
Abstract
Proanthocyanidins (PAs) play significant roles in plants and are bioactive compounds with health benefits. The polymerization mechanism has been debated for decades. Here we show that laccases (LACs) are involved in PA polymerization and miR397a is a negative regulator of PA biosynthesis in Salvia miltiorrhiza and Populus trichocarpa. Elevation of miR397a level causes significant downregulation of LACs, severe reduction of polymerized PAs, and significant increase of flavan-3-ol monomers in transgenic S. miltiorrhiza and P. trichocarpa plants. Enzyme activity analysis shows that miR397a-regulated SmLAC1 catalyzes the polymerization of flavan-3-ols and the conversion of B-type PAs to A-type. Both catechin and epicatechin can serve as the starter unit and the extension unit during PA polymerization. Overexpression of SmLAC1 results in significant increase of PA accumulation, accompanied by the decrease of catechin and epicatechin contents. Consistently, CRISPR/Cas9-mediated SmLAC1 knockout shows the opposite results. Based on these results, a scheme for LAC-catalyzed PA polymerization is proposed. The work provides insights into PA polymerization mechanism.
Similar content being viewed by others
Introduction
Proanthocyanidins (PAs, condensed tannins) are the most abundant secondary metabolites and the second most abundant plant phenolic polymers after lignin1,2. They widely exist in seeds, fruits, leaves, flowers, stems and roots to protect plants from biotic and abiotic stresses, control soil nitrogen cycling, and affect seed coat color and seed dormancy, germination, and longevity2,3,4,5. PAs are flavor constituents and astringency of wine, beer, tea and fruit juice, and functional ingredients in various beverages, fruits, vegetables, foods, herbal remedies and dietary supplements2,6,7. They have antioxidant, anti-bacterial, anti-diabetic, anti-viral, anti-inflammatory, and anti-cancer activities, show health beneficial effects on urinary tract, cardiovascular, stomach, brain and bone, and may protect dentin collagen7,8,9,10,11,12,13. In addition, PAs in forages are associated with improved ruminal digestibility and availability of by-pass protein and reduced detrimental effects of internal parasites and risk of bloat in animals14,15.
PAs are oligomers or polymers of flavan-3-ol monomers. The degree of polymerization (DP) ranges from 2 to 30 or more monomers1. Flavan-3-ols can be divided into 2, 3-cis-flavan-3-ols and 2,3-trans-flavan-3-ols. The former ones are represented by dominant (−)-epicatechin and (−)-epigallocatechin, featured by a 2,3-cis-2R, 3R configuration and minor (+)-epicatechin and (+)-epigallocatechin featured by a 2,3-cis-2S, 3S configuration, whereas the latter ones include (+)-catechin and (+)-gallocatechin characterized by a 2,3-trans-2R, 3S configuration and (−)-catechin and (−)-gallocatechin characterized by a 2,3-trans-2S, 3R configuration16. Both 2,3-cis-flavan-3-ols and 2,3-trans-flavan-3-ols are synthesized via the flavonoid pathway. They share the same upstream biosynthetic pathway, from phenylalanine to leucoanthocyanidins (flavan-3,4-diols), but have different downstream pathways2,6,17,18,19,20. Of the four configurations, only 2,3-trans-2R, 3S-flavan-3-ols are produced from leucoanthocyanidins under the catalysis of NADPH-dependent leucoanthocyanidin reductase (LAR)21, whereas three other configurations of flavan-3-ols are synthesized from anthocyanidins under the catalysis of NADPH-dependent anthocyanidin reductase (ANR)22,23. In addition, (+)-catechin was converted to (−)-epicatechin via flav-2-en-3-ol catalyzed by leucoanthocyanidin dioxygenase (LDOX) and ANR24. In wheat and maize that do not accumulate PAs, ANRs predominantly generate (+)-epicatechin19,24.
Although the biosynthesis of flavan-3-ol monomers has been intensively studied, the mechanism of their polymerization is still at a hypothetical stage6,18,25,26,27. There are three hypotheses for the formation of PAs6. In the first hypothesis, flavan-3-ol monomers serve as the starter units to be condensed with carbocations converted from leucoanthocyanidins through nonenzymatic oxidation16,28,29. This hypothesis was partially supported by recent results showing that leucocyanidin could be converted into carbocations in vitro under weakly acidic conditions29. In the second one, anthocyanins, instead of leucoanthocyanidins, are converted to quinone methides probably under the catalysis of polyphenol oxidase (PPO). The resulting quinone methides are then converted to carbocations through enzymatic or nonenzymatic oxidation. Finally, flavan-3-ols attach the carbocations to yield PAs6. In the third hypothesis, flavan-3-ols are converted to o-quinone by PPO and then to carbocations directly through coupled nonenzymatic oxidation or via flav-3-en-3-ols through nonenzymatic reduction and rearrangement6,30,31. In addition, laccases (LACs) and peroxidases (PERs) could be also involved in this process32. PAs are finally produced through the condensation of flavan-3-ols and their carbocations through nonenzymatic reactions6. This hypothesis was partially supported by recent results from Medicago truncatula, Vitis vinifera, and Camellia sinensis. In M. truncatula, carbocation converted from 4β-(S-cysteyl)-epicatechin (Cys-EC) could be a potential extension unit. Cys-EC could act as precursor of extension unit for nonenzymatic polymerization of PAs in vitro under neutral pH. Loss of LAR function led to the accumulation of Cys-EC and the loss of extractable epicatechin-derived PAs in plants33. In V. vinifera, both Cys-EC and 4β-(S-cysteyl)-catechin (Cys-C) are present. Cys-EC could be converted to epicatechin under the catalysis of VvLARs to serve as the starter unit, whereas Cys-C could be converted to catechin carbocation in vitro under neutral conditions to serve as the extension unit34. In Camellia sinensis, incubation of (−)-epicatechin or (+)-catechin with the extracts of CsANRa transgenic flowers formed dimeric procyanidin B1 or B229.
LACs are multicopper oxidoreductases widely existing in plants and other organisms, such as fungi, bacteria, and insects. They catalyze oxidative coupling reaction of phenol derivatives, resulting in the formation of polymer compounds, such as lignans, stilbenes, and hydroxycinnamic acid derivatives35,36,37,38,39,40,41,42,43. The involvement of LACs in PA polymerization has been implied in various studies. Commercial LAC produced by the fungus Aspergillus oryzae was found to catalyze the oxidation of (+)-catechin to oligomeric products in vitro, although the products generated were structurally different from the natural PAs44. Arabidopsis thaliana TRANSPARENT TESTA 10 (TT10), a LAC-like PPO, was shown to catalyze the oxidation of epicatechin into brown epicatechin oligomers different from procyanidin oligomers45. LACs have been found to catalyze the oxidative polymerization of phenolic monomers46,47,48,49,50,51 and were proposed to be candidates for PA polymerization32,42.
In plants, LACs exist as a large multigene family48,49,52. Approximately half of the family members are post-transcriptionally regulated by the conserved miRNA family, MIR39748,49. In order to test the hypothesis that LACs are involved in PA polymerization in planta, we generated miR397a-overexpressed Salvia miltiorrhiza, a model system for medicinal plants and a popular material in numerous prescriptions of traditional Chinese medicine widely used for treating cardiovascular and cerebrovascular diseases53. With the downregulation of miR397a-targeted SmLACs, the content of flavan-3-ol monomers, such as catechin and epicatechin, was significantly increased, whereas the content of PAs was significantly decreased in the flowers of transgenic S. miltiorrhiza. Similar results were also found in the leaves of transgenic Populus trichocarpa overexpressing miR397a. Enzyme activity analysis of recombinant SmLAC1 that was encoded by a gene targeted by miR397a and highly expressed in S. miltiorrhiza flowers showed that SmLAC1 could catalyze the polymerization of flavan-3-ol monomers to produce PAs. Knockout of SmLAC1 using CRISPR/Cas9 editing technology resulted in a significant decrease in PA accumulation and significant increase of catechin and epicatechin. Overexpression of SmLAC1 increased the content of PAs. Based on these results, we conclude that LACs are involved in PA polymerization in S. miltiorrhiza and P. trichocarpa, and miR397a is a negative regulator of PA biosynthesis.
Results
The elevation of miR397a level downregulates of a subset of SmLACs in S. miltiorrhiza
We previously showed that 23 of 65 SmLACs were targets of MIR397, a conserved miRNA family across different species49. The 21nt sequence of mature S. miltiorrhiza miR397 is identical to P. trichocarpa miR397a48,49. In order to downregulate SmLACs, the Ptr-MIR397a-overexpressing vector constructed before48 was introduced into S. miltiorrhiza (Fig. 1a). Seven transgenic lines (397a-1–397a-7) were obtained. All transgenic plants were cultivated in a greenhouse and flowered at the age of 2 years. No obvious phenotypic differences were observed between the transgenics and the wild type plants. Analysis of miR397a expression showed that the levels of miR397a were significantly elevated in all seven transgenic lines, ranging from a 3.2-fold increase in line 397a-7 to a 20.5-fold in 397a-1 as compared with the wild type (Fig. 1b). On the contrary, miR397a-targeted SmLACs, such as SmLAC1, SmLAC23, SmLAC24, and SmLAC32, were significantly downregulated (Fig. 1c). Consistent with the reduction of SmLAC gene expression, total LAC activity in seven transgenic lines was reduced to 37.3%, 40.2%, 55.9%, 39.5%, 52.3%, 37.4%, and 53.0% of the wild type, respectively (Fig. 1d). These data indicated that the increase of miR397a reduced expression of SmLACs and their protein’s activity in S. miltiorrhiza.

a Generation of miR397a-overexpressed S. miltiorrhiza transgenics. b MiR397a levels in three wild type plants (WT1–WT3) and seven transgenic lines (n = 3 biologically independent experiments). Expression levels are relative with wild type (set as 1). Data are means ± SD of three biologically independent experiments (n = 3). ** indicates P < 0.01 (One-way ANOVA). c Relative expression levels of target genes, including SmLAC1, SmLAC23, SmLAC24, and SmLAC32, in the flowers of wild type and transgenic plants. The expression levels were analyzed using qRT-PCR. SmUBQ10 was used as a control. Expression levels in wild type (WT) were set to 1. Data are means ± SD of three biologically independent experiments (n = 3). d Enzyme activity of LACs in the flowers of wild type and transgenic plants. ** indicates P < 0.01 (One-way ANOVA). Source data are provided in the Source Data file.
The overexpression of miR397a increases flavan-3-ol monomers and decrease PAs in the flowers of S. miltiorrhiza
Since transgenic S. miltiorrhiza lines 397a-1, 397a-4 and 397a-6 showed the most significant accumulation of miR397a and reduction of LAC activity, we then focused on the flowers of these lines for metabolome analysis. The contents of 57 flavonoids and phenolic acids showed significant difference in the transgenic and the wild type flowers (p < 0.05). Among them, 50 metabolites, such as catechin and epicatechin, were significantly increased, whereas the other 12, such as procyanidin B1, B2, and B3, were significantly decreased (Supplementary Fig. 1 and Supplementary Table 1). The highest accumulation of catechin and epicatechin was found in transgenic line 397a-6, in which the level of catechin and epicatechin reached to 9.5-fold and 5.9-fold of that in wild type flowers, respectively. The average fold change of catechin and epicatechin was 7.2 and 4.9, respectively (Supplementary Table 1). The largest decrease of PAs was found for procyanidin B3 in transgenic line 397a-6. The level of procyanidin B3 was about 18.1% of that in wild type flowers. The average levels of procyanidin B1, B2, and B3 in transgenic flowers were 40.3%, 37.3%, and 25.7% of that in wild type flowers, respectively (Supplementary Table 1).
To further confirm the accumulation of flavan-3-ol monomers and the reduction of polymerized PAs in the flowers of miR397a-overexpressed transgenic S. miltiorrhiza, the content of total extractable and non-extractable PAs was determined using a spectrophotometer, and the content of individual component was analyzed using ultra-performance liquid chromatography (UPLC). The results showed that the average levels of extractable and non-extractable PAs in seven transgenic lines were about 58.1% and 52.8% of that in wild type, respectively (Fig. 2a, b). Extraction of polymerized PAs with 70% acetone showed that the color of extracts from the flowers of miR397a-overexpressed transgenic S. miltiorrhiza was relatively lighter than that from wild type flowers (Fig. 2c). All of the polymerized PAs were significantly decreased in all seven transgenic lines (Fig. 2d–h). The average levels of procyanidins A1, A2, B1, B2, and B3 in transgenic flowers were 46.0%, 52.2%, 53.6%, 22.3%, and 41.7% of that in wild type flowers, respectively. On the contrary, the contents of catechin and epicatechin were significantly increased in all seven transgenic lines (Fig. 2i, j). Catechin content increased to 2.9–10.5-fold of wild type, and epicatechin content increased to 4.1–11.3-fold. The decrease of procyanidin contents and the increase of flavan-3-ol monomer contents from UPLC analysis were consistent with those from metabolome analysis. It suggests that elevation of miR397a level may cause significant accumulation of catechin and epicatechin and significant reduction of polymerized PAs in S. miltiorrhiza.

a Extractable PA contents determined using the dimethylaminocinnamaldehyde (DMACA) method and shown as epicatechin equivalents. b Non-extractable PA contents measured using the butanol/HCl method and shown as procyanidin B1 equivalents. c Coloration of PA extracts from wild type and transgenic S. miltiorrhiza flowers. d–h Procyanidins A1, A2, B1, B2, and B3 contents analyzed using UPLC. i, j Catechin and epicatechin contents analyzed using UPLC. All data are the average of three independent biological replicates (n = 3). Error bars show the standard deviations. The mean contents were analyzed using one-way ANOVA test. * and ** indicate P < 0.05 and P < 0.01, respectively. Source data are provided in the Source Data file.
Procyanidin A1, A2, B1, B2, and B3 identified in S. miltiorrhiza
To investigate whether procyanidins A1, A2, B1, B2, and B3 were produced in S. miltiorrhiza, proanthocyanidin compounds were extracted from S. miltiorrhiza flowers and analyzed using UPLC-MS. Chromatograms were scanned and extracted for the compounds with mass/ratio (m/z) = 575 and 577, which corresponded to A- (Fig. 3a) and B-type procyanidins (Fig. 3b), respectively. The results showed that five peaks were found at m/z 575 and three peaks were scanned at m/z 577. Since there were multiple peaks for the compounds, subsequent MS/MS analyses were performed to identify their molecular compositions. MS/MS spectra showed that two compounds with an m/z value of 575 exhibited main fragment ions at m/z 125, 285, 289, 407, 423, and 449 (Fig. 3c), which were characteristic of A-type PAs. Among them, the fragments at m/z 125 and 449 resulted from Heterocyclic Ring Fission (HRF) cleavage of A-type PAs. The fragment at m/z 285 resulted from QM cleavage of A-type PAs. The fragment at m/z 423 resulted from Retro-Diels-Alder (RDA) cleavage (Fig. 3d). MS spectra showed that there were three compounds having an m/z value of 577. MS/MS spectra showed that all of them could be further broken into the same fragment ions at m/z 125, 289, 407, 425, and 451 (Fig. 3e), which were characteristics of B-type PAs. The fragments at m/z 125 and 451 were derived from the cleavage of the HRF (Fig. 3f). Similarly, the fragments at m/z 289 and 425 resulted from the cleavage of QM and RDA (Fig. 3f), respectively. Additionally, the fragment at m/z 407 was formed due to the loss of water from the fragment with m/z 425 (Fig. 3f). The results suggest that both A- and B-type procyanidins, including procyanidins A1, A2, B1, B2, and B3, exist in S. miltiorrhiza.

a Extracted ion chromatogram (EIC) at m/z = 575 showing two peaks of procyanidin A1 and A2. b Extracted ion chromatogram (EIC) at m/z = 575 showing two peaks of procyanidin B1, B2, and B3. c MS/MS spectra of the m/z = 575 peaks of procyanidin A1 and A2 in (b). d MS/MS spectra of the m/z = 577 peaks from procyanidin B1, B2, and B3 in (b). e Fragmentation pattern of procyanidin A1 and A2 in (a). f Fragmentation patterns of procyanidin B1, B2, and B3 in (a). Source data for Fig. 3a–d are provided in the Source Data file.
Overexpression of miR397a increases the contents of flavan-3-ol monomers and decreases the contents of PAs in transgenic P. trichocarpa
Because the sequence of mature S. miltiorrhiza miR397 is identical to P. trichocarpa miR397a, we tested whether miR397 has the conserved function in these two distinct species. We overexpressed Ptr-MIR397a in P. trichocarpa48. Similar to S. miltiorrhiza, no phenotypic differences were observed between transgenic and wild type P. trichocarpa plants. Leaves from four transgenic lines were quantified for contents of total extractable, total non-extractable, and individual PA components. Similar to the results from S. miltiorrhiza, elevation of miR397a level significantly decreased the accumulation of extractable and non-extractable PAs (Fig. 4a, b). The content of total extractable PAs in four transgenic lines (397a-1, 397a-6, 397a-9, and 397a-10) was reduced by 52.4%, 50.0%, 46.7%, and 43.2% as compared to the wild type (Fig. 4a). The total non-extractable PA content was reduced by 52.5%, 52.3%, 49.2%, and 40.4% (Fig. 4b). After extraction with 70% acetone, we could detect procyanidins A1, A2, B1, B2, and B3 in leaf extracts of P. trichocarpa by negative ion mode electrospray MS (Fig. 4c, d). UPLC analysis showed that the contents of procyanidins A1, A2, B1, B2, and B3 were all dramatically reduced in leaves of all the four transgenics compared with wild type. On average, procyanidins A1, A2, B1, B2, and B3 were decreased to 40.6%, 65.5%, 54.5%, 30.5%, and 40.6% of the wild type, respectively (Fig. 4e–i). UPLC analysis also showed that the content of catechin and epicatechin was significantly increased in the leaves of transgenic P. trichocarpa (Fig. 4j, k). On average, catechin and epicatechin increased to 4.4- and 5.5-fold of wild type, respectively. Therefore, the elevation of miR397a level causes significant increase of flavan-3-ol monomers and significant decrease of polymerized PAs in both S. miltiorrhiza and P. trichocarpa.

a Extractable PA contents determined using the dimethylaminocinnamaldehyde (DMACA) method and shown as epicatechin equivalents. b Non-extractable PA contents determined using the butanol/HCl method and shown as procyanidin B1 equivalents. c, d Detection of procyanidins A1, A2, B1, B2, and B3 in leaf extracts of P. trichocarpa through elective ion chromatography (EIC) analysis. e–k The contents of procyanidins A1, A2, B1, B2, B3, catechin and epicatechin in wild type and transgenic P. trichocarpa leaves. All data are the average of three independent biological replicates (n = 3). Error bars show the standard deviations. The mean contents were analyzed using one-way ANOVA test. * and ** indicates P < 0.05 and P < 0.01, respectively. Source data are provided in the Source Data file.
Overexpression of miR397a alternates expression profiles of the PA biosynthetic genes in transgenic S. miltiorrhiza
To elucidate global changes of gene expression in miR397a-overexpressed S. miltiorrhiza, transcriptome was analyzed for flowers from six transgenic lines and three wild type plants. We identified 1857 genes differentially expressed in transgenic lines (transcript abundance ratio <0.5 or >2; p < 0.05) with 1056 genes being downregulated and 801 upregulated (Supplementary Data 1).
Our previous studies identified a total of 65 SmLAC genes, of which 23 were targets of miR397 in S. miltiorrhiza49 and 18 of these 23 targets were expressed in the flowers. These 18 targets were downregulated by 30–87% in transgenics (Supplementary Fig. 2a and Supplementary Data 1). The results are consistent with those from qRT-PCR analysis (Fig. 1c), confirming the roles of miR397a in regulating a subset of SmLAC targets in S. miltiorrhiza flowers. By contrast, seven SmLACs, such as SmLAC8, SmLAC9 and SmLAC38, that were not the targets of miR397a, were upregulated in transgenics (Supplementary Fig. 2a). In addition, nine peroxidase (SmPER) genes were upregulated more than twofold and five SmPPO genes that were thought to be the most likely condensing enzyme for PA polymerization were increased more than threefold in transgenic flowers (Supplementary Fig. 2b). Upregulation of SmPERs, SmPPOs and non-miR397a-targeted SmLACs could be a response of plant cells in alleviating the effect of oxidative stress caused by the reduction of miR397a-targeted SmLACs and the accumulation of flavan-3-ol monomers. Moreover, there were 15 uridine diphosphate glycosyltransferase genes upregulated in transgenic flowers (Supplementary Fig. 2b and Supplementary Data 1). It could represent a mechanism for reducing the over-accumulation of flavan-3-ol monomers by converting their precursors into anthocyanidins.
Transcriptome analysis showed that the expression of phenylpropanoid and flavonoid biosynthetic genes, such as SmPALs, Sm4CLs, SmCHSs, SmDFRs, and SmANS, was decreased in transgenics (Supplementary Fig. 2b). Downregulation of these genes could be resulted from the feedback regulation of flavan-3-ol monomer accumulation through unknown route(s). This suggests that the increase of flavan-3-ol monomer content in transgenics was caused by downregulation of the SmLACs involved in monomer polymerization but not by upregulation of the genes involved in monomer biosynthesis.
SmLAC1 catalyzes the polymerization of flavan-3-ol monomers in vitro
Through transcriptome analysis, we found that 18 miR397a-targeted SmLACs were expressed in the flowers of wild type S. miltiorrhiza and downregulated in transgenics (Supplementary Data 1). Among them, SmLAC1 exhibited the highest expression in the flowers of wild type plants and the most significant downregulation in transgenics. In addition, SmLAC1 was predominantly expressed in the flowers of S. miltiorrhiza49. Thus, we selected SmLAC1 for further analysis to confirm the role of miR397a-regulated SmLACs in PA polymerization and elucidate the underlying mechanism of PA polymerization.
SmLAC1 recombinant proteins were expressed in Escherichia coli and purified by MBP-Tag affinity chromatography (Fig. 5a). Inoculation of SmLAC1 recombinant proteins with (+)-catechin (2,3-trans), (−)-epicatechin (2,3-cis) or both showed the generation of oxidative yellowish products (Fig. 5b), while the control reactions remained colorless, and no products were formed when the purified SmLAC1 recombinant proteins were replaced with the cellular extracts from empty vector controls (Fig. 5b, c). The analysis suggests that, in our reaction conditions, (+)-catechin and (−)-epicatechin cannot be polymerized through nonenzymatic oxidation.

a An SDS-PAGE gel image shows the purified recombinant MBP-SmLAC1 protein. b MBP-SmLAC1 protein catalyzed the conversion of catechin (C) and epicatechin (EC) to yellowish compounds. The reactions without SmLAC1 were used as the controls. c UPLC analysis of the products from catechin and epicatechin under the catalysis of SmLAC1. d MS/MS spectra of the products C1, E1, and CE1. e MS/MS spectra of the products C2, E2, and CE2. Source data for Fig. 5a–c are provided in the Source Data file.
We then elucidated the structure of oxidative products generated under the catalysis of SmLAC1 recombinant proteins using UPLC/Q-TOF-MS/MS. UPLC analysis showed that, when (+)-catechin, (−)-epicatechin and a mixture of (+)-catechin and (−)-epicatechin were used as the substrate, products C1–C10, E1–E10, and CE1–CE7 could be detected at 280 nm (Fig. 5c). Analysis of MS spectra in negative mode [M-H]– showed that the m/z value was 573 for products E3 and E7, 575 for products C2, C3, C4, C6, C8, C10, E2, and CE2, 577 for products C1, E1, E6, CE1, and CE3, 579 for products E9, 861 for products C7 and CE4, 863 for products C9, E4, E5, and CE5, and 865 for product E8, CE6, and CE7 respectively (Fig. 5c). MS/MS spectrum analysis showed that three products with the m/z value of 577, including C1, E1 and CE1 (Fig. 5d), were broken into the main fragment ions at m/z 125, 289, 407, 425, and 451 (Fig. 5d). These fragment ions were characteristic of B-type Pas54,55,56,57,58. The fragments at m/z 125 and 451 resulted from HRF cleavage. The fragments at m/z 289 and 425 resulted from QM cleavage and RDA cleavage, respectively. The fragment at m/z 407 resulted from water loss of the fragment m/z 425. Products C1, E1, and CE1 had the same retention time (RT) with procyanidin B3 (RT 3.3 min), B2 (RT 5.1 min) and B1 (RT 3.4 min) standards, respectively (Fig. 5c). Based on these results, we annotated C1 to procyanidin B3, E1 to procyanidin B2, and CE1 to procyanidin B1.
The products C2, E2, and CE2 had the m/z value of 575 (Fig. 5e). MS/MS spectrum analysis showed that it was broken into the main fragment ions at m/z 125, 285, 289, 407, 423, and 449 (Fig. 5e). These fragment ions were characteristic of A-type PAs and resulted from HRF, RDA and QM cleavages57,58. The retention times of products E2 and CE2 were 7.8 min and 6.6 min, respectively, which were the same as procyanidin A2 and A1.
The products C3, C4, C6 and C8 (m/z 575) exhibited the same MS/MS spectra (Supplementary Fig. 3a–c). These products, together with the products E3 (m/z 573) (Supplementary Fig. 4a), E7 (m/z 573) (Supplementary Fig. 4e), C5 (m/z 575) (Supplementary Fig. 3c), E6 (m/z 577) (Supplementary Fig. 4d), and E9 (m/z 579) (Supplementary Fig. 4g), could be produced from the polymerization of two flavan-3-ol monomers. The products C7 and CE4 had the m/z value of 861 (Supplementary Figs. 3e and 5b). The products C9, E5 and CE5 had the m/z value of 863 (Supplementary Figs. 3g, 4c and 5c). The products E8, CE6 and CE7 had the m/z value of 865 (Supplementary Figs. 3f and 5d, e). The product E10 has the m/z value of 859 (Supplementary Fig. 4h). These products had different MS/MS spectra data and could be produced from the polymerization of three flavan-3-ol monomers. The MS/MS spectra of these products were different from C-type trimers. Polymerization of flavan-3-ol monomers to the products other than C-type trimers had been observed before44. The m/z value of products CE6 and CE7 was 865 (Supplementary Fig. 5d, e). Its MS/MS spectrum is illustrated in Supplementary Fig. 5d. The primary signals were exhibited at m/z 125, 289, 407, 575, and 739 (Supplementary Fig. 5d). These fragment ions were characteristic of PAs and resulted from HRF, RDA and QM cleavage57,58. The MS/MS spectra indicated that CE6 was a natural polymer of three flavan-3-ol monomers. Its chemical structure remains to be elucidated.
We next determined the Michaelis-Menten kinetic parameter (Km), the turnover number (kcat), and the catalytic efficiency (kcat/Km) of SmLAC1 using catechin or epicatechin as the substrate (Supplementary Table 2). The Km, Vmax and Kcat/Km values for epicatechin and catechin were 14.8 and 21.2 µM, 102.3 and 68.0 nkat mg−1, 2.4 × 105 and 2.9 × 105 M−1 S−1, respectively. The results indicated that SmLAC1 had high affinity towards both substrates tested, with the more favorable substrates to be epicatechin.
1H NMR and 13C NMR analysis determines C1, E1, CE1, C2, and E2 to be dimeric B-type and A-type procyanidins
Products C1, E1, CE1, C2, and E2 were purified using semi-preparative HPLC, followed by analysis using 1H NMR and 13C NMR spectroscopy to determine their structures. The MS/MS spectra of C1, E1, and CE1 suggested that they were B-type procyanidin dimers. The parts per million (PPM) values for the 26 protons and 30 carbons in C1, E1 and CE1 at −40 °C were successfully obtained and found to be similar to those of procyanidin B259 (Supplementary Table 3 and Supplementary Fig. 6a). In order to assign protons and carbons, procyanidin B2 was used as a reference due to its shared structural features with C1, E1, and CE1. Specifically, sixteen protons were designated for both the A unit and B unit, namely A2, A3, A4a, A5, A6, A7, A8a, A1’, A2’, A3’, A4’, A5’, A6’, B2, B3, B4a, B5, B6, B7, B8a, B1’, B2’, B3’, B4’, B5’, and B6 (Fig. 6a–c). In addition, ten protons were identified for the ten hydroxyl groups within C1, E1, and CE1 (Fig. 6a–c). To assign all carbons, 13C NMR analysis was performed using procyanidin B2 as the reference, resulting in 30 PPM values for the carbons C1, E1, and CE1 (Fig. 6d–f). These PPM values closely matched those of the corresponding 30 carbons in procyanidin B259. Based on these data, the 30 carbons of C1, E1, and CE1 were established (Fig. 6d–f).

a–c 1H NMR assignments of 15 protons on the six rings and 10 hydroxyl groups in products C1, E1, and CE1, respectively. d–f 13C NMR assignments of 30 carbons in products C1, E1, and CE1, respectively. g, h 1H NMR assignments of 14 protons on the six rings of products C2 and E2, respectively. i, j 13C NMR assignments of 30 carbons in products C2 and E2, respectively.
In 1H NMR spectrum of C1, two AMX systems were observed in the aromatic region (δH 5.8–6.9) with large coupling constants in the region of δH 4.5–3.7 (H-2/H-3/H-4) (Supplementary Fig. 6a). Additionally, the 13C NMR spectrum showed two carbon signals at 83.9 and 82.4, which correspond to C-2 of the C and F rings (Supplementary Fig. 6b). These results indicate the presence of two catechin units. Comparison of the 1H NMR and 13C NMR spectroscopic data with the data reported previously60 suggests that C1 is procyanidin B3, a dimer of catechin ((+)-catechin-(4α→8)-(+)-catechin).
Based on the 1H and 13C NMR spectra, E1 contained an (−)-epicatechin unit as A unit and B unit, respectively. The 1H NMR spectrum of E1 showed two proton signals (4.24, brs and 3.78, brs) assigned to H-3 of A unit and B unit. Along with a set of signals (4.96, brs and 5.08, brs) corresponding to H-2 of A unit and B unit, E1 was confirmed to contain two epicatechin units (Supplementary Fig. 7a). In the 13C NMR spectrum of E1, two carbon signals at 76.9 and 79.4 were observed (Supplementary Fig. 7b). It further supported the presence of two epicatechin units. Compared with the data reported previously59, E1 was unequivocally identified as procyanidin B2, a dimer of epicatechin ((−)-epicatechin-(4β→8)-(−)-epicatechin).
The 11H NMR spectrum of CE1 exhibited methylene proton signals (2.52, brs and 2.60, brs), which were assigned to the H-4 of the A unit as a (+)-catechin moiety (Supplementary Fig. 8a). The 13C NMR spectrum showed that CE1 had an (−)-epicatechin moiety as the B unit, based on the assignment of the carbon signal (36.6) at the C-4 of the B unit, the carbon signal (76.9) at the C-2 of the B unit, and the carbon signal (157.7) at the C-5 of the B unit (Supplementary Fig. 8b). Comparison of the 1H NMR and 13C NMR spectroscopic data with previously reported data59 revealed that CE1 was procyanidin B1, a dimer of epicatechin and catechin ((−)-epicatechin-(4β→8)-(+)-catechin).
MS/MS analysis of products C2 and E2 in negative-ion modes revealed a deprotonated ion [M-H]− at m/z 575, indicating the molecular formula C30H24O12. Based on the m/z ratios and MS/MS fragment profiles, C2 and E2 were classified as A-type procyanidins. The 1H NMR and 13C NMR spectral data were obtained at 25 °C. They provided PPM values for 15 protons and 30 carbons of C2 and E2, respectively (Supplementary Table 4 and Supplementary Fig. 9). Integration and inspection of the changes in chemical shift upon 25 °C failed to identify the hydroxyl protons, which were confirmed by H/D exchange with methanol-d4. The 1H NMR (CD3OD) spectra of C2 and E2 were highly similar to each other (Supplementary Table 4 and Supplementary Fig. 9). Since the PPM values of 15 protons and 30 carbons were similar to those of A1, we used this standard as a reference to assign protons and carbons. The 15 protons were assigned to A and B units, including A3, A4, A6, A8, A2’, A5’, A6’, B2, B3, B4, B6, B2’, B5’, and B6 (Fig. 6g, h). To assign all carbons, 13C NMR analysis was performed using procyanidin A1 as the reference, resulting in 30 PPM values for the carbons C2 and E2 (Fig. 6i, j).
The presence of the isolated AB coupling system at δ 4.0–4.5 with J3,4 = 3.4–3.5 Hz (Supplementary Fig. 9a, c) was ascribed as a diagnostic feature of the C-ring protons of A-type proanthocyanidins61,62. The meta-substitution pattern revealed by the two coupled doublets at around δH 6.0 (H-6, and H-8 in A unit), a residual one proton singlet at δH 6.08 and d 6.09 (H-6 in B unit), and the two AMX coupling systems in the aromatic region (δH 6.7–7.2) suggested a dimeric flavanol (Supplementary Fig. 9a, c). The AB coupling system, the meta-substitution pattern and the two AMX coupling systems confirmed C2 and E2 of A-type procyanidins. The characteristic acetal carbon at 100.16 and 100.41 ppm in 13C NMR spectrum suggested the presence of a doubly linked (A-type) subunit in the structures of C2 and E2 (Supplementary Fig. 9b, d), further identifying them as A-type procyanidins. Direct comparison of E2 spectral data with literature established data63 and comparison of its RT with A-type procyanidin standards showed that E2 was identified as procyanidin A2. C2 had a different RT compared with procyanidin A1 and A2, but its 1H NMR and 13C NMR spectra were similar to those of procyanidin A1 and A2. Based on these results, C2 was deduced to be an A-type procyanidin.
SmLAC1 catalyzes the conversion of B-type to A-type procyanidins
The production of procyanidin A in the reaction with SmLAC1 and substrates indicated that SmLAC1 could catalyze the conversion of B-type procyanidin to A-type. In order to verify this catalytic activity of SmLAC1, in vitro assays were performed using procyanidins B1, B2, and B3 as the substrate. UPLC/Q-TOF-MS/MS analysis of the products generated under the catalysis of SmLAC1 showed that B-type procyanidins were indeed converted to A-type (Fig. 7a–c). In addition, various oxidative products other than the dimers of A-type procyanidins could be produced as described before ref. 64.

a–c UPLC analysis of the products from procyanidins B1, B2, and B3 under the catalysis of SmLAC1. d MS/MS spectra of the products 1, 2, and 3. e–g 1H NMR assignments of 14 protons on the six rings of products 1, 2, and 3, respectively. h–j 13C NMR assignments of 30 carbons in products 1, 2, and 3, respectively.
UPLC detection at 280 nm revealed a total of eleven products, of which one (product 1) was produced with procyanidin B1 as a substrate (Fig. 7a), three (products 2, 4, and 5) were generated from procyanidin B2 (Fig. 7b), and six (products 3, 6–11) were generated with procyanidin B3 as a substrate (Fig. 7c). Analysis of MS spectra in negative mode [M-H]– showed that six of the products, including 1–3 and 6–10 had the m/z value of 575 (Fig. 7d and Supplementary Fig. 10). MS/MS spectrum analyses showed that products 1–3 had the same MS/MS spectra as A-type procyanidin (Fig. 7d). The UPLC RT for products 1 and 2 was 6.3 and 7.8 min, which were identical to procyanidin A1 and A2 standards, respectively. Products 4 (RT 2.2 min) and 5 (RT 3.3 min) produced using procyanidin B2 as a substrate had the m/z value of 573. They exhibited the different MS/MS fragment ions. The product 5 had the same MS/MS fragment ions as the product E3 generated using flavan-3-ol monomer as a substrate (Supplementary Figs. 4a and 10b). In addition, they had the same RT (Fig. 5c). Products 6–10 produced using procyanidin B3 as a substrate had the m/z value of 575, but they had different MS/MS fragment ions (Supplementary Fig. 10c, g). Product 7 exhibited the same MS/MS spectra as C3, C4, and C6 generated using catechin as a substrate, and it had the same UPLC RT as C4. The chemical structures of these products are currently unknown.
In order to further investigate the catalytic activity of recombinant SmLAC1, its enzyme reaction kinetic properties were tested in vitro for procyanidins B1, B2, and B3 and determined from plots of reaction velocity versus substrate concentration. As shown in Supplementary Table 2, the Km value varied from 18.2 µM for procyanidin B2 to 32.1 µM for procyanidin B1. The Vmax value varied from 61.2 nkat mg−1 for procyanidin B2 to 30.6 nkat mg−1 for procyanidin B1. The Kcat/Km value varied from 3.0 × 105 M−1 S−1 for procyanidin B1 to 5.0 × 106 M−1 S−1 for procyanidin B3. Taken together, the results indicated that SmLAC1 had high affinity towards procyanidins B1, B2 and B3, with the more favorable substrates to be procyanidin B2.
Unambiguous assignment of protons and carbons of products 1, 2 and 3 through 1H NMR and 13C NMR spectrum analysis
Products 1, 2, and 3 were further purified by semi-preparative HPLC. They structures were determined using 1H NMR and 13C NMR, which were performed in CD3OD at room temperature. 1H NMR and 13C NMR spectra of all three products showed great similarity to each other and were similar to those of C2 and E2 (Supplementary Table 5, Supplementary Figs. 11 and 12). The m/z value of 575 indicates that products 1, 2, and 3a had molecular formula of C30H24O12 and could be a dimer proanthocyanidins. The chemical shifts of protons and carbons were compared to those of A1, C2, and E2 as references for assignment (Fig. 7e–j). The existence of A-type proanthocyanidin was supported by an isolated AB coupling system at δ 4.0–4.3 with J3,4 = 3.4–3.5 Hz, the meta-coupled doublets at around δ 6.0 (H-6, and H-8 in A unite), a residual one aromatic proton singlet at δ 6.08 and d 6.09 (H-6 in B unite), and two AMX systems (δ 6.8–7.2) in 1H NMR (Supplementary Table 5 and Supplementary Fig. 11). The structures of products 1, 2, and 3 were also confirmed by the presence of one methylene, thirteen methines and sixteen quaternary carbons in the 13C NMR spectra (Supplementary Table 5 and Supplementary Fig. 12). Comparison of their 1H NMR and 13C NMR spectral data with those previously reported61 showed that products 1 and 2 were procyanidin A1 and A2, respectively. Product 3 was generated using procyanidin B3 as a substrate and exhibited similar NMR spectra to products 1 and 2, with some characteristic differences in the chemical shifts (Supplementary Table 5). It suggests that product 3 is an A-type procyanidin derived from procyanidin B3. However, its chemical structure remains to be elucidated.
SmLAC1 polymerizes flavan-3-ol monomers in S. miltiorrhiza
To further confirm the involvement of SmLAC1 in PA biosynthesis, transgenic lines overexpressing SmLAC1 were generated. The transcription level of SmLAC1 was significantly elevated in five overexpression lines (SmLAC-1, SmLAC-2, SmLAC-3, SmLAC-4, and SmLAC-5). The fold changes were 18.9, 32.8, 12.8, 19.4, and 10.1, respectively (Fig. 8a). The average contents of total extractable and non-extractable PAs in transgenic plants were increased by 62.5% and 55.0%, respectively (Fig. 8b, c). UPLC analysis of the leaves of SmLAC1-overexpressed transgenic S. miltiorrhiza showed significant increases of procyanidins A1, A2, B1, B2, and B3 contents (Fig. 8d, e). The average contents of procyanidins A1, A2, B1, B2, and B3 were increased from 1.1, 2.2, 1.8, 1.9, and 2.3 μg g−1 FW in the control to 2.9, 4.8, 6.2, 9.1, and 6.2 μg g−1 FW in transgenic leaves, respectively (Fig. 8g–k). Additionally, the levels of catechin and epicatechin were decreased in all five SmLAC1-overexpressed lines (Fig. 8f, m, l). The results suggest that SmLAC1 plays a significant role in PA polymerization.

a Transcript levels of SmLAC1 in different transgenic lines. Transcripts were determined by qPCR and normalized relative to the expression of SmUBQ10. b Extractable PA levels in leaves of the control and SmLAC1 transgenic lines quantified using DMACA reagent and expressed as epicatechin equivalents. c Non-extractable PA levels evaluated by the butanol-HCl method and expressed as procyanidin B1 equivalents. d–f Elective ion chromatography (EIC) analysis of procyanidins A1, A2, B1, B2, B3, catechin and epicatechin in leaf extracts of the control and SmLAC1-overexpressed transgenic S. miltiorrhiza. g–k. The contents of procyanidins A1, A2, B1, B2, and B3 analyzed using UPLC. l, m Catechin and epicatechin contents in leaves of the control and SmLAC1-overexpressed transgenic S. miltiorrhiza. All data are the average of three independent biological replicates (n = 3). * and ** indicates P < 0.05 and P < 0.01 (One-way ANOVA), respectively. Source data are provided in the Source Data file.
To further investigate the function of SmLAC1 in PA polymerization, we used the CRISPR/Cas9 system to knock out SmLAC1. It resulted in the generation of five positive lines, including smlac1-1, smlac1-2, smlac1-3, smlac1-4, and smlac1-5. Sanger sequencing chromatograms indicated that all five transgenic lines were homozygous and had mutations in both alleles at the same DNA locus (Fig. 9a). UPLC analysis showed that total extractable PA content in the leaves of mutant lines was significantly reduced to 56.5%, 66.7%, 60.9%, 60.8%, and 52.2% of the control, respectively (Fig. 9b). In addition, the average non-extractable PA content in the mutants decreased to 33.9% of the control (Fig. 9c). Furthermore, the levels of procyanidins A1, A2, B1, B2, and B3 in mutant lines were reduced to 49.5%, 33.2%, 41.7%, 35.0%, and 39.7%, respectively (Fig. 9d, e, g–k). Notably, the content of catechin and epicatechin in the leaves of smlac1 mutants was significantly increased (Fig. 9f, l, m). Catechin and epicatechin levels were up to 15 and 16 times higher than those in the control. The results provide strong evidence to support the involvement of SmLAC1 in PA polymerization.

a The sgRNA-edited nucleotide sequences in smlac1 mutants. b Extractable PA levels in leaves of the control and smlac1 mutants quantified using DMACA reagent and expressed as epicatechin equivalents. c Non-extractable PA levels quantified by the butanol-HCl method and expressed as procyanidin B1 equivalents. d, e EIC profiles of procyanidin A1, A2, B1, B2, and B3 contents in the control and smlac1 mutants. f EIC profiles of catechin and epicatechin contents in the control and smlac1 mutants. g, k UPLC analysis of procyanidin A1, A2, B1, B2, and B3 contents in the control and five smlac1 mutants. l, m Catechin and epicatechin contents in leaves of the control and five smlac1 mutants. Values represent means ± SD from three biological replicates (n = 3). * and ** indicate P < 0.05 and P < 0.01 (One-way ANOVA), respectively. Source data are provided in the Source Data file.
Discussion
Plant miRNAs are a class of small noncoding RNAs of about 21 nucleotides in length. They play significant regulatory roles in many aspects of a plant mainly through direct cleavage of gene transcripts with perfect or near-perfect complementarity to the miRNA48,65,66,67,68. MiR397, a miRNA molecule highly conserved in many plant species, has been shown to target LAC gene transcripts for cleavage in P. trichocarpa48, S. miltiorrhiza49, rice69, and A. thaliana70. Elevation of miR397a level severely reduced the expression and total enzyme activity of LACs in company with the reduction of lignin content in the stem differentiating xylem of P. trichocarpa48. Overexpression of OsmiR397 in rice downregulated OsLAC gene expression, increased rice grain size, promoted branch number, accelerated spikelet meristem differentiation, and reduced lignin deposition in stems65,71,72. In Arabidopsis, miR397 targeted LACs to regulate seed number, cadmium tolerance, and lignin biosynthesis73. These results suggest the regulatory role of miR397 in lignin polymerization.
Through genetic transformation and subsequent analysis, we revealed a role of miR397a in regulating PA polymerization in S. miltiorrhiza and P. trichocarpa. We found that elevation of miR397a level caused significant downregulation of SmLACs in the flowers of S. miltiorrhiza enriched with PAs. Total extractable and non-extractable PAs in transgenic flowers were reduced to as low as 38.1% and 37.0% of those in wild type flowers (Fig. 2). Procyanidins B1, B2, and B3, were reduced to as low as 37.7%, 24.4%, and 18.1% when analyzed using an untargeted LC-MS approach (Supplementary Table 1) and to as low as 42.6%, 15.0%, and 28.5% when analysis using UPLC (Fig. 2). Severe reduction of PA content in transgenics suggested the significance of miR397a in PA polymerization. Similar results were also observed in miR397a-overexpressed P. trichocarpa leaves enriched with PAs (Fig. 4). Our work suggested that the regulatory role of miR397 in PA polymerization could be highly conserved in plants.
One hypothesis well accepted is that PAs are formed by a nonenzymatic condensation of flavan-3-ol monomers as the starter units with carbocations derived from leucoanthocyanidin, anthocyanin or flavan-3-ol as the extension units6,16,20,25,26,28,29,30,31,74,75. The formation of these carbocations could be either nonenzymatic26,29 or under the catalysis of PPO6,30,31. The direct evidence included non-enzymatic condensation of dimeric PAs from flavan-3-ols or leucoanthocyanidins via chemical synthesis of quinone methides and carbocation in vitro76 or via direct incubation77.
The other hypothesis is that PPOs, LACs, and PERs are involved in the polymerization of PAs6,32. PPOs are copper-binding enzymes utilizing oxygen to catalyze the conversion of o-diphenols to the corresponding o-quinones. They can also convert monophenols to o-diphenol intermediates and then to o-quinones78,79. The generated o-quinones are highly reactive compounds that can polymerize or react with themselves and other compounds to produce pigments80,81. For PA polymerization, PPOs were proposed to catalyze the oxidation of anthocyanins and flavan-3-ols. Then, the resulting quinone methides were converted to carbocations to serve as the extension units for B-type PAs6,25. The generated B-type PAs could be further converted to A-type under the catalysis of PPO6,82. PERs and LACs were proposed to catalyze the formation of quinone methides, which were further converted to carbocations for PA polymerization32. PERs use hydrogen peroxide to oxidize their substrates. NtFP, a member of the class III peroxidase subfamily, catalyzed the polymerization of flavanols to produce a yellowish dimeric flavonoid called papanridin59. Commercial horseradish PERs were found to catalyze the oxidative coupling of (+)-catechin to form three different biphenyl dimers83. The PERs from Fragaria ananassa catalyzed oxidation of (+)-catechin to yield dehydrodicatechin and (+)-catechin oligomers84. Since these products were not natural PAs, it is still unknown whether PERs are involved in PA polymerization in plants.
As described in the introduction section, previous studies have indicated that LACs are probably involved in PA polymerization in plants44,45,46,47,48. Here we showed that SmLAC1, a miR397a-regulated LAC specifically and highly produced in the flowers of S. miltiorrhiza49, had a high affinity towards flavan-3-ol monomers. SmLAC1 could catalyze the polymerization of flavan-3-ol monomers and the conversion of B-type PAs to A-type in vitro. Both catechin and epicatechin could serve as the starter unit and the extension unit during PA polymerization. Downregulation of a subset of LACs in S. miltiorrhiza and P. trichocarpa through miR397a overexpression caused significant increase in catechin and epicatechin contents and severe decrease of total extractable PA, total non-extractable PA, and procyanidin B1, B2, and B3 contents. P. trichocarpa has five SmLAC1 orthologs (PtLAC13, PtLAC37–PtLAC39, and PtLAC48)49. All of them are targets of miR397a48. Although we have not determined their enzymatic functions, these PtLACs may be also involved in PA polymerization. In addition, overexpression of SmLAC1 resulted in significant increase of extractable and non-extractable PA contents and decrease of catechin and epicatechin contents in S. miltiorrhiza leaves. Consistently, knockout of SmLAC1 led to significant decrease of total extractable PA, non-extractable PAs and polymerized PA contents and increase of catechin and epicatechin contents. Based on the degree of PA content reduction, we conclude that LACs are involved in the polymerization of PA and this polymerization is regulated by miR397a in S. miltiorrhiza and P. trichocarpa. Moreover, we propose that the catalysis of LACs is a main mechanism of PA formation in these two plant species.
In previous hypotheses, carbocations were proposed to play a crucial role in PA polymerization. Various indirect evidences have been obtained to support this viewpoint. For instance, leucocyanidin could be converted into carbocations in vitro through nonenzymatic oxidation under weakly acidic conditions29. Cys-EC could serve as an extension unit for nonenzymatic polymerization of B-type PAs in vitro under neutral pH33. Cys-EC could be converted to epicatechin under the catalysis of V. vinifera LARs and Cys-C could be converted to catechin carbocation in vitro under neutral conditions34. Incubation of (−)-epicatechin or (+)-catechin with Camellia sinensis ANRa transgenic petal extracts resulted in the formation of dimeric procyanidins in nucleophilic solvents29. However, it is still unknown how those carbocations are generated in vivo and what the contribution degree of carbocations to the total PA content is in plants.
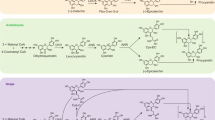
In this study, we found that PAs were polymerized from flavan-3-ols. Both catechin and epicatechin could serve as the starter unit and the extension unit during PA polymerization. The polymerization of B-type and the conversion of B-type PAs to A-type could be mainly catalyzed by LACs. Since LAC-catalyzed phenol coupling is typically processed through the generation of phenol radical intermediates85,86,87,88, PAs may be also polymerized through the same mechanism. Based on these results, we propose a PA polymerization model, in which intermediate radicals play a critical role (Fig. 10).

MiR397a-regulated LACs (e.g., SmLAC1) initiate the polymerization reaction. Both (+)-catechin and (−)-epicatechin can serve as the starter and the extension units.
As depicted in Fig. 10, flavan-3-ols (e.g., (+)-catechin and (−)-epicatechin) act as the starter and the extension units. Ptr-miR397a-targeted LACs (e.g., SmLAC1) initiate the reaction through oxidation of the meta-hydroxyl group on the A-ring. It results in the formation of a free radical. Through resonance, the free radical delocalizes to different positions of the flavan-3-ol. The free radical is stabilized through coupling with the other radical to form a dimer. Among the dimers yielded, those formed through coupling of the free radical at the C8 position of a flavan-3-ol molecule with the free radical at position C4 of the other flavan-3-ol molecule are C8-C4-linked B-type PA dimers (e.g., procyanidin B1, B2, and B3).
A-type PAs are epimers of B-types. They are formed through an oxidative intramolecular reaction of B-type PAs. SmLAC1 oxidizes B-type PAs at the ortho-hydroxyl group on the B-ring of the extension unit and the meta-hydroxyl group on the A-ring of the starter unit to form two free radicals. The free radical generated at the ortho-hydroxyl group delocalizes to the C2 position at the C-ring of the extension unit through resonance. Coupling of the free radical at the C2 position of the C-ring of the extension unit and the free radical at the hydroxyl group of the C7 position of the A-ring of the starter unit forms the 2-O-7 linkage, resulting in the generation of A-type PAs.
Methods
Genetic transformation of S. miltiorrhiza
Ptr-MIR397a and SmLAC1 overexpression vector were introduced into Agrobacterium tumefaciens strain GV3101 using the freeze-thaw method48,89. Agrobacterium-mediated leaf disk transformation method was performed to obtain transgenic S. miltiorrhiza90,91,92. Leaf explants were co-cultivated with agrobacteria in the dark for two days and then transferred to Murashige-Skoog agar media supplemented with 1.0 mg L−1 6-BA, 0.1 mg L−1 NAA, 300 mg L−1 cefotaxime and 30 mg L−1 kanamycin to induce shoots. Transgenic shoots were excised and then transferred to 1/2 Murashige-Skoog agar media supplemented with antibiotics for rooting. Transgenics were verified by PCR analysis on genomic DNA using primers 5′-CGCACAATCCCACTATCCTTCGCAA-3′ and 5′-CGCTGCACTCAATGATGGTTCTCCA-3′.
RNA isolation and qRT-PCR
Total RNA and miRNA were extracted from powdered samples using the EASYspin Plus Plant Kit (Aidlab, Beijing, China) and the EASYspin Plant microRNA Kit (Aidlab) according to the manufacturer’s protocol, respectively. Quantitation of miR397a was determined using qRT-PCR. Approximately 2 μg of RNA per sample were reversely transcribed to first-strand cDNA using the SuperScript II First-Strand Synthesis System (Invitrogen). Reverse transcription of miRNA was performed using the Mir-X miRNA First-Strand Synthesis Kit (Takara). qRT-PCR was conducted with three biological replicates using SYBR Green Master Mix (Applied Biosystems) on a CFX96IM Real-Time PCR Detection System (Bio-Rad). All primers used are listed in Supplementary Table 6. Relative transcript levels were calculated using the formula for comparative Ct value.
SmLAC enzyme assays
SmLAC purification and enzyme assay were carried out48. Three grams of flowers were homogenized in 15 mL extraction buffer (50 mM Tris-HCl, pH 6.8, 1 mM EDTA, 2% SDS, 10% (V/V) glycerol). Proteins were concentrated using Amicon Ultra-4 Centrifugal Filter-100K (Millipore). Soluble protein extract was loaded into a Sephadex G200 column and sequentially eluted with 0.2 M NaCl in phosphate buffer (pH 7.0) at a flow rate of 2 min ml−1. The elution was collected. LAC activity was determined in a 200 μl reaction mixture composed of 100 mM buffer (pH 5.0), 1 mM 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid) (ABTS), and 50 μl of protein extract. The reaction was incubated at 30 °C for 20 min and then stopped by adding 100 μl SDS. Generated cationic radicals were determined by measuring the increase of absorbance at 420 nm. The activity values are expressed in enzymatic activity units (U) as the amount of substrate converted per minute.
Metabolic profiling
Fresh flowers were freeze-dried under vacuum and ground into powder using a mixer mill. Total 100 mg of powder was extracted overnight at 4 °C with 1 ml of 70% aqueous methanol. After centrifugation at 10,000 × g for 10 min, the extracts were filtered using a 0.22 μm membrane. Metabolome analyses were performed using a UPLC system coupled with a tandem mass spectrometer. The separation of metabolites was carried out on a WATERS ACQUITY UPLC HSST3 C18 column (1.8 μm, 2.1 mm × 50 mm). The injection volume was 5 μl. Gradient elution was performed at a flow rate of 0.4 ml/min with 0.04% acetic acid-water (solvent A) and 0.04% acetic acid-acetonitrile (solvent B). The elution procedure was from 5% B at 11.0 min to 95% B at 12.0 min to 5% B at 12.1 min and then held at 5% B until 15 min.
Mass spectrometry was carried out using API 4500 QTRAP LC/MS/MS mode equipped with an ESI Turbo Ion-Spray interface. The analysis was operated in a negative ion mode and controlled by Analyst 1.6 software (AB Sciex). ESI was performed with the following parameters: capillary voltage of 5500 V, fragmentation voltage of 135 V, and source temperature of 550 °C. Ion source gas I (GSI), gas II (GSII) and curtain gas (CUR) were set at 55.0, 60.0, and 25.0 psi, respectively.
All mass spectral data were imported into Progenesis QI. Quantitative peak area information was extracted and exported for data plotting and principal component analysis (PCA). The data set was log-normalized. PCA and PLS-DA analyses were performed to determine the overall differences of metabolites. ANOVA test was performed to identify differential metabolites based on fold change ≥ 2 or ≤ 0.5 and P-value < 0.01.
RNA sequencing and transcriptome analysis
Total RNA was extracted from nine S. miltiorrhiza flower samples using the EASYspin Plus Plant Kit (Aidlab). Sequencing libraries were prepared using the NEBNext Ultra™ RNA Library Prep Kit for Illumina® (NEB) following the manufacturer’s recommendations. The libraries were sequenced on an Illumina Hiseq 2500 platform to generate 150 bp paired-end reads. Clean reads were obtained after removing low-quality reads, reads with adaptor sequences, and reads with more than 5% unknown bases in raw reads. The data were then aligned with S. miltiorrhiza genome using Hisat2. Differential expression analysis was performed using Cuffdiff (v2.1.1) software with the default parameters. Differentially expressed genes were identified with the threshold of fold-change ≥ 2 or ≤ 0.5 and adjusted p-value < 0.05.
Determination of PA content
About 0.1 g of samples was ground into powder in liquid nitrogen. The powder was ultrasonically extracted with 1 mL of PA extraction solvent (PES; 70% [v/v] acetone with 0.5% [v/v] acetic acid) in ice water for 30 min93. The obtained homogenate was centrifuged at 15,000 × g for 5 min. The supernatant was collected, whereas the remaining residues were re-extracted for twice. Supernatants from three extraction processes were pooled for further extraction of extractable PAs. An equal volume of chloroform was added to the supernatants, vortexed for 30 s, and centrifuged at 5000 × g for 5 min. Then, the supernatant was extracted twice with chloroform and twice with hexane. The resulting aqueous phase (the extractable PA) was dissolved in 50% methanol after freeze-drying. Total extractable PA content was quantified by spectrophotometer after reaction with DMACA (0.2% [w/v] DMACA in methanol/HCl (7:1, v/v)), and UPLC analysis93.
Non-extractable PA was analyzed using the butanol-HCl method93. One ml of butanol-HCl (95:5, v/v) reagent was added to the dried residues after PES extraction (70% [v/v] acetone with 0.5% [v/v] acetic acid). The mixture was sonicated in ice water for 1 h, boiled for 1 h, and then centrifuged at 15,000 × g for 5 min after cooling to room temperature. The content of non-extractable PAs was measured by absorbance at 550 nm. Procyanidin B1 was used as a standard.
Expression and purification of recombinant SmLAC1 proteins
The open reading frame of SmLAC1 was amplified using primer F (5′- ttcagggagaattggccatggATGGAGCGCTCTTTGATGTTTT-3′) as the forward primer and primer R (5′-gacgaattccccgggggatccGCACTTGGGAAAATCAGCCG-3′) as reverse primer. Purified PCR products were ligated into XF00245 vector to fuse with a MBP tag at the N-terminus using ClonExpress Ultra One Step Cloning Kit (Vazyme, China), and transformed into competent E. coli strain DH5α, and then subjected to sequencing analysis. The correct sequence was transformed into E. coli strain Rosetta (DE3) for protein expression. E. coli cells were cultivated in LB medium at 37 °C until the OD600 reached 0.6–0.8. To induce the expression of SmLAC1 recombinant protein, 0.5 mM IPTG was added. The cells were then cultivated at 16 °C for 24 h. MBP-tagged SmLAC1 proteins were purified using MBP-TAG Dextrin Resin as per the manufacturer’s manual. Purified proteins were concentrated and desalted in 100 mM acetate buffer (pH 5.0) using Amicon Ultra-4 Centrifugal Filter-100K (Millipore). Aliquots were stored at −80 °C. The purity of recombinant SmLAC1 was examined by SDS-PAGE. Protein concentration was quantified by Bradford assays.
Analysis of enzyme activity and reaction kinetics
Enzyme activity assay was performed in a 100 μL reaction mixture consisting of 100 mM acetate buffer (pH 5.0), 10–20 μg of recombinant SmLAC1, and 200 μM substrate (catechin or epicatechin). The reaction was incubated at 28 °C for 2 h and then terminated by adding 100 μL of methanol. The reaction products were injected into a UPLC or an LC-MS/MS system for analysis. Enzyme activity was calculated by measuring the increase of product content. To determine the kinetic parameters, SmLAC1 was incubated with different concentrations of catechin, epicatechin, or catechin and epicatechin, ranging from 5 to 300 μM. Enzyme assays were performed in triplicate at each substrate concentration. The Vmax and Km values were calculated using Graphpad Prism 9 with nonlinear regression analysis.
UPLC analysis
UPLC analysis was carried out on UPLC (Waters) equipped with an Acquity BEH C18 1.7μm, 2.1 × 100 mm column. The mobile phase consisted of 0.1% formic acid in water (solvent A) and 0.1% formic acid in methanol (solvent B). The flow rate was 0.2 mL min−1. The detection was monitored at 280 nm wavelength. The column was maintained at ambient temperature. The gradient procedure was 0.00–1.50 min, 10%–20% B; 1.50–3.00 min, 20%–20% B; 3.00–7.00 min, 20%–40% B; 7.00–9.50 min, 40%–90% B; 9.50–12.00 min, 90%–100% B. Compounds were identified based on the comparison of retention times and ultraviolet spectra with those of authentic standards.
Q-TOF analysis
MS and MS/MS data of oxidative products were acquired using an Acquity UPLC system coupled to a Xevo G2-XS Q-TOF mass spectrometer (Waters) operatedin negative electrospray mode with a capillary voltage of 2.0 kV. The mobile phase consisted of 0.1% (v/v) formic acid in water (A) and 0.1% (v/v) formic acid in methanol (B). Two μL of sample was injected. The flow rate was 0.2 mL min−1. The gradient procedure was as follows: 0.00–1.50 min, 10%–20% B; 1.50–3.00 min, 20%–20% B; 3.00–7.00 min, 20%–40% B; 7.00–9.50 min, 40%–90% B; 9.50–12.00 min, 90%–100% B. Instrument parameters, data acquisition and data processing were controlled by Masslynx 4.1 and Xcalibur. The ESI source parameters were set as follows: sample cone, 30 V; source temperature, 100 °C; desolvation temperature, 20 °C; desolvation gas (N2) flow, 900 L h−1; cone gas (N2) flow, 50 L h−1. Compounds were detected by MSE centroid analysis at a resolving power of 30,000 with a scan time of 1 s. For full-scan MS analysis, the data acquisition range of mass-to-charge ratio (m/z) was from 50 to 1000. Leucine encephalin [M-H]– ion as lock mass (m/z 554.2615) was infused via the LockSpray ion source to ensure accuracy and reproducibility. Accurate mass and fragment ions were obtained using MassLynx™ software (version 4.1, Waters).
Structure analysis by NMR
Enzyme activity assay was expanded from 100 μL to 100 ml. The products were purified using semipreparative HPLC on Waters 2535Q instrument equipped with an Agilent Zorbax SB C18 column (5 μm, 9.4 × 250 mm). 1H NMR and 13C NMR of the purified products were obtained on Bruker Avance III-400 or III-600 spectrometer. Spectra were recorded at 25 °C and −40 °C. The isolated compounds were identified by comparison of spectra data with literatures reported previously.
Overexpression and CRISPR/cas9-mediated knockout of SmLAC1
Full-length SmLAC1 open-reading frame was amplified using 2× Phanta Master (Vazyme, China) with primer F (5′-tcttagaattcccggggatccATGGAGCGCTCTTTGATGTTTT-3′) as the forward primer and primer R (5′-caccacgtgtgaattggtgaccGCACTTGGGAAAATCAGCCG-3′) as the reverse primer. The obtained cDNA was cloned into a pCAMBIA1391 vector between BamH I and BstE II sites using the ClonExpress MultiS One Step Cloning kit (Vazyme, Nanjing, China). A SmLAC1-knockout vector was constructed using the CRISPR/Cas9 system94. To design the sgRNA sequences, two putative target sites located at the first exon of SmLAC1-coding sequence were selected. PCR amplification was performed using pDT1T2 vector as a template with two primer pairs containing two individual guide RNA (sgRNA) sequences of SmLAC1 (Supplementary Table 6). The products were purified and digested with BsaI and ligated into pHEE401E binary vector. All constructs were confirmed by sequencing.
All plasmids were isolated and transferred into A. tumefaciens strain GV3101, which was then used to transform S. miltiorrhiza leaves to generate transgenic plantlets using leaf disk transformation method95. Transgenics were verified by PCR using primers listed in Supplementary Table 6. The fragments surrounding the two target sites were amplified by PCR using gene-specific primers Mut-F and Mut-R. Purified PCR products were sequenced using the primer Mut-F. Positive transgenic shoots were cultured in 1/2 MS medium at 25 °C with a photoperiod of 16 h of light and 8 h of dark.
Statistical analysis
To test for statistical significance, two tailed unpaired student’s t test was used to determine the significance between wild type and transgenic plants in the PA composition, as well as the kinetics of SmLAC1 toward catechin and epicatechin. Statistics were performed in SPSS Statistics (v22; IBM) using one-way ANOVN at significance level of 0.05.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data supporting the findings of this work are available within the paper and the Supplementary Information files. A reporting summary for this article is available as a Supplementary Information file. The transcriptome datasets are available at NCBI BioProject PRJNA1210854. The mass spectrometry data have been deposited into the Science Data Bank [https://doi.org/10.57760/sciencedb.19944]. Source data are provided with this paper.
References
Constabel, C. P. Molecular controls of proanthocyanidin synthesis and structure: prospects for genetic engineering in crop plants. J. Agric. Food Chem. 66, 9882–9888 (2018).
Jun, J. H., Xiao, X., Rao, X. & Dixon, R. A. Proanthocyanidin subunit composition determined by functionally diverged dioxygenases. Nat. Plants. 4, 1034–1043 (2018).
Debeaujon, I., Leon-Kloosterziel, K. M. & Koornneef, M. Influence of the testa on seed dormancy, germination, and longevity in Arabidopsis. Plant. Physiol. 122, 403–414 (2000).
Joanisse, G. D., Bradley, R. L., Preston, C. M. & Bending, G. D. Sequestration of soil nitrogen as tannin-protein complexes may improve the competitive ability of sheep laurel (Kalmia angustifolia) relative to black spruce (Picea mariana). N. Phytol. 181, 187–198 (2009).
Deng, Y. & Lu, S. Biosynthesis and regulation of phenylpropanoids in plants. Crit. Rev. Plant. Sci. 36, 257–290 (2017).
Dixon, R. A., Xie, D. Y. & Sharma, S. B. Proanthocyanidins–a final frontier in flavonoid research? N. Phytol. 165, 9–28 (2005).
Aron, P. M. & Kennedy, J. A. Flavan-3-ols: nature, occurrence and biological activity. Mol. Nutr. Food Res. 52, 79–104 (2008).
Prior, R. L. & Gu, L. Occurrence and biological significance of proanthocyanidins in the American diet. Phytochemistry 66, 2264–2280 (2005).
Kawahara, S. I. et al. Identification and characterization of oligomeric proanthocyanidins with significant anti-cancer activity in adzuki beans (Vigna angularis). Heliyon 5, e02610 (2019).
Odai, T., Terauchi, M., Kato, K., Hirose, A. & Miyasaka, N. Effects of grape seed proanthocyanidin extract on vascular endothelial function in participants with prehypertension: a randomized, double-blind, placebo-controlled study. Nutrients 11, 2844 (2019).
Rauf, A. et al. Proanthocyanidins: a comprehensive review. Biomed. Pharmacother. 116, 108999 (2019).
Jekabsone, A. et al. Investigation of antibacterial and antiinflammatory activities of proanthocyanidins from Pelargonium sidoides DC root extract. Nutrients 11, 2829 (2019).
Yang, H. et al. Effect of tea root-derived proanthocyanidin fractions on protection of dentin collagen. J. Int. Med. Res. 48, 300060519891303 (2020).
Aerts, R. J., Barry, T. N. & McNabb, W. C. Polyphenols and agriculture: beneficial effects of proanthocyanidins in forages. Agric. Ecosyst. Environ. 75, 1–12 (1999).
Dixon, R. A., Liu, C. G. & Jun, J. H. Metabolic engineering of anthocyanins and condensed tannins in plants. Curr. Opin. Biotechnol. 24, 329–335 (2013).
Stafford, H. A. Pathway to proanthocyanidins (condensed tannins), flavan-3-ols, and unsubstituted flavans. In: Flavonoid Metabolism (ed. Stafford, H. A.) 63–99 (Boca Raton, FL, USA: CRC Press, 1990).
Rousserie, P., Rabot, A. & Geny-Denis, L. From flavanols biosynthesis to wine tannins: what place for grape seeds? J. Agric. Food Chem. 67, 1325–1343 (2019).
Eichenberger, M. et al. The catalytic role of glutathione transferases in heterologous anthocyanin biosynthesis. Nat. Catal. 6, 927–938 (2023).
Lu, N. Revisiting decade-old questions in proanthocyanidin biosynthesis: current understanding and new challenges. Front. Plant. Sci. 15, 1373975 (2024).
Lu, N., Jun, J. H., Li, Y. & Dixon, R. A. An unconventional proanthocyanidin pathway in maize. Nat. Commun. 14, 4349 (2023).
Tanner, G. J. et al. Proanthocyanidin biosynthesis in plants. Purification of legume leucoanthocyanidin reductase and molecular cloning of its cDNA. J. Biol. Chem. 278, 31647–31656 (2003).
Xie, D. Y., Sharma, S. B., Paiva, N. L., Ferreira, D. & Dixon, R. A. Role of anthocyanidin reductase, encoded by BANYULS in plant flavonoid biosynthesis. Science 299, 396–399 (2003).
Peng, Q. Z. et al. An integrated approach to demonstrating the ANR pathway of proanthocyanidin biosynthesis in plants. Planta 236, 901–918 (2012).
Jun, J. H., Lu, N., Docampo-Palacios, M., Wang, X. & Dixon, R. A. Dual activity of anthocyanidin reductase supports the dominant plant proanthocyanidin extension unit pathway. Sci. Adv. 7, eabg4682 (2021).
Xie, D. Y. & Dixon, R. A. Proanthocyanidin biosynthesis–still more questions than answers? Phytochemistry 66, 2127–2144 (2005).
Dixon, R. A. & Sarnala, S. Proanthocyanidin biosynthesis-a matter of protection. Plant. Physiol. 184, 579–591 (2020).
Yu, K., Dixon, R. A. & Duan, C. A role for ascorbate conjugates of (+)-catechin in proanthocyanidin polymerization. Nat. Commun. 13, 3425 (2022).
Creasey, L. L. & Swain, T. Structure of condensed tannins. Nature 208, 151–153 (1965).
Wang, P. et al. Functional demonstration of plant flavonoid carbocations proposed to be involved in the biosynthesis of proanthocyanidins. Plant. J. 101, 18–36 (2020).
Haslam, E. Symmetry and promiscuity in procyanidin biochemistry. Phytochemistry 16, 1625–1640 (1977).
Xie, D. Y., Sharma, S. B. & Dixon, R. A. Anthocyanidin reductases from Medicago truncatula and Arabidopsis thaliana. Arch. Biochem. Biophys. 422, 91–102 (2004).
Peng, Q. Z. & Xie, D. Y. Progress on the polymerization mechanism of proanthocyanidins during their biosynthesis. Acta Bot. Boreal. -Occident. Sin. 32, 624–632 (2012).
Liu, C., Wang, X., Shulaev, V. & Dixon, R. A. A role for leucoanthocyanidin reductase in the extension of proanthocyanidins. Nat. Plants. 2, 16182 (2016).
Yu, K., Jun, J. H., Duan, C. & Dixon, R. A. VvLAR1 and VvLAR2 are bifunctional enzymes for proanthocyanidin biosynthesis in grapevine. Plant. Physiol. 3, 1362–1374 (2019).
Gianfreda, L., Xu, F. & Bollag, J. M. Laccases: a useful group of oxidoreductive enzymes. Bioremed. J. 3, 1–26 (1999).
Canas, A. I. & Camarero, S. Laccases and their natural mediators: biotechnological tools for sustainable ecofriendly processes. Biotechnol. Adv. 28, 694–705 (2010).
Chaurasia, P. K., Bharati, S. L., Yadava, S. & Yadav, R. S. Purification, characterization and synthetic application of a thermally stable laccase from Hexagonia tenuis MTCC-1119. Indian. J. Biochem. Biophys. 52, 60–67 (2015).
Yonekura-Sakakibara, K. et al. Seed-coat protective neolignans are produced by the dirigent protein AtDP1 and the laccase AtLAC5 in Arabidopsis. Plant. Cell. 33, 129–152 (2021).
Al-Khayri, J. M. et al. Stilbenes, a versatile class of natural metabolites for inflammation-an Overview. Molecules 28, 3786 (2023).
Marrone, G. et al. Possible beneficial effects of hydrolyzable tannins deriving from Castanea sativa L. in internal medicine. Nutrients 16, 45 (2023).
Tan, Y. Z. et al. Lignans from the root of Valeriana jatamansi and their biological evaluation. J. Asian Nat. Prod. Res. 25, 810–817 (2023).
Ube, N. et al. Molecular identification of a laccase that catalyzes the oxidative coupling of a hydroxycinnamic acid amide for hordatine biosynthesis in barley. Plant. J. 115, 1037–1050 (2023).
Zhao, Y., Jiang, C., Lu, J., Sun, Y. & Cui, Y. Research progress of proanthocyanidins and anthocyanidins. Phytother. Res. 37, 2552–2577 (2023).
Osman, A. M., Wong, K. & Fernyhough, A. The laccase/ABTS system oxidizes (+)-catechin to oligomeric products. Enzyme Microb. Technol. 40, 1272–1279 (2007).
Pourcel, L., Routaboul, J. M., Cheynier, V., Lepiniec, L. & Debeaujon, I. Flavonoid oxidation in plants: from biochemical properties to physiological functions. Trends Plant. Sci. 12, 29–36 (2007).
Freudenberg, K. Biosynthesis and constitution of lignin. Nature 183, 1152–1155 (1959).
Berthet, S. et al. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems. Plant. Cell. 23, 1124–1137 (2011).
Lu, S. et al. Ptr-miR397a is a negative regulator of laccase genes affecting lignin content in Populus trichocarpa. Proc. Natl. Acad. Sci. USA. 110, 10848–10853 (2013).
Li, C., Li, D., Zhou, H., Li, J. & Lu, S. Analysis of the laccase gene family and miR397-/miR408-mediated posttranscriptional regulation in Salvia miltiorrhiza. PeerJ 7, e7605 (2019).
Cardullo, N., Muccilli, V. & Tringali, C. Laccase-mediated synthesis of bioactive natural products and their analogues. RSC Chem. Biol. 3, 614–647 (2022).
Zoclanclounon, Y. A. B. et al. Characterization of peroxidase and laccase gene families and in silico identification of potential genes involved in upstream steps of lignan formation in sesame. Life (Basel). 12, 1200 (2022).
Turlapati, P. V., Kim, K. W., Davin, L. B. & Lewis, N. G. The laccase multigene family in Arabidopsis thaliana: towards addressing the mystery of their gene function(s). Planta 233, 439–470 (2011).
Lu, S. Biosynthesis and regulatory mechanisms of bioactive compounds in Salvia miltiorrhiza, a model system for medicinal plant biology. Crit. Rev. Plant. Sci. 40, 243–283 (2021).
Smith, C. A. et al. METLIN: a metabolite mass spectral database. Ther. Drug. Monit. 27, 747–751 (2005).
Huang, Y., Chen, L., Feng, L., Guo, F. & Li, Y. Characterization of total phenolic constituents from the stems of Spatholobus suberectus using LC-DAD-MS(n) and their inhibitory effect on human neutrophil elastase activity. Molecules 18, 7549–7556 (2013).
Poupard, P., Sanoner, P., Baron, A., Renard, C. M. & Guyot, S. Characterization of procyanidin B2 oxidation products in an apple juice model solution and confirmation of their presence in apple juice by high-performance liquid chromatography coupled to electrospray ion trap mass spectrometry. J. Mass. Spectrom. 46, 1186–1197 (2011).
Lin, L. Z., Sun, J., Chen, P., Monagas, M. J. & Harnly, J. M. UHPLC-PDA-ESI/HRMSn profiling method to identify and quantify oligomeric proanthocyanidins in plant products. J. Agric. Food Chem. 62, 9387–9400 (2014).
Yuzuak, S., Ballington, J. & Xie, D. Y. HPLC-qTOF-MS/MS-based profiling of flavan-3-ols and dimeric proanthocyanidins in berries of two muscadine grape hybrids FLH 13-11 and FLH 17-66. Metabolites 8, 57 (2018).
Zhu, Y., Yuzuak, S., Sun, X. & Xie, D. Y. Identification and biosynthesis of plant papanridins, a group of novel oligomeric flavonoids. Mol. Plant. 16, 1773–1793 (2023).
Huang, H. M. et al. HPLC/ESI-MS and NMR analysis of chemical constitutes in bioactive extract from the root nodule of Vaccinium emarginatum. Pharm. (Basel). 14, 1098 (2021).
Nam, J. W. et al. Subtle chemical shifts explain the NMR fingerprints of oligomeric proanthocyanidins with high dentin biomodification potency. J. Org. Chem. 80, 7495–7507 (2015).
Nam, J. W. et al. Absolute configuration of native oligomeric proanthocyanidins with dentin biomodification potency. J. Org. Chem. 82, 1316–1329 (2017).
Lou, H. et al. A-type proanthocyanidins from peanut skins. Phytochemistry 51, 297–308 (1999).
Wang, C. M. et al. Structure elucidation of procyanidins isolated from rhododendron formosanum and their anti-oxidative and anti-bacterial activities. Molecules 20, 12787–12803 (2015).
Rhoades, M. W. et al. Prediction of plant microRNA targets. Cell 110, 513–520 (2002).
Palatnik, J. F. et al. Control of leaf morphogenesis by microRNAs. Nature 425, 257–263 (2003).
Sunkar, R. & Zhu, J. K. Novel and stress-regulated micro-RNAs and other small RNAs from. Arabidopsis. Plant. Cell. 16, 2001–2019 (2004).
Lu, S., Sun, Y. H. & Chiang, V. L. Stress-responsive microRNAs in Populus. Plant. J. 55, 131–151 (2008).
Zhang, Y. C. et al. Overexpression of microRNA OsmiR397 improves rice yield by increasing grain size and promoting panicle branching. Nat. Biotechnol. 31, 848–852 (2013).
Wang, C. Y. et al. MiR397b regulates both lignin content and seed number in Arabidopsis via modulating a laccase involved in lignin biosynthesis. Plant. Biotechnol. J. 12, 1132–1142 (2014).
Swetha, C. et al. Major domestication-related phenotypes in Indica rice are due to loss of miRNA-mediated laccase Silencing. Plant. Cell. 30, 2649–2662 (2018).
Lian, J. P. et al. MicroRNA397 promotes rice flowering by regulating the photorespiration pathway. Plant. Physiol. 194, 2101–2116 (2024).
Ali, S. et al. miR397-LACs mediated cadmium stress tolerance in Arabidopsis thaliana. Plant. Mol. Biol. 8, 11 (2023).
Wang, P. et al. Comprehensive analysis of metabolic fluxes from leucoanthocyanins to anthocyanins and proanthocyanidins (PAs). J. Agric. Food Chem. 68, 15142–15153 (2020).
Lu, N., Jun, J. H., Liu, C. G. & Dixon, R. A. The flexibility of proanthocyanidin biosynthesis in plants. Plant. Physiol. 190, 202–205 (2022).
Ohmori, K., Ushimaru, N. & Suzuki, K. Oligomeric catechins: an enabling synthetic strategy by orthogonal activation and C(8) protection. Proc. Natl. Acad. Sci. USA 101, 12002–12007 (2004).
Jiang, X. et al. Analysis of accumulation patterns and preliminary study on the condensation mechanism of proanthocyanidins in the tea plant [Camellia sinensis]. Sci. Rep. 5, 8742 (2015).
Sánchez-Ferrer, A., Rodríguez-López, J. N., García-Cánovas, F. & García-Carmona, F. Tyrosinase: a comprehensive review of its mechanism. Biochim. Biophys. Acta. 1247, 1–11 (1995).
McLarin, M. A. & Leung, I. K. H. Substrate specificity of polyphenol oxidase. Crit. Rev. Biochem. Mol. Biol. 55, 274–308 (2020).
Bachem, C. W. B. et al. Antisense expression of polyphenol oxidase genes inhibits enzymatic browning in potato tubers. Nat. Biotechnol. 12, 1101–1105 (1994).
Bittner, S. When quinones meet amino acids: chemical, physical and biological consequences. Amino Acids. 30, 205–224 (2006).
Tanaka, T., Kondou, K. & Kouno, I. Oxidation and epimerization of epigallocatechin in banana fruits. Phytochemistry 53, 311–316 (2000).
Hosny, M. & Rosazza, J. P. Novel oxidations of (+)-catechin by horseradish peroxidase and laccase. J. Agric. Food Chem. 50, 5539–5545 (2002).
López-Serrano, M. & Barceló, A. R. Comparative study of the products of the peroxidase-catalyzed and the polyphenoloxidase-catalyzed (+)-catechin oxidation. Their possible implications in strawberry (Fragaria x ananassa) browning reactions. J. Agric. Food Chem. 50, 1218–1224 (2002).
Mattinen, M. L. et al. Laccase-catalyzed polymerization of tyrosine-containing peptides. FEBS J. 272, 3640–3650 (2005).
Pourcel, L. et al. TRANSPARENT TESTA10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat. Plant. Cell. 17, 2966–2980 (2005).
Mnich, E. et al. Phenolic cross-links: building and de-constructing the plant cell wall. Nat. Prod. Rep. 37, 919–961 (2020).
Hüttel, W. & Müller, M. Regio- and stereoselective intermolecular phenol coupling enzymes in secondary metabolite biosynthesis. Nat. Prod. Rep. 38, 1011–1043 (2021).
Holsters, M. et al. Transfection and transformation of Agrobacterium tumefaciens. Mol. Gen. Genet. 163, 181–187 (1978).
Wang, M. et al. ARGONAUTE genes in Salvia miltiorrhiza: identification, characterization, and genetic transformation. Methods Mol. Biol. 1640, 173–189 (2017).
Liu, M., Chen, X., Wang, M. & Lu, S. SmPPT, a 4-hydroxybenzoate polyprenyl diphosphate transferase gene involved in ubiquinone biosynthesis, confers salt tolerance in Salvia miltiorrhiza. Plant. Cell Rep. 38, 1527–1540 (2019).
Liu, M. et al. Functional analysis of polyprenyl diphosphate synthase genes involved in plastoquinone and ubiquinone biosynthesis in Salvia miltiorrhiza. Front. Plant. Sci. 10, 893 (2019).
Pang, Y., Peel, G. J., Sharma, S. B., Tang, Y. & Dixon, R. A. A transcript profiling approach reveals an epicatechin-specific glucosyltransferase expressed in the seed coat of Medicago truncatula. Proc. Natl. Acad. Sci. USA. 105, 14210–14215 (2008).
Wang, C. et al. FIONA1 is an RNA N6-methyladenosine methyltransferase affecting Arabidopsis photomorphogenesis and flowering. Genome Biol. 23, 40 (2002).
Zhou, H. et al. Genome-wide identification and functional analysis of Salvia miltiorrhiza microRNAs reveal the negative regulatory role of Smi-miR159a in phenolic acid biosynthesis. Int. J. Mol. Sci. 25, 5148 (2024).
Acknowledgements
This work was supported by the CAMS Innovation Fund for Medical Sciences (CIFMS) [grant No. 2021-I2M-1-029].
Author information
Authors and Affiliations
Contributions
S.L. conceived and designed the study. C.L. and S.L. analyzed and interpreted data. C.L., X.Q., M.J., X.H., D.L., X.C., X.P., and F.S. acquired data. C.L. and S.L. wrote the original draft. Q.L., D.Y.X., V.L.C., and S.L. discussed, reviewed and edited the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks William Laing and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Li, C., Qiu, X., Hou, X. et al. Polymerization of proanthocyanidins under the catalysis of miR397a-regulated laccases in Salvia miltiorrhiza and Populus trichocarpa. Nat Commun 16, 1513 (2025). https://doi.org/10.1038/s41467-025-56864-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-56864-0
