
Abstract
Membrane contact sites are hubs for interorganellar lipid transport within eukaryotic cells. As a principal tether bridging the endoplasmic reticulum (ER) and the plasma membrane in Saccharomyces cerevisiae, the protein IST2 has a major role during lipid transport between both compartments. Here, we show a comprehensive investigation elucidating the structural and mechanistic properties of IST2 and its interaction with the soluble lipid transfer protein OSH6. The ER-embedded transmembrane domain of IST2 is homologous to the TMEM16 family and acts as a constitutively active lipid scramblase. The extended C terminus binds to the plasma membrane and the phosphatidylserine–phosphatidylinositol 4-phosphate exchanger OSH6. Through cellular growth assays and biochemical and structural studies, we characterized the interaction between both proteins and show that OSH6 remains associated with IST2 during lipid shuttling between membranes. These results highlight the role of the IST2–OSH6 complex in lipid trafficking and offer initial insights into the relevance of scramblases for carrier-like lipid transport mechanisms.
Similar content being viewed by others
Main
The lipid composition of membranes surrounding cellular compartments is highly heterogeneous, resulting from a combination of localized biosynthesis and complex transport processes1,2,3. The negatively charged phosphatidylserine (PS), for example, is synthesized in the endoplasmic reticulum (ER), while it is accumulated in the plasma membrane (PM)4,5,6. In eukaryotic cells, the transport of PS occurs through both vesicular and nonvesicular mechanisms. The latter are largely mediated by members of the oxysterol-binding protein-related protein family (ORP), some of which possess targeting motifs that localize them to membrane contact sites (MCSs) where they promote lipid exchange7,8,9,10,11. In the yeast Saccharomyces cerevisiae, nonvesicular transport of PS to the PM is facilitated by the ORP homologs OSH6 and OSH7 (refs. 4,12), which accumulate PS on the PM in exchange for phosphatidylinositol 4-phosphate (PtdIns4P)9,11. PtdIns4P is transferred from the PM to the ER, where its dephosphorylation by the phosphatase SAC1 helps to maintain a concentration gradient that drives transport13. This process takes place at the cortical ER, which is abundant in S. cerevisiae and stabilized by different tethering proteins that keep both membranes at distances ranging between 10 and 30 nm, as revealed from electron microscopy (EM) data14,15,16. Because OSH6 and OSH7 themselves lack domains localizing them to the cortical ER, they bind to IST2, which is one of the primary tethers found to form ER–PM contact sites in this unicellular organism17,18,19,20.
The 946-residue-long IST2 is a member of the TMEM16 family, encompassing ion channels and lipid scramblases in eukaryotic cells that are regulated by intracellular calcium21,22,23. IST2 distinguishes itself from other family members by its modular organization, consisting of a membrane-inserted N-terminal half and an extended C-terminal cytoplasmic region of comparable length. The sequence of the N-terminal part resembles other fungal TMEM16 scramblases and the mammalian paralogs TMEM16H and TMEM16K, albeit without the residues of the Ca2+-binding site, which are strongly conserved among other family members24. In contrast, the following region is predicted to be unstructured and to bridge the ER with the PM by binding to the inner leaflet of the latter through a cluster of positively charged residues at the C-terminal end20. The cytoplasmic region of IST2 was shown to localize OSH6 and OSH7 to MCSs. Both bind to a stretch in the center of the elongated tail, which is conserved within the family of Saccharomycetaceae17,18. In this study, we focus on OSH6 but one can expect our findings to also extend toward the highly homologous OSH7. While the coarse region interacting with OSH6 has been identified, the details of the IST2–OSH6 interaction and its role in nonvesicular lipid transport have remained elusive. Open questions concern the function of IST2 as a lipid scramblase, the structural basis of its interaction with OSH6 and whether the two proteins remain associated during lipid shuttling. To address these questions, we engaged in extended investigations using a combination of biochemical, structural and cellular approaches to understand the intricate interplay between IST2 and OSH6 at MCSs.
Results
Structural features of the IST2–OSH6 complex
In our study, we aimed for a detailed comprehension of the IST2–OSH6 system for lipid transport. To characterize the complex in vitro, we separately overexpressed full-length IST2 (IST2fl) and OSH6 in S. cerevisiae. After their purification and reconstitution, we found both proteins to coelute as a complex on size-exclusion chromatography (SEC) (Fig. 1a). A structural characterization of this sample by cryo-EM provided distinct reconstructions that are seemingly not connected, which can be attributed to the transmembrane domain of IST2 (Fig. 1b,c and Extended Data Fig. 1) and bound OSH6 (Fig. 1b,d and Extended Data Fig. 1), respectively. These features are in accordance with the predicted modular organization of IST2 consisting of a folded N-terminal and a largely unstructured C-terminal half (Fig. 1b, Extended Data Fig. 1, Table 1 and Supplementary Fig. 1). The membrane-inserted domain of IST2 assembles as a dimer, showing the characteristic architecture of a TMEM16 protein24 (Fig. 1c and Extended Data Fig. 1a). In the case of bound OSH6, the small size of the protein compromised particle alignment, which consequently limited the obtained resolution (Fig. 1d and Extended Data Fig. 1a). In this dataset, we also found two-dimensional (2D) classes containing multiple particles at short distances (Extended Data Fig. 1a), which could account for potential interactions between OSH6 and IST2, although the resolution of these classes is insufficient to draw a definitive conclusion.

a, SEC profile of a sample containing IST2 and OSH6 at a 1.5× molar excess (yellow). Inset, the peak fraction (*) contains the complex of both proteins as shown by SDS–PAGE. The red trace represents free OSH6. b, Scheme composed of the cryo-EM density of IST2 and OSH6 as observed in their complex with the flexible linker that is not defined in the structure sketched as line. The C-terminal parts interacting with the PM are shown in green and the membrane boundaries are indicated. c, Cryo-EM density of the N-terminal membrane-inserted part of the IST2 dimer at 4.9 Å. d, Cryo-EM density of OSH6 at 10.5 Å with a placed ribbon representation of its structure (PDB 4B2Z) shown for comparison. e–h, Lipid-scrambling properties of the N-terminal construct IST2716 assayed in liposomes by the irreversible bleaching of fluorescent lipids facing the outside. e, Absence of lipid-scrambling activity in murine TMEM16A-containing proteoliposomes. Data show averages of three measurements obtained from one reconstitution. f, Ca2+-dependent scrambling in proteoliposomes containing nhTMEM16. In e,f, liposomes not containing any protein (neg.) are shown as a control. g, Ca2+-independent scrambling in proteoliposomes containing IST2716. Traces of empty liposomes and nhTMEM16 (displayed in b) are displayed as dotted lines for comparison. In f,g, data show the averages of nine measurements obtained from three independent reconstitutions. In e–g, traces depict the fluorescence decrease in tail-labeled NBD–PE lipids after the addition of dithionite (t = 0) in the absence and presence of 1 mM Ca2+. The dashed gray line indicates the 0.5 threshold. h, Comparison of fluorescence 150 s after the addition of dithionite in proteoliposomes containing nhTMEM16 or IST2716. Bars show the average of nine experiments (displayed as dots). The fluorescence difference in the presence of Ca2+ is significant in the case of nhTMEM16 (P < 0.001) but not IST2716 (P > 0.9999) as calculated by a two-way ANOVA using Šidák’s multiple comparisons test. For the statistical analysis, values from the same reconstitution were matched to account for differences in the reconstitution efficiency. In e–h, errors are the s.e.m.
The N-terminal domain (NTD) of IST2 forms a lipid scramblase
Next, we were interested whether the N-terminal part of IST2 retained its capability to mediate lipid transfer between both leaflets of the membrane in spite of a previous publication refuting scrambling activity of the protein25. We thus prepared a construct containing the first 716 amino acids (IST2716), which lacks the bulk of the C-terminal region including the proposed OSH6-binding sequence (Extended Data Fig. 2a and Supplementary Fig. 1a). To investigate its scrambling capability, we reconstituted the purified IST2716 into liposomes containing trace amounts of fluorescent lipids (Extended Data Fig. 2b). Scrambling was assayed by following the time-dependent decrease in fluorescence upon bleaching of labeled lipids residing in the outer leaflet of the bilayer25 (Extended Data Fig. 2c). In the case of empty liposomes or proteoliposomes containing TMEM16A, which is unable to scramble lipids, the fluorescence decayed to about 50% of its initial value (Fig. 1e). For the fungal scramblase nhTMEM16, we observed a slow fluorescence decay to 15–30% of its initial value in the absence of Ca2+ and faster kinetics upon Ca2+addition, reflecting the increased transfer rates of lipids between both leaflets upon ligand binding24 (Fig. 1f and Extended Data Fig. 2d–f). Reconstituted IST2716 displayed pronounced scrambling activity in the absence of Ca2+, which was not further enhanced upon Ca2+ addition, distinguishing the protein as a constitutively active lipid scramblase (Fig. 1g,h and Extended Data Fig. 2b,d–f). This feature aligns with the absence of the conserved Ca2+-binding site in homologs from Saccharomycetaceae, which are all supposed to participate in similar processes at MCSs (Supplementary Fig. 1b).
Structure of the IST2 scramblase unit
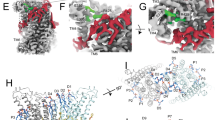
After identifying the membrane-inserted part of IST2 as a constitutively active scramblase, we investigated how this functional feature would be manifested in its architecture. We used the well-behaved IST2716 construct for cryo-EM sample preparation, for which we obtained high-resolution reconstructions in the detergent glyco-disogenin (GDN) at a global resolution of 2.84 Å and in small-sized lipid nanodiscs (assembled with the lipid scaffolding protein 1E3D1) at 3.35 and 3.8 Å, respectively (Fig. 2a, Extended Data Figs. 2–4 and Table 1). As observed for the transmembrane domain of IST2fl, IST2716 is a homodimer that shares the general features of known structures of TMEM16 family members24,26,27,28 (Fig. 2a,b). Each subunit consists of a cytosolic NTD that is followed by a unit comprising ten membrane-spanning segments and a disordered C terminus encompassing residues 596–716 (Fig. 2b,c and Supplementary Fig. 1a). Although the NTD of IST2 resembles the equivalent region of nhTMEM16, it is considerably smaller and restricted to the core ferredoxin fold consisting of a four-stranded antiparallel β-sheet and two interspersed α-helices (Fig. 2c and Extended Data Fig. 5a). This cytoplasmic domain tightly interacts with intracellular helices and loops at the periphery of IST2 remote from the dimer interface to stabilize the observed structure in this region (Fig. 2c and Extended Data Fig. 5b). The dimer interface burying 1,516 Å2 of the combined molecular surface involves contacts between the symmetry-related α10 helices located in the outer leaflet and between α10 and α3 in the inner leaflet of the bilayer (Extended Data Fig. 5c). The low-resolution density of the detergent micelle surrounding the membrane-embedded part of the protein shows a pronounced distortion that delineates a trail of lipid-like densities surrounding the transmembrane helices that is still visible at high map contour (Fig. 2d and Extended Data Fig. 5d,e). A similar distortion is also observed in two distinct reconstructions from a dataset of IST2716 in lipid nanodiscs (Extended Data Figs. 4 and 6a), which further underlines the role of the protein in destabilizing its environment. Bound detergent molecules and lipids follow the arrangement of transmembrane helices, resulting in an apparent thinning of the bilayer at both entrances of the subunit cavity, closely resembling previously described features observed in the fungal scramblases afTMEM16 and nhTMEM16 (refs. 27,29,30) (Fig. 2d and Extended Data Fig. 5d,e). Similarly, the part of the protein defining the subunit cavity, which comprises α-helices 3–8, displays a striking resemblance to the Ca2+-bound active conformations of nhTMEM16, afTMEM16 and human TMEM16K (refs. 24,29,31) (Fig. 2e and Extended Data Fig. 6b,c). By forming a continuous polar furrow exposed to the core of the membrane, which is confined by α4 and α6, this unit appears of appropriate width to accommodate a lipid headgroup, presumably lowering the barrier for lipid flip-flop, as suggested in computer simulations of nhTMEM16 (refs. 32,33,34,35) (Fig. 2e and Extended Data Fig. 6d). Despite the described similarities, IST2 contains several unique structural hallmarks, which presumably stabilize its conformation to impede a collapse of the polar furrow as observed in other fungal family members in absence of Ca2+ (ref. 27,29,31). In nhTMEM16 and its orthologs, the open conformation is stabilized by Ca2+ binding to an extended site located within the inner membrane leaflet that is strongly conserved within the TMEM16 family24. This site contains five acidic residues on α6–α8 that coordinate two Ca2+ ions to rigidify the intracellular part of α6 in its interaction with α8 (Fig. 2f,g). In IST2, three of the conserved acidic residues located on α7 and α8 are substituted. In particular, the swap of a glutamate (E506 in nhTMEM16) with a lysine (K443 in IST2) is striking, as it introduces a positively charged side chain that replaces Ca2+ by forming a bifurcated salt bridge with the conserved residues E379 on α6 and D476 on α8 (Fig. 2f,g). Additionally, the observed conformation of α6, the lower part of which shows a stronger bending toward the membrane plane compared to other homologs of known structure, is stabilized by a successive short cytoplasmic helix (α6i) in a region that is disordered in other family members (Fig. 2h).

a, Cryo-EM density of IST2716 at 2.84 Å viewed from within the membrane. The relationship between views is indicated. Subunits are colored in yellow and violet, with residual density presumably corresponding to detergent or lipids in gray. b, Ribbon representation of IST2716 with colors and orientation as in a. Membrane boundaries are indicated and selected regions of the structure are labeled. c, Ribbon representation of the IST2 subunit at indicated orientations, with the NTD colored in red. Transmembrane helices are labeled. d, Cryo-EM density of IST2716 with residual density corresponding to detergent and lipids colored in violet. A lowpass-filtered map at low contour showing the distortion in the detergent micelle is superimposed. ‘#’ and ‘*’ indicate sites of deformation. e, Subunit cavity viewed from within the membrane. Shown are α3–α8 from a single subunit. Superposition of IST2716 (yellow) and the Ca2+-bound open state of nhTMEM16 (left, brown; PDB 6QM9), IST2716 (center) and nhTMEM16 (right). The molecular surface in the region of the subunit cavity is displayed and colored according to the properties of contacting residues (polar, green; acidic, red; basic, blue). f, Region of IST2 corresponding to the regulatory Ca2+-binding site of the TMEM16 family. Left, cryo-EM density superimposed on the model. Right, ribbon with the corresponding side chains of IST2 displayed. g, Comparison of the regulatory region between nhTMEM16 and IST2. Left, nhTMEM16 with two Ca2+ ions bound, Right, superposition of equivalent regions of IST2 and nhTMEM16. Bottom, sequence alignment of relevant regions in fungal and murine TMEM16 proteins. The numbering refers to IST2. h, Superposition of transmembrane α6 and α7 and their connecting region in IST2 and nhTMEM16. Inset, zoomed-in view of the interacting cytoplasmic helices α6i and α6’. In e–g, Ca2+ ions bound to nhTMEM16 are displayed as blue spheres. i, Conformation of α4 and α6 lining the edge of the subunit cavity in a superposition of the subunits of IST2 and nhTMEM16 in the presence (open; PDB 6QM9) and absence (closed; PDB 8TPM) of Ca2+. The protein is shown as a ribbon with the equivalent positions of α4 occupied by glycine residues in nhTMEM16 marked as spheres.
In nhTMEM16 and afTMEM16, the absence of Ca²⁺ causes α6 to detach from the binding site, triggering a conformational change in α4 that collapses the subunit cavity on the extracellular side27,29. In nhTMEM16, α4 contains multiple glycine residues that influence its conformational flexibility and, thus, facilitate the collapse29 (Fig. 2i). In IST2, these glycines are not conserved, presumably increasing the stability of α4 in the observed conformation (Fig. 2i). Consequently, the cavity is found open in all determined structures of IST2716. For nanodisc-reconstituted IST2716, we obtained two reconstructions with different disc sizes, which we termed ‘narrow’ and ‘wide’ according to the width of the lipid layer surrounding the protein (Extended Data Fig. 4). In both subunits of the narrow and in one subunit of the wide reconstruction, the proximity of the scaffold protein to IST2716 appears to constrain α6, leading to a loss of density in its lower half and the following α6i (Extended Data Fig. 6a,b). Conversely, the density of the entire α6 region is retained in the second subunit of the wide reconstruction, where the bilayer surrounding the cavity is larger (Extended Data Fig. 6a,b). Despite these differences, both structures recapitulate features found in IST2716GDN, where α4 and α6 adopt an open groove conformation (Extended Data Fig. 6b). However, even in the case of IST2, this open conformation would be intrinsically destabilized by the interaction of the hydrophilic surface inside the cavity with the hydrophobic core of a membrane or detergent micelle. This is reflected in the structure of the membrane-inserted unit of IST2fl in complex with OSH6, where the cavity has rearranged to shield its hydrophilic surface by movements of α3, α4 and α6 (Extended Data Fig. 6e,f).
In summary, the structural and functional characterization of the membrane-inserted part of IST2 defines its function as a constitutively active lipid scramblase, exhibiting all hallmarks associated with a lowered energy barrier for lipid flip-flop.
The C-terminal region of IST2 and its interaction with OSH6
The absence of structured elements following the membrane-inserted part of IST2 (that is, after residue 595) confirms previous predictions of the C-terminal linker as an intrinsically disordered protein sequence20 (Figs. 1 and 2, Extended Data Figs. 1, 3 and 4 and Supplementary Fig. 1a). To obtain further insight into the properties of this region and its interaction with lipid transfer proteins (LTPs), we first determined the minimal sequence required for OSH6 binding by pulldown assays. We initially based our construct design on previously published results, which suggested the core binding region to be located between residues 727 and 776 (refs. 17,18) (Supplementary Fig. 1a,c). This conserved stretch is situated close to the center of the 333-residue-long linker (excluding the C terminus tethered to the PM), measuring about 100 nm in a fully extended state and severalfold exceeding the 10–30-nm-wide gap between the ER and the PM. Through systematic truncation of the sequence on both ends of the binding site, we found an unperturbed interaction with a peptide encompassing residues 732–759, followed by a pronounced decline in shorter sequence stretches (Extended Data Fig. 7a). To determine the strength of the interaction, we quantified the binding of peptides comprising IST2 residues 732–761 (IST2732−761) and 732–756 (IST2732−756) to OSH6 by isothermal titration calorimetry (ITC) and obtained affinities in the high nanomolar to low micromolar range (694.4 ± 54.2 nM and 714.0 ± 86.7 nM in the case of IST2732−761 and 1.1 ± 0.17 μM in the case of IST2732−756) (Fig. 3a and Extended Data Fig. 7b–d). To understand the structural basis of the interaction, we cocrystallized both peptides with an OSH6 variant lacking the first 30 aa (termed OSH6ΔN), which were disordered in previously determined crystal structures10,11. Data collected from complexes with either peptide extend beyond 2.0-Å resolution and provide a detailed view of the LTP in complex with its interacting region on the IST2 C terminus (Fig. 3b–g, Extended Data Fig. 7e–g and Table 2). In both structures, OSH6 adopts a similar curved β-sheet conformation with a central a single lipid-binding site that is closed by an N-terminal lid, as observed in previous complexes with PS and PtdIns4P10,11 (Fig. 3b). An exception concerns the N terminus, where α1 is extended, presumably as a consequence of crystal packing interactions (Fig. 3b and Extended Data Fig. 7e). The map obtained for the OSH6ΔN–IST2732−761 complex shows well-defined density for residues 732–758 of IST2, which bind to OSH6 on the opposite side of the lid covering the entrance to the lipid-binding pocket, burying 1,516 Å2 of the combined molecular surface (Fig. 3b–d and Extended Data Fig. 7f–h). The shorter peptide IST2732−756 shows an equivalent binding mode and is defined until R750 (Fig. 3c). The interacting residues on OSH6 are strongly conserved in homologs that presumably form similar complexes with IST2-like proteins but not in other family members that do not bind this tethering protein17 (Extended Data Fig. 7i and Supplementary Fig. 2). The N-terminal part of the peptide occupies a groove lined by residues 408–434 at the C terminus of OSH6, which is stabilized by intermolecular hydrogen bonds between backbones and side chains. The latter include interactions of the side chains of Y391, Q417 and T420 of OSH6 and T736 of IST2 (Fig. 3e and Extended Data Fig. 7g,h). Aliphatic contacts are pronounced among residues 737–742 containing two proline residues (P738 and P742), perturbing the extended conformation of the peptide in a region that is unusually hydrophobic compared to the remainder of the IST2 linker (Fig. 3f, Extended Data Fig. 7g,h and Supplementary Fig. 1a,c). At the C terminus of the interaction region, residues S744–R750 form a short helical segment, which is stabilized by an R750–T743 hydrogen bond and the stacking of the aromatic side chains of Y747 and F751 (Fig. 3g and Extended Data Fig. 7g). Our results align with previously published data, in which substitution of T736 and T743 compromised the recruitment of OSH6 to contact sites17. In addition to the observed stabilization of the peptide conformation, R750 might also engage in long-range water-mediated interactions with the more distant D141 of OSH6, which was also described to impair binding when substituted to alanine17 (Fig. 3g). Substitution of R750 to alanine did indeed severely compromise binding to OSH6 in a pulldown assay and ITC, emphasizing the importance of this residue for the interaction (Extended Data Fig. 7j–l). In addition to a detailed view of the interaction with its epitope on IST2, the crystal structure revealed well-resolved density of bound PS in the conserved binding pocket of OSH6, which was confirmed by mass spectrometry (Fig. 3h). Because OSH6 was overexpressed in S. cerevisiae, the lipid corresponds to the endogenous substrate and demonstrates that binding of PS, which is considerably more abundant than its higher affinity substrate PtdIns4P36, and IST2 are mutually compatible. Together, our results confirm the direct interaction between OSH6 and IST2 through a conserved region in the center of the disordered tail involving residues 732–751 (Supplementary Fig. 1a,c).

a, Binding of a peptide encompassing residues 732–761 of IST2 (IST2732−761) to OSH6. Binding isotherms were obtained from the titration of IST2732−761 to OSH6. The data show a fit to a single-binding-site model line with a Kd of 694.4 ± 54.2 nM. Errors represent fitting errors. Inset, thermogram. b, Ribbon representation of the structure of the OSH6–IST2732−761 complex determined by X-ray crystallography at 1.84 Å. IST2732−761 is colored in yellow and the N-terminal lid helix of OSH6 is colored in green. A bound PS molecule is displayed in light blue. c, The 2Fo − Fc density of the refined structure superimposed on a model of the IST2-binding region. Left, density from the OSH6–IST2732−761 complex defining the conformation of residues 732–758 of the peptide. Right, density of the OSH6 complex with the shorter IST2732−756 peptide determined at 1.94 Å defining the conformation of residues 732–750 of the peptide. The sequence of the IST2732−761 peptide is shown below. d, Sections of the OSH6–IST2 interactions revealed in the OSH6–IST2732−761 complex. A model of the well-defined parts of the IST2732−756 peptide in stick representation, with its interacting region of OSH6 represented as ribbon. Different regions of the interaction are indicated as shaded areas and shown as zoomed-in views in e–g with OSH6 represented as sticks. e–g, Sections of the OSH6–IST2 interactions highlighted in d in relation to the OSH6-binding sequence on IST2: N-terminal region (e), center (f) and C-terminal region (g). h, The 2Fo − Fc density of the bound PS lipid (left) and lipid-binding region of OSH6, with interacting side chains shown as sticks.
OSH6 interaction with membranes
After characterizing its binding to IST2, we investigated the interaction of OSH6 with lipid bilayers, following up on earlier studies that described this process as a critical step during the extraction and release of its lipid cargo37. To monitor its avidity for negatively charged lipid patches, we mixed fluorescently labeled OSH6 with lipid nanodiscs composed of the anionic lipid POPG (phosphatidylglycerol) and the neutral lipid POPE (phosphatidylethanolamine) and were able to detect its coelution with the nanodisc fraction in SEC (Fig. 4a). As the binding of its specific substrates PS or PtdIns4P decreases its affinity to lipid bilayers, we subsequently overexpressed and purified OSH6 from Escherichia coli (Extended Data Fig. 7b), whose membranes have a low PS content, and confirmed the absence of bound lipids by mass spectrometry. With this sample, we proceeded with a structural characterization by cryo-EM (Fig. 4b and Extended Data Fig. 8). In 2D reconstructions of this dataset, distinct classes show the density of single OSH6 molecules bound to the edge of a lipid nanodisc (Extended Data Fig. 8b). After intensive classification, a three-dimensional (3D) reconstruction permitted the placement of OSH6 with its loop region leading to the PS-binding site in presumed contact with lipids (Fig. 4c,d and Extended Data Fig. 8c–g). In this binding mode, the IST2-binding epitope is located on the opposite face of the protein remote from the membrane, permitting simultaneous binding to its tether (Fig. 4d). The location of OSH6 at the edge of nanodiscs illustrates its preference for distorted membrane regions, as no density was found in the center of the disc, nor did we detect interactions with the isolated scaffold protein (Extended Data Fig. 8b,h,i). This preference is remarkable considering the observed membrane distortion by the transmembrane part of IST2, which could facilitate OSH6 binding, although such a mechanism is still hypothetical (Fig. 2d and Extended Data Figs. 5d,e and 6a).

a, SEC profiles of OSH6 in absence and presence of 2N2 lipid nanodiscs monitored by the fluorescence of the labeled LTP. The comigration with nanodiscs is reflected in the decreased elution volume. b, SEC profile and SDS–PAGE gel (inset; n = 1) of the OSH6–2N2 lipid nanodisc sample used for cryo-EM analysis. c, Ab initio reconstruction of OSH6 (red) bound to the edge of a lipid nanodisc (white) obtained from a cryo-EM dataset. d, Model of OSH6 fitted into the masked and refined cryo-EM density. The lid helix α1 is colored in green. The interacting IST2 peptide (yellow), which was not included in the sample, is displayed for reference. The bound POPS is shown as space-filling model and the relationship between views is indicated.
Properties of the IST2 linker relevant for lipid transfer
After characterizing the binding of OSH6 to IST2 on a biochemical and structural level, we investigated the role of the IST2 linker and its interaction with OSH6 for lipid transport. We were particularly interested in the influence of linker length, the impact of the relative location of the binding site and whether OSH6 remains bound to the cytoplasmic tail of IST2 during lipid transfer.
To investigate these questions, we used a previously established cellular assay based on the rescue of a growth defect in an ist2;psd1-knockout (KO) strain, as a consequence of phosphatidylcholine (PC) limitation18. The described growth deficit can be alleviated either by transformation with a construct encoding wild-type (WT) IST2 or by supplementation of choline, a PS precursor of the Kennedy pathway (Fig. 5a and Extended Data Fig. 9a–c). In contrast, growth is not rescued by overexpression of the truncated construct IST2716, with no OSH6-binding site, or the longer construct IST2921, with the binding site but without the C-terminal cortical localization sequence, which were both previously shown to mislocalize to the perinuclear ER38 (Fig. 5a, Extended Data Fig. 9c and Supplementary Fig. 3a). Using this assay, we assessed the ability of modified IST2 constructs to form a functional lipid transport system at MCSs in yeast. We first probed the effect of an alteration of the OSH6-binding site by introducing the R750A substitution, which strongly weakened binding (Extended Data Fig. 7j–l), or by scrambling the sequence of the binding site without altering its composition (Supplementary Fig. 3b). In both cases, we found a severely compromised complementation of growth, which was pronounced in the mutant and completely abolished in case of the scrambled binding site, underlining the role of the interaction for lipid transport (Fig. 5b and Extended Data Fig. 9d). We then probed the impact of the relative location of the binding site on the linker by its displacement by 83 residues toward the N or C terminus of the protein while preserving its overall length (Supplementary Fig. 3c). In this case, we found a severely reduced growth phenotype, which is more pronounced upon movement of the site toward the N terminus than toward the C terminus (Fig. 5c and Extended Data Fig. 9c), as expected from the different lengths of the respective linker regions.

Cell growth assayed upon overexpression of IST2 constructs in a S. cerevisiae ist2;psd1-KO strain. a, Comparison of WT (IST2) to the constructs IST2921 lacking 25 residues of the C terminus including the PM-binding region and IST2716 lacking 230 residues including the OSH6-binding site (P < 0.0001 for both). b, Comparison of WT to a protein carrying the R750A substitution in the OSH6-binding site of IST2 (P = 0.001), which strongly reduced the interaction, and another construct where the sequence of the binding site was scrambled without changing its composition (P < 0.001). c, Comparison of WT to constructs where the OSH6-binding site was moved by 83 residues toward the N or C terminus of the protein (P < 0.0001 for both). d, Comparison of WT to constructs where the IST2 linker was symmetrically shortened by removal of residues from the center of both sequence regions located N-terminally and C-terminally to the binding site to reach the indicated fraction of its original length (IST2 versus neg., P < 0.0001; IST2 versus 75%, P = 0.9994; IST2 versus 50%, P < 0.0001; IST2 versus 33%, P < 0.0001; IST2 versus 25%, P < 0.0001). NS, not significant. e, Comparison of WT to constructs with the OSH6-binding site removed and OSH6 fused into the equivalent position into the linker (fusion, P = 0.0137), a fusion construct where the linker was symmetrically truncated by 50% in the center of both regions flanking the fused LTP (fusion trunc., P < 0.0001) or expanded by 50% by introducing a G-S peptide sequence (Fusion ext., P = 0.9951). In a–e, schemes illustrate the properties of IST2 constructs. Bars display the mean normalized growth compared to WT (n = 3 × 3 replicates for most constructs and 2 × 3 replicates for fusion trunc. and fusion ext.). Errors are the s.e.m. Values of individual experiments are shown as dots. Statistical analysis was performed using a one-way ANOVA and Tukey’s multiple comparisons test. Values from the same experimental replicate were matched in the analysis. f, Potential energy of the C-terminal region of IST2 modeled as an entropic spring during the shuttle of OSH6 between the ER and the PM in constructs of different length. Top, correspondence of energies to states during transport.
Next, we investigated the effect of linker length on complementation upon shortening of the C terminus by a symmetric removal of residues from the center of both regions flanking each side of the OSH6-binding sequence (Supplementary Fig. 3d). Assuming complete flexibility of the linker on both sides of the binding site, its length would exceed the experimentally observed distance between the PM and the ER at least by a factor of two20, which would permit a shuttle mechanism of OSH6 between both membranes while remaining bound to IST2. In our studies, we observed pronounced complementation in constructs containing up to three quarters of the residues and a sharp decline in shorter constructs with an apparent minimal length of 170–250 residues, suggesting that shorter constructs would be insufficient to permit bound OSH6 to reach both membranes (Fig. 5d and Extended Data Fig. 9e). Lastly, we investigated a fusion construct where the OSH6-binding site was replaced by a covalent insertion of OSH6 into the linker (Supplementary Fig. 3e). Remarkably, this construct, which is biochemically well behaved, led to a nearly WT-like complementation of the growth phenotype, which we found unaltered upon extension and considerably reduced upon shortening of the linker (Fig. 5e and Extended Data Fig. 9f,g). To better understand the results of the complementation studies, we analyzed our constructs by approximating the two flexible linkers of IST2 flanking the bound OSH6 as entropic springs, which allowed an estimation of the energies along its trajectory between opposite membranes (Extended Data Fig. 10). These calculations suggest that both linker length and binding site location have evolved to ensure a shallow energy profile, which is strongly increased upon shortening of the linker or displacement of the binding site, compromising the ability to shuttle lipids (Fig. 5f and Extended Data Fig. 10b–d). The results from the yeast complementation assays in conjunction with a mathematical model, thus, demonstrate the importance of the OSH6–IST2 interaction for lipid transport between apposing membranes of the contact site, suggesting that the complex remains intact during lipid transfer.
Discussion
As the main hub for lipid synthesis in eukaryotes, the ER has an important role in the distribution of lipids to other cellular compartments. Recently, nonvesicular lipid transport at MCSs of the ER has gained considerable attention39 and was shown to proceed through two general mechanisms. Channel-like transport processes, as described for VPS13 (ref. 40,41), the autophagy-related protein ATG2 (ref. 42,43) and the ERMES complex44,45, are mediated by continuous hydrophobic conduits, which bridge the membranes of contacted compartments and facilitate lipid diffusion with low specificity. A second mechanism mediates selective transport by carrier proteins, such as members of the ORP family, which contain a specific and saturable lipid-binding site7,46. These proteins are tethered to a flexible linker attached to a membrane-embedded domain, which localizes the lipid carriers to the MCS to take up and release their cargo at the respective target membranes47,48 (Fig. 6). The tether can either be covalently attached to the lipid carrier, as in the case of certain mammalian ORP homologs, or be a distinct protein as in the case of the yeast IST2–OSH6/7 system46,49.

a, Transport mechanism of the IST2–OSH6 system. Center, general organization of the complex consisting of the transmembrane domain of IST2 located in the ER that is tethered to the PM through a lipid-binding region attached to the C terminus of a flexible linker, which contains a binding site for the LTP OSH6 in its center. Left, close-up view of the transport cycle illustrating the shuttling of OSH6 between membranes to mediate the exchange of the lipids PS and PtdIns4P with the IST2 linker acting as an entropic spring. Right, function of the membrane-inserted part to distort the membrane and mediate lipid scrambling between both leaflets to facilitate lipid exchange. b, Analogous processes in higher eukaryotes mediated by the mammalian homologs ORP5 and ORP8. In both cases, the interaction with a lipid scramblase is still hypothetical.
In our study, we investigated the IST2–OSH6 system in S. cerevisiae with a focus on the role of the membrane protein IST2, a homolog of the TMEM16 family that encompasses ion channels and lipid scramblases23. We were particularly interested in the structural properties of IST2 and whether it has retained the lipid transport activity found in other family members. Although its ability to scramble lipids was challenged previously25, we here confirmed this function using in vitro lipid transport assays and a thorough structural characterization, which together provide a mechanistic explanation for its constitutive activity (Figs. 1g,h and 2). In our data, IST2 shows the structural hallmarks of a TMEM16 lipid scramblase with unique features in the catalytic subunit cavity region that lead to a stabilization of the ‘open’ conformation (Fig. 2e–i). Similar to structures obtained for the Ca2+-bound active form of its fungal homologs afTMEM16, nhTMEM16 and the mammalian TMEM16K (refs. 24,27,31), this cavity exposes a hydrophilic surface to the membrane (Fig. 2e). Unlike in most family members, the regulatory Ca2+-binding site is altered in IST2 and its close homologs in Saccharomycetaceae to stabilize the observed conformation even in the absence of Ca2+ (Fig. 2f,g and Supplementary Fig. 1b).
The identification of IST2 as a lipid scramblase aligns with their proposed role as essential components of nonvesicular lipid transport systems50. In channel-like systems, scramblases can relax tensions created by a lipid net flux between two compartments through extraction from a single leaflet. Conversely, their role is less obvious in carrier-like systems, where transport occurs as a 1:1 lipid exchange. In the case of OSH6, PS transport from the ER to the PM is compensated by PtdIns4P countertransport, preventing a pronounced change in the membrane surface of either compartment11. However, even in this case, scrambling may be necessary to relax local lipid imbalances arising from the accumulation of the bulky inositol headgroup of PtdIns4P in the vicinity of IST2. Additionally, our data suggest that the membrane distortion by IST2 could facilitate the access of OSH6 to its lipid cargo. In both channel and carrier models of lipid transfer, lipid extraction from the bilayer is considered the rate-limiting step40,51 and local membrane perturbations could help to lower this energy barrier. In our cryo-EM studies of OSH6–nanodisc complexes, we observe OSH6 binding to the periphery of the lipid bilayer (Fig. 4d,e), suggesting its preference for distorted membrane regions, although this hypothesis requires further validation.
In addition to the characterization of its transmembrane domain, we investigated the extended unstructured cytosolic region of IST2, which functions as a tether at ER–PM contact sites that recruits the soluble OSH6 through a conserved binding site located in its center. Our results reveal the structural basis of the interaction with the IST2 C terminus, showing that its affinity is strong enough to support sustained attachment to the linker in a cellular environment (Fig. 3a–g and Extended Data Fig. 7). Cellular complementation assays further demonstrate that this persistent binding of OSH6 is crucial for efficient lipid transport (Fig. 5), despite the intrinsic membrane avidity of OSH6 observed in vitro (Fig. 4). The permanent attachment of OSH6 to IST2 during lipid transport is reminiscent of the homologous OSBP-related lipid transport systems found in mammals52, where the PS–PtdIns4P exchangers ORP5 and ORP8 directly act as membrane tethers through an N-terminal PH domain, which binds to PtdIns molecules in the PM, and a C-terminal transmembrane helix, which anchors the protein to the ER49,53. Both systems share equivalent features of a lipid carrier that is anchored to the center of a flexible tether, which can be modeled as two flanking entropic springs (Fig. 6 and Extended Data Fig. 10). These springs compact to minimize their potential energy, which in turn increases upon extension54,55 (Extended Data Fig. 10b). The resulting restriction of off-axis displacements essentially limits the movement of OSH6 to a one-dimensional diffusion. In this scenario, one of the two springs is loaded, as OSH6 binds to either the PM or the ER while the other remains relaxed (Fig. 6a). Together, this system resembles an oscillator-like mechanism, which could enhance the rate of lipid exchange and compensate for the comparably low transport capacity of a single lipid molecule per carrier.
Because the covalent association of a membrane tether to a lipid scramblase is a unique feature of Saccharomycetaceae, whereas ORP systems in higher eukaryotes only contain a simple membrane anchor, it will be interesting to investigate whether these proteins recruit accessory scramblases to take over the role of IST2 in mammalian systems (Fig. 6b). Interestingly, the human homologs TMEM16K and TMEM16H were shown to localize to MCSs between the ER and endosomes, although their exact function remains elusive56,57. While our data clearly demonstrate that the ER domain exerts a functional role beyond anchoring, previously published results suggest that the TM unit of IST2 is, to a large extent, dispensable18. Consequently, the question of whether this role is of immediate relevance for PS transport or whether it facilitates other cellular processes still needs to be resolved.
In summary, our results provide detailed mechanistic insight into lipid transport at MCSs mediated by carrier-like mechanisms. Our data extend the previous notion of IST2 as a simple membrane tether and demonstrate that the continued binding of OSH6 is indispensable for its efficiency as a lipid transporter. While additional experiments will be necessary to better understand the interplay between both parts of the complex, our study opens up a compelling view on the role of intracellular TMEM16 proteins at MCSs and demonstrates the association of an active scramblase with a carrier-like lipid transport system.
Methods
Cloning of expression constructs
All expression constructs were generated by FX cloning58 unless specified otherwise. For constructs expressed in yeast, the sequences of nhTMEM16, IST2fl and IST2 containing residues 1–716 (IST716) were cloned into a modified FX-compatible pYES2 vector with an N-terminal SBP–myc fusion followed by a 3C recognition site (pYESnSM3). IST2 tail variants used in the yeast complementation assay (Supplementary Fig. 3) were either ordered as synthetic DNA constructs from Genscript (IST250%, Move-C, Move-N and IST2–OSH6 fusion) or cloned by a seamless exchange of synthetically ordered tail fragments into a ccdb cassette flanked by SapI cleavage sites introduced between residues 590–591 (serine) and 926–927 (alanine) in IST2fl (ISTScr, IST225%, IST233%, IST275%, IST2–OSH6 fusion extended and IST2–OSH6 fusion truncated). The sequences of OSH6 and OSH6ΔN were cloned into pYESnSM3. The substitutions R750A in IST2 and C389S in OSH6were introduced using a modified QuickChange protocol59. For bacterial expression constructs, the sequence of OSH6 was cloned into a pGEX-4T vector using BamHI and EcoRI. The forward primer was extended in the 5′ end to introduce an N-terminal 3C cleavage sequence following the thrombin recognition site. TMEM16A was inserted into a pcDXc3GMS vector using FX cloning58. The sequences for MSP2N2 and MSP1E3D1 were purchased from Addgene. All constructs were verified by Sanger sequencing. All oligonucleotides used in this study can be found in the Supplementary Information.
Protein expression and purification
Recombinant IST2 and nhTMEM16 constructs were transformed into the S. cerevisiae expression strain FGY217 using the LiAc–SS carrier DNA–PEG method60. Positive clones were grown in SC −Ura selection medium at 30 °C in a fermenter until an optical density at 600 nm (OD600) of 0.7 was reached. Protein expression was induced by the addition of 2% galactose and the temperature was reduced to 25 °C. After 40 h, the cells were harvested at 4,000g for 10 min.
IST2716 and nhTMEM16
For IST2716 and nhTMEM16, cells were resuspended in buffer A (20 mM HEPES pH 7.6 and 150 mM NaCl), supplemented with 1 mM MgCl2, protease inhibitors (PMSF, benzamidine, pepstatin A and leupeptin), DNAse and lysozyme and lysed in a HPL6 high-pressure cell disruptor at 40 kpsi. Cell debris was removed by centrifugation at 6,000g for 30 min at 4 °C. Membranes were isolated from the supernatant by ultracentrifugation at 150,000g for 1 h at 4 °C. The membrane pellet was resuspended in minimal volumes of buffer A using a Dounce homogenizer, flash-frozen in liquid nitrogen and stored at −80 °C until further use.
For protein purification, membranes were resuspended in 5× pellet volume of buffer A supplemented with protease inhibitors, 5% glycerol and 2% GDN (Anatrace). Extraction was carried out for 1.5 h at 4 °C. The insoluble fraction was removed by ultracentrifugation at 85,000g for 1 h and the solubilized protein was bound in batch to Streptactin resin (IBA) for 2 h at 4 °C. The resin was washed with 60 column volumes (CVs) of buffer B (20 mM HEPES pH 7.6, 150 mM NaCl, 5% glycerol and 0.03% GDN) using gravity flow and bound protein was eluted in 5 CVs of buffer B supplemented with 5 mM d-desthiobiotin. Protein was concentrated to 2 mg ml−1 and loaded onto a Superose 6 10/300 GL column pre-equilibrated with buffer C (20 mM HEPES pH 7.6, 150 mM NaCl and 0.03% GDN). Peak fractions were pooled and concentrated to 1–2.5 mg ml−1.
IST2fl
Purification of IST2fl was performed similarly with the exception of buffer A consisting of 40 mM HEPES pH 7.6, 500 mM NaCl, 5% glycerol and GDN being replaced by 2% tridecylmaltoside and CHS at a molar ratio of 9:1 during extraction.
TMEM16A
Purification of TMEM16A for reconstitution was performed similarly to IST2716, in the presence of 2 mM EGTA to remove free Ca2+, with the attached GFP tag cleaved using 3C protease. Murine TMEM16A was expressed in HEK293 GnTI− cells (American Type Culture Collection; tested negative for Mycoplasma contamination) grown in Hyclone medium with 1% penicillin–streptomycin, 1% FBS, 2% glutamine and 0.15% Kollophor p188 by transient transfection in suspension culture. The expression vector was mixed with PEI MAX in a 1:2.5 ratio and valproic acid was added to a final concentration of 3.5 mM after transfection. The protein was expressed at 37 °C for 48 h. Cells were harvested by centrifugation and washed once with PBS. TMEM16A was extracted in 20 mM HEPES pH 7.4, 150 mM NaCl, 2 mM EGTA and protease inhibitors (described above) by the addition of 2% GDN for 2 h at 4 °C and the insoluble fraction was removed by ultracentrifugation at 85,000g for 30 min at 4 °C. The supernatant was bound in batch to Streptactin resin (IBA) for 2 h at 4 °C and washed with 60 CVs of SEC buffer (20 mM HEPES pH 7.4, 150 mM NaCl, 2 mM EGTA and 0.03% GDN). Bound protein was eluted in wash buffer containing 5 mM d-desthiobiotin and concentrated to 1.5 mg ml−1. The c-terminal SBP–myc–GFP tag was cleaved by the addition of 0.1 mg ml−1 3C protease for 45 min at 4 °C. The cleaved protein was loaded onto a Superose 6 10/300 GL column equilibrated in SEC buffer and peak fractions were pooled and concentrated to 1 mg ml−1.
OSH6
Recombinant OSH6 was purified from S. cerevisiae (for crystallization, OSH6Δ1–30; for ITC experiments, WT OSH6) and from E. coli (for cryo-EM experiments and pulldowns). In S. cerevisiae, OSH6 was expressed similarly to IST2. Cells were resuspended in buffer D (40 mM HEPES pH 7.6, 150 mM NaCl, 1 mM MgCl2 and 0.5 mM TCEP) supplemented with protease inhibitors, DNAse and lysozyme and lysed with an HPL6 at 40 kpsi. The cytosolic fraction was cleared by ultracentrifugation at 150,000g for 1 h and bound to Streptactin resin (IBA) for 2 h as described above. The resin was washed with 60 CVs of buffer E (25 mM HEPES pH 7.6, 150 mM NaCl and 0.5 mM TCEP) and eluted with buffer E supplemented with 5 mM d-desthiobiotin. The protein was concentrated to 3 mg ml−1 and loaded onto a Superdex200 10/300 GL column equilibrated in buffer E. Peak fractions were pooled, supplemented with 5% glycerol, concentrated, aliquoted and flash-frozen. Recombinant OSH6ΔN was purified similarly, except that the pH of all buffers was adjusted to 7.8 and TCEP was omitted from the final gel-filtration step.
For OSH6 expression and purification in E. coli, BL21 transformants were grown in Luria–Bertani (LB) medium at 37 °C. Upon reaching an OD600 of 0.8, the temperature was reduced to 25 °C and gene expression was induced by the addition of 0.3 mM IPTG at an OD600 of 1.2. The protein was expressed overnight at 18 °C. Cells were harvested, resuspended in buffer D and lysed by sonication. The insoluble fraction was removed by ultracentrifugation at 150,000g for 1 h at 4 °C and the expressed protein in the supernatant was bound in batch to GSH–agarose (Genscript) for 1.5 h at 4 °C. The resin was washed with 50 CVs of buffer E and 50 µM 3C protease was added to cleave the protein from the resin for 45 min at 4 °C. The flowthrough containing the protein was concentrated and loaded onto a Superdex200 10/300 GL column equilibrated in buffer E.
Lipid preparation and nanodisc reconstitution
All lipids were purchased from Avanti, mixed in chloroform at the desired molar ratio, dried under nitrogen and washed in diethylether. After forming a thin lipid film under nitrogen, residual traces of solvent were removed in a desiccator overnight. The film was rehydrated in SEC buffer (25 mM HEPES pH 7.4 and 150 mM NaCl) for 2 h at room temperature, sonicated briefly, subjected to three freeze–thaw cycles, aliquoted and stored at −80 °C. For Nanodisc reconstitution of IST2716, the purified protein was concentrated to 0.87 mg ml−1 and reconstituted using the scaffold protein MSP1E3D1 and a POPC:POPG 3:1 lipid mix at a protein-to-lipid ratio of 1:50 with a 5× molar excess of empty nanodiscs. The protein and lipid mix were incubated in buffer C for 30 min at 4 °C. Scaffold protein was added and incubated for another 30 min. Clean SM-II Biobeads were added at a ratio of 20 mg per mg of detergent. The detergent was removed overnight at 4 °C by gentle agitation. Excess empty nanodiscs were removed by SEC on a Superose 6 10/300 GL column. The fractions containing the reconstituted protein were pooled and concentrated to 1.1 mg ml−1.
Empty nanodiscs for OSH6 binding were formed in SEC buffer using a 1:200 protein-to-lipid ratio of MSP2N2 and a 3:1 POPC:POPG lipid mix. Nanodiscs were reconstituted as described above. The detergent-free supernatant was concentrated to 3.6 mg ml−1 and incubated with a 1.2× molar excess of purified OSH6 at a final concentration of 28 µM. The sample was injected multiple times on a Superose 6 5/150GL column equilibrated in SEC buffer. The peak fractions were pooled and concentrated to 1.63 mg ml−1. Purity of the sample was assessed by SDS–PAGE.
Protein reconstitution and lipid-scrambling assay
For lipid-scrambling experiments, IST2716, nhTMEM16 and TMEM16A were reconstituted into destabilized liposomes as described previously61. In brief, liposomes at 20 mg ml−1 in resuspension buffer (20 mM HEPES pH 7.5, 300 mM KCl and 2 mM EGTA) composed of a 8:2 mixture of soybean polar extract and cholesterol with 0.5 mol.% tail-labeled NBD–PE (Avanti) were destabilized by adding 0.02% aliquots of Triton X-100. The onset of liposome destabilization was observed by the decrease of scattering at 540 nm in an ultraviolet–visible light spectrofluorometer. Protein was added at a protein-to-lipid ratio of 1:300 (nhTMEM16) or 1:200 (IST2716 and TMEM16A) to account for different reconstitution efficiencies. Reconstitution efficiency was assessed by western blot against the N-terminal myc tag of the proteins using a monoclonal anti-c-myc primary antibody (Sigma; 1:5,000 dilution) and an horseradish-peroxidase-conjugated goat anti-mouse secondary antibody (Jackson ImmunoResearch; 1:10,000 dilution). For empty liposomes, buffer C was added at an equivalent volume. Detergent was removed by four sequential additions of 20 mg SM-II Biobeads per mg of lipids overnight at 4 °C and liposomes were obtained at 170,000g for 30 min. The liposome pellet was resuspended to a final lipid concentration of 10 mg ml−1, subjected to three freeze–thaw cycles and stored at −80 °C until further use. For the assay, liposomes were diluted to a final concentration of 20 µM in assay buffer (80 mM HEPES pH 7.5, 300 mM KCl and 2 mM EGTA) in a quartz glass cuvette. After recording the fluorescence of NBD for 60 s, 30 mM dithionite was added to bleach the fluorescence of NBD in the outer leaflet of the liposomes. The fluorescence was recorded for an additional 6 min. Each trace was normalized to the NBD fluorescence level right before the addition of dithionite. Data were recorded using FluorEssence 3.0 (Horiba Scientific). Statistical analysis was performed using analysis of variance (ANOVA) in GraphPad Prism. Because the final plateau differs depending on the reconstitution efficiency, samples reconstituted from the same batch of destabilized liposomes on the same day were matched for statistical analysis.
Pulldown assays
Constructs encoding N-terminally GST tagged peptides of different length encompassing the OSH6-binding epitope of IST2 were transformed into E. coli BL21 cells. Bacteria were grown in LB medium until an OD600 of 0.8 and gene expression was induced upon addition of IPTG to a final concentration of 0.3 mM. Expression was carried out at 25 °C for 2.5 h. Cells were harvested, washed in PBS and resuspended in lysis buffer (25 mM HEPES pH 7.5, 300 mM NaCl, 10% glycerol w/v, 0.5 mM TCEP, 0.1% Triton X-100 and 1 mM MgCl2) supplemented with protease inhibitors and DNAse. Cells were lysed by sonication and the insoluble fraction was removed by centrifugation at 150,000g for 1 h at 4 °C. The cleared lysate was frozen in liquid nitrogen and stored at −80 °C until further use. The thawed lysate was diluted 1:10 in binding buffer (25 mM HEPES pH 7.4, 150 mM NaCl, 0.5 mM TCEP and 1 mg ml−1 BSA), mixed with glutathione agarose (Genscript) at a final ratio of 1:4 and incubated for 2 h at 4 °C under gentle agitation. Beads were washed five times with a 10× volume excess of binding buffer to remove unbound proteins. Then, 20 µl of coated beads were incubated with a final concentration of 5 µM OSH6 in a total volume of 40 µl for 1 h at 4 °C under gentle agitation in the presence of 1 mg ml−1 BSA. Beads were washed five times with a 50× volume excess of binding buffer without BSA and mixed with SDS sample buffer. Samples were boiled and of 5 µl of beads were analyzed by SDS–PAGE. Stained gels were scanned and band intensity was quantified by measuring the mean gray value in FIJI.
Cofiltration of nanodiscs with OSH6
The OSH6 mutant C389S was purified from S. cerevisiae as described for WT and the remaining surface-accessible cysteine (C62) was labeled with the fluorophore Cy5 using the Red-Maleimide second-generation protein labeling kit (Nanotemper). Labeled OSH6 was incubated with a 1.2× molar excess of nanodiscs composed of a MSP2N2 and POPG:POPC (3:1) mixture at a molar ratio of 1:200 and injected on a Superose 6 5/150 GL column. The eluate was collected in 100-µl fractions and the Cy5 fluorescence of the fractions was measured in a TECAN Sparc plate reader. To test for interactions between OSH6 and the scaffold protein MSP2N2 in the absence of lipids, unlabeled OSH6 was incubated with a 1.2× molar excess of MSP2N2 for 30 min and injected on a Superose 200 5/150 GL column. Peak fractions were collected and analyzed by SDS–PAGE. The band intensities of OSH6 and MSP2N2 were quantified for each fraction and plotted to confirm the lack of comigration of both proteins in the absence of lipids.
ITC
ITC measurements were performed with a MicroCal iTC200 calorimeter at 25 °C with an initial delay of 60 s and stirring at 1,000 rpm. IST2 peptides at a stock concentration of 240 µM were titrated with 20 injections of 2-µl aliquots (except the first injection containing 1.4 µl) with a spacing of 180 s, reference power of 3DP and a filter period of 5 s at high feedback mode and gain. OSH6 was filled in the reaction cell aiming at a concentration of 20 µM; the exact concentration used for data fitting was determined for each measurement from the superfluous liquid after filling the reaction cell. The reference cell contained H2O. OSH6 and the IST2732−761 peptide were in the same buffer (25 mM HEPES pH 7.5, 150 mM NaCl and 5% glycerol). The baseline was subtracted by titrating the analyte into buffer. Data were fitted in Origin using a single-site sigmoidal fit in an iterative manner until the χ2 values converged.
X-ray crystallography
Peptides used for crystallography and ITC (IST2732–761 and IST2732–757) were purchased from Genscript. OSH6ΔN was concentrated to 200 µM after affinity purification and cleaved overnight by the addition of 3C protease. The cleaved protein was incubated for 30 min with a 1.2× molar excess of the respective peptide and injected onto a Superdex 200 10/300 GL column equilibrated in 15 mM HEPES pH 7.8 and 150 mM NaCl. OSH6 peak fractions were pooled and concentrated to 18.5 mg ml−1. The presence of the peptide was confirmed by mass spectroscopy. Crystallization experiments were performed at the protein crystallization facility of the Department of Biochemistry of the University of Zurich (UZH). The final sample was supplemented with an additional 0.5× molar excess of peptide and mixed with the crystallization liquor at a ratio of 1:1 at 20 °C. OSH6–IST732−757 crystallized at 20 °C in 100 mM Bis–Tris pH 5.5, 200 mM KCl, 75 mM MgCl2 and 28–29% PEG 400. OSH6–IST732−761 crystallized at 4 °C in 100 mM Bis–Tris pH 5.5, 200 mM KCl, 20 mM MgCl2 and 28–30% PEG400. Crystals formed within 10 days and were cryoprotected by a stepwise increase in the PEG400 concentration to 35% before flash-freezing in liquid nitrogen. X-ray diffraction data were collected at the Petra III p13 beamline at European Molecular Biology Laboratory Hamburg and processed with XDS62 and EDNA63. The crystals were of space group C2221 with one copy of the complex in the asymmetric unit and data extended to beyond 2.0 Å for both datasets (Table 2). The structure was solved using molecular replacement with Phaser64 implemented in the PHENIX software package65 using the coordinates of OSH6 (Protein Data Bank (PDB) 4B2Z) as the start model. Iterative model building and refinement were performed in Coot66 and PHENIX65, respectively.
Sample preparation for cryo-EM and data collection
For the assembly of the IST2–OSH6 complex, affinity-purified IST2fl was mixed with a 1.2× molar excess of OSH6 purified from E. coli and the mixture was loaded onto a Superose 6 5/150 GL column equilibrated in 20 mM HEPES pH 7.6, 150 mM NaCl and 0.03% GDN. The peak fractions corresponding to the complex were pooled and concentrated to 1.08 mg ml−1; then, 2.5 µl of sample was applied to a Quantifoil UltrAuFoil R1.2/1.2 300-mesh grid, blotted for 3 s and plunge-frozen in a liquid ethane and propane mix using a Mark V Vitrobot. For IST2716 in GDN, the protein was purified as described above and concentrated to 2.56 mg ml−1; then, 2.5 µl of sample was applied to a Quantifoil R1.2/R1.3 Au 200-mesh grid and blotted for 2 s before plunge-freezing. Further samples were prepared similarly with the following modifications: IST2716–MSP1E3D1 was concentrated to 1.09 mg ml−1 and applied to a Quantifoil UltrAuFoil R1.2/1.2 300-mesh grid; OSH6–MSP2N2 was concentrated to 1.63 mg ml−1 and blotted for 3 s before plunge-freezing. Data were collected on a Titan Krios G3i equipped with a postcolumn energy filter and a Bioquantum K3 detector at a physical pixel size of 0.651 Å at a magnification of 130,000 using EPU version 2.9 (Thermo Fisher).
Cryo-EM data processing, model building and refinement
All raw videos were corrected for gain, motion and contrast transfer function (CTF) in cryoSPARC version 4 (ref. 67). Micrographs with poor CTF estimation, high drift and ice thickness were discarded.
IST2–OSH6 complex
For the IST2–OSH6 dataset, the blob picker function was used to generate templates from a random subset of 1,000 micrographs, which represented different particle sizes and the heterogeneity of the dataset; these micrographs were subsequently used for template picking. After an initial 2D classification, the particles were sorted into three subsets on the basis of their size and the number of particles in the box. Classes exhibiting features of the IST2 transmembrane domain were subjected to multiclass ab initio reconstruction yielding a single class with well-defined transmembrane density. Particles sorted into this class were used to train a Topaz model to increase the number of picks68. After re-extraction of the picked particles and further cleaning of the particle set using 2D classification, the particles were again subjected to ab initio reconstruction and the single good class was further refined using heterogeneous refinement with a decoy class and homogeneous refinement in C1. Because the map appeared symmetric, a final round of nonuniform refinement with imposed C2 symmetry was performed to obtain an improved final reconstruction. Classes with small particles were subjected to separate ab initio reconstruction, resulting in two classes with triangular shape, which resembled previously solved crystal structures of OSH6 in shape and size. Because pooling of those classes did not improve the result, the best class was chosen for homogeneous refinement with a large, soft mask, resulting in an envelope of the 50-kDa protein at 11-Å resolution.
IST2716–GDN
For the IST2716 structure in GDN, templates for template picking were generated from a subset of 100 micrographs using the blob picker function. Picked particles were subjected to several rounds of 2D classification until the resolution of classes did not improve further and particles were no longer sorted into junk classes. The clean particle set was subjected to ab initio reconstruction with two classes, resulting in a single good ab initio model, which was refined using heterogeneous refinement with a decoy class generated from the same dataset and subjected to nonuniform refinement in C1. Upon reaching the Nyquist limit, particles contributing to the final reconstruction were unbinned and refined with global CTF refinement and C2 symmetry imposed to obtain the final reconstruction.
IST2716–nanodisc
For the IST2716 structure in 1E3D1 nanodiscs, templates were generated from a subset of micrographs using blob picker. The initial particle set was cleaned by using iterative rounds of 2D classification. Particles sorted into 2D classes with visible transmembrane density were unbinned through re-extraction and, after further 2D classification, subjected to multiclass ab initio reconstruction, resulting in two classes with well-defined transmembrane density differing in the apparent size of the nanodisc, as well as visibility of the cytosolic bridging helix and features of the lower half of α6. The two distinct classes and two junk classes obtained from the ab initio reconstruction were used as input models for the sorting of all particles by heterogeneous refinement and then refined separately using nonuniform refinement. In the case of the narrow conformation, C2 symmetry improved the resolution and was, therefore, applied in the final refinement step. In the cases of the IST2fl–OSH6 complex and IST2716ND structures, neither local nor global CTF refinement improved the maps.
OSH6–nanodisc
For the OSH6–nanodisc dataset, the initial particle picking was performed using blob picker in cryoSPARC. The particle selection was improved by sequential rounds of 2D classification and Topaz picking. Particles were subjected to ab initio classification, which resulted in classes showing free nanodiscs and nanodiscs bound to a molecule with approximate size of OSH6. The particle set was further sorted in 3D using heterogeneous refinement to exclude unbound nanodiscs and subjected to an initial refinement in cryoSPARC67. The refined particles were exported and re-extracted in RELION version 5 (ref. 69,70) with a smaller box size. The particles were then subjected to two iterative rounds of unmasked 3D classification without alignment. Particles from classes showing higher resolution features of OSH6 were re-extracted with recentering on OSH6 and subjected to another round of 3D classification without alignment with a soft mask including the OSH6 density. The two best classes were pooled and aligned using the autorefine option of RELION version 5. The density corresponding to the nanodisc was subtracted by masking the OSH6 density. The particles were subjected to a final refinement and RELION postprocessing. For all described datasets, structures were built into cryo-EM maps in Coot66 and refined in PHENIX71.
To identify the optimal placement of the crystal structure of OSH6 within the low-resolution OSH6–nanodisc map, a model map from the OSH6 structure (PDB 9HDK), excluding the IST2 peptide and the lid helix, was calculated at 8-Å resolution. This map was used for a global search in ChimeraX72 to find the best correlation with the OSH6–nanodisc map. After optimization, fits were considered unique if they differed by a cluster angle greater than 6° or a shift greater than 3 Å. For each unique fit, the correlation was plotted against the proportion of the model map located within the reference map. Upon inspection of the four placements with highest correlation, only the top-scoring position showed optimal overlap with the protein part of the density. The best fit was then used to position the atomic model of OSH6 within the reoriented model map and subsequently refined using rigid-body refinement in PHENIX71, which did not visibly alter the position of the initial placement. This fit is shown in Fig. 4d. Figures containing molecular structures and densities were prepared with DINO (http://www.dino3d.org) and ChimeraX72. Details of the data-processing workflow are shown in the Extended Data figures.
Yeast growth assays
IST2 constructs were transformed into a S. cerevisiae ist2;psd1-KO strain (AWY327), generously provided by C. Loewen, using the LiAc–SS carrier DNA–PEG method60 and plated onto SC −Ura selection medium containing 1 mM choline bitartrate. Plates were kept at 4 °C for a maximum of 4 weeks. Single colonies were picked and grown in liquid SC −Ura medium containing 1 mM choline to mid-log phase. Cells were harvested by centrifugation and the pellet was washed three times with sterile water to remove all traces of choline. Cells were subsequently diluted to an OD600 of 0.35 and a tenfold dilution series was spotted in three replicates on solid medium containing 2% dextrose and 2% galactose or 1 mM choline. Plates were incubated at 30 °C for 3–4 days. The experiment was repeated three times with different transformants for each construct. For evaluation, plates were imaged in a Vilber Fusion FX using bright-field illumination. The background was subtracted in FIJI and the mean gray value of each spot was quantified using a circular mask of equal size for all replicates and all constructs, unless otherwise stated. The quantification of the 1:100 dilution was chosen for comparison between constructs as it was found in the linear range of the dilution series (Extended Data Fig. 9b). For each replicate, all constructs were normalized to IST2 WT. Statistical analysis was performed in GraphPad Prism using ANOVA.
Mathematical modeling
For the mathematical modeling of the linkers as entropic springs, the potential energy associated with loading and unloading the worm-like chains was calculated using Eq. (1) adapted from previous studies55,73:
In this equation, lp corresponds to the persistence length of a single amino acid (0.38 nm) as determined by atomic force microscopy and spectroscopy54, k is the Boltzmann constant, Lc is the contour length of the respective linker segment (calculated as nresidues × lp), T is a temperature of 303.15 K and L0 is a value of 0 nm, reflecting a fully relaxed state of the linker. The potential energy (U) was calculated for extensions along the trajectory between the ER and PM (with Li ranging from 0 to 22 nm in 0.1-nm increments). The total potential energy was calculated as the sum of the potential energies for SpringN and SpringC at the respective position of Li.
The relative distribution P of OSH6 between the two membranes was derived from Eq. (2).
The values were normalized for each construct and plotted.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Cryo-EM density maps and atomic models were deposited to the EM Data Bank and the PDB, respectively, under accession codes EMD-52170 (IST2716GDN), EMD-53210 (IST2716ND wide), EMD-53211 (IST2716ND narrow) and EMD-53200 (IST2fl) and 9HDH (OSH6ΔN/IST2732-761), 9HDK (OSH6ΔN/IST2732-756), 9HHE (IST2716GDN), 9QJT (IST2716ND wide), 9QJU (IST2716ND narrow) and 9QJA (IST2fl), respectively. Raw data of lipid-scrambling and yeast growth assays were deposited to the DRYAD database (https://doi.org/10.5061/dryad.8w9ghx40q). Source data are provided with this paper.
References
Drin, G. Creating and sensing asymmetric lipid distributions throughout the cell. Emerg. Top. Life Sci. 7, 7–19 (2023).
Holthuis, J. C. & Menon, A. K. Lipid landscapes and pipelines in membrane homeostasis. Nature 510, 48–57 (2014).
van Meer, G., Voelker, D. R. & Feigenson, G. W. Membrane lipids: where they are and how they behave. Nat. Rev. Mol. Cell Biol. 9, 112–124 (2008).
Lenoir, G., D’Ambrosio, J. M., Dieudonne, T. & Copic, A. Transport pathways that contribute to the cellular distribution of phosphatidylserine. Front. Cell Dev. Biol. 9, 737907 (2021).
Harayama, T. & Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell Biol. 19, 281–296 (2018).
Leventis, P. A. & Grinstein, S. The distribution and function of phosphatidylserine in cellular membranes. Annu. Rev. Biophys. 39, 407–427 (2010).
Raychaudhuri, S. & Prinz, W. A. The diverse functions of oxysterol-binding proteins. Annu. Rev. Cell Dev. Biol. 26, 157–177 (2010).
Pietrangelo, A. & Ridgway, N. D. Bridging the molecular and biological functions of the oxysterol-binding protein family. Cell. Mol. Life Sci. 75, 3079–3098 (2018).
Im, Y. J., Raychaudhuri, S., Prinz, W. A. & Hurley, J. H. Structural mechanism for sterol sensing and transport by OSBP-related proteins. Nature 437, 154–158 (2005).
Maeda, K. et al. Interactome map uncovers phosphatidylserine transport by oxysterol-binding proteins. Nature 501, 257–261 (2013).
Moser von Filseck, J. et al.Phosphatidylserine transport by ORP/Osh proteins is driven by phosphatidylinositol 4-phosphate. Science 349, 432–436 (2015).
Hankins, H. M., Baldridge, R. D., Xu, P. & Graham, T. R. Role of flippases, scramblases and transfer proteins in phosphatidylserine subcellular distribution. Traffic 16, 35–47 (2015).
von Filseck, M., Vanni, J., Mesmin, S., Antonny, B. & Drin, B. G.A phosphatidylinositol-4-phosphate powered exchange mechanism to create a lipid gradient between membranes. Nat. Commun. 6, 6671 (2015).
Hoffmann, P. C. et al. Tricalbins contribute to cellular lipid flux and form curved ER–PM contacts that are bridged by rod-shaped structures. Dev. Cell 51, 488–502 (2019).
Manford, A. G., Stefan, C. J., Yuan, H. L., Macgurn, J. A. & Emr, S. D. ER-to-plasma membrane tethering proteins regulate cell signaling and ER morphology. Dev. Cell 23, 1129–1140 (2012).
Saheki, Y. & De Camilli, P. Endoplasmic reticulum–plasma membrane contact sites. Annu. Rev. Biochem. 86, 659–684 (2017).
D’Ambrosio, J. M. et al. Osh6 requires Ist2 for localization to ER–PM contacts and efficient phosphatidylserine transport in budding yeast. J. Cell Sci. 133, jcs243733 (2020).
Wong, A. K. O., Young, B. P. & Loewen, C. J. R.Ist2 recruits the lipid transporters Osh6/7 to ER–PM contacts to maintain phospholipid metabolism. J. Cell Biol. 220, e201910161 (2021).
Babu, M. et al. Interaction landscape of membrane-protein complexes in Saccharomyces cerevisiae. Nature 489, 585–589 (2012).
Kralt, A. et al. Intrinsically disordered linker and plasma membrane-binding motif sort Ist2 and Ssy1 to junctions. Traffic 16, 135–147 (2015).
Brunner, J. D., Schenck, S. & Dutzler, R. Structural basis for phospholipid scrambling in the TMEM16 family. Curr. Opin. Struct. Biol. 39, 61–70 (2016).
Suzuki, J. et al. Calcium-dependent phospholipid scramblase activity of TMEM16 protein family members. J. Biol. Chem. 288, 13305–13316 (2013).
Whitlock, J. M. & Hartzell, H. C. Anoctamins/TMEM16 proteins: chloride channels flirting with lipids and extracellular vesicles. Annu Rev. Physiol. 79, 119–143 (2017).
Brunner, J. D., Lim, N. K., Schenck, S., Duerst, A. & Dutzler, R. X-ray structure of a calcium-activated TMEM16 lipid scramblase. Nature 516, 207–212 (2014).
Malvezzi, M. et al. Ca2+-dependent phospholipid scrambling by a reconstituted TMEM16 ion channel. Nat. Commun. 4, 2367 (2013).
Paulino, C., Kalienkova, V., Lam, A. K. M., Neldner, Y. & Dutzler, R. Activation mechanism of the calcium-activated chloride channel TMEM16A revealed by cryo-EM. Nature 552, 421–425 (2017).
Falzone, M. E. et al. Structural basis of Ca2+-dependent activation and lipid transport by a TMEM16 scramblase. eLife 8, e43229 (2019).
Alvadia, C. et al. Cryo-EM structures and functional characterization of the murine lipid scramblase TMEM16F. eLife 8, e44365 (2019).
Kalienkova, V. et al. Stepwise activation mechanism of the scramblase nhTMEM16 revealed by cryo-EM. eLife 8, e44364 (2019).
Falzone, M. E. et al. TMEM16 scramblases thin the membrane to enable lipid scrambling. Nat. Commun. 13, 2604 (2022).
Bushell, S. R. et al. The structural basis of lipid scrambling and inactivation in the endoplasmic reticulum scramblase TMEM16K. Nat. Commun. 10, 3956 (2019).
Bethel, N. P. & Grabe, M. Atomistic insight into lipid translocation by a TMEM16 scramblase. Proc. Natl Acad. Sci. USA 113, 14049–14054 (2016).
Jiang, T., Yu, K., Hartzell, H. C. & Tajkhorshid, E.Lipids and ions traverse the membrane by the same physical pathway in the nhTMEM16 scramblase. eLife 6, e28671 (2017).
Khelashvili, G. et al. Membrane lipids are both the substrates and a mechanistically responsive environment of TMEM16 scramblase proteins. J. Comput. Chem. 41, 538–551 (2020).
Stansfeld, P. J. et al. MemProtMD: automated insertion of membrane protein structures into explicit lipid membranes. Structure 23, 1350–1361 (2015).
Fairn, G. D., Curwin, A. J., Stefan, C. J. & McMaster, C. R. The oxysterol binding protein Kes1p regulates Golgi apparatus phosphatidylinositol-4-phosphate function. Proc. Natl Acad. Sci. USA 104, 15352–15357 (2007).
Lipp, N. F. et al. An electrostatic switching mechanism to control the lipid transfer activity of Osh6p. Nat. Commun. 10, 3926 (2019).
Maass, K. et al. A signal comprising a basic cluster and an amphipathic α-helix interacts with lipids and is required for the transport of Ist2 to the yeast cortical ER. J. Cell Sci. 122, 625–635 (2009).
Reinisch, K. M. & Prinz, W. A.Mechanisms of nonvesicular lipid transport. J. Cell Biol. 220, e202012058 (2021).
Melia, T. J. & Reinisch, K. M.A possible role for VPS13-family proteins in bulk lipid transfer, membrane expansion and organelle biogenesis. J. Cell Sci. 135, jcs259357 (2022).
Li, P., Lees, J. A., Lusk, C. P. & Reinisch, K. M.Cryo-EM reconstruction of a VPS13 fragment reveals a long groove to channel lipids between membranes. J. Cell Biol. 219, e202001161 (2020).
Wang, Y. et al. Structural basis for lipid transfer by the ATG2A–ATG9A complex. Nat. Struct. Mol. Biol. 32, 35–47 (2024).
Valverde, D. P. et al. ATG2 transports lipids to promote autophagosome biogenesis. J. Cell Biol. 218, 1787–1798 (2019).
Jeong, H., Park, J., Jun, Y. & Lee, C. Crystal structures of Mmm1 and Mdm12–Mmm1 reveal mechanistic insight into phospholipid trafficking at ER–mitochondria contact sites. Proc. Natl Acad. Sci. USA 114, E9502–E9511 (2017).
Wozny, M. R. et al. In situ architecture of the ER–mitochondria encounter structure. Nature 618, 188–192 (2023).
Wong, L. H., Gatta, A. T. & Levine, T. P. Lipid transfer proteins: the lipid commute via shuttles, bridges and tubes. Nat. Rev. Mol. Cell Biol. 20, 85–101 (2019).
Olkkonen, V. M. OSBP-related protein family in lipid transport over membrane contact sites. Lipid Insights 8, 1–9 (2015).
de Saint-Jean, M. et al. Osh4p exchanges sterols for phosphatidylinositol 4-phosphate between lipid bilayers. J. Cell Biol. 195, 965–978 (2011).
Chung, J. et al. PI4P/phosphatidylserine countertransport at ORP5- and ORP8-mediated ER–plasma membrane contacts. Science 349, 428–432 (2015).
Ghanbarpour, A., Valverde, D. P., Melia, T. J. & Reinisch, K. M.A model for a partnership of lipid transfer proteins and scramblases in membrane expansion and organelle biogenesis. Proc. Natl Acad. Sci. USA 118, e2101562118 (2021).
Dittman, J. S. & Menon, A. K. Speed limits for nonvesicular intracellular sterol transport. Trends Biochem. Sci. 42, 90–97 (2017).
Jamecna, D. et al. An intrinsically disordered region in OSBP acts as an entropic barrier to control protein dynamics and orientation at membrane contact sites. Dev. Cell 49, 220–234 (2019).
Du, X. et al. A role for oxysterol-binding protein-related protein 5 in endosomal cholesterol trafficking. J. Cell Biol. 192, 121–135 (2011).
Rico, F., Gonzalez, L., Casuso, I., Puig-Vidal, M. & Scheuring, S. High-speed force spectroscopy unfolds titin at the velocity of molecular dynamics simulations. Science 342, 741–743 (2013).
Scheuring, S. Forces and energetics of the canonical tetrameric cation channel gating. Proc. Natl Acad. Sci. USA 120, e2221616120 (2023).
Petkovic, M., Oses-Prieto, J., Burlingame, A., Jan, L. Y. & Jan, Y. N. TMEM16K is an interorganelle regulator of endosomal sorting. Nat. Commun. 11, 3298 (2020).
Jha, A. et al. Anoctamin 8 tethers endoplasmic reticulum and plasma membrane for assembly of Ca2+ signaling complexes at the ER/PM compartment. EMBO J. 38, e101452 (2019).
Geertsma, E. R. & Dutzler, R. A versatile and efficient high-throughput cloning tool for structural biology. Biochemistry 50, 3272–3278 (2011).
Zheng, L., Baumann, U. & Reymond, J. L. An efficient one-step site-directed and site-saturation mutagenesis protocol. Nucleic Acids Res. 32, e115 (2004).
Gietz, R. D. & Schiestl, R. H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2, 31–34 (2007).
Geertsma, E. R., Nik Mahmood, N. A., Schuurman-Wolters, G. K. & Poolman, B. Membrane reconstitution of ABC transporters and assays of translocator function. Nat. Protoc. 3, 256–266 (2008).
Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 66, 125–132 (2010).
Incardona, M. F. et al. EDNA: a framework for plugin-based applications applied to X-ray experiment online data analysis. J. Synchrotron Radiat. 16, 872–879 (2009).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Adams, P. D. et al. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr. D Biol. Crystallogr. 58, 1948–1954 (2002).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132 (2004).
Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296 (2017).
Bepler, T. et al. Positive-unlabeled convolutional neural networks for particle picking in cryo-electron micrographs. Nat. Methods 16, 1153–1160 (2019).
Zivanov, J. et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife 7, e42166 (2018).
Burt, A. et al. An image processing pipeline for electron cryo-tomography in RELION-5. FEBS Open Bio 14, 1788–1804 (2024).
Afonine, P. V. et al. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr. D Struct. Biol. 74, 531–544 (2018).
Pettersen, E. F. et al. UCSF ChimeraX: structure visualization for researchers, educators, and developers. Protein Sci. 30, 70–82 (2021).
Bustamante, C., Marko, J. F., Siggia, E. D. & Smith, S. Entropic elasticity of λ-phage DNA. Science 265, 1599–1600 (1994).
Smart, O. S., Neduvelil, J. G., Wang, X., Wallace, B. A. & Sansom, M. S.HOLE: a program for the analysis of the pore dimensions of ion channel structural models. J. Mol. Graph. 14, 354–360 (1996).
Acknowledgements
This research was supported by a grant from the Swiss National Science Foundation (310030_215236 to R.D.). M.A. was supported by a Candoc grant of the University of Zurich (project number K-41106-05-01). We thank S. Scheuring for making us aware of the analogy of the IST2 C terminus to an entropic spring and for his help in the analysis. Cryo-EM data were collected at the Center for Microscopy and Image Analysis of UZH. P. Szwedziak is acknowledged for support during microscope operation. We thank B. Blattman and G. Kurduldu of the Protein Crystallization Center at UZH for their support with crystallization. X-ray crystallography data were collected at beamline P13 at the Deutsches Elektronen-Synchrotron in Hamburg. We acknowledge the support of F. Nai, R. Bedi and staff of the P13 beamline during data collection. We thank B. Wimmer for advice regarding cryo-EM data processing. The yeast strains used for complementation experiments were generously provided by C. J. R. Loewen (University of British Columbia). We acknowledge members of the R.D. group for their help at various stages of the project.
Author information
Authors and Affiliations
Contributions
M.A. expressed and purified the proteins, collected and analyzed the cryo-EM and X-ray data and carried out the lipid transport and yeast complementation assays. A.S. established the yeast complementation assay, performed the ITC measurements and supported M.A. in various experiments. M.A. and R.D. jointly planned the experiments, analyzed the data and wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Structural & Molecular Biology thanks David Sauer, Anant Menon and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available. Primary Handling Editor: Katarzyna Ciazynska, in collaboration with the Nature Structural & Molecular Biology team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Cryo-EM data processing of IST2 in detergent in complex with OSH6.
a, Representative micrograph (n = 13,000), 2D classes and data processing workflow for IST2/OSH6 complex. Particles were sorted in 2D into three categories based on size. IST2 and OSH6 particles were then processed separately as indicated. Data was processed using cryoSPARC. b, Local resolution of the final IST2 map computed in cryoSPARC. c, GSFSC curve used for resolution estimation of final maps of IST2. d, GSFSC curve used for resolution estimation of final maps of OSH6. e, Angular viewing direction distribution of particles contributing to the final map of IST2fl. f, Angular viewing direction distribution of particles contributing to the final map of OSH6.
Extended Data Fig. 2 Biochemical properties of IST2716 and lipid transport experiments.
a, Size exclusion chromatogram of a sample of IST2716 and nhTMEM16 used for liposome reconstitution. b, Western Blots against n-terminal myc tag of nhTMEM16 and IST2716 reconstituted into liposomes for scrambling assays (n = 3, all shown). c, Scheme of the lipid scrambling assay. ‘red.’ refers to the addition of dithionite to the outside of liposomes. Fluorescent lipids are indicated as yellow, lipids with bleached fluorophore as grey spheres, a scramblase mediating the bidirectional diffusion of lipids between leaflets as cyan box. d-f, Scrambling data from three independent reconstitutions that are combined in Fig. 1f, g. The displayed nhTMEM16 and IST2716 preparations were reconstituted into the same batch of destabilized liposomes. Empty liposomes of the same batch are shown as negative control (neg.). Recordings show mean of three technical replicates, errors are s.d. g-h, Size exclusion chromatograms and SDS PAGE (inset, n = 1) of IST2716 in detergent (g) and after reconstitution in lipid nanodiscs (h) used as samples for the cryo-EM datasets shown in Extended Data Figs. 3 and 4, respectively. Dashed lines indicate pooled fractions.
Extended Data Fig. 3 Cryo-EM data processing of the IST2 transmembrane domain in detergent.
a, Representative CTF-corrected micrograph for IST2716GDN (n = 22,507). b, Representative final 2D classes for IST2716GDN. c, Data processing workflow for IST2716GDN performed in cryoSPARC. d, GSFSC curve used for resolution estimation of final map of IST2716GDN. e, Angular viewing direction distribution of particles contributing to the final map of IST2716GDN. f, Local resolution estimation for IST2716GDN computed in cryoSPARC.
Extended Data Fig. 4 Cryo-EM data processing of the IST2 transmembrane domain in lipid nanodiscs.
a, Representative CTF-corrected micrograph for IST2716ND. b, Representative final 2D classes for IST2716ND. c, Data processing workflow for IST2716ND performed in cryoSPARC leading to two distinct reconstructions based on nanodisc size, termed ‘narrow’ and ‘wide’. d, GSFSC curve used for resolution estimation of final map of IST2716ND ‘narrow’ conformation. e, Angular viewing direction distribution of particles contributing to the final map of IST2716ND ‘narrow’. f, Local resolution estimation for IST2716ND ‘narrow’ computed in cryoSPARC. g, GSFSC curve used for resolution estimation of final map of IST2716ND ‘wide’ conformation. h, Angular viewing direction distribution of particles contributing to the final map of IST2716ND ‘wide’. i, Local resolution estimation for IST2716ND ‘wide’ computed in cryoSPARC.
Extended Data Fig. 5 Features of the IST2 transmembrane domain.
a, Superposition of NTDs of IST2 and nhTMEM16. Secondary structure elements of IST2 are labeled. b, Location of the NTD (red) in the IST2 subunit. Ribbon representation of the subunit is shown left. Blowup of the marked region right. c, Dimer interface. Left, IST2 subunit with α10 of the contacted subunit displayed as ribbon. Boxes indicate magnified regions shown right. Top, interacting residues between symmetry related helices α10 and bottom, the α10-α3 interface. Sidechains of interacting residues are shown as sticks. d, Features of the detergent micelle displaying a pronounced distortion at both entrances to the lipid permeation path. The view is towards the subunit cavity from within the membrane. The blurred density corresponding to the protein envelope including the detergent micelle is superimposed on the high-resolution density of the protein (grey) and colored according to its distance to the center of the presumed membrane plane (yellow, distances in nm). The distorted and undistorted widths of the detergent micelle are indicated by dashed and solid lines, respectively. Right panel shows view with the front part clipped. e, Bulky lipid-like density (magenta) surrounding the membrane-inserted region of IST2716 detergent dataset.
Extended Data Fig. 6 Conformational properties of the IST2 transmembrane domain.
a, Cryo-EM densities of IST2716 constructs in nanodiscs showing ‘narrow’ (left) and ‘wide’ (right) particle reconstructions. A view of the narrow reconstruction along the dimer (bottom) at larger contour illustrates presumable density of the scaffolding protein. Membrane deformations are indicated (#, *). b, View towards the subunit cavity of IST2 structures obtained from the ‘narrow’ (top) and ‘wide’ (bottom) nanodisc reconstructions. Left, cryo-EM density superimposed on the refined structure, right, superposition with the corresponding structure in detergent. Proteins are shown as ribbons. c, Superposition of subunits of the ‘open’ conformations of IST2 and nhTMEM16 (PDBID 6QM9). d, Diameter of the open subunit cavity in IST2 and nhTMEM16 measured by Hole74. Inset (right) indicates the start and end of mapped positions in the cavities of both proteins. e, Superposition of the densities of the membrane inserted part of the IST2/OSH6 complex and the IST2716 subunit (low pass filtered to the same resolution) illustrates a different conformation of α4 and α6. A blowup of the marked region is shown right. f, View towards the subunit cavity of the collapsed conformation of IST2 observed in the IST2/OSH6 complex. Left, cryo-EM density superimposed on the refined structure, right, superposition with IST2716 in detergent. Proteins are shown as ribbons.
Extended Data Fig. 7 Biochemical and structural characterization of the IST2/OSH6 interaction.
a, Left, SDS-PAGE gel of pulldowns of OSH6 with fusion-constructs of peptides carrying the indicated region of IST2 fused to GST by binding to GSH resin (n = 1). Neg. refers to GST not containing any attached peptide. Right, intensity ratio of bands corresponding to OSH6 and the respective GST-IST2 fusion constructs. Dashed line indicates value of negative control (neg.). b, Comparison of size exclusion chromatography profiles of purified OSH6 expressed in S. cerevisiae and E. coli showing the same elution volume of the main protein peak. Inset shows SDS-PAGE gel of OSH6 purified from E. coli (n = 1). c, d, Binding of a peptide encompassing residues 732-761 of IST2 (IST2732-761, showing an independent biological replicate from data displayed in Fig. 3a) (c) or residues 732-756 (IST2732-756) (d) to OSH6. Binding isotherms were obtained from titrations of either peptide to OSH6. The data show fits to a model assuming a single binding site with Kd valued of 714.0 ± 86.7 nM (IST2732-761) and 1.1 ± 0.17 μM (IST2732-756). The dashed line in b refers to the fit of IST2732-761. Errors represent fitting errors. The thermograms are shown as inset (right). e, Superposition of OSH6 structures obtained in absence (cyan, PDBID 4B2Z) and in complex with IST2732-761 (red). The displayed region illustrates conformational differences in the lid helix α1 covering the lipid binding site. f, 2Fo-Fc density (grey mesh) superimposed on different regions of the OSH6/IST2 interface. g, Structural details of IST2/OSH6 interactions. h, Sequence alignment of OSH6 binding region of IST2 of three Saccheromyces species (S. paradoxus, S. kudriavzeii, S. cerevisiae). Residues in contact with OSH6 are colored in green. Numbering refers to the S. cerevisiae IST2. i, Sequence alignment of IST2 interactions of OSH6 and OSH7 of the same strains. Residues in contact with IST2 are colored in yellow. Numbering refers to the S. cerevisiae OSH6. j, Intra- and intermolecular interactions of the Arg 750 sidechain of IST2. k, Pulldown of OSH6 by a peptide of the IST2732-761 region carrying the point mutation R750A in comparison to WT. Experiments were caried out as described for a. Bars show mean of three technical replicates, errors are s.d. l, Binding of a peptide encompassing residues 732-761 of IST2 carrying the mutation R750A to OSH6. Binding isotherms were obtained from isothermal titrations of the peptide to OSH6. Dashed line refers to the fit of IST2732-761. The thermogram is shown as inset (right).
Extended Data Fig. 8 Structural characterization of the OSH6 interaction with lipid nanodiscs.
a, Representative CTF-corrected micrograph for OSH6ND (n = 19.283). b, 2D classes for OSH6ND calculated in cryoSPARC representing the OSH6-bound fraction of nanodiscs used for initial 3D reconstruction. c, Data processing workflow for OSH6ND performed in cryoSPARC and Relion v5. d, Results of 2D classification without alignment of refined particles in the final map performed in Relion v5. e, GSFSC curve of final map displayed in (c) computed in Relion v5. f, Euler angle distribution for final 3D reconstruction shown for two orientations of the map. g, Positioning of the OSH6 model into the OSH6ND map. Left, correlation between the experimentally determined map (shown in c) and 100 representative placements covering all orientations of a density map calculated from the OSH6 structure. The initial placement was followed by a local optimization to improve the orientation resulting in 50 unique clusters. The red data point represents the fit shown in Fig. 4d. Center, close-up of the top four hits obtained. Right, histogram of hits within the top four unique clusters showing the largest population for the top hit. h, i, Size-exclusion chromatography analysis of OSH6 and MSP2N2 in the absence of lipids. h, Data points show normalized band intensity from SDS-PAGE analysis of SEC fractions shown in i, collected at the indicated retention volumes. h,i, The experiment was performed once.
Extended Data Fig. 9 Functional characterization of IST2 constructs by yeast complementation assays.
a, Scheme of the IST2/OSH6 complex bridging the ER and the PM. The residue range of the indicated parts of IST2 is shown. b, Normalized mean gray value quantification of spot assay dilution series of IST2 transformed into ist2/psd1 KO cells under inducing conditions (2% Galactose, n = 27). Dashed line indicates 1:100 dilution, which was used for evaluation and comparison of all other constructs. For each dilution series, values were normalized to the maximum and minimum mean gray value. Data was fit with a sigmoidal model using GraphPad prism. c-f, Yeast spot dilution series used for FIJI-based evaluation shown in Fig.6. Shown are c, WT in comparison to the construct IST2921 lacking the C-terminal attachment site to the PM and constructs where the OSH binding site was moved by 83 residues in N or C-terminal direction of the linker, d, mutations in the OSH6 binding region, e, constructs with decreasing liker length, f, fusions of OSH6 into the linker. Images show representative 10-fold dilution series of n = 3×3 (n = 2×3 for Fusion trunc. and Fusion ext.) replicates for plates containing 2% galactose and 1 mM choline for each IST2 construct transformed into ist2/psd1 KO cells. Neg. refers to cells transfected with pYESnSM3 vector only. Plates were imaged with a Vilber Fusion FX detection system and the background was subtracted using FIJI. g, Size exclusion chromatogram of the purified IST2-OSH6 fusion construct (Fusion) in comparison to IST2 WT.
Extended Data Fig. 10 Mathematical Model of the IST2 linker.
a, Scheme illustrating the setup for the mathematical modelling of the IST2 C-terminal region as two entropic springs (SpringN and SpringC). b, Elastic worm-like chain (WLC) potential energy for SpringN (left) and SpringC (right) upon their extension in wild-type (WT, IST2) and mutant constructs. c, Distribution of bound OSH6 along the axis between the endoplasmic reticulum (ER) and the plasma membrane (PM) in constructs of different lengths (100% refers to WT). d, Left, potential energy of the C-terminal region of IST2 during the shuttle of OSH6 between the ER and the PM in IST2 constructs with a relocated OSH6 binding site. Right, distribution of bound OSH6 along the z-axis between ER and PM in constructs with a relocated OSH6 binding site. WT (IIST2) is indicated as dashed line. c, d, The correspondence of energies or distributions to states during transport is indicated (top).
Supplementary information
Supplementary Information
Supplementary Figs. 1–3 and Table 1 (list of primers).
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 1
Unprocessed SDS–PAGE gels.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 4
Unprocessed SDS–PAGE gels.
Source Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 2
Unprocessed blots and SDS–PAGE gels.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 7
Unprocessed SDS–PAGE gels and thermograms.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 8
Unprocessed SDS–PAGE gels.
Source Data Extended Data Fig. 9
Statistical source data.
Source Data Extended Data Fig. 9
Unprocessed SDS–PAGE gels and yeast growth plate images.
Source Data Extended Data Fig. 10
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Arndt, M., Schweri, A. & Dutzler, R. Structural basis for lipid transport at membrane contact sites by the IST2–OSH6 complex. Nat Struct Mol Biol 32, 2219–2230 (2025). https://doi.org/10.1038/s41594-025-01660-z
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41594-025-01660-z
