
Abstract
Here we use magnetic resonance imaging to study the impact of marathon running on brain structure in humans. We show that the signal for myelin water fraction—a surrogate of myelin content—is substantially reduced upon marathon running in specific brain regions involved in motor coordination and sensory and emotional integration, but recovers within two months. These findings suggest that brain myelin content is temporarily and reversibly diminished by severe exercise, a finding consistent with recent evidence from rodent studies that suggest that myelin lipids may act as glial energy reserves in extreme metabolic conditions.
Similar content being viewed by others


Main
Prolonged endurance exercise mobilizes energy stores throughout the body to meet energy demands. Marathon runners primarily rely on carbohydrates as the main energy source during a race1. Once glycogen becomes low in muscle, liver and other organs, including brain, marathon runners utilize fat as an energy source1,2,3,4. Fat is more abundant in the body than carbohydrates, and it can provide a sustained source of energy for prolonged endurance exercise.
Myelin surrounds and enwraps axons in the central and peripheral nervous systems, providing electrical insulation and metabolic support to axons. The primary components of myelin are lipids, which make up 70–80% of myelin, whereas myelin proteins compact and stabilize its multilayered structure. While oligodendroglial fatty acid metabolism has been proposed to serve as energy reserves during glucose deprivation5, we further hypothesize that myelin lipids may contribute to brain activity as does body fat to fuel muscle.
To test our hypothesis we evaluated brain myelin content by multicomponent relaxometry magnetic resonance imaging (MRI) in city and mountain marathon runners. To that end, we acquired multi-echo T2-weighted MRI sequences post-processed with the DECAES algorithm for multicomponent relaxometry analysis6, thus providing three-dimensional (3D) parametric maps of the myelin water fraction (MWF), a non-invasive surrogate imaging biomarker of myelin6,7,8. MWF measures pools of water molecules within myelin lamellae as a proxy, faithfully reflecting histological staining for myelin, and has high sensitivity to detect subtle variations in the myelin content9. In this manner, we estimated water trapped between the lipid bilayers of the myelin sheath around axons and excluded luminal water that accounts for intra- and extracellular water.
MRI-based MWF maps were constructed from n = 10 subjects within 48 h before running a marathon (Extended Data Fig. 1). All individuals showed a similar distribution of MWF across the brain, with naturally occurring, slight, interindividual variabilities. Maps of the luminal water fraction (LWF), myelin fraction T2 relaxation times and luminal fraction T2 relaxation times were also obtained in the fitting process. High-resolution 3D T1-weighted MRI images of all subjects showed no significant interindividual variability for the different imaging sessions (Extended Data Fig. 2). The absence of large differences among subjects allowed us to build up averaged MWF maps of the brain (n = 10; value ranges between 0 and 0.25). These value ranges are in line with MWF values found in healthy subjects over a wide range of ages (Extended Data Fig. 3)10,11.
Similarly, averaged maps of MWF values were constructed for the second (n = 10 individuals), third (n = 2 individuals) and fourth (n = 6 individuals) imaging sessions, corresponding to post-run (24–48 h after the marathon), two weeks and two months (recovery) after the marathon, respectively. Coronal sections integrating imaging maps from a representative individual show a clear post-run reduction in MWF in motor descending pathways (Fig. 1, top).

Top, slab volumes (average intensity projection) of MWF maps from five consecutive coronal slices centred in motor descending pathways from an individual before the marathon (pre-run), two days after the marathon (post-run, 2 d) and two weeks after the marathon (post-run, 2 w), as well as after two months (2 m) of recovery. Bottom, histogram with mean ± s.e.m. values from all subjects (n = 10 for pre- and post-run, n = 6 for recovery) showing significant MWF reduction in white and grey matter areas 1–2 days after the marathon, and subsequent recovery to pre-run levels during two months of recovery. Individual data points for each histogram bar (n = 20 for pre- and post-run, n = 12 for recovery) originated from the two hemispheres in each subject. Notice that values return to pre-run levels in all areas. *P < 0.05 and ***P < 0.005, one-tailed paired Student’s t-test. See Extended Data Fig. 4 for exact P values and its Source Data for individual data points.
Next, to obtain a comprehensive view of MWF changes, we analysed its values after segmentation of the entire brain into different regions with the help of an atlas-aided 3D morphometric analysis. Thus, we divided the brain into 50 white matter regions using the JHU brain atlas of white matter tracks12 and 56 grey matter regions using the LPBA40 collection of the SRI24 brain atlas13. Mean MWF values were recorded for these 106 brain regions for each subject at each imaging session (https://doi.org/10.5281/zenodo.14726926 (ref. 14) and Fig. 2).

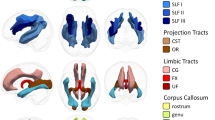
White matter areas that show a significant reduction in the MWF post-run (n = 10). Segmented white matter regions are displayed using the JHU atlas (https://neurovault.org/collections/264/ collection:264).
We focused on the regions with higher MWF values, as variations in this parameter are reflected better in more densely myelinated regions15. We observed consistent and significant region-variable MWF reductions in 12 white matter areas (Fig. 1, bottom, and Extended Data Fig. 4). In addition, MWF was lower in a few grey matter regions (Extended Data Fig. 4). However, lower myelin content in grey matter limits resolution to monitor MWF and those changes are uncertain.
MWF can be regarded as a strong correlate of myelin content reduction15,16,17,18; nevertheless, the exact correlation factor is to be determined, and, therefore, MWF-based studies should be considered as a semiquantitative evaluation of myelin content (see ref. 15 for a comprehensive analysis of MWF as a myelin imaging biomarker). An estimate of MWF changes versus ex vivo histopathological myelin quantification suggests that the former are typically larger than the latter17, indicating that the reduction in myelin after marathon running should indeed be lower (approximately 40–50%) than the measured decrease in MWF.
Bilateral MWF loss in white matter was extensive after completion of the marathon, and its extent was similar in both hemispheres (Fig. 2). MWF was lower in the corticospinal tract, pontine crossing tract, cerebral and all three cerebellar peduncles, as well in the anterior and posterior corona radiata (for a complete list see Extended Data Fig. 4). Quantification of the changes illustrates that the MWF signal decreases significantly by up to 28% and 26% in the pontine crossing and corticospinal tracts, respectively (Figs. 1 and 2 and Extended Data Fig. 4). Thus, MWF robustly diminished in axonal tracts involving motor function and coordination along with sensory and emotional integration19,20.
As MWF represents water trapped between myelin lamellae, a confounding factor of these changes could be dehydration. However, global dehydration would not change MWF by definition (for details see Extended Data Fig. 1). To rule out partial, regional dehydration, we measured total and partial central nervous system volumes. We found that the total brain, cerebrospinal fluid (CSF) (ventricles), grey matter, white matter, deep brain, brainstem and cerebellum volumes did not significantly change intra-individually across imaging sessions. Thus, P values for comparisons of paired brain regions post-run versus pre-run scans indicate, in all instances, absences of significant differences in the volume of all compared structures (Extended Data Fig. 5). These data are in line with previous studies showing that extensive exercise does not alter brain volumes independently of the hydration state, as measured by MRI21.
These observations are also consistent with previous findings showing that the level of hydration does not change MWF levels22. In turn, brain water volumes remain unaltered after extensive exercise21. Furthermore, MWF loss was not observed along the entire brain volume, as it would be as a consequence of dehydration, remaining unaltered in some axonal tracts (for example, corpus callosum), as well as in nearly all grey matter areas. Yet, and to exclude the possibility of selective dehydration in heavily myelinated territories, we calculated regional water content levels throughout the entire brain utilizing the grey and white matter atlases used to analyse MWFs. We found no significant differences in water content between pre- and post-run values in those areas showing significant reduction in MWF (Extended Data Figs. 6–8).
Collectively, these data provide solid evidence of bilateral MWF decrease after marathon running in motor- and non-motor-related brain connections, with a similar amplitude in each hemisphere. Notwithstanding, it is important to emphasize that the overall MWF reduction in the whole brain is limited and occurs in restricted areas, thus leaving the bulk of myelin unaffected.
We next examined whether MWF levels recover at rest after completion of the endurance effort. To that end, we performed MRI scans on runners (n = 2) two weeks after the run and found that, although there was a substantial partial increase of MWF values they did not reach pre-run levels (Fig. 1). These findings prompted us to measure MWF maps at later stages. We found that MWF values fully recovered from completion of the marathon two months after the run in all areas with MWF reduction (Fig. 1 and Extended Data Figs. 4 and 9; https://doi.org/10.5281/zenodo.14726926 (ref. 14). Thus, MWF status goes back to normal in myelinated tracts involved in motor and non-motor functions after two months of strenuous exercise.
Together, these findings provide compelling evidence that white matter tracts undergo a reduction in MWF that is later restored after recovery from prolonged endurance exercise. It would therefore be important to evaluate whether these changes transiently affect neurophysiological and cognitive functions associated with those regions.
Our findings show that running a marathon reduces runners’ MWF levels in white matter areas, with a similar impact in both hemispheres. MWF values recover thereafter and reach pre-run values two months after the event. This reversible reduction in MWF upon prolonged exercise and recovery after lowering physical activity strongly suggests changes in myelin structure and content that may open up a new view of myelin as an energy store ready to use when common brain nutrients are in short supply. We define this process as metabolic myelin plasticity.
To assess myelin content we used MWF, an MRI imaging surrogate of myelin that faithfully represents its levels with a near linear relationship with ex vivo histopathological myelin quantification15,16,17,18. Accordingly, MWF values correlate with changes in myelin content, although they represent semiquantitative measurements of myelin. This limits an accurate account of the actual extent of myelin content changes. In turn, measured MWF may vary for reasons other than changes in myelin content, including increases in permeability of myelin membranes, alterations in iron concentration or axonal swelling. However, the impact of these features on MWF reduction in marathon runners is unlikely (see ‘MWF confounding factors’ and references therein).
Hitherto, changes in MWF have not been evaluated following strenuous, prolonged exercise. However, ultramarathon multistage runners may experience subtle reversible brain structural changes without neurological complications23,24,25,26,27,28 along with increasing activity29 because of energy deficits. This might result in usage of myelin-derived fatty acids for energy production. This idea is plausible, as glial cells can fuel neural metabolism by β-oxidation, and myelin peroxisomes are essential for white matter maintenance in flies and mice5,30,31,32. Thus, oligodendrocytes may survive glucose deprivation by local utilization of myelin lipids, which in turn allows oligodendrocytes to share energy substrates with axons to support their function5. Indeed, myelin-derived fatty acids can be β-oxidized and transferred into mitochondria for ATP production by CPT1A transport5,33,34. The amphipathic nature of these fatty acids makes them easily transferable to axons as well as to the surrounding glia and neurons and to support energy demands at various subcellular compartments. Thus, myelin turnover under low glucose conditions can transiently support axonal energy metabolism, a feature that may explain the gradual loss of myelin in neurodegenerative diseases with underlying hypometabolism35.
Oligodendrocytes are particularly resistant to glucose withdrawal36,37, and therefore use alternative mitochondria-dependent energy sources that may be relevant in pathology. Thus, continuous myelin turnover and fatty acid metabolism can help myelinated nerves to overcome a transient loss of glucose supply, thereby providing an energy reserve5. Indeed, ongoing inflammatory activity in multiple sclerosis lesions may compromise the energy support for (re)myelinating cells, while neuropathological findings in this demyelinating disease probably reflect defective oligodendroglial energy homeostasis38.
Rather limited reduction in MWF, and presumably myelin, after completion of the marathon makes it unlikely that consumed myelin contributes to whole-body energy balance. However, we cannot rule out the possibility that it does in even more extreme exercise conditions and/or disease states, where nutrition is at stake (for example, anorexia nervosa and famine). Indeed, animal and human studies show that undernutrition negatively influences myelination39,40,41,42. In particular, dietary scarcity in anorexia nervosa leads to disturbances in cognition43, paralleled with altered grey and white matter myelin and connectivity44,45,46,47,48,49.
Regular, moderate-intensity, physical activity is a key factor in maintaining brain health across the lifespan50. By contrast, strenuous physical activity is a risk factor for amyotrophic lateral sclerosis (ALS) in individuals with a genetic predisposition to the disease51,52. Therefore, endurance exercise may pose an additional danger to heavily myelinated disease-vulnerable motor areas (for example, corticospinal tract) in individuals with ALS genetic risk, as myelin itself and myelin-producing oligodendrocytes are also vulnerable to glutamate excitotoxicity53.
Myelin plasticity is fundamental to brain adaptation to neuronal activity, as it modifies myelin structure by increasing or decreasing the thickness of the myelin sheath54,55. Our findings strongly suggest that widespread reversible MWF reduction in endurance physical exercise represents a new form of plasticity to support brain function at the expense of myelin lipid use.
This pilot observational imaging study has limitations, including a relatively small number of participants that will require validation in a larger cohort, difficulty in assessing grey matter MWF changes due to relatively low myelin content that compromises the signal-to-noise ratio and limited spatial resolution due to long scanning times and the inherent slight movement of subjects. Nonetheless, the potential relevance of the findings described here to brain energy metabolism warrant additional studies.
Methods
Subjects
Runners were recruited at the Donostia city marathon 2022 (n = 2) and 2023 (n = 2), the Zegama-Aizkorri mountain marathon 2023 (n = 2), Hiru Handiak mountain marathon 2023 (n = 2) and Valencia city marathon (n = 2). No financial compensation was given to volunteers. All of them were well-trained men (n = 8) and woman (n = 2) aged 45–73-years-old who completed the trial in a healthy state. All subjects provided written informed consent before participation, including consent to share the information provided in the study, and the experiments were conducted in accordance with the Helsinki Declaration (2001). The procedure was approved by Comité de Etica de Investigación de Euskadi. Imaging sessions were carried out 24–48 hours before and after the marathon (n = 10), two weeks (n = 2) and two months later (n = 6) (Extended Data Fig. 10). Randomization was not necessary as we studied a single group of subjects. In all instances, individuals were well hydrated.
Data acquisition
MRI scans were acquired on a 3T whole-body MRI system (Achieva Dstream, Philips Medical System) using the internal quadrature body coil for transmission and a 32-channel phased array coil for reception. Each subject underwent an imaging protocol that included: (1) a multi-echo 3D gradient and spin–echo sequence (GRASE) for myelin water imaging, with the following parameters: TR = 2,000 ms; 32 echoes with a minimum echo time of 9.3 ms and maximum of 298 ms; SENSE 2.5; flip angle 90°, bandwidth in EPI frequency direction 2,591; field of view (FOV) 230 mm2; 78 slices in transverse orientation; voxel size 1.2 × 1.2 × 1.8 mm3; total scan time 7:08 min; and (2) a high-resolution T1w anatomical image in a sagittal orientation with the following parameters: TR = 7.4 ms; TE = 3.4 ms; matrix size 228 × 228; flip angle 9°; FOV 250 × 250 × 180 mm; slice thickness 1.1 m; 300 slices; acquisition time 4:55 min.
Image processing
Image processing and data analysis were performed blinded to group allocation during data collection. Raw data were exported in DICOM format for offline processing. First, data were converted into NIFTI format using dcm2niix56. Anatomical brain images (skull stripped out) were obtained from T1-weighted images applying BET57 implementation from the FSL v.6.0 distribution58. Population distributions of T2 values were computed voxelwise from multi-echo data using Julia implementation DECAES software6. For the analysis of interindividual and intra-individual changes in MWF maps, all MWF maps for subjects at different imaging sessions (pre-run, post-run and recovery) were registered to a T2-weighted anatomical image of the brain of subject 1 (echo 8 of the echo train, with TE = 74.48 ms) at session 1 using ANT Syn registration59. For image segmentation and assessment of MWF values in different regions of the brain, T1-weighted images were nonlinearly registered to the JHU DTI-based white matter atlas for white matter track segmentation12 and in parallel to the LPBA40 collection of the SRI24 brain atlas13 for segmentation of grey matter regions. Transformation matrices were later applied to the MWF maps, previously linearly registered to the corresponding T1W image. Further image processing for the creation of figures was performed with the freely available software FIJI (ImageJ v.2.0) and ITK-Snap v.4.0.1. Individual MWF data are provided in https://doi.org/10.5281/zenodo.14726926 (ref. 14).
Statistics
We used Microsoft Excel (Office 365, 2024 version) for statistical analyses, and values were calculated as mean ± standard deviation of the mean (s.d.) or standard error of the mean (s.e.m.). Statistical significance was defined as *P < 0.05, **P < 0.01, ***P < 0.005 (or as otherwise indicated in figure captions). No statistical methods were used to predetermine sample sizes but our sample sizes are similar to those reported in previous publications60. Data distribution was assumed to be normal but this was not formally tested. Post-run versus pre-run groups, or recovery versus post-run groups, were compared using a one-tailed paired Student’s t-test. Each participant was analysed separately and measurements across imaging sessions normalized to pre-run data. We made all statistical analysis and compared subjects post versus pre (n = 10), and recovery versus pre (2 months, n = 6). For the comparison of the different brain regions, we used MWF mean values obtained for both left and right hemispheres. For the analysis of percentage of change in MWF values, pretreatment of the data was performed to eliminate outliers using the interquartile (IQR) method61, discarding all data below Q1 – 1.5 × IQR or over Q3 + 1.5 × IQR limits.
Technical considerations and limitations
Multi-echo T2 sequences allow differentiation between water with different T2 relaxation times
Water in the brain is located in the CSF, extracellular space, intracellular space and between myelin layers. Approximately 70–85% of the brain mass is water, and approximately 40% of the mass in myelinated axonal tracts is compartmentalized water.
In human white matter, the MRI signal from water can be separated into three pools on the basis of the T2 relaxation time. The longest T2 component is due to CSF. The intermediate component arises from both intracellular and extracellular water (LWF), and the shortest T2 component is due to water trapped between myelin bilayers (or MWF). CSF is often neglected in myelin imaging with MRI because of the low amount of CSF in the white matter62. MWF is defined as the ratio of the area of short T2 components to the area of all T2 components7,16. The ranges of T2 relaxation time used for the assignment of MWF and LWF where 0–30 ms and 30–200 ms, respectively, in line with current standards7. This fully distinguished the myelin-bound and luminal populations.
T2 mapping is a well-established method for myelin quantification. Originally, a T2 multi-echo spin–echo sequence was developed, but the main drawback was the long scanning time, which prevented its application in clinical scenarios. In recent years, together with other methods, a combined GRASE sequence has been adopted for myelin water imaging (MWI), reducing the acquisition time to less than 10 min with full brain coverage63. Reproducibility of MWI with this technique has been demonstrated in a multisite, multivendor study64 and with different reconstruction methods65.
However, MWI with T2 mapping is not without challenges, and several factors need to be considered. The signal-to-noise ratio in MWI with T2 mapping is low. The myelin signal decays rapidly as the MWF is approximately 10%, and the relaxation time of myelin water is relatively short at 10 ms, which makes changes in MWF difficult to detect66.
MWF confounding factors
A possible confounding effect on assimilating MWF to myelin content is brain oedema, which may occur in strenuous exercise, such as high mountain ultramarathons, but has not been observed in 42.195-km marathons25,67. Moreover, oedema/inflammation during new lesion formation in multiple sclerosis has a minor impact, if any, on MWF60. These previous reports indicate that even if minor oedema appears in a standard marathon run, the impact on MWF would be marginal.
Another confounding factor can be iron, the main source of paramagnetic susceptibility in the brain68. Iron shortens T2 components and thus potentially increases MWF values69. Thus, total iron depletion in experimental animals reduces MWF values by one-fourth69. However, serum iron levels increase or remain similar after ultramarathon and marathon running, respectively70,71. Therefore, it is unlikely that changes in iron homeostasis contribute significantly to MWF reduction after prolonged endurance exercise, and, if so, it may underestimate real myelin content reduction.
Lastly, fibre orientation may also affect myelin water T2, as well as the intracellular and extracellular water T2; however, MWF has a weaker innate orientation dependence9. Moreover, MWF value comparisons within and among subjects at all stages were made at similar MRI plane levels and orientations.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Numerical data and materials used in this work are available via Zenodo at https://doi.org/10.5281/zenodo.14726926 (ref. 14). Images from subjects are available upon request (contact: Pedro Ramos-Cabrer, pramos@cicbiomagune.es). Source data are provided with this paper.
Code availability
Code used in this work is available via Zenodo at https://doi.org/10.5281/zenodo.14726926 (ref. 14).
References
Burke, L. M. & Hawley, J. A. Swifter, higher, stronger: what’s on the menu? Science 362, 781–787 (2018).
Alghannam, A. F., Ghaith, M. M. & Alhussain, M. H. Regulation of energy substrate metabolism in endurance exercise. Int. J. Environ. Res. Public Health 18, 4963 (2021).
Matsui, T. et al. Astrocytic glycogen-derived lactate fuels the brain during exhaustive exercise to maintain endurance capacity. Proc. Natl Acad. Sci. USA 114, 6358–6363 (2017).
Magistretti, P. J. & Allaman, I. Acellular perspective on brain energy metabolism and functional imaging. Neuron 86, 883–901 (2015).
Asadollahi, E. et al. Oligodendroglial fatty acid metabolism as a central nervous system energy reserve. Nat. Neurosci. 27, 1934–1944 (2024).
Doucette, J., Kames, C. & Rauscher, A. DECAES—decomposition and component analysis of exponential signals. Z. Med. Phys. 30, 271–278 (2020).
MacKay, A. L. & Laule, C. Magnetic resonance of myelin water: an in vivo marker for myelin. Brain Plast. 2, 71–91 (2016).
Caverzasi, E. et al. MWF of the corpus callosum is a robust measure of remyelination: results from the ReBUILD trial. Proc. Natl Acad. Sci. USA 120, e2217635120 (2023).
Morris, S. R. et al. Myelin biomarkers in the healthy adult brain: correlation, reproducibility, and the effect of fiber orientation. Magn. Reson. Med. 89, 1809–1824 (2023).
Dvorak, A. V. et al. An atlas for human brain myelin content throughout the adult life span. Sci. Rep. 11, 269 (2021).
Kiely, M. et al. Insights into human cerebral white matter maturation and degeneration across the adult lifespan. NeuroImage 15, 118727 (2022).
Mori, S., Wakana, S., van Zijl, P. C. M. & Nagae-Poetscher, L. M. MRI Atlas of Human White Matter (Elsevier, 2005).
Rohlfing, T., Zahr, N. M., Sullivan, E. V. & Pfefferbaum, A. The SRI24 multichannel atlas of normal adult human brain structure. Hum. Brain Mapp. 31, 798–819 (2010).
Ramos-Cabrer, P. et al. Source data and code. Zenodo https://doi.org/10.5281/zenodo.14726926 (2025).
Laule, C. et al. Magnetic resonance imaging of myelin. Neurotherapeutics 4, 460–484 (2007).
Mackay, A. et al. In vivo visualization of myelin water in brain by magnetic resonance. Magn. Reson. Med. 31, 673–677 (1994).
Laule, C. et al. Myelin water imaging in multiple sclerosis: quantitative correlations with histopathology. Mult. Scler. 12, 747–753 (2006).
Laule, C. et al. Myelin water imaging of multiple sclerosis at 7 T: correlations with histopathology. NeuroImage 40, 1575–1580 (2008).
Bubb, E. J., Nelson, A. J. D., Cozens, T. C. & Aggleton, J. P. Organisation of cingulum bundle fibres connecting the anterior thalamic nuclei with the rodent anterior cingulate and retrosplenial cortices. Brain Neurosci. Adv. 4, 2398212820957160 (2020).
Verhulst, M. M. L. H., Glimmerveen, A. B., van Heugten, C. M., Helmich, R. C. G. & Hofmeijer, J. MRI factors associated with cognitive functioning after acute onset brain injury: Systematic review and meta-analysis. NeuroImage Clin. 38, 103415 (2023).
Dieleman, N., Koek, H. L. & Hendrikse, J. Short-term mechanisms influencing volumetric brain dynamics. NeuroImage Clin. 16, 507–513 (2017).
Meyers, S. M. et al. Does hydration status affect MRI measures of brain volume or water content? J. Magn. Reson. Imaging 44, 296–304 (2016).
Gliner, J. A., Matsen-Twisdale, J. A., Horvath, S. M. & Maron, M. B. Visual evoked potentials and signal detection following a marathon race. Med. Sci. Sports 11, 155–159 (1979).
Freund, W. et al. Substantial and reversible brain gray matter reduction but no acute brain lesions in ultramarathon runners: experience from the TransEurope-FootRace Project. BMC Med. 10, 170 (2012).
Herm, J. et al. MRI brain changes after marathon running: results of the Berlin Beat of Running study. Int. J. Sports Med. 40, 856–862 (2019).
Ozdurak Singin, R. H., Duz, S. & Kiraz, M. Cortical and subcortical brain volume alterations following endurance running at 38.6 km and 119.2 km in male athletes. Med. Sci. Monit. 27, e926060 (2021).
Bailey, S. P., Hall, E. E., Folger, S. E. & Miller, P. C. Changes in EEG during graded exercise on a recumbent cycle ergometer. J. Sports Sci. Med. 7, 505–511 (2008).
Brümmer, V., Schneider, S., Strüder, H. K. & Askew, C. D. Primary motor cortex activity is elevated with incremental exercise intensity. Neuroscience 181, 150–162 (2011).
Saab, A. S. et al. Oligodendroglial NMDA receptors regulate glucose import and axonal energy metabolism. Neuron 91, 119–132 (2016).
Kassmann, C. M. Myelin peroxisomes – essential organelles for the maintenance of white matter in the nervous system. Biochimie 98, 111–118 (2014).
McMullen, E. et al. Glycolytically impaired Drosophila glial cells fuel neural metabolism via β-oxidation. Nat. Commun. 14, 2996 (2023).
Kassmann, C. M. et al. Axonal loss and neuroinflammation caused by peroxisome-deficient oligodendrocytes. Nat. Genet. 39, 969–976 (2007).
Bolaños, J. P. Bioenergetics and redox adaptations of astrocytes to neuronal activity. J. Neurochem. 139, 115–125 (2016).
Morant-Ferrando, B. et al. Fatty acid oxidation organizes mitochondrial supercomplexes to sustain astrocytic ROS and cognition. Nat. Metab. 5, 1290–1302 (2023).
Zilberter, Y., Tabuena, D. R. & Zilberter, M. NOX-induced oxidative stress is a primary trigger of major neurodegenerative disorders. Prog. Neurobiol. 231, 102539 (2023).
Narine, M. & Colognato, H. Current insights into oligodendrocyte metabolism and its power to sculpt the myelin landscape. Front. Cell. Neurosci. 16, 892968 (2022). 28.
Fern, R., Davis, P., Waxman, S. G. & Ransom, B. R. Axon conduction and survival in CNS white matter during energy deprivation: a developmental study. J Neurophysiol. N79N, 95–105 (1998).
Tepavčević, V. Oligodendroglial energy metabolism and (re)myelination. Life 11, 238 (2021).
Laczkovics, C. et al. White matter integrity is disrupted in adolescents with acute anorexia nervosa: a diffusion tensor imaging study. Psychiatry Res. Neuroimaging 320, 111427 (2022).
Krigman, M. R. & Hogan, E. L. Undernutrition in the developing rat: effect upon myelination. Brain Res. 107, 239–255 (1976).
Wiggins, R. C. & Fuller, G. N. Relative synthesis of myelin in different brain regions of postnatally undernourished rats. Brain Res. 162, 103–112 (1979).
Connor, J. R. & Menzies, S. L. Relationship of iron to oligodendrocytes and myelination. Glia 17, 83–93 (1996).
Diagnostic and Statistical Manual of Mental Disorders (DSM-5) (American Psychiatric Association, 2013).
Katzman, D. K. et al. Cerebral gray matter and white matter volume deficits in adolescent girls with anorexia nervosa. J. Pediatr. 129, 794–803 (1996).
Boto, J. et al. Cerebral gray and white matter involvement in anorexia nervosa evaluated by T1, T2, and T2* mapping. J. Neuroimaging 29, 598–604 (2019).
Gaudio, S., Carducci, F., Piervincenzi, C., Olivo, G. & Schiöth, H. B. Altered thalamo–cortical and occipital–parietal–temporal–frontal white matter connections in patients with anorexia and bulimia nervosa: a systematic review of diffusion tensor imaging studies. J. Psych. Neurosci. 44, 324 (2019).
Lloyd, E. C. et al. Large-scale exploration of whole-brain structural connectivity in anorexia nervosa: alterations in the connectivity of frontal and subcortical networks. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 8, 864–873 (2023).
Pappaianni, E. et al. Initial evidence of abnormal brain plasticity in anorexia nervosa: an ultra-high field study. Sci. Rep. 12, 2589 (2022).
Watson, H. J. et al. Genome-wide association study identifies eight risk loci and implicates metabo-psychiatric origins for anorexia nervosa. Nat. Genet. 51, 1207–1214 (2019).
Kirkinezos, I. G., Hernandez, D., Bradley, W. G. & Moraes, C. T. Regular exercise is beneficial to a mouse model of amyotrophic lateral sclerosis. Ann. Neurol. 53, 804–807 (2003).
Chapman, L., Cooper-Knock, J. & Shaw, P. J. Physical activity as an exogenous risk factor for amyotrophic lateral sclerosis: a review of the evidence. Brain 146, 1745–1757 (2023).
Zheng, X. et al. Physical activity as risk factor in amyotrophic lateral sclerosis: a systematic review and meta-analysis. J. Neurol. 270, 2438–2450 (2023).
Matute, C., Sánchez-Gómez, M. V., Martínez-Millán, L. & Miledi, R. Glutamate receptor-mediated toxicity in optic nerve oligodendrocytes. Proc. Natl Acad. Sci. USA 94, 8830–8835 (1997).
Fields, R. D. & Dutta, D. J. Treadmilling model for plasticity of the myelin sheath. Trends Neurosci. 42, 443–447 (2019).
Knowles, J. K., Batra, A., Xu, H. & Monje, M. Adaptive and maladaptive myelination in health and disease. Nat. Rev. Neurol. 18, 735–746 (2022).
Li, X., Morgan, P. S., Ashburner, J., Smith, J. & Rorden, C. The first step for neuroimaging data analysis: DICOM to NIfTI conversion. J. Neurosci. Methods 264, 47–56 (2016).
Smith, S. M. Fast robust automated brain extraction. Hum. Brain Mapp. 17, 143–155 (2002).
Jenkinson, M., Beckmann, C. F., Behrens, T. E., Woolrich, M. W. & Smith, S. M. FSL. NeuroImage 62, 782–790 (2012).
Avants, B. B., Epstein, C. L., Grossman, M. & Gee, J. C. Symmetric diffeomorphic image registration with cross-correlation: evaluating automated labeling of elderly and neurodegenerative brain. Med. Image Anal. 12, 26–41 (2008).
Vavasour, I. M. et al. Water content changes in new multiple sclerosis lesions have a minimal effect on the determination of myelin water fraction values. J. Neuroimaging 31, 1119–1125 (2021).
Smiti, A. A critical overview of outlier detection methods. Comp. Sci. Rev. 38, 100306 (2020).
Henkelman, R. M., Stanisz, G. J. & Graham, S. J. Magnetization transfer in MRI: a review. NMR Biomed. 14, 57–64 (2001).
Prasloski, T. et al. Rapid whole cerebrum myelin water imaging using a 3D GRASE sequence. NeuroImage 63, 533–599 (2012).
Lee, L. E. et al. Inter-vendor reproducibility of myelin water imaging using a 3D gradient and spin echo sequence. Front. Neurosci. 12, 854 (2018).
Fischi-Gomez, E. et al. Variability and reproducibility of multi-echo T2 relaxometry: insights from multi-site, multi-session and multi-subject MRI acquisitions. Front. Radiol. 2, 930666 (2022).
van der Weijden, C. W. J. et al. Quantitative myelin imaging with MRI and PET: an overview of techniques and their validation status. Brain 146, 1243–1266 (2023).
Zanchi, D. et al. Extreme mountain ultra-marathon leads to acute but transient increase in cerebral water diffusivity and plasma biomarkers levels changes. Front. Physiol. 7, 664 (2017).
Deistung, A., Schweser, F. & Reichenbach, J. R. Overview of quantitative susceptibility mapping. NMR Biomed. 30, e3569 (2017).
Birkl, C. et al. The influence of brain iron on myelin water imaging. NeuroImage 199, 545–552 (2019).
Kaufmann, C. C. et al. Effect of marathon and ultra-marathon on inflammation and iron homeostasis. Scand. J. Med. Sci. Sports 31, 542–552 (2021).
Kłapcińska, B. et al. Metabolic responses to a 48-h ultra-marathon run in middle-aged male amateur runners. Eur. J. Appl. Physiol. 113, 2781–2793 (2013).
Acknowledgements
We thank runner volunteers for their generosity and patience. A. Planas, J. J. Lucas, F. M. Goñi, F. Kirchhoff, G. Foffani, J. P. Bolaños, A. Volterra, M. M. Panicker and F. Pérez-Cerdá for insightful discussions and suggestions on the manuscript. We also thank Osatek-Galdakao for providing human imaging facilities for MRI scans. Funding: Spanish Ministry of Science and Innovation and Universities grants PID2022-143020OB-I00 (C.M.), PID2023-152005OB-I00 (P.R.-C.), Basque Government grant IT-1551-22 (C.M.) Basque Government grant KK-2021/0009 (P.R.-C.). CIBERNED Network, Instituto Carlos III, grant CB06/05/0076 (C.M.), Ikerbasque Foundation, the Ikerbasque Research Professors Program (P.R.-C.).
Author information
Authors and Affiliations
Contributions
Conceptualization: P.R.-C., C.M. Methodology: P.R.-C., A.C.-Z., D.P., M.M.-G., A.R.-A., C.M. Investigation: P.R.-C., A.C.-Z., D.P., M.M.-G., A.R.-A., C.M. Funding acquisition: P.R.-C., C.M. Supervision: P.R.-C., C.M. Writing—original draft: P.R.-C., C.M. Writing—review and editing: P.R.-C., A.C.-Z., D.P., M.M.-G., A.R.-A., C.M.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Metabolism thanks Alex MacKay and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Alfredo Giménez-Cassina, in collaboration with the Nature Metabolism team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Schematic representation of the DECAES procedure to extract myelin water fraction (MWF) values.
MRI T2 weighted multi-echo images were processed using decomposition and component analysis of exponential signals (DECAES). This procedure uses the relaxation curves in each pixel to deconvolve the signal from 3 different pools of water: 1) water protons in myelin sheaths (short T2 values), 2) inter- and intracellular water molecules (luminal water fraction, with intermediate T2 values), and 3) signal from CSF (unrestricted water pool, with very large T2 values). Thus, a set of two T2 maps (short T2 population and intermediate T2 population, the CSF contribution is generally obviated) are generated, along with two maps of water fraction (% of contribution of that fraction to the total signal), one reflecting MWF and the other reflecting the luminal water fraction (LWF). The approach used in this study computes MWF derived from the area of myelin protons without differentiating luminal protons and unrestricted water pools whose combined area is estimated, as established by Doucette et al.6, MacKay et al.7, Caverzasi et al.8 ms, millisecond.
Extended Data Fig. 2 High-resolution 3D T1-weighted MRI images of four representative subjects.
Sagittal, axial and coronal views of the 3D images acquired with T1 weighting for anatomical reference, pre- and post-run. Images show no significant interindividual differences for the different imaging sessions. Volumes of whole brain and structures was unaltered (see also Extended Data Fig. 5).
Extended Data Fig. 3 Myelin water fraction (MWF) parametric maps from representative subjects before the marathon challenge.
Top) Pre-exercise MWF maps for subjects scanned in this study. From top to bottom, arbitrarily chosen axial, coronal and sagittal planes. Bottom) Average maps obtained after ANTs Syn registration of all images to a common spatial frame.
Extended Data Fig. 4 Myelin water fraction changes (%) in brain regions post-run and after 2 months of recovery.
one-tail paired Student’s t-test.
Extended Data Fig. 5 Volumes of brain regions across subjects and pre-run and post-run MRI sessions.
Student’s t values were calculated using a one-tail paired t-test.
Extended Data Fig. 6 Regional S0 images from T2 fitting of multi-echo data maps are unaltered after marathon running.
To assess further the lack of post-run dehydration as a confounding factor in MWF values, we analyzed images as described below: 1) We carried out a bi-exponential decay analysis on acquired T2 multi-echo sets of images to obtain (a) 2 sets of T2 maps (low and high mobility protons) and (b) 2 sets of S0 images (Signal extrapolated at TE=0 ms). (c) Total S0 maps were obtained by adding the two individual S0 components. 2) CSF regions were segmented out, registering pre- (c) and post-run images (d). 3) Percentage of change of S0 images were determined pixel by pixel for each subject using the following image operation 100*(pre-post)/pre (e). Notice that maps reflect % changes. Subjects were scanned with identical acquisition sequence in the same scanner, with equal repetition time and flip angle. At fixed T1 weighting changes on water content should be reflected as change in S0 maps between pre- and post-run.
Extended Data Fig. 7 Regional S0 images from bi-exponential fitting of T2 multi-echo data maps of white matter regions.
Values (mean ± stdev) derived from S0 images obtained after bi-exponential fitting of T2 multi-echo data images for all subjects. White matter regions were mapped using atlas in Ref. 18. Comparisons were made using one-tail paired Student’s t-test.
Extended Data Fig. 8 Regional S0 images from bi-exponential fitting of T2 multi-echo data maps of gray matter regions.
Values (mean ± stdev) derived from S0 images obtained after bi-exponential fitting of T2 multi-echo data images for all subjects. Gray matter regions were mapped using atlas in Ref. 19. Comparisons were made using one-tail paired Student’s t-test.
Extended Data Fig. 9 Myelin water fraction (MWF) imaging from all subjects together showing post-run signal decay and recovery thereafter.
Left, Slab volumes (average intensity projection) of MWF maps from 5 consecutive coronal slices centered in motor descending pathways from all subjects (mean MWF of scanned individuals) at imaging sessions before the marathon (pre-exercise), after (post-exercise) (n=10) and at 2-months of recovery (n=6). Color code bar displays represented values. Notice that MWF values return to pre-run values at 2-months of recovery.
Extended Data Fig. 10 CONSORT diagram showing number of MRI scanned subjects at all stages.
Diagram illustrates the time points at which the runners were scanned. Imaging sessions were carried out 24–48 hours before (Pre-run) and after (Post-run) the marathon (n = 10), two weeks (n = 2) and two months later (n = 6).
Supplementary information
Supplementary Code
1. Code used in Python to calculate MWF values for each individual and each image session. 2. Code used in Python to calculate MWF values for each individual and each image session. 3. Code used for bi-exponential fit of T2 MRI data to calculate proton densities for each individual and each image session.
Source data
Source Data Fig. 1
Raw MWF data for all individuals and percentage changes from baseline per brain region. Data used to build bar plot of Fig. 1.
Source Data Fig. 2
Text file with label description of the JHU atlas for ITK-Snap.
Source Data Extended Data Fig. 4
Raw data for construction of Extended Data Fig. 4.
Source Data Extended Data Fig. 5
Raw data for construction of Extended Data Fig. 5.
Source Data Extended Data Fig. 7
Raw data for construction of Extended Data Fig. 7.
Source Data Extended Data Fig. 8
Raw data for construction of Extended Data Fig. 8.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ramos-Cabrer, P., Cabrera-Zubizarreta, A., Padro, D. et al. Reversible reduction in brain myelin content upon marathon running. Nat Metab 7, 697–703 (2025). https://doi.org/10.1038/s42255-025-01244-7
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s42255-025-01244-7
This article is cited by
-
Mild chronic hypoxia and the brain: an ambiguous relationship
Journal of Neuroinflammation (2026)
-
Integrated respiratory functions predict myelin status in the mouse brain
npj Biomedical Innovations (2026)
-
Aerobic glycolysis in Schizophrenia: Developmental rescue or energetic breakdown?
Molecular Psychiatry (2026)
-
Signaling roles for astrocytic lipid metabolism in brain function
EMBO Reports (2026)
-
Myelin Lipid Reserves as a Conditional Metabolic Buffer: Implications for Alzheimer’s Disease and Ischemic Stroke
NeuroMolecular Medicine (2026)
