
Abstract
Progress in garlic cultivation requires the evaluation and selection of superior clones adapted to diverse agroecological conditions in Iran. This study aimed to assess the adaptability of six promising garlic clones in comparison with three local garlic populations over a two-year period (2020–2021). The experiment was conducted in Hamedan, Zanjan, and Kerman provinces using a randomized complete block design with four replications at each location. Significant differences were observed in yield and associated yield-related traits among the evaluated clones and parental populations. Importantly, the selected clones outperformed their respective parental populations, with clones 7.15 and 8.11 achieving average yields of 13,018 kg/ha and exhibiting superior performance across all locations. Traits most strongly influencing yield included neck diameter, plant biomass, fresh and dry bulb weight, bulb length and diameter, bulb weight, dry plant yield, allicin yield, pseudo-stem height, total plant height, and bulb height. Allicin content varied significantly across regions, with most of the selected clones showing higher concentrations compared to the parental populations. Principal component and cluster analyses grouped the six clones and three parental populations into two distinct clusters. Clones 7.8, 1.11, 8.11, 1.15, and 7.15 were classified together due to their superior yield potential and favorable yield components. Overall, the findings indicated that clones 7.15 and 8.11 are highly adaptable and are recommended for large-scale cultivation in all three studied regions.
Similar content being viewed by others

Introduction
Garlic (Allium sativum L.) is a diploid species of the family Alliaceae, characterized as a diploid plant. It is widely cultivated worldwide and ranks as the second most important allium crop after onion, owing to its significant nutritional and medicinal properties1,2,3. Native to Central Asia4, garlic has been domesticated for approximately 3,000 years5. According to FAOSTAT6, global garlic production in 2023 was projected at 28 million tons, cultivated across 1.6 million hectares, with an average yield of 17 t/ha. In Iran, annual garlic production is about 59,900 tons from 9,000 hectares, averaging 12.3 t/ha. China, India, Bangladesh, and Egypt are the leading producers, with China and India together contributing nearly 80% of global output. Garlic is among the oldest domesticated crops, with historical records of its use dating back 5,000 years, including prominent references in ancient Egypt and India7. Despite its long cultivation history and high economic and culinary value, garlic farming faces major constraints, particularly infertility in commercial cultivars and the gradual accumulation of pathogens. The foundation of any breeding program lies in the availability of genetically diverse germplasm for effective selection8. However, due to the progressive loss of flowering ability and sexual reproduction, garlic is propagated exclusively by vegetative means. As a result, crop improvement has primarily depended on exploiting existing genetic diversity, while efforts to expand it through mutation breeding and laboratory techniques have achieved only limited success9. Globally, approximately 600 garlic cultivars are currently recognized10.
Morphological traits such as bulb structure, leaf size, scape height, and the number of bulb scales are polygenically controlled, whereas traits including clove number per bulb and clove weight exhibit greater variability, suggesting stronger environmental influence or weaker genetic regulation11,12. A study by Alam Khomram et al.13 reported that three maternal garlic populations from Urmia, Arak, and Kerman performed well under the climatic conditions of Hamedan. Within these populations, Urmia, Solan, and Hidareh showed the highest allicin yields per hectare, measured via bulb yield and enzymatic pyruvic acid activity. Bulb weight was found to have a strong direct correlation with yield, while neck diameter exerted an indirect but significant influence through its association with bulb weight. Additional traits, including leaf area index, harvest index, leaf dry weight, and bulb diameter, were positively correlated with allicin production.
Evaluations of garlic ecotypes collected from different regions of Iran revealed substantial variation in bulb and clove characteristics. Bulb weight ranged from 16 to 62.34 g, while clove weight varied from 0.7 to 3.77 g. The number of cloves per bulb ranged widely, from 7 to 40. Most ecotypes exhibited purple outer skins, although a few clones displayed red or white coloration14. Similarly, studies have documented significant variability in bulb weight, fresh biomass, clove size (length, diameter, and width), neck diameter, plant height at harvest, and leaf width, indicating a high degree of morphological diversity among Iranian garlic ecotypes15,16. Germplasm studies on native Iranian garlic populations from Varamin, Khorasan, Zanjan, Hamedan, Mazandaran, Arak, and Ahvaz revealed that the populations from Arak (Tafresh), Tarom, and Urmia exhibited higher yields compared to those from other regions17. Research on the genetic diversity of Iranian garlic further indicated no significant correlation between genetic variation and geographical origin. Instead, genetic factors were shown to exert a stronger influence on the biochemical properties of garlic than environmental conditions14. In particular, allicin content in native Iranian populations appeared to be more strongly determined by genetic factors than by environmental influences14. Moreover, studies have reported that the allicin levels of many Iranian ecotypes surpass the medicinal threshold of 4.5 mg/g fresh weight14,18. In an evaluation of 10 ecotypes from Hamedan, total pyruvate content varied among populations. Shurin and Toin ecotypes exceeded 80 µmol/g fresh weight, Barfjin, Moeen, Aliabad, Bahar, and Solan ranged between 75 and 80 µmol/g fresh weight, while Tuyserkan, Merianj, and Hidareh fell below 75 µmol/g fresh weight19. Similarly, an analysis of 200 garlic samples collected from eastern Australia showed that allicin concentrations ranged from 0.5 to 9 mg/g fresh weight, with approximately half of the samples exceeding the pharmaceutical threshold20. Previously, cultivars with the highest recorded allicin levels included Romanian Red (7.7 mg/g fresh weight)21, Spanish Roja (7.6 mg/g), and Laotouxu (6.6 mg/g)22,23.
Despite this variability, the average garlic yield in Iran remains below the global standard, primarily due to the limited availability of improved cultivars. To date, the only registered improved cultivar in Iran is ‘Mazand,’ officially released in 200213. In an effort to address this limitation, 18 native garlic parental populations were collected from different regions of Iran during 2014–2015 and subsequently evaluated. Several promising clones were selected from these populations, and their yield performance was tested across multiple locations. The present study aims to assess the yield and yield components of six selected clones relative to their parental populations across diverse regions, with the ultimate goal of identifying the most promising garlic clones for large-scale cultivation.
Materials and methods
Site of the experiment and characteristics of garlic clones and indigenous populations
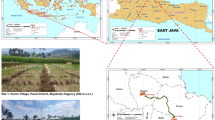
The present study was conducted over a two-year period (2020–2021) at research stations located in Ekbatan (Hamedan), Tarom (Zanjan), and Bardsir (Kerman). The research focused on three indigenous garlic populations derived from Bardsir (No. 8), Hidareh (No. 11), and Hosein Abad (No. 15) in Hamedan. In addition to these, two promising clones selected from each of the three populations were included, resulting in a comprehensive evaluation of nine clones together with their associated maternal populations (Fig. 1).
Between 2014 and 2015, nine local populations from Hamedan were sampled, originating specifically from Merianaj, Bahar, Shorin, Solan, Tuyserkan, Hidareh, Tuin, Hossein Abad, and Ali Abad. In addition, nine further local populations were collected from other provinces, including Gorgan (Golestan), Urmia (West Azerbaijan), Arak (Markazi), Bardsir (Kerman), Amol (Mazandaran), Ramhormoz (Khuzestan), Isfahan (Isfahan), Tarom (Zanjan), and Zabol (Sistan). These populations were initially assessed in Hamedan with respect to yield and its components, and subsequently, the top individual plants were selected through clonal selection13. Each clone was developed and propagated using the individual clonal selection breeding method employed in previous years. From each mother plant, eight garlic bulbs were selected on the basis of their desirable appearance and size. These bulbs were then propagated along separate lines for a duration of two years. During the third and fourth years, the clones were cultivated in separate plots, and once propagation had reached the desired scale, the clones were evaluated in both preliminary and advanced trials conducted over an additional two-year period.
In preparation for planting in the fall, several land management activities, including plowing, discing, and troweling, were carried out. Comprehensive soil testing was conducted at all three locations, following which ammonium phosphate fertilizer was incorporated into the soil at a rate of 100 kg/ha at planting. Urea fertilizer was applied at a total rate of 200 kg/ha across three stages during land preparation: after cultivation, prior to discing, and during ridge formation. In early spring, coinciding with the initiation of plant growth, two-thirds of the remaining urea fertilizer was applied in two cases as a slurry mixed with irrigation water, administered just prior to bulb formation. Each experimental plot consisted of a ridge measuring six meters in length and one meter in width, with four rows of garlic planted on each ridge at intervals of 25 cm. Prior to planting, all garlic bulbs were disinfected using Rural TS fungicide. Planting was carried out in late November, with bulbs from each clone or variety placed along the cultivation lines at a spacing of 10 cm and at a depth of 7–10 cm. The experimental layout therefore consisted of plots six meters long and one-meter wide, with four rows per plot and a row spacing of 25 cm, while an inter-row spacing of 10 cm was maintained.

Maternal populations from the provinces of Kerman (No. 8), Hamedan (Nos. 11 and 15), as well as locations in Zanjan, Kerman, and Hamedan, designated for the assessment of the adaptability of clones and parent populations. This map was generated utilizing ArcGIS version 10.8.2 software24.
The spacing of the plants along the rows was established at 10 cm, while the planting depth ranged from 5 to 7 cm. The distance between the experimental blocks (replications) was set at 3.5 m. Consequently, the populations and selected clones were planted uniformly at a density of 400,000 plants per hectare. The climatic conditions of the experimental sites are presented in Table 1.
As the physiological maturation of the plants advanced, as indicated by yellowing, visible color changes, and drying of the aerial parts, with approximately 70% of the leaves drying out, harvesting was performed between the latter half of May and the latter half of July, adjusted according to the specific regional conditions. At the initial stage, five carefully selected plants were harvested in their entirety. Each harvested plant was placed in a separate envelope, which was labeled with the corresponding sample code, plot number, block number (replication), and the year of the experiment. Following this initial collection, the plants within each experimental unit were topped prior to carrying out the final harvest. The following attributes were assessed in detail: neck diameter, pseudo stem height, apparent plant height, dry weight of both the shoot and the root, total garlic weight, clove moisture percentage, dry matter content of the garlic, the ratio of bulb dry matter to total plant dry matter, and garlic dimensions including length, width, diameter, and the sphericity coefficient. Pseudo stem height was determined by recording the average height, measured from the soil surface to the last leaf, in five selected plants.
Morphological characteristics
Neck diameter
During the period of maximum plant growth, and based on the cultivation area, five plants were selected. The maximum and minimum neck diameters were measured approximately one centimeter above the soil surface using a digital caliper. The average of these measurements was calculated to represent the neck diameter.
Apparent plant height
The average apparent height was determined from five selected plants.
Dry weight of shoots and roots
The average dry weight of leaves, pseudo stem, and roots within a radius of five centimeters around the garlic was measured in five selected plants.
Garlic weight
The average garlic weight from five selected plants was recorded in grams, measured using a digital scale with an accuracy of 0.1 g.
Percentage of clove moisture
This parameter was calculated based on the relative loss of clove slices, each 2–3 mm thick, after drying in an oven (Memmert UNB 100, Germany) at 75 °C for 48 h.
Garlic dry matter
Garlic dry matter was determined by calculating the average product of garlic weight and dry matter percentage for five selected plants.
Garlic length
The average maximum width of garlic, measured perpendicular to the direction of grain arrangement, was recorded in five selected plants using calipers.
Garlic width
The average minimum width of garlic, also measured perpendicular to the direction of grain arrangement, was determined in five selected plants using calipers.
Garlic diameter
The average thickness (or height) of garlic, measured in the direction of the grain arrangement, was recorded in five selected plants using calipers.
Sphericity coefficient
To determine the sphericity of the garlic, geometric dimensions were measured using calipers with an accuracy of 0.02 mm. The sphericity coefficient was then calculated according to the formula established by Mohsenin25.
The dimensions of garlic bulbs were denoted by the variables a, b, and c, corresponding to height, length, and width, respectively. A sphericity coefficient approaching unity indicated that the garlic bulb possessed a shape increasingly similar to a perfect sphere. In addition to these dimensional traits, data were systematically recorded on the quantity and weight of garlic skins, the number and weight of cloves, overall garlic yield, and dry matter yield.
Total pyruvic acid content
For sample preparation, 50 g of garlic were blended with 100 ml of distilled water to obtain a homogenate. Following ten minutes of blending, the extract was filtered through filter paper and diluted tenfold. To develop the colored complex, 50 µL of the diluted extract were combined with 2 ml of distilled water and 2 ml of 2,4-dinitrophenylhydrazine reagent in a test tube. The mixture was incubated in a water bath at 37 °C for eight minutes, after which the reaction was neutralized by adding 2 ml of 1.5 M sodium hydroxide. Sample concentrations were quantified against a standard curve prepared from sodium pyruvate solutions (1, 2, 3, 4, and 5 µL/ml), using a spectrophotometer (Pharmacia Novaspec II, Netherlands) at a wavelength of 515 nm, according to the method of Anthon and Barrett26.
Allicin estimation
Allicin content was quantified using a modified version of the procedure described by Khar et al.27. The calculation was based on the conversion of the measured amount of pyruvate, expressed as micromoles per gram of fresh weight. The final allicin content was expressed as milligrams per gram of fresh weight. In formula (2) for calculating the amount of allicin synthesized in fresh garlic, Khar et al.27 utilized alliin as the reference compound. In contrast, the present study employed pyruvate in place of alliin. In both formulations, the synthesis of one (µmol) of either alliin or pyruvate per (g) of fresh garlic corresponds to the production of one (mg) of allicin per (g) of fresh garlic.
Allicin yield
The yield of allicin was determined by applying the 2:1 molar ratio between the concentration of pyruvic acid and allicin, in conjunction with the molar weight of allicin, using the following equation for calculation.
Statistical analysis
The experiment was conducted using a randomized complete block design with four replications at each site. Data collection was carried out in Microsoft Excel 2016, which was used for recording raw observations. An analysis of variance (ANOVA) was performed, and mean comparisons were conducted using MiniTab 19.11 software. Differences among means were further examined using Tukey’s multiple range test at a significance level of p < 0.05. With the increasing number of germplasm samples incorporated into plant breeding programs, the classification and ranking of genetic variability have become increasingly important. Multivariate statistical methods provide a powerful framework for germplasm classification, ranking variability among numerous samples, and assessing genetic relationships among study materials. These approaches, which allow for the simultaneous evaluation of multiple measurements for each individual, are widely employed in studies of genetic diversity across different data types, including morphological, biochemical, and molecular traits. Among these, cluster analysis and principal component analysis are among the most frequently applied methods28,29. In the present study, principal component, correlation and cluster analysis were conducted using JMP 13 software.
Results
Agro-morphological and yield traits
The combined analysis of variance (Table 2) indicated mean comparisons and coefficients of variation (CV) for traits recorded over the two years (2021–2022) across three locations, i.e., Ekbatan (Hamedan), Tarom (Zanjan), and Bardsir (Kerman). The variance analysis revealed significant differences among locations, as well as significant interaction effects for year × environment, genotype × environment, and genotype × environment × year. However, no significant differences were observed for the main effect of year or for the genotype × year interaction in any of the measured traits (Table 2). Clones 7.15 and 8.11, with an average garlic yield of 13,018 kg/ha, were grouped in category A. In contrast, the three maternal populations, i.e., Bardsir Kerman (8), Hidareh Hamedan (11), and Hossein Abad Hamedan (15), produced an average yield of 11,390 kg/ha and were classified in category D. The yield performance of group A was 14.3% higher than that of group D, which represented the maternal populations. Among the three experimental sites, Ekbatan (Hamedan) exhibited the highest yields for both selected clones and maternal populations (Fig. 2).
Genotype × environment interactions, manifested as changes in genotype ranking across environments, represent a complex phenomenon shaped by environmental factors. These included agroecological conditions, climate, and agronomic practices, as well as the genetic determinants of plant growth and development30. Such evaluations require assessments across both temporal and spatial scales within the target environments where genotypes are intended for cultivation. Multi-environment trials remain the most effective approach for elucidating genotypic responses to environmental variation. The combined analysis of variance is the standard statistical method for detecting genotype × environment interactions in multi-environment experimental frameworks31,32.

The yield (kg/ha) of selected clones and garlic mother populations across three locations: Ekbatan, Tarom, and Bardsir.
The combined analysis of variance revealed that both genotype and the genotype × region interaction had a significant effect on neck diameter (p < 0.05). Furthermore, the interactions of year × region and genotype × year × region were also significant (p < 0.01) (Table 2). With the exception of clone 5.8 from the three parental populations, all garlic clones exhibited larger neck diameters. Among these, four clones, i.e., 7.8, 1.15, 7.15, and 1.11, had an average neck diameter of 11.01 mm and were classified into group A. In contrast, the three maternal populations, together with clone 5.8, which had an average neck diameter of 10.04 mm, were placed in group C (Fig. 3).

Neck diameter (mm) of selected garlic clones and their corresponding maternal populations.
The combined analysis of variance demonstrated that the effect of region on the pseudo-stem height of garlic clones and maternal populations was statistically significant (p < 0.05). In addition, the interaction effects of year × region, genotype × region, and genotype × year × region on pseudo-stem height were highly significant (p < 0.01) (Table 2). In this study, the pseudo-stem length of all selected clones exceeded that of the maternal populations. Clone 7.15, with an average pseudo-stem height of 136 mm, was placed individually in group A, whereas the three maternal populations, with an average pseudo-stem height of 116 mm, were classified in group D (Table 3).
Similarly, the combined analysis of variance indicated that the effect of region on plant height was significant (p < 0.05), while the effects of year × region, genotype, and genotype × year × region were highly significant (p < 0.01) (Table 2). Mean comparisons showed that three clones, i.e., 1.15, 7.15, and 7.8, had an average plant height of 554.9 mm and were assigned to group A. In contrast, two maternal populations, Bardsir (8) and Hidareh (11), with an average plant height of 517.8 mm, were categorized in group D (Fig. 4).

Plant height (mm) of selected garlic clones and their corresponding maternal populations.
Furthermore, the comprehensive analysis of variance revealed that both genotype and the interaction effects of year × region and genotype × year × region significantly influenced bulb weight (p < 0.01) (Table 2). With the exception of clone 5.8 from the three parental populations, all garlic clones exhibited higher bulb weight. Among them, clones 7.15, 8.11, and 7.8 achieved an average bulb weight of 57.92 g and were categorized in group A. Conversely, the three maternal populations, together with clone 5.8, which had an average bulb weight of 49.41 g, were classified in group C, reflecting lower bulb weight (Fig. 5).

Bulb weight (g) of selected garlic clones and their corresponding maternal populations.
The combined analysis of variance indicated that the factor of genotype had a significant effect on bulb dry matter (p < 0.05). Moreover, the interaction effects of year × region and genotype × year × region on bulb dry matter were highly significant (p < 0.01) (Table 2). In the evaluation of bulb dry matter content, all clones, with the exception of clone 5.8, showed higher dry matter values compared with the parental populations Bardsir (8), Hidareh (11), and Hossein Abad (15). Three clones, 7.15, 8.11, and 1.15, recorded an average bulb dry matter of 23.74 g and were grouped in category A. In contrast, the three parental populations, together with clone 5.8, which had an average bulb dry matter of 20.20 g, were classified in group D (Table 3).
The findings further showed that the interaction effects of year × region, genotype × region, and genotype × year × region on the proportion of bulb dry matter to total plant dry matter were highly significant (p < 0.01) (Table 2). Mean comparison (Table 3) indicated that the parental populations Hidareh (11) and Hossein Abad (15), along with clones 1.11, 1.15, and 8.11, were categorized in group A, with an average of 83.62%. Conversely, the Bardsir (8) population and clones 7.15, 7.8, and 5.8, which averaged 82.35% for this trait, were placed in group D.
For bulb length, the combined analysis of variance revealed significant effects of region, genotype, and genotype × region, as well as highly significant interaction effects of year × region and genotype × year × region (p < 0.01) (Table 2). According to the mean data comparison (Table 3), clones 7.15 and 7.8 exhibited the greatest average bulb length (58.88 mm) and were placed in group A, whereas the maternal populations Hidareh (11) and Hossein Abad (15) showed the lowest average bulb length (53.58 mm) and were assigned to group G.
The analysis of variance further demonstrated that genotype had a highly significant effect on bulb diameter (p < 0.01) (Table 2). Mean comparisons (Table 3) revealed that clones 7.8, 1.11, and 7.15, with an average bulb diameter of 36.12 mm, were categorized in group A. In contrast, the three parental populations had the smallest average diameter (33.40 mm) and were classified in group D.
Finally, the combined analysis of variance indicated that both region and the genotype × region interaction significantly affected the number of tunics (p < 0.05). In addition, the interaction effects of year × region and genotype × year × region on tunic number were highly significant (p < 0.01) (Table 2). The mean data comparison (Table 3) showed that clones 1.15 and 8.11, with an average of 6.78 tunic layers, were classified in group A. By contrast, all three parental populations together with clones 7.8 and 7.15, which averaged 6.32 tunic layers, were placed in group E.
Allicin and total pyruvate traits
Table 3 presents the average allicin yield per hectare across the evaluated clones. Clones 7.8 and 8.11 produced the highest yields, averaging 65.40 kg/ha, and were therefore classified in group A. In contrast, the Hosein Abad maternal population exhibited the lowest yield, 52.18 kg/ha, and was assigned to group C. Furthermore, among the cultivation regions, the Ekbatan region of Hamadan recorded the highest allicin yields for both selected clones and maternal populations, outperforming the other two regions (Fig. 6).

Allicin yield per hectare across various selected clones, their parental populations, and distinct garlic cultivation sites.
The analysis of variance revealed a significant genotype × region interaction effect on total pyruvate levels (p < 0.05), while the interaction effects of year × region and genotype × year × region were highly significant (p < 0.01) (Table 2). Mean comparison results showed that clones 7.8 and 5.8 had the highest average total pyruvate concentration (61.29 µmol/g fresh bulb) and were classified in group A. In contrast, the Hossein Abad (15) population together with clone 7.15 exhibited the lowest average concentration (57.65 µmol/g fresh bulb) and were assigned to group D (Fig. 7).

The total pyruvate levels (µmol/g) for selected garlic clones and their respective mother populations.
Correlation between traits and yield
The correlation analysis (Fig. 8) revealed significant interrelationships among the evaluated traits, highlighting their importance for breeding programs and for selecting superior clones from maternal stocks. Garlic yield showed very strong positive correlations with several traits, including clove weight (r = 0.982), bulb weight (r = 0.982), plant biomass (r = 0.960), dry matter yield (r = 0.960), bulb dry matter (r = 0.930), bulb length (r = 0.932), allicin yield (r = 0.922), and neck diameter (r = 0.910), all highly significant at p < 0.001. Additional significant correlations (p < 0.01) were observed with bulb width (r = 0.870), plant height (r = 0.800), bulb height (r = 0.770), and pseudo-stem height (r = 0.770). As expected, larger bulbs, together with greater bulb and clove weight, were associated with higher yields.
Allicin yield was also strongly correlated with several traits. It showed very strong positive associations with plant biomass (r = 0.940), garlic yield (r = 0.922), and bulb length (r = 0.920), all significant at p < 0.001. Significant correlations (p < 0.01) were detected with bulb height (r = 0.880), clove weight (r = 0.870), bulb weight (r = 0.870), neck diameter (r = 0.850), bulb width (r = 0.820), and dry matter yield (r = 0.810). At p < 0.05, allicin yield was positively correlated with bulb dry matter (r = 0.740) and plant height (r = 0.690). Collectively, these results indicate that allicin yield is closely associated with garlic yield and its key yield-dependent components (Fig. 8).

Pearson’s correlation coefficients between the studied traits. In this figure, Y1 through Y19 represent various parameters including yield, neck diameter, pseudo stem height, plant height, plant biomass, bulb weight, percentage of bulb humidity, bulb dry matter, percentage ratio of bulb dry matter to plant dry matter, bulb length, bulb width, bulb height, sphericity coefficient, number of tunics, clove number, clove weight, dry matter yield, total pyruvate content, and allicin yield, respectively. ns, *, ** and *** are non- significant, significant at the 5, 1 and 0.1% probability levels, respectively.
Principal component analysis and cluster analysis
Principal Component Analysis (PCA) was applied to distinguish selected clones from their corresponding mother populations, effectively grouping them based on trait performance. The first two principal components explained 79.11% of the total variance, with PC1 accounting for 61.12% and PC2 for 11.61%. The PCA results indicated that seven garlic clones and three mother populations clustered into two distinct groups (Fig. 9). Clones 7.8, 1.11, 8.11, 1.15, and 7.15 were grouped together due to their superior yield and yield components relative to the mother populations and clone 5.8. Notably, clone 7.8 was separated from clones 1.11, 8.11, 1.15, and 7.15 on the basis of its elevated total pyruvate concentration and higher bulb moisture content. Among the mother populations, Hidareh (11) and Hossein Abad (15) were distinguished from Bardsir (8) and clone 5.8 due to their higher ratio of bulb dry matter to plant dry matter. Differentiation along PC1 was primarily driven by yield-related traits, including neck diameter, pseudo-stem height, plant height, plant dry matter, bulb weight, bulb dry matter, bulb diameter, bulb height, dry matter yield, and allicin content. These variables are generally classified as yield traits and components. In contrast, PC2 was influenced negatively by bulb moisture percentage, the ratio of bulb dry matter to plant dry matter, number of cloves, and total pyruvate concentration (Table 4).

Principal component analysis of the selected clones and parental garlic populations.
Cluster analysis supported the results of the principal component analysis, grouping seven clones and three parental garlic populations into two main clusters. The first cluster comprised the three maternal populations, i.e., Hidareh (11), Hossein Abad (15), and Bardsir (8), together with clone 5.8. This group was characterized by higher values for the sphericity coefficient, bulb moisture percentage, and the ratio of bulb dry matter to plant dry matter. Within this cluster, clone 5.8 differed from the three parental populations by exhibiting a particularly high bulb moisture percentage and a reduced ratio of bulb dry matter to plant dry matter. The second cluster included clones 7.8, 1.11, 8.11, 1.15, and 7.15. These clones were distinguished by superior yield performance and enhanced yield components, including neck diameter, plant height, plant dry matter, bulb weight, bulb dry matter, bulb diameter, dry matter yield, and allicin content, all of which were greater than those of the parental populations and clone 5.8. Traits related to yield and its components had a strong and positive influence on the formation of this cluster. Within the second cluster, two subgroups were identified: clones 7.8 and 7.15 formed one subgroup due to their higher performance-related trait values, while clones 1.11, 8.11, and 1.15 formed the second subgroup (Fig. 10).

Cluster analysis illustrating the grouping of selected clones alongside their garlic parental populations.
Discussion
Genetic diversity is fundamental to the success of any breeding program, whether it occurs naturally within a population or is deliberately introduced by breeders, typically through traditional crossbreeding techniques. In the case of garlic populations, however, crossbreeding is not feasible, making it necessary to optimize the use of existing natural diversity. Although asexual reproduction prevents the generation of new genetic combinations through recombination, clonal selection strategies can still be applied effectively. These strategies focus on a single garlic variant while exploiting the limited but enduring diversity within populations that have persisted and expanded over many years under a range of biotic and abiotic stresses. Through this process, cultivars may be developed that exhibit relative resistance to the predominant stresses of specific regions. The clonal selection method for asexually propagated crops is a long-established and highly effective approach33. Moreover, Kallo34 emphasized that mass clonal selection is particularly effective for maintaining varieties within subspecies and is also beneficial during the early stages of population development and selection. The present study revealed significant differences in yield and a range of yield-associated traits, including neck diameter, plant height, fresh and dry weight of garlic, garlic length and height, bulb weight, and dry matter yield, between selected clones and their parental populations. The results further demonstrated that individual clonal selection from native Iranian garlic populations can produce offspring that display diversity in several agronomic traits, despite the sterility and asexual mode of reproduction characteristic of garlic. This observed diversity is most likely attributable to the accumulation of advantageous random mutations over long periods of traditional cultivation, combined with the adaptive efforts of local farmers who have worked extensively with indigenous garlic varieties. Supporting evidence from previous research underscores the presence of considerable diversity within garlic populations. Mohammadi et al.35 examined the agronomic and morphological diversity of 16 native Iranian garlic populations collected from five regions, i.e., Hamedan, Gilan, Isfahan, Markazi, and Mazandaran, and reported significant variation in traits such as plant height, plant weight, bulb weight, bulb quantity, as well as both fresh and dry bulb weights and bulb dimensions. Likewise, Sood et al.36 investigated genetic diversity among five garlic cultivars in India and found substantial variation in yield-related characteristics, including fresh and dry yield, bulb diameter, bulb length, and bulb weight. Del-Pozo et al.37 also identified extensive variability in garlic clump production timing and reported significant genetic diversity in photoperiod and temperature responses. Furthermore, Gomez-Riera38 evaluated the agronomic performance of 37 garlic clones and documented notable differences in dormancy periods across these clones.
The findings of the experiment demonstrated that the performance of garlic clones and their parental populations varied across different regions and climatic conditions. Specifically, the performance recorded in the Hamedan region was superior to that observed in the Tarom and Bardsir regions. In addition, the yield of the selected clones exceeded that of the parental populations. Among these, clones 7.15 and 8.11 achieved an average yield of 13,018 kg/ha, representing the highest yield recorded across all three regions evaluated. Findings reported by Mengistu et al.39 also confirmed substantial variability in bulb yield and related traits among garlic genotypes, which were influenced by both genetic factors and environmental conditions. The interaction between genotype and environment was shown to be significant, underscoring the importance of evaluating genotypes across diverse environments to identify those that combine stability with high yield potential. The development of garlic cultivars that demonstrate broad adaptability and consistent performance is therefore essential for enhancing productivity and ensuring economic viability. Furthermore, the location of cultivation plays a crucial role in determining the characteristics of a cultivar, as numerous studies have indicated that variations in climate exert a pronounced effect on flower stalk formation and flavor40. Research conducted by Jabbes et al.41 on the diversity of 31 garlic populations in Tunisia revealed that several traits, including bulb weight, bulb diameter, number of leaves per plant, and stem length, had a significant influence on yield. Similarly, a study by Alam et al.42 on garlic germplasm in Bangladesh demonstrated that yield and its associated traits, including plant height, number of leaves, and both fresh and dry bulb weights, were strongly determined by genotype. Among the genotypes examined, G19 was identified as the most productive in terms of yield and related agronomic characteristics. Supporting this, Khadi et al.43 reported significant differences among 25 garlic genotypes in relation to growth and yield parameters. Within their study, the native genotype Gulbarga exhibited superior growth characteristics, including plant height, leaf length, and both fresh and dry weights, as well as enhanced yield traits such as bulb height, bulb diameter, bulb weight, and overall yield per hectare.
Further investigations by Popa et al.44 focused on the morphological traits of autumn garlic genotypes and demonstrated significant variability among 14 Romanian genotypes in relation to bulb weight, bulb size, and bulb number. Their findings emphasized that both genotype and environmental factors, including the year of assessment, influenced these traits, with native genotypes performing more favorably than registered varieties. In the correlation matrix (Table 4), the majority of the traits examined exhibited strong intercorrelations, suggesting that particular characteristics may be targeted in garlic breeding programs to optimize both time and labor efficiency. The correlation analysis conducted in this study provided important insights into the traits most critical for estimating yield. These traits included neck diameter, plant biomass, fresh and dry bulb weights, bulb length and diameter, bulb weight, bulb dry yield, plant dry yield, allicin yield, pseudo stem height, plant height, and bulb height. Each of these traits was found to exert a direct influence on garlic yield, with increases in these components being positively correlated with enhanced yield. Supporting these findings, Jabbes et al.41 confirmed that bulb weight, bulb diameter, number of leaves per plant, and stem length significantly influence yield. Similarly, Wang et al.45 reported a strong positive correlation between bulb yield and both bulb weight and bulb diameter, indicating that larger bulb size and greater weight are strongly associated with higher yields. In relation to Nepali climatic conditions, Panthee et al.46 identified clove diameter and number, bulb weight and diameter, number of leaves, and plant height as the primary determinants of garlic yield.
Allicin is the principal bioactive compound produced by the garlic plant and is recognized as the most biologically active sulfur-containing compound, responsible for garlic’s characteristic aroma and flavor. Within the garlic bulb, it initially exists in the form of alliin, which is converted into allicin when the bulb is cut or crushed. The results of this study demonstrated that the allicin content of garlic clones and populations was strongly influenced by geographical location and climatic conditions. Significant variation in allicin levels was observed across the three experimental sites, with Hamedan showing the highest concentrations, followed by Tarom, and Bardsir exhibiting the lowest. Furthermore, the selected garlic clones, except for clone 5.8, displayed higher allicin concentrations compared with their parental populations. Allicin yield was also found to have a strong and significant positive correlation with several agronomic traits, including garlic yield per hectare, neck diameter, plant biomass, bulb weight, bulb length and diameter, bulb height, clove dry weight, and dry matter yield. Additional significant positive correlations were observed with pseudo-stem height, bulb dry matter, and plant height. By contrast, allicin yield did not exhibit a significant correlation with total pyruvate content. Previous studies have likewise reported that allicin levels are influenced by both geographical and genetic factors14,47, corroborating the findings of the present work. For example, Akbarpour et al.48, in an evaluation of the yield and phytochemical composition of eight Iranian garlic ecotypes, observed that the Hamedan and Kerman ecotypes contained higher concentrations of allicin, alliin, total phenolics, total flavonoids, and total antioxidant capacity compared with the other ecotypes examined. In another study, Wang et al.45 reported that the allicin content of 212 garlic cultivars grown under identical environmental conditions ranged from 0.81 to 3.01%, indicating that genetic factors contribute substantially to the observed variation. Similarly, Jabbes et al.41 noted pronounced differences in allicin, isoalliin, GluAlCs, IsoGluAlCs, and total allicin content among native Tunisian garlic cultivars.
A broad consensus among researchers suggests that, despite variations in allicin content reported across studies, considerable genetic diversity exists among garlic cultivars in relation to allicin levels, thereby providing opportunities for the genetic improvement of garlic quality18,23,45,47,49,50. Wang et al.45 further documented a significant positive correlation between allicin content and pseudo-stem diameter, a finding that supports the present study and suggests that this trait could serve as a practical indicator for identifying garlic genotypes with elevated allicin levels. Consistent with the results obtained here, they also reported no significant correlation between allicin yield and total pyruvate content, a weak association that may be explained by the existence of distinct biochemical pathways involved in allicin synthesis. In general, the biochemical process underlying allicin formation occurs when garlic cloves are crushed, thereby activating the enzyme alliinase, which catalyzes the conversion of the precursor alliin into allicin51,52.
The present study employed Principal Component Analysis (PCA) to distinguish between selected garlic clones and their corresponding mother populations, effectively grouping them into distinct categories based on 17 morphological traits in addition to total pyruvate and allicin content. By applying this approach, clones characterized by both high yields and elevated allicin content were identified in comparison with their parental populations and Clone 5.8. PCA is a central technique in multivariate statistical analysis, designed to reduce the dimensionality of large datasets composed of correlated variables, thereby generating new uncorrelated components. These principal components encapsulate the comprehensive variation within the dataset12,53. In related research, Eghlima et al.54 applied PCA to evaluate various ecotypes of Equisetum arvense, where agro-morphological traits were classified into three principal components that together explained 96.77% of the total variance. Complementing PCA, cluster analysis is an approach that utilizes morphological traits and functions as a traditional breeding tool for assessing biological diversity while facilitating the large-scale selection of genetic variation in breeding programs12. In the current study, cluster analysis reinforced the results obtained through PCA, categorizing seven clones along with three parental garlic populations into two distinct groups. This method effectively differentiated the maternal populations and Clone 5.8, both of which exhibited comparatively lower yields and reduced allicin levels, from the remaining clones. Further supporting the utility of multivariate techniques, Eghlima et al.55 demonstrated that PCA combined with heatmap clustering can be employed to identify optimal cultivation regions for Glycyrrhiza glabra and to predict phytochemical production. Similarly, Panthee et al.46 conducted a PCA of 190 garlic genotypes collected from diverse regions of Nepal, classifying them into three groups on the basis of 17 morphological traits. Their analysis revealed that bulb weight and diameter, together with the number of cloves per bulb, were the most influential traits affecting yield. In addition, Ayed et al.56 carried out a cluster analysis of 36 native Tunisian garlic genotypes, dividing them into three groups according to 14 morpho-agronomic traits. One of these groups was distinguished by the inclusion of 20 of the most important Tunisian genotypes, which were characterized by superior yield performance, bulb weight, clove weight and length, leaf dimensions, and pseudo-stem diameter.
Conclusion
In the present study, the yield and associated traits, including neck diameter, plant height, fresh and dry weight of garlic, bulb length and diameter, clove weight, and dry matter yield, exhibited significant variability among the clones and their corresponding parental populations. Several traits were identified as critical for yield estimation, including neck diameter, plant biomass, fresh and dry bulb weight, bulb dimensions, clove weight, dry plant yield, allicin yield, pseudo-stem height, total plant height, and bulb elevation. Each of these traits exerted a direct influence on yield outcomes. The performance of garlic clones and their parental populations varied across regions and climatic conditions. Among the study sites, the performance recorded in the Hamedan region was superior to that observed in both Tarom and Bardsir. The yield of the selected clones consistently surpassed that of the parental populations, with Clones 7.15 and 8.11 achieving the highest performance, recording an average yield of 13,018 kg/ha across all three regions. The allicin content of garlic clones and populations was also shown to be influenced by geographical location and climatic conditions, with significant differences documented across the three experimental sites. Allicin concentrations were highest in Hamedan, followed by Tarom, and lowest in Bardsir. Moreover, the selected garlic clones, except for clone 5.8, exhibited higher allicin levels compared to their parental populations. In addition, allicin yield demonstrated a strong and significant positive correlation with a range of agronomic traits, including garlic yield per hectare, neck diameter, plant biomass, bulb weight, bulb length and diameter, bulb height, clove dry weight, and dry matter yield. These findings highlight the integral role of both morphological and biochemical traits in determining garlic yield potential. In future research concerning clonal selection programs utilizing garlic parental populations, it is advisable to make selections grounded in these traits to identify promising clones with elevated allicin content.
Data availability
The data used in this study are available the corresponding author on reasonable request.
References
Singh Batth, G., Kumar, H., Gupta, V. & Singh Brar, P. GGe biplot analysis for characterization of Garlic (Allium sativum L.) germplasim based on Agro-Morphological traits. IJPB 7 (2), 106–110 (2013).
Kamenetsky, R., Khassanov, F., Rabinowitch, H. D., Auger, J. & Kik, C. Garlic biodiversity and genetic resources. Med. Aromatic Plant. Sci. Biotechnol. 1 (1), 1–5 (2007).
Nosraty, A. E. Effect of planting method, plant density and seed clove size on yield of Hamedan Garlic. SPJ 20, 401–404. https://doi.org/10.22092/spij.2017.110592 (2004).
Kamenetsky, R. Garlic: Botany and horticulture. In: Plant Breeding Reviews 29. Janick J (eds). 123–172. https://doi.org/10.1002/9780470168011.ch2 (John Wiley & Sons, 2005).
Ovesná, J. et al. Diversity of S-alkyl cysteine sulphoxide content within a collection of Garlic (Allium sativum L.) and its association with the morphological and genetic background assessed by AFLP. Horti Sci. 129, 541–547. https://doi.org/10.1016/j.scienta.2011.06.009 (2011).
F. A. O. S. T. A. T. fao.org/faostat/en/#data/QC. (2023).
Parreño, R. et al. Turning Garlic into a modern crop: state of the Art and perspectives. Plants (6), 12. https://doi.org/10.3390/plants12061212 (2023).
Swarup, S. et al. Genetic diversity is indispensable for plant breeding to improve crops. Crop Sci. 61 (2), 839–852. https://doi.org/10.1002/csc2.20377 (2021).
Zheng, S. J. et al. Garlic breeding system innovations. Med. Aromat. Plant. Sci. Biotechnol. 1 (1), 6–15 (2007).
Zheng, S. J. et al. Shemesh-Mayer E, Kamenetsky-Goldstein R Traditional and novel approaches in Garlic (Allium sativum L.) breeding. In (eds Al-Khairy, J. M., Mohan Jain, S. & Jhonson, D. V.) Advances in Plant Breeding Strategies: Vegetable Crops. Bulb, Roots and Tubers. Springer Nature Switzerland, Switzerland, 3–49. (2021).
Polyzos, N., Papasotiropoulos, V., Lamari, F. N., Petropoulos, S. A. & Bebeli, P. J. Phenotypic characterization and quality traits of Greek Garlic (Allium sativum L.) germplasm cultivated at two different locations. Genet. Resour. Crop Evol. 66, 1671–1689. https://doi.org/10.1007/s10722-019-00831-4(01234567 (2019).
Tesfaye, A. Genetic divergence and cluster analysis for bulb yield and related traits in Garlic (Allium sativum) accessions at dorze, Southern Ethiopia. Agrosystems Geosci. Environ. 5, e20269. https://doi.org/10.1002/agg2.20269 (2022).
Alemkhoumaram, M. H., Keshtkar, A. H. & Mirzaie Asl, A. Study of agromorphological diversity, and path analysis of bulb yield and allicin content of Iranian Garlic landraces under Hamedan region. JHORTS 33 (3–3), 481–497 (2019).
Baghalian, K., Ziai, S. A., Naghavi, M. R., Naghdi Badi, H. & Khalighi, A. Evaluation of allicin content and botanical traits in Iranian Garlic (Allium sativum L.) ecotypes. Sci. Hortic. 103, 155–166. https://doi.org/10.1016/j.scienta.2004.07.001 (2005).
Abasifar, A. R. & Dashti, F. Study of relationship between morphological traits and flowering in Iranian garlic clones. IJHS 46 (1), 63–75. http://doi.10.22059/ijhs.2015.54126 (2015).
Kakaei, M. Morphological evaluation of different local ecotypes of Iranian Garlic (Allium sativum L.) under nonirrigated conditions in Asadabad region (Hamadan province). Taxonomy Biosystematics. 15 (56), 95–112. https://doi.org/10.22108/TBJ.2024.141167.1255 (2024).
Zarbakhsh, A. Investigating the Collection of Native Garlic Populations in Different Regions and Studying the Effects of Climate on its Morphological Characteristics Frost number 81/586. (Seed and Plant Improvement Institute, 2002).
Mirzaei, R., Liaghati, H. & Mahdavi Damghani, A. Evaluating yield quality and quantity of Garlic as affected by different farming systems and Garlic clones. PJBS 10, 2219. https://doi.org/10.3923/pjbs.2007.2219.2224 (2007).
Abedi, M., Bayat, F. & Nosrati, A. E. Evaluation of agronomical traits and pyruvic acid content in Hamedan garlic (Allium Sativum L.) ecotypes. Eur. J. Exp. Biol. 3 (2), 541–544. http://doi.10.5829/idosi.wasj.2013.22.05.1371 (2013).
Eagling, D. & Sterling, S. A cholesterol-lowering Extract from Garlic: a Report for the Rural Industries Research and Development Corporation (RIRDC publication, 2000).
Koch, H. P. & Lawson, L. D. Garlic. The Science and Therapeutic Application of Allium Sativum and Related Species 2nd edn (Williams and Wilkins, 1996).
Lawson, L. D., Wood, S. G. & Hughes, B. G. HPLC analysis of allicin and other thiosulfinates in Garlic clove homogenates. Planta Med. 57, 263–270. https://doi.org/10.1055/s-2006-960087 (1991).
Camargo, A., Masuelli, R. & Burba, J. L. Characterization of Argentine Garlic cultivars for their allicin content. Acta Hortic. 688, 309–312. https://doi.org/10.17660/ActaHortic.2005.688.44 (2005).
Anonymous. https://community.esri.com/t5/data-loading-tools-blog/arcgis-desktop-arcmap-10-8-2-the-latest-and-last/ba-p/1368034 (2024).
Mohsenin, N. N. Physical Properties of Plant and Animal Materials. Structure, Physical Characteristics and Mechanical Properties 2nd Edn. 11 https://doi.org/10.1002/food.19870310724 (Gordon and Breach Science, 1986).
Anthon, G. E. & Barrett, D. M. Modified method for the determination of pyruvic acid with dinitrophenylhydrazine in the assessment of onion pungency. J. Sci. Food Agric. 83 (12), 1210–1213. https://doi.org/10.1002/jsfa.1525 (2003).
Khar, A., Banerjee, K., Jadhav, M. R. & Lawande, K. E. Evaluation of Garlic ecotypes for allicin and other allyl thiosulphinates. Food Chem. 128, 988–996. https://doi.org/10.1016/j.foodchem.2011.04.004 (2011).
Manly, B. F. J. Multivariate Statistical Methods: A Primer. 2nd Edition. (Chapman & Hall, 1994).
Brown-Guedira, G. L., Thompson, J. A., Nelson, R. L. & Warburton, M. L. Evaluation of genetic diversity of soybean introductions and North American ancestors using RAPD and SSR markers. Crop Sci. 40 (3), 815–823. https://doi.org/10.2135/cropsci2000.403815x (2000).
Annicchiarico, P. Genotype × Environment Interactions: Challenges and Opportunities for Plant Breeding and Cultivar Recommendations. FAO Plant Production and Protection Paper No. 174 (FAO, 2002).
Cochran, W. G. & Cox, G. M. Experimental Design. 2nd Edition, 615. (John Wiley and Sons, 1957).
Yazdi Samadi, B., Rezaei, A. & Valyzadeh, M. Statistical Designs in Agricultural Research 764 (Tehran University Publications, 1997).
Herdia-Zepeda, A. Number of cloves per bulb: selection criteria for Garlic improvement. I. Results with ‘chileno’ type. Acta Hortic. 433, 265–270. https://doi.org/10.17660/ActaHortic.1997.433.26 (1997).
Kallo, G. Vegetable Breeding I (CRC Press, 1988).
Mohammadi, B., Khodadadi, M., Karami, E. & Shaaf, S. Variation in Agro-morphological characters in Iranian Garlic landraces. Int. J. Veg. Sci. 20, 202–215. https://doi.org/10.1080/19315260.2013.788594 (2014).
Sood, D. R., Chokar, V. & Singh, J. Studies on growth, pungency and flavor characteristics of five varieties of Garlic (Allium sativum L.) bulbs during development. Veg. Sci. 27 (2), 180–184 (2000).
Ines-Gonzalez, D. P. A., Barraza, M. & C Phenological development of 13 clones of Garlic (Allium sativum L.): influence of temperature, photoperiod and cold storage. Acta Hortic. 433, 389–394. https://doi.org/10.17660/ActaHortic.1997.433.40 (1997).
Gomez-Riera, J. L. B. Characterization, adaptation and selection of Garlic germplasm (Allium sativum L.) through the management of dormancy, in mendoza, Argentina. Acta Hortic. 433, 151–164. https://doi.org/10.17660/ActaHortic.1997.433.14 (1997).
Mengistu, F. G., Mossie, G. A. & Fita, G. T. Evaluation of Garlic genotypes for yield performance and stability using GGE biplot analysis and genotype by environment interaction. Plant. Genet. Res. Charact. Utili. 1–7. https://doi.org/10.1017/S1479262123000904 (2023).
Khar, A., Hirata, S., Abdelrahman, M., Shigyo, M. & Singh, H. Breeding and genomic approaches for climate-resilient garlic. In: Genomic Designing of Climate-Smart Vegetable Crops (Kole C. (eds). https://doi.org/10.1007/978-3-319-97415-6_8 (Springer, 2020).
Jabbes, N., Arnault, I., Auger, J., Al Mohandes Dridi, B. & Hannachi, C. Agro-morphological markers and organo-sulphur compounds to assess diversity in Tunisian Garlic landraces. Sci. Hortic. 148, 47–54. https://doi.org/10.1016/j.scienta.2012.08.013 (2012).
Alam, M. S., Rahim, M. A. & Simon, P. W. Performance evaluation of Garlic germplasms under dry land condition. J. Agrofor. Environ. 3 (2), 43–45 (2010).
Khadi, P., Hiremath, S. M., Mannikeri, I. M. & Tippannavar, P. S. Evaluation of Garlic (Allium sativum L.) genotypes for growth and yield under Northern zone of Karnataka. J. Farm. Sci. 35 (2), 247–249. https://doi.org/10.61475/jfm.v35i02.82 (2022).
Popa, P. M. & Cosmulescu, S. Evaluation and comparison of morphological traits in Garlic as influenced by year and genotypic diversity. J. Cent. Eur. Agric. 25 (4), 978–987. https://doi.org/10.5513/JCEA01/25.4.4321 (2024).
Wang, H., Li, X., Shen, D., Oiu, Y. & Song, J. Diversity evaluation of morphological traits and allicin content in Garlic (Allium sativum L.) from China. Euphytica 198, 243–254. https://doi.org/10.1007/s10681-014-1097-1 (2014).
Panthee, D. R. et al. Diversity analysis of Garlic (Allium sativum L.) germplasms available in Nepal based on morphological characters. Gen. Resour. Crop Evol. 53, 205–212. https://doi.org/10.1007/s10722-004-6690-z (2006).
Sterling, S. J. & Eagling, D. R. Agronomics and allicin yield of Australian grown Garlic (Allium sativum). Acta Hortic. 555, 63–73. https://doi.org/10.17660/ActaHortic.2001.555.6 (2001).
Akbarpour, A., Kavoosi, B., Hosseinifarahi, M., Tahmasebi, S. & Gholipour, S. Evaluation of yield and phytochemical content of different Iranian Garlic (Allium sativum L.) ecotypes. Int. J. Hortic. Sci. 8 (4), 385–400. https://doi.org/10.22059/ijhst.2020.303657.373 (2021).
González, R. E., Soto, V. N. C., Sance, M. A. M., Camargo, A. B. & Galmarini, C. R. Variability of solids, organosulfur compounds, pungency and health-enhancing traits in Garlic (Allium sativum L.) cultivars belonging to different ecophysiological groups. J. Agri Food Chem. 57 (21), 10282–10288. https://doi.org/10.1021/jf9018189 (2009).
Soto Vargas, V. C., González, R., Sance, M., Burba, J. & Camargo, A. Genotype-environment interaction on the expression of allicin and pyruvic acid in Garlic (Allium sativum L). Rev. Fac. Cienc. Agrar. 42 (2), 15–22 (2010).
Tresina, P. S., Santhiya Selvam, M., Doss, A. & Mohan, V. R. Antidiabetic bioactive natural products from medicinal plants. In: Atta-ur Rahman (ed) Studies in Natural Products Chemistry. Science Direct 75 75–118. https://doi.org/10.1016/B978-0-323-91250-1.00004-5 (2022).
Yamaguchi, Y. & Kumagai, H. Characteristics, biosynthesis, decomposition, metabolism and functions of the Garlic odour precursor, S–allyl–L–cysteine sulfoxide (Review). Exp. Ther. Med. 19 (2), 1528–1535. https://doi.org/10.3892/etm.2019.8385 (2019).
Bhardwaj, N., Parashar, B., Kumar, A., Verma, D. & Jaslam, P. K. M. Principal component analysis for the assessment of genetic diversity. AIP Conf. Proc. 2357. https://doi.org/10.1063/5.0080566 (2022).
Eghlima, G., Esmaeili, H., Frzaneh, M. & Mirjalili, M. H. Multivariate analysis of Equisetum arvense L. ecotypes based on silicon content, phytochemical and morphological characterization. Silicon 16, 115–122. https://doi.org/10.1007/s12633-23-02660-8 (2024).
Eghlima, G., Mashhadi Tafreshi, Y., Aghamir, F., Ahadi, H. & Hatami, M. Regional environmental impacts on growth traits and phytochemical Profles of Glycyrrhiza glabra L. for enhanced medicinal and industrial use. BMC Plant Biol. 25, 116. https://doi.org/10.1186/s12870-025-06147-z (2025).
Ayed, C., Mezghani, N., Rhimi, A. & Dridi, B. A. L. M. Morphological evaluation of Tunisian Garlic (Allium sativum L.) landraces for growth and yield traits. JHPR 2 (1), 43–52. https://doi.org/10.22077/jhpr.2018.1838.1033 (2019).
Acknowledgements
This paper is a component of a research project designated by the approval number 0-63-33-123-991333, which was conducted with financial assistance from the Agricultural Research, Education, and Extension Organization (AREEO).
Funding
The present work financially was supported by the Agricultural Research, Education and Extension Organization (Areeo).
Author information
Authors and Affiliations
Contributions
MA, MHA and BD: Designed and performed the experiments; MA, MHA and BD: Recorded the data; MHA: Analyzed the data; MA: Wrote the paper, read and approved the manuscript; FB: Labored in the quantification of allicin.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Azimi, M., Alemkhoumaram, M.H., Damankeshan, B. et al. Adaptability of parental populations and selected garlic (Allium sativum L.) clones for introducing promising clones in Iran. Sci Rep 15, 34950 (2025). https://doi.org/10.1038/s41598-025-18841-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-18841-x
