
Abstract
Inflammation is critical for controlling infections but can cause disease when unchecked. During Mycobacterium tuberculosis (Mtb) infection, neutrophil-dominated inflammation is associated with exacerbated disease. ATG5 expression by neutrophils mediates autophagy-independent control of infection but mechanistic understanding of how this regulates protective neutrophil function is lacking. Using genetic mouse models along with in vivo and in vitro infection systems, we report herein that ATG5 is required in neutrophils to suppress type I interferon-induced PAD4-mediated histone citrullination and neutrophil extracellular trap (NET) release. In addition, ATG5 suppresses type I interferon-induced CXCL2 secretion and neutrophil swarming during Mtb infection. Elevated type I IFN signalling and NET release contribute to the early susceptibility of Atg5fl/fl-LysM-Cre mice during infection. These findings identify ATG5 as a master regulator of how type I interferon influences neutrophil responses during infection, revealing a potential target for host-directed therapies.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others


Data availability
The raw RNA sequencing data have been deposited in the Gene Expression Omnibus under accession number GSE280193. Fold differences of transcripts of interest are in Supplementary Tables 1 and 2. Source files for the western blot uncropped raw data are in the Source Data Files Section of the Supplementary Information.
References
Global Tuberculosis Report (World Health Organization, 2024).
Condos, R., Rom, W. N., Liu, Y. M. & Schluger, N. W. Local immune responses correlate with presentation and outcome in tuberculosis. Am. J. Respir. Crit. Care Med. 157, 729–735 (1998).
Han, Y. et al. High blood neutrophil-lymphocyte ratio associated with poor outcomes in miliary tuberculosis. J. Thorac. Dis. 10, 339–346 (2018).
Eum, S.-Y. et al. Neutrophils are the predominant infected phagocytic cells in the airways of patients with active pulmonary TB. Chest 137, 122–128 (2010).
Berry, M. P. R. et al. An interferon-inducible neutrophil-driven blood transcriptional signature in human tuberculosis. Nature 466, 973–977 (2010).
Moreira-Teixeira, L. et al. Type I IFN exacerbates disease in tuberculosis-susceptible mice by inducing neutrophil-mediated lung inflammation and NETosis. Nat. Commun. 11, 5566 (2020).
Kimmey, J. M. et al. Unique role for ATG5 in neutrophil-mediated immunopathology during M. tuberculosis infection. Nature 528, 565–569 (2015).
Muefong, C. N. & Sutherland, J. S. Neutrophils in tuberculosis-associated inflammation and lung pathology. Front. Immunol. 11, 962 (2020).
Nair, S. et al. Irg1 expression in myeloid cells prevents immunopathology during M. tuberculosis infection. J. Exp. Med. 215, 1035–1045 (2018).
Mishra, B. B. et al. Nitric oxide prevents a pathogen-permissive granulocytic inflammation during tuberculosis. Nat. Microbiol. 2, 17072 (2017).
Castillo, E. F. et al. Autophagy protects against active tuberculosis by suppressing bacterial burden and inflammation. Proc. Natl Acad. Sci. USA 109, E3168–E3176 (2012).
Watson, R. O., Manzanillo, P. S. & Cox, J. S. Extracellular M. tuberculosis DNA targets bacteria for autophagy by activating the host DNA-sensing pathway. Cell 150, 803–815 (2012).
Golovkine, G. R. et al. Autophagy restricts Mycobacterium tuberculosis during acute infection in mice. Nat. Microbiol. https://doi.org/10.1038/s41564-023-01354-6 (2023).
Kinsella, R. L. et al. Autophagy prevents early proinflammatory responses and neutrophil recruitment during Mycobacterium tuberculosis infection without affecting pathogen burden in macrophages. PLoS Biol. 21, e3002159 (2023).
Kim, K. H. & Lee, M.-S. Autophagy—a key player in cellular and body metabolism. Nat. Rev. Endocrinol. 10, 322–337 (2014).
Lahiri, V., Hawkins, W. D. & Klionsky, D. J. Watch what you (self-) eat: autophagic mechanisms that modulate metabolism. Cell Metab. 29, 803–826 (2019).
Liu, K., Sutter, B. M. & Tu, B. P. Autophagy sustains glutamate and aspartate synthesis in Saccharomyces cerevisiae during nitrogen starvation. Nat. Commun. 12, 57 (2021).
Onodera, J. & Ohsumi, Y. Autophagy is required for maintenance of amino acid levels and protein synthesis under nitrogen starvation. J. Biol. Chem. 280, 31582–31586 (2005).
Yang, Z. & Klionsky, D. J. Permeases recycle amino acids resulting from autophagy. Autophagy 3, 149–150 (2007).
Kinsella, R. et al. Perspectives and advances in the understanding of tuberculosis. Annu. Rev. Pathol. 16, 377–408 (2021).
Manca, C. et al. Hypervirulent M. tuberculosis W/Beijing strains upregulate type I IFNs and increase expression of negative regulators of the Jak-Stat pathway. J. Interferon Cytokine Res. 25, 694–701 (2005).
Dorhoi, A. et al. Type I IFN signaling triggers immunopathology in tuberculosis‐susceptible mice by modulating lung phagocyte dynamics. Eur. J. Immunol. 44, 2380–2393 (2014).
Kimmey, J. M. et al. The impact of ISGylation during Mycobacterium tuberculosis infection in mice. Microbes Infect. 19, 249–258 (2017).
Naik, S. K. et al. Type I IFN signaling in the absence of IRGM1 promotes M. tuberculosis replication in immune cells by suppressing T cell responses. Mucosal Immunol. https://doi.org/10.1016/j.mucimm.2024.07.002 (2024).
Huynh, J. P. et al. Bhlhe40 is an essential repressor of IL-10 during Mycobacterium tuberculosis infection. J. Exp. Med. 215, 1823–1838 (2018).
Poli, V. & Zanoni, I. Neutrophil intrinsic and extrinsic regulation of NETosis in health and disease. Trends Microbiol. https://doi.org/10.1016/j.tim.2022.10.002 (2022).
Pylaeva, E., Lang, S. & Jablonska, J. The essential role of type I interferons in differentiation and activation of tumor-associated neutrophils. Front. Immunol. 7, 629 (2016).
Pylaeva, E. et al. Detrimental effect of type I IFNs during acute lung infection with Pseudomonas aeruginosa is mediated through the stimulation of neutrophil NETosis. Front. Immunol. 10, 2190 (2019).
Gul, E. et al. Type I IFN–related NETosis in ataxia telangiectasia and Artemis deficiency. J. Allergy Clin. Immunol. 142, 246–257 (2018).
Garcia-Romo, G. S. et al. Netting neutrophils are major inducers of type I IFN production in pediatric systemic lupus erythematosus. Sci. Transl. Med. 3, 73ra20 (2011).
Chowdhury, C. S. et al. Type I IFN-mediated NET release promotes Mycobacterium tuberculosis replication and is associated with granuloma caseation. Cell Host Microbe 32, 2092–2111.e7 (2024).
Thiam, H. R. et al. NETosis proceeds by cytoskeleton and endomembrane disassembly and PAD4-mediated chromatin decondensation and nuclear envelope rupture. Proc. Natl Acad. Sci. USA 117, 7326–7337 (2020).
Filio-Rodríguez, G. et al. In vivo induction of neutrophil extracellular traps by Mycobacterium tuberculosis in a guinea pig model. Innate Immun. 23, 625–637 (2017).
Kotov, D. I. et al. Early cellular mechanisms of type I interferon-driven susceptibility to tuberculosis. Cell 186, 5536–5553.e22 (2023).
Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 18, 134–147 (2018).
Branzk, N. et al. Neutrophils sense microbe size and selectively release neutrophil extracellular traps in response to large pathogens. Nat. Immunol. 15, 1017–1025 (2014).
Wang, F. et al. ATG5 provides host protection acting as a switch in the atg8ylation cascade between autophagy and secretion. Dev. Cell https://doi.org/10.1016/j.devcel.2023.03.014 (2023).
Tatsiy, O. & McDonald, P. P. Physiological stimuli induce PAD4-dependent, ROS-independent NETosis, with early and late events controlled by discrete signaling pathways. Front. Immunol. 9, 2036 (2018).
Lewis, H. D. et al. Inhibition of PAD4 activity is sufficient to disrupt mouse and human NET formation. Nat. Chem. Biol. 11, 189–191 (2015).
Darrah, E., Rosen, A., Giles, J. T. & Andrade, F. Peptidylarginine deiminase 2, 3 and 4 have distinct specificities against cellular substrates: novel insights into autoantigen selection in rheumatoid arthritis. Ann. Rheum. Dis. 71, 92–98 (2012).
Su, R. et al. Mycobacterium tuberculosis infection induces low-density granulocyte generation by promoting neutrophil extracellular trap formation via ROS pathway. Front. Microbiol. 10, 1468 (2019).
Parker, H., Dragunow, M., Hampton, M. B., Kettle, A. J. & Winterbourn, C. C. Requirements for NADPH oxidase and myeloperoxidase in neutrophil extracellular trap formation differ depending on the stimulus. J. Leukoc. Biol. 92, 841–849 (2012).
Neeli, I., Khan, S. N. & Radic, M. Histone deimination as a response to inflammatory stimuli in neutrophils. J. Immunol. 180, 1895–1902 (2008).
Pieterse, E., Rother, N., Yanginlar, C., Hilbrands, L. B. & van der Vlag, J. Neutrophils discriminate between lipopolysaccharides of different bacterial sources and selectively release neutrophil extracellular traps. Front. Immunol. 7, 484 (2016).
Yipp, B. G. et al. Infection-induced NETosis is a dynamic process involving neutrophil multitasking in vivo. Nat. Med. 18, 1386–1393 (2012).
Fuchs, T. A. et al. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 176, 231–241 (2007).
Song, Z. et al. NADPH oxidase controls pulmonary neutrophil infiltration in the response to fungal cell walls by limiting LTB4. Blood 135, 891–903 (2020).
Hopke, A. et al. Neutrophil swarming delays the growth of clusters of pathogenic fungi. Nat. Commun. 11, 2031 (2020).
Lämmermann, T. et al. Neutrophil swarms require LTB4 and integrins at sites of cell death in vivo. Nature 498, 371–375 (2013).
Subramanian, B. C., Majumdar, R. & Parent, C. A. The role of the LTB4-BLT1 axis in chemotactic gradient sensing and directed leukocyte migration. Semin. Immunol. 33, 16–29 (2017).
Afonso, P. V. et al. LTB4 is a signal-relay molecule during neutrophil chemotaxis. Dev. Cell 22, 1079–1091 (2012).
Sawant, K. V. et al. Chemokine CXCL1-mediated neutrophil trafficking in the lung: role of CXCR2 activation. J. Innate Immun. 7, 647–658 (2015).
Mihlan, M., Glaser, K. M., Epple, M. W. & Lämmermann, T. Neutrophils: amoeboid migration and swarming dynamics in tissues. Front. Cell Dev. Biol. 10, 871789 (2022).
Poplimont, H. et al. Neutrophil swarming in damaged tissue is orchestrated by connexins and cooperative calcium alarm signals. Curr. Biol. 30, 2761–2776.e7 (2020).
Vieira, S. et al. A crucial role for TNF-α in mediating neutrophil influx induced by endogenously generated or exogenous chemokines, KC/CXCL1 and LIX/CXCL5. Br. J. Pharmacol. 158, 779–789 (2009).
Chou, R. C. et al. Lipid-cytokine-chemokine cascade drives neutrophil recruitment in a murine model of inflammatory arthritis. Immunity 33, 266–278 (2010).
Kienle, K. et al. Neutrophils self-limit swarming to contain bacterial growth in vivo. Science 372, eabe7729 (2021).
Brandt, S. L. et al. Excessive localized leukotriene B4 levels dictate poor skin host defense in diabetic mice. JCI Insight 3, e120220 (2018).
Carter, G. W. et al. 5-lipoxygenase inhibitory activity of zileuton. J. Pharmacol. Exp. Ther. 256, 929–937 (1991).
Lv, J. et al. BLT1 mediates bleomycin-induced lung fibrosis independently of neutrophils and CD4 + T cells. J. Immunol. 198, 1673–1684 (2017).
Isles, H. M. et al. Pioneer neutrophils release chromatin within in vivo swarms. eLife 10, e68755 (2021).
Apel, F. et al. The cytosolic DNA sensor cGAS recognizes neutrophil extracellular traps. Sci. Signal. 14, eaax7942 (2021).
Glennon-Alty, L., Moots, R. J., Edwards, S. W. & Wright, H. L. Type I interferon regulates cytokine-delayed neutrophil apoptosis, reactive oxygen species production and chemokine expression. Clin. Exp. Immunol. 203, 151–159 (2021).
Mutua, V. & Gershwin, L. J. A review of neutrophil extracellular traps (NETs) in disease: potential anti-NETs therapeutics. Clin. Rev. Allergy Immunol. 61, 194–211 (2021).
Pham, D. L. et al. Neutrophil autophagy and extracellular DNA traps contribute to airway inflammation in severe asthma. Clin. Exp. Allergy 47, 57–70 (2017).
Veras, F. P. et al. SARS-CoV-2–triggered neutrophil extracellular traps mediate COVID-19 pathology. J. Exp. Med. 217, e20201129 (2020).
Pulavendran, S. et al. Production of neutrophil extracellular traps contributes to the pathogenesis of Francisella tularemia. Front. Immunol. 11, 679 (2020).
Zhu, S. et al. Neutrophil extracellular traps contribute to immunothrombosis formation via the STING pathway in sepsis-associated lung injury. Cell Death Discov. 9, 315 (2023).
Sorgi, C. A. et al. Eicosanoid pathway on host resistance and inflammation during Mycobacterium tuberculosis infection is comprised by LTB4 reduction but not PGE2 increment. Biochim. Biophys. Acta 1866, 165574 (2020).
Mayer-Barber, K. D. et al. Host-directed therapy of tuberculosis based on interleukin-1 and type I interferon crosstalk. Nature 511, 99–103 (2014).
Zhao, Z. et al. Autophagosome-independent essential function for the autophagy protein Atg5 in cellular immunity to intracellular pathogens. Cell Host Microbe 4, 458–469 (2008).
Hwang, S. et al. Nondegradative role of Atg5-Atg12/ Atg16L1 autophagy protein complex in antiviral activity of interferon gamma. Cell Host Microbe 11, 397–409 (2012).
Hara, T. et al. Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 441, 885–889 (2006).
Gawriluk, T. R. et al. Autophagy is a cell survival program for female germ cells in the murine ovary. Reproduction 141, 759–765 (2011).
Malhotra, R., Warne, J. P., Salas, E., Xu, A. W. & Debnath, J. Loss of Atg12, but not Atg5, in pro-opiomelanocortin neurons exacerbates diet-induced obesity. Autophagy 11, 145–154 (2015).
Steed, A. L. et al. The microbial metabolite desaminotyrosine protects from influenza through type I interferon. Science 357, 498–502 (2017).
Aronesty, E. Comparison of sequencing utility programs. Open Bioinform. J. 7, 1–8 (2013).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Patro, R., Duggal, G., Love, M. I., Irizarry, R. A. & Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 14, 417–419 (2017).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Liberzon, A. et al. The molecular signatures database hallmark gene set collection. Cell Syst. 1, 417–425 (2015).
Gillespie, M. et al. The reactome pathway knowledgebase 2022. Nucleic Acids Res. 50, D687–D692 (2022).
Mócsai, A., Zhou, M., Meng, F., Tybulewicz, V. L. & Lowell, C. A. Syk is required for integrin signaling in neutrophils. Immunity 16, 547–558 (2002).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Steger, C. An unbiased detector of curvilinear structures. IEEE Trans. Pattern Anal. Mach. Intell. 20, 113–125 (1998).
Najmeh, S., Cools-Lartigue, J., Giannias, B., Spicer, J. & Ferri, L. E. Simplified human neutrophil extracellular traps (NETs) isolation and handling. J. Vis. Exp. https://doi.org/10.3791/52687 (2015).
Acknowledgements
This work was supported by NIH grants (R01 AI179024, R01 AI184666, R01 AI132697 and U19 AI142784), a Burroughs Wellcome Fund Investigators in the Pathogenesis of Infectious Disease Award, and the Philip and Sima Needleman Center for Autophagy Therapeutics and Research to C.L.S., a Potts Memorial Foundation postdoctoral fellowship to R.L.K., and a National Science Foundation Graduate Research Fellowship DGE-1143954 and the NIGMS Cell and Molecular Biology Training Grant GM007067 to J.M.K. We acknowledge the assistance of S. Sviben and J. Fitzpatrick at the Washington University Center for Cellular Imaging (WUCCI) in electron microscopy studies, which is supported by the Washington University School of Medicine, The Children’s Discovery Institute of Washington University and St Louis Children’s Hospital (CDI-CORE-2015-505 and CDI-CORE-2019-813), and the Foundation for Barnes-Jewish Hospital (3770). We also thank the Genome Technology Access Center at the McDonnell Genome Institute at Washington University School of Medicine for RNA sequencing and analysis. The Center is partially supported by NCI Cancer Center Support Grant P30 CA91842 to the Siteman Cancer Center and by ICTS/CTSA Grant UL1TR002345 from the National Center for Research Resources (NCRR), a component of the National Institutes of Health (NIH), and NIH Roadmap for Medical Research. Atg5fl/fl, Becn1fl/fl, Atg16l1fl/fl, Atg12fl/fl and Ifnar1−/− mice were kindly provided by N. Mizushima (The University of Tokyo), E. Rucker (University of Kentucky), S. Virgin (Washington University School of Medicine), J. Debnath (UCSF) and A. Steed (Washington University School of Medicine), respectively. This publication is solely the responsibility of the authors and does not necessarily represent the official view of NCRR or NIH. The model in Extended Data Fig. 7 was created with BioRender.com.
Author information
Authors and Affiliations
Contributions
R.L.K. and C.L.S. wrote the manuscript. Experiments were designed and performed by R.L.K., C.S.C., A.S. and J.M.K., with assistance from S.R.M. and A.P. under the guidance of C.L.S. RNA sequencing data were analysed by Y.M., E.E. and M.N.A. Mice were bred and maintained by D.K. Funding and direction were provided by C.L.S.
Corresponding authors
Ethics declarations
Competing interests
All authors declare no competing interests.
Peer review
Peer review information
Nature Microbiology thanks Lu Huang and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Loss of type I IFN signaling in Atg5fl/fl-LysM-Cre mice alters lesion architecture and neutrophil distribution in the lung without affecting bacterial burdens or neutrophil accumulation.
(A) Median fluorescence intensity of IFNAR1 on neutrophils in the lung at 14 dpi with Mtb quantified by flow cytometry. (B) The amount of IFNβ in the lungs from GFP-Mtb infected mice at 14 dpi quantified by ELISA. (C-L) Immune cell abundance in the lungs of GFP-Mtb infected mice at 14 or 21 dpi determined by flow cytometry. Each datapoint represents one biological replicate from at least 2 independent experiments. (M) Additional representative H&E-stained lung sections from GFP-Mtb infected mice at 21 dpi. Scale bar is 1 mm. (N) Additional representative merged tiled confocal immunofluorescence images of lung sections from GFP-Mtb infected mice at 21 dpi that were probed with antibodies to detect citrullinated histone H3 (H3Cit; red), Ly6G (neutrophil marker; cyan), and DNA (Hoechst; blue). GFP-Mtb is shown in green. Scale bar is 1 mm. (O) Representative confocal immunofluorescence images of lung sections at 60X magnification from GFP-Mtb infected mice at 14 dpi that were probed with antibodies to detect Ly6G (neutrophil marker; cyan), and DNA (Hoechst; blue). GFP-Mtb is also shown. Scale bar is 50μm. (P) The average size of contiguous Ly6G+ regions in lung tissue sections at 14 dpi. Each data point is from one field of view where data from 4 mice from at least two independent infection experiments were compiled. (Q) The proportion of Ly6G+ pixel area across complete lung sections (Fig. 1j) determined by Fiji analysis of immunofluorescence histology images of lung sections from mice infected with GFP-Mtb for 21 dpi. Each data point is from one lung section from 3 mice from at least 2 independent experiments. All graphs report the mean ± SD. Statistical differences were determined by an unpaired two-tailed student t-test (A-B, P) or one-way ANOVA and Šídák multiple comparison test (C-L, Q) (only comparing within a single timepoint). * P < 0.05, ** P < 0.01, and **** P < 0.0001. Differences that are not statistically significant are designated as ns. Complete statistical analysis including the number of samples used is in Supplementary Table 4.
Extended Data Fig. 2 ATG5 functions independent of autophagy to suppress NET release during M. tuberculosis infection in vivo and in vitro.
(A) Representative confocal immunofluorescence images of lung sections at 60X magnification from GFP-Mtb infected mice at 14 dpi that were probed with antibodies to detect citrullinated histone H3 (H3Cit; red), Ly6G (neutrophil marker; cyan), and DNA (Hoechst; blue). GFP-Mtb is also shown. Scale bar is 50μm. (B) The % of H3Cit+ pixel area per field and (C) the percent of Ly6G+ pixel area that was H3Cit+ per lung section in confocal immunofluorescence images of lung sections from GFP-Mtb infected mice at 14 dpi. (B-C) Each data point is from one field of view from 4 mice from at least two separate infection experiments. (D) The proportion of H3Cit+ neutrophils in the lungs of GFP-Mtb infected mice at 21 dpi that are GFP-Mtb- (bystander) and GFP-Mtb+ determined by flow cytometry. Each data point is from one mouse, compiled from 2 independent experiments. (E) Representative SEM images of mouse neutrophils infected with GFP-Mtb in vitro at 3000x magnification. Yellow arrows indicate NETs being released from cells. Scale bar is 10μm. (F) The number of NETs released per 10 cells quantified from SEM images at 1000X magnification of neutrophils infected with GFP-Mtb for 4 or 18 hpi. Each datapoint represents one field with 3 fields containing 10-40 cells/field compiled from 3 independent experiments. (G) Proportion of neutrophils infected in vitro with GFP-Mtb that are GFP-Mtb+ at 4 hpi, and (H) GFP-Mtb MFI in GFP-Mtb+ neutrophils at 4 hpi measured by flow cytometry. (I) The proportion of GFP-Mtb- (bystander) and GFP-Mtb+ neutrophils from GFP-Mtb infected neutrophil cultures at 4 hpi that are H3Cit+ as determined by flow cytometry. (J) Representative confocal immunofluorescence microscopy images of neutrophils mock treated in vitro for 4 or 18 h and stained for H3Cit (red), neutrophil marker myeloperoxidase (MPO), and DNA (Hoechst, blue) (60X magnification). Scale bar is 50μm. (K) The proportion of Hoechst+ cells that were H3Cit+ and (L) the number of extracellular H3Cit+ NETs released per 100 cells after mock-infection of neutrophils for 4 or 18 h, as quantified from fluorescence microscopy experiments. (M) Representative confocal immunofluorescence microscopy images of neutrophils infected in vitro with GFP-Mtb for 4 or 18 h and stained for H3Cit (red) and DNA (Hoechst, blue) (60X magnification). Scale bar is 50μm. (N) The proportion of Hoechst+ cells that were H3Cit+ and (O) the number of extracellular H3Cit+ NETs released per 100 cells after GFP-Mtb infection of neutrophils for 4 or 18 h, as quantified from fluorescence microscopy experiments. (P-R) Assessment of single copy LysM-Cre mediated Cre-Lox recombination efficiency of Atg5, Becn1 and Atg16l1 in bone marrow neutrophils from 3 biological replicates reported as percent recombination by calculating the percentage of the deletion PCR product intensity over the combined intensity of the floxed product and the deleted product. (P) Samples are from Atg5fl/fl and Atg5fl/fl-LysM-Cre mice, where Atg5fl/fl-LysM-Cre neutrophils exhibit 96.30%, 94.91% and 97.05% recombination efficiency. (Q) Samples are from Becn1fl/fl and Becn1fl/fl-LysM-Cre mice, where Becn1fl/fl-LysM-Cre neutrophils exhibit 94.54%, 90.02% and 95.32% recombination efficiency. (R) Samples are from Atg16l1fl/fl and Atg16l1fl/fl-LysM-Cre mice, where Atg16l1fl/fl-LysM-Cre neutrophils exhibit 94.07%, 93.33% and 92.93% recombination efficiency. (S) Mtb CFU recovered from cultures of neutrophils infected with GFP-Mtb in vitro for 4 or 18 hpi, reported as a ratio to 0 hr CFU. Each datapoint is from an independent well of infected cells compiled from at least 2 independent experiments. (T) Kaplan Meier curve of survival proportions during Mtb infection of mice. The median survival time for each genotype is reported. (U) Log transformed graph of the colony forming units (CFU) in the right lung of mice at 21 dpi. (V) Proportion of CD45+ cells that are neutrophils (Ly6G+ CD11b+) in the lungs of mice at 21 dpi. (W) Proportion of neutrophils (CD45+ Ly6G+ CD11b+) infected in vitro with GFP-Mtb at 4 hpi that are positive for surface staining for the primary granule marker CD63, and (X) the MFI of CD63 on CD63+ neutrophils at 4 hpi determined by flow cytometry. Each datapoint represents one biological replicate compiled from 2 independent experiments. All graphs report the mean ± SD. Statistical differences were determined by an unpaired two-tailed student t-test (B-C, F-H, K-L, W-X), one-way ANOVA and Šídák multiple comparison test (I, N-O, S, U-V) or a log-rank Mantel-Cox test (T). * P < 0.05, ** P < 0.01, and **** P < 0.0001. Differences that are not statistically significant are designated as ns. Complete statistical analysis including the number of samples used is in Supplementary Table 4.
Extended Data Fig. 3 ATG5 suppresses type I IFN-induced PAD4-mediated histone citrullination during M. tuberculosis infection.
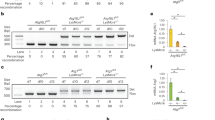
(A, D) Mtb CFU recovered from cultures of mouse bone marrow neutrophils infected with GFP-Mtb in vitro for 4 and 18 hpi. Each datapoint is from a single well compiled from at least 2 independent experiments. (B) Proportion of neutrophils (CD45+ Ly6G+ CD11b+) infected in vitro with GFP-Mtb that are GFP-Mtb+, and the GFP-Mtb MFI in infected neutrophils (C) at 4 hpi determined by flow cytometry. (E) Representative confocal immunofluorescence microscopy images at 60X magnification of neutrophils treated with ionomycin for 4 or 18 h and stained for DNA (Hoechst), H3Cit, and neutrophil marker MPO. Scale bar is 50μm. (F) The proportion of Hoechst+ cells that are H3Cit+ and (G) the number of H3Cit+ NETs released per 100 cells from neutrophils treated with ionomycin for 4 or 18 h, based on fluorescence microscopy experiments. (F-G) Each datapoint represents data from one field, with a minimum of 6 fields containing 40-150 cells/field each compiled from at least 2 independent experiments. (H) The proportion of viable neutrophils (CD45+, Ly6G+, CD11b+, Zombie-) in the lungs of GFP-Mtb infected mice at 21 dpi determined by flow cytometry. (I) The proportion of neutrophils in the lungs of GFP-Mtb infected mice at 21 dpi that are H3Cit+ split into Zombie-viability dye positive and Zombie-negative determined by flow cytometry. There is no significant difference between the proportion of Zombie+ H3Cit+ neutrophils in the lungs of Atg5fl/fl and Atg5fl/fl-LysM-Cre mice. There are significantly more Zombie- H3Cit+ neutrophils in the lungs of Atg5fl/fl-LysM-Cre mice relative to Atg5fl/fl controls (P < 0.05). (J) The proportion of GFP-Mtb infected neutrophils that are H3Cit+ at 4 hpi split into Zombie-viability dye positive and Zombie-negative determined by flow cytometry. There are significantly more Atg5fl/fl-LysM-Cre Zombie+ H3Cit+ neutrophils than Atg5fl/fl (P < 0.05). There are significantly more Atg5fl/fl-LysM-Cre Zombie- H3Cit+ neutrophils relative to Atg5fl/fl controls (P < 0.01). Agarose gel analysis of PCR products targeting nuclear or mitochondrial genes, amplified from extracellular DNA released by Atg5fl/fl and Atg5fl/fl-LysM-Cre neutrophils after Mtb infection (K), ionomycin treatment (K), or mock infection (L). All graphs report the mean ± SD. Statistical differences were determined by one-way ANOVA and Šídák multiple comparison test other than (H) where an unpaired two-tailed student t-test was performed. * P < 0.05, ** P < 0.01. Differences that are not statistically significant are designated as ns. Complete statistical analysis including the number of samples used is in Supplementary Table 4.
Extended Data Fig. 4 ATG5 mediates protection during M. tuberculosis infection by restricting PAD4-dependent NET release.
(A-L) Flow cytometry quantification of immune cell abundance in the lungs of Mtb infected mice at 21 dpi. Each datapoint represents one biological replicate from at least 2 independent experiments. Open blue squares, Atg5fl/fl; closed blue squares, Atg5fl/fl-LysM-Cre; open pink squares, Padi4−/−/Atg5fl/fl; closed pink squares, Padi4−/−/Atg5fl/fl-LysM-Cre. (J) Additional representative H&E-stained lung sections from GFP-Mtb infected mice at 21 dpi. Scale bar is 500μm. (K) Additional representative merged tiled confocal immunofluorescence images at 10X magnification of lung sections from GFP-Mtb infected mice at 21 dpi that were probed with antibodies to detect citrullinated histone H3 (H3Cit; red), Ly6G (neutrophil marker; cyan), and DNA (Hoechst; blue). GFP-Mtb is shown in green. Scale bar is 1 mm. All graphs report the mean ± SD. Statistical differences were determined by or one-way ANOVA and Šídák multiple comparison test. * P < 0.05, ** P < 0.01. Differences that are not statistically significant are designated as ns. Complete statistical analysis including the number of samples used is in Supplementary Table 4.
Extended Data Fig. 5 ATG5 suppresses type I IFN-induced neutrophil swarming independently from the regulation of NET release.
The Fiji particle analysis parameters used to measure the average size of neutrophil aggregates. The average neutrophil aggregate size at 4 hpi of mock treated mouse neutrophils based on fluorescence microscopy (n = 8 for each genotype). Each datapoint represents the average neutrophil aggregate size per field, with a minimum of 8 fields containing 100-250 cells/field each compiled from at least 2 independent experiments. All graphs report the mean ± SD. Statistical differences were determined by an unpaired two-tailed student t-test. Differences that are not statistically significant are designated as ns. Complete statistical analysis including the number of samples used is in Supplementary Table 4.
Extended Data Fig. 6 Model of ATG5 activity in neutrophils.
Model depicting how ATG5 functions in neutrophils to regulate type I IFN-induced NET release and swarming. Mtb infection in neutrophils activates type I IFN signaling and PAD4-mediated histone citrullination, where type I IFN signaling promotes the extracellular release of NETs3. ATG5 functions downstream of type I IFN signaling to suppress type I IFN-induced PAD4-mediated histone citrullination and CXCL2 production during Mtb infection. ATG5 also limits histone citrullination in bystander neutrophils in culture with GFP-Mtb+ neutrophils. Created with BioRender.com.
Extended Data Fig. 7 Flow cytometry assessment of neutrophil purity following anti-Ly6G selection or 62% percoll enrichment and flow cytometry gating strategies to identify immune cells in the lung.
(A) The gating strategy used to identify neutrophils (GR1+Ly6G+CD11b+) after anti-Ly6G enrichment from the lung at 14 dpi. (B) The proportion of single cells that are neutrophils from Atg5fl/fl and Atg5fl/fl-LysM-Cre mice after anti-Ly6G enrichment from the lung at 14 dpi. Bar graphs report the mean ± SD where each data point is from an individual mouse. (C) The gating strategy used to identify immune cells in the lung during Mtb infection at 14 or 21 dpi. The Lymphoid markers were only included at 21 dpi. Also depicted is the gating used to assess GFP-Mtb positivity, H3Cit-positivity and Zombie viability dye positivity of neutrophils in the lungs of mice infected with GFP-Mtb at 21 dpi. (D) Representative confocal immunofluorescence microscopy images at 60x magnification of lung sections from GFP-Mtb infected Atg5fl/fl-LysM-Cre mice at 21 dpi probed with antibodies to detect citrullinated histone H3 (H3Cit; red), Ly6G (neutrophil marker; cyan), and DNA (Hoechst; blue). GFP-Mtb is also shown. A consecutive section was stained with Hoechst and probed with anti-Ly6G, and anti-rabbit IgG-AF555 antibodies but was not probed with the primary anti-H3Cit antibody. Scale bar is 50μm. (E) The gating strategy used to assess neutrophil purity and identify contaminating cell types following 62% percoll gradient enrichment from the bone marrow, where most cells are neutrophils and B cells are the most abundant contaminating cell. Also depicted is the gating used to assess GFP-Mtb positivity, H3Cit-positivity and Zombie viability dye positivity of neutrophils following in vitro infection with GFP-Mtb. (F) Neutrophil purity in bone marrow neutrophil isolations using 62% percoll assessed by flow cytometry. (G) Representative confocal microscopy image of bone marrow neutrophils stained with Hoechst to reveal nuclear morphology (20x magnification). Scale bar is 50 μm.
Supplementary information
Supplementary Information
Supplementary information protocols, legends for supplementary tables and source data of gel images.
Supplementary Tables 1–4
1: Differentially expressed genes in neutrophils isolated from the lungs of Atg5fl/fl-LysM-Cre and Atg5fl/fl mice at 14 dpi. 2: Differentially expressed genes in the type I IFN and RIG-I signalling pathways identified by GSEA analysis of transcripts from neutrophils isolated from the lungs of Atg5fl/fl-LysM-Cre relative to Atg5fl/fl mice at 14 dpi. 3: Primer sequences used to amplify nuclear and mitochondrial genes from extracellular DNA released by neutrophils. 4: Statistical comparisons from all graphed data, exported from GraphPad Prism.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kinsella, R.L., Sur Chowdhury, C., Smirnov, A. et al. ATG5 suppresses type I IFN-dependent neutrophil effector functions during Mycobacterium tuberculosis infection in mice. Nat Microbiol 10, 1323–1339 (2025). https://doi.org/10.1038/s41564-025-01988-8
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41564-025-01988-8
