
Abstract
Interleukin-9 (IL-9) is a multifunctional cytokine with roles in a broad cross-section of human diseases. Like many cytokines, IL-9 is transcriptionally regulated by a group of noncoding regulatory elements (REs) surrounding the IL9 gene. These REs modulate IL-9 transcription by forming 3D loops that recruit transcriptional machinery. IL-9-promoting transcription factors (TFs) can bind REs to increase locus accessibility and permit chromatin looping, or they can be recruited to already accessible chromatin to promote transcription. Ample mechanistic and genome-wide association studies implicate this interplay between IL-9-modulating TFs and IL9 cis-REs in human physiology, homeostasis, and disease.
Similar content being viewed by others


Introduction
Interleukin 9 (IL-9), first discovered in the 1980s, is a pleiotropic cytokine with roles in type 2 immunity, autoimmunity, antitumor immunity, and other cellular processes1. IL-9 has diverse sources that include CD4+ T cells, CD8+ T cells, innate lymphoid cells (ILCs), mast cells, and basophils1. While the sources and functions of IL-9 are tissue- and disease-specific, T helper 9 (Th9) cells are the best characterized and most abundant source of IL-9, particularly in humans1. Indeed, IL-9 is the hallmark effector cytokine of Th9 cells and defines them as a discrete T helper subset1.
Like many other cytokines, IL-9 is controlled by noncoding regulatory elements (REs) surrounding the IL9 gene2,3,4,5. These include the IL9 promoter and several enhancer elements. Enhancers modulate transcription by forming 3D interactions, or loops, that recruit transcriptional machinery5. Some transcription factors (TFs) can act as “pioneer” factors, binding to a promoter or enhancer to increase chromatin accessibility within a locus, permitting the formation of loops and recruitment of downstream TFs6. This interplay between accessibility, chromatin remodeling, looping, and TF recruitment regulates IL-9 production through the actions of several REs within the extended locus3,4,7,8.
Because IL-9 regulation is best characterized in CD4+ T cells, most epigenetic IL-9 studies have focused on Th9 cells. Relative to other T helper subsets, Th9 cells display several unique features associated with IL9 epigenetic regulatory mechanisms. Most strikingly, Th9 cells exhibit transcriptional and epigenetic instability, which may be a negative checkpoint on sustained STAT5- and STAT6-dependent bystander activation2,9. Stable circulating Th9 cells are not detected in healthy subjects but can be found in patients with inflammatory diseases, suggesting that IL9 epigenetic regulation is aberrant in these individuals. This aberrant regulation could be related to cell-intrinsic differences or downstream of microenvironmental cues specific to the inflammatory microenvironment that alter Il9/IL9 epigenetics in CD4+ T cells.
Here, we review fundamental mechanisms of epigenetic IL-9 regulation, which are largely derived from studies in murine models. We then analyze genome-wide association studies to extrapolate known links between IL-9 epigenetics and human disease. Finally, we review mechanistic evidence for IL-9 epigenetic regulatory mechanisms in disease pathogenesis, while describing ongoing and future areas of investigation in this field.
Structure of the extended IL9 locus and the function of key regulatory elements
The Il9/IL9 locus occupies an ~45-kb region comprising a promoter and several critical downstream and upstream cis-regulatory elements (REs) with enhancer activity3,4,7,8,10. One unique feature of the extended Il9/IL9 locus is its dynamic accessibility in Th9 cells, which is associated with Th9 lineage instability2. Like other cytokine loci, the Il9/IL9 locus becomes accessible during differentiation, with accessibility and TF binding beginning to increase within 12 hours of activation6. During differentiation, this programming of the Il9/IL9 locus depends on a complex network of TFs that includes several TCR-induced factors. Once the extended locus has become accessible, a smaller set of TFs is critical for acute activation of the locus. We and others have shown that STAT5 and STAT6 have a critical role downstream of IL-2 and IL-42,11. However, after removing TCR stimulation, accessibility and histone architecture change slowly over time, rendering the locus refractory to transcriptional activation. While the human IL9 locus is more stable than the murine Il9 locus, it also exhibits dynamic accessibility and TF binding2.
The murine Il9 locus
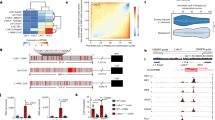
The murine Il9 gene is located on chromosome 13, with the extended Il9 locus characterized based on CTCF binding sites that mark the boundaries3,4,7,8. Within this locus, six putative cis- REs have been identified (Fig. 1a). The best characterized of these is the Il9 promoter (Il9p), also known as conserved noncoding sequence 1 (CNS1). Most Th9-promoting TFs have been shown to target the promoter directly. These include STAT5, STAT6, ETS family TFs (PU.1, ETV5, and ERG), SMAD2, SMAD3, TAK1, OX40, NF-κB, BATF, AP-1, GATA3, IRF1, IRF4, RARα, and DBP12. Moreover, many IL-9 enhancers act at least in part by forming 3D interactions, or loops, with the Il9 promoter4,7,8.

a Gene tracks show the extended murine Il9 locus, including the promoter and surrounding putative enhancers. b Gene tracks show the extended human IL9 locus, including the promoter and surrounding putative enhancers. For both human and murine loci, regulatory elements (REs) were identified based on accessibility (ATAC-seq, green), poised enhancer marks (H3K4M1 ChIP-seq, red), active promoter marks (H3K4M3 ChIP-seq, blue), and active enhancer marks (H3K27Ac ChIP-seq, purple). Human and murine orthologs are marked with black triangles. The data were obtained from GSE222910.
Downstream of the promoter, CNS + 5.5 (also called DS or SEa) is positioned ~5.4 kb from the transcriptional start site (TSS) (Fig. 1a)3,4,7,8. CNS + 5.5 promotes IL-9 production through OX40 signaling, primarily through RelB/p300-mediated chromatin acetylation. Deletion of CNS + 5.5 significantly reduces IL-9 production in OX40-stimulated murine Th9 cells, while loops between CNS1 (promoter) and CNS + 5.5 form in OX40-induced Th9 cells4.
CNS-6 (also referred to as E1 or SEb) is located -6 kb upstream of the Il9 TSS and binds STAT5, STAT6, and IRF4 to promote IL-9 production (Fig. 1a)7,13. Notably, IRF1 also binds to CNS-6, thereby displacing IRF4 to block activating histone modifications and recruit transcriptional repressors13. IRF1-mediated repression of IL-9 production occurs downstream of IFN-γ and is STAT1-dependent13. CNS-6 deletion reduces IL-9 production in cells differentiated under Th9-promoting conditions in vitro, although the in vivo functions of this cis-RE are not yet known7. Further upstream, CNS-20 (also E2) and CNS-25 (also E3 or SEc) are located 20 and 25 kb upstream of the Il9 TSS, respectively (Fig. 1a). CNS-20 is bound by STAT5 and STAT6, while CNS-25 is bound by many transcription factors, including BATF, IRF4, STAT5, STAT6, GATA3, and Foxo1. Deletion of CNS-20 and CNS-25 reduces Th9 differentiation; CNS-25 is also critical for IL-9 production by mast cells and basophils but does not affect IL-9 production in innate lymphoid cells (ILCs)3,7,8. Nonetheless, CNS-25-deficient mice are protected from chronic airway inflammation and anaphylaxis3,8. Importantly, looping between CNS-25 and the Il9 promoter is stronger in Th9 cells than in Th2 cells. Moreover, while CNS-25 is an Il9 enhancer in Th9 and Th17 cells, it acts as an epigenetic silencer in Th2 cells8. The mechanisms underlying this bifunctionality are unclear, but they could involve context-dependent binding of Th9-promoting TFs and Th2-promoting/Th9-repressing TFs.
The human IL9 locus
The human IL9 locus is located on chromosome 5 and has about 55% homology to the murine Il9 locus14. The extended locus contains six putative cis-REs (Fig. 1b). A downstream RE (CNS + 4.5, also known as DS) is located 4.5 kb upstream of the Transcription Start Site (TSS)2,13. In addition to the promoter (IL9p), there are four upstream elements2. CNS-4.5 (also known as E1) is located 4.5 kb upstream of the TSS and is homologous to murine CNS-6, while CNS-14 (also known as E2) is positioned 14 kb from the TSS and shares homology with murine E22. The upstream RE CNS-18 (also known as E3) is homologous to murine CNS-25. Deletion of CNS-18 in human Th9 cells reduced IL-9 expression, while the production of other cytokines was unaffected8. This included other type 2 cytokines (IL-4, IL-5, IL-13), although the genes encoding these cytokines are in a cluster approximately 3 mb downstream of the IL9 locus. Another previously uncharacterized element (CNS-20, also known as E4) is located 20 kb upstream of the TSS (Fig. 1b) and demonstrated Th9 cell-specific accessibility compared to naive cells and other subsets2. While Th9-specific chromatin architecture and homology to murine functional enhancers suggest that CNS + 4.5, CNS-4.5, CNS-14, and CNS-20 have critical regulatory roles, functional studies will be needed to address this question more definitively.
Interactions between IL-9-modulating transcription factors and the extended IL9 locus
One of the key mechanisms of enhancer-mediated gene regulation involves the recruitment of activating TFs15. TF binding to these noncoding DNA sequences results in formation of 3D loops and recruitment of transcriptional machinery. These mechanisms play a pivotal role in the control of lineage-specific gene expression in T cells15. A large network of IL-9-inducing TFs binds to the Il9/IL9 locus to modulate transcription; this includes engagement of key noncoding cis-REs.
STAT family
STATs are critical signal-dependent TFs that guide T cell development and subset commitment; STAT5 and STAT6 strongly induce Th9 differentiation downstream of IL-2 and IL-4, as well as promoting IL-9 production from other cell types like ILC2s6,7,10,15,16,17,18,19. STAT5 is also activated downstream of other Th9-promoting signals, including Itk, although this is also at least partly mediated by IL-218. STAT5 and STAT6 promote not only Th9 differentiation but also IL-9 production in committed Th9 cells, including innate-like bystander activation of recently activated resting human and mouse Th9 cells2,6. IL-2/STAT5 signaling also induces IL-9 in ILC2s, while STAT5 promotes IL-9 production downstream of TNF-α in CD8+ Tc9 cells19,20. Other STAT5-inducing cytokines like IL-7 and IL-15 have not been found to directly induce IL-9, although IL-7 can indirectly induce IL-9 in ILC2s by enhancing the effect of IL-3321.
STAT5 binds all IL9 REs, including the promoter and upstream/downstream enhancers (Fig. 2)6,7. Consecutively activated STAT5 stimulated >3-fold induction of the Il9 promoter element, >2-fold induction of CNS-6 and CNS-20, and <1.5-fold induction of CNS-25, establishing that these are STAT5-responsive REs6,7. In mast cells, STAT5 binds CNS-25 preferentially to the Il9 promoter, suggesting that STAT5-enhancer interactions may be different in various cell types3. Mechanistically, STAT5 promotes Il9 locus accessibility and modulates the binding and expression of other transcription factors. STAT5 acts as a “pioneer” TF that is required for other IL-9-inducing TFs like BATF to bind the Il9 locus (Fig. 3)6. STAT5 signaling also controls the balance between Th9 and Th17 differentiation. Blocking STAT5 promotes Th9 plasticity towards a Th17-like phenotype via STAT3-independent and RoRrץt- and BATF-dependent mechanisms10. STAT5 binding to the Il9 promoter also competes with the actions of the IL-9-repressing TF BCL622.

Schematic shows the 5 putative regulatory elements (REs) that promote Il9 transcription. Various transcription factors (TFs) that bind to Il9-promoting REs are shown, together with their reported RE associations. Il9-modulating TFs are reported to promote (green) or repress (red) IL-9 production, or to have context-dependent effects (yellow). Some IL-9-modulating TFs are not reported to directly target the Il9 locus; these TFs are schematically represented as proximal to, but not associated with, cis-REs.

IL-9-regulating transcription factors (TFs) are reported to regulate IL-9 expression via several mechanisms of action. These include direct binding, modulation of accessibility, histone modification, and induction or repression of other Il-9-modulating TFs. In this schematic, the curved arrows indicate that a TF directly targets—or binds to—the Il9 locus. Activating or inhibitory effects on other proteins are shown by straight arrows or lines, respectively.
STAT6 is activated downstream of IL-4 signaling and binds at REs throughout the Il9 locus, like STAT5 (Fig. 2)6,7. At the Il9 promoter, STAT6 inhibits the expression and binding of Th9-repressing transcription factors like T-bet and Foxp3 (Fig. 3)6,12,23. STAT6 is also required to induce BATF, which promotes Th9 development6,12,23. IRF4 directly promotes Th9- and ILC2-derived IL-9 production: STAT6 increases IRF4 expression and function (Fig. 3)23,24.
The effects of STAT1 and STAT3 on IL-9 are less straightforward and may be context- or species-dependent. In some murine models, STAT1 induces IRF1, which targets the Il9 promoter to induce IL-9 (Fig. 2)25. Conversely, IL-27/STAT1 signaling represses the differentiation of IL-9-producing T cells, while human STAT1 gain-of-function mutations repress IL-9 production via induction of T-bet12,26. STAT3 interferes with STAT5 activation to suppress murine Th9 differentiation; it is not clear whether STAT3 targets Il9 REs16. By contrast, STAT3 promotes the differentiation of IL-9+ human T cells26. The reasons for these species-specific effects are unclear but could involve repressing STAT1 to induce IL-926.
ETS family
The ETS (E26 transformation-specific) family includes three TFs that promote the differentiation of IL-9-producing CD4+ T cells: PU.1, ETV5 (ETS translocation variant 5), and ERG (Ets-related gene)12,27,28. PU.1 is the first TF found to directly interact with the Il9 promoter (Fig. 2) and was initially designated as a master TF for the Th9 lineage29. However, subsequent studies revealed that Th9 differentiation can proceed in the absence of PU.1, albeit at a much lower efficiency than in PU.1-sufficient cells27. PU.1 acts primarily during the early stage of Th9 differentiation; its expression decreases during the later stage of differentiation12. PU.1 binds the Il9 promoter and induces GCN5, a histone acetyltransferase (HAT) that acetylates key lysines on histones H3 and H4, thereby activating critical cis-REs (Fig. 2)30. PU.1 may also promote Th9 identity by suppressing genes essential for the development of other subsets, including Th2 and Tfh (T follicular helper) cells12.
ETV5 is another ETS family TF with a role in the differentiation of IL-9+ T cells: deletion of both PU.1 and ETV5 reduces IL-9 production more than individual deletion of each gene27. While Spi1 (encodes PU.1) expression peaks early during Th9 development, Etv5 expression increases downstream of IRF4 and STAT6 during the later stages of Th9 differentiation27. In addition to binding the Il9 promoter, ETV5 also targets CNS + 5.5 (DS) and CNS-6 (E1) and recruits p300 to activate transcription (Figs. 2 and 3)27.
A third member of the ETS family, ETS-related gene (ERG), also uses epigenetic mechanisms to induce IL-9 production and differentiation of IL-9-producing T cells28. ERG targets the Il9 promoter, CNS + 5.5 (DS) and CNS-25 (E3) in naive T cells; recruitment increases at CNS-6 (E1) during Th9 differentiation (Fig. 2). In late stages of differentiation (d5, mouse), ERG recruitment at the Il9 promoter is significantly higher in Th9 cells than in Th0 or Th2 cells, which do not produce substantial amounts of IL-9. Like ETV5, ERG promotes p300 recruitment during the later stages of Th9 differentiation (Fig. 2)28. Conversely, during the early stages of Th9 differentiation, the expression of other ETS-family TFs like PU.1 is upregulated. These TFs can partially compensate for ERG, suggesting that they cooperatively regulate IL-9 production28.
SMADs
Like IL-2 and IL-4, TGF-β is a critical Th9-promoting cytokine12. When TGF-β engages its receptor, SMAD2 and SMAD3 are phosphorylated, associate with their common partner SMAD4, and translocate to the nucleus to drive the expression of downstream genes12. SMAD2/3 are critical for TGFβ-dependent IL-9 production but are dispensable for other IL-9-inducing cytokines like IL-4 and IL-1β31. Downstream of TGF-β, SMAD2 and SMAD3 do not directly bind the Il9 locus but are still required to displace EZH2 and remove suppressive H3K27 modifications at the Il9 promoter and CNS + 5.5 (DS) (Fig. 2)32. Several indirect mechanisms have been implicated in SMAD2/3-mediated Il9 epigenetic regulation. Phosphorylation of Serine 213 within the linker region of SMAD3 induces IL-9 by inducing the Albumin D-site-Binding Protein (DBP), a Proline and Acidic amino acid-Rich basic leucine ZIPper (PAR bZIP) TF that directly targets the Il9 promoter33. SMAD2 and SMAD/3 also modulate the binding of PU.1 and IRF4 to the extended Il9 locus, although they do not directly induce these TFs32. Indeed, IRF4 cannot induce Il9 transcription in the absence of SMAD2/3, suggesting that SMAD2 and SMAD/3 are required for IRF4 to regulate Il9 locus epigenetics (Fig. 3)32.
TAK1
TGF-β not only phosphorylates Smad2/3 but also triggers the activation of TGF-β activated kinase 1 (TAK1)34. Inhibition of TAK1 blocks the development of IL-9-producing CD4+ T cells but not the differentiation of other subsets34. TAK1 does not regulate PU.1 or IRF4 but rather inhibits the DNA-binding Il9 transcriptional repressor ID3, thereby promoting IL-9 production34. ID3 prevents E2A and GATA3 from binding the Il9 promoter, although it is unclear whether these mechanisms also regulate other Il9 cis-REs34. TAK1 also inhibits the histone deacetylase SIRT1, which represses the IL-9-inducing mTOR-HIF1α pathway35. Like E2A and GATA3, HIF1α directly targets the Il9 promoter (Fig. 2); its role at other CNS regions could be an area of future investigation35.
NF-κB
Multiple NF-κB-activating signals promote Th9 differentiation and induce IL-9 in other cell types12. For example, lipopolysaccharide enhances IL-9 production in activated mast cells36. In T cells, NF-κB (p65) binds the Il9 promoter to induce transcription downstream of T cell receptor (TCR) activation (Fig. 2)37. IL-9-promoting cytokines that signal through NF-κB include IL-1β, IL-33, and IL-36; IL-2 is also reported to promote IL-9 production via NF-κB induction31,38,39,40,41,42. In addition to directly targeting the IL9 locus, IL-1β promotes IL-9 production by suppressing the expression of the IL-9-inhibitory TF BCL639,43.
The TNFR superfamily contains many IL-9-inducing factors that act through NF-κB20,44,45,46,47. TNF-α induces IL-9 in CD4+ T cells, CD8+ T cells, and eosinophils20,48. OX40 is a costimulatory molecule expressed by activated CD4+ and CD8 + T cells; its ligand (OX40L) is exclusively expressed by antigen-presenting cells (APCs). Downstream of OX40L-OX40, p52-RelB directly binds the Il9 promoter, inducing IL-9 independent of the PU.1 and STAT TFs (Fig. 2)44. Although OX40 also promotes canonical (p50-RelA) NF-κB activation, this pathway is dispensable for IL-9 production44. OX40-OX40L signaling also induces Batf3, another IL-9-promoting TF (Fig. 2)49,50. Other IL-9-inducing TNFR family members include Fas, TL1A, and GITR (glucocorticoid-induced TNF receptor-related)20,44,45,46,47. Like OX40, GITR is a costimulatory molecule that induces IL-9 through TRAF6-NF-κB47. TL1A acts via STAT5 and BATF family TFs; although TL1A also activates NF-κB, this pathway may be less critical for TL1A-induced IL-9 production46. The TNFR family member Fas activates PKC-β, which both activates NF-κB and inactivates NFAT1. This has the dual effect of enhancing IL-9 production and creating a negative feedback loop that constrains Fas-induced Th9 differentiation45.
BATF family
The BATF family is composed of three members: BATF, BATF2, and BATF3, all expressed primarily in T and B cells6,49,51. IL-9-promoting inducers of BATF and BATF3 include TCR, OX40, TL1A, and IL-4/STAT66,49,50,51. BATF and BATF3 are highly homologous and can compensate for each other to induce IL-9 through similar epigenetic mechanisms49. BATF acts by binding the Il9 promoter and CNS-25 but cannot increase the accessibility of Il9 chromatin (Fig. 2)6. Consequently, BATF can only activate transcription when the locus is already poised by other TFs like STAT56. BATF3 also targets the Il9 promoter, where it interacts with IRF4 to form a complex that collaboratively enhances transcription (Fig. 2)49. Notably, PU.1 binding remains unaltered in Batf-deficient mice, indicating that PU.1 and BATF function independently50.
FOX family
Foxo1, a member of the forkhead box O (Foxo) family, plays a pivotal role in diverse cellular processes like cell survival, apoptosis, and T cell development52. During T cell differentiation, Foxo1 is phosphorylated and inactivated by PI(3)K/AKT signaling; IL-7-driven dephosphorylation activates Foxo1, as does TGF-β/Smad3 signaling52,53. After activation, Foxo1 binds and transactivates the Il9 and Irf4 promoters (Fig. 2), a process synergistically enhanced by IRF4-Foxo1 binding in a feed-forward loop52. STAT5 can also interact with Foxo1 signaling to induce IL-9 production by promoting permissive epigenetic changes like p300 recruitment and histone acetylation at the Il9 locus54. Simultaneously, Foxo1 inhibits the Th17 cell program by targeting Rorγt to modulate Th9-Th17 plasticity. Other FOX family TFs with a role in IL-9 regulation include Foxp1, which binds and negatively regulates the Il9 promoter, competing with Foxo154. Foxp3 can also suppress IL-9 via incompletely understood mechanisms involving GITR and STAT6 (Figs. 2 and 3)12.
IRFs
Interferon-regulatory factor 4 (IRF4) induces IL-9 in multiple cell types downstream of diverse stimuli including IL-33, thymic stromal lymphopoietin (TSLP), TGF-β, TL1A, and IL-412,24,55. IRF4 directly targets the Il9 promoter and CNS + 5.5, increasing locus accessibility (Fig. 2)13,50. Downstream of Smad3, IRF8 also targets the Il9 promoter and forms complexes with other TFs like BATF and PU.1 (Fig. 2)56. These complexes induce IL-9 production in Th9 cells and may have a role in other cell types56. IRF1 directly induces IL-9 downstream of IL-1β but represses IL-9 downstream of IFN-γː in both cases, IRF1 modulates accessibility of the Il9 locus13,25. These disparate results may be related to interactions with other TFs like NF-κB.
GATA3
In addition to activating STAT6, IL-4 induces GATA3, which promotes IL-9 production in T cells and potentially in other cell types23. Other TFs like the DNA-binding inhibitor Id3 modulate IL-9 indirectly through GATA3. Like many other TFs, GATA3 targets the Il9 promoter, although its binding has not been investigated at other cis-REs (Fig. 2)34.
PPAR-γ
Peroxisome proliferator-activated receptor γ (PPAR-γ) is a lipid-activated TF with a well-established role in Th2 cells57,58. Because of the overlap in Th2/Th9 identity and function, several studies have subsequently investigated the role of PPAR-γ in Th2-Th9 plasticity. In allergic subjects, IL-9 production can define a subpopulation characterized by an activation-induced signature and high PPAR-γ expression58. In this subpopulation, IL-9 production is transient, with IL-9+ cells developing into conventional Th2 cells. Notably, PPAR-γ agonists do not modulate IL-9 or Th2 cytokine production, implicating a ligand-independent mechanism58. The mTORC1 signaling pathway may modulate the effects of PPAR-γ on IL-9 production57.
HIF1α
Metabolic regulation is essential for T cell activation and differentiation due to complex and shifting energy requirements, mainly through aerobic glycolysis and oxidative phosphorylation35,57,59. HIF1α, a well-known TF, plays a pivotal role in orchestrating metabolic pathways in T cells, transitioning from aerobic to anaerobic metabolism, reducing ATP depletion, preventing the apoptosis of inflammatory effector cells, and modulating inflammatory capacity60. In CD4+ and CD8+ T cells, extracellular ATP promotes IL-9 production through nitric oxide (NO), mTOR, and HIF-1α61,62. HIF-1α directly targets the Il9 promoter (Fig. 2) and regulates Nos2 to induce NO in a feed-forward loop60. HIF-1α can also be activated by EGFR (epidermal growth factor receptor), and the EGFR ligand amphiregulin strongly induces Th9 differentiation downstream of IL-33 and TSLP60.
Vitamin receptors (RAR and VDR)
Vitamins A and D metabolites signal through nuclear receptors that act as TFs to modulate gene expression7. The vitamin D metabolite calcitriol engages VDR (vitamin D receptor), which interacts with PU.1 to block its binding to the Il9 promoter and repress the differentiation of IL-9-producing T cells (Fig. 2)63. Vitamin A metabolism produces several immunomodulatory metabolites, including retinoic acid (RA), which represses Th9 differentiation through RA receptor alpha (RARα)7. RARα recruits the corepressor NRIP1 to the Il9 promoter and upstream enhancers, reducing chromatin accessibility. Concurrently exposing Th9 cells to both metabolites negates this epigenetic repression by altering VDR-RXR (retinoic X receptor) interactions and HDAC recruitment64.
Other repressive TFs: TBX21, BCL6, and E2F8
TBX21, or T-bet, suppresses Th2 and Th9 differentiation while promoting Th1 differentiation; T-bet also suppresses IL-9 in ILCs26,65. While T-bet binds to key enhancers in different T helper subsets, its actions at the Il9 locus are unknown66. BCL6 is an IL-9-repressing TF that binds the Il9 promoter near the STAT5/6 binding site (Fig. 2)22,39,43. IL-2, IL-1β, and IL-21 modulate IL-9 expression by altering the relative expression of BCL6 and STAT522,39,43. Similarly, the IL-9-repressive TF E2F8 competes with DBP at the Il9 promoter: Smad3 represses E2F8 while inducing DBP (Fig. 2)33.
Together, these studies demonstrate that modulation of cis-REs within the Il9 locus underlies critical regulatory mechanisms involving a broad cross-section of TFs and stimuli.
Clinical/epidemiological associations of the extended IL9 locus with human disease
Allergy and atopic dermatitis
An extensive body of literature links IL-9 to type 2 (allergic) inflammation (Table 1). Expression levels of IL-9 and its receptor are increased in both murine allergy/asthma models and human subjects with allergy1. Blockade of IL-9 and its receptor improve pathology in murine disease models, whereas transfer of IL-9-producing cells exacerbates pathology1. While IL-9 blockade was not successful in early asthma clinical trials, many of these trials were performed in the pre-endotyping era, leading some investigators to propose newer trials in “IL-9high/Th9high” subjects2. Various single nucleotide polymorphisms (SNPs) in IL9 and the gene encoding its receptor, IL9R, are linked to allergic disease risk – including SNPs within putative IL9 cis-REs, although linkage disequilibrium can make it difficult to identify disease-causal SNPs within an extended locus67. For example, one disease-associated polymorphism, rs11741137, is located within CNS + 4.5 (DS)68. In subjects with proven housedust mite allergy, the T allele is associated with an increased risk of housedust mite induced severe asthma exacerbation68. Similar results were observed for patients with the A allele of rs2069885, located within the IL9 promoter. Also within the IL-9 promoter, the G allele/GG genotype at rs1859430 is associated with increased asthma risk, whereas the C allele/CC genotype at rs2066758 reduces disease risk69. The IL9 promoter SNP rs2069885 and intronic SNP rs2069882 are both associated with sex-specific differences in asthma risk70.
While IL9 locus genetics have been most extensively studied in asthma, increased IL-9 expression is also seen in atopic dermatitis (AD, Table 1)1. The rs31563 SNP located within the IL9 promoter increases allergic-type AD susceptibility, whereas the rs3093467 IL9R SNP is associated with non-allergic dermatitis71. Gene-gene interaction analysis suggests these two SNPs synergistically promote AD by combining the rs31563 GG and rs3093467 TT genotypes71.
Autoimmunity
In addition to allergic disease, increased IL-9 expression, Th9 cell expansion, and IL-9+ ILC2 expansion have also been reported in autoimmune diseases like inflammatory bowel disease (IBD), systemic sclerosis, rheumatoid arthritis (RA), psoriatic disease, and vasculitis72,73,74,75,76. IL-9 promotes antibody production and B cell proliferation, suggesting it may have a pathogenic role in autoimmune diseases75. In IBD, Th9 cells are elevated within the intestinal mucosa and prevent wound healing by promoting intestinal cell permeability74. In psoriatic disease, IL-9+ cells infiltrate skin and joints, and IL-9 induces pathogenic IL-17 expression77,78. The role of IL-9 in RA and scleroderma is less clear: IL-9 may promote wound healing and reduce fibrosis in some contexts, but IL-9 is also reported to induce fibrosis and worsen joint inflammation72,75,76,79,80,81. In multiple sclerosis, IL-9 is elevated within the central nervous system and reduces macrophage activation82.
Because ongoing studies are still defining the role of IL-9 in autoimmune diseases, the role of IL9 epigenetics in autoimmunity is not as well investigated as it is in allergy (Table 1). In patients with axial spondyloarthropathy, the IL9 promoter SNP rs2069885 A allele is associated with a favorable response to tumor necrosis factor (TNF) inhibitors83. Epigenetic modulators of IL-9 like PU.1 are implicated in murine models of autoimmunity including IBD and RA74,80. The rs3093457 IL9R SNP is also associated with RA, and further studies are needed to probe whether interactions with IL9 SNPs promote RA, as seen for IL9R SNPs in allergy84.
Cancer
IL-9 has both tumorigenic and antitumorigenic properties depending on the type of cancer and the tumor microenvironment. As a lymphocyte growth factor, IL-9 promotes tumorigenesis in many hematological malignancies85. These include T-cell malignancies like anaplastic large-cell lymphoma and human T-cell leukemia virus type 1 (HTVL-1) transformation, as well as B cell malignancies like Hodgkin’s lymphoma85. Yet, in the setting of hematopoietic stem cell transplantation, IL-9 can promote graft vs. leukemia-mediated antitumor effects86. IL-9 also has profound antitumor activity against solid tumors87. This is at least partly through stimulation of cytotoxic CD8+ lymphocytes, an effect that has been leveraged to engineer T cells bearing synthetic orthogonal IL-9 receptors with enhanced antitumor activity88.
Supporting the critical role of IL-9 in antitumor immunity, five SNPs within the IL9 promoter and CNS + 4.5 have been linked to laryngeal squamous cell carcinoma (LSCC, Table 1)89. The A-G-C-G-G haplotype of rs1859430-rs2069870-rs11741137-rs2069885-rs2069884 reduces disease risk, while the AA genotype at rs1859430 is associated with poor survival89. The G allele and GG genotype of rs1859430 in the IL9 promoter also increase the risk of recurrent pituitary adenoma (PA)90. Upstream of the IL9 promoter, rs740002 and rs1859428 within CNS-14 and CNS-20 (E2-E3) are associated with malignant melanoma, particularly in patients with pro-oncogenic CDKN2A mutations91. Together, these studies suggest that noncoding cis-REs modulate IL-9 expression, thereby affecting homeostatic tumor surveillance.
Other diseases
In addition to modulating immune cell signaling and function, IL-9 can also affect non-hematopoietic cells like keratinocytes, smooth muscle cells, and glial cells – although these roles are not well characterized1. These effects may underlie the associations of IL-9 with non-immunologic diseases; alternatively, IL-9 may promote subclinical immune dysfunction in patients with these diseases1. Age-related macular degeneration is negatively associated with haplotypes A-G-C-G-G and G-A-T-A-T in rs1859430, rs2069870, rs11741137, rs2069885, and rs2069884 (Table 1)92. Three of these SNPs are in the IL9 promoter, one is in CNS + 4.5, and one is in an intronic noncoding region. Cholesterol levels are associated with two IL9 promoter SNPs, while coronary artery disease is associated with SNPs in the IL9 promoter and intronic noncoding regions93,94. This is consistent with the observation that IL-9 worsens murine atherogenesis, while serum IL-9 is elevated in patients with atherosclerosis95.
Conclusions
The IL9 locus is a complex regulatory region comprising multiple cis-REs that interact with each other to modulate IL-9 production. Although most studies have been done in murine cells, the human IL9 locus contains putative REs homologous to major murine Il9 promoter and enhancer regions. Nonetheless, the human IL9 locus has some unique REs and structural features, so further studies should focus on characterizing IL-9 epigenetic regulation in human cells. Although several cell types can produce IL-9, most epigenetic studies have been done in Th9 cells. In Th9 cells, the IL9 locus exhibits a unique dynamic accessibility that permits innate-like IL-9 production in recently activated Th9 cells but also prevents sustained nonspecific IL-9 production over time. A complex network of TFs bind to and regulate the IL9 locus, including ETS family TFs, STATs, IRFs, NF-kB, SMADs, FOX family TF, and IRFs1. These TFs almost universally bind the IL9 promoter; some also engage upstream and downstream enhancers, suggesting that they may be involved in 3D chromatin looping.
Epigenetic regulation of IL-9 is seen in murine models of allergic asthma, autoimmunity, antihelminth defense, and antitumor immunity. SNPs within IL9 cis-REs are associated with a broad spectrum of inflammatory diseases, as well as nonimmunologic conditions. Together, these studies implicate epigenetic modulation as a core regulatory mechanism governing IL-9 production in the setting of human disease. However, many questions remain unanswered: these include the possibility of cell-specific REs, RE-specific TF binding, and the role of REs found exclusively in the human IL9 locus. Further investigations of these unanswered questions will provide a more complete view of epigenetic mechanisms governing IL-9 production and a broader understanding of noncoding REs in human physiology, homeostasis, and disease.
References
Pajulas, A., Zhang, J. & Kaplan, M. H. The World according to IL-9. J Immunol. 211, 7–14 (2023).
Son, A. et al. Dynamic chromatin accessibility licenses STAT5- and STAT6-dependent innate-like function of T(H)9 cells to promote allergic inflammation. Nat. Immunol. 24, 1036–1048 (2023).
Abdul Qayum, A. et al. The Il9 CNS-25 regulatory element controls mast cell and basophil IL-9 production. J. Immunol. 203, 1111–1121 (2019).
Xiao, X. et al. Guidance of super-enhancers in regulation of IL-9 induction and airway inflammation. J. Exp. Med. 215, 559–574 (2018).
Vahedi, G. et al. Super-enhancers delineate disease-associated regulatory nodes in T cells. Nature 520, 558–562 (2015).
Fu, Y. et al. STAT5 promotes accessibility and is required for BATF-mediated plasticity at the Il9 locus. Nat. Commun. 11, 4882 (2020).
Schwartz, D. M. et al. Retinoic acid receptor alpha represses a Th9 transcriptional and epigenomic program to reduce allergic pathology. Immunity 50, 106–120 e110 (2019).
Koh, B. et al. A conserved enhancer regulates Il9 expression in multiple lineages. Nat. Commun. 9, 4803 (2018).
Tan, C. et al. Antigen-specific Th9 cells exhibit uniqueness in their kinetics of cytokine production and short retention at the inflammatory site. J. Immunol. 185, 6795–6801 (2010).
Canaria, D. A. et al. STAT5 represses a STAT3-independent Th17-like program during Th9 cell differentiation. J. Immunol. 207, 1265–1274 (2021).
Olson, M. R. et al. Paracrine IL-2 is required for optimal type 2 effector cytokine production. J. Immunol. 198, 4352–4359 (2017).
Kaplan, M. H. The transcription factor network in Th9 cells. Semin. Immunopathol. 39, 11–20 (2017).
Campos Carrascosa, L. et al. Reciprocal regulation of the Il9 locus by counteracting activities of transcription factors IRF1 and IRF4. Nat. Commun. 8, 15366 (2017).
Noelle, R. J. & Nowak, E. C. Cellular sources and immune functions of interleukin-9. Nat. Rev. Immunol. 10, 683–687 (2010).
Kanno, Y., Vahedi, G., Hirahara, K., Singleton, K. & O’Shea, J. J. Transcriptional and epigenetic control of T helper cell specification: molecular mechanisms underlying commitment and plasticity. Annu. Rev. Immunol. 30, 707–731 (2012).
Olson, M. R., Verdan, F. F., Hufford, M. M., Dent, A. L. & Kaplan, M. H. STAT3 impairs STAT5 activation in the development of IL-9-secreting T cells. J. Immunol. 196, 3297–3304 (2016).
Yang, X. P. et al. Opposing regulation of the locus encoding IL-17 through direct, reciprocal actions of STAT3 and STAT5. Nat. Immunol. 12, 247–254 (2011).
Gomez-Rodriguez, J. et al. Itk is required for Th9 differentiation via TCR-mediated induction of IL-2 and IRF4. Nat. Commun. 7, 10857 (2016).
Wilhelm, C. et al. An IL-9 fate reporter demonstrates the induction of an innate IL-9 response in lung inflammation. Nat. Immunol. 12, 1071–1077 (2011).
Yang, S. et al. TNF-a is a potent stimulator of Tc9-cell differentiation. J. Immunother. 43, 265–272 (2020).
Gurram, R. K. et al. Crosstalk between ILC2s and Th2 cells varies among mouse models. Cell Rep. 42, 112073 (2023).
Liao, W. et al. Opposing actions of IL-2 and IL-21 on Th9 differentiation correlate with their differential regulation of BCL6 expression. Proc. Natl Acad. Sci. USA 111, 3508–3513 (2014).
Goswami, R. et al. STAT6-dependent regulation of Th9 development. J. Immunol. 188, 968–975 (2012).
Mohapatra, A. et al. Group 2 innate lymphoid cells utilize the IRF4-IL-9 module to coordinate epithelial cell maintenance of lung homeostasis. Mucosal Immunol. 9, 275–286 (2016).
Vegran, F. et al. The transcription factor IRF1 dictates the IL-21-dependent anticancer functions of TH9 cells. Nat. Immunol. 15, 758–766 (2014).
Zhang, Y. et al. Human TH9 differentiation is dependent on signal transducer and activator of transcription (STAT) 3 to restrain STAT1-mediated inhibition. J. Allergy Clin. Immunol. 143, 1108–1118 e1104 (2019).
Koh, B. et al. The ETS family transcription factors Etv5 and PU.1 function in parallel to promote Th9 Cell development. J. Immunol. 197, 2465–2472 (2016).
Kharwadkar, R. et al. ERG functionally overlaps with other Ets proteins in promoting TH9 cell expression of Il9 during allergic lung inflammation. J. Immunol. 210, 537–546 (2023).
Chang, H. C. et al. The transcription factor PU.1 is required for the development of IL-9-producing T cells and allergic inflammation. Nat. Immunol. 11, 527–534 (2010).
Goswami, R. & Kaplan, M. H. Gcn5 is required for PU.1-dependent IL-9 induction in Th9 cells. J. Immunol. 189, 3026–3033 (2012).
Xue, G., Jin, G., Fang, J. & Lu, Y. IL-4 together with IL-1beta induces antitumor Th9 cell differentiation in the absence of TGF-beta signaling. Nat. Commun. 10, 1376 (2019).
Wang, A. et al. Cutting edge: Smad2 and Smad4 regulate TGF-beta-mediated Il9 gene expression via EZH2 displacement. J. Immunol. 191, 4908–4912 (2013).
Park, S. A. et al. Opposing functions of circadian protein DBP and atypical E2F family E2F8 in anti-tumor Th9 cell differentiation. Nat. Commun. 13, 6069 (2022).
Nakatsukasa, H. et al. The DNA-binding inhibitor Id3 regulates IL-9 production in CD4(+) T cells. Nat. Immunol. 16, 1077–1084 (2015).
Wang, Y. et al. Histone deacetylase SIRT1 negatively regulates the differentiation of interleukin-9-producing CD4(+) T cells. Immunity 44, 1337–1349 (2016).
Stassen, M. et al. IL-9 and IL-13 production by activated mast cells is strongly enhanced in the presence of lipopolysaccharide: NF-kappa B is decisively involved in the expression of IL-9. J. Immunol. 166, 4391–4398 (2001).
Jash, A. et al. Nuclear factor of activated T cells 1 (NFAT1)-induced permissive chromatin modification facilitates nuclear factor-kappaB (NF-kappaB)-mediated interleukin-9 (IL-9) transactivation. J. Biol. Chem. 287, 15445–15457 (2012).
Olguin-Martinez, E. et al. IL-33 and the PKA pathway regulate ILC2 populations expressing IL-9 and ST2. Front. Immunol. 13, 787713 (2022).
Canaria, D. A. et al. IL-1beta promotes IL-9-producing Th cell differentiation in IL-2-limiting conditions through the inhibition of BCL6. Front. Immunol. 13, 1032618 (2022).
Harusato, A. et al. IL-36gamma signaling controls the induced regulatory T cell-Th9 cell balance via NFkappaB activation and STAT transcription factors. Mucosal Immunol. 10, 1455–1467 (2017).
Chen, J. et al. Interleukin-33 contributes to the induction of Th9 cells and antitumor efficacy by dectin-1-activated dendritic cells. Front. Immunol. 9, 1787 (2018).
Blom, L., Poulsen, B. C., Jensen, B. M., Hansen, A. & Poulsen, L. K. IL-33 induces IL-9 production in human CD4+ T cells and basophils. PLoS One 6, e21695 (2011).
Bassil, R. et al. BCL6 controls Th9 cell development by repressing Il9 transcription. J. Immunol. 193, 198–207 (2014).
Xiao, X. et al. OX40 signaling favors the induction of T(H)9 cells and airway inflammation. Nat. Immunol. 13, 981–990 (2012).
Shen, Y. et al. Fas signaling-mediated TH9 cell differentiation favors bowel inflammation and antitumor functions. Nat. Commun. 10, 2924 (2019).
Richard, A. C. et al. The TNF-family ligand TL1A and its receptor DR3 promote T cell-mediated allergic immunopathology by enhancing differentiation and pathogenicity of IL-9-producing T cells. J. Immunol. 194, 3567–3582 (2015).
Kim, I. K. et al. Glucocorticoid-induced tumor necrosis factor receptor-related protein co-stimulation facilitates tumor regression by inducing IL-9-producing helper T cells. Nat. Med. 21, 1010–1017 (2015).
Gounni, A. S. et al. IL-9 expression by human eosinophils: regulation by IL-1beta and TNF-alpha. J. Allergy Clin. Immunol. 106, 460–466 (2000).
Lee, W. H. et al. BATF3 is sufficient for the induction of Il9 expression and can compensate for BATF during Th9 cell differentiation. Exp. Mol. Med. 51, 1–12 (2019).
Tsuda, M. et al. A role for BATF3 in TH9 differentiation and T-cell-driven mucosal pathologies. Mucosal Immunol. 12, 644–655 (2019).
Jabeen, R. et al. Th9 cell development requires a BATF-regulated transcriptional network. J. Clin. Investig. 123, 4641–4653 (2013).
Malik, S. et al. Transcription factor Foxo1 is essential for IL-9 induction in T helper cells. Nat. Commun. 8, 815 (2017).
Buttrick, T. S. et al. Foxo1 promotes Th9 cell differentiation and airway allergy. Sci. Rep. 8, 818 (2018).
Bi, E. et al. Foxo1 and Foxp1 play opposing roles in regulating the differentiation and antitumor activity of TH9 cells programmed by IL-7. Sci. Signal. 10, https://doi.org/10.1126/scisignal.aak9741 (2017).
Staudt, V. et al. Interferon-regulatory factor 4 is essential for the developmental program of T helper 9 cells. Immunity 33, 192–202 (2010).
Humblin, E. et al. IRF8-dependent molecular complexes control the Th9 transcriptional program. Nat. Commun. 8, 2085 (2017).
Bertschi, N. L. et al. PPAR-gamma regulates the effector function of human T helper 9 cells by promoting glycolysis. Nat. Commun. 14, 2471 (2023).
Micosse, C. et al. Human “TH9” cells are a subpopulation of PPAR-gamma(+) TH2 cells. Sci. Immunol. 4, https://doi.org/10.1126/sciimmunol.aat5943 (2019).
Alladina, J. et al. A human model of asthma exacerbation reveals transcriptional programs and cell circuits specific to allergic asthma. Sci. Immunol. 8, eabq6352 (2023).
Roy, S. et al. EGFR-HIF1alpha signaling positively regulates the differentiation of IL-9 producing T helper cells. Nat. Commun. 12, 3182 (2021).
Roy, S. & Awasthi, A. ATP triggers human Th9 cell differentiation via nitric oxide-mediated mTOR-HIF1alpha pathway. Front. Immunol. 10, 1120 (2019).
Li, Y., Li, L., Zhang, W. & Gao, Y. Amphiregulin/epidermal growth factor receptor/hypoxia-inducible factor-1alpha pathway regulates T helper 9 and T cytotoxic 9 cell response in adult patients with infectious mononucleosis. Biomol. Biomed. 23, 63–72 (2023).
Vyas, S. P., Hansda, A. K., Kaplan, M. H. & Goswami, R. Calcitriol regulates the differentiation of IL-9-secreting Th9 cells by modulating the transcription factor PU.1. J. Immunol. 204, 1201–1213 (2020).
Vyas, S. P. & Goswami, R. Calcitriol and Retinoic acid antagonize each other to suppress the production of IL-9 by Th9 cells. J. Nutr. Biochem. 96, 108788 (2021).
Matsuki, A. et al. T-bet inhibits innate lymphoid cell-mediated eosinophilic airway inflammation by suppressing IL-9 production. J. Allergy Clin. Immunol. 139, 1355–1367 e1356 (2017).
Vahedi, G. Remodeling the chromatin landscape in T lymphocytes by a division of labor among transcription factors. Immunol. Rev. 300, 167–180 (2021).
Koch, S., Sopel, N. & Finotto, S. Th9 and other IL-9-producing cells in allergic asthma. Semin. Immunopathol. 39, 55–68 (2017).
Sordillo, J. E. et al. Genome-wide expression profiles identify potential targets for gene-environment interactions in asthma severity. J. Allergy Clin. Immunol. 136, 885–892 e882 (2015).
Chen, L. X. et al. Associations of IL-18 and IL-9 expressions and gene polymorphisms with asthma. Eur. Rev. Med. Pharmacol. Sci. 24, 6931–6938 (2020).
Aschard, H. et al. Sex-specific effect of IL9 polymorphisms on lung function and polysensitization. Genes Immun. 10, 559–565 (2009).
Namkung, J. H. et al. An association between IL-9 and IL-9 receptor gene polymorphisms and atopic dermatitis in a Korean population. J. Dermatol. Sci. 62, 16–21 (2011).
Rauber, S. et al. Resolution of inflammation by interleukin-9-producing type 2 innate lymphoid cells. Nat. Med. 23, 938–944 (2017).
Ohtsuki, S. et al. Deficiency of the CD155-CD96 immune checkpoint controls IL-9 production in giant cell arteritis. Cell Rep. Med. 4, 101012 (2023).
Gerlach, K. et al. TH9 cells that express the transcription factor PU.1 drive T cell-mediated colitis via IL-9 receptor signaling in intestinal epithelial cells. Nat. Immunol. 15, 676–686 (2014).
Ciccia, F. et al. Interleukin-9 and T helper type 9 cells in rheumatic diseases. Clin. Exp. Immunol. 185, 125–132 (2016).
Yanaba, K., Yoshizaki, A., Asano, Y., Kadono, T. & Sato, S. Serum interleukin 9 levels are increased in patients with systemic sclerosis: association with lower frequency and severity of pulmonary fibrosis. J. Rheumatol. 38, 2193–2197 (2011).
Schlapbach, C. et al. Human TH9 cells are skin-tropic and have autocrine and paracrine proinflammatory capacity. Sci. Transl. Med. 6, 219ra218 (2014).
Ciccia, F. et al. Interleukin-9 overexpression and th9 polarization characterize the inflamed gut, the synovial tissue, and the peripheral blood of patients with psoriatic arthritis. Arthritis Rheumatol. 68, 1922–1931 (2016).
Guggino, G. et al. Interleukin-9 over-expression and T helper 9 polarization in systemic sclerosis patients. Clin. Exp. Immunol. 190, 208–216 (2017).
Tu, J. et al. PU.1 promotes development of rheumatoid arthritis via repressing FLT3 in macrophages and fibroblast-like synoviocytes. Ann. Rheum. Dis. 82, 198–211 (2023).
Zhuang, R. et al. Perivascular fibrosis is mediated by a KLF10-IL-9 signaling axis in CD4+ T cells. Circ. Res. 130, 1662–1681 (2022).
Donninelli, G. et al. Interleukin-9 regulates macrophage activation in the progressive multiple sclerosis brain. J. Neuroinflamm. 17, 149 (2020).
Bialy, S. et al. Th2 Cytokines (Interleukin-5 and -9) polymorphism affects the response to anti-TNF treatment in Polish patients with ankylosing spondylitis. Int. J. Mol. Sci. 23, https://doi.org/10.3390/ijms232113177 (2022).
Burkhardt, J. et al. Association of the X-chromosomal genes TIMP1 and IL9R with rheumatoid arthritis. J. Rheumatol. 36, 2149–2157 (2009).
Cannon, A., Pajulas, A., Kaplan, M. H. & Zhang, J. The dichotomy of interleukin-9 function in the tumor microenvironment. J. Interferon Cytokine Res. 43, 229–245 (2023).
Ramadan, A. et al. Specifically differentiated T cell subset promotes tumor immunity over fatal immunity. J. Exp. Med. 214, 3577–3596 (2017).
Purwar, R. et al. Robust tumor immunity to melanoma mediated by interleukin-9-producing T cells. Nat. Med. 18, 1248–1253 (2012).
Kalbasi, A. et al. Potentiating adoptive cell therapy using synthetic IL-9 receptors. Nature 607, 360–365 (2022).
Pasvenskaite, A. et al. The role of IL-9 polymorphisms and serum IL-9 levels in carcinogenesis and survival rate for laryngeal squamous cell carcinoma. Cells 10, https://doi.org/10.3390/cells10030601 (2021).
Mickevicius, T., Vilkeviciute, A., Glebauskiene, B., Kriauciuniene, L. & Liutkeviciene, R. Do TRIB1 and IL-9 gene polymorphisms impact the development and manifestation of pituitary adenoma? In Vivo 34, 2499–2505 (2020).
Yang, X. R. et al. Identification of modifier genes for cutaneous malignant melanoma in melanoma-prone families with and without CDKN2A mutations. Int. J. Cancer 125, 2912–2917 (2009).
Vilkeviciute, A., Cebatoriene, D., Kriauciuniene, L., Zemaitiene, R. & Liutkeviciene, R. IL-9 and IL-10 single-nucleotide variants and serum levels in age-related macular degeneration in the caucasian population. Mediators Inflamm. 2021, 6622934 (2021).
Zha, L. et al. Genetic association analysis between IL9 and coronary artery disease in a Chinese Han population. Cytokine 150, 155761 (2022).
Chen, X., Wang, X., Zhang, Z., Chen, Y. & Wang, C. Role of IL-9, IL-2RA, and IL-2RB genetic polymorphisms in coronary heart disease. Herz 46, 558–566 (2021).
Zhang, W. et al. IL-9 aggravates the development of atherosclerosis in ApoE-/- mice. Cardiovasc. Res. 106, 453–464 (2015).
Funding
D.M.S. is funded in part by the National Psoriasis Foundation, the Jeffrey Modell Foundation, the Samuel and Emma Winters Foundation, and Eli Lilly. None of these funders contributed to this manuscript.
Author information
Authors and Affiliations
Contributions
A.S., I.B.: data review, writing; G.H.F.: figures, writing; and D.M.S.: supervision, writing, editing.
Corresponding author
Ethics declarations
Competing interests
D.M.S. consults for Sobi Pharmaceuticals and receives grant support from Eli Lilly.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Son, A., Baral, I., Falduto, G.H. et al. Locus of (IL-9) control: IL9 epigenetic regulation in cellular function and human disease. Exp Mol Med 56, 1331–1339 (2024). https://doi.org/10.1038/s12276-024-01241-y
Received:
Revised:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s12276-024-01241-y
This article is cited by
-
Targeting inflammation in cancer therapy: from mechanistic insights to emerging therapeutic approaches
Journal of Translational Medicine (2025)
-
Th9/IL-9 axis mediates airway fibrosis in traumatic tracheal stenosis via TGF-β1/SMAD2/3 signaling
Respiratory Research (2025)
-
Food allergy: begin at the skin, end at the mast cell?
Nature Reviews Immunology (2025)
