
Abstract
During COVID-19 pandemic, cases of postvaccination infections and restored SARS-CoV-2 virus have increased after full vaccination, which might be contributed to by immune surveillance escape or virus rebound. Here, artificial linear 9-mer human leucocyte antigen (HLA)-restricted UC peptides were designed based on the well-conserved S2 region of the SARS-CoV-2 spike protein regardless of rapid mutation and glycosylation hindrance. The UC peptides were characterized for its effect on immune molecules and cells by HLA-tetramer refolding assay for HLA-binding ability, by HLA-tetramer specific T cell assay for engaged cytotoxic T lymphocytes (CTLs) involvement, by HLA-dextramer T cell assay for B cell activation, by intracellular cytokine release assay for polarization of immune response, Th1 or Th2. The specific lysis activity assay of T cells was performed for direct activation of cytotoxic T lymphocytes by UC peptides. Mice were immunized for immunogenicity of UC peptides in vivo and immunized sera was assay for complement cytotoxicity assay. Results appeared that through the engagement of UC peptides and immune molecules, HLA-I and II, that CTLs elicited cytotoxic activity by recognizing SARS-CoV-2 spike-bearing cells and preferably secreting Th1 cytokines. The UC peptides also showed immunogenicity and generated a specific antibody in mice by both intramuscular injection and oral delivery without adjuvant formulation. In conclusion, a T-cell vaccine could provide long-lasting protection against SARS-CoV-2 either during reinfection or during SARS-CoV-2 rebound. Due to its ability to eradicate SARS-CoV-2 virus-infected cells, a COVID-19 T-cell vaccine might provide a solution to lower COVID-19 severity and long COVID-19.
Similar content being viewed by others


Introduction
The rapid transmission of SARS-CoV-2 since the initial outbreak in December 2019 has caused more than 775.6 million infections and 7.1 million deaths as of July 20241. Its continued spread is due to its nature as an RNA virus that is prone to mutation. To date, the SARS-CoV-2 virus has evolved several variants of concern (VOCs) since the emergence of alpha, omicron and omicron subvariants such as BA2.75, BA.4 and BA.5. To date, many COVID-19 vaccines have been approved to vaccinate people; however, SARS-CoV-2 viruses are still evolving2. As it evolves, SARS-CoV-2 becomes more contagious and less lethal, but it can still cause vulnerable populations to develop multi-inflammatory syndrome (MIS)3,4.
To prevent SARS-CoV-2 infection, vaccination is an important strategy to generate neutralizing antibodies to stop SARS-CoV-2 transmission. At the beginning of the COVID-19 pandemic, the SARS-CoV-2 spike protein was regarded as the primary vaccine antigen target, especially due to the high antigenicity of the receptor-binding domain (RBD) of the spike protein. However, the spike protein is a rapidly mutating SARS-CoV-2 virus protein, whose mutation sites frequently occur on the RBD of SARS-CoV-25. Although the original spike antigen vaccine can successfully be effective against wild-type SARS-CoV-2, the subsequent evolution of SARS-CoV-2 variants, such as alpha, beta, gamma, delta, and omicron variants, has been reported to cause gradual loss of virus neutralization protection. The virus neutralization titres against the BA.4/BA.5 variants was dramatically reduced 21-fold compared to that of the wild type observed in people after booster vaccination6. Postvaccination infections (PVI) were frequently diagnosed after a new SARS-CoV-2 variant arose, which may be caused in part by the timing of vaccination, the vaccine-induced antibody level and waning immunity7. It raises the question of whether current vaccines are insufficient to protect against all SARS-CoV-2 variant infections and conveys an basic need to develop an effective vaccine to generate broadly neutralizing antibodies and long-term memory cellular immunity against SARS-CoV-2 variants and subvariants8,9. Despite the 60–80% vaccination coverage rate in developing and developed countries, the goal of herd immunity is still far from being achieved because of the instability in SARS-CoV-2 that accumulates massive mutations10. Furthermore, a new VOC of COVID-19 pandemic has appeared approximately every 6 months, as reported by WHO. Although there is a high demand for vaccine products against the new rise of VOCs of SARS-CoV-2, there is no effective vaccine product yet available. A new vaccine product requires the completion of vaccine design, proof of concept, validation of the pharmacology, manufacturing, safety evaluation and regulatory approval. Therefore, a universal vaccine conveying wide protection against all kinds of variants is a basic need for controlling the COVID-19 disease11. According to a WHO survey report, most mutations occur in the RBD region of the spike protein, which is a potential target antigen for developing COVID-19 vaccines. It is difficult to develop a vaccine specific for the mutation in the RBD region of the SARS-CoV-2 spike protein. The S2 region of the SARS-CoV-2 spike protein possesses fewer mutations than the RBD region, which is regarded as a “universal antigen”12,13. It has caused a change in focus to develop vaccines targeting the S2 region rather than against variable mutations in the RBD domain14.
A neutralizing antibody against SARS-CoV-2 can stop its entry into cells via the ACE2 receptor. However, if the SARS-CoV-2 virus breaks through the neutralizing antibody protection and infects cells, the SARS-CoV-2 virus is replicated in the cells and secretes numerous viral particles into the blood circulation. When immunity declines, the endogenous virus can infect other cells, and symptoms become obvious. Activated T-cells can eradicate virus-infected cells to stop the risk of endogenous infection15,16. T-cell vaccines have been widely developed as therapeutic strategies in oncology due to their high specificity to tumour cells. The HLA-I peptide derived from tumour-associated antigens activates CTL cells against tumour cells through the MHC/peptide-restricted pathway17. However, a subset of CTLs bearing γδTCR, so-called γδT-cells, can recognize target cells without MHC peptide restriction and are mostly involved in antiviral activity18. The activation of γδT-cells is regarded as a potential therapeutic method in antitumour and antiviral treatment19,20.
Compared to humoral immunity, cellular immunity contributes to long-lasting immunity, in which specific memory T-cells can be detected after 10 years, whereas specific antibodies decrease to low neutralizing levels within 2–3 months and undetectable levels within 2–3 years21,22. It might be proposed that cellular immunity is critical for preventing SARS-CoV-2 infection rather than humoral immunity, especially in postvaccine COVID-19 cases23. Many studies and surveys have been published to support that the cellular immunity-inducing vaccine against SARS-CoV-2 is more effective than that of humoral immunity at the stage of initial virus infection and long COVID24,25,26.
The similar concept of using the HLA-restricted peptide as vaccines for COVID-19 have been proposed and tested in clinical studies, including CoVac-1 from University Hospital Tübingen in Germany, CoVepiT from OSE Immunotherapeutics in France, and PepGNP-SARSCoV2 from Emergex Vaccines in the UK24. For CoVac-1, it was designed for HLA-restriction immune response, which HLA-DR+ T cells were directly loaded with six epitope peptides for consequent T-cell and B cell activation27. By means of loading of HLA-restricted peptides to antigen-presenting cells, the cellular and humoral immune responses were evoked, which was able to react to foreign antigen without antigen processing28.
In our study, we demonstrated a novel design of an HLA-restricted peptide that could activate CTLs to eradicate virus-infected mimic cells and activate immunity to produce antibodies against the SARS-CoV-2 virus spike protein without adjuvant formulation. It has also been shown to provoke mucosal immunity when it is delivered orally, and higher levels of IgA may be secreted to mucosal sites for protection from viral infection. The HLA-restricted peptide vaccine provides a new potential scheme for developing vaccines against viral infection.
Results
UC peptides designed from the well-conserved region of the SARS-CoV-2 spike protein for HLA restriction
The UC-100 and UC-152 epitopes were designed from the S2 domain of the SARS-CoV-2 virus spike protein, for which the well-conserved region was selected based on the side chain of a specific amino acid due to its steric protein structure (Fig. 1a). The order of the UC peptide sequence was directly assigned based on the exposed amino acids on the surface of the SARS-CoV-2 spike S2-well-conserved region in order to avoid virus mutations and glycosylation. The side chain of each amino acid was aligned with a structural linear sequence according to a distance of approximately 3.8 Å of Cα–Cα between 2 amino acids on the protein surface. Artificial UC-100 peptides comprising 9 amino acids were designed and expected to bind with the major histocompatibility complex (MHC)-class I molecule. The binding affinity to various MHC-I molecules was predicted, as shown in Table 1. The artificial UC-100 antigen was predicted to not only strongly bind to HLA-A201 but also bind to other major histocompatibility molecules. The artificial UC-100 epitope sequence aligned with the surface of the SARS-CoV-2 spike protein structure, which was discontinuous in the primary sequence and assembled into a linear arrangement on the surface (Fig. 1b). The separated sequence could not be processed as a known mechanism of antigen presentation by immune cells, but these separated short peptides were assembled as 9-mer amino acid immunogens, which might provoke cellular immunity by means of HLA-I molecule binding. Due to spike polypeptide refolding, the discontinued primary sequence of the spike protein formed a linear alignment on the surface. To obtain a higher binding activity of antigen peptides to MHC molecules, artificial UC-152 was modified with one amino acid substitution from the UC-100 peptide. The UC-152 peptide was predicted to have a higher binding affinity to MHC-I A201, as shown in Table 1. Both the UC-100 and UC-152 peptides were designed to boost cellular immunity.

The design of the HLA-restricted UC peptides of the SARS-CoV-2 spike protein. (a) The structure of the SARS-CoV-2 spike protein (PDB entry 6VSB) is highlighted in red with mutation hotspots of major pandemic VOCs (alpha, beta, gamma, delta, omicron BA.1/BA.2 and BA.4/BA.5). The predicted glycosylation sites of the SARS-CoV-2 spike protein are highlighted in blue. The UC peptides were designed from a well-conserved region of the SARS-CoV-2 spike protein (green shadow region). (b) The UC peptides were designed with a nonlinear sequence of the SARS-CoV-2 spike S2 domain primary structure, which folded to a linear arrangement on the surface of the tertiary structure of the SARS-CoV-2 spike protein.
UC peptides can be primed onto CTLs and engaged with the corresponding CTLs
To evaluate the binding affinity of UC-100 and UC-152 to MHC-I molecules, the HLA-A201 complex was analysed in an HLA refolding assay. The anti-β2m complex antibody can recognize the correctly folded structure when the matched peptide is loaded on the MHC-I molecule along with β2m. In the refolding assay, both UC-100 and UC-152 showed higher binding activity to HLA-A201 in a dose-dependent manner than a cytomegalovirus (CMV)-positive antigen (Fig. 2a). The results showed that both the 9-mer UC-100 and UC-152 peptides could refold with the HLA-A201 molecule and β2m. UC-100 and UC-152 derived from the assembly of the separated surface SARS-CoV-2 spike S2 region could be loaded on the HLA class I molecule and recognized by corresponding CTLs. To verify whether corresponding CTLs existed, the UC-100 and UC-152 antigens were subjected to a tetramer assay, and the corresponding CTLs were stained with the labelled tetramer complex. In the tetramer assay, HLA-A201 molecules refolded with UC-100 (4 μM) and UC-152 (4 μM) showed increases of 2.2% and 3.3% in positive CTLs compared with tetramers without loading peptide (Fig. 2b). These correctly refolded complexes would be engaged with the corresponding T-cell receptor in CD8+ CTLs in human peripheral blood mononuclear cells (PBMCs). These results showed that both UC-100 and UC-152 peptides could form a complex with HLA and trigger the activation of CTLs through the MHC/TCR complex.

The mechanistic immune responses of 9-mer UC peptides. (a) The UC-100 and UC-152 peptides were analysed in duplicate and represented as the MFI ± S.D. for peptide/HLA-A201/β2m complex formation at 1, 3 and 9 nM, respectively. CMVpp65 peptide was used as a positive control, and no peptide was used as a negative control. (b) The binding of human CD8+ cells (HLA-A201) was stained with the labelled tetramer complex (peptide/HLA-A201/β2m complex) in triplicate and represented as the MFI ± S.E. Tetramer binding of CTLs (CD8+ lymphocytes) was expressed as a percentage of gated with unstained CD8+ cells, and none of the peptides were added as a negative control. (c) Human T-helper cells bound to peptide/HLA-DR were assayed in triplicate with the indicated level of labelled dextramer, which is expressed as MFI ± S.E. (d,e) Human PBMCs were treated with various concentrations of UC-100 and UC-152 peptides (0.4, 2, 10, 50, 250 nM) in triplicate. The intracellular cytokines IL-4 and IFN-γ were double-stained and analysed by flow cytometry, and the levels of IL-4 and IFN-γ are expressed as the MFI ± S.E. The statistically significant differences are indicated by asterisks (unpaired one-tailed Student’s t test; *p < 0.05, **p < 0.01, ***p < 0.001).
UC peptides can be presented to MHC class II molecules and interact with T-helper cells
Although the HLA-DR molecule can be loaded with up to a 15-mer antigenic peptide in length, it can present a loaded antigen to the responsible T-helper cells. Thus, the 9-mer UC peptides were evaluated for the engagement of T-helper cells with the formation of the HLA-DR/peptide complex. To verify the roles of antigen peptides involved in the interaction between HLA-DR and TCRs, the peptides (200 μM) were subjected to a dextramer assay to understand the engagement of human T-helper cells. Human PBMCs were used for the dextramer assay, and the mean fluorescence intensity (MFI) of the labelled dextramer was analysed to determine the level of engaged CD4+ cells. In the dextramer assay, the results showed that UC-100 and UC-152 peptides could form a complex with HLA-DR and CD4+ T-helper cells that were increased 3.1-fold and 2.7-fold compared to the nonpeptide loaded group, respectively (Fig. 2c). The 9-mer UC-100 and UC-152 peptides formed HLA DR/peptide complexes and interacted with T-helper cells via HLA DR/TCR engagement. This finding suggests that 9-mer UC peptides might also be presented to immune cells via HLA DR/TCR molecules.
UC peptides polarized the Th1 cytokine release of human PBMCs
Immune cells, such as T-helper cells, can secrete cytokines to support an immune response towards a Th1 (IFN-γ-secreting) or Th2 (IL-4-secreting) phenotype. To understand the cytokine release pattern of the immune response in response to UC peptides, human PBMCs were treated with different concentrations of UC-100 and UC-152 peptides. The intracellular levels of the cytokines IFN-γ and IL-4 from T-helper cells were analysed to represent the Th1 and Th2 responses. The results showed that both UC-100 and UC-152 preferably stimulated IFN-γ cytokine secretion rather than IL-4 cytokine secretion (Fig. 2d,e). The level of IFN-γ was elevated in response to both UC-100 and UC-152 in a dose-dependent manner. This indicated that the 9-mer UC peptides preferably support the Th1 response.
UC peptides can directly activate the specific lysis activity of CTLs
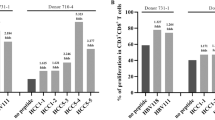
Based on previous results, UC peptides can form the MHC-I/peptide complex, and numerous CTLs were determined to be engaged. The complex might directly activate CTLs to attack antigen-bearing cells. To verify the effect of peptide-primed immune cells, the lytic activity of CTLs was assessed by targeting spike-expressing cells. In a CTL lysis assay, spike-expressing cells cocultured with different ratios of PBMCs were treated with 9 nM and 27 nM UC-100 peptides. The UC-100 peptide increased the lytic activity of spike-expressing cells in a dose-dependent manner (Fig. 3a). Similar results were observed in experiments with UC-152 peptide-treated PBMCs (Fig. 3b). These results support that spike-expressing cells can be recognized by and are cytotoxic to CTLs, and that CTLs were primed by the UC peptides. It was also demonstrated that UC peptides can activate immune cells to lyse virus-infected cells.

In vitro specific lysis activity of CTL by 9-mer UC peptides. Human PBMCs treated with 9 nM and 27 nM UC-100 (a) and UC-152 (b) peptides were cocultured with SARS-CoV-2 spike-expressing HEK 293 cells at ratios of 27:1, 9:1, 3:1 and 1:1 for 5 days. The lytic activity was measured by LDH release, total target cell death was set as 100% cytotoxicity, and no peptide treatment was used as the negative control. The assay was performed in duplicate and is represented as the mean ± S.E. Significant differences are indicated by asterisks (unpaired one-tailed Student’s t test; *p < 0.05, **p < 0.01, ***p < 0.001).
The nonadjuvanted UC peptides provoked immunogenicity in vivo
Mice were immunized by intramuscular injection with UC-100 and UC-152 without adjuvant formulation (Fig. 4a). The total levels of IgG, anti-peptide (UC-100 or UC-152)-specific IgG and anti-spike trimer were analysed to indicate the immunogenicity of UC peptides. The results showed that total mouse IgG could be increased from 186 to 339 µg/mL by UC-100 peptides and from 127 to 340 µg/mL by UC-152 peptides (Fig. 4b,e). There was a 1.8–2.7-fold increase in IgG production following UC peptide challenge. The immunised serum was also subjected to a specific binding assay using UC-100 and UC-152 peptide-coated microplates. The results showed that serum antibody was significantly increased by approximately 1.6-fold and 1.8-fold by binding to plates coated with UC-100 and UC-152 peptides, respectively (Fig. 4c,f). This result indicated that antibody production induced by UC-100 and UC-152 peptides can specifically recognize antigenic peptide sequences. The results also showed that both antibodies induced by UC-100 and UC-152 peptides could also bind to the SARS-CoV-2 spike trimer protein, similar to the peptide binding results (Fig. 4d,g). In the comparison of predose and postdose serum, an approximate 4.0–4.5-fold increase in specific anti-spike trimer IgG was induced. It was shown that 9-mer UC peptides not only showed immunogenicity in animals but also induced antibodies to recognize the matched synthetic peptide and structural sequence of the spike protein on the surface.

In vivo immune responses of injected UC peptides and complement activity of immunized serum. (a) BALB/c mice were intramuscularly administered 3 doses of 50 µg of UC-100 (n = 3) or 400 µg of UC-152 (n = 4) 14 days apart. (b–d) The serum of UC-100 mice was collected pre-(Day-1) and posttreatment (Day41) and tested in duplicate and represented as the mean for the levels of (b) total IgG; (c) specific IgG against UC-100 peptide; (d) specific IgG against SARS-CoV-2 spike trimer protein. (e–g) The serum of UC-152 mice was collected pre-(Day-1) and posttreatment (Day41) and tested in duplicate and represented as the mean for the levels of (e) total IgG; (f) specific IgG against UC-152 peptide; (g) specific IgG against SARS-CoV-2 spike trimer protein. (h,i) Antibody-mediated complement activity was tested in duplicate and is represented as the mean for pre- and postserum of mice vaccinated with UC-100 and UC-152 peptides, respectively. The complement activity was expressed as the IC50 of cytotoxicity of the (h) UC-100-vaccinated group and (i) UC-152-vaccinated group. Significant differences are indicated by asterisks (unpaired one-tailed Student’s t test; *p < 0.05, **p < 0.01, ***p < 0.001).
The UC peptide-induced antibody can activate complement to lyse epitope-bearing cells
To verify the complement effects of the vaccinated serum antibodies on virus-infected cells, the serum was tested in a complement-dependent cytotoxicity (CDC) assay using spike-expressing cells. In the CDC assay, spike-expressing HEK293 cells cultured in serum-free medium were treated with serially diluted serum supplemented with young rabbit complement. Complement cytotoxicity was measured by lactate dehydrogenase (LDH) activity. The results showed that complement-induced activity was increased approximately 2.1-fold in serum vaccinated with UC-152 peptide compared to the prevaccinated serum level, but not in UC-100 peptide-vaccinated mice (Fig. 4h,i). This result indicated that the UC peptide-induced antibody might target the intact spike protein of infected cells depending on the binding affinity of the induced antibody.
Oral administration of UC peptides provokes immunogenicity in mice
Due to the digestion resistance of artificial UC peptides, immunogenicity by oral administration was evaluated to understand whether the peptide could stimulate gut immunity. Five BALB/c mice were orally administered 200 μg of polymer-formulated UC-100. After 3 doses administered 2 weeks apart, predose and postdose serum were analysed for the levels of total IgG, specific anti-peptide IgG and anti-SARS-CoV-2 spike IgG (Fig. 5a). The results showed that total IgG was increased from 136 to 388 μg/mL by oral UC-100 peptides (Fig. 5b). Regarding the specificity of the induced antibody, an approximately 1.6-fold increase in anti-peptide IgG and a 4.4-fold increase in anti-spike IgG were measured (Fig. 5c,d). These results suggested that oral administration of UC-100 peptide could induce an immune response through gut immunity. To understand the CDC function of the induced antibody, serum was tested for complement activity, and the results showed that antibody-mediated complement activity was slightly increased by approximately 27% in vaccinated serum compared with the prevaccinated serum level (Fig. 5e). These results indicated that complement activity mediated by antibodies might be involved in the eradication of virus-infected cells. To further understand the possibility of mucosal immunity from an HLA-restricted peptide vaccine, sera were used to determine the levels of total IgA and specific IgA. The results showed that total IgA was significantly increased from 13.50 to 33.04 μg/mL by oral UC-100 peptides, which represents an approximately 2.4-fold increase in IgA (Fig. 5f). Regarding the specificity of the induced antibody, there was an approximately 1.8-fold increase in the specific anti-peptide IgA (Fig. 5g). These results suggested that oral administration of short UC peptides could induce mucosal immunity.

In vivo immune responses of oral UC peptides and complement activity of immunized serum. (a) BALB/c mice (n = 5) were orally administered 3 doses of 200 µg UC-100 14 days apart. (b–d) The serum of UC-100 mice was collected pre-(Day-1) and posttreatment (Day 41) and tested in duplicate and represented as the mean for the levels of (b) total IgG; (c) specific IgG against UC-100 peptide; (d) specific IgG against SARS-CoV-2 spike trimer protein; (e) The antibody-mediated complement activity was tested in duplicate and represented as the mean for pre- and postserum of mice vaccinated with UC-100. The complement activity was expressed as the IC50 of cytotoxicity. (f,g) The serum of UC-100 mice was collected pre-(Day-1) and posttreatment (Day 41) and tested in duplicate and represented as the mean for the levels of (f) total IgA and (g) specific IgA against UC-100 peptide. Significant differences are indicated by asterisks (unpaired one-tailed Student’s t test; *p < 0.05, **p < 0.01, ***p < 0.001).
Discussion
In this study, artificial 9-mer UC peptides were designed to enhance T-cell immunity. We successfully demonstrated the antigenicity of these UC peptides and the possible mechanism of the UC peptides in T-cell and B-cell activation. These data support the development of a 9-mer peptide as a T-cell vaccine accompanying a B-cell response (Fig. 6). This explains how the 9-mer short peptide activates CTLs through MHC-I recognition and activates B-cells to produce antibodies via MHC-DR engagement. Based on these observations, the 9-mer HLA-restricted peptide has the potential to be used to develop a next-generation vaccine against SARS-CoV-2. To date, the COVID-19 is still a global potential threat due to several variants’ waves causing PVI, including Omicron and its subvariants. These virus variants escape from human immunity and exist even in fully vaccinated or SARS-CoV-2 convalescent human serum29. There is still a basic need to develop an effective COVID-19 vaccine to stop this disease. In fact, PVI following new VOCs frequently occurs in nations or areas with high vaccination coverage rates30. Restored SARS-CoV-2 viruses have been observed in approximately 10% of convalescent patients31. It is implied that SARS-CoV-2 viruses are not eradicated and persistently exist in a latent phase. It can be deduced that there is a balance of viral load between the function of antibody-producing B-cells and virus-infected cells. When the balance collapses, the viruses rebound and become contagious. It is suggested that the activation of cellular immunity would be beneficial for complete recovery from the health conditions caused by long COVID-19 and in virus-restored patients32. However, scientists still struggle to improve neutralizing antibodies as a major function of vaccines to stop the new SARS-CoV-2 variant infection and virus recovery. Unfortunately, loss of neutralization activity and immune surveillance escape caused by virus mutations destroy the line of defence by neutralizing antibodies, which raises the need for long-term T-cell immunity33. According to a survey, current vaccines can provoke a high amount of neutralizing antibodies but do not stop PVI. Recently, several studies were conducted to develop broad-spectrum COVID-19 vaccines to address the issue of immune surveillance escape, including using multiple T-cell epitopes, multiple spike proteins loaded on virus-like particles, or using conserved regions outside the RBD for targeting epitopes or spike S2 domains as antigens34,35,36,37. Nearly 1 year after the emergence of the COVID-19 pandemic, scientists noticed that T-cell immunity played a critical protective role in SARS-CoV-2 convalescent patients and appealed to regulatory affairs that measurement of T-cell immunity should be included in COVID-19 vaccine clinical trials38. T-cell vaccines are supposed to be effective against newly evolved variants of SARS-CoV-2 and long COVID-19.

Summary of the SARS-CoV-2 HLA-restricted peptide T-cell vaccine. The possible mechanistic immune response to HLA-restricted peptides has been elucidated. The 9-mer UC peptides can be primed onto both HLA class I and II molecules, and the immune response is triggered for both humoral and cellular immunity. Through HLA class I molecule binding, CTLs can be activated, and an increase in IFN-γ cytokines and specific lytic activity have been observed. Through HLA class II molecule binding, B-cells are activated, and specific antibodies are produced in accordance with complement cytotoxicity activity.
Eradicating virus-infected cells is the key to success in stopping PVI. For this purpose, many scientists and institutions have altered vaccine development strategies to use short HLA-restricted peptides against SARS-CoV-2 virus infection and have claimed broad protection from SARS-CoV-2 variants and subvariants39. However, the response of CD8+ T-cells may be lost if the HLA-I-restricted antigen contains mutations40,41. It has been suggested that HLA-restricted peptides should avoid targets including areas of potential mutations and glycosylation obstructions. Otherwise, it faces the same problem of activity loss due to rapid evolution mutations.
Another achievable strategy for lowering PVI rates is to generate mucosal immunity42,43. To strengthen COVID-19 protection, IgA antibodies were generated through mucosal immunity. It could be provoked by vaccination via nasal spray or oral delivery. It is beneficial to generate IgA in the mucosa to defend against viral infection44. It has been reported that a high level of IgA can be induced through nasal and oral administration for mucosal IgA responses45. Interestingly, the only peptide inoculated without adjuvant formulation provoked a serum IgG as well as a secretory IgA response. It was deduced that short peptides provoke the immune response through Peyer’s patches in the intestine.
Despite the effective use of current COVID-19 vaccines, including mRNA- and virus-based formats, the stability of the vaccines is a critical issue for maintaining efficacy during transportation46. Because of their structural stability, short peptide vaccines are more stable and can include any formulation to fulfil the need for fast distribution. Moreover, alongside the study on the efficacy of UC peptides, mice were monitored for body weight and observed grossly. There was no apparent change in body weight and food consumption in the oral or intramuscular group of mice during the study period (see Supplementary Tables S1, S2 online). In a pilot toxicology rat model study, no changes were observed in the serum biochemistry and haematology after a five-fold dosage was administered intramuscularly. These observations suggest the high safety profile of UC peptides as a vaccine antigen. To achieve a good immune response, most effective vaccines are formulated with adjuvants. Adjuvants can amplify and modulate immune responses. The CpG adjuvant is widely used in modulating the immune response to enhance Th1 cellular immunity47. However, it often causes unexpected side effects, such as autoimmune disease in susceptible individuals48. In this study, we demonstrated the potential use of an HLA-restricted peptide COVID-19 vaccine without formulation with an adjuvant. It was shown that only short peptides are sufficient to trigger cellular and humoral immunity. It is also thought that intact immunity, including humoral and cellular immunity, can be induced to protect against virus infection and that unwanted side effects can be strongly reduced.
In conclusion, we designed an HLA-I-restricted peptide based on a well-conserved S2 domain of the SARS-CoV-2 spike protein to avoid the highly mutated RBD domain and potential glycosylation interference with antibody binding. It might provide broad-spectrum neutralizing antibody protection against RBD-mutated SARS-CoV-2 viruses. In addition, cellular immunity and CTLs are activated by HLA-I-restricted peptides to eliminate virus-infected cells and stop internal production of viruses. T-cell vaccines provide a solution to end the COVID-19 disease.
Methods
HLA-restricted peptides
The HLA-restricted peptides UC-100 (KLNSGDSKV) and UC-152 (KLNSGLSKV) were designed by Vacino Biotech Taiwan based on the S2 domain of the SARS-CoV-2 spike protein. The UC-100 and UC-152 peptides were synthesized by GenScript (NJ, USA) with 98% purity, and the pH value was adjusted to 7.2–7.4 for the in vitro and in vivo studies.
HLA-A201-peptide complex refolding assay
The easYmer kit (ImmunAware Aps, Copenhagen, Denmark) was used to determine the binding ability of the peptide to the HLA-A201 molecule. The detailed methods were performed according to the kit’s manual instructions. Briefly, the peptide was incubated with the β2m-microglobulin (β2m) light chain subunit and biotin-tagged recombinant HLA-A201 heavy chain at 18 °C for 48 h to form a 500 nM folded HLA complex. The folded complexes were diluted with dilution phosphate-buffered saline (PBS) buffer containing 5% glycerol to serial concentrations of 8.9 nM, 3.0 nM and 1.0 nM. Then, 20 µL of 45-fold diluted streptavidin-coated beads (6–8 µm beads, Cat SVP-60-5, Spherotech, IL, USA) was added to each well. The beads were suspended in the solution with PE-labelled anti-human β2m monoclonal antibody and incubated for 30 min at 4 °C. The fluorescence intensity was measured by flow cytometry (BD LSRFortessa™X20, Franklin Lakes, NJ, USA). An HLA-A201 binding peptide, NLVPMVATV (CMV pp65495–503), which was reported to have an IC50 of 45 nM for HLA-A201, was used as the positive control49. Complex formation without peptide addition was used as the negative control.
In vivo study of HLA-restricted peptides
The mouse immunogenicity studies were performed by Super Laboratory Co., Ltd. (Taipei, Taiwan), and the animals were supplied by BioLASCO Taiwan Co., Ltd. (Taipei, Taiwan). For the immunogenicity study of the UC-100 peptide antigen, three female BALB/c mice (7–9 weeks old) were injected intramuscularly with 3 doses of 50 µg peptide 14 days apart. For the immunogenicity study of UC-152 peptide antigen, four female BALB/c mice (7–9 weeks old) were injected intramuscularly with 3 doses of 400 µg peptide 14 days apart. For the oral ability study of the UC-100 peptide antigen, five female BALB/c mice (7–9 weeks old) were orally administered 3 doses of 200 µg of peptide formulated with 1% PLCL-PEG-PLCL polymer (Sigma‒Aldrich, USA). The serum was collected before the first dose of immunization (predose) and at 14 days after the last dose immunization (postdose). All animals were euthanized with the pre-anaesthetization of CO2 and the withdrawn of whole blood via cardiopuncture. The serum was stored at − 80 °C prior to antibody measurement and functional assay. The animal protocol was reviewed and approved by the Super Laboratory Company's Institutional Animal Care and Use Committee (approval IACUC number 111-14c). All animal experiments in this study were performed in accordance with the relevant guidelines and regulations. This study was reported in accordance with the ARRIVE guidelines.
Measurement of IgG and IgA levels
To determine the total IgG and IgA levels, the IgG (Total) Mouse Uncoated ELISA Kit (Cat., No. 88-50400, Thermo Fisher Scientific, MA, USA) and Mouse IgA ELISA Kit (Cat., E-90A, Immunology Consultants Laboratory, Inc., OR, USA) were used according to the manufacturers’ protocols. For total IgG measurement, a Nunc™ MaxiSorp™ 9018 ELISA plate was coated with 100 μL of the coating buffer containing capture antibody and incubated overnight at 4 °C. The plate was then washed twice with 400 μL of wash buffer. After the wells were blocked with 250 μL of blocking buffer (PBS, 1% bovine serum albumin (BSA), 0.1% Tween 20) and incubated at RT for 2 h, the plate was washed twice. Then, 100 μL of the diluted samples and standards were added to the reaction wells. Then, 50 μL of diluted detection antibody was added to all wells. The plate was covered and incubated at RT for 2 h. The samples were washed four times, 100 μL of substrate solution provided in the kit was added, and the samples were incubated at RT for 15 min. Finally, 100 μL of stop solution provided in the kit was added, and the plate was read at an optical density (OD) of 450 nm by a microplate reader (SpectraMax iD5, Molecular Devices, CA, USA). For total IgA measurement, 4000-fold diluted samples (100 μL) were added to an IgA capture antibody-precoated plate. The procedures followed the kit’s instructions. The signal was read at OD 450 nm by a microplate reader (SpectraMax iD5, Molecular Devices, CA, USA).
To measure the levels of anti-UC peptide-specific IgG, a 96-well plate was coated with 100 μL of PBS buffer containing 5 µg of UC-100 and UC-152 peptide and incubated overnight at 4 °C. The following steps for anti-peptide IgG measurement were performed as described for total IgG measurement. Briefly, after incubation with anti-mouse HRP antibody, the reaction microplate was washed four times, 100 μL of TMB Solution (ab171522, Abcam, Cambridge, UK) was added, and the plate was incubated at RT for 20 min. Finally, 100 μL of Stop Solution (Cat No. 5150-0020, SeraCare, MA, USA) was added, and the plate was read at OD 450 nm by a microplate reader (SpectraMax iD5, Molecular Devices, CA, USA). For specific IgA antibody measurement, UC-100 was coated onto a 96-well microplate. The experimental procedures were performed as previously described for IgG.
To measure the anti-spike trimer IgG antibody, a mouse SARS-CoV-2 spike trimer-coated 96-well plate kit (RAS-T023, Acro Biosystems, DE, USA) was used. The procedure was performed according to the kit’s instructions. 100 μL of diluted sample was added to an anti-spike trimer-coated plate, and 100 μL dilution buffer was used as a blank. The reaction was stopped by adding 50 μL of stop solution provided in the kit and read at OD 450 nm by a microplate reader (SpectraMax iD5, Molecular Devices, CA, USA).
Th1 and Th2 cytokine release assay
Briefly, 1 × 105 HLA-A201+ human PBMCs (STEMCELL Technologies Inc., Vancouver, British Columbia, Canada) were seeded in a 96-well microplate50. The PBMCs were treated with 5 different concentrations of antigen peptides at final concentrations of 0.4, 2, 10, 50 and 250 nM and cultured with X-VIVO 15 medium for 12 days. The culture medium was refreshed every 5 days. To analyse the surface marker and intracellular cytokine levels, the suspended cells were collected and stained with anti-CD4-PerCP-Cy5.5 conjugated (Cat. 566923, BD Pharmingen, USA), IFN-γ-FITC chrome conjugated (Cat. 554700, BD Pharmingen, USA), and IL4-PE chrome conjugated (Cat. 559333, BD Pharmingen, USA) antibodies. The cytokine levels in CD4+ T-helper cells was measured by flow cytometry (BD LSRFortessa™X20, Franklin Lakes, NJ, USA).
HLA-DR-peptide engagement assay
A U-Load Dextramer kit MHC II HLA-DRB1*0101/APC (Immudex, Denmark) was used to determine the binding ability of a peptide to the HLA-DR molecule. The method was performed according to the kit’s protocol. The 10 µL MHC-II-peptide monomer was prepared by mixing the reagents of a 2 µL of 1 mM peptide (UC-100 or UC-152), a 3 µL of dissolved U-load MHC-II peptide loading component and a 5 µL of U-load MHC-II (1 mg/mL). PBMCs (2 × 105) suspended in 50 µL of wash buffer composed of 1% FCS in phosphate-buffered saline (PBS), pH 7.4. The 10 µL of prepared MHC dextramer as described in the protocol was added to the PBMCs and incubated in the dark for 30 min at RT, followed by staining with the anti-CD4-PerCP-Cy5.5 conjugated antibody and further incubation in the dark for 20 min at RT. The cells were washed twice with 2 mL of wash buffer, and the cell pellet was resuspended in wash buffer for analysis by flow cytometry (BD LSRFortessa™X20, Franklin Lakes, NJ, USA).
Peptide-specific CTL engagement assay
The easYmer kit (ImmunAware Aps, Denmark) was used for a tetramer assay to determine the peptide-specific CTL binding level to the peptide-HLA-A201 complex51. The methods were performed according to the kit’s protocol. Briefly, the peptide was incubated with the β2-microglobulin (β2m) light chain subunit and biotin-tagged recombinant HLA-A201 heavy chain for 48 h at 18 °C to form a 50 µL 500 nM folded HLA complex. A tetramer was produced by incubating the equivalent of 2.1 µL of 0.2 mg/mL streptavidin-APC (Cat., 554067, BD Pharmingen, USA) and 50 µL of 500 nM folded HLA complex in the dark at 4 °C overnight. PBMCs (1 × 105, HLA-A201+) (STEMCELL Technologies Inc., Vancouver, British Columbia, Canada) were costained with the APC-labelled tetramer complex and FITC-conjugated anti-CD8 antibody (Cat., 555634, BD Pharmingen, USA) and analysed by flow cytometry (BD LSRFortessa™X20, Franklin Lakes, NJ, USA) for the percentage of tetramer-stained CD8+ cells.
Cytotoxic activity assay of specific T-cells
HEK293 cells expressing the SARS-CoV-2 spike protein (D614) (Cat No. 293-cov2-s, InvivoGen, CA, USA) were used as target cells, which were seeded into a 96-well plate52. HLA-A201+ PBMCs (STEMCELL Technologies Inc., Vancouver, British Columbia, Canada) were cocultured with 3000 SARS-CoV-2 spike-expressing cells at ratios of 27:1, 9:1, 3:1 and 1:1, and the culture was treated with UC-100 and UC-152 peptides at a final concentration of 9 nM or 27 nM for 5 days at 37 °C and 5% CO2. Cytotoxicity was measured by using an LDH-Cytox™ assay kit (Cat No. 426401, BioLegend, CA, USA). SARS-CoV-2 spike-expressing cells without PBMCs and peptide treatment were used as the negative control, while lysis buffer added to the well containing only spike-expressing cells was used as the positive control. Fifty microlitres of LDH substrate was added to each well and incubated for 30 min at RT. Finally, the OD at 490 nm was read by a microplate reader (EnVision Xcite Multilabel Reader, PerkinElmer, MA, USA). The percentage of specific cell lysis was calculated according to the manufacturer’s instructions.
Complement-dependent cytotoxicity assay
To determine the functions of the antibodies induced by the UC peptide, complement cytotoxicity was measured in SARS-CoV-2 spike protein (D614)-expressing HEK293 cells (Cat No. 293-cov2-s, InvivoGen CA, USA)53. The cytotoxicity mediated by the antibodies was measured by using an LDH-Cytox™ assay kit (BioLegend, CA, USA). The 4 × 103 SARS-CoV-2 spike-expressing cells were suspended in 45 µL of serum-free DMEM in each well of a 96-well microplate. The serum samples were serially diluted twofold for 9 dilutions with serum-free DMEM starting at a ratio of 1:20 to 1:5120. Five microlitres of diluted serum was added to the wells containing SARS-CoV-2-expressing cells and incubated for 15 min at 37 °C and 5% CO2. A final concentration of 10% young rabbit complement (CL3441-S50-R, Cedarlane, Ontario, Canada) was added to each well and incubated for an additional 30 min at 37 °C and 5% CO2. The plate was then cooled at RT for 15 min for measurement of the cytotoxicity by LDH release. The sample wells containing serum were used as background controls, while lysis buffer was added to the sample wells containing SARS-CoV-2-expressing cells, and complement was used as the positive control. Then, 50 µL of LDH substrate was added to each well and incubated for 30 min at RT. Finally, the OD at 490 nm was read by a microplate reader (EnVision Xcite Multilabel Reader, PerkinElmer, MA, USA). The percentage of specific cell lysis was calculated according to the manufacturer’s instructions.
Statistical analysis
Groups were compared using an unpaired one-tailed Student’s t test embedded in Prism 5 (GraphPad Software Inc., San Diego, CA, USA) for the measurement of IgG antibody, IgA antibody, tetramer and dextramer. A p value < 0.05 was considered significant (*p < 0.05, **p < 0.01, and ***p < 0.001).
Data availability
Data is provided within the supplementary information files.
References
WHO Coronavirus Disease (COVID-19) Dashboard. In: World Health Organization (2024). https://covid19.who.int. Accessed 02 July 2024.
Kyriakidis, N. C., López-Cortés, A., González, E. V., Grimaldos, A. B. & Prado, E. O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. npj Vaccines 6, 28 (2021).
Sansone, M., Studahl, M., Berg, S., Gisslén, M. & Sundell, N. Severe multisystem inflammatory syndrome (MIS-C/A) after confirmed SARS-CoV-2 infection: A report of four adult cases. Infect. Dis. 54, 378–383 (2022).
Stafie, C. S. et al. Pathogenic connections in post-COVID conditions: What do we know in the large unknown? A narrative review. Viruses 14, 1686 (2022).
Abavisani, M. et al. Mutations in SARS-CoV-2 structural proteins: A global analysis. Virol. J. 19, 220 (2022).
Hachmann, N. P. et al. Neutralization escape by SARS-CoV-2 Omicron subvariants BA.2.12.1, BA.4, and BA.5. N. Engl. J. Med. 387, 86–88 (2022).
Hu, J. et al. Reduced neutralization of SARS-CoV-2 B.1.617 variant by convalescent and vaccinated sera. Genes Dis. 9, 1290–1300 (2022).
Morens, D. M., Taubenberger, J. K. & Fauci, A. S. Universal coronavirus vaccines—An urgent need. N. Engl. J. Med. 386, 297–299 (2022).
Wherry, E. J. & Barouch, D. H. T cell immunity to COVID-19 vaccines. Science 377, 821–822 (2022).
Kumar, S., Karuppanan, K. & Subramaniam, G. Omicron (BA.1) and sub-variants (BA.1.1, BA.2, and BA.3) of SARS-CoV-2 spike infectivity and pathogenicity: A comparative sequence and structural-based computational assessment. J. Med. Virol. 94, 4780–4791 (2022).
Vashishtha, V. et al. Looking to the future: Is a universal coronavirus vaccine feasible?. Expert Rev. Vaccines 21, 277–280 (2022).
Guo, L. et al. Targetable elements in SARS-CoV-2 S2 subunit for the design of pan-coronavirus fusion inhibitors and vaccines. Signal Transduct. Target. Ther. 8, 197 (2023).
Harvey, W. T. et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 7, 409–424 (2021).
Shah, P., Canziani, G. A., Carter, E. P. & Chaiken, I. The case for S2: The potential benefits of the S2 subunit of the SARS-CoV-2 spike protein as an immunogen in fighting the COVID-19 pandemic. Front. Immunol. 12, 637651 (2021).
Thomas, P. G., Keating, R., Hulse-Post, D. J. & Doherty, P. C. Cell-mediated protection in influenza infection. Emerg. Infect. Dis. 12, 48–54 (2006).
Toor, S. M., Saleh, R., Nair, V. S., Taha, R. Z. & Elkord, E. T-cell responses and therapies against SARS-CoV-2 infection. Immunology 162, 30–43 (2021).
Want, M. Y., Bashir, Z. & Najar, R. A. T cell based immunotherapy for cancer: Approaches and strategies. Vaccines (Basel) 11, 835 (2023).
Shin, S. et al. Antigen recognition determinants of gammadelta T cell receptors. Science 308, 252–255 (2005).
Vyborova, A. et al. γ9δ2T cell diversity and the receptor interface with tumor cells. J. Clin. Investig. 130, 4637–4651 (2020).
von Massow, G., Oh, S., Lam, A. & Gustafsson, K. Gamma delta T-cells and their involvement in COVID-19 virus infections. Front. Immunol. 12, 741218 (2021).
Le Bert, N. et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 584, 457–464 (2020).
Ng, O.-W. et al. Memory T cell responses targeting the SARS coronavirus persist up to 11 years post-infection. Vaccine 34, 2008–2014 (2016).
Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 23, 186–193 (2022).
Dolgin, E. T-cell vaccines could top up immunity to COVID, as variants loom large. Nat. Biotechnol. 40, 3–4 (2022).
Zhang, Q. et al. Identification of SARS-CoV-2-specific T cell and its receptor. J. Hematol. Oncol. 17, 15 (2024).
Tarke, A. et al. SARS-CoV-2 vaccination induces immunological T cell memory able to cross-recognize variants from Alpha to Omicron. Cell 185, 847–859 (2022).
Heitmann, J. S. et al. A COVID-19 peptide vaccine for the induction of SARS-CoV-2 T cell immunity. Nature 601, 617–622 (2022).
Somogyi, E. et al. A peptide vaccine candidate tailored to individuals’ genetics mimics the multi-targeted T cell immunity of COVID-19 convalescent subjects. Front. Genet. 12, 684152 (2021).
Planas, D. et al. Considerable escape of SARS-CoV-2 Omicron to antibody neutralization. Nature 602, 671–675 (2022).
Lipsitch, M., Krammer, F., Regev-Yochay, G., Lustig, Y. & Balicer, R. D. SARS-CoV-2 breakthrough infections in vaccinated individuals: Measurement, causes and impact. Nat. Rev. Immunol. 22, 57–65 (2022).
Callaway, E. COVID rebound is surprisingly common—Even without Paxlovid. Nature https://doi.org/10.1038/d41586-022-02121-z (2022).
Wiech, M. et al. Remodeling of T cell dynamics during long COVID is dependent on severity of SARS-CoV-2 infection. Front. Immunol. 13, 886431 (2022).
Riou, C. et al. Escape for recognition of SARS-CoV-2 variant spike epitopes but overall preservation of T cell immunity. Sci. Transl. Med. 14, eabj6824 (2022).
Rouzbahani, A. K., Kheirandish, F. & Hosseini, S. Z. Design of a multi-epitope-based peptide vaccine against the S and N proteins of SARS-COV-2 using immunoinformatics approach. Egypt. J. Med. Hum. Genet. 23, 16 (2022).
Dolgin, E. Pan-coronavirus vaccine pipeline takes form. Nat. Rev. Drug Discov. 21, 324–326 (2022).
Fang, Y. et al. An antibody that neutralizes SARS-CoV-1 and SARS-CoV-2 by binding to a conserved spike epitope outside the receptor binding motif. Sci. Immunol. https://doi.org/10.1126/sciimmunol.abp9962 (2022).
Ng, K. W. et al. SARS-CoV-2 S2-targeted vaccination elicits broadly neutralizing antibodies. Sci. Transl. Med. 14, eabn3715 (2022).
Wherry, E. J. et al. Letter to FDA—T-cell assessment in vaccine studies_4–21–22_FINAL.pdf. https://drive.google.com/file/d/1OPfStqOnuKAEUkrjfFUouXMjDB_-tnmV/edit (2022).
Tarke, A. et al. Comprehensive analysis of T cell immunodominance and immunoprevalence of SARS-CoV-2 epitopes in COVID-19 cases. Cell Rep. Med. 2, 100204 (2021).
Agerer, B. et al. SARS-CoV-2 mutations in MHC-I-restricted epitopes evade CD8 + T cell responses. Sci. Immunol. 6, eabg6461 (2021).
Hamelin, D. J. et al. The mutational landscape of SARS-CoV-2 variants diversifies T cell targets in an HLA-supertype dependent manner. Cell Syst. 13, 143–157 (2022).
Lavelle, E. C. & Ward, R. W. Mucosal vaccines—Fortifying the frontiers. Nat. Rev. Immunol. 22, 236–250 (2022).
Russell, M. W., Moldoveanu, Z., Ogra, P. L. & Mestecky, J. Mucosal immunity in COVID-19: A neglected but critical aspect of SARS-CoV-2 infection. Front. Immunol. 11, 611337 (2020).
Sheikh-Mohamed, S. et al. Systemic and mucosal IgA responses are variably induced in response to SARS-CoV-2 mRNA vaccination and are associated with protection against subsequent infection. Mucosal Immunol. 15, 799–808 (2022).
Boyaka, P. N. Inducing mucosal IgA: A challenge for vaccine adjuvants and delivery systems. J. Immunol. 199, 9–16 (2017).
Uddin, M. N. & Roni, M. A. Challenges of storage and stability of mRNA-based COVID-19 vaccines. Vaccines (Basel) 9, 1033 (2021).
Millan, C. L. B., Weeratna, R., Krieg, A. M., Siegrist, C.-A. & Davis, H. L. CpG DNA can induce strong Th1 humoral and cell-mediated immune responses against hepatitis B surface antigen in young mice. Proc. Natl. Acad. Sci. USA 95, 15553–15558 (1998).
Segal, B. M., Chang, J. T. & Shevach, E. M. CpG oligonucleotides are potent adjuvants for the activation of autoreactive encephalitogenic T-cells in vivo. J. Immunol. 164, 5683–5688 (2000).
Hassan, C. et al. Naturally processed non-canonical HLA-A*02:01 presented peptides. J. Biol. Chem. 290, 2593–2603 (2015).
Tapia-Calle, G. et al. A PBMC-based system to assess human T cell response to influenza vaccine candidates in vitro. Vaccines (Basel) 7, 181 (2019).
Bonté, P.-E. et al. Single-cell RNA-seq-based proteogenomics identifies glioblastoma-specific transposable elements encoding HLA-I-presented peptides. Cell Rep. 39, 110916 (2022).
Babaer, D., Zheng, M., Ivy, M. T., Zent, R. & Tiriveedhi, V. Methylselenol producing selenocompounds enhance the efficiency of mammaglobin-A peptide vaccination against breast cancer cells. Oncol. Lett. 18, 6891–6898 (2019).
Osada, T. et al. Polyfunctional anti-human epidermal growth factor receptor 3 (anti-HER3) antibodies induced by HER3 vaccines have multiple mechanisms of antitumor activity against therapy resistant and triple negative breast cancers. Breast Cancer Res. 20, 90 (2018).
Acknowledgements
We thank Super Laboratory Co., Ltd. (Taiwan) for animal experimentation and serum collection in this study.
Author information
Authors and Affiliations
Contributions
J.M.C. and Y.R.L. designed the HLA-restricted peptide and supervised this study. C. W.L. performed the mechanistic immune response of the in vitro assays and complement activity of the induced ex vivo assay. I.H.L. performed the measurement of the level of total antibody, specific antibody against peptide and spike trimer protein. All authors participated in analysing the data, J.M.C. and Y.R.L. wrote the manuscript, and all authors helped revise the manuscript.
Corresponding author
Ethics declarations
Competing interests
This research was financially supported by Vacino Biotech Co., Ltd. (Taiwan). Vacino Biotech has patents (US 17/726,688, PCT/CN2022/089266, ROC 111115497) concerning the therapeutic compounds used in this study, for which Yi-Ru Lee and Jia-Ming Chang are also the authors of the patents. Yi-Ru Lee, Chiung-Wen Liou, I-Hua Liu, and Jia-Ming Chang are employees of Vacino Biotech Company and belong to this company.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lee, YR., Liou, CW., Liu, IH. et al. A nonadjuvanted HLA-restricted peptide vaccine induced both T and B cell immunity against SARS-CoV-2 spike protein. Sci Rep 14, 20579 (2024). https://doi.org/10.1038/s41598-024-71663-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-71663-1
