
Abstract
Human-wildlife conflict (HWC) involving megafauna is a global conservation concern. In the tropical lowlands of Nepal (Terai), the megafauna populations are increasing due to successful conservation efforts, introducing the potential for greater HWC. We analyzed the spatio-temporal trend of reported HWC incidents from 2013 to 2022 in the buffer zones of Bardia and Chitwan National Parks in the Terai. Of the total 14,989 reported HWC incidents, crop raiding (n = 8,129) and livestock depredation (n = 4,611) were the most common. Elephants were responsible for 42.8% of all incidents. Total HWC incidents increased over the 10-year period, and there was a significant association between total population estimates of rhino and tiger and total HWC incidents involving those species. At the site level, this association held except for rhino HWC in Bardia. Similarly, incidents of livestock depredation driven by leopard and tiger, and crop damage incidents from elephant, rhino, and boar, increased over the study period. In contrast, property damage—exclusively by elephants—remained relatively stable in Chitwan but decreased in Bardia, possibly reflecting seasonal and transboundary movement patterns of elephants, or the effectiveness of targeted mitigation measures that have reduced elephant intrusions into settlements. Conflict hotspots—defined as the wards in the top quantile of HWC incidents—indicated priority areas for focused mitigation and remedial efforts. Habitat restoration in the buffer zones and corridors outside PAs could potentially mitigate HWC, but it might simultaneously export HWC to new areas outside the PAs. Targeted community-based mitigation measures should be implemented in the peak raiding periods, and predator-proof sheds would reduce livestock losses; both initiatives require substantial external financial support. Finally, the widespread adoption of livestock insurance that responds to spatiotemporal risk should be considered, along with funding and coordination by government and conservation organizations. Overall, the evidence suggests that successful megafauna conservation may be associated with increased HWC. Nevertheless, our findings offer practical insights to guide species-specific conflict mitigation and HWC management in tropical landscapes where megafauna populations are recovering, and conflicts are intensifying.
Similar content being viewed by others

Introduction
Human-wildlife conflict (HWC) is a globally prevalent challenge for conservation authorities managing protected areas (PAs) and for people living in the surrounding settlements1,2,3,4. HWC is especially severe in rural communities of low-income countries, where the costs in terms of both finance and food security are disproportionately high5,6. In the buffer zones or areas surrounding PAs - particularly in low-income countries like Nepal - the burden of conservation success can be substantial7,8,9,10. This underscores the importance of developing policies that sustain conservation success while addressing the economic challenges faced by rural communities11,12.
Populations of large mammals have been increasing in some countries13,14and HWC involving megafauna species has similarly been increasing (e.g4,15,16,17,18. In Nepal, early research in two important PAs of the tropical lowlands (Terai) indicated that there has been an increase in the number of reported HWC incidents in the buffer zones7,19. While studies have provided compelling qualitative data to suggest that the rise in conflict incidents may be linked to growing wildlife populations (e.g20, to our knowledge research on quantifying this association is very limited. Understanding this relationship is important because management efforts focus on large flagship species as key contributors to tourism revenue in several countries (e.g21,22, and yet increasing HWC incidents may manifest in negative attitudes among local communities toward conservation, potentially resulting in retaliatory killings23,24,25,26.
Beyond critical population-conflict associations, a greater understanding of species- and site-specific conflict dynamics is essential in assessing potential HWC mitigation responses. For example, the type of damage incurred by wildlife species varies across both ecological and geographic contexts. Similarly, activity patterns may differ across species, leading to different hotspots of HWC (e.g1,4,7,19,27. A clearer understanding of the spatiotemporal dynamics of HWC is crucial to inform policymakers and managers prioritizing conservation activities to not only restore megafauna populations, but also to develop mitigation strategies and reduce HWC incidents, thereby contributing to human-wildlife coexistence and long-term conservation success.
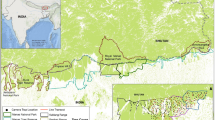
Among the seven PAs in Nepal’s Terai, Bardia National Park (BNP) and Chitwan National Park (CNP) are the largest, comprising more than 50% of the protected landscapes of the Terai and protecting significant populations of megafauna (See Fig. 1)28. Both PAs lie within the Terai Arc Landscape (TAL), a priority landscape for tiger conservation29,30. These PAs are the biodiversity hotspots that harbor some of the most iconic species of Asia31. The core area of the PAs is strictly protected by the national army, whereas the multiple-use buffer zone forests are restored and managed by the buffer zone communities32. In addition to the strict protection of protected areas by the national army, community volunteers have made significant contributions to wildlife protection, management, and conflict mitigation efforts33. These strategies have facilitated the recovery of globally recognized megafauna species such as tiger (Panthera tigris tigris), leopard (P. pardus), rhino (Rhinoceros unicornis), and elephant (Elephas maximus)28,29,34,35. Preliminary evidence suggests that HWC may be increasing7 possibly in association with the successful conservation outcomes in those PAs.

Location map of Bardia and Chitwan National Parks and their buffer zones in Nepal. The land cover map highlights the Terai region of Nepal, showing its connection to other protected areas within Nepal and India. Map generated using QGIS version 3.28 (https://qgis.org); Land cover data87; protected areas in Nepal88; Indian PAs: Protected Planet [https://www.protectedplanet.net/country/IND]; administrative boundaries: GADM [https://gadm.org].
In this research, we utilized a decade-long dataset (2013–2022) to test the following hypotheses about HWC in BNP and CNP: (1) HWC has increased over the past decade; (2) the temporal trends in HWC incidents are associated with the population trends of megafauna; (3) the incident rates for crop species and livestock prey vary across megafauna species; (4) there is a detectable spatiotemporally-explicit pattern in HWC incidents (“hotspots”) in the PA buffer zones.
Results
Temporal trends in wildlife populations and HWC incidents
Combined across both PAs, a total of 14,989 HWC incidents was reported from 2013 to 2022 (Table 1), increasing from 1,061 in 2013 to 1,939 in 2022 (Fig. 2). Summed across both PAs, both tiger and rhino populations increased between 2005 and 2022, with tigers more than doubling from an estimated 100 to ~ 250 (r = 0.96, P < 0.01), and rhino populations increasing from 400 to more than 700 (r = 0.99, P < 0.001) (Fig. 3). Concomitantly, the number of reported incidents involving tigers increased from < 100 in 2013 to more than 200 in 2022 (Pearson r = 0.78, P < 0.01) and incidents involving rhino increased from < 100 to ~ 400 (Pearson r = 0.75, P < 0.05). Leopard incidents also increased from < 300 to more than 600 (r = 0.80, P < 0.01) (Fig. 3). Elephant incidents, on the contrary, showed a declining trend over the 10 years, although there was no statistically significant trend owing to high interannual variation (Fig. 3).

Total HWC incidents reported in Bardia and Chitwan National Parks of Nepal over 10 years (2013–2022). The dotted line is the linear trend.

Trends in rhino and tiger populations combined across both sites, and separately for Bardia and Chitwan (left), alongside reported incident counts caused by four megafauna species over the decade 2013–2022, combined and separately for Bardia and Chitwan (right). The significance level for correlation between population or incidents and year is denoted by ‘NS’, ‘*’, ‘**’, and ‘***’ which represent non-significant, 90%, 95%, and 99% significance levels respectively.
When analyzed by site, in Bardia tiger populations increased steadily from approximately 60 to over 120 individuals between 2005 and 2022 (r = 0.75, P < 0.01), while rhino populations remained relatively stable over the same period (Fig. 3). Also, reported incidents involving tigers and leopards exhibited significant increasing trends (r = 0.70–0.75, P < 0.05), whereas rhino and elephant-related incidents showed a declining trend. In Chitwan, both tiger and rhino populations showed strong positive increases, with tiger numbers rising from ~ 30 to over 100 (r = 0.82, P < 0.01), and rhino from ~ 400 to nearly 700 (r = 0.99, P < 0.001). In addition, incidents involving rhinos and leopards increased significantly (r = 0.83, P < 0.001 and r = 0.79, P < 0.001 respectively), while tiger-related incidents showed non-significant increase, and elephant incidents remained stable with no discernible trend (Fig. 3).
Temporal trends of HWC incidents differed across incident types and between Bardia and Chitwan Parks (Fig. S1). Average annual crop raiding rates were similar between the two sites (424.9/year in Bardia and 388/year in Chitwan). They increased significantly over time in Chitwan but showed no temporal trend in Bardia. At both sites, livestock depredation increased significantly over the 10 years, although the average annual rates were several times greater in Bardia as compared to Chitwan (366/year versus 95.1/year). The average rate of human attacks differed between Bardia and Chitwan, and neither PA showed any temporal trend in this HWC incident type. Property damage showed a significant decline in Bardia, whereas Chitwan had very low annual rates of property damage and no trend.
Species-specific incident dynamics
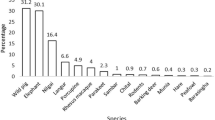
Seven wildlife species were involved in four incident types (Fig. 4). The most common incident type was crop raiding (n = 8,129), especially of paddy rice (48.3%), wheat (24.9%) and maize (18.6%) (Table 1). Elephants were responsible for 56.1% of crop-raiding incidents, with the remainder of raids evenly split between rhino and wild boar (Fig. 4). The crop raiding frequency was variable across these three species: while elephants mostly raided paddy, which accounted for 68% of their raids (Fig. 5a), rhino mostly raided wheat followed by paddy, and wild boar mostly raided maize, followed by wheat and paddy (Fig. 5a).

Proportion of contribution of wildlife species to conflict incidents in Bardia and Chitwan National Parks.

(a) Incident rates involving crop types by elephant, rhino, and wild boar. (b) Incident rates involving livestock types by leopard and tiger.
There were 4,611 incidents of livestock depredation, dominated by goat predation (60.9%) followed by pig (16.7%) and cattle (15.2%) predation (Table 1). Livestock depredation was largely attributable to leopards (> 75%) (Fig. 4), which strongly preferred goats, with the remainder of wildlife depredation due to tigers, which preferred cattle over goats (Fig. 5b). Buffalo, which are very large prey, accounted for less than 15% of all predation cases, and very small prey such as rabbits and fowl, which are kept in enclosures, were rarely preyed upon (Table 1).
Among the four reported incident types, human attacks were the least frequent (n = 506), with fatalities occurring in 26% of incidents (Table S1). Seven wildlife species attacked humans, with rhino accounting for 31.8% of all cases, followed by elephant (21.5%) and tiger (17.3%) (Fig. 4). Mortality from wildlife attack varied greatly depending on the wildlife species encountered: 45% percent of encounters with tigers and elephants and 19.9% of encounters with rhino resulted in a human fatality (Table S1).
Elephants were involved in all 1,743 property damage incidents, and they were also the only species involved in three types of incidents (Table 2). Of the property damage incidents, 96.8% involved damage to housing, likely because the animals were seeking stored food (e.g. harvested rice) within the structure (Table 1).
Conflict hotspots and spatial heterogeneity of HWC incidents
The cumulative incident density in Bardia (range: 30.5–163 km− 2; mean ± SE = 79.97 ± 15.99 km− 2) was notably higher than in Chitwan (range: 7.6–88.2 km−2; mean ± SE = 20.31 ± 3.96 km− 2) (Table S2). Spatially, the hotspot wards of incident densities accounted for 14% of buffer zone area (8 wards) in Bardia and 25% of the buffer zone area (20 wards) in Chitwan (Fig. 6a). The hotspot wards in Bardia were clustered in the southwestern sector of the buffer zone, and in Chitwan there was a cluster along the southern border along with scattered wards along the northern buffer (Fig. 6a). When stratified by megafauna species, some regions or wards showed particularly high density of conflict: tigers and leopards had high incident densities in the NW buffer zone of Bardia and scattered wards of high incident rates for leopards around Chitwan (Fig. 6b). On the contrary, elephants and rhinos showed strong clustering of hotspots in SW Bardia and in the southern buffer of Chitwan (Fig. 6b).

(a) Spatial distribution of cumulative incident density across wards in the buffer zones of Bardia and Chitwan National Parks, caused by four megafauna species (tiger, leopard, rhino, and elephant). (b) Species-specific incident density (number of incidents per square kilometer) by ward. Each polygon within the buffer zone represents a ward—the smallest administrative unit in Nepal. Dark red polygons indicate conflict hotspots, green polygons indicate no reported incidents (“0”), and brown represents the core protected areas. Breaks were defined using the “classInt” package in R with the “quantile” method86. Map generated using QGIS version 3.28 (https://qgis.org); Protected area boundaries88; administrative boundaries of Nepal: Open Data Nepal [https://opendatanepal.com/dataset/nepal-municipalities-wise-geographic-data-shp-geojson-topojson-kml]. This map figure was made by BN (first author).
Discussion and recommendations
In this study, we found a significant positive correlation between rhino and tiger population estimates and the number of incidents reporting their involvement. Overall, conservation intervention in both Bardia and Chitwan has increased tiger population estimates from approximately 100 to 250, and for rhino from 400 to over 700 between 2005 and 202234,36. Correspondingly, the number of incidents involving tigers increased from 71 to 219, and those involving rhinos increased from 86 to 395 between 2013 and 2022. Site-specific results are also consistent with population-incident trend associations except for rhino in Bardia. Thus, while conservation policies, and government and local community efforts have led to the recovery of globally important megafauna in Nepal’s PAs, this case highlights the significant long-term challenges of managing such conservation gains, viz. rising wildlife populations and the associated increase in HWC. The challenge extends beyond tiger and rhino, however: though Nepal lacks official census data for leopards (a co-habiting species with tiger), the sharp rise in livestock depredation (less than 300 to over 600 in the last decade 2013–2022) suggests that their numbers have also grown. As megafauna populations grow, they may reach or exceed the carrying capacity of a PA, especially when crops and livestock supplement their diets. This increases competition for limited habitat and resources, pushing megafauna into settlement areas and potentially leading to more HWC incidents37,38. Consequently, higher losses (e.g. of high-value livestock) can foster negative attitudes towards wildlife conservation and raise concerns about the sustainability of conservation efforts39.
In contrast to rising conflict trends involving tiger, rhino and leopard, we observed a declining trend in elephant-related conflict incidents in our study. This decline likely reflects the seasonal and transboundary movement patterns of elephants40,41which migrate between Nepal and adjoining Indian protected areas, including Katarniaghat Wildlife Sanctuary and Valmiki Tiger Reserve (Fig. 1). Their sporadic presence in the study area may lead to fewer conflict reports in certain years. Additionally, field observations and discussions with local people in Bardia indicated that reinforced concrete fences prevented elephants from entering settlements along with community-based initiatives, including the mobilization of rapid response teams and awareness programs like “Hatti Mero Sathi”. These initiatives may have contributed to the decline in property damage incidents in Bardia, in contrast to Chitwan, where such incidents did not decline. However, to better understand the elephant movements and associated incidents, we recommend real-time migration tracking along transboundary routes. In addition, further studies are required to evaluate the effectiveness of concrete fences and community-based initiatives in the study area.
When comparing the two sites, Chitwan had a higher number of human attacks, while Bardia had more crop raiding and livestock depredation cases (Fig. S1). This may be due to differences in wildlife population densities. Chitwan holds over 90% of Nepal’s rhino population, and rhino were responsible for the highest proportion of human attacks combined across both PAs (Fig. 4), likely reflecting higher human attacks due to their high local abundance in Chitwan. In contrast, there was higher livestock depredation and crop damage in Bardia, possibly due to greater densities of leopards and elephants. Leopards, which caused the majority of livestock losses (Fig. 4), were reported to depredate livestock about seven times more frequently than tigers in Bardia42. Additionally, tiger density in Bardia (7.15/100 km²) exceeds that of Chitwan (4.06/100 km²)43possibly indicating higher densities of coexisting leopards because in prey-rich areas like Bardia, leopards can coexist with tigers through spatial or dietary partitioning44. Similarly, Bardia’s estimated resident elephant population—reported to be more than double that of Chitwan—may explain the higher frequency of elephant-related crop damage observed in Bardia45. Despite these differences, HWC was found to be increasing on both sites.
To address the conservation success challenges, key conservation efforts should be focused on habitat expansion outside PA boundaries through habitat restoration, particularly in buffer zones and biological corridors46. Past studies demonstrate that prey densities are lower in corridor forests outside protected areas compared to core zones47. In addition, previous studies from both Nepal and other regions have documented that conservation efforts such as maintaining habitat connectivity through forest corridors and protecting remnant forest patches have facilitated habitat use by megafauna not only within protected areas but also in human-modified landscapes outside these boundaries48,49. Restoring habitat outside protected areas can increase forage availability during resource-scarce periods, which may reduce megafauna reliance on agricultural lands and decrease crop raiding incidents. Additionally, enhancing prey density outside PAs may help reduce large carnivore (e.g. tiger and leopard) movements into human-dominated landscapes50. In Nepal’s Terai, WWF has initiated habitat restoration activities in buffer zone forests51 and expanding those efforts will further benefit megafauna with large home ranges. Afforestation or reforestation of corridors can effectively connect with protected areas, facilitating the movement of megafauna between PAs, including across borders, which will also contribute to increasing carrying capacity of a particular PA52. For example, in Nepal, protecting and effectively managing the Khata Corridor and forests surrounding Bardia could enhance the frequency of megafauna movement between Bardia and India’s Katerniaghat Wildlife Sanctuary. Similarly, safeguarding the Shikaribas Corridor would enhance connectivity between Chitwan and India’s Balmiki Tiger Reserve. This connectivity is also essential for maintaining genetic diversity and ensuring the long-term sustainability of species52. It is recognized, however, that by increasing habitat area and connectivity outside of PAs, there is the likelihood that HWC will be exported further afield from the PAs themselves, as wildlife populations expand into those new habitats and corridors2. Thus, additional interventions need to safeguard lives and property of rural residents as wildlife populations continue to expand.
Beyond habitat expansion through restorations, species-specific conflict rates by crop and livestock type should be considered to effectively mitigate losses to crops and livestock. For example, understanding the feeding frequencies of wildlife species is crucial for designing mitigation measures. We found that certain herbivorous megafauna species raid particular crop more frequently than others: while elephants raid paddy rice more often than other crops, rhinos raid wheat, and wild boar raid maize more often than other crops. Further, our results revealed the intra-annual or monthly patterns of crop raiding for these three major crops, highlighting their distinct peak raiding periods (Figure S2). Previous studies have also reported that elephants tend to raid paddy fields during the peak harvesting period (Sept-Nov) in the Terai, when the paddy is most nutritious and palatable8; see also53,54. Rhinos, on the other hand, target wheat during young and intermediate growth stages55,56which in the Terai is Dec-Feb, when less forage is available in the natural habitats. Wild boar was found to raid maize during the maturing stage when it is ripe and easily digestible57. In our study, we did not use camera traps and telemetry to assess the species-specific habitat uses or ranging patterns of megafauna involved in conflicts. Thus, understanding wildlife behavior—such as why certain crop types are preferred over others—remains a critical research need. However, the temporal raiding patterns shown by our study can be addressed efficiently through temporal deterrent strategies. For example, community-based guarding activities involving watchtowers (Machans) during peak seasons of crop raiding can be an effective and low-cost solution for protecting crops from wildlife58, especially when local governments and organizations employ local people for such guarding activities. From our field observations and evaluations, this mitigation approach would be more affordable than fencing and could both enhance crop security and provide livelihood support for low-income and unemployed individuals in buffer zone communities.
Livestock depredation is a function of both the ecological niche of the predator, as well as the availability of preferred prey species. Leopards depredate mostly goats (see59,60,61,62 and tiger attack larger-sized prey species (cattle) (see63,64,65. In the buffer zones of Bardia and Chitwan, most livestock depredations occurred in livestock sheds rather than in grazing lands and forests7,66which suggests that the type of shed structures can mitigate predation risk. Cattle are typically kept in open or unprotected sheds with minimal or no walls, making them more vulnerable to tiger attacks. In contrast, goats and sheep are usually kept in closed sheds, constructed from traditional materials such as brittle wood and bamboo, or more sturdy materials like hardwood and gabion wire, which are considered “predator-proof” (see Fig. S3). An obvious remedial measure would be to build protective structures (predator-proof sheds) for the livestock. However, while buffer zone programs receive government support (30–50% of park revenue) along with grants and subsidies from conservation organizations, only a small portion (13.7%) of this budget was allocated to HWC prevention and mitigation67,68. Non-government conservation organizations have supported some households to build predator-proof sheds for small livestock in the buffer zones of Bardia and Chitwan51,69. Overall, however, the support is insufficient to meet the needs of all victims affected by livestock depredation, especially those of lower-income households who struggle to afford mitigation measures. Thus, increased funding and support for construction of predator-proof sheds is required from both the government and conservation organizations70.
Combined with habitat restoration and species-specific mitigation, a conflict hotspot-based management strategy could contribute to enhanced HWC mitigation. The southwestern sector of Bardia and the southern region of Chitwan have emerged as conflict hotspots, characterized by overlapping conflicts involving multiple species. The movement of megafauna from protected areas of India connecting to the southern region of Bardia and Chitwan through corridors may be the cause of spatial overlap of incident hotspots71. The connectivity provided by wildlife corridors, such as the Khata Corridor from the Katarniaghat Wildlife Sanctuary and the Shikaribas Corridor from the Valmiki Tiger Reserve, facilitates movement between habitats, increasing the pressure in conflict zones in both PAs72,73. We also found higher incident rates involving tigers and leopards in the northern buffer zone of Bardia, and higher incident rates scattered along the northern Chitwan, specifically for leopards. This pattern can be associated with livelihood strategies of local people. Based on our fieldwork in Bardia, we observed that the northern region of Bardia exhibited a high concentration of livestock holdings, which might contribute to an increased frequency of livestock depredation by tigers and leopards66. However, detailed studies are required to further understand the relationship between the density of livestock and predation risk. Besides, studies have reported that seasonal trans-border migratory movements of elephants can cause damages to crop and property40,41so the clustering of their conflict hotspots in SW Bardia and the southern buffer of Chitwan may result from their migration from Indian’s PAs. We recommend fine-scale research to understand megafauna movement and damage patterns in corridors and buffer zone forests.
Given the variable, but ever-present, risk of wildlife predation throughout the buffer zone, the broader adoption of community insurance could be a relevant strategy for buffer zone households. Insurance is currently adopted by only a few commercial livestock farmers in buffer zones. Our informal interviews and discussions with local stakeholders during fieldwork revealed that the widespread adoption of insurance by households has been limited by unaffordable annual premiums. Research should investigate the potential to create a spatially explicit community-based, multi-stakeholder insurance compensation model to compensate for high-value crop and livestock losses by wildlife (see74. Stakeholders include local governments, park authorities, Buffer Zone User Committees (BZUCs), and conservation organizations such as WWF, NTNC (National Trust for Nature Conservation), and ZSL (Zoological Society of London), all of which have been supporting mitigation efforts in the study area. To mitigate the high premiums, policy can further consider levying fees on ecotourism to help fund community-based insurance or relief from livestock depredation.
In conclusion, our study highlights the need for balancing conservation success and mitigation of HWC in rural communities. We provide the first quantitative association between wildlife population estimates and HWC for tigers and rhinos, suggesting that this trend is likely to be experienced where megafauna populations are rebounding or restored. Further, our work strengthens the call for greater attention on habitat expansion through restoration outside the protected areas in the buffer zones and corridor forest, which can help to extend wildlife habitat and enhance connectivity of populations, thus increasing carrying capacity and potentially mitigating HWC in the short term. In addition, mitigation must be enhanced: cost-effective community-based mitigation measures should be implemented in the peak raiding periods and predator-proof sheds are important to save the large livestock with high market value. Additionally, establishing community-based, multi-stakeholder livestock insurance schemes, is required in conflict hotspots to optimize resources and mitigate HWC impacts, which will secure the long-term conservation efforts.
Methods
Study area
The study was conducted in the buffer zones of BNP (28° 15’ to 28° 35.5’ N and 80° 10’ to 81° 45’ E, established in 1988 with an area of 968 km2 and CNP (27˚16.56’ to 27˚42.14’N and 83˚50.23’ to 84˚46.25’E; established in 1973 with an area of 953 km2 (Fig. 1). Both protected areas lie within the Terai Arc Landscape (TAL) of the tropical lowland (Terai) region of Nepal, a priority landscape for tiger conservation29,30. The Terai region has a subtropical climate characterized by a mean annual temperature range of 20–28 °C and annual rainfall of 1500–2000 mm75. The dominant forest type of lowland Terai is dry seasonal Sal (Shorea robusta) forest followed by riverine forest and mixed hardwood forest. Both protected areas have similar vegetation, landscape features, and rich biodiversity including several threatened megafauna species of global concern like Royal Bengal tiger (Panthera tigris tigris), common leopard (P. pardus), Asian elephant (Elephas maximus), and one-horned rhino (Rhinoceros unicornis), etc. These megafaunas have been frequently involved in HWC incidents within the buffer zones7,9,19. There is a high human population density within the buffer zone with the record of 263 people km− 1 in BNP and 297 people km−1 in CNP76,77. BNP lies within the western region and is connected to the Katerniaghat Wildlife Sanctuary in India via the Khata corridor, ungazetted national forest, and community forests72. CNP lies within the central region of Nepal and is connected to the Balmiki Tiger Reserve of India via the Shikaribas corridor (Fig. 1)73. Each buffer zone is subdivided into wards, which is the smallest unit of local government in Nepal. Within the two buffer zones, agriculture is a major livelihood strategy. In the Terai region, the primary crops cultivated include paddy, wheat, maize, and mustard for both subsistence and commercial use78. Major livestock raised by local communities include goat, sheep, pig, cattle, and buffalo, primarily for milk, meat, manure, and draught power79.
Data collection
After obtaining the research permit from the Department of National Parks and Wildlife Conservation (DNPWC), Nepal, we visited the central offices of BNP and CNP in January 2023 to obtain the park-specific research permits. These permits allowed us to access the records of the respective PA while adhering to the protocols and guidelines of the National Park and Wildlife Conservation Act 197380.
We reviewed and extracted data from the relief claims of victims, a method that has been employed in similar studies investigating HWC7,81. Each PA headquarter or main office (Thakurdwara in BNP and Kasara in CNP) has a “Buffer Zone and Monitoring Section” for collecting claims of wildlife victims and processing them for the distribution of relief funds. Victims must file their relief claims within 35 days of the incident occurrence for the approval of processing their paper-based relief claim applications82. Between January and April 2023, we gathered all the paper-based relief claims from 2013 to 2022 from the PA central offices. We then entered all claims into a database and classified each claim into one of the four main types of HWC incidents recognized by the PA main offices: human attack, livestock depredation, crop raiding, or structural property damage. From each incident type, we recorded the victim’s municipality and ward number, the damage or loss, the wildlife species involved, and the date of occurrence. Since our study focused on officially reported relief claims, it does not capture unreported cases of human fatalities and livestock depredation occurring within national park boundaries. Additionally, it excludes crop-raiding incidents on public lands cultivated by landless communities, who are ineligible for government relief under the current legal provisions.
We also obtained population census data of tigers and rhinos from the annual reports of BNP and CNP, species conservation action plans for tigers and rhinos, and relevant literature76,77,83,84,85. These two megafauna species (tiger and rhino) are the only ones for which periodic population censuses are conducted within our two study sites.
Data management and analysis
We quantified the total number of HWC incidents by species and by incident type across all years. Within each incident type the livestock species killed, crop species damaged, human casualty and injury incurred, or property damaged was further identified. A Pearson correlation was applied to both the tiger and rhino populations over time, to test for association between population estimates and the number of reported incidents. Similarly, a correlation test was run on the number of HWC incidents across years for leopard and elephant. We also computed incident density (no. incidents/km2) in each buffer zone ward for tiger, leopard, rhino, and elephant. Then, we joined a table of cumulative incident density of these four megafaunas for each ward to the shapefile of the ward. We defined the breaks for cumulative incident density using the “classInt” package in R, using the “quantile” method86. Conflict hotspots were defined as those wards in the top quantile, i.e. those with the highest density of incidents involving four megafauna species3. These spatial analyses were conducted using QGIS version 3.22. We adhered to the data storage and management protocols established by DNPWC, Nepal, and the University of Helsinki, Finland.
Data availability
The datasets generated and/or analyzed during the current study are not publicly available to protect the anonymity of individuals affected by wildlife incidents but are available from the corresponding author upon reasonable request.
References
Bagheriyan, E., Karimi, A. & Yazdandad, H. Assessing spatio-temporal patterns of human-wildlife conflicts in a human-dominated landscape: a case study from Iran. Biodivers. Conserv. 32, 4239–4257 (2023).
Meyer, M. & Börner, J. Rural livelihoods, community-based conservation, and human–wildlife conflict: scope for synergies? Biol. Conserv. 272, 109666 (2022).
Sharma, P., Chettri, N. & Wangchuk, K. Human–wildlife conflict in the roof of the world: Understanding multidimensional perspectives through a systematic review. Ecol. Evol. 11, 11569–11586. https://doi.org/10.1002/ece3.7980 (2021).
Tiller, L. N. et al. Changing seasonal, Temporal and Spatial crop-raiding trends over 15 years in a human-elephant conflict hotspot. Biol. Conserv. 254, 108941 (2021).
Barua, M., Bhagwat, S. A. & Jadhav, S. The hidden dimensions of human–wildlife conflict: health impacts, opportunity and transaction costs. Biol. Conserv. 157, 309–316 (2013).
Braczkowski, A. R. et al. The unequal burden of human-wildlife conflict. Commun. Biol. 6, (2023).
Lamichhane, B. R. et al. Spatio-temporal patterns of attacks on human and economic losses from wildlife in Chitwan National park, Nepal. PLoS One 13 (2018).
Pant, G., Dhakal, M., Pradhan, N. M. B., Leverington, F. & Hockings, M. Nature and extent of human-elephant Elephas maximus conflict in central Nepal. ORYX 50, 724–731. https://doi.org/10.1017/S0030605315000381 (2016).
Shahi, K. et al. Characterizing damages caused by wildlife: learning from Bardia National park, Nepal. Hum. Dimensions Wildl. 27, 173–182 (2022).
Silwal, T. et al. When, where and whom: assessing wildlife attacks on people in Chitwan National park, Nepal. ORYX 51, 370–377 (2017).
Tian, D., Xie, Y., Barnosky, A. & Wei, F. Defining the balance point between conservation and development. Conserv. Biol. 33 (2018).
Wei, F. et al. Balancing community livelihoods and biodiversity conservation of protected areas in East Africa. Curr. Opin. Environ. Sustain. 33, 26–33 (2018).
Chapron, G. et al. Recovery of large carnivores in europe’s modern human-dominated landscapes. Sci. (1979). 346, 1517–1519 (2014).
Duangchantrasiri, S. et al. Rigorous assessment of a unique tiger recovery in Southeast Asia based on photographic capture-recapture modeling of population dynamics. Glob Ecol. Conserv. 53 (2024).
Buchholtz, E. K., McDaniels, M., McCulloch, G., Songhurst, A. & Stronza, A. A mixed-methods assessment of human-elephant conflict in the Western Okavango panhandle, Botswana. People Nat. 5, 557–571 (2023).
Claude Twahirwa, J. et al. Positive population trends among meso- and megaherbivores follow intensive conservation efforts in volcanoes National park, Rwanda. Wildl. Biol. https://doi.org/10.1002/wlb3.01118 (2023).
Dellinger, J. A., Macon, D. K., Clifford, D. L. & Torres, S. G. Temporal trends and drivers of mountain Lion depredation in california, USA. 15, 162–177 (2021).
Mackenzie, C. A. & Ahabyona, P. Elephants in the garden: financial and social costs of crop raiding. Ecol. Econ. 75, 72–82 (2012).
Dhakal, S., Rimal, S., Paudel, P. & Shrestha, A. Spatio-Temporal patterns of livestock predation by leopards in Bardia National park, Nepal. Land (Basel) 12 (2023).
Gargallo, E. Human-Wildlife conflict in a ‘successful’ community conservation programme: economic and territorial impacts on namibia’s conservancies. J. Arid Environ. 193, 104591 (2021).
Okello, M. M., Manka, S. G. & D’Amour, D. E. The relative importance of large mammal species for tourism in Amboseli National park, Kenya. Tour Manag. 29, 751–760 (2008).
Van Der Meer, E., Badza, M. N. & Ndhlovu, A. Large carnivores as tourism flagship species for the Zimbabwe component of the Kavango Zambezi transfrontier conservation area. Afr. J. Wildl. Res. 46, 121–134 (2016).
Abdullah, A., Sayuti, A., Hasanuddin, H., Affan, M. & Wilson, G. People’s perceptions of elephant conservation and the human-elephant conflict in Aceh jaya, sumatra, Indonesia. Eur. J. Wildl. Res 65 (2019).
Kissui, B. M. Livestock predation by lions, leopards, spotted hyenas, and their vulnerability to retaliatory killing in the Maasai steppe, Tanzania. Anim. Conserv. 11, 422–432 (2008).
Oli, M. K., Taylor, I. R. & Rogers, M. E. Snow Leopard Panthera Uncia predation of livestock: an assessment of local perceptions in the annapurna conservation area, Nepal. Biol. Conserv. 68, 63–68 (1994).
Baral, K. et al. Anthropogenic mortality of large mammals and trends of conflict over two decades in Nepal. Ecol. Evol. 12 (2022).
Miller, J. R. B., Jhala, Y. V., Jena, J. & Schmitz, O. J. Landscape-scale accessibility of livestock to tigers: implications of Spatial grain for modeling predation risk to mitigate human-carnivore conflict. Ecol. Evol. 5, 1354–1367 (2015).
DNPWC. Protected Areas of Nepal. Department of National Parks and Wildlife Conservation, Babarmahal, Kathmandu, Nepal (2018).
Thapa, K. et al. Tigers in the Terai: strong evidence for meta-population dynamics contributing to tiger recovery and conservation in the Terai Arc landscape. PLoS One 12 (2017).
Thapa, K. et al. Assessment of genetic diversity, population structure, and gene flow of Tigers (Panthera Tigris Tigris) across nepal’s Terai Arc landscape. PLoS One 13 (2018).
Chaudhary, S., Uddin, K., Chettri, N., Thapa, R. & Sharma, E. Protected areas in the Hindu Kush himalaya: A regional assessment of the status, distribution, and gaps. Conserv. Sci. Pract 4 (2022).
Aryal, K., Dhungana, R. & Silwal, T. Understanding policy arrangement for wildlife conservation in protected areas of Nepal. Hum. Dimens. Wildl. 26, 1–12. https://doi.org/10.1080/10871209.2020.1781983 (2021)
Lamichhane, S., Joshi, R., Poudel, B. & Subedi, P. Role of community in leading conservation: effectiveness, success and challenges of community-Based Anti-Poaching unit in Nepal. Grassroots J. Nat. Resour. 3, 94–109 (2020).
DNPWC. Annual Report 2077/2078. Department of National Parks and Wildlife Conservation 154 (Government of Nepal, 2022).
Maron, D. F. Tigers have nearly tripled in nepal, but at what cost? National Geographic (2022).
DNPWC. Annual Report 2069/2070. Department of National Parks and Wildlife Conservation 89 (Government of Nepal, 2013).
Caro, T. M. & Stoner, C. J. The potential for interspecific competition among African carnivores. Biol. Conserv. 110, 67–75 (2003).
Young, T. P., Palmer, T. M. & Gadd, M. E. Competition and compensation among cattle, zebras, and elephants in a semi-arid savanna in laikipia, Kenya. Biol. Conserv. 122, 351–359 (2005).
Abie, G. G., Goudar, K. S., Ibrahim, H. & Getachew, G. Livestock Depredation and Pastoralist Attitudes towards Carnivore in the Landscape of Lions Associated with Gambella National Park, Abobo District, Western Ethiopia. Int. J. Zool. (2024).
Gross, E. M. et al. Elephants in the village: causes and consequences of property damage in Asia and Africa. Conserv. Sci. Pract. 3 (2021).
Neupane, D., Johnson, R. L. & Risch, T. S. How do land-use practices affect human-elephant conflict in nepal? Wildl. Biol. 2017 (2017).
Sijapati, R. K., Sharma, H. P., Sharma, S., Subedi, J. R. & Belant, J. L. Livestock depredation by leopards and tigers near Bardia national park, nepal. Animals 11 (2021).
DNPWC & DFSC. Status of Tigers and Prey in Nepal 2022. (p. 148). (2022).
Lovari, S., Pokheral, C. P., Jnawali, S. R., Fusani, L. & Ferretti, F. Coexistence of the tiger and the common Leopard in a prey-rich area: the role of prey partitioning. J. Zool. 295, 122–131 (2015).
Ram, A. & Acharya, H. B. Status Distribution and Habitat Use by Asian Elephants in Nepal. A Compendium of Conservation Bulletin Published by Department of National Parks and Wildlife Conservation, Nepal (2020).
Baral, K. et al. Characterization and management of human-wildlife conflicts in mid-hills outside protected areas of Gandaki province, Nepal. PLoS One 16 (2021).
Wegge, P., Yadav, S. K. & Lamichhane, B. R. Are corridors good for Tigers Panthera Tigris but bad for people? An assessment of the Khata corridor in lowland Nepal. ORYX 52, 35–45 (2018).
Athreya, V., Odden, M., Linnell, J. D. C., Krishnaswamy, J. & Karanth, U. Big cats in our backyards: persistence of large carnivores in a human dominated landscape in India. PLoS One 8 (2013).
Ram, A. K. et al. Dynamic occupancy modelling of Asian elephants (Elephas maximus) reveals increasing landscape use in Nepal. Sci. Rep. 14 (2024).
Kolipaka, S. S. Can Tigers survive in human-dominated landscapes. Leiden Univ. Leiden (2018).
WWF. World Wide Fund for Nature Nepal (WWF-Nepal). Annual Report 2023 (p.31). Baluwatar, Kathmandu, Nepal. (2023).
Beier, D. et al. The Role of Landscape Connectivity in Planning and The Role of Landscape Connectivity in Planning and Implementing Conservation and Restoration Priorities. Issues in Implementing Conservation and Restoration Priorities. Issues in Ecology Ecology. https://scholars.unh.edu/geog_facpub
Naha, D., Dash, S. K., Chettri, A., Roy, A. & Sathyakumar, S. Elephants in the neighborhood: patterns of crop-raiding by Asian elephants within a fragmented landscape of Eastern India. PeerJ 8, e9399 (2020).
Webber, C. E., Sereivathana, T., Maltby, M. P. & Lee, P. C. Elephant crop-raiding and human-elephant conflict in cambodia: crop selection and seasonal timings of raids. ORYX 45, 243–251 (2011).
Studsrød, J. E., Wegge, P., Park-People & Relationships The case of damage caused by park animals around the Royal Bardia National park, Nepal. Environ. Conserv. 22, 133–142 (1995).
Gross, E. M. et al. Seasonality, crop type and crop phenology influence crop damage by wildlife herbivores in Africa and Asia. Biodivers. Conserv. 27, 2029–2050 (2018).
Cai, J., Jiang, Z., Zeng, Y., Li, C. & Bravery, B. D. Factors affecting crop damage by wild Boar and methods of mitigation in a giant panda reserve. Eur. J. Wildl. Res. 54, 723–728 (2008).
Thapa, S. Effectiveness of crop protection methods against wildlife damage: A case study of two villages at Bardia National park, Nepal. Crop Prot. 29, 1297–1304 (2010).
Dar, N. I., Minhas, R. A., Zaman, Q. & Linkie, M. Predicting the patterns, perceptions and causes of human–carnivore conflict in and around Machiara National park, Pakistan. Biol. Conserv. 142, 2076–2082 (2009).
Dhungana, R. et al. Livestock depredation by leopards around Chitwan National park, Nepal. Mammalian Biology. 96, 7–13 (2019).
Kandel, S. R. et al. An Emerging Issue of Human-Leopard Conflict in the Human-Dominated Landscape of Mid-Hills: A Case Study from Tanahun District of Nepal. Int. J. Zool. (2023).
Pant, B., Sharma, H. P., Dahal, B. R., Regmi, S. & Belant, J. L. Spatio-temporal patterns of human-wildlife conflicts and effectiveness of mitigation in shuklaphanta National park, Nepal. PLoS One 18 (2023).
Karanth, K. & Sunquist, M. Behavioural correlates of predation by tiger (Panthera tigris), Leopard (Panthera pardus) and Dhole (Cuon alpinus) in nagarahole, India. J. Zool. 250, 255–265 (2000).
Tamang, B. & Baral, N. Livestock depredation by large cats in Bardia National park, nepal: implications for improving park-people relations. Int. J. Biodivers. Sci. Manage. 4, 44–53 (2008).
Ramesh, T. et al. Patterns of livestock predation risk by large carnivores in india’s Eastern and Western Ghats. Glob Ecol. Conserv. 24, e01366 (2020).
Upadhyaya, S. et al. (ed, K.) Determining the risk of predator attacks around protected areas: the case of Bardia National park, Nepal. ORYX 54 670–677 (2020).
DNPWC. Buffer Zone Management Guidelines 1999. Department of National Park and Wildlife Conservation (DNPWC), Kathmandu, Nepal. (1999).
Lamichhane, B. R. et al. Contribution of buffer zone programs to reduce Human-Wildlife impacts: the case of the Chitwan National park, Nepal. Hum. Ecol. 47, 95–110 (2019).
NTNC. Annual Report 2022. National Trust for Nature Conservation (NTNC) Khumaltar, Lalitpur, Nepal. (2022).
Thapa, K., Baral, S. & Rayamajhi, S. Effectiveness of Human-Snow Leopard co-existence measure- a systematic analysis. J. Nat. Conserv. 76, 126511 (2023).
Talukdar, B. & Sinha, S. Challenges and opportunities of transboundary rhino conservation in India and Nepal. Pachyderm 54, 45–51 (2013).
Bhatt, T. R., Castley, J. G., Sims-Castley, R., Baral, H. S. & Chauvenet, A. L. M. Connecting tiger (Panthera tigris) populations in nepal: identification of corridors among tiger-bearing protected areas. Ecol Evol 13 (2023).
Poudel, P. Shikaribas Corridor: Connecting Tri-Boundary for Wildlife Movement. NepalPress (2023).
Chen, S., Yi, Z. F., Campos-Arceiz, A., Chen, M. Y. & Webb, E. L. Developing a spatially-explicit, sustainable and risk-based insurance scheme to mitigate human–wildlife conflict. Biol. Conserv. 168, 31–39 (2013).
Karki, R., Talchabhadel, R., Aalto, J. & Baidya, S. K. New Climatic classification of Nepal. Theor. Appl. Climatol. 125, 799–808 (2016).
CNP. Chitwan National Park (CNP): Annual Report- Fiscal Year 2078/79 (2021/2022). Department of National Parks and Wildlife Conservation, Chitwan, Nepal. (2022).
BNP. Management Plan of Bardia National Park and Its Buffer Zone (2079/80-2083/84). (p. 300) (Bardia National Park Office, 2022).
Shrestha, N., Raes, D., Vanuytrecht, E. & Sah, S. K. Cereal yield stabilization in Terai (Nepal) by water and soil fertility management modeling. Agric. Water Manag. 122, 53–62 (2013).
Thapa Karki, S. Do protected areas and conservation incentives contribute to sustainable livelihoods? A case study of Bardia National park, Nepal. J. Environ. Manage. 128, 988–999 (2013).
GON. The National Parks and Wildlife Conservation Act 1973 (4th Amendment in 1993). Government of Nepal, Kathmandu, Nepal. (1973).
Acharya, K. P., Paudel, P. K., Neupane, P. R. & Köhl, M. Human-wildlife conflicts in nepal: patterns of human fatalities and injuries caused by large mammals. PLoS One 11 (2016).
MOFE. Guidelines for Compensation Payments on Damages from Wildlife (Third Amendment). Ministry of Forests and Environment, Kathmandu. (2017).
DNPWC. Tiger Conservation Action Plan (2023–2032) (Department of National Parks and Wildlife Conservation, Babarmahal, Kathmandu, 2023).
DNPWC. The Greater One-Horned Rhinoceros Conservation Action Plan for Nepal (2017–2021) Department of National Parks and Wildlife Conservation, Kathmandu, Nepal. (2017).
Thapa, K., Nepal, S., Thapa, G., Bhatta, S. R. & Wikramanayake, E. Past, present and future conservation of the greater one-horned rhinoceros Rhinoceros unicornis in Nepal. ORYX 47, 345–351. https://doi.org/10.1017/S0030605311001670 (2013).
Bivand, R. classInt: choose univariate class intervals. R package version 0.4-3. Computer software]. Retrieved from (2020). https://cranr-project.org. .
FRTC. Land Cover of Nepal (2000–2019). Forest Research and Training Centre, Kathmandu, Nepal (2022). RDS.1972729).
UNEP-WCMC & IUCN. Protected Planet: Protected Areas of Nepal; The World Database on Protected Areas (WDPA)/the Global Database on Protected Areas Management Effectiveness (GD-PAME) (UNEP-WCMC and IUCN, 2017).
Acknowledgements
Research was conducted under permit numbers 079080-954 and 079080-477 for Department of National Park and Wildlife Conservation and Department of Forests and Soil Conservation, Nepal. The authors thank Bardia and Chitwan National Parks for granting park-specific research permits. Financial support for the research came from the Russell E. Train Education for Nature Program (EFN) WWF US (grant no. EF14061), Rufford Foundation (grant no. 39423-2), and WWF Nepal (grant no. NP15629). BN was supported by grant number 20230015 from the Suomen Metsätieteellinen Seura. The authors gratefully acknowledge the work of the dedicated field assistants along with the support provided by local stakeholders throughout our fieldwork. Institutional Review Data collection protocols and survey tools were reviewed and approved by Ethical Review Board, Tribhuvan University, Office of the Rector, Research Directorate, Kathmandu, Nepal (ERBTU-081-004). The University of Helsinki Library provided Open Access funding.
Author information
Authors and Affiliations
Contributions
Conceptualization and methodology design – B.N. and E.L.W.; Data collection, material acquisition, and data curation – B.N.; Data analysis and visualization – B.N., J.J., and E.L.W.; Writing – original draft – B.N.; Supervision (lead) – E.L.W.; Writing – review and editing – led by E.L.W., with equal contributions from J.J., A.P.G., D.S. and N.J.H.; Funding acquisition – B.N.; Finalization and approval for submission – all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Neupane, B., Jamaludin, J., Gautam, A.P. et al. Balancing act: navigating increasing human-wildlife conflict amidst megafauna recovery in the tropical lowlands of Nepal. Sci Rep 15, 31201 (2025). https://doi.org/10.1038/s41598-025-16707-w
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-16707-w
