
Abstract
Euchromatic histone-lysine N-methyltransferase 2 (EHMT2) has been implicated in cardiovascular diseases, yet its role in vascular remodeling remains incompletely understood. Here we investigated the contribution of EHMT2 to vascular smooth muscle cell (VSMC) proliferation, migration and neointima formation following vascular injury using carotid artery injury models and in vitro VSMC studies. Transcriptomic (RNA sequencing) and epigenomic (CUT&Tag) profiling revealed that EHMT2 levels were elevated in injured arteries and growth-stimulated VSMCs, whereas EHMT2 deletion attenuated injury-induced neointima formation. Mechanistically, EHMT2 methyltransferase activity promoted VSMC proliferation and migration, with pathway analyses implicating cell cycle and growth programs as major downstream targets. We further identified GADD45G as a critical EHMT2-regulated gene characterized by H3K9me2 enrichment, and demonstrated that GADD45G enforced G1-phase arrest by suppressing cyclinB1, cyclinD1, CDK2 and CDK4. Importantly, both genetic and pharmacological inhibition of EHMT2, through GADD45G knockdown or administration of the EHMT2 inhibitor BIX-01294, significantly reduced neointimal lesion formation in injury models. These findings collectively establish EHMT2 as a key epigenetic driver of vascular remodeling by repressing GADD45G and facilitating cell cycle progression, highlighting EHMT2 as a potential therapeutic target for vascular proliferative diseases.
Similar content being viewed by others

Translational perspective
In this study, we demonstrate that EHMT2 plays a critical role in regulating VSMC proliferation and migration and in promoting injury-induced vascular neointima formation. Mechanistically, EHMT2 drives the pathological proliferation and migration of VSMCs through an epigenetic mechanism, namely lysine 9 on histone 3 (H3K9) dimethylation at the GADD45G locus. Collectively, these findings identify the EHMT2–GADD45G axis as a potential therapeutic target for preventing or treating pathological vascular remodeling.
Introduction
Neointima formation, also referred to as intimal hyperplasia, is a common feature of vascular remodeling, including postangioplasty stenosis1, atherosclerosis2 and vein bypass graft failure3,4. Compelling evidence has demonstrated that, in vascular remodeling, the vascular smooth muscle cell (VSMC) phenotype switches from a contractile phenotype to a synthetic phenotype while exhibiting decreased expression of contractile proteins and increased proliferation and migration abilities5. Therefore, an improved understanding of the mechanisms underlying VSMC proliferation and migration may be crucial for the development of novel therapeutics for vascular pathologies.
Accumulating evidence highlights the importance of epigenetic modifications in the pathological processes of VSMC proliferation and neointima formation6. Histone methylation is an important determinant of chromatin spatial structure, and the regulation of chromatin conformation and transcription factor accessibility to DNA is crucial in VSMC fate switching7,8. However, the nature of histone methylation modification factors and how such factors favor VSMC proliferation during neointima formation are still poorly understood.
The histone methyltransferase euchromatic histone-lysine N-methyltransferase 2 (EHMT2), also known as G9a, is a su(var)3-9/enhancer of zeste/trithorax (SET) domain-containing protein with histone lysine methyltransferase activity9. EHMT2 is responsible for the monomethylation and dimethylation of lysine 9 on histone H3 (H3K9me1 and H3K9me2), resulting in transcriptional gene activation or repression10. EHMT2 is involved in cell development, pluripotency, cell proliferation and migration11. Aberrant EHMT2 expression is frequently observed in multiple tumors and contributes to cancer pathogenesis and progression12. Recently, EHMT2 was shown to play a critical role in cardiovascular remodeling. For example, EHMT2 expression was increased in advanced human carotid atherosclerotic plaques, and EHMT2 inhibitors blocked the proliferation and migration of arterial smooth muscle cells (SMCs)13. EHMT2 and its partner protein GLP have been shown to ameliorate pulmonary vascular remodeling14. In addition, EHMT2 inhibits aortic SMC death by suppressing autophagy activation15. However, the role of EHMT2 in VSMC proliferation in vascular neointima formation remains unknown.
In this study, we explored the role of EHMT2 in the proliferation and migration regulation of VSMCs and vascular intimal hyperplasia. SMC-specific EHMT2 deletion in mice inhibited injury-induced neointima formation in wire-injured carotid arteries. In vitro and in vivo studies revealed that EHMT2 modulated VSMC proliferation and migration. Transcriptome and cleavage under targets and tagmentation (CUT&Tag) profiling revealed that EHMT2 exerted its effects by directly catalyzing the H3K9 dimethylation of GADD45G and eliminating the GADD45G-induced VSMC cell cycle arrest. Our findings provide evidence that EHMT2 plays a crucial role in the epigenetic regulation of GADD45G during neointima formation in response to vascular injury.
Materials and methods
Expanded methods are available in the Supplementary Information.
Mouse carotid artery wire injury model
The carotid arteries of these mice were subjected to wire injury as previously described16,17. In brief, 10–12-week-old male mice weighing 25–30 g were anesthetized by inhalation of 2% isoflurane. A 0.38-mm guidewire (C-SF-15-15; Cook) was inserted through the external carotid artery incision into the arterial lumen toward the aortic arch and pulled back and forth five times with a rotating motion. The sham littermate control mice underwent the same procedures without the incision and passage of the wire. The mice were intraperitoneally injected with BIX-01294 at an established dose of 5 mg/kg (once a week) for 28 days18,19. Dimethyl-sulfoxide-treated mice served as the control mice. Adeno-associated virus 9 (AAV) for SMC-specific transfection with a mouse GADD45G small interfering RNA (siRNA) (AAV-si-GADD45G) or negative control (AAV-siScrambled) was generated by Genechem Biotech. AAV-si-GADD45G titers (5.0 × 1012 vg/ml) or negative control virus was injected into the tail veins of male EHMT2SMCKO mice20.
Rat carotid artery injury model
Rat carotid artery neointima formation was induced by balloon angioplasty to the left common carotid artery as previously described21. The procedures were performed under sodium pentobarbital (50 mg/kg) anesthesia. To denudate the endothelium, a 2-Fr Fogarty balloon catheter (Cordis) was inserted into the common carotid, advanced to the aortic arch, inflated to produce moderate resistance and gradually withdrawn three times. The sham-operated rat carotid arteries served as the control. The lentivirus transfection into the injured vessels was performed as previously described22. In brief, for in vivo EHMT2-knockdown studies, 100 μl of sh-EHMT2 lentivirus (1 × 108 TU/ml, transducing units per ml) and the GFP lentivirus (1 × 108 TU/ml) (GeneChem, Shanghai, China) were luminally delivered into the injured carotid artery and incubated for 30 min to enhance transfection. After the lentivirus solution removal, the external carotid artery was ligated to restore the blood flow. The arteries were collected at the indicated time points.
Cell culture
VSMCs were isolated from the thoracic aorta of male Sprague–Dawley rats (150–180 g) using a collagenase I digestion method, as previously described23. The VSMCs were maintained in Dulbecco’s modified Eagle medium containing 10% fetal bovine serum, 100 U/ml penicillin and 100 μg/ml streptomycin. VSMCs at passages 3–8 and approximately 80% confluency were used for experiments.
Vector construction and cell transfection
Lentivirus particles carrying either EHMT2 cDNA, its variant p.Ala1130Ser, si-EHMT2, GADD45G cDNA and si-GADD45G were generated commercially (GeneChem). In brief, for construction of the lentiviral vector carrying the EHMT2 p.Ala1130Ser variant, the p.Ala1130Ser mutation was introduced using the QuikChange II XL Site-Directed Mutagenesis Kit (Agilent Tech).
For in vitro transfection, the recombinant lentivirus was directly transfected into VSMCs 72 h before the experiment. For the siRNA transfection, VSMCs cultured in six-well plates were transfected with 20 nM siRNA per well using Lipofectamine RNAiMAX reagent. The siRNAs targeting rat GADD45G, EHMT2 and scrambled sequence were synthesized by Sangon Biotech.
RNA-seq analysis
Total RNA was extracted from four independent samples of rat VSMCs subjected to indicated treatment. RNA sequencing (RNA-seq) analysis was performed at Majorbio Bio-Pharm Tech, with differential gene expression analysis performed using DESeq2 (adjusted P value <0.05 and |fold change| >1.5). The functional enrichment analyses of Gene Ontology (GO) terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways was subsequently carried out.
CUT&Tag assay
CUT&Tag experiments were performed using the Hyperactive In-Situ ChIP Library Prep Kit for Illumina (TD901-02, Vazyme), as previously described with modifications24. In brief, 100,000 cells were incubated with activated concanavalin A-coated magnetic beads. The bead-bound cells were permeabilized and successively incubated with H3K9me2 primary antibody (ab176882, Abcam) and secondary antibody. The beads were resuspended in pAG-Tn5-containing wash buffer. Tagmentation reaction was then performed. Extracted DNA fragments were used for polymerase chain reaction (PCR) amplification to prepare the DNA library. The CUT&Tag libraries were sequenced on an Illumina NovaSeq platform at Novogene.
CUT&Tag data analysis
The CUT&Tag sequencing generated the PE150 sequencing data. FASTQ files were processed to assess the quality of the raw sequencing data. Clean reads were aligned to the rat genome (National Center for Biotechnology Information reference) using Bowtie2. The aligned reads were indexed and sorted with samtools. Duplicate reads in the input were removed using Picard, and peak calling was performed with MACS2, excluding blacklist regions of the rat genome. DeepTools was used to calculate scores for genomic regions of interest and to generate heatmaps. BigWig tracks were viewed in the Integrative Genomics Viewer (IGV) browser.
Statistical analysis
All the experimental data are expressed as means ± standard errors of the mean. Statistical analysis was performed using GraphPad Prism (GraphPad, version 9.4.0). The Shapiro–Wilk normality test was performed before data comparison. Differences in means between two groups were analyzed via an unpaired two-tailed Student’s t-test or the Mann–Whitney U test as appropriate. Comparisons among more than two groups were performed using one-way analysis of variance (ANOVA) or the nonparametric Kruskal–Wallis test. A value of P < 0.05 was considered statistically significant.
Results
EHMT2 expression is upregulated in carotid artery neointima
To investigate the potential involvement of EHMT2 in the process of VSMC proliferation and neointima formation, we first analyzed EHMT2 expression in the rat balloon-injured carotid artery model. Cryosection staining of the carotid artery revealed that EHMT2 was mainly localized in the neointima and that EHMT2 expression was highly upregulated in the balloon-injured and wire-injured carotid neointima compared with the control at days 7 and 14 postinjury (Fig. 1a and Supplementary Fig. 1). Meanwhile, we found that the mRNA level of EHMT2 was significantly increased on day 7 postoperation and was maintained at that level until day 28 (Fig. 1b). Similarly, the protein levels of EHMT2 and the proliferation markers PCNA and p-H3 were markedly increased in balloon-injured carotid arteries at days 7, 14 and 28 postinjury (Fig. 1c).

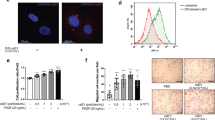
a Immunofluorescence staining shows the expression of EHMT2 and SM22α in the carotid arteries (n = 4) at day 14 after balloon injury. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI). Scale bar, 200 µm. The intensity was normalized to that of DAPI. b Relative EHMT2 mRNA levels in rat carotid arteries (n = 6) at different time points after balloon injury. The mRNA level was normalized to that of β-actin and quantified. c Relative protein levels of EHMT2, PCNA and p-H3 in balloon-induced carotid artery injury model rats (n = 4) at days 7, 14 and 28 postinjury were evaluated via western blot. d EHMT2 mRNA expression was quantified in rat VSMCs (n = 4) stimulated with different concentrations of PDGF-BB for 24 h. e Relative protein levels of EHMT2, PCNA and p-H3 in rat VSMCs (n = 4) stimulated with PDGF-BB at concentrations of 20 ng/ml or 40 ng/ml were evaluated via western blot. f Immunofluorescence staining showed the localization and expression of EHMT2 in VSMCs in rat VSMCs (n = 4). Nuclei were stained with DAPI. Scale bar, 20 µm. The intensity was normalized to that of DAPI. g Quantitative analysis of EHMT2 mRNA levels in rat VSMCs (n = 3) and human VSMCs (n = 3) obtained from the GEO database after PDGF-BB treatment. h Protein levels of H3K9me2 and EHMT2 in cultured VSMCs (n = 4) stimulated with PDGF-BB at a concentration of 20 ng/ml were evaluated via western blot. The data are presented as mean ± s.e.m. P values were calculated using an unpaired two-tailed t-test (a and f–h) or a one-way ANOVA (b–e).
To recapitulate those findings in vitro, we analyzed EHMT2 mRNA expression in cultured rat VSMCs in the presence of proliferative stimulus. Notably, platelet-derived growth factor-BB (PDGF-BB) is recognized as one of the most potent mitogens and chemokines acting on VSMCs and plays a central role in neointima formation after vascular injury25,26. Upon stimulation with different doses of PDGF-BB, EHMT2 mRNA expression was upregulated in a dose-dependent manner (Fig. 1d). PDGF-BB promoted the upregulation of PCNA and p-H3, accompanied with elevated EHMT2 protein levels in a dose-dependent manner in cultured rat VSMCs (Fig. 1e). Immunofluorescence staining of cultured rat VSMCs revealed that EHMT2 was located in the nucleus and further condensed upon PDGF-BB stimulation (Fig. 1f). Furthermore, our observation was supported in Gene Expression Omnibus datasets. EHMT2 mRNA expression was upregulated in PDGF-BB-stimulated human VSMCs in the GSE77280 dataset and in rat VSMCs in the GSE241545 dataset (Fig. 1g). In line with the H3K9me2 enzymatic activity of EHMT2, PDGF-BB treatment promoted the protein expression of both EHMT2 and H3K9me2 in VSMCs (Fig. 1h). Together, these results suggest that EHMT2 was aberrantly upregulated in synthetic VSMCs and neointima formation.
SMC-specific EHMT2 deficiency attenuates neointima formation following vascular injury
To investigate whether EHMT2 contributes to neointima formation, SMC-specific EHMT2-knockout mice were constructed, followed by injury of the carotid artery with a wire (Supplementary Fig. 2). The SMC-specific deletion of EHMT2 significantly decreased neointima formation in wire-injured carotid arteries in mice at day 14 postinjury (Fig. 2a), with a reduced area of neointima formation (Fig. 2b) and ratio of neointimal thickness to medial thickness (Fig. 2c) in the wire-injured carotid arteries compared with the control; however, the medial area was not notably altered (Fig. 2d).

a–d Representative hematoxylin and eosin (HE) staining images (a) and quantification of the neointima area (b) intima-to-media ratio (c) and media area (d) on day 14 after carotid artery wire injury in EHMT2flox/flox or EHMT2SMCKO mice (n = 6). Scale bar, 100 µm. e–h Representative HE staining images (e) and quantification of the neointimal area (f) intima-to-media ratio (g) and media area (h) of the balloon-injured rat carotid artery (n = 6) with or without intraarterial transfection of lentivirus carrying vector or sh-EHMT2 sequences at day 14 postinjury. Scale bar, 200 µm. i,j Immunofluorescence staining (i) and quantification of PCNA and SM22α in the carotid neointima (j) at day 14 after artery wire injury in EHMT2flox/flox and EHMT2SMCKO mice (n = 4). k,l The expression of EHMT2, H3K9me2, PCNA and SM22α was evaluated via western blot (k) and quantified (l) in the neointima of carotid artery after wire injury at day 14 postinjury in EHMT2flox/flox or EHMT2SMCKO mice (n = 4). m,n The expression of EHMT2, H3K9me2, PCNA and SM22α was evaluated via western blot (m) and quantified (n) in the neointima of carotid artery after balloon injury at day 14 postinjury in rats (n = 4) treated with lentivirus carrying vector or sh-EHMT2 sequence. The data are presented as mean ± s.e.m. P values were calculated using an unpaired two-tailed t-test (b–e, g, h and j) or a one-way ANOVA (l and n).
In the rat balloon-injured carotid artery model, the local delivery of lentivirus carrying an EHMT2 knockdown or empty vector was successfully performed. Compared with those in the empty vector group at day 14 postinjury, balloon injury-induced neointimal lesions were significantly reduced in the EHMT2-knockdown group after EHMT2 virus transduction (Fig. 2e), as determined by decreases in both the neointimal area and the intima-to-media ratio (Fig. 2f,g). Moreover, the media area of the rat carotid artery was unaffected after EHMT2 knockdown (Fig. 2h).
Meanwhile, immunofluorescence staining of PCNA and SM22α revealed that absence of EHMT2 decreased PCNA expression while increasing SM22α in carotid neointima (Fig. 2i,j). Western blot analysis confirmed that SMC-specific deletion of EHMT2 decreased the expression of H3K9me2 and PCNA but increased the expression of the VSMC contractile marker SM22α in mouse carotid arteries at day 14 (Fig. 2k,l). Consistently, the local delivery of EHMT2-knockdown lentivirus resulted in suppressed H3K9me2 and PCNA expression at day 14 in rat balloon-injured arteries (Fig. 2m,n). Taken together, these results suggest that EHMT2 acts as an important regulator of neointima formation.
EHMT2 regulates the proliferation and migration of VSMCs
To validate the abovementioned in vivo observations, we investigated whether EHMT2 affects VSMC proliferation in vitro. The siRNA effectively knocked down EHMT2 expression in VSMCs (Supplementary Fig. 3), and EHMT2 knockdown significantly suppressed the expression of H3K9me2 and PCNA, which was accompanied by the upregulation of SM22α expression (Fig. 3a). Furthermore, immunofluorescence assays for Ki-67, another well-recognized proliferation marker, revealed that EHMT2 knockdown dramatically decreased the proliferation rate of VSMCs (Fig. 3b). Cell Counting Kit-8 (CCK-8) proliferation assay showed that EHMT2 knockdown prevented PDGF-BB-evoked VSMC proliferation (Supplementary Fig. 4). Both scratch and transwell assays were performed to examine the role of EHMT2 in cell migration. Scratch assays revealed that the healing area was dramatically smaller in the EHMT2-knockdown group than in the control group upon PDGF-BB stimulation (Fig. 3c). Similarly, the transwell assay revealed fewer migratory cells in the EHMT2-knockdown group than the control group (Fig. 3d), suggesting that EHMT2 inhibition impairs cell migration ability.

a Representative western blot analysis and quantification of EHMT2, H3K9me2, PCNA and SM22α expression in VSMCs transfected with si-EHMT2 or si-Scr (n = 5). b VSMC proliferation (n = 4) was assessed via Ki-67 immunofluorescence. Nuclei were stained with DAPI. Scale bar, 100 μm. c Representative images of wound-healing assays. Images of si-EHMT2 or si-Scr control-treated VSMCs (n = 4) were taken at 0 and 12 h after scratching. Scale bar, 200 µm. d Representative images and quantitative analysis of the migration of VSMCs (n = 5) treated with si-EHMT2 or si-Scr control. Scale bar, 400 µm. e Representative western blot analysis and quantification of EHMT2, H3K9me2, PCNA and SM22α expression in VSMCs (n = 5) transfected with lentivirus carrying EHMT2 cDNA (oe-EHMT2) or control vectors. f VSMC proliferation (n = 4) was assessed via Ki-67 immunofluorescence, and the nuclei were stained with DAPI. Scale bar, 100 μm. g Representative images of wound-healing assays. Images of oe-EHMT2 or vector control lentivirus-transfected VSMCs (n = 3) were taken at 0 and 6 h after scratching. Scale bar, 200 µm. h Transwell assays of VSMCs (n = 5) were performed after treatment with and without oe-EHMT2. Scale bar, 400 µm. The data are presented as mean ± s.e.m. P values were calculated using an unpaired two-tailed t-test (a–h).
The results of the above loss-of-function experiments prompted us to investigate whether EHMT2 overexpression promoted the dedifferentiation of SMCs into a more proliferative state. Protein level analysis and fluorescence imaging revealed that EHMT2 was successfully overexpressed in VSMCs through lentiviral transduction (Supplementary Fig. 5). EHMT2 overexpression increased levels of H3K9me2 and PCNA (Fig. 3e) and enhanced VSMC proliferation, as determined by Ki-67 immunofluorescence staining (Fig. 3f), while lowering the level of contractile marker SM22α (Fig. 3e). Compared with the control, the overexpression of EHMT2 also promoted the migration of VSMCs, as determined by scratch (Fig. 3g) and transwell assays (Fig. 3h).
Moreover, two distinct EHMT2 chemical inhibitors, BIX-01294 and UNC0642, were used to determine the effects of EHMT2 inhibition. BIX-01294 and UNC0642 substantially suppressed EHMT2 enzymatic activity in cell-free systems27 (Supplementary Fig. 6). As expected, both BIX-01294 and UNC0642 effectively attenuated EHMT2 enzymatic activity in VSMCs (Supplementary Fig. 7a,b). BIX-01294 decreased VSMC proliferation as detected by CCK-8 analysis in a concentration-dependent manner in the presence of PDGF-BB (Supplementary Fig. 8). Furthermore, both BIX-01294 and UNC0642 inhibited the expression of VSMC proliferation markers PCNA and Ki-67, as well as VSMC migration in transwell assays, whereas EHMT2 overexpression partially reversed the inhibitory effects of these two inhibitors on proliferation and migration (Supplementary Fig. 7). Together, these observations suggest that EHMT2 mediates the proliferation and migration of VSMCs.
EHMT2 promotes VSMC proliferation in a methyltransferase activity-dependent manner
EHMT2 is a histone methyltransferase that functions primarily through H3K9me2 in mammals28. To elucidate whether the role of EHMT2 in VSMCs is dependent on its methyltransferase activity, the key amino acid in the catalytic domain of EHMT2 was predicted and mutated. According to the published EHMT2 catalytic domain structure in complex with the H3 tail29, Ala 1130 of rat EHMT2, which is homogeneous to Ala1077 in humans, was predicted to be a mutation candidate to alter the catalytic activity of EHMT2 (Fig. 4a). Therefore, rat EHMT2 Ala 1130, which is located in the catalytic SET domain, was substituted with serine (p.Ala1130Ser; Fig. 4b). A lentivirus carrying the p.Ala1130Ser variant was transfected into cultured rat VSMCs. Compared with the EHMT2 group, the p.Ala1130Ser group presented significantly decreased total H3K9me2 levels (Fig. 4c). Moreover, p.Ala1130Ser overexpression led to a decrease in the VSMC proliferation marker PCNA and an increase in the contractile marker SM22α (Fig. 4c). Furthermore, the p.Ala1077Ser variant reduced the percentage of Ki-67-positive VSMCs (Fig. 4d). Similarly, the results of the scratch and transwell assays revealed that the p.Ala1130Ser variant inhibited the migratory ability of VSMCs (Fig. 4e,f). Overall, the regulatory effect of EHMT2 on VSMC proliferation and migration is dependent on its methyltransferase activity.

a Analysis of histone methyltransferase sequence alignment across different species, with a specific focus on the rat p.Ala1130 residue and other residues linked to enhanced functional activity. b Structure of EHMT2 SET domain dimers interacting with histone H3 and SAM. Magnification revealing that the formation of hydrogen bonds between residue p.Ala1130 in EHMT2 and the histone tail residue and other EHMT2 residues. c Representative western blot analysis and quantification of H3K9me2, SM22α and PCNA expression in oe-EHMT2, oe-EHMT2(A1130S) and control lentivirus vector-transfected VSMCs (n = 4). d VSMC (n = 4) proliferation was assessed via a Ki-67 assay, and nuclei were stained with DAPI. Scale bar, 100 μm. e Representative images of wound-healing assays. Images were taken at 0 and 12 h after scratching monolayers of oe-EHMT2 VSMCs, oe-EHMT2 (A1130S) VSMCs and control VSMCs (n = 3). f Representative images and quantitative analysis of the migration of VSMCs (n = 5) treated with oe-EHMT2 and oe-EHMT2 (A1130S). Scale bar, 400 µm. The data are presented as mean ± s.e.m. P values were calculated using an unpaired two-tailed t-test (d–f) or a two-way ANOVA (c).
Identification of GADD45G as an EHMT2-mediated H3K9me2-targeted protein in VSMCs
To elucidate the underlying mechanisms by which EHMT2 regulates VSMC proliferation, the transcriptome of VSMCs treated with PDGF-BB in the presence or absence of EHMT2 siRNA was profiled via RNA-seq. Notably, principal component analysis revealed significant differences in gene expression patterns between the control and EHMT2-knockdown groups (Supplementary Fig. 9). PDGF-BB-treated VSMCs presented 1,382 differentially expressed genes (DEGs), whereas VSMCs cotreated with PDGF-BB and EHMT2 siRNA presented 752 genes with altered expression levels, resulting in 316 genes reciprocally upregulated by PDGF-BB and downregulated by EHMT2 siRNA (Fig. 5a). Genes ranked according to their association with proliferation or functional relevance to proliferation are displayed in a heatmap (Fig. 5b). KEGG and GO enrichment analyses revealed that multiple proliferation-related pathways, including cell cycle, cell growth, cell differentiation and apoptosis, were influenced by EHMT2 (Fig. 5c,d). Subsequent gene set enrichment analysis (GSEA) revealed that cell death and growth pathways, including MAPK cascade and JNK signaling, were enriched upon EHMT2 knockdown (Fig. 5e). These data further support that EHMT2 facilitates the growth of VSMCs.

a Venn diagram showed the number of DEGs uniquely identified in each comparison. b Heatmap of the transcriptomic profiling of the DEGs. c,d KEGG (c) and GO (d) analyses of DEGs in VSMCs. e GSEA of DEGs in VSMCs treated as indicated within pathways related to cell death and growth. f Pie chart displaying the genome-wide distribution of H3K9me2 occupancy in promoter, exon, intron and intergenic regions in VSMCs derived from CUT&Tag data. g Heatmap of the target genes associated with differential H3K9me2 peak binding sites in si-EHMT2 and si-Scr VSMCs following PDGF-BB stimulation. h Venn diagram depicting the intersection of genes under EHMT2 regulation, genes bound by H3K9me2 and target genes identified via GO pathway analysis of the RNA-seq data. i Analysis of motif interactions with enriched H3K9me2 peaks at the GADD45G and Klf15 loci. j IGV analysis of H3K9me2 peaks at the GADD45G and Klf15 loci in VSMCs. k ChIP–qPCR assay was performed to detect the relative binding level of H3K9me2 at the promoter region of GADD45G in VSMCs under different treatments.
Given that EHMT2 regulates H3K9me2 in mammals, CUT&Tag analysis was performed to identify candidate genes downstream of EHMT2. Global mapping analysis revealed that EHMT2 knockdown altered the H3K9me2 binding region distribution in the genome (Fig. 5f). In 3 replicates, 51,749 H3K9me2 interaction sites were identified after normalization with CUT&Tag. EHMT2 knockdown decreased the H3K9me2 signals of VSMCs, as shown in the general CUT&Tag data heatmaps (Fig. 5g), with a decrease of 13,958 H3K9me2 peak binding sites.
To further identify potential target genes of H3K9me2, we intersected genes showing hypomethylation in CUT&Tag data with upregulated genes from RNA-seq analysis. This approach identified 23 DEGs with associated H3K9me2 binding peaks (Fig. 5h). GO analysis for proliferation-related functions among these 23 DEGs highlighted four candidate genes: GADD45G, Bmp6, Nox4 and Klf15 (Fig. 5h). Subsequent motif analysis of H3K9me2 CUT&Tag-seq peaks revealed enrichment for canonical motifs corresponding to GADD45G and Klf15 (Fig. 5i), and IGV analysis confirmed H3K9me2 binding at the promoter regions of these two genes (Fig. 5j). While Klf15 is known to maintain the contractile phenotype of VSMCs and inhibit neointima formation, bioinformatic analysis indicated that EHMT2 binds specifically to the GADD45G promoter (Fig. 5j). This binding was functionally relevant, as EHMT2 knockdown reduced H3K9me2 enrichment at the GADD45G promoter (Fig. 5j). Based on these findings, we performed chromatin immunoprecipitation followed by quantitative PCR (ChIP–qPCR), which confirmed that EHMT2 directly regulates GADD45G transcription via H3K9me2 (Fig. 5k and Supplementary Fig. 10). Collectively, these data indicate that GADD45G is a downstream target modulated by EHMT2-mediated H3K9me2 during vascular remodeling.
GADD45G, which is negatively regulated by EHMT2, induces G1 cell cycle arrest during VSMC proliferation
To validate the link between EHMT2 and GADD45G, we further investigated whether GADD45G expression was negatively associated with EHMT2 in VSMCs. Western blot and reverse transcription qPCR revealed that EHMT2 knockdown increased GADD45G expression (Fig. 6a), whereas EHMT2 overexpression downregulated GADD45G expression (Fig. 6b). In murine carotid arteries with SMC-specific EHMT2 deficiency, GADD45G expression was elevated (Fig. 6c). Immunofluorescence staining indicated that the GADD45G protein was mainly localized in the cytoplasm (Fig. 6d). Together with the CUT&Tag analysis results, we concluded that GADD45G is a target gene regulated by EHMT2-mediated H3K9 methylation in VSMCs.

a The mRNA and protein levels of GADD45G in rat VSMCs (n = 4) treated with si-Scr or si-EHMT2. b The mRNA and protein levels of GADD45G in rat VSMCs (n = 4) treated with lentivirus vector (Vector) or carrying EHMT2 (oe-EHMT2). c Representative western blot analysis and quantification of GADD45G expression in EHMT2flox/flox and EHMT2SMCKO mice (n = 3). d Immunofluorescence staining of GADD45G in VSMCs (n = 4) with si-Scr or si-EHMT2. Nuclei were stained with DAPI. Scale bar, 20 µm. The intensity was normalized to that of DAPI. e The expression of GADD45G in rat VSMCs (n = 4) stimulated with PDGF-BB or the control was evaluated via western blot. f VSMC proliferation (n = 4) was detected via Ki-67 immunofluorescence, and nuclei were stained with DAPI. Scale bar, 100 μm. g Percentages of VSMCs in different phases of the cell cycle after being treated or untreated with si-EHMT2. h Percentages of VSMCs in different phases of the cell cycle after being treated or untreated with oe-EHMT2. i Representative western blot analysis and quantification of cyclin B1, cyclin D1, CDK2, CDK4 and p21 expression in si-GADD45G-treated VSMCs and control VSMCs (n = 4). j Representative western blot analysis and quantification of cyclin B1, cyclin D1, CDK2, CDK4 and p21 expression in oe-GADD45G VSMCs and control VSMCs (n = 4). P values were calculated using an unpaired two-tailed t-test (a–j).
Although GADD45G was shown to be involved in multiple GSEA pathways related to cell growth, it remains unknown whether GADD45G participates in the proliferation of VSMCs. GADD45G expression was downregulated upon PDGF-BB stimulation in VSMCs and in balloon injury-induced neointima as determined by western blot and immunostaining (Fig. 6e and Supplementary Fig. 11). GADD45G knockdown increased the number of Ki-67-positive VSMCs (Fig. 6f). Deregulation of the cell cycle in VSMCs is a universal hallmark of intimal hyperplasia and is considered a potential mechanism for inhibiting VSMC proliferation30. The cell cycle distribution of VSMCs was assessed via flow cytometry analysis, and we found that GADD45G knockdown increased the percentage of cells in the S phase (Fig. 6g). By contrast, the proportion of VSMCs in the S phase was significantly lower in the GADD45G-overexpressing group than in the vector control group (Fig. 6h). Moreover, the expression of the cell cycle markers cyclin B1, cyclin D1, CDK2, CDK4 and p21 was screened. GADD45G knockdown suppressed p21 expression and promoted cyclin B1, cyclin D1, CDK2 and CDK4 expression (Fig. 6i). GADD45G overexpression induced the expression of p21 and decreased the levels of cyclin B1, cyclin D1, CDK2 and CDK4, leading to cell cycle arrest in VSMCs (Fig. 6j). These data demonstrate that GADD45G, regulated by EHMT2, arrests the VSMC cell cycle at the G1 phase, preventing progression into the S phase.
EHMT2 regulates VSMC proliferation and migration through the negative regulation of GADD45G
To determine whether GADD45G mediates the inhibition of proliferation induced by EHMT2 silence, GADD45G was knocked down in VSMCs. GADD45G silencing abrogated the inhibitory effects of si-EHMT2 on VSMC proliferation, as determined by PCNA expression, EdU incorporation and CCK-8 analysis (Fig. 7a,b and Supplementary Fig. 12). Similarly, scratch and transwell assays revealed that the simultaneous knockdown of GADD45G and EHMT2 restored the decreased migratory ability of VSMCs after EHMT2 knockdown (Fig. 7c,d).

a Representative western blot analysis and quantification of PCNA and SM22α expression in VSMCs treated with si-Scr, si-EHMT2 and combined EHMT2/GADD45G knockdown (si-EHMT2 + si-GADD45G) (n = 5 in each group). b The proliferation of VSMCs (n = 4) was detected via immunostaining of EdU, and nuclei were stained with DAPI. Scale bar, 50 μm. c Representative images of wound-healing assays. Images were taken at 0 and 12 h after scratching monolayers in VSMCs treated with si-Scr, si-EHMT2 and combined EHMT2/GADD45G knockdown (si-EHMT2 + si-GADD45G) (n = 3 in each group). d Representative images and quantitative analysis of transwell migration assay in VSMCs treated with si-Scr, si-EHMT2 and combined EHMT2/GADD45G knockdown (si-EHMT2 + si-GADD45G) (n = 5 in each group). Scale bar, 400 µm. e Percentages of VSMCs in different phases of the cell cycle after treatment with si-Scr, si-EHMT2 and combined EHMT2/GADD45G knockdown (si-EHMT2 + si-GADD45G) (n = 5 in each group). f,g Representative western blot analysis (f) and quantification (g) of cyclin B1, cyclin D1, CDK2, CDK4 and p21 expression in VSMCs treated with si-Scr, si-EHMT2 and combined EHMT2/GADD45G knockdown (si-EHMT2 + si-GADD45G) (n = 4 in each group). h Immunofluorescence staining showed the expression of GADD45G and cyclin D1 in si-Scr, si-EHMT2 and combined EHMT2/GADD45G knockdown (si-EHMT2 + si-GADD45G) transfected VSMCs (n = 4 in each group). Nuclei were stained with DAPI. Scale bar, 100 µm. The intensity was normalized to that of DAPI. P values were calculated using a one-way ANOVA (a–h).
Given the regulation of cell cycle by GADD45G, we investigated whether EHMT2 and GADD45G are involved in the process of VSMC cell cycle arrest. Flow cytometry analysis of proportions of VSMCs revealed that EHMT2 knockdown induced G1 cell cycle arrest in VSMCs, an effect that was alleviated by the transfection with GADD45G siRNA (Fig. 7e). The screening of cell cycle markers revealed that EHMT2 knockdown led to the upregulation of p21 expression and the downregulation of cyclin B1, cyclin D1, CDK2 and CDK4 expression in VSMCs (Fig. 7f,g). Concomitant knockdown of GADD45G partially reversed these changes induced by EHMT2 silencing alone (Fig. 7f,g). Immunostaining of cyclin D1 suggested that GADD45G knockdown reversed the suppressed expression of cyclin D1 after EHMT2 knockdown (Fig. 7h). Taken together, these results suggest that the epigenetic suppression of GADD45G is an important downstream mechanism for EHMT2-mediated VSMC proliferation and migration.
Potential therapeutic effects of targeting the EHMT2–GADD45G axis in vascular remodeling
To test the potential therapeutic value of EHMT2 for vascular remodeling, EHMT2 was pharmacologically inhibited by BIX-01294 to assess changes in intimal hyperplasia. Consistent with the results of the genetic insufficiency experiments, BIX-01294 treatment, compared with control treatment, led to a substantial decrease in the neointima area and the intima-to-media ratio at 28 days postinjury (Fig. 8a,b). Meanwhile, protein levels of H3K9me2 and PCNA were downregulated in murine carotid arteries following BIX-01294 treatment (Supplementary Fig. 13).

a,b Representative images of HE-stained tissues (a) and quantification of neointima area and the intima-to-media ratio (b) on day 28 after wire injury of the carotid arteries of mice (n = 6) treated with or without BIX-01294. Scale bar, 100 µm. c,d Representative images of HE-stained tissues (c) and quantification of neointima area and the intima-to-media ratio (d) on day 14 after wire injury of the carotid arteries of the EHMT2flox/flox or EHMT2SMCKO mice (n = 6) transfected with AAV-si-scrambled or AAV-si-GADD45G. Scale bar, 100 µm. e,f Expression of GADD45G, cyclin D1, PCNA and SM22α in the carotid neointima (n = 3) after wire injury was evaluated via western blot (n = 3). g Immunofluorescence staining and quantification of PCNA and cyclin D1 in the carotid neointima (n = 4) at day 14 after wire injury. Scale bar, 20 µm. All data are presented as mean ± s.e.m. P values were calculated using an unpaired two-tailed t-test (b) or a one-way ANOVA (d, f and g).
Moreover, to validate the involvement of GADD45G in neointima formation after carotid artery injury, we intravenously injected adeno-associated viruses targeting VSMCs carrying siRNA against GADD45G into tail veins of EHMT2-knockout mice. We found that neointima formation was significantly restored by GADD45G knockdown at day 14 postinjury in EHMT2-knockout mice (Fig. 8c,d), together with increased protein levels of PCNA and cyclin D1, as well as decreased protein levels of GADD45G and SM22α, in injured carotid arteries (Fig. 8e,f). In addition, the inhibition of VSMC proliferation after EHMT2 knockout was alleviated after GADD45G knockdown, as shown by immunofluorescence staining for PCNA and cyclin D1 (Fig. 8g), suggesting that GADD45G overexpression might be a potential therapeutic strategy for neointima formation. Collectively, these findings indicate that the EHMT2–GADD45G axis represents a promising avenue for the development of novel preventive and therapeutic strategies for the treatment of vascular remodeling.
Discussion
Preventing restenosis following vascular injury is crucial for enhancing the therapeutic efficacy, prognosis and quality of life of patients with coronary artery disease after stent implantation. Here, we demonstrated that the targeted modulation of EHMT2 significantly reduces neointima formation induced by carotid artery injury. In this study, we confirmed that EHMT2 expression was upregulated in PDGF-BB-stimulated VSMCs and injured carotid arteries. In vivo, the targeted depletion of VSMC EHMT2 resulted in a reduction in injury-induced neointima formation in the carotid artery. Our findings revealed that the inhibition of EHMT2 led to a decrease in the proliferation and migration functions of VSMCs and a reduction in the proportion of cells in S phase; moreover, these biological functions were strongly dependent on the methyltransferase activity of EHMT2. Our data also demonstrated that EHMT2 modulates VSMC growth partially through histone methylation-mediated regulation of GADD45G and its involvement in cell cycle-related biological processes. In addition, the inhibition of GADD45G rescued VSMC proliferation and neointima formation ability after carotid artery injury in the context of EHMT2 depletion or deficiency. In conclusion, EHMT2 may represent a novel therapeutic target for vascular diseases associated with the abnormal proliferation and migration of VSMCs.
The expression of H3K9me2 has been reported to be lower in atherosclerotic plaques from patients compared with healthy human arteries13. Furthermore, Liang et al. profiled histone methylation via quantitative mass spectrometry and revealed that H3K9me2 expression was downregulated in cultured human VSMCs upon PDGF-BB stimulation31. Specifically, H3K9me2 was found to be reduced within VSMCs in atherosclerosis and injury-induced arterial remodeling via the use of in vivo VSMC lineage tracing systems32. Similarly, H3K9me2 levels were lower in VSMCs from patients with diabetes than in those from control patients33,34. These observations consistently illustrate the loss of H3K9me2 in VSMCs during atherosclerosis and vascular neointimal hyperplasia. Moreover, H3K9me2 is enriched in silenced genes and is related to decreased chromatin accessibility and transcriptional repression28. A reduction in H3K9me2 denotes activation of a series of genes favoring a more proliferative state in VSMCs. H3K9me2 is catalyzed mainly by EHMT2 rather than by EHMT1 (GLP) in mammals, and EHMT2 expression is upregulated during vascular remodeling. Data from the GEO database and the observations in our study, including those from VSMCs under PDGF stimulation and after balloon-induced carotid artery injury, were consistent with a previous report on the upregulation of EHMT2 expression in VSMCs from human carotid atherosclerotic lesions12. The apparent contradiction between decreased H3K9me2 and increased EHMT2 might be attributed to the fine and complicated balance between H3K9 methylation and demethylation by groups of histone lysine methylation modifiers. In addition, EHMT1 (GLP) and EHMT2 are structurally homologous and can form a functional heterodimer that catalyzes H3K9me235. EHMT1 might therefore partially compensate for H3K9me2 deposition when EHMT2 is deleted or downregulated. Genetic or pharmacological strategies targeting both EHMT1 and EHMT2 have been developed to achieve more complete suppression of H3K9me236. Accordingly, the use of BIX-01294 and UNC0642 in this study cannot fully rule out their potential inhibitory effects on EHMT1. Given the dynamic balance between methylation and demethylation and the functional redundancy between EHMT1 and EHMT2, we focused on EHMT2 to clarify its specific contribution to vascular remodeling.
Epigenetic regulation has been reported to be crucial during VSMC differentiation and neointima formation37,38. DNA methylation and DNA methyltransferases play pivotal roles in regulating vascular neointima formation and SMC plasticity39. Histone modifications, particularly acetylation and methylation, have been shown to be vital elements in the determination of VSMC plasticity. A growing body of evidence has shown that HDACs, histone acetyltransferases and methyltransferases contribute to injury-induced neointimal hyperplasia38,40. Notably, histone 3 lysine modifications have been shown to participate in VSMC plasticity. For example, H3K4me2 removal in VSMCs results in the loss of differentiation and reduced contractility due to the impaired recruitment of TET241. The overexpression of KDM3a, an H3K9me2-specific demethylase, accelerates neointima formation, whereas KDM3a knockdown reduces neointima formation33. In particular, other histone arginine methylations that are primarily mediated by PRMT5 were recently shown to regulate SMC marker gene transcription39,42. Together, these studies revealed the pivotal role of histone methylation in the proliferation and migration process of VSMCs and prompted us to investigate whether EHMT2 and its histone methyltransferase activity regulate the VSMC proliferation.
In this study, by constructing an EHMT2SMCKO mouse model, we found that EHMT2 deficiency led to reduced neointima formation. In addition, the downregulation of PCNA and upregulation of SM22α after the genetic deletion or chemical inhibition of EHMT2 indicated that the loss of EHMT2 inhibited the SMC transit into a proliferative state. In addition, it has been reported that EHMT2 is recruited by the transcription factor BACH1 and facilitates the BACH1-mediated modulation of VSMC phenotypic switching43. Here, our study further provided explicit and in-depth evidence on how EHMT2 itself participated the VSMC proliferation and neointima formation. Mechanistically, our RNA-seq analysis revealed that, in addition to genes related to the cell cycle and DNA synthesis, the underlying genes are related to cell death and apoptosis, findings that are consistent with the observation that EHMT2 inhibits aortic SMC death by suppressing autophagy activation. Moreover, GO analysis indicated that EHMT2 mediates important biological processes in SMCs, such as cell differentiation and development, the MAPK cascade and the JNK cascade, a finding that is consistent with the well-established role of EHMT2 in cancer cell proliferation44,45. Furthermore, the methyltransferase activity required for EHMT2-induced VSMC proliferation might suggest the importance of gene silencing during SMC transdifferentiation and neointima formation.
Furthermore, CUT&Tag screening revealed that GADD45G is di-methylated at H3K9 in its promoter region and may serve as a key mediator of EHMT2’s role in vascular remodeling. GADD45G has been reported to play key roles in the cellular stress response, DNA repair, autoimmunity and cell cycle regulation46,47. Specifically, GADD45G acts as a novel tumor suppressor, and GADD45G insufficiency leads to significantly enhanced tumor growth through Janus kinase/signal transducer and activator of transcription 3 activation48,49,50. GADD45G is widely recognized as a tumor suppressor. Loss of GADD45G has been shown to facilitate tumor growth, in part through activation of the JAK–STAT3 pathway. Beyond JAK–STAT3, GADD45G can restrain cell proliferation through multiple signaling cascades48,49,50. For example, in myeloproliferative neoplasms, GADD45G was reported to bind RAC2 to inhibit the RAC2–PAK1–PI3K/AKT axis. In acute myeloid leukemia, GADD45G activates p38 MAPK and thereby downregulates E2F1, leading to impaired DNA repair capacity and reduced proliferation50. In addition, GADD45G can interact with MAP3K4 to regulate p38 phosphorylation in astrocytes. However, only a handful of studies have investigated the role of GADD45G in CVD. GADD45G may be involved in myocardial cell death and left ventricular remodeling after myocardial infarction51. GADD45G overexpression induces significant alterations in cardiac morphology and histology, ultimately leading to cardiac remodeling and heart failure51. With respect to the role of GADD45G in the cellular stress response and cell cycle regulation, a regulatory function of GADD45G on the VSMC proliferation and vascular intimal hyperplasia is plausible. Given its established role in stress response and cell cycle regulation, GADD45G is hypothesized to modulate VSMC proliferation and intimal hyperplasia50. Its antiproliferative effects have been linked to DNA damage-repair mechanisms, and it can also regulate NF-κB-dependent inflammatory responses in a cell type-specific manner50. Collectively, these observations support a broader model in which GADD45G functions as a central integrator of DNA damage repair, proliferative signaling and inflammatory pathways to fine-tune cellular proliferation—potentially including VSMCs during vascular remodeling50. This study demonstrates that GADD45G acts as an inhibitor of VSMC proliferation, as indicated by the accelerated neointimal hyperplasia following GADD45G knockdown. Importantly, we provide the first evidence of the epigenetic regulation of GADD45G by histone methyltransferase EHMT2, showing that EHMT2-mediated enrichment of H3K9me2 on GADD45G exons represses its transcriptional activity. Overall, these findings expand the regulatory landscape of GADD45G in cell proliferation by linking its established role in cell cycle control to a previously underappreciated vascular context. These observations further identify the EHMT2–H3K9me2–GADD45G axis as a key epigenetic mechanism driving VSMC proliferation and injury-induced vascular remodeling.
Beyond its well-characterized role in histone methylation, accumulating evidence indicates that EHMT2 also regulates cellular processes through nonhistone mechanisms across diverse biological contexts. EHMT2 has a broad repertoire of nonhistone targets and has been shown to activate the tumor suppressor p53 via a mechanism independent of its methyltransferase activity52,53. Similarly, EHMT2 can repress MyoD-dependent transcription by depositing H3K9me2 at MyoD target promoters, and by directly interacting with MyoD and methylating specific lysine residues, thereby constraining its transcriptional activity54. EHMT2 has been implicated in promoting de novo DNA methylation55. Here, the nonhistone targets of EHMT2 were not explored, and it remains elusive whether EHMT2 directly methylates GADD45G or modulates GADD45G function through nonhistone pathways. Nevertheless, we speculate that nonhistone actions of EHMT2 may synergize with the EHMT2–H3K9me2–GADD45G axis, together forming an integrated regulatory network that shapes VSMC behavior during vascular remodeling.
In conclusion, the aberrant upregulation of EHMT2 expression promotes neointima formation in animal models. EHMT2 acts as a crucial epigenetic regulator of VSMC proliferation and migration in a methyltransferase activity-dependent manner. Mechanistically, EHMT2 controls the histone methylation status at the promoter region of GADD45G and represses GADD45G transcription. Thus, EHMT2 and GADD45G could be promising potential therapeutic targets for the development of novel strategies to treat vascular remodeling.
Data availability
The authors declare that all the data and methods supporting the findings of this study are available in the manuscript or Supplementary Material or from the corresponding authors upon reasonable request.
References
Gomez, D. & Owens, G. K. Smooth muscle cell phenotypic switching in atherosclerosis. Cardiovasc. Res. 95, 156–164 (2012).
Liu, M. W., Roubin, G. S. & King, S. B. 3rd Restenosis after coronary angioplasty. Potential biologic determinants and role of intimal hyperplasia. Circulation 79, 1374–1387 (1989).
de Vries, M. R., Simons, K. H., Jukema, J. W., Braun, J. & Quax, P. H. Vein graft failure: from pathophysiology to clinical outcomes. Nat. Rev. Cardiol. 13, 451–470 (2016).
Chinese College of Cardiovascular Physicians Chinese expert consensus on anti-thrombotic therapy for pan-vascular diseases (2023 edition). Cardiol. Plus 9, 49–69 (2024).
Motwani, J. G. & Topol, E. J. Aortocoronary saphenous vein graft disease: pathogenesis, predisposition, and prevention. Circulation 97, 916–931 (1998).
McDonald, O. G. & Owens, G. K. Programming smooth muscle plasticity with chromatin dynamics. Circ. Res. 100, 1428–1441 (2007).
McDonald, O. G., Wamhoff, B. R., Hoofnagle, M. H. & Owens, G. K. Control of SRF binding to CArG box chromatin regulates smooth muscle gene expression in vivo. J. Clin. Invest. 116, 36–48 (2006).
Blackledge, N. P. & Klose, R. J. The molecular principles of gene regulation by Polycomb repressive complexes. Nat. Rev. Mol. Cell Biol. 22, 815–833 (2021).
Ni, Y. et al. G9a in cancer: mechanisms, therapeutic advancements, and clinical implications. Cancers 16, 2175 (2024).
Bobyyleva, P. I., Tyrina, E. A., Lobanova, M. V. & Buravkova, L. B. Histone H3K9 methylation features under hypoxic conditions after the HIF1A knockdown in mesenchymal stromal cells in vitro. Bull Exp Biol Med 177, 431–435 (2024).
Scheer, S. & Zaph, C. The lysine methyltransferase G9a in immune cell differentiation and function. Front Immunol 8, 429 (2017).
Padeken, J., Methot, S. P. & Gasser, S. M. Establishment of H3K9-methylated heterochromatin and its functions in tissue differentiation and maintenance. Nat. Rev. Mol. Cell Biol. 23, 623–640 (2022).
Greissel, A. et al. Alternation of histone and DNA methylation in human atherosclerotic carotid plaques. Thromb. Haemost. 114, 390–402 (2015).
Awada, C. et al. G9a/GLP targeting ameliorates pulmonary vascular remodeling in pulmonary arterial hypertension. Am. J. Respir. Cell Mol. Biol. 68, 537–550 (2023).
Chen, T. Q. et al. EHMT2/G9a inhibits aortic smooth muscle cell death by suppressing autophagy activation. Int. J. Biol. Sci. 16, 1252–1263 (2020).
Zhang, S. M. et al. Interferon regulatory factor 9 is critical for neointima formation following vascular injury. Nat. Commun. 5, 5160 (2014).
Lindner, V., Fingerle, J. & Reidy, M. A. Mouse model of arterial injury. Circ. Res. 73, 792–796 (1993).
Kim, J. H. et al. BIX01294 inhibits EGFR signaling in EGFR-mutant lung adenocarcinoma cells through a BCKDHA-mediated reduction in the EGFR level. Exp. Mol. Med. 53, 1877–1887 (2021).
Irifuku, T. et al. Inhibition of H3K9 histone methyltransferase G9a attenuates renal fibrosis and retains klotho expression. Kidney Int. 89, 147–157 (2016).
Zeng, Z. et al. Circular RNA circMAP3K5 acts as a microRNA-22-3p sponge to promote resolution of intimal hyperplasia via TET2-mediated smooth muscle cell differentiation. Circulation 143, 354–371 (2021).
Zhang, W. & Trebak, M. Vascular balloon injury and intraluminal administration in rat carotid artery. J. Vis. Exp. 94, 52045 (2014).
Fan, Y. et al. Protective role of RNA helicase DEAD-box protein 5 in smooth muscle cell proliferation and vascular remodeling. Circ. Res. 124, e84–e100 (2019).
Wu, S. et al. Sodium–glucose cotransporter 2 inhibitors attenuate vascular calcification by suppressing endoplasmic reticulum protein thioredoxin domain containing 5 dependent osteogenic reprogramming. Redox Biol. 73, 103–183 (2024).
Kaya-Okur, H. S. et al. CUT&Tag for efficient epigenomic profiling of small samples and single cells. Nat. Commun. 10, 1930 (2019).
Contreras, O., Cordova-Casanova, A. & Brandan, E. PDGF–PDGFR network differentially regulates the fate, migration, proliferation, and cell cycle progression of myogenic cells. Cell Signal 84, 110036 (2021).
Li, L., Blumenthal, D. K., Terry, C. M., He, Y. & Carlson, M. L. PDGF-induced proliferation in human arterial and venous smooth muscle cells: molecular basis for differential effects of PDGF isoforms. J. Cell Biochem. 112, 289–298 (2011).
Mozzetta, C. et al. The histone H3 lysine 9 methyltransferases G9a and GLP regulate polycomb repressive complex 2-mediated gene silencing. Mol. Cell 53, 277–289 (2014).
Tachibana, M. et al. G9a histone methyltransferase plays a dominant role in euchromatic histone H3 lysine 9 methylation and is essential for early embryogenesis. Genes Dev. 16, 1779–1791 (2002).
Jayaram, H. et al. S-adenosyl methionine is necessary for inhibition of the methyltransferase G9a by the lysine 9 to methionine mutation on histone H3. Proc. Natl Acad. Sci. USA 113, 6182–6187 (2016).
Li, W. et al. The multifunctional Ca2+/calmodulin-dependent kinase II delta (CaMKIIdelta) controls neointima formation after carotid ligation and vascular smooth muscle cell proliferation through cell cycle regulation by p21. J. Biol. Chem. 286, 7990–7999 (2011).
Liang, J. et al. Inhibition of polycomb repressor complex 2 ameliorates neointimal hyperplasia by suppressing trimethylation of H3K27 in vascular smooth muscle cells. Br. J. Pharmacol. 176, 3206–3219 (2019).
Harman, J. L. et al. Epigenetic regulation of vascular smooth muscle cells by histone H3 lysine 9 dimethylation attenuates target gene-induction by inflammatory signaling. Arterioscler. Thromb. Vasc. Biol. 39, 2289–2302 (2019).
Chen, J. et al. Histone demethylase KDM3a, a novel regulator of vascular smooth muscle cells, controls vascular neointimal hyperplasia in diabetic rats. Atherosclerosis 257, 152–163 (2017).
Zhang, B. F. et al. KDM3A inhibition attenuates high concentration insulin‑induced vascular smooth muscle cell injury by suppressing MAPK/NF‑kappaB pathways. Int. J. Mol. Med. 41, 1265–1274 (2018).
Kramer, J. M. Regulation of cell differentiation and function by the euchromatin histone methyltranserfases G9a and GLP. Biochem. Cell Biol. 94, 26–32 (2016).
Jing, R. et al. Maturation and persistence of CAR T cells derived from human pluripotent stem cells via chemical inhibition of G9a/GLP. Cell Stem Cell 32, 71–85 (2025).
Elia, L. et al. UHRF1 epigenetically orchestrates smooth muscle cell plasticity in arterial disease. J. Clin. Invest. 128, 2473–2486 (2018).
Chakraborty, R. et al. Histone acetyltransferases p300 and CBP coordinate distinct chromatin remodeling programs in vascular smooth muscle plasticity. Circulation 145, 1720–1737 (2022).
Zhu, N. et al. Epigenetic regulation of vascular smooth muscle cell phenotypic switch and neointimal formation by PRMT5. Cardiovasc. Res. 119, 2244–2255 (2023).
Zhong, X. et al. The lysine methyltransferase SMYD2 facilitates neointimal hyperplasia by regulating the HDAC3–SRF axis. Acta Pharm. Sin. B 14, 712–728 (2024).
Liu, M. et al. H3K4 di-methylation governs smooth muscle lineage identity and promotes vascular homeostasis by restraining plasticity. Dev. Cell 56, 2765–2782 (2021).
Liu, H. et al. Protein arginine methyltransferase 5-mediated arginine methylation stabilizes Kruppel-like factor 4 to accelerate neointimal formation. Cardiovasc. Res. 119, 2142–2156 (2023).
Guo, J. et al. BACH1 deficiency prevents neointima formation and maintains the differentiated phenotype of vascular smooth muscle cells by regulating chromatin accessibility. Nucleic Acids Res. 51, 4284–4301 (2023).
Chae, Y. C. et al. FOXO1 degradation via G9a-mediated methylation promotes cell proliferation in colon cancer. Nucleic Acids Res. 47, 1692–1705 (2019).
Rao, V. K. et al. G9a promotes proliferation and inhibits cell cycle exit during myogenic differentiation. Nucleic Acids Res. 44, 8129–8143 (2016).
Liu, L. et al. Gadd45 beta and Gadd45 gamma are critical for regulating autoimmunity. J. Exp. Med. 202, 1341–1347 (2005).
Takekawa, M. & Saito, H. A family of stress-inducible GADD45-like proteins mediate activation of the stress-responsive MTK1/MEKK4 MAPKKK. Cell 95, 521–530 (1998).
Zhang, L. et al. Growth arrest and DNA damage 45G down-regulation contributes to Janus kinase/signal transducer and activator of transcription 3 activation and cellular senescence evasion in hepatocellular carcinoma. Hepatology 59, 178–189 (2014).
Zhang, P. et al. Gadd45g insufficiency drives the pathogenesis of myeloproliferative neoplasms. Nat. Commun. 15, 2989 (2024).
Guo, D. et al. GADD45g acts as a novel tumor suppressor, and its activation suggests new combination regimens for the treatment of AML. Blood 138, 464–479 (2021).
Lucas, A. et al. Gadd45gamma regulates cardiomyocyte death and post-myocardial infarction left ventricular remodelling. Cardiovasc. Res. 108, 254–267 (2015).
Rada, M. et al. Human EHMT2/G9a activates p53 through methylation-independent mechanism. Oncogene 36, 922–932 (2017).
Huang, J. et al. G9a and Glp methylate lysine 373 in the tumor suppressor p53. J. Biol. Chem. 285, 9636–9641 (2010).
Ling, B. M. et al. Lysine methyltransferase G9a methylates the transcription factor MyoD and regulates skeletal muscle differentiation. Proc. Natl Acad. Sci. USA 109, 841–846 (2012).
Leung, D. C. et al. Lysine methyltransferase G9a is required for de novo DNA methylation and the establishment, but not the maintenance, of proviral silencing. Proc. Natl Acad. Sci. USA 108, 5718–5723 (2011).
Acknowledgements
We appreciate Y. Sun at Xuzhou Medical University for critically reading the manuscript. We thank H. Kang, M. Deng and K. Li for their technical assistance.
Funding
This work was supported by the National Key Research and Development Program of China (grant no. 2022YFA1104204), the Natural Science Foundation of Chongqing, China (grant no. cstc2021jcyj-msxmX0716), Acute Radiation Syndrome Medical and Pharmaceutical Basic Research Innovation Center, Ministry of Education of China (ARSBIC-A-2026), the Chongqing Youth High-End Talent Project, and Special Project for Talent Cultivation of Xinqiao Hospital of Army Medical University (2022YQB001).
Author information
Authors and Affiliations
Contributions
Z.Q., J.R. and Y.Z. designed the study. Z.Q. and C.L. acquired the funding. Z.Q. and J. Zhang. supervised the research. Z.W., J. Zhao, W.L., X.M., B.W., F.Z. and H.T. performed the research and analyzed the data. N.S., K.N., J.H., X.W. and K.Z. assisted in performing the experiments. Z.Q., Z.W. and Y.Z. wrote the manuscript. All authors reviewed the paper and approved the final draft.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
All animal procedures conformed to the guidelines from Directive 2010/63/EU of the European Parliament on the protection of animals used for scientific purposes and were approved by the Laboratory Animal Welfare and Ethics Committee of Army Medical University (AMUWEC20234765/20245305). SMC-specific EHMT2-KO (EHMT2SMCKO) mice were created by crossing EHMT2flox/flox mice (GemPharma Tech Company) with Tagln-cre mice (B6.Cg-Tg[Tagln-cre]1Her/J; Jackson Laboratory). All animals were kept on standard condition and given access to chow and water ad libitum. Age- and sex-matched littermate SMC-Cre mice on a C57BL/6J background were used as control mice.
Consent for publication
Not applicable.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wang, Z., Zhao, J., Luo, W. et al. EHMT2 aggravates vascular remodeling via epigenetic inhibition of GADD45G. Exp Mol Med (2026). https://doi.org/10.1038/s12276-026-01702-6
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s12276-026-01702-6
