFisher Scientific). Then, horseradish peroxidase-conjugated goat anti-rabbit IgG (Ref. 7074, Cell Signaling Technology) was used. Bond antibodies were visualized using the Clarity Western-ECL Blotting Substrate (Bio-Rad Laboratories) and scanned using ImageQuant Las 4000 system (GE Healthcare Europe GmbH). Images were analyzed using ImageLab 5.0 software.
Immunohistochemical analysis
Immunohistochemistry (IHC) analyses were performed on a representative collection of FFPE samples, selecting a subset of them showing the highest cellularity. Therefore, we analyzed 3 control, 2 LG and 2 HG samples. Briefly, deparaffinized sections were incubated overnight (4 °C) with the primary antibodies against the proteins of interest: HNRNPK Polyclonal Antibody (1:50, PA5-95866, ThermoFisher Scientific), MBNL1 Polyclonal Antibody (1:50, PA5-96911, ThermoFisher Scientific), PTBP1 Polyclonal Antibody (1:500, PA5-81297, ThermoFisher Scientific) and RAVER1 Polyclonal Antibody (1:30, PA5-60424 and PA5-61977, ThermoFisher Scientific), as recommended in each datasheet. Then, samples were incubated with anti-rabbit horseradish peroxidase antibodies at 1:250 dilution (#7074; Cell Signaling, Danvers, MA, USA). Finally, sections were developed with diaminobenzidine (Envision system 2-Kit Solution DAB) and contrasted with hematoxylin.
Statistic and bioinformatic analysis
Data were evaluated for heterogeneity of variance and normal distribution using the Kolmogorov-Smirnov test. Statistical differences were assessed by t-test, Mann-Whitney U test or by one-way ANOVA followed by Fisher’s correction exact test. Correlations were studied using Pearson or Spearman correlation test, depending on the normality. RStudio (v4.2.2) was used for survival and relapse analysis (package survival v3.4.0), to represent the linear correlations (package ggplot2 v3.5.1), as well as to analyze and represent the RNA-seq and proteomic data using the packages tidyverse v2.0.0, dplyr v1.1.4 and ggplot2 v3.5.1; enrichment analysis of the RNA-seq and proteomics data were performed using fgsea v1.24.0 package and the hallmark gene set from the Human MSigDB Collections (GSEA H gene set). Clustering analyses and PLS-DA (Partial Least Squares-Discriminant Analysis) were performed with MetaboAnalyst v5.0 (McGill University, Quebec, Canada). The rest of the statistical analyses were performed using Prism v9.0 (GraphPad, La Jolla, CA, USA). To normalize data within treatment and control groups and minimize intragroup differences, acquired values were compared to controls (set at 100%). In vitro results were generated from at least three separate independent experiments. Data are represented with median and interquartile range or mean ± standard error of the mean (SEM). *p < 0.05; **p < 0.01; ***p < 0.001, ****p < 0.0001 indicated statistically significant differences.
Results
Patient characteristics and treatment
A total of 29 patient samples from our discovery cohort were studied, including 11 control samples used as reference, consisting of non-tumor tissue from patients subjected to PMP surgery or patients underwent appendicectomy for benign causes, and 18 PMP samples: 10 LG and 8 HG according to the PSOGI classification [5] (Table 1). Percentage of women was 51.72% (15 of 29) and median age at diagnosis 60 (46–70) years. Median perioperative PCI (Peritoneal Cancer Index) was 28 (19–39) and 27.8% of the patients received adjuvant treatment. Completeness cytoreduction 0 and 1 (CC0–CC1) was achieved in 77.8% of patients, and CC2–CC3 was achieved in 22.2%. A 16.7% of patients presented a major morbidity (Clavien-Dindo ≥ 3). At the end of the follow-up, 17.6% of patients suffered from relapse after the treatment with a median RFP of 20 (14–28) months and 76.5% of patients were alive, with a median follow-up of 21 (15–28) months. Demographic and clinical characteristics of patients included in the RNA-seq analyses are available at GEO (GSE228375, GSE228376). Patients included in the proteomic analyses have been previously characterized [19].
Splicing, RNA exosome and NMD machineries are dysregulated in PMP
Expression level of 110 genes were evaluated in the discovery cohort using a qPCR microfluidic array. No differences were detected among the control samples, suggesting that the RNA processing machineries under study are stable in healthy tissue. Regarding the LG and HG samples, while marked differences in the expression of several genes were observed with respect to the controls, we only detected minimal variances between the LG and HG, and therefore we decided to consider the 18 PMP samples as a joint group henceforth (Fig. S1). Close analysis of results revealed notable differences between the expression of these machineries’ components in the control and PMP samples, suggesting a dysregulation of these genes at the RNA level. Specifically, 24.19% of studied genes were significantly altered (Fig. 1A), which allowed to cluster samples in distinct groups (Fig. 1B). Furthermore, PLS-DA demonstrated that dysregulated RNA-related gene expression can distinguish between most of the control and PMP samples (Fig. 1B). The splicing-related genes HNRNPK, RAVER1, MBNL1, RNU6ATAC and PTBP1 contributed the most to the discrimination model according to the Variable Importance in Projection (VIP) Score analysis (VIP-Score > 1.5). Splicing-related genes were the highest altered between control and PMP (24.19%), highlighting that most of them were downregulated in the tumor, except for two that were upregulated: PTBP1 and RAVER1 (Fig. 1A). Most of the altered genes were splicing factors, but it was also striking the contribution to this dysregulation of some of the RNUs core of the major spliceosome (RNU6) and, specially, the minor spliceosome (RNU4ATAC, RNU6ATAC), which catalyzes the splicing of less than the 1% of human introns [26]. 18.52% genes were found altered in the RNA exosome complex, all of them downregulated in PMP; and 23.81% of NMD genes were dysregulated, only one of them upregulated, SMG9, and the rest downregulated, in line with what was observed in the other machineries. The disruption in the expression of splicing machinery, RNA exosome and NMD components was corroborated in the two external RNA-seq validation cohorts analyzed (Fig. S2, Fig. S3A), in which not only downregulation of HNRNPK, MBNL1, NOVA1 and RNU4ATAC (splicing machinery), C1D (RNA exosome) and ETF1 (NMD) were confirmed, but also other alterations were uncovered. It is worth noting that the results from the FFPE RNA-seq were more similar to those obtained in our discovery cohort than the ones from the fresh frozen samples RNA-seq, probably due to the more comparable composition of the type of samples (LG and HG PMP). Of note, also, only 3 genes were differentially expressed among PMP grades, the splicing factors HNRNPK, MBNL1 and RBMX, whose expression was decreased in HG (Fig. S3B). Proteomic data analysis only pointed to 2 statistically significant alterations: overexpression of the splicing factor HNRNPA1 and the NMD component ALYREF in PMP (Fig. S4A). Nevertheless, comparison of PMP with different grades also revealed that HNRNPK splicing factor tended to be downregulated in LG and PTBP1 was overexpressed in HG compared to control samples, while RBMX expression was decreased in HG as compared to LG (Fig. S4B). It is worth to highlight the difficulty of a proteomic study in this mucinous tumor, which makes it extremely hard to identify proteins expressed at low levels. In fact, only 1025 proteins were found in PMP, with only 8 of them belonging to the splicing machinery, 1 to the RNA exosome complex and 1 to the NMD, and therefore it was not feasible to perform a standard validation of the differences found at the RNA levels of the rest of the proteins of interest (Fig. S4).

A RNA expression level of each machinery individually as Log2 of the Fold-Change Tumor/Control. In the upper panel the expression of all genes is represented showing upregulated (colored in red) and downregulated genes (colored in blue) as the Log2 of the Fold-Change of Tumor/Control (n = 11 controls; n = 18 tumors); in the graph below are represented the statistically significant genes altered as violin plots as Log2 of normalized copy number, showing the median expression and the first and third quartiles (thin lines). T-test or Mann–Whitney U test were used to compare means between the two groups. Asterisks represent significant differences: *p < 0.05; **p < 0.01; ****p < 0.0001. B PLS-DA and VIP-Score of all significantly altered genes. PLS-DA shows two different clusters: control tissue (“0”, green) and PMP tissue (“1”, red); in the right panel is the score of each of these genes (top 15) contribution to the discrimination model, VIP-Score, and the mean expression of each gene in the control, “0”, and PMP, “1”, groups (“high expression” in red, “low expression” in blue).
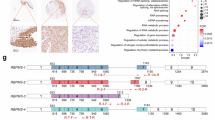
The typical low cellularity that characterizes PMP also hinders its study with IHC techniques. This notwithstanding, to demonstrate the presence and precise distribution of some of the genes altered in the tumors at protein level in our discovery cohort and assess their potential contribution to discriminate between control and PMP, we applied IHC to study HNRNPK, MBNL1, PTBP1 and RAVER1 (VIP-Score > 1.5) in a selected set of controls and PMP samples showing the highest number of cells (Fig. 2). This approach allowed us to visualize the expression of three of these genes and determine their specific tissue and cellular distribution, although a quantitative analysis of their protein levels was precluded by the marked differences in cellularity between the control tissue and PMP. We found that in the control samples, HNRNPK, MBNL1 and PTBP1 proteins were mostly uniformly distributed in the nucleus of all cells in the tissue, in contrast with the PMP tissue, wherein each factor was expressed with a higher intensity in goblet cells that give rise to the tumor; HNRNPK, MBNL1 in the nucleus and cytoplasm, and PTBP1 mostly in the nucleus. In contrast, RAVER1 was not detected by IHC despite testing different antibodies and protocols, which, according to the manufacturer information, successfully stained other tissues. To rule out technical issues, we optimized the staining conditions, tested different antibody concentrations, and validated the protocol with other markers under the same conditions. Unfortunately, none of these approaches allowed for RAVER1 detection in either PMP or control samples. This suggests a specific issue with the antibodies in the context of our samples or technical limitations inherent to IHC for this protein (Fig. S5).

Representative IHC images (10X) showing the distribution of HNRNPK, MBNL1 and PTBP1 in FFPE control samples (appendix) and in LG and HG PMP. Zoomed images of the regions framed in blue (20X) and pink (40X) are shown in the bottom panel.
Expression of splicing, RNA exosome and NMD genes possesses discriminant potential and seems relevant in PMP prognosis
To determine whether alterations found in RNA biology-related machineries could be clinically relevant in PMP patients, correlations between expression of these genes and key clinical parameters of PMP aggressiveness were assessed (Fig. 3A). This revealed associations in the three machineries, particularly to OS and RFP, suggesting that dysregulations in these genes may be linked to patient clinical outcomes. Expression of 48.39% of the evaluated splicing-related genes was associated with poor OS, and that of 9.68% with decreased RFP, being remarkable the cases of HNRNPA3, HNRNPK, RBM17 and SRSF3, which are downregulated in PMP, and its lower expression associated to lower OS (Fig. S6). Expression of 22.58% genes was inversely correlated to Ki67, which may suggest that the general downregulation of splicing-related genes is linked to worse prognosis in PMP. Correlations with other parameters were less frequent/intense, with only certain associations potentially relevant to p53 expression level by IHC (3.23%), CK7 positivity (6.45%) and postoperative morbidity, Clavien-Dindo ≥ 3, (3.23%) (Fig. S7). Expression of RNA exosome and NMD machineries also showed associations to OS and RFP. In the first case, 74% of assessed RNA exosome genes were significantly associated with poor OS, where, interestingly, lower gene expression was always linked to reduced OS, suggesting that the general downregulation observed in the RNA exosome complex machinery may be connected to poor prognosis. In addition, 40% of RNA exosome genes correlated to lower RFP (Fig. S8). In contrast to the splicing machinery, increased expression of 18.52% of RNA exosome related genes was associated with higher Ki67 levels. Moreover, 14.81% of RNA exosome related genes were inversely associated with PCI score; SKIC3 expression was directly correlated to p53 level and EXOSC9 expression to CK7 positivity (Fig. S9). Finally, most of NMD genes (80.95%) were also associated with OS; in all cases, the low expression of these genes was related to low OS, highlighting GSPT1, RBM8A and SMG8. RFP was significantly associated with expression levels of 42.86% of NMD genes (Fig. S10). The correlation observed between expression of 28.57% of the assessed NMD genes and PCI score was noteworthy, in that, as observed in RNA exosome-related genes, downregulation was linked to high PCI in all cases. Also, the expression of three genes (ETF1, RBM8A, UPF2) was directly correlated to Ki67 (Fig. S10). Despite the limited clinical data available in the GEO RNA-seq validation cohorts, analysis of the correlations between the expression of the dysregulated RNA-related genes and relevant clinical characteristics revealed interesting associations with PCI, recurrence and comorbidity (Fig. S11). Among them, association of lower NOVA1 expression with comorbidity is highlighted by its significant downregulation at the RNA level in the three studied cohorts.

A Clinical correlations of all genes measured. Relevant clinical parameters in PMP as OS, RFP, Ki67, p53 levels, PCI, CK7 presence and morbidity were associated to some splicing, RNA exosome and NMD genes expression. These correlations are represented in the graph as the p-value in blue (for splicing machinery), green (RNA exosome complex), and orange (NMD) being the symbol bigger as the p-value is smaller. The upward triangle represents direct correlations, the inverted triangle represents inverse correlations. Logrank test, Pearson or Spearman correlation and t-test or Mann–Whitney U test were used for survival and relapse analysis, Ki67, p53 and PCI, and CK7 positivity and morbidity, respectively. B Best ROC curves for each machinery. In the splicing machinery (blue), RNU4ATAC was the best gene to discriminate between PMP and control tissue; RBM7 was the best among the RNA exosome (green) and SMG8 in the NMD (orange).
The clinical-molecular associations found supported our initial hypothesis about the possible role of RNA biology machineries in the pathobiology of PMP. Therefore, we decided to evaluate the discriminant potential of these genes between control and PMP patient tissues. Indeed, Receiver Operating Characteristic (ROC) analyses proved that a discrete set of the above-listed genes could be potentially helpful to distinguish between control and PMP samples (Fig. 3B, Fig. S12A). Specifically, 13 of the dysregulated splicing genes showed an Area Under the Curve (AUC) above 0.7, with RNU4ATAC standing out as the best gene for discriminating between the two groups; while 5 RNA exosome and 5 NMD genes displayed an AUC above 0.7, highlighting RBM7 and SMG8, respectively (Fig. 3B, Fig. S12A). In addition, to provide a more clinically relevant perspective, we further evaluated whether these genes could distinguish between tumor grades by applying the same analysis to differentially expressed genes between LG and HG PMP. In our cohort, EXOSC2 stood out as a key marker, successfully differentiating between LG and HG (Fig. S12B). We also assessed the discriminatory power of differentially expressed genes across the three additional available cohorts (Fig. S13), obtaining striking results, such as ESPR1 achieving a perfect ROC curve with an AUC = 1, and several other genes displaying AUC > 0.9, including PRPF40A and DPC2 in the fresh frozen cohort (Fig. S13A), as well as CELF1, HNRNPK, MBNL1, PRPF8, RNU11, RNU12, RBM25, RBMX, SRSF1, TIA1, DIS3, EXOSC8, DPC2, NCBP1, and XNR1, in the FFPE cohort (Fig. S13B), and HNRNPA1 and ALYREF in the proteomic cohort (Fig. S13C). Additionally, some of these genes demonstrated the ability to distinguish between LG and HG PMP, with HNRNPK and MBNL1 showing an AUC > 0.9 in the FFPE cohort and RBMX reaching an AUC > 0.8 in the proteomic dataset (Fig. S14A, Fig. S14B).These results underscore the substantial discriminant potential of some of the genes tested and thereby reinforce the idea that the machineries regulating RNA biology could be involved in PMP clinical evolution.
Cancer progression related genes were altered in PMP and correlated to splicing-related genes
To further examine the relationship between alterations in RNA biology and tumor progression, we evaluated a selected set of cell cycle, proliferation, immunomodulation, inflammation and angiogenesis genes in our discovery cohort. We observed differences in 9 genes evaluated: CDKN2A, CDKN2D, CDK4, CDK6, ATM (cell cycle and proliferation); CXCL3, CCL5, CXCR2 (immunomodulation and inflammation) and KDR (angiogenesis) (Fig. 4A, Supplemental Table 3). All cell cycle and proliferation genes but CDKN2D were upregulated in PMP. Two immunomodulation and inflammation genes (CXCL3 and CCL5) were also upregulated whereas CXCR2 was downregulated. KDR was also found upregulated in PMP. Then, we assessed the relationship between these genes and RNA biology-related genes. This revealed certain significant correlations in the case of RNA exosome (EXOSC2, MTREX and RBM7) and NMD (ETF1, GSPT1, RBM8A, SMG8, SMG9), while, in splicing-related genes, we identified 20 relationships with cell cycle and proliferation genes (CDKN2A, CDK4 and CDK6) and 2 with the immunoregulation and inflammation gene CCL5 (Fig. 4B, Figs. S15, S16). Interestingly, downregulated genes correlated inversely with CDKN2A, CDK6, and CCL5 whereas their correlation with CDK4 was direct. On the other hand, the two splicing factors overexpressed in PMP, PTBP1 and RAVER1 showed a direct correlation with CDK4. Additionally, to enhance our search for genes related to relevant pathways in cancer, we explored different panels of genes (cell cycle and proliferation, migration and invasion, immunoregulation and inflammation, mucin biosynthesis and secretion, angiogenesis and DNA-repair; according to GO terms) in the two external RNA-seq datasets, examining their putative relationships with splicing machinery, RNA exosome and NMD components expression. This strategy unveiled a substantial number of significant correlations between spliceosome and splicing factors expression and these cancer-related genes, with some of them specifically validating the correlations found in our discovery cohort (e.g., direct correlation of HNRNPF with CDK4, inverse correlation between NOVA1 and CDK6, and MBNL1 inverse correlation with CDKN2A), while other being completely new (Figs. S17, S18). Although less abundant, relevant correlations were also found between these genes and RNA exosome (Fig. S19) and NMD (Fig. S20) components. To a lesser extent, likely due to the technical limitations of studying this mucinous tumor at the protein level, applying a similar approach to the proteomic data yielded comparable results, where a small number of interesting correlations between the levels of components of the splicing machinery and those of pathways of cell cycle and proliferation, migration and invasion, immunoregulation and inflammation, and mucin biosynthesis and secretion were identified, at the protein level (Fig. S21). These results suggest a potential link between RNA biology regulatory mechanisms and these cancer-related pathways’ regulation in PMP, although further studies are needed to elucidate the underlying causes of these associations.

A Significantly altered cancer-related genes in PMP. CDKN2A, CDKN2D, CDK4, CDK6, ATM, CXCL3, CCL5, CXCR2 and KDR expression are represented as Log2 of normalized copy number in the control (white violin, n = 11) and the PMP (colored violin, n = 18) tissues. T-test or Mann–Whitney U test were used and asterisks represent significant differences: *p < 0.05; **p < 0.01; *** p < 0.001. B Significant correlations found between these cancer-related altered genes and splicing-related dysregulated genes. Significant correlations are shown together with Pearson or Spearman correlation coefficient (r) and p value in each case.
Splicing inhibition reduces functional parameters of tumor aggressiveness in PMP in vitro
We sought to ascertain if the associations observed in human samples correspond to a functional role of the splicing process in PMP aggressiveness. To this end, we first evaluated cell proliferation, a key cancer feature clearly associated with RNA biology-related genes in our PMP cohort, by measuring cell viability in a suitable PMP cell model, N14A cells, in response to the splicing inhibitor Pladienolide-B. Of the three doses tested, 10-10 M, 10-9 M, 10-8 M, a strong time-dependent reduction in cell viability was observed with Pladienolide-B at 10-9 M and 10-8 M ( > 40% at 24 h, > 80% at 72 h), but not at 10-10 M; accordingly, a submaximal 10-9 M dose was selected for the rest of the experiments (Fig. 5A). After this clear reduction in cell viability due to the splicing process inhibition, we sought to determine whether this approach could benefit the effect of the current therapeutic strategies for PMP, chemotherapeutics such as mitomycin-C and cisplatin. Thus, we tested the impact of the combination of these drugs with Pladienolide-B over cell viability in N14A cells. Pladienolide-B, mitomycin-C and cisplatin significantly decreased cell viability after 24 to 72 h acting by their own, however, the combination of Pladienolide-B with each one reached a higher cell viability reduction at every time checked comparing with Pladienolide-B alone, at 48 h respect to cisplatin alone and at 72 h in comparison with mitomycin-C alone (Fig. 5B).

A Cell viability assay using Pladienolide-B at 10-10 M, 10-9 M, and 10-8 M. The graph shows cell viability percentage respect to the control of N14A in crescent doses of Pladienolide-B at the different times of the experiment (24, 48 and 72 h). The dotted line at Y = 100 represents the control (vehicle). B Cell viability assay using Pladienolide-B (10-9 M) in combination with mitomycin-C (4 µg/mL) and cisplatin (0.5 × 10-3 M). The graph shows the percentage of cell viability relative to the control (vehicle, dotted line at Y = 100) in N14A cells under different treatments: Pladienolide-B, mitomycin-C, cisplatin, Pladienolide-B + mitomycin-C, and Pladienolide-B + cisplatin at 24, 48, and 72 h. C Cell migration using Pladienolide-B at 10-9 M in the wound-healing assay. The left panel represents the quantification of the area of the scratch in the cells treated respect to the control in percentage in N14A cell line after 16 h. The dotted line at Y = 100 represents control. In the right panel are the pictures of the scratch at 16 h for the control (up) and the treatment (down). D Turbidity assay using Pladienolide-B at 10-9 M. OD at 900 nm is represented for the control (white box) and treated (colored box) cells. E RNA expression level of mucin 2 splicing variants using Pladienolide-B at 10-9 M. The RNA expression levels of all MUC2 isoforms, MUC2-204, MUC2-205 and MUC2-206 are represented in the control and treated cells. Data are represented as mean ± SEM, n = 3 in viability, migration and qPCR assays and n = 6 in turbidity assay. For cell viability, mean differences were assessed using ANOVA, with post hoc Tukey’s test to compare between group pairs. In the rest of the cases, t-test or Mann–Whitney U test were used. Asterisks represent significant differences compared to the control: *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001; symbol plus represents significant differences between the different treatments: Pladienolide-B vs Pladienolide-B + mitomycin-C; Pladienolide-B vs Pladienolide-B + cisplatin; mitomycin-C vs Pladienolide-B + mitomycin-C; cisplatin vs Pladienolide-B + cisplatin; +p < 0.05; ++p < 0.01; ++++p < 0.0001. F MUC-205 and MUC-206 splice variants relative expression in N14A. PSI (percent spliced-in) is represented for MUC-205 and MUC-206 splice variants in the control and after treatment (Pladienolide-B at 10-9 M). The structure of each mucin 2 variant is represented in the right scheme. Pd: Pladienolide-B.
Unlike most tumors, PMP cells rarely invade surrounding tissues or form conventional metastases. Instead, they migrate through the mucinous matrix within the peritoneal cavity, leading to the formation of new tumor foci [27]. To investigate this unique migratory behavior, we used a wound-healing assay to assess the effect of Pladienolide-B on PMP cell migration. We detected a slight reduction of cell migration in the presence of Pladienolide-B in N14A cells after 16 h, although no significant effects were observed (Fig. 5C). Then, to test whether Pladienolide-B influences the mucus secretion, the cell culture medium was collected, and OD was measured. This showed that Pladienolide-B significantly reduces the mucus secretion of N14A cells into the medium (Fig. 5D). This intriguing observation prompted us to investigate possible mechanisms mediating the reduction in mucus production by Pladienolide-B. Mucin 2 is the predominant mucin secreted by PMP cells, as reported in > 95% of PMP cases, followed by mucins 5AC, 5B and 1 [28]. By blocking the splicing process in the cell, Pladienolide-B may be interfering mucin variants production and, consequently, secretion by N14A cells. Thus, we evaluated the splicing variants of the main mucin in PMP, MUC2, in response to treatment (Fig. 5E). MUC2-206, the canonical variant, is the longest and was the main one in vehicle-treated cells; MUC2-204 is the shortest one, spans exons 34 and 35 of the long variant, and may not be considered a true splicing variant, as it may be generated through alternative promoter use; and MUC2-205 is a splicing variant that cuts exon 30 into 3, changing the reading frame. Of note, we observed that Pladienolide-B treatment drastically reduced the expression of MUC2-206 in N14A cells. Interestingly, expression of MUC2-205, although low, remained stable, thus suggesting a change in the relative proportion of the mucin 2 splicing variants by an alteration of the splicing process (Fig. 5F), which may also explain the observed reduction of OD of the medium.
After observing the functional effect of broad splicing inhibition on the PMP cell model, we sought to take it a step further by focusing on some of the splicing factors that were revealed to be dysregulated in our PMP patient cohort. Specifically, we selected two downregulated splicing factors HNRNPK and MBNL1 and one upregulated, PTBP1, as they were on the top of the VIP-Score analysis and its dysregulation was validated in at least one other external cohort. To study the potential role of these splicing factors in the pathology we modulated their expression to reverse the dysregulation found in our PMP samples, overexpressing HNRNPK and MBNL1 and silencing PTBP1, validated by qPCR (Fig. 6A). Then, we studied the effect of this modulation over one of the principal cancer characteristics, cell viability, which was clearly reduced at 48 h after HNRNPK overexpression, at 48 and 72 h after MBNL1 overexpression and at 24 and 72 h after PTBP1 silencing (Fig. 6B). Results from this approach suggested that the expression of these splicing factors may have an implication in cell proliferation, as shown by measuring by qPCR relevant cell cycle and proliferation genes, together with other genes involved in cancer-related pathways that correlated with the splicing machinery RNA expression in the PMP cohorts (Fig. 4B, Figs. S17, S18, S21). Specifically, HNRNPK and MBNL1 overexpression, as well as PTBP1 silencing, similarly reduced cyclins levels, which nicely fits with the decrease observed in cell viability. This finding additionally strengthens certain correlations observed in the human PMP cohorts: CDK4 correlated positively with PTBP1, and its knockdown reduced this cyclin level; CDK6 was inversely correlated to HNRNPK and MBNL1 expression and the overexpression of the two splicing factors resulted in a CDK6 downregulation. HNRNPK overexpression also reduced CDKN2A level, whereas the modulation of the other splicing factors did not affect expression of this gene. CDKN2D expression also decreased significantly with PTBP1 silencing and HNRNPK overexpression. Moreover, AKT1, ATM and MKI67 levels tended to decrease after modulation of the studied genes, matching with the decline in cell viability. Expression of the chemokine CCL5 increased after each modulation (although it was only significant with PTBP1 silencing), which could be related to a more inflammatory and immune microenvironment in the tumor. In contrast, expression of the VEGF receptor gene, KDR, was not altered following splicing factors modulation. Similarly, the cytokine CXCL3 and its receptor CXCR2 showed extremely low expression levels in the cells studied, with no detectable changes between control and splicing factors modulation (Fig. 6C). Having observed these changes at the RNA level, we aimed to assess whether these alterations extended to the protein level. However, obtaining sufficient protein for reliable Western blot analysis from PMP cells proved to be challenging. This model presents several limitations due to its sensitivity and complexity, as well as the scarcity of comparable studies in bibliography. Despite these challenges, we successfully measured two of the modulated splicing factors (MBNL1 and PTBP1) and evaluated the subsequent activation of two key proliferation pathways: AKT and ERK. We observed a reduction in ERK phosphorylation after transfection, with a significantly lower pERK/ERK ratio following PTBP1 silencing. In this condition, AKT phosphorylation was also reduced (Fig. S22). Nevertheless, activation of these pathways after HNRNPK modulation could not be confirmed at protein levels, as Western blot did not allow to validate its overexpression, likely due to limitations related to the antibody or the technique itself. (see raw Western blot images). The interesting changes observed in this relevant set of selected genes prompted us to use the RNA-seq and proteomic data to further explore HNRNPK, MBNL1 and PTBP1 role in the pathology. To this end, samples from PMP patients were divided into two groups, with high or low expression of HNRNPK, MBNL1 or PTBP1, and their expression profiles were explored by an enrichment analysis for the most represented pathways in each group. PMP samples with lower expression of HNRNPK and MBNL1 displayed relevant cancer-related pathways enriched, as angiogenesis, apoptosis, Epithelial–Mesenchymal Transition (EMT), inflammatory and immune responses, MTOR, MYC, NOTCH, TGF-β or WNT signaling, among others, as evidenced by the results from the FFPE RNA-seq, where HNRNPK and MBNL1 were significantly downregulated. In the case of PTBP1, patient samples with higher levels were enriched in MTORC1 and NOTCH signaling, both related to cell growth and proliferation. On the other hand, lower PTBP1 expression was linked to an enrichment in inflammatory and immune pathways, as highlighted by proteomic data where PTBP1 is significantly upregulated (Fig. S23). Finally, to determine whether the modulation of these splicing factors could also affect mucin 2 splicing pattern, as observed with Pladienolide-B-mediated splicing inhibition, we measured MUC2 variants. HNRNPK and MBNL1 overexpression showed similar effects, leading to a reduced expression of all MUC2 variants compared to the control, although significant differences were only observed for MUC2-205 upon HNRNPK overexpression. In contrast, PTBP1 silencing resulted in a distinct splicing pattern. MUC2-204 and MUC2-206 variants were downregulated upon PTBP1 knockdown, with a significant reduction in MUC2-206 expression. Interestingly, MUC2-205 showed a tendency toward increased expression under this silencing (Figs. 6D, E). These findings suggest that specific splicing factor modulation differentially impacts MUC2 isoform expression, reinforcing the notion that alternative splicing regulation plays a critical role in PMP pathophysiology.

A Validation of each modulation at the RNA level by qPCR. The graphs show the expression levels of each splicing factor relative to the control (CMV6 plasmid for overexpression; scramble for silencing) 24 h post-transfection. Expression was normalized using GAPDH as the housekeeping gene. The dotted line at Y = 100 represents the control. B Cell viability assay after each transfection. Cell viability rate is shown as percentage of the control (dotted line at Y = 100) at the different times (24, 48, 72 h). C Cancer-related genes altered after modulation of HNRNPK, MBNL1, and PTBP1, measured by qPCR. Gene expression is presented as Log2 of the Fold-Change modulation/control. Upregulated genes are shown in red, downregulated genes in blue. D RNA expression level of mucin 2 splicing variants after HNRNPK, MBNL1 and PTBP1 modulation. Expression levels of MUC2 isoforms (MUC2-204, MUC2-205, and MUC2-206) are shown in the control transfection (CMV6 plasmid for HNRNPK and MBNL1, or scramble for PTBP1), normalized to GAPDH expression. Data are presented as mean ± SEM, n = 3. For cell viability, differences were assessed using ANOVA with post hoc Tukey’s test for pairwise comparisons. For other cases, t-test or Mann–Whitney U test were used. Asterisks represent significant differences compared to the control: *p < 0.05; **p < 0.01; *** p < 0.001; ****p < 0.0001. E Relative expression of MUC2-205 and MUC2-206 splice variants in N14A after modulation of HNRNPK, MBNL1, and PTBP1. PSI (percent spliced-in) for MUC2-205 and MUC2-206 splice variants are shown for control and different modulations.
Discussion
PMP is a poorly studied disease, due both to its rarity, which makes sample collection difficult, and to the intrinsic complexity of these samples, mostly composed by mucus with low cellularity, which complicates its study at the molecular level [29]. Only few studies have investigated PMP at transcriptional level, most of them comparing PMP gene expression profiles with those of related tumors, like appendiceal mucinous neoplasms and/or colorectal cancer, which revealed interesting pathways altered at the transcriptomic level in PMP, as TGF-β signaling, EMT, extracellular matrix and angiogenesis [30]. In this context, our study, which represents the first approach to precisely explore central regulatory systems of RNA biology in PMP, reveals a severe dysregulation of the splicing machinery, RNA exosome complex and NMD in this tumor, which may entail pathophysiological implications. Specifically, our study illustrates a general alteration in the expression of the components of these machineries that is strongly associated with prognostic parameters, as well as demonstrates that inhibiting the splicing process in vitro, by using a generic splicing inhibitor and modulating the expression of specific splicing factors, significantly reduces key functional parameters of aggressiveness in PMP cells.
Our results are consistent with recent evidence that dysregulation in RNA regulatory processes is a widespread feature in cancer [31]. Changes in the RNA expression of genes belonging to splicing machinery have been observed in pancreatic adenocarcinoma [24], pancreatic neuroendocrine tumors [32, 33] and lung carcinoids [34], among others, demonstrating that general dysregulation of these genes may be related to tumor aggressiveness. Even though RNA exosome complex and NMD are less studied in cancer, similar results have been found studying RNA exosome genes in prostate cancer [35], among others; and in NMD components in lung adenocarcinoma, hepatocellular carcinoma, prostate cancer, breast cancer or colorectal cancer [36, 37]. Here, we demonstrate for the first time that the effectors of these processes are dysregulated and associated with a poor prognosis in PMP, which could aid in understanding the molecular functioning of this rare, understudied disease, opening new research pathways. Among this general dysregulation, it was unexpected to find almost all genes downregulated in our discovery cohort, as only two of the splicing factors studied, PTBP1 and RAVER1, were upregulated. PTBP1 was also found upregulated in HG PMP at the protein level. This factor is a regulatory splicing repressor whose upregulation is well described in various cancers, where it plays an oncogenic role, and has been proposed as a potential therapeutic target, with promising results in glioblastoma [38], glioma [39],hepatocellular carcinoma [40], bladder [41], and pancreatic ductal adenocarcinoma [42]. RAVER1 acts as a PTB corepressor for splicing, known to interact with PTBP1 [43]; however, its implication in cancer has only been reported recently, linked to bad prognosis and with an oncogenic role in lung adenocarcinoma [44]. PTBP1 and RAVER1 overexpression in PMP could indicate these two factors are acting together repressing the splicing process in PMP. Only one other gene was upregulated in our discovery cohort, SMG9, that encodes a key protein of the NMD complex [12]. SMG9 upregulation has also been studied in recent investigations in glioma [45] and hepatocellular carcinoma [46]. Nevertheless, the genes more associated to PMP progression were downregulated in PMP tissues in our discovery cohort, particularly HNRNPA3, HNRNPK, MBNL1 and SRSF3, which were linked to lower OS. High HNRNPA3 expression is linked to poor prognosis in bladder cancer [47]; HNRNPK downregulation was also confirmed in the data from the two external RNA-seq. This splicing factor acts as a repressor of the process. This gene is downregulated in most tumors and, as in our study, its lower expression linked to worse prognosis, although, in some cancers can be upregulated and play an oncogenic role [13, 48]; MBNL1 downregulation was validated in the FFPE external cohort and is a well-known tumor suppressor gene, downregulated in breast cancer [49] and glioblastoma [50]. This splicing factor can act promoting or repressing the splicing process, changes in its expression are reported to affects the splicing pattern during EMT in pancreatic adenocarcinoma [51]; and SRSF3 is also downregulated in hepatocellular carcinoma [52], where its low expression is linked to bad prognosis, whereas it is upregulated and plays oncogenic roles in glioblastoma [53] among other cancers. Exosome and NMD genes associated to clinical parameters are less studied in cancer, and no relevant coincidences were found with our results.
Interestingly, our study unveiled that key genes related to cell cycle and proliferation, immunomodulation and inflammation, angiogenesis, migration, DNA-repair and mucin secretion are altered in PMP, as had been previously reported in other tumors but not in this rare neoplasia [54, 55]. Particularly, CDKN2A and CDKN2D are critical checkpoints, preventing cell cycle progression from G1 to S phase; CDK4 and CDK6 drive the transition from G1 to S; while ATM is a crucial regulator at the G1/S and G2/M checkpoints, blocking the cycle in response to DNA damage [56]. The expression profile of these genes is meaningful, as the low expression of the suppressor CDKN2D, along with the high expression of the cyclins CDK4 and CDK6 suggests that the tumor is highly proliferative, as these kinases drive cells through a critical checkpoint in the cell cycle. However, the combination of high CDKN2A and low CDKN2D could indicate a complex regulatory environment within the tumor. While the tumor experiences signals or stresses that lead to CDKN2A upregulation, possibly as an attempt to control cell proliferation, simultaneously low expression of CDKN2D could undermine this control, allowing PMP cells to bypass this checkpoint and continue to proliferate. On the other hand, high expression of ATM may indicate that the cells are experiencing significant DNA damage or replication stress [56]. In this context, high ATM expression might be a response to the high proliferative activity driven by CDK4 and CDK6, which can increase replication stress and the accumulation of DNA damage. This could lead to a scenario where PMP cells are replicating but also accumulating genetic damage, potentially leading to a more aggressive and therapy-resistant phenotype. The interplay between these pathways highlights the complexity of PMP, which warrants further investigations in the future to fully elucidate the underlying mechanisms involved. Beyond cell cycle and DNA damage response alterations, our analysis also revealed dysregulation of key genes involved in immunomodulation and inflammation, further supporting the complexity of PMP. Specifically, we found CXCL3 and CCL5 upregulated and CXCR2 downregulated in PMP samples. CXCL3, a pro-inflammatory chemokine, has been implicated in tumor progression, angiogenesis, immune evasion, and metastasis in various cancers. CXCL3 exerts its effects through CXCR2, triggering the PI3K/AKT and MAPK/ERK pathways, and promoting an immunosuppressive microenvironment by recruiting neutrophils. The CXCL3-CXCR2 axis has been widely studied in cancer [57, 58]. On the other hand, CCL5 is a chemokine that causes inflammation and functions as a chemoattractant for monocytes, memory T helper cells, and eosinophils, fostering an inflammatory tumor microenvironment [59]. Inflammation and angiogenesis are tightly linked processes in the tumor microenvironment, both contributing to PMP progression [30]. KDR encodes the VEGF tyrosine kinase receptor, acting as the main mediator of VEGF-induced endothelial proliferation, survival, and migration, playing an essential role in the regulation of angiogenesis, a process widely reported in different tumor types, including colorectal cancer [60]. Of note, we discovered that the expression of these genes was tightly linked to splicing, RNA exosome and NMD-related altered genes in PMP, uncovering an interesting, previously unrecognized relationship between altered splicing and these cancer-related genes in PMP, which could help to understand the changes observed by inhibiting the splicing process in vitro.
PMP is characterized by a slow but persistent growth that, coupled to the high motility of its cells, which facilitates their distribution through the peritoneal cavity, results in enhanced tumor progression [1, 27, 29]. We assessed the inhibition of the splicing process using Pladienolide-B, a small molecule that specifically targets SF3B1, a core component of the spliceosome [21], which we have successfully employed to this end in previous studies [24]. This drug enables to block the splicing process in a controlled manner to assess the effect of reducing this process activity on different functional parameters of tumor behavior using N14A cells as a PMP in vitro model [20]. Results revealed that inhibiting the splicing process clearly reduced functional aggressiveness characteristics, which is in complete concordance with numerous studies in different cancers [24]. Moreover, we demonstrated that the modulation of specific splicing factors altered in PMP, HNRNPK, MBNL1 and PTBP1, lead to the alteration of genes related to cell cycle and proliferation, among others, and reduced cell viability in vitro. Furthermore, we observed that combining Pladienolide-B with chemotherapeutic agents commonly used in PMP treatment enhanced their efficacy, suggesting that splicing inhibition could potentiate the therapeutic effects of these drugs. These findings highlight the potential of targeting the spliceosome as a complementary strategy to current PMP therapies. This approach is increasingly being explored in the context of various cancers, with several clinical trials currently underway to evaluate its impact on standard cancer therapies [61, 62]. Importantly, besides growth and dissemination, PMP progression is determined by the excessive secretion by tumor cells of mucin, which fills the peritoneal cavity with mucus and allows tumor cells to spread throughout the peritoneum, reducing the effect of chemotherapy [1, 29]. The role of secretion of different types of mucins has been amply studied in diverse cancer types. Mucin glycosylation profile of cancer cells is specifically changed in comparison with healthy ones, related to poor clinical outcomes and cancer progression [63]. Different drugs have been used to target these mucins in cancer, being those that degrade mucin the most promising [64]. Our cell model, N14A, is reported to secrete mucin, which accumulates in the flask and forms a mucous layer [20], and can be measured by assessing the turbidity of the media after decanting the cells. Moreover, splicing variants of mucin genes have also been studied and related to cancer progression [65]. In line with this, our present results revealed that inhibiting splicing with Pladienolide-B reduced the turbidity of the media and also altered the pattern of MUC2 splicing variants. In line with this, our results revealed that splicing inhibition using Pladienolide-B reduced the turbidity of the media and also altered the MUC2 splicing pattern. HNRNPK and MBNL1 overexpression led to a reduction in all MUC2 variants, while PTBP1 silencing resulted in a shift in the MUC2 splicing variants, with MUC2-205 tending to be more abundant in silenced cells than MUC2-206, the canonical, that is more expressed in the control. This, coupled to the observed reduction in cell viability, confirmed that hampering the splicing process could represent a new therapeutic opportunity in PMP.
Taken together, the results of this study demonstrate the dysregulation of different RNA biology machineries in PMP, linked to poor prognosis, tumor aggressiveness and cancer-related pathways as cell cycle and proliferation or immunomodulation and inflammation. The inhibition of splicing using Pladienolide-B and the modulation of specific splicing factors (HNRNPK, MBNL1 and PTBP1), reduced cell viability in vitro, altered mucin secretion and MUC2 splicing pattern. Furthermore, combining splicing inhibition with conventional chemotherapies enhanced their therapeutic effects, offering a potential new treatment strategy for PMP. These original findings document for the first time the importance of splicing, RNA exosome and NMD alterations in this rare, understudied tumor, inviting further exploration of these targetable processes to improve and personalize the treatment of PMP.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Patrick-Brown TDJH, Carr NJ, Swanson DM, Larsen S, Mohamed F, Flatmark K. Estimating the Prevalence of Pseudomyxoma Peritonei in Europe Using a Novel Statistical Method. Ann Surg Oncol. 2021;28:252–7.
Govaerts K, Lurvink RJ, De Hingh IHJT, Van der Speeten K, Villeneuve L, Kusamura S, et al. Appendiceal tumours and pseudomyxoma peritonei: Literature review with PSOGI/EURACAN clinical practice guidelines for diagnosis and treatment. Eur J Surg Oncol. 2021;47:11–35.
Kusamura S, Barretta F, Yonemura Y, Sugarbaker PH, Moran BJ, Levine EA, et al. The Role of Hyperthermic Intraperitoneal Chemotherapy in Pseudomyxoma Peritonei After Cytoreductive Surgery. JAMA Surg. 2021;156:e206363.
Baratti D, Kusamura S, Nonaka D, Cabras AD, Laterza B, Deraco M. Pseudomyxoma peritonei: biological features are the dominant prognostic determinants after complete cytoreduction and hyperthermic intraperitoneal chemotherapy. Ann Surg. 2009;249:243–9.
Rufián-Andujar B, Valenzuela-Molina F, Rufián-Peña S, Casado-Adam Á, Sánchez-Hidalgo JM, Rodríguez-Ortiz L, et al. From the Ronnett to the PSOGI Classification System for Pseudomyxoma Peritonei: A Validation Study. Ann Surg Oncol. 2021;28:2819–27.
Murage NW, Ahmed NM, Underwood TJ, Walters ZS, Breininger SP. The genetic profile and molecular subtypes of human pseudomyxoma peritonei and appendiceal mucinous neoplasms: a systematic review. Cancer Metast Rev. 2023;42:335–59.
Arjona-Sanchez A, Martinez-López A, Moreno-Montilla MT, Mulsow J, Lozano-Lominchar P, Martínez-Torres B, et al. External multicentre validation of pseudomyxoma peritonei PSOGI-Ki67 classification. Eur J Surg Oncol J Eur Soc Surg Oncol Br Assoc Surg Oncol. 2023;49:1481–8.
Blázquez-Encinas R, Moreno-Montilla MT, García-Vioque V, Gracia-Navarro F, Alors-Pérez E, Pedraza-Arevalo S, et al. The uprise of RNA biology in neuroendocrine neoplasms: altered splicing and RNA species unveil translational opportunities. Rev Endocr Metab Disord. 2023;24:267–82.
Marasco LE, Kornblihtt AR. The physiology of alternative splicing. Nat Rev Mol Cell Biol. 2023;24:242–54.
Makino DL, Halbach F, Conti E. The RNA exosome and proteasome: common principles of degradation control. Nat Rev Mol Cell Biol. 2013;14:654–60.
Hentze MW, Kulozik AE. A perfect message: RNA surveillance and nonsense-mediated decay. Cell. 1999;96:307–10.
Kurosaki T, Popp MW, Maquat LE. Quality and quantity control of gene expression by nonsense-mediated mRNA decay. Nat Rev Mol Cell Biol. 2019;20:406–20.
Bradley RK, Anczuków O. RNA splicing dysregulation and the hallmarks of cancer. Nat Rev Cancer. 2023;23:135–55.
Tao Y, Zhang Q, Wang H, Yang X, Mu H. Alternative splicing and related RNA binding proteins in human health and disease. Signal Transduct Target Ther. 2024;9:26.
Anczukow O, Allain FH-T, Angarola BL, Black DL, Brooks AN, Cheng C, et al. Steering research on mRNA splicing in cancer towards clinical translation. Nat Rev Cancer. 2024;24:887–905.
Rogalska ME, Mancini E, Bonnal S, Gohr A, Dunyak BM, Arecco N, et al. Transcriptome-wide splicing network reveals specialized regulatory functions of the core spliceosome. Science. 2024;386:551–60.
Karam R, Wengrod J, Gardner LB, Wilkinson MF. Regulation of nonsense-mediated mRNA decay: implications for physiology and disease. Biochim Biophys Acta. 2013;1829:624–33.
Chlebowski A, Lubas M, Jensen TH, Dziembowski A. RNA decay machines: the exosome. Biochim Biophys Acta. 2013;1829:552–60.
Romero-Ruiz A, Granados-Rodríguez M, Bura FI, Valenzuela-Molina F, Rufián-Andújar B, Martínez-López A, et al. Breaking the Mucin Barrier: A New Affinity Chromatography-Mass Spectrometry Approach to Unveil Potential Cell Markers and Pathways Altered in Pseudomyxoma Peritonei. Biol Proced Online. 2024;26:13.
Roberts DL, O’Dwyer ST, Stern PL, Renehan AG. Global gene expression in pseudomyxoma peritonei, with parallel development of two immortalized cell lines. Oncotarget. 2015;6:10786–800.
Kotake Y, Sagane K, Owa T, Mimori-Kiyosue Y, Shimizu H, Uesugi M, et al. Splicing factor SF3b as a target of the antitumor natural product pladienolide. Nat Chem Biol. 2007;3:570–5.
Rossi A, Biancalana L, Vančo J, Malina T, Zacchini S, Dvořák Z, et al. The effect of a varying pyridine ligand on the anticancer activity of Diiron(I) bis-cyclopentadienyl complexes. Chem Biol Interact. 2025;406:111318.
Martínez-Quintanilla J, Cabot D, Sabia D, Arqués O, for the PMPnet Group, Vergés J, et al. Precision Oncology and Systemic Targeted Therapy in Pseudomyxoma Peritonei. Clin Cancer Res. 2024;30:4082–99.
Alors-Perez E, Blázquez-Encinas R, Alcalá S, Viyuela-García C, Pedraza-Arevalo S, Herrero-Aguayo V, et al. Dysregulated splicing factor SF3B1 unveils a dual therapeutic vulnerability to target pancreatic cancer cells and cancer stem cells with an anti-splicing drug. J Exp Clin Cancer Res. 2021;40:382.
Vandesompele J, Preter KD, Roy NV, Paepe AD. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genom Biol. 2002;3:90.
Ding Z, Meng Y-R, Fan Y-J, Xu Y-Z. Roles of minor spliceosome in intron recognition and the convergence with the better understood major spliceosome. Wiley Interdiscip Rev RNA. 2023;14:e1761.
Lazzari N, Rigotto G, Montini B, Del Bianco P, Moretto E, Palladino F, et al. Stemness and hybrid epithelial-mesenchymal profiles guide peritoneal dissemination of malignant mesothelioma and pseudomyxoma peritonei. Int J Cancer. 2025;156:201–15.
Amini A, Masoumi-Moghaddam S, Ehteda A, Morris DL. Secreted mucins in pseudomyxoma peritonei: pathophysiological significance and potential therapeutic prospects. Orphanet J Rare Dis. 2014;9:71.
Mittal R, Chandramohan A, Moran B. Pseudomyxoma peritonei: natural history and treatment. Int J Hyperth. 2017;33:511–9.
Pretzsch E, Neumann J, Nieß H, Pretzsch CM, Hofmann FO, Kirchner T, et al. Comparative transcriptomic analyses reveal activation of the epithelial-mesenchymal transition program in non-metastasizing low grade pseudomyxoma peritonei. Pathol Res Pr. 2024;254:155129.
Bonnal SC, López-Oreja I, Valcárcel J. Roles and mechanisms of alternative splicing in cancer - implications for care. Nat Rev Clin Oncol. 2020;17:457–74.
Pedraza-Arevalo S, Alors-Pérez E, Blázquez-Encinas R, Herrera-Martínez AD, Jiménez-Vacas JM, Fuentes-Fayos AC, et al. Spliceosomic dysregulation unveils NOVA1 as a candidate actionable therapeutic target in pancreatic neuroendocrine tumors. Transl Res J Lab Clin Med. 2023;251:63–73.
Alors-Pérez E, Pedraza-Arevalo S, Blázquez-Encinas R, García-Vioque V, Agraz-Doblas A, Yubero-Serrano EM, et al. Altered CELF4 splicing factor enhances pancreatic neuroendocrine tumors aggressiveness influencing mTOR and everolimus response. Mol Ther Nucleic Acids. 2024;35:102090.
Blázquez-Encinas R, García-Vioque V, Caro-Cuenca T, Moreno-Montilla MT, Mangili F, Alors-Pérez E, et al. Altered splicing machinery in lung carcinoids unveils NOVA1, PRPF8 and SRSF10 as novel candidates to understand tumor biology and expand biomarker discovery. J Transl Med. 2023;21:879.
Sáez-Martínez P, Porcel-Pastrana F, Montero-Hidalgo AJ, Lozano de la Haba S, Sanchez-Sanchez R, González-Serrano T, et al. Dysregulation of RNA-Exosome machinery is directly linked to major cancer hallmarks in prostate cancer: Oncogenic role of PABPN1. Cancer Lett. 2024;584:216604.
Nogueira G, Fernandes R, García-Moreno JF, Romão L. Nonsense-mediated RNA decay and its bipolar function in cancer. Mol Cancer. 2021;20:72.
Lindeboom RGH, Vermeulen M, Lehner B, Supek F. The impact of nonsense-mediated mRNA decay on genetic disease, gene editing and cancer immunotherapy. Nat Genet. 2019;51:1645–51.
Wang K, Pan S, Zhao P, Liu L, Chen Z, Bao H, et al. PTBP1 knockdown promotes neural differentiation of glioblastoma cells through UNC5B receptor. Theranostics. 2022;12:3847–61.
Song X, Tiek D, Miki S, Huang T, Lu M, Goenka A, et al. RNA splicing analysis deciphers developmental hierarchies and reveals therapeutic targets in adult glioma. J Clin Invest. 134:e173789.
Shen L, Lei S, Zhang B, Li S, Huang L, Czachor A, et al. Skipping of exon 10 in Axl pre-mRNA regulated by PTBP1 mediates invasion and metastasis process of liver cancer cells. Theranostics. 2020;10:5719–35.
Bielli P, Panzeri V, Lattanzio R, Mutascio S, Pieraccioli M, Volpe E, et al. The Splicing Factor PTBP1 Promotes Expression of Oncogenic Splice Variants and Predicts Poor Prognosis in Patients with Non-muscle-Invasive Bladder Cancer. Clin Cancer Res J Am Assoc Cancer Res. 2018;24:5422–32.
Calabretta S, Bielli P, Passacantilli I, Pilozzi E, Fendrich V, Capurso G, et al. Modulation of PKM alternative splicing by PTBP1 promotes gemcitabine resistance in pancreatic cancer cells. Oncogene. 2016;35:2031–9.
Rideau AP, Gooding C, Simpson PJ, Monie TP, Lorenz M, Hüttelmaier S, et al. A peptide motif in Raver1 mediates splicing repression by interaction with the PTB RRM2 domain. Nat Struct Mol Biol. 2006;13:839–48.
Wedler A, Bley N, Glaß M, Müller S, Rausch A, Lederer M, et al. RAVER1 hinders lethal EMT and modulates miR/RISC activity by the control of alternative splicing. Nucleic Acids Res. 2024;52:3971–88.
Dai Y, Zhang H, Feng S, Guo C, Tian W, Sun Y, et al. SMG9 is a novel prognostic-related biomarker in glioma correlating with ferroptosis and immune infiltrates. Heliyon. 2024;10:e25716.
Jin X, Yin J, Zhu H, Li W, Yu K, Liu M, et al. SMG9 Serves as an Oncogene to Promote the Tumor Progression via EMT and Wnt/β-Catenin Signaling Pathway in Hepatocellular Carcinoma. Front Pharm. 2021;12:701454.
Amano N, Matsumoto K, Shimizu Y, Nakamura M, Tsumura H, Ishii D, et al. High HNRNPA3 expression is associated with lymph node metastasis and poor prognosis in patients treated with radical cystectomy. Urol Oncol. 2021;39:196.e1–196.e7.
Escobar-Hoyos LF, Penson A, Kannan R, Cho H, Pan C-H, Singh RK, et al. Altered RNA Splicing by Mutant p53 Activates Oncogenic RAS Signaling in Pancreatic Cancer. Cancer Cell. 2020;38:198–211.e8.
Fish L, Pencheva N, Goodarzi H, Tran H, Yoshida M, Tavazoie SF. Muscleblind-like 1 suppresses breast cancer metastatic colonization and stabilizes metastasis suppressor transcripts. Genes Dev. 2016;30:386–98.
Voss DM, Sloan A, Spina R, Ames HM, Bar EE. The Alternative Splicing Factor, MBNL1, Inhibits Glioblastoma Tumor Initiation and Progression by Reducing Hypoxia-Induced Stemness. Cancer Res. 2020;80:4681–92.
Xiao L, Sun H, Cheng R, Yang R, Jin X, Xu Z, et al. Functional requirement of alternative splicing in epithelial-mesenchymal transition of pancreatic circulating tumor. Mol Ther Nucleic Acids. 2024;35:102129.
Sen S, Langiewicz M, Jumaa H, Webster NJG. Deletion of serine/arginine-rich splicing factor 3 in hepatocytes predisposes to hepatocellular carcinoma in mice. Hepatol Balt Md. 2015;61:171–83.
Fuentes-Fayos AC, Vázquez-Borrego MC, Jiménez-Vacas JM, Bejarano L, Pedraza-Arévalo S, L-López F, et al. Splicing machinery dysregulation drives glioblastoma development/aggressiveness: oncogenic role of SRSF3. Brain J Neurol. 2020;143:3273–93.
Spring LM, Wander SA, Andre F, Moy B, Turner NC, Bardia A. Cyclin-dependent kinase 4 and 6 inhibitors for hormone receptor-positive breast cancer: past, present, and future. Lancet Lond Engl. 2020;395:817–27.
Shiloh Y. ATM and related protein kinases: safeguarding genome integrity. Nat Rev Cancer. 2003;3:155–68.
Ameratunga M, Kipps E, Okines AFC, Lopez JS. To Cycle or Fight-CDK4/6 Inhibitors at the Crossroads of Anticancer Immunity. Clin Cancer Res J Am Assoc Cancer Res. 2019;25:21–8.
Sun X, He X, Zhang Y, Hosaka K, Andersson P, Wu J, et al. Inflammatory cell-derived CXCL3 promotes pancreatic cancer metastasis through a novel myofibroblast-hijacked cancer escape mechanism. Gut. 2022;71:129–47.
Ren H, Yang X, Hou W, Meng J, Luo D, Zhang C. Comprehensive analysis of the clinical and biological significances for chemokine CXCL3 in cholangiocarcinoma. Med (Balt). 2024;103:e37460.
Zeng Z, Lan T, Wei Y, Wei X. CCL5/CCR5 axis in human diseases and related treatments. Genes Dis. 2021;9:12–27.
Mohammad Rezaei F, Hashemzadeh S, Ravanbakhsh Gavgani R, Hosseinpour Feizi M, Pouladi N, Samadi Kafil H, et al. Dysregulated KDR and FLT1 Gene Expression in Colorectal Cancer Patients. Rep. Biochem Mol Biol. 2019;8:244–52.
Ottawa Hospital Research Institute. A Phase II Randomized Window of Opportunity Trial Evaluating Clinical and Biological Effects of PRMT5 Inhibitor, GSK3326595. Early Stage Breast Cancer. 2022. https://clinicaltrials.gov/study/NCT04676516
Janssen Research & Development, LLC. A Phase 1, First-in-Human, Open-Label Study of the Safety, Pharmacokinetics, and Pharmacodynamics of JNJ-64619178, an Inhibitor of Protein Arginine Methyltransferase 5 (PRMT5). Subjects Adv Cancers. 2025 https://clinicaltrials.gov/study/NCT03573310
Hollingsworth MA, Swanson BJ. Mucins in cancer: protection and control of the cell surface. Nat Rev Cancer. 2004;4:45–60.
Beatson R, Burchell JM. A mucin degrader for cancer therapy. Nat Biotechnol. 2024;42:572–3.
Kumar S, Cruz E, Joshi S, Patel A, Jahan R, Batra SK, et al. Genetic variants of mucins: unexplored conundrum. Carcinogenesis. 2017;38:671–9.
Acknowledgements
We are gratefully indebted to all the patients and their families for generously donating the samples and clinical data for research purposes. We would like to express our sincere gratitude to Dr. A. Renehan and Dr. P. Caswell for providing us with N14A cell line. Their generosity and support were crucial for the completion of this study. We also acknowledge the technical help of the personnel of core services of the IMIBIC (UCAIB) and University of Cordoba (SCAI).
Funding
This work was supported by the Plan Estatal de I + D + i 2017-2020 and co-financed by the Instituto de Salud Carlos III Subdirección General de Evaluación y Fomento de la Investigación and the Fondo Europeo de Desarrollo Regional (FEDER) [PI19/01603 and PI22/01213 (to A.A.S.)]. Spanish Ministry of Science and Innovation [MICINN; PID2019‐105201RB‐I00, AEI/10.13039/501100011033 and PID2022-136227OB-I00 (to JPC)]. Instituto de Salud Carlos III, co‐funded by European Union (ERDF/ESF, “Investing in your future”); Predoctoral contract FI17/00282 (to E.A.P.)]. Society for Endocrinology Early Career Grant (to A.I.C.). Spanish Ministry of Universities Predoctoral contracts FPU18/02275 (to R.B.E.) and FPU20/03958 (to V.G.V.); postdoctoral contract under the program María Zambrano funded by the European Union Next Generation-EU (to S.P.A.). Junta de Andalucía (BIO‐0139); FEDER UCO-202099901918904 (to J.P.C. and A.I.C.). Grupo Español de Tumores Neuroendocrinos y Endocrinos (G2016 and G2019 Research grants, to J.P.C.). University of Cordoba, Predoctoral Contract BOUCO2021/00110 (to M.T.M.M.). CIBERobn Fisiopatología de la Obesidad y Nutrición. CIBER is an initiative of Instituto de Salud Carlos III.
Author information
Authors and Affiliations
Contributions
The work reported in the paper has been performed by the authors, unless clearly specified in the text: J.P.C., A.A.S., A.I.C. conceived and designed the project. M.T.M.M., E.A.P., A.M.L., R.B.E., V.G.V., M.G.R., M.C.V.B. performed the experiments. M.T.M.M., S.P.A., E.A.P., A.M.L., R.B.E., V.G.V., M.G.R., R.O.S., M.C.V.B., A.R.R., J.P.C., A.A.S., A.I.C. analyzed data and interpreted the results. M.T.M.M., S.P.A., E.A.P., A.R.R., J.P.C., A.A.S., A.I.C. prepared figures. A.M.L., R.O.S., L.R.O., F.V.M., B.R.A., A.A.S. acquired clinical/pathological data and samples. M.T.M.M., S.P.A., J.P.C., A.A.S., A.I.C. wrote the manuscript. M.T.M.M., S.P.A., A.M.L., R.B.E., V.G.V., L.R.O., F.V.M., B.R.A., M.G.R., R.O.S., E.A.P., M.C.V.B., A.R.R., J.P.C., A.A.S., A.I.C. critically revised the manuscript and approved final version.
Corresponding authors
Ethics declarations
Competing interests
All the authors of the manuscript, have no relevant financial or non-financial interests to disclose, no competing interests to declare that are relevant to the content of this article, no affiliations with or involvement in any organization or entity with any financial interest or non-financial interest in the subject matter or materials discussed in this manuscript and no financial or proprietary interests in any material discussed in this article.
Ethics approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards. The study was approved by the Bioethics Committee of the Medical University of Cordoba/IMIBIC and Ethics Committees from all the Hospitals involved in the study (protocol code PI19/01603, version 1 – 13/03/2019). Informed consent was obtained through the Andalusian Biobank (Servicio Andaluz de Salud), from all individual participants included in the study and all of them are available from the corresponding author on reasonable request.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Moreno-Montilla, M.T., Pedraza-Arevalo, S., Martínez-López, A. et al. Exploring RNA biology in pseudomyxoma peritonei uncovers splicing dysregulation as a novel, targetable molecular vulnerability. Cancer Gene Ther 32, 721–736 (2025). https://doi.org/10.1038/s41417-025-00911-x
Received:
Revised:
Accepted:
Published:
Issue date:
DOI: https://doi.org/10.1038/s41417-025-00911-x
