
Abstract
The consolidation process stabilizes a new initially labile memory. This consolidation could operate on a shorter timescale during wakefulness after initial motor learning. Within micro-offline learning states, sequences of simple individual actions learned through interleaved practice are condensed into a unified skill through a time-dependent consolidation process occurring during wakeful periods. While emerging evidence links Glutamate and GABA modulations in the primary motor cortex (M1) to motor learning, its relationship with micro-offline consolidation processes in brief resting states during motor learning is unclear. To investigate this issue, we employed Transcranial magnetic stimulation (TMS) to evaluate whether interindividual variation of different neurotransmitters at rest influences motor learning consolidation in humans. Our results point to the role of GABAB in micro-offline motor consolidation processes during motor learning in M1. This finding could have an important impact on planning neuropharmacology or non-invasive brain stimulation approaches in clinical domains, such as post-stroke rehabilitation.
Similar content being viewed by others
Introduction
The ability to perform complex sequences of movements quickly and accurately is critical for many motor skills. Although training improves performance in a large variety of motor sequence tasks, a growing body of research suggests that incorporating periods of waking rest alongside practice1,2 can also contribute to significant performance improvements. Interestingly, while these periods of rest were traditionally considered as wasted or unfruitful3, recent researchers have suggested that the newly formed memory undergoes an “offline” process during intervals of wakefulness, during which the initially unstable memory is consolidated and converted into an engram4,5 that could withstand interferences and the elapse of time3,6,7. The gains observed during brief rest periods referred to as micro-offline gains (i.e., the performance boost from the last practice trial to the first trial after the rest), suggest that motor consolidation takes place on a much shorter timescale than previously believed1,8,9, even before the sleep-related consolidation process1. In terms of behavior, micro-offline improvements align closely with the initial online learning curve until reaching a performance plateau, contributing to overall early learning. However, the process through which the brain consolidates sequences of distinct action representations into refined and temporally precise skills during these micro-offline gains remains unclear. One potential mechanism being considered involves neuroplastic changes occurring at different levels of the central nervous system10,11 with a primary focus on the functional reorganization of the primary motor cortex (M1), which seems to have a role in motor learning consolidation3,12. Such plastic reorganization has been hypothesized to be facilitated by reduced cortical inhibition via modulation of γ-aminobutyric acid (GABA) neurotransmission13,14,15,16,17. More specifically, it has been reported that a decrease in GABA after acute deafferentation results in the facilitation of LTP-like plasticity within M118. Supporting this link, it was shown that the administration of GABA receptor agonists impairs both the retention in visuomotor learning19 and the plasticity induced by motor practice13.
In agreement with such findings, decreases in GABA concentration have also been demonstrated using proton magnetic resonance spectroscopy (MRS) both in the acute phase of recovery after stroke20, and during motor learning15,21. With regards to this last aspect, a study by Kolasinski et al. required participants to perform a visually cued serial reaction time task21. The authors measured the GABA concentration in three different groups: the “learning group” was explicitly informed to expect a cue-related repeating sequence; the “movement group” was informed the cue was not predictive of a sequence, and the “rest group” was asked to perform a passive resting task. Results showed a reduction in GABA concentration in the learning group but not in either the motor or the resting group. No changes in Glutamate were observed in any group. Furthermore, M1 GABA measured early in task performance was correlated with the degree of subsequent learning. These findings indicated that higher GABA concentrations were linked to poorer motor learning outcomes.
Despite being a promising technique, one limitation of MRS is its current inability to distinguish between the various functional GABAergic pools (i.e., GABAA and GABAB) and their temporal dynamics, which are believed to play different roles in metabolism, synaptic neurotransmission, and cortical inhibition22,23. Thus, despite the existence of previous neuroimaging findings, which support the hypothesis that LTP-like plasticity within M1 is critically dependent on GABA modulation15,16,17,24, to date, no existing studies have investigated whether distinct forms of inhibition (i.e., GABAA or GABAB mediated) measured within M1 before motor learning in wakefulness can predict offline motor consolidation in humans. To address this issue, we used transcranial magnetic stimulation (TMS) to investigate at-rest motor-evoked potentials (MEPs)—a probe of corticospinal excitability (CSE)—as well as short-interval and long-interval intracortical inhibition (SICI/LICI) and facilitation (sICF/ICF). While short intracortical inhibition (SICI) is supposed to be mediated by post-synaptic potentials via fast ionotropic GABAA receptors, long intracortical inhibition (LICI) seems to reflect inhibitory post-synaptic potentials produced via slower metabotropic GABAB receptors25,26,27. Intracortical facilitation (ICF) is believed to be mediated by NMDA glutamatergic receptors28,29,30,31,32, and short intracortical facilitation (sICF) seems to depend on trans-synaptic excitation of intracortical interneurons that project to the corticospinal neurons33,34,35. Our findings suggest intriguing and distinctive roles for neurotransmitters within M1 in facilitating the consolidation of motor learning on a microscale, intricately linked to temporal mechanistic plasticity in the human motor system. They also highlight a role for neurochemical modifications to both motor consolidation and plasticity in the human brain, which may hold important clinical implications in rehabilitative settings such as in stroke and brain injury.
Results
Sixteen right-handed healthy adults were requested to tap an imposed sequence on a response pad with their left hand (Fig. 1a). Firstly, we evaluated the improvement in skill learning performance by comparing the mean tapping speed of the first trial block (T0) with the tapping speed at the block (T) in which 95% of the performance gains are first achieved at the individual level. Student’s t-test showed an improvement in motor learning performance (t(15) = −6.88, p < 0.01 Cohen’s d = −1.72) when 95% of the learning had been achieved compared to the first block (Fig. 2).

Panel a Participants learned the motor-skill task consisting of alternating practice and rest periods of 10 s duration for a total of 36 blocks. In each practice task block, participants were asked to repeatedly tap the sequence 4-1-3-2-4 as quickly and accurately as possible using their left, non-dominant hand. Practice task blocks were interleaved with 10-s waking rest intervals (inter-practice rest). Note that motor learning performance increases occur during the intervening periods of waking rest (micro-offline gains), but not during the active practice task (micro-online learning). Panel b Mean performance of all participants. Skill was measured as the correct sequence typing speed (sequences [seq]/s). Mean average performance (mean ± SEM) increased rapidly during early learning (the set of trials within which 95% of total learning occurred, trials 1–11 (left panel) extracted by means of the curve fitting performance (right panel). The vertical black line in the right panel corresponds to the block T where 95% of total learning occurred. Micro-online changes were calculated as the difference in tapping speed (keypresses/s) between the first and last correct sequence within a practice period (black in inset) and micro-offline changes as the difference between the last correct sequence within a practice period and the first of the next practice period (red in inset).

Data points in the violin plot depict the sum of changes in performance measured as the average inter-tap interval within correct sequences (tapping speed measured in keypresses/s) in the first block (T0) and in the block (T) where 95% of total learning occurred in each participant.
Secondly, we aimed to identify whether the amount of both inhibitory and facilitatory (i.e., ICF and sICF) neurotransmitters, assessed by TMS on the motor area before motor learning, were associated with both the performance of learning that occurred in block T (micro-online-T) and the brief motor consolidation during the rest period in the same block (micro-offline-T). Notably, the different inhibitory intracortical paired-pulse TMS (pp-TMS) protocols, allow us to discriminate between GABAA (SICI) and GABAB (LICI) neurotransmitters. Using Bayesian correlation matrix analyses, we found very strong evidence toward the H1 for a negative relationship between LICI and Micro-offline-T learning (R = −0.834; BF10 = 468 Table 1; Fig. 3), such that participants who showed a lower level of GABAB neurotransmitter (disinhibition) have a better motor consolidation. No other significant correlation was reported (see Table 1). To test the GABAB contribution, we compared the correlation with the dependent correlations of GABAA, using Hittner’s test. The correlation of GABAB with micro-offline performance differed significantly from equivalent correlations between GABAA and the same behavioral measure: Hittner’s Z p < 0.05 (z = −2.8535, p-value = 0.0043).

Relationship between baseline GABA concentrations and micro-offline learning consolidation when 95% of total learning occurred. In the left panel, the short intracortical inhibition corresponding to GABAA modulation is reported; in the right panel, the long intracortical inhibition corresponds to GABAB neurotransmitter modulation.
In line with the previous analysis, a linear regression model was tested between the GABAB and the estimates of the individual motor learning fitting coefficients, and participant performance (micro-online-T and micro-offline-T) to evaluate the predictability of the factors used. The model with Micro-offline-T learning explained a moderate proportion of the variance R2 = 0.69 (BF10 > 100) (Table 2). Micro-offline-T learning (B = −0.58, BFinclusion = 52.6) provides moderate evidence for being included in the model, while there is moderate evidence supporting that the other variables (BFinclusion < 0.5) should not be included (Table 3; Fig. 4).
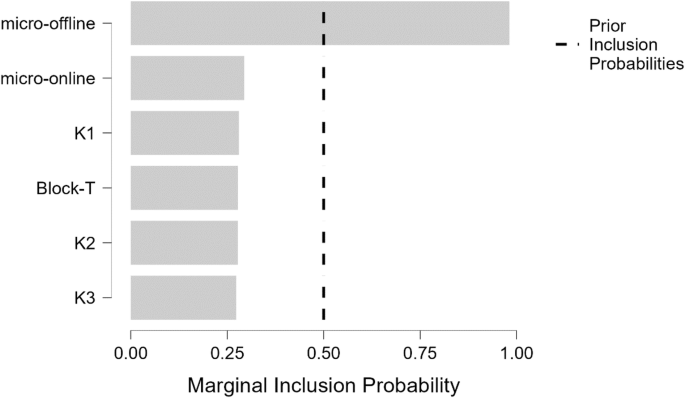
Bar graph of posterior inclusion probabilities for the Bayesian linear regression of the LICI data. The dashed line represents the prior inclusion probabilities.
Discussion
Proficiency in motor skills is essential for navigating daily tasks in challenging environments. Through consistent practice, actions become faster, more precise, and easier to perform. This process, known as the consolidation process, transforms an initially fragile memory into a resilient and enduring one36,37,38. Recently, evidence has suggested that the consolidation operates on a shorter timescale, such as between blocks in the initial phase of learning, and this could potentially dramatically shorten the time required for motor learning and consolidation1,3,39. Interestingly, an enhancement in motor learning has been reported after a period of waking rest, and this phenomenon has been defined as off-line learning1,2,8,9. The aim of the current study was to reveal the underlying neurotransmitter mechanisms subtending such rapid and offline enhancement of motor performance. To investigate this issue, we used a motor consolidation task that has previously been shown to promote continuous motor learning1,2,8, with the aim of assessing whether the GABAergic or glutamatergic concentrations in M1 may support offline learning gains following online motor learning. Although the reduction of intracortical inhibition is an important substrate for optimal motor learning in humans, most studies, including ours, did not find any correlation between the degree of SICI changes and learning13,40,41. Specifically, our data revealed that only GABAB and not GABAA modulation (at rest) correlates with a rapid form of motor consolidation when early motor learning has occurred. Our results are in line with several evidence suggesting a pivotal role of GABAB in motor memory consolidation42,43,44.
All this evidence suggests that while both types of GABA receptors are involved in motor learning processes, GABAB receptors, with their slower and prolonged action, may play a more significant role in motor memory consolidation than GABAA receptors, which act on a faster timescale. Our results not only discriminate the functional role of GABAB in motor learning consolidation but additionally provide evidence that the amount of GABAB can predict the micro-offline-T gain when 95% of total learning has occurred. Interestingly, this result is explained by micro-offline, but not micro-online, improvements. Supporting this concept, evidence suggests that improvements in performance while acquiring a new skill predominantly happen during the rest periods coupled with practice, rather than during the active practice itself. This supports the hypothesis that “much, if not all” skill learning occurs offline during rest rather than during online practice1,2,39. In line with our results, previous TMS45,46,47, pharmacological14, and MRS15,21 studies have shown a reduction in GABAergic inhibition following motor learning. This increase in excitability caused by learning is frequently linked to enhancements in motor performance and provides evidence for the involvement of the GABA inhibitory system in driving M1 plasticity during the acquisition of skills. In summary, inhibitory networks on M1 can influence neural plasticity48 specifically by GABAB mediation49.
A key role in determining the degree of cortical plasticity is played by the balance between excitation and inhibition (E-I). Indeed, an increased excitation or disinhibition enhances neuronal responses to external stimuli and facilitates synaptic plasticity processes50,51. Moreover, E–I balance has been suggested as a factor that can influence significant aspects of cognition and behavior51. The increased association between Glutamate and GABA suggests a learning-related phenomenon, possibly signifying a more precise and regulated processing of the acquired experience in the motor cortex during consolidation stages. It is crucial to emphasize that the absence of evidence of a glutamatergic correlation with motor consolidation does not rule out the potential contribution of glutamatergic mechanisms to motor learning. Indeed, different GABA-modulated inhibitory neurons selectively affect specific domains of excitatory neurons, allowing them to control different aspects of the spatial and temporal activity of the Glutamate receptors52. While we did not assess alterations in Glutamate and GABA levels during the learning phase, it is plausible that the upswing in Glutamate and GABA coupling after the task might signify a physiological “rebound” effect. This effect could be an effort to restore physiological functions that might have been disrupted during the learning process, to their normal state24. On the other hand, the E–I balance may be linked to the storage of short-term memory. In particular, given the heightened vulnerability of motor memories to interference immediately after learning, enhancing the coupling between Glutamate and GABA may play a role in fortifying the initial stages of motor consolidation, thereby stabilizing the trace of the motor memory53,54. Thus, greater GABA inhibitory tone could strongly stabilize the newly learned skill by protecting it from interference55,56. Interestingly, it has recently been hypothesized that inhibitory activity during retrieval is crucial for memory selectivity; and inhibitory synaptic plasticity during memory consolidation plays a key role in the formation of selective engrams4.
An alternative hypothesis is based on how the information is stored. In fact, besides the enhancement of synaptic strength induced by learning, synapses also store information by altering their structure through rapid Hebbian plasticity57,58,59,60,61, mainly similar to long-term potentiation (LTP-like)62,63,64. In this context, GABA may have the role of regulating Hebbian plasticity by offering a comprehensive assessment of the overall performance of the entire network65. Increased protein synthesis and dendritic spine remodeling have been well documented during the late phases of LTP, resulting in the expansion of current spines or the creation of new ones50. Furthermore, motor learning has been shown to promote the formation and stabilization of new spines in M1, which is expressed specifically in M1 engram cells5,66. This hypothesis is in agreement both with studies that have shown how intense local release of Glutamate or GABA can induce post-synaptic dendritic spine formation67 and studies that have illustrated the pivotal role of GABAergic disinhibition in initiating long-term potentiation (LTP)-like plasticity and fostering LTP-like synaptic responses in M168,69,70. A potential limitation of our study is that pp-TMS measures represent an indirect way to measure GABA and Glutamate levels within M1, and studies investigating the associations between MRS-assessed GABA levels and TMS-assessed GABA measures have reported mixed findings (for a comprehensive review on the topic see ref. 71). Thus, future studies should combine MRS and TMS measures to provide complementary and comprehensive information on glutamatergic and GABAergic neurotransmission. Moreover, our study does not address two key factors in motor learning: the strengthening of motor memories over time72 and the role of repetition in increasing resistance to interference73. As our neurophysiological measures were limited to a single session, the findings were restricted to the pre-skill acquisition phase, and potential consolidation effects were not assessed. Given that skill improvements can occur both during and after motor practice, future research could explore learning-induced cortical changes during these later stages of motor learning, which are likely to engage broader neural networks beyond the primary motor cortex (M1)74. Our data have significant implications for the creation of effective interventions designed to enhance motor skills and address motor impairments in rehabilitation settings. The motor consolidation phenomenon, facilitated by a decrease in cortical inhibition via modulation of the neurotransmitter GABA75,76 could provide a model in the acute phase of recovery after stroke17,20. Indeed, widespread alterations of inhibitory neurotransmission after stroke seem to be a crucial potential mechanism77 in functional recovery and deserve special attention78. Exploring the function of GABA in learning mechanisms could offer insights for innovative interventions targeting GABA modulation aimed at enhancing functional recovery.
Methods
Participants
The study cohort consisted of 16 right-handed participants between the ages of 27 and 43 (mean age 30.25 [SD: 5]; 10 females and 6 males). The number of participants was determined on the basis of a power analysis based on a similar correlation between GABA and behavioral indices of motor performance79. This analysis indicated that a sample size of ~16 participants is necessary to achieve a statistical power (1 − β) of 0.90 (two-tailed α = 0.05; H1 corr p_ac = −0.69). Analysis was performed with G*Power software80. All participants were right-handed, as assessed by the Edinburgh handedness inventory81 (EHI). A minimum EHI score of 40 was required to confirm right-handedness (EHI: M = 92.5, SD = 31). Participants had no prior history of drug or alcohol abuse or neurological, psychiatric, or sleep disorders. Prior to the TMS studies, participants had to be free of any drugs and were instructed to be, alcohol-, and caffeine-free for 12 h before and during the study period. The experimental setting was approved by the Ethics Committee of the Department of Psychology of Bologna University (Prot. 0210065, 27/07/23) and was carried out in accordance with the ethical standards of the revised Declaration of Helsinki (2013). All participants gave their written and informed consent to take part in the study.
Motor learning task
The sequential finger tapping task required participants to press four keys on a response pad (Cedrus LS-LINE, Cedrus Corp) with the fingers of their left (nondominant) hand, repeating the five-element sequence (trials): 4-1-3-2-4, as quickly and as accurately as possible for a period of 10 s. Each block consisted of a 10 s practice period followed by a 10 s rest period1,8,82,83. The numeric sequence (4-1-3-2-4) was displayed at the top of the screen at all times to exclude any working memory component. Each key press produced a white dot on the screen, forming a row from left to right, rather than the number itself, so as not to provide accurate feedback. After completion of a full iteration of the sequence, the dots were removed. During the 10 s interleaved rest periods, the sequence was replaced with a string of five “X” symbols, which participants were instructed to fixate on. Individual Keypress timing (ms) and a number of sequences were recorded for behavioral data analysis. Participants were asked to perform 36 blocks. Rest periods (10 s) were interleaved between blocks. The task lasted for a total of 12 min. Visual stimuli and task instructions were presented, and keypress responses were recorded using a custom script running in Matlab84,85.
Electromyography recording
Surface electromyography (EMG) was recorded from the left first dorsal interosseous (FDI) muscle by means of a wireless system (Zerowire EMG, Aurion, Italy) with a tendon-belly montage. EMG signals were digitized (2 kHz) and acquired with a CED Power1401-3A board (Cambridge Electronic Design, Cambridge, UK). All the acquired data were stored for offline analysis using the Signal 3.09 software (Cambridge Electronic Design, Cambridge, UK).
Transcranial magnetic stimulation (TMS)
Participants were seated on a comfortable armchair during all the experimental sessions, with their left arm on an armrest. Paired-pulse TMS (pp-TMS) protocols were administered through a 70 mm figure-of-eight focal coil connected to a Magstim BiStim2 monophasic stimulator (The Magstim Company, Whitland, UK). The FDI optimal scalp position (OSP) was found by moving the coil in 0.5 cm steps over the right M1 hand area and using a slightly suprathreshold stimulus. Resting motor threshold (rMT) was defined as the lowest intensity that evoked a motor-evoked potential (MEP) with >50 μV amplitude in 5 out of 10 consecutive trials while the participants kept the FDI muscle relaxed86,87. For each participant, the FDI hotspot was marked on a head cap to maximize coil placement consistency through the testing sessions. The coil was positioned tangentially to the scalp following the orthodox method, with the handle pointed backward, angled 45° from the midline, perpendicular to the central sulcus. Motor evoked potentials (MEPs) were measured using 120% of the individual rMT. Each neurophysiological index was recorded at rest in a separate session immediately before the behavioral session, to avoid any sleep effects on motor acquisition and consolidation88. Inhibition and facilitation indices, measured by paired-pulse TMS protocols, were determined from the ratio of the mean MEP amplitude.
Paired-pulse TMS measures: intracortical inhibition indices (SICI and LICI)
Short intracortical inhibition (SICI) is measured via a pp-TMS paradigm with a first subthreshold conditioning stimulus (CS) followed, 2.5 ms later, by a second suprathreshold test stimulus (TS). Here we set the CS at 80% and the TS at 120% of individual rMT. SICI may represent short-lasting inhibitory postsynaptic potentials in corticospinal neurons28,89 by means of GABAA receptor-mediated fast intracortical inhibition in M128,90.
Long intracortical inhibition (LICI) consists of two suprathreshold stimuli; the first pulse (CS) elicits an MEP that inhibits the second pulse (TS). We set both the CS and the TS at 120% of the individual rMT with an ISI of 100 ms. It has been proposed that the LICI reflects GABAB receptor-mediated slow inhibition in M128,91. LICI was expressed as the ratio between TS and the previous CS MEP amplitude, to avoid modulations driven by local excitability changes in M1.
Paired-pulse TMS measures: intracortical facilitation indices
Short intracortical facilitation (sICF) was obtained by a pp-TMS protocol, with the first stimulus at 120% followed by a second stimulus at 80% of the rMT, with 1.4 ms ISIs70,92.
Intracortical facilitation (ICF) was elicited by a pp-TMS protocol with the CS set at 80% and the TS at 120% of the rMT, with an ISI of 15 ms. This protocol normally shows a facilitation, which is believed to be mediated by NMDA glutamatergic receptors93,94.
Behavioral data processing
Analysis of motor performance was analyzed as previously described1,8 .
Tapping speed was found using Eq. (1)1, and was quantified as the average of the time intervals between adjacent key presses within correct sequences8,95,96 divided by 1000 (key presses/s).
where intervali represents the time interval between keypresses i and i + 1 in a given sequence.
Performance within each trial was calculated as the mean tapping speed of all correctly performed sequences, including correct sequences the participant has not completed by the end of the trial8,97,98. The number of correct sequences was calculated per trial, also including correct sequences the participant had not completed by the end of the trial.
Modeling of performance curves and the determination of the early learning block cutoff
For each participant, we determined the cutoff block, T, in which 95% of the performance gains are achieved. First, we fit the Individual participant performance curves of mean tapping speed and number of correct sequences per block, B(t), with the following exponential function Eq. (2):
where k1−3 represent the initial performance (function intercept), maximum performance (function plateau) and learning rate (function slope) respectively; and t ∈ [1, +∞) represents trial. Parameters k1−3 were estimated by gradient descent, with the objective function defined as the root mean square error between B and L functions (see Eq. (3)):
where \(\lambda {||}{K}_{2}^{2}{||}\) is the L2 regularization term with λ = 0.1.
A constrained nonlinear least-squares method (MATLAB’s lsqcurvefit, trust-region-reflective algorithm) was then used to estimate parameters K1 (boundary constraints = [0,5]), K2 ([0,15]) and K3 ([0,2]) from these observed tapping speed data. Boundaries remained constant throughout all following experiments to avoid overfitting within each experimental condition. Next, for each participant, from Eq. (3) we estimated the end of early learning as the block T after 95% of the total learning had occurred. In practice, this value can be estimated as Eq. (4):
Thus, we set T to the first block where 95% of the learning had been achieved.
Calculation of micro-online and micro-offline gains during early learning
We studied trial-by-trial early learning by dissecting performance improvements occurring during practice (micro-online) and during rest (micro-offline) periods, as previously reported1,8,99.
Micro-online learning refers to the difference in tapping speed (keypresses/second) between the last and the first block of a practice block (Fig. 1) and quantified learning that occurred during short periods of practice. Specifically, we focused on the micro-online learning (micro-online-T) of the first trial where 95% of the learning had been achieved (T) (Fig. 2a, b).
Micro-offline learning refers to the difference in tapping speed between the first trial of a practice block and the last trial of the previous block (Fig. 1) and quantified learning that occurred during short periods of rest. In particular, we investigated the micro-offline (micro-offline-T) learning blocks of early learning cutoff corresponding to the first block where 95% of the learning had been achieved (T) (Fig. 1b).
Motor learning analysis
To verify motor learning, we compared the mean trial tapping speed of the first block (T0) and the block (T), in which 95% of the performance gains are achieved at the individual level. First, we checked all data for normality using the Shapiro–Wilk test. Since the data does not differ significantly from a normal distribution (Tapping speed_T0: W = 0.94 p = 0.45; Tapping speed_T: W = 0.96 p = 0.78), the comparison was performed by two-tailed paired sample Student’s t-test.
Bayesian Pearson correlation analysis
Bayesian analysis used JASP software (JASP Team, version 1.18.03) with conventional thresholds for Bayes factors (BF) representing substantial (>3), strong (>10), and very strong (>30) evidence in favor of hypotheses100,101,102. Due to the relatively small sample of participants tested, we employed Bayesian correlation analysis to obtain more robust results103. We assessed the Bayesian scores of each correlation using Bayesian correlation pairs in JASP on the mean values for each participant, to assess whether evidence supports the presence or absence of a correlation. Finally, we compared all possible GABA correlations using the cocor toolbox, developed by104. Comparison of correlation statistics was undertaken using Hittner’s test105: Hittner, May, and Silver’s (2003) modification of Dunn and Clark’s z106 using a back transformed average Fisher’s (1921) Z procedure.
Bayesian linear regression
We conducted a linear regression, examining whether LICI (GABAB neurotransmission) predicts the individual subjective performance fitting estimates k1−3, the cutoff T, Micro-online and Micro-offline learning in which 95% of the performance gains are first achieved at the individual level. We applied a beta (a = b = 1) prior to the models, which assumes that the model sizes are equally likely before observing any data, and we applied a JZS prior (r scale = 0.354) to the regression coefficients.
Data availability
The electromyographic and behavioral datasets used and/or analyzed during the current study are available at the link https://osf.io/xbdwu/.
Code availability
Custom written code is available upon request by contacting the Lead Contact, Pasquale Cardellicchio (pasquale.cardellicchio@g(oogle)mail.com).
References
Bönstrup, M. et al. A rapid form of offline consolidation in skill learning. Curr. Biol. 29, 1346–1351.e4 (2019).
Buch, E. R., Claudino, L., Quentin, R., Bönstrup, M. & Cohen, L. G. Consolidation of human skill linked to waking hippocampo-neocortical replay. Cell Rep. 35, 109193 (2021).
Wang, Y. et al. Memory consolidation of sequence learning and dynamic adaptation during wakefulness. Cereb. Cortex 34, bhad507 (2024).
Tomé, D. F. et al. Dynamic and selective engrams emerge with memory consolidation. Nat. Neurosci. 27, 561–572 (2024).
Josselyn, S. A. & Tonegawa, S. Memory engrams: recalling the past and imagining the future. Science 367, eaaw4325 (2020).
Robertson, E. M. & Cohen, D. A. Understanding consolidation through the architecture of memories. Neuroscientist 12, 261–271 (2006).
Criscimagna-Hemminger, S. E. & Shadmehr, R. Consolidation patterns of human motor memory. J. Neurosci. 28, 9610–9618 (2008).
Bönstrup, M., Iturrate, I., Hebart, M. N., Censor, N. & Cohen, L. G. Mechanisms of offline motor learning at a microscale of seconds in large-scale crowdsourced data. npj Sci Learn 5, 1–10 (2020).
Jacobacci, F. et al. Rapid hippocampal plasticity supports motor sequence learning. Proc. Natl Acad. Sci. USA 117, 23898–23903 (2020).
Nuara, A. et al. The value of corticospinal excitability and intracortical inhibition in predicting motor skill improvement driven by action observation. Neuroimage 266, 119825 (2023).
Caroni, P., Donato, F. & Muller, D. Structural plasticity upon learning: regulation and functions. Nat. Rev. Neurosci. 13, 478–490 (2012).
Mizes, K. G. C., Lindsey, J., Escola, G. S. & Ölveczky, B. P. The role of motor cortex in motor sequence execution depends on demands for flexibility. Nat. Neurosci. 27, 2466–2475 (2024).
Ziemann, U., Muellbacher, W., Hallett, M. & Cohen, L. G. Modulation of practice-dependent plasticity in human motor cortex. Brain 124, 1171–1181 (2001).
Bütefisch, C. M. et al. Mechanisms of use-dependent plasticity in the human motor cortex. Proc. Natl Acad. Sci. USA 97, 3661–3665 (2000).
Floyer-Lea, A., Wylezinska, M., Kincses, T. & Matthews, P. M. Rapid modulation of GABA concentration in human sensorimotor cortex during motor learning. J. Neurophysiol. 95, 1639–1644 (2006).
Stagg, C. J., Bachtiar, V. & Johansen-Berg, H. The role of GABA in human motor learning. Curr. Biol. 21, 480 (2011).
Bachtiar, V. & Stagg, C. J. The role of inhibition in human motor cortical plasticity. Neuroscience 278, 93–104 (2014).
Levy, L. M., Ziemann, U., Chen, R. & Cohen, L. G. Rapid modulation of GABA in sensorimotor cortex induced by acute deafferentation. Ann. Neurol. 52, 755–761 (2002).
Johnstone, A., Grigoras, I., Petitet, P., Capitão, L. P. & Stagg, C. J. A single, clinically relevant dose of the GABAB agonist baclofen impairs visuomotor learning. J. Physiol. 599, 307–322 (2021).
Głodzik-Sobańska, L. et al. GABA in ischemic stroke. Proton magnetic resonance study. Med. Sci. Monit. 10, 88–93 (2004).
Kolasinski, J. et al. The dynamics of cortical GABA in human motor learning. J. Physiol. 597, 271–282 (2019).
Belelli, D. et al. Extrasynaptic GABAa receptors: form, pharmacology, and function. J. Neurosci. 29, 12757–12763 (2009).
Martin, D. L. & Rimvall, K. Regulation of gamma-aminobutyric acid synthesis in the brain. J. Neurochem. 60, 395–407 (1993).
Eisenstein, T., Furman-Haran, E. & Tal, A. Early excitatory-inhibitory cortical modifications following skill learning are associated with motor memory consolidation and plasticity overnight. Nat. Commun. 15, 1–17 (2024).
Cardellicchio, P., Dolfini, E., Hilt, P. M., Fadiga, L. & D’Ausilio, A. Motor cortical inhibition during concurrent action execution and action observation. Neuroimage 208, 116445 (2020).
Cardellicchio, P., Dolfini, E., Fadiga, L. & D’Ausilio, A. Parallel fast and slow motor inhibition processes in Joint Action coordination. Cortex. 133, 346–357 (2020).
Cardellicchio, P., Koch, G., Fadiga, L. & D’Ausilio, A. Motor overload: GABAergic index of parallel buffer costs. Brain Stimul. 14, 1106–1108 (2021).
Ziemann, U. et al. TMS and drugs revisited 2014. Clin. Neurophysiol. 126, 1847–1868 (2015).
Ziemann, U. TMS and drugs. Clin. Neurophysiol. 115, 1717–1729 (2004).
Borgomaneri, S., Vitale, F. & Avenanti, A. Early changes in corticospinal excitability when seeing fearful body expressions. Sci. Rep. 5, 14122 (2015).
Borgomaneri, S., Vitale, F. & Avenanti, A. Behavioral inhibition system sensitivity enhances motor cortex suppression when watching fearful body expressions. Brain Struct. Funct. 222, 3267–3282 (2017).
Borgomaneri, S., Vitale, F., Gazzola, V. & Avenanti, A. Seeing fearful body language rapidly freezes the observer’s motor cortex. Cortex 65, 232–245 (2015).
Di Lazzaro, V. et al. Chapter 12 Generation of I waves in the human: spinal recordings. Suppl. Clin. Neurophysiol. 56, 143–152 (2003).
Ziemann, U. et al. Demonstration of facilitatory I wave interaction in the human motor cortex by paired transcranial magnetic stimulation. J. Physiol. 511, 181–190 (1998).
Ziemann, U. I-waves in motor cortex revisited. Exp. Brain Res. 238, 1601–1610 (2020).
Borgomaneri, S., Battaglia, S., Sciamanna, G., Tortora, F. & Laricchiuta, D. Memories are not written in stone: Re-writing fear memories by means of non-invasive brain stimulation and optogenetic manipulations. Neurosci. Biobehav. Rev. 127, 334–352 (2021).
Battaglia, S., Nazzi, C., Fullana, M. A., di Pellegrino, G. & Borgomaneri, S. ‘Nip it in the bud’: low-frequency rTMS of the prefrontal cortex disrupts threat memory consolidation in humans. Behav. Res. Ther. 178, 104548 (2024).
Borgomaneri, S. et al. State-dependent TMS over prefrontal cortex disrupts fear-memory reconsolidation and prevents the return of fear. Curr. Biol. 30, 3672–3679.e4 (2020).
Robertson, E. M. Skill memory: mind the ever-decreasing gap for offline processing. Curr. Biol. 29, R287–R289 (2019).
Paparella, G. et al. Relationship between the interlimb transfer of a visuomotor learning task and interhemispheric inhibition in healthy humans. Cereb. Cortex 33, 7335–7346 (2023).
Berghuis, K. M. M., Semmler, J. G., Opie, G. M., Post, A. K. & Hortobágyi, T. Age-related changes in corticospinal excitability and intracortical inhibition after upper extremity motor learning: a systematic review and meta-analysis. Neurobiol. Aging 55, 61–71 (2017).
Heaney, C. F. & Kinney, J. W. Role of GABA(B) receptors in learning and memory and neurological disorders. Neurosci. Biobehav. Rev. 63, 1–28 (2016).
Vlachou, S. GABAB receptors and cognitive processing in health and disease. Curr. Top. Behav. Neurosci. 52, 291–329 (2021).
Kohl, M. M. & Paulsen, O. The roles of GABAB receptors in cortical network activity. Adv. Pharmacol. 58, 205–229 (2010).
Ho, K., Cirillo, J., Ren, A. & Byblow, W. D. Intracortical facilitation and inhibition in human primary motor cortex during motor skill acquisition. Exp. Brain Res. 240, 3289–3304 (2022).
Liepert, J., Classen, J., Cohen, L. G. & Hallett, M. Task-dependent changes of intracortical inhibition. Exp. Brain Res. 118, 421–426 (1998).
Smyth, C., Summers, J. J. & Garry, M. I. Differences in motor learning success are associated with differences in M1 excitability. Hum. Mov. Sci. 29, 618–630 (2010).
Lang, N. et al. Circadian modulation of GABA-mediated cortical inhibition. Cereb. Cortex 21, 2299–2306 (2011).
McDonnell, M. N., Orekhov, Y. & Ziemann, U. Suppression of LTP-like plasticity in human motor cortex by the GABA B receptor agonist baclofen. Exp. Brain Res. 180, 181–186 (2007).
Feldman, D. E. Synaptic mechanisms for plasticity in neocortex. Annu. Rev. Neurosci. 32, 33–55 (2009).
Carcea, I. & Froemke, R. C. Cortical plasticity, excitatory-inhibitory balance, and sensory perception. Prog. Brain Res. 207, 65–90 (2013).
Markram, H. et al. Interneurons of the neocortical inhibitory system. Nat. Rev. Neurosci. 5, 793–807 (2004).
Lim, S. & Goldman, M. S. Balanced cortical microcircuitry for maintaining information in working memory. Nat. Neurosci. 16, 1306–1314 (2013).
Rubin, R., Abbott, L. F. & Sompolinsky, H. Balanced excitation and inhibition are required for high-capacity, noise-robust neuronal selectivity. Proc. Natl Acad. Sci. USA. 114, E9366–E9375 (2017).
Eisenstein, T., Furman-Haran, E. & Tal, A. Increased cortical inhibition following brief motor memory reactivation supports reconsolidation and overnight offline learning gains. Proc. Natl Acad. Sci. 120, e2303985120 (2023).
Shibata, K. et al. Overlearning hyperstabilizes a skill by rapidly making neurochemical processing inhibitory-dominant. Nat. Neurosci. 20, 470–475 (2017).
Fiebig, F. & Lansner, A. A spiking working memory model based on Hebbian short-term potentiation. J. Neurosci. 37, 83–96 (2017).
Fiebig, F., Herman, P. & Lansner, A. An indexing theory for working memory based on fast Hebbian plasticity. eNeuro 7, 2 (2020).
Lansner, A., Fiebig, F. & Herman, P. Fast Hebbian plasticity and working memory. Curr. Opin. Neurobiol. 83, 102809 (2023).
Huang, Q. S. & Wei, H. A computational model of working memory based on spike-timing-dependent plasticity. Front. Comput. Neurosci. 15, 630999 (2021).
Masse, N. Y., Rosen, M. C. & Freedman, D. J. Reevaluating the role of persistent neural activity in short-term memory. Trends Cogn. Sci. 24, 242–258 (2020).
Ziemann, U., Ilić, T. V., Pauli, C., Meintzschel, F. & Ruge, D. Learning modifies subsequent induction of long-term potentiation-like and long-term depression-like plasticity in human motor cortex. J. Neurosci. 24, 1666–1672 (2004).
Rosenkranz, K., Kacar, A. & Rothwell, J. C. Differential modulation of motor cortical plasticity and excitability in early and late phases of human motor learning. J. Neurosci. 27, 12058–12066 (2007).
Stefan, K. et al. Temporary occlusion of associative motor cortical plasticity by prior dynamic motor training. Cereb. Cortex 16, 376–385 (2006).
Kuśmierz, Ł., Isomura, T. & Toyoizumi, T. Learning with three factors: modulating Hebbian plasticity with errors. Curr. Opin. Neurobiol. 46, 170–177 (2017).
Hwang, F. J. et al. Motor learning selectively strengthens cortical and striatal synapses of motor engram neurons. Neuron 110, 2790–2801.e5 (2022).
Runge, K., Cardoso, C. & de Chevigny, A. Dendritic spine plasticity: function and mechanisms. Front. Synaptic Neurosci. 12, 566347 (2020).
Trepel, C. & Racine, R. J. GABAergic modulation of neocortical long-term potentiation in the freely moving rat. Synapse 35, 120–128 (2000).
Castro-Alamancos, M. A., Donoghue, J. P. & Connors, B. W. Different forms of synaptic plasticity in somatosensory and motor areas of the neocortex. J. Neurosci. 15, 5324–5333 (1995).
Casarotto, A. et al. Mechanisms of Hebbian-like plasticity in the ventral premotor—primary motor network. J. Physiol. 601, 211–226 (2023).
Cuypers, K. & Marsman, A. Transcranial magnetic stimulation and magnetic resonance spectroscopy: ōpportunities for a bimodal approach in human neuroscience. Neuroimage 224, 117394 (2021).
Brashers-Krug, T., Shadmehr, R. & Bizzi, E. Consolidation in human motor memory. Nature 382, 252–255 (1996).
Krakauer, J. W., Ghez, C. & Ghilardi, M. F. Adaptation to visuomotor transformations: consolidation, interference, and forgetting. J. Neurosci. 25, 473–478 (2005).
Baraduc, P., Lang, N., Rothwell, J. C. & Wolpert, D. M. Consolidation of dynamic motor learning is not disrupted by rTMS of primary motor cortex. Curr. Biol. 14, 252–256 (2004).
Maruyama, S. et al. Cognitive control affects motor learning through local variations in GABA within the primary motor cortex. Sci. Rep. 11, 1–14 (2021).
Peters, A. J., Liu, H. & Komiyama, T. Learning in the rodent motor cortex. Annu. Rev. Neurosci. 40, 77–97 (2017).
Redecker, C., Wang, W., Fritschy, J.-M. & Witte, O. W. Widespread and long-lasting alterations in GABA(A)-receptor subtypes after focal cortical infarcts in rats: mediation by NMDA-dependent processes. J. Cereb. Blood Flow Metab. 22, 1463–1475 (2002).
Paparella, I., Vanderwalle, G., Stagg, C. J. & Maquet, P. An integrated measure of GABA to characterize post-stroke plasticity. NeuroImage. Clin. 39, 103463 (2023).
Loomes, M. et al. Is cortical inhibition in primary motor cortex related to executive control? Cortex 160, 100–114 (2023).
Erdfelder, E., FAul, F., Buchner, A. & Lang, A. G. Statistical power analyses using G*Power 3.1: tests for correlation and regression analyses. Behav. Res. Methods 41, 1149–1160 (2009).
Oldfield, R. C. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9, 97–113 (1971).
Rickard, T. C., Cai, D. J., Rieth, C. A., Jones, J. & Ard, M. C. Sleep does not enhance motor sequence learning. J. Exp. Psychol. Learn. Mem. Cogn. 34, 834–842 (2008).
Censor, N., Dayan, E. & Cohen, L. G. Cortico-subcortical neuronal circuitry associated with reconsolidation of human procedural memories. Cortex 58, 281–288 (2014).
Sigmon, K. & Davis, T. A. MATLAB Primer. Natick, MA, USA. https://doi.org/10.1201/9781420034950 (Chapman and Hall/CRC, 2004).
Brainard, D. H. The psychophysics toolbox. Spat. Vis. 10, 433–436 (1997).
Rossi, S., Hallett, M., Rossini, P. M. & Pascual-Leone, A. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 120, 2008–2039 (2009).
Rossini, P. M. et al. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application: an updated report from an I.F.C.N. Committee. Clin. Neurophysiol. 126, 1071–1107 (2015).
Truong, C. et al. Time of day and sleep effects on motor acquisition and consolidation. npj Sci. Learn. 8, 1–9 (2023).
Ilić, T. V. et al. Short-interval paired-pulse inhibition and facilitation of human motor cortex: the dimension of stimulus intensity. J. Physiol. 545, 153–167 (2002).
Di Lazzaro, V. et al. GABAA receptor subtype specific enhancement of inhibition in human motor cortex. J. Physiol. 575, 721–726 (2006).
McDonnell, M. N., Orekhov, Y. & Ziemann, U. The role of GABA(B) receptors in intracortical inhibition in the human motor cortex. Exp. brain Res. 173, 86–93 (2006).
Siebner, H. R. et al. Transcranial magnetic stimulation of the brain: what is stimulated?—A consensus and critical position paper. Clin. Neurophysiol. 140, 59–97 (2022).
Schwenkreis, P. et al. Riluzole suppresses motor cortex facilitation in correlation to its plasma level. Exp. Brain Res. 135, 293–299 (2000).
Ziemann, U., Chen, R., Cohen, L. G. & Hallett, M. Dextromethorphan decreases the excitability of the human motor cortex. Neurology 51, 1320–1324 (1998).
Vahdat, S., Fogel, S., Benali, H. & Doyon, J. Network-wide reorganization of procedural memory during NREM sleep revealed by fMRI. Elife 6, e24987 (2017).
Fioravante, D. & Regehr, W. G. Short-term forms of presynaptic plasticity. Curr. Opin. Neurobiol. 21, 269–274 (2011).
Walker, M. P., Brakefield, T., Hobson, J. A. & Stickgold, R. Dissociable stages of human memory consolidation and reconsolidation. Nature 425, 616–620 (2003). 2003 4256958.
Hardwicke, T. E., Taqi, M. & Shanks, D. R. Postretrieval new learning does not reliably induce human memory updating via reconsolidation. Proc. Natl Acad. Sci. USA 113, 5206–5211 (2016).
Johnson, B. P. et al. Generalization of procedural motor sequence learning after a single practice trial. npj Sci. Learn. 8, 1–8 (2023).
van Doorn, J. et al. The JASP guidelines for conducting and reporting a Bayesian analysis. Psychon. Bull. Rev. 28, 813–826 (2021).
Keysers, C., Gazzola, V. & Wagenmakers, E.-J. Using Bayes factor hypothesis testing in neuroscience to establish evidence of absence. Nat. Neurosci. 23, 788–799 (2020).
Cardellicchio, P., Dolfini, E. & D’Ausilio, A. The role of dorsal premotor cortex in joint action stopping. iScience 24, 103330 (2021).
Sanchez-Taltavull, D. et al. Bayesian correlation is a robust gene similarity measure for single-cell RNA-seq data. NAR Genomics Bioinforma. 2, lqaa002 (2020).
Diedenhofen, B. & Musch, J. cocor: A comprehensive solution for the statistical comparison of correlations. PLoS One 10, e0121945 (2015).
Hittner, J. B., May, K. & Silver, N. C. A Monte Carlo evaluation of tests for comparing dependent correlations. J. Gen. Psychol. 130, 149–168 (2003).
Dunn, O. J. & Clark, V. Correlation coefficients measured on the same individuals. J. Am. Stat. Assoc. 64, 366–377 (1969).
Acknowledgements
Sara Borgomaneri was supported by Next Generation EU (NGEU) and funded by the Ministry of University and Research (MUR), National Recovery and Resilience Plan (NRRP) PRIN 2022 (grant No. 2022XKZBFC—CUP J53D23008340001): The influence of emotions on action control: brain network plasticity and potential trans-diagnostic applications (D DN. 104 02.02.2022) and Bial Foundation, Portugal (033/22). This work was supported by the Italian Ministry of Health (Ricerca Corrente). The views and opinions expressed are solely those of the authors and do not necessarily reflect those of the European Union, nor can the European Union be held responsible for them.
Author information
Authors and Affiliations
Contributions
P.C. designed the experiment. P.C. recorded and analyzed the data. P.C. and S.B. wrote the paper. All authors read and approved the final paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Cardellicchio, P., Borgomaneri, S. Level of M1 GABAB predicts micro offline consolidation of motor learning during wakefulness. npj Sci. Learn. 10, 10 (2025). https://doi.org/10.1038/s41539-025-00299-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41539-025-00299-1
