
Abstract
Anxiety disorders are a prevalent public health burden that significantly impair daily functioning and decrease quality of life. A growing body of research suggests DNA methylation (DNAm), an epigenetic modification that can impact gene expression, may be altered in anxiety disorders. The current review used a systematic approach to identify and synthesize the literature regarding epigenome-wide association studies (EWASs) of anxiety disorders in humans. We screened 804 articles returned by a search in PubMed in May 2025 and identified 12 studies for inclusion. All included studies examined DNAm in blood. In total, 2023 DNAm sites corresponding to 985 genes were significantly associated with anxiety disorders. Three DNAm sites significantly replicated across studies and four nominally replicated, meeting methylome-wide significance in one study and nominal significance (p < 0.05) in at least one other. This low replicability is likely a result of phenotypic heterogeneity, small sample sizes, the use of different multiple testing correction methods, and inconsistent adjustment for relevant comorbidities. Findings suggest DNAm associated with anxiety disorders may promote dysregulation of immune and inflammatory processes, some possibly sex-dependent. Collectively, the findings from studies included in this review provide preliminary evidence of DNAm alterations related to anxiety in whole blood and multiple blood cell-types. Future EWASs of anxiety disorders could benefit from using more ethnically diverse participants and longitudinal study designs.
Similar content being viewed by others
Introduction
Anxiety is a normative response to perceived and actual threats that, when dysregulated, can transform into a debilitating disorder characterized by worry and fear so excessive it is disturbing to one’s daily life [1]. Anxiety disorders are prevalent with over 300 million people worldwide experiencing one or more in 2019 [2]. The Diagnostic and Statistical Manual of Mental Disorders 5th edition (DSM-5) defines seven primary anxiety disorders with the prevalence of each ranging as follows: generalized anxiety disorder (GAD) (3%), panic disorder (2–3%), social anxiety disorder (SAD) (7%), specific phobia (8–12%), agoraphobia (1–2%), separation anxiety (2%), and selective mutism (< 1%) [3]. Research into the patho-etiology of anxiety disorders lags somewhat behind other mental disorders despite its high prevalence and widespread impacts. Heterogeneous clinical presentations and extensive comorbidity with other mental disorders complicate investigations of the etiology of anxiety disorders [4,5,6]. Like most medical phenotypes, risk for anxiety disorders is largely attributable to a combination of genetic and environmental factors [7, 8]. Anxiety disorders are highly polygenic, and heritability estimates based on known genetic variants range from 5–20% [9,10,11]. Yet, twin and family studies report higher heritability in the range of 20–60% [7, 12, 13], which leaves 15–40% of the heritability unaccounted for or “missing”. In comparison to other psychiatric disorders, a substantial portion of the heritability for anxiety disorders remains unexplained by known genetic variants [14, 15]. One proposed reason for this “missing” heritability is gene-environment interactions. While environmental factors, such as adverse life events, contribute independently to anxiety disorders [16, 17], they may also reveal genetic effects that only manifest in specific environmental contexts. This suggests genetic contributions to anxiety disorders may be underestimated when the role of environmental factors is ignored [18]. Regardless, the biological mechanisms through which genetic and environmental factors interact to exert their effects remain elusive.
Suggested biological mechanisms include epigenetic modifications that influence gene expression without altering the underlying DNA nucleotide sequence. We focus on DNA methylation (DNAm), the most studied epigenetic modification in humans. DNAm involves the addition of a methyl group to the carbon 5 position of a cytosine base and is most commonly found in the sequence context CpG (i.e., cytosine-phosphate-guanine). DNAm is maintainable across cell division and largely conserved throughout the human population with minimal interindividual variation at most CpGs, highlighting the integral role DNAm plays in proper cell and tissue functioning [19,20,21,22]. DNAm alterations occur in response to genetic and environmental triggers which change overtime across cells and tissues, suggesting DNAm may play a role in dynamically adapting to changing environments [23]. It is thought DNAm may confer increased risk for anxiety disorders by causing a maladaptive response to genetic and environmental factors [24]. In comparison to other psychiatric disorders where genetic contributions are stronger (e.g., schizophrenia), DNAm and other epigenetic modifications are expected to contribute to the etiology of anxiety disorders, particularly in the context of gene-environment interactions. A growing body of research has found evidence of altered DNAm in depression and post-traumatic stress disorder which are closely related to anxiety disorders [25, 26]; however DNAm alterations in anxiety disorders are far less studied and understood.
Early DNAm studies of anxiety disorders focused on CpGs in promoter regions of candidate genes involved in stress response and neurotransmission, but evidence of anxiety-related DNAm at these candidate genes is inconsistent. For example, the glucocorticoid receptor gene NR3C1 exhibited hypomethylation in panic disorder [27, 28], while hypermethylation was seen in adolescents and adults with GAD [29, 30]. Similarly, the oxytocin receptor gene OXTR was hypermethylated in females with GAD, panic disorder, agoraphobia, or specific and social phobia [31], but hypomethylated in males and females with social phobia [32]. The opposing directions of effect may indicate these genes are differentially impacted in individual anxiety disorders. However, candidate gene DNAm studies have been broadly criticized for their lack of replicability and high likelihood of false positive associations [33]. Several reasons have been suggested for this including small sample sizes which lack power to detect effects, no correction for multiple testing, and lack of adjustment for relevant confounders. For example, candidate gene DNAm studies typically do not control for differences in cell-type composition despite known impacts on comparisons of DNAm between samples [21, 34]. Not controlling for cell-type composition produces inflated test statistics and, therefore, a large number of seemingly significant associations. Thus, significant findings in candidate gene DNAm studies may reflect changes in cell-type composition related to anxiety disorders rather than true associations [21]. Further, candidate gene approaches typically examine a small number of CpGs in the promoter region of one gene with a posited role in anxiety disorders. Given the known polygenicity of anxiety disorders, methylomic associations are unlikely to be restricted to promoters or the few examined candidate genes. Such limited approaches put DNAm studies in danger of repeating the long, largely unsuccessful history of psychiatric candidate gene association studies [35,36,37,38].
Epigenome-wide association studies (EWASs) analyzing associations between observable traits and CpGs across the methylome are preferred, as they can overcome some limitations of candidate gene studies. EWASs are hypothesis-free and not limited to promoter regions of specific genes, allowing for a more comprehensive investigation of the methylomic landscape in anxiety disorders. Routine inclusion of relevant covariates, such as cell-type composition, and correction for multiple testing further decreases the likelihood EWAS findings are false positives [39]. These advances mirror those seen over the past decade in psychiatric genetic association studies [8, 13]. Reviews of EWASs of depression and PTSD have been conducted [25, 26], yet an in-depth examination of the methylomic landscape in anxiety disorders has not been investigated. Here, we conduct a systematic literature review to identify EWASs of anxiety disorders, describe study characteristics, and synthesize major themes among findings.
Methods
Search strategies and study selection
This systematic review was conducted in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [40]. The outcome of ‘anxiety disorders’ was defined as a formal or self-reported diagnosis or symptom count of any of the seven primary anxiety disorders. To be included, studies must have evaluated DNAm on a methylome-wide scale in humans, used a phenotype of anxiety disorders as defined above, and published in English in a peer-reviewed journal (i.e., preprints were not considered). If a study evaluated DNAm associations with additional phenotypes, we only extracted and synthesized information pertaining to anxiety disorders. We also only considered studies correcting for cell-type composition to ensure findings reviewed reflect true associations instead of differences in cell-type composition.
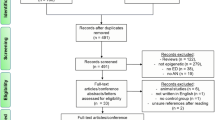
Studies were identified by searching PubMed for empirical articles meeting inclusion criteria in May 2025. The search term used was (“anxiety” OR “generalized anxiety” OR “specific phobia” OR “social phobia” OR “social anxiety” OR “agoraphobia” OR “panic” OR “panic disorder” OR “separation anxiety” OR “mutism”) AND (“methylome” OR “methylome wide” OR “methylome-wide” OR “epigenome” OR “epigenome wide” OR “epigenome-wide” OR “methylation” OR “DNA methylation”). Titles and abstracts were reviewed by SJI and SLC. Discrepancies were resolved by consensus. Duplicates, reviews, pre-prints, retractions, studies conducted in non-human or non-biological models, studies not examining anxiety disorders, and studies not examining DNAm on a methylome-wide scale were excluded (Fig. 1).

A Pubmed search in May 2025 yielded 799 unique articles after removal of duplicates. During abstract screening 783 articles were excluded because they were non-empirical articles, not conducted in humans, did not assess DNA methylation or did not assess it on a methylome-wide scale, or examined an outcome other than anxiety. The remaining 16 articles underwent full-text elligibility screening and 4 articles were excluded for having small sample sizes, not presenting or discussing methylome-wide association study results, or not correcting for cell-type composition. This lead to the inclusion of 12 studies in this report.
For the sixteen studies passing initial screening, data were extracted on study sample characteristics, study design, covariates included, multiple testing correction methods, main EWAS findings including the location of significant findings, and findings from replication attempts in independent samples. When reported, information on potential comorbidities were also extracted. Four studies were excluded for the following reasons: 1) conducted an EWAS in six participants [41], which is drastically underpowered to detect effects in an EWAS [42], 2) only evaluated downstream impacts of DNAm associated with anxiety disorders on biological pathways and did not report main EWAS findings [43], and 3) did not account for cell-type composition [44, 45].
We conducted a cross-study replication to assess the reproducibility of anxiety disorder EWAS findings and identify potential CpGs or genes warranting further investigation. For this purpose, we extracted information on CpGs from each study that met methylome-wide significance as defined by that study’s criterion (e.g., Bonferroni, false discovery rate or FDR) or were nominally significant (p < 0.05). Replication was evaluated at the positional level to enable the direct comparison of array- and sequencing-based findings when the same CpGs were assayed. CpGs at the same positional location meeting methylome-wide significance criteria in at least two studies were considered significantly replicated. Sites were considered nominally replicated if they were at the same positional location and met methylome-wide significance in one study and nominal significance defined as p < 0.05 in another. The same direction of effect was not used as a replication criterion as DNAm associations with anxiety may vary across individual disorders. We also assessed if there was replication on the gene-level by examining if CpGs that met methylome-wide significance criteria in more than one study were annotated to the same gene. Annotations provided by the original studies were used for this purpose.
Results
The search yielded 804 articles of which 12 met criteria for inclusion [23, 46,47,48,49,50,51,52,53,54,55,56]. These studies are summarized in Table 1. All studies evaluated DNAm associated with anxiety disorders cross-sectionally with 11 studies having a case-control design. Discovery samples ranged in size from 96 to 1522 participants. Most studies examined a single anxiety disorder or its symptoms, namely panic disorder, GAD, and SAD. The remaining studies evaluated associations with composite phenotypes of multiple anxiety disorders, either current or lifetime, though the disorders included in each study varied. The majority of participants across studies were of European ancestry, however multiple Asian sub-populations were also represented. Participants from all studies were mostly female consistent with the sex imbalance in anxiety disorder prevalence. Adolescents were considered in one study while adults were the focus of the remaining eleven.
EWAS findings reflect DNAm associations with anxiety disorders in blood only, as no studies used alternate tissues, and only one study conducted additional evaluations in different blood cell-types. DNAm was quantified with the Illumina 450k or EPIC arrays in 10 studies and sequencing-based methods in the remaining two, namely methyl-binding domain sequencing and targeted methyl-seq. Multiple testing correction was performed in all studies, though the choice of correction method and threshold for declaring methylome-wide significance varied. Although the covariates included were study-specific, potential confounding from age, sex, and cell-type composition was consistently controlled for across all studies. Potentially relevant psychiatric comorbidities were less consistently controlled for as only two studies accounted for depression [47, 50]. However, several studies used psychiatric comorbidities as an exclusion criterion [47, 49, 51]. Smoking was accounted for in seven studies. Four studies attempted to replicate their significant findings in an independent sample, with four CpGs replicating within-study. Potential test statistic inflation was reported in five studies, all of which used the genomic inflation factor λ. Six studies provided statements regarding the availability of raw data and/or summary statistics.
In total, 2023 CpGs reached methylome-wide significance after correction for multiple testing, which corresponded to 985 unique genes (Table S1). The literature collectively suggests anxiety disorders are associated with altered DNAm; however cross-study replicability was minimal as only cg03019505, cg25372841, and cg25947600 met methylome-wide significance thresholds in more than one study [47, 49] (Table 2). Four CpGs met methylome-wide significance in one study and nominal significance (p < 0.05) in another. Additional CpGs may meet nominal replication criteria, however we could not evaluate this possibility as most studies only reported methylome-wide significant findings or findings related to candidate genes. All significantly and nominally replicating CpGs were from array-based studies. A full list of all CpGs reported by the studies reviewed here and the reporting threshold (i.e., the criterion a finding must meet to be reported) used by each study are in Table S2. At the gene-level, 23 genes annotated to a significant finding in more than one study (Table S3). A binomial test confirmed this level of gene-overlap was more than is expected by chance (expected = 11 genes, p = 6.9 × 10−4), indicating a true enrichment.
Discussion
This systematic review examined methylome-wide DNAm associations with anxiety disorders, focusing on 12 empirical studies in humans. Notable strengths of the literature included consistent use of multiple testing correction and inclusion of some relevant confounders (e.g., age, sex, cell-type composition). Anxiety disorder phenotypes were clearly defined in all studies, which is critical for drawing meaningful conclusions. Findings from reviewed studies provide preliminary evidence of DNAm associations with anxiety disorders, however overall cross-study replicability was minimal, and the handful of findings implicated across multiple studies have only tentative links to anxiety. As such, it remains difficult to identify potential CpGs or genes warranting further investigation.
Phenotypic heterogeneity likely contributed to the lack of replication across studies, as phenotypes ranged from recent symptoms of generalized anxiety [46] to composites of either current or lifetime diagnosis of multiple anxiety disorders [47, 50, 52]. Most studies examined single anxiety disorders, and as such their findings likely reflect unique aspects of each anxiety disorder that may be unlikely to replicate across disorders. However, the results of single disorder studies can potentially be used to distinguish between anxiety diagnoses. For example, Ohi et al. [53] found DNAm in ANAPC16 was associated with SAD but not panic disorder, suggesting this finding may be an identifying marker for SAD. While clinically valuable, such disorder-specific signals offer limited insight into the potential role of DNAm across anxiety diagnoses. Studies using composite phenotypes comprised of multiple anxiety disorders are better positioned to identify DNAm with broad anxiety relevance. Three studies using composite phenotypes each reported significant associations [47, 50, 52], providing evidence that some DNAm may be common to multiple anxiety disorders. We note that there were no replicating findings between these studies, which may have been driven by the use of three different platforms for assaying DNAm. Each platform varies in their coverage of the genome and therefore may have limited overlap with one another. It also suggests DNAm could differ between current and lifetime anxiety disorder status. One EWAS found high correlations between analyses conducted using participants with a lifetime anxiety disorder versus a subset of participants with a current diagnosis [50], suggesting some sites may be useful for diagnostic support and risk prediction. However, preliminary evidence of dynamic DNAm changes following cognitive behavioral therapy [47] suggests that some DNAm could partially or fully return to baseline levels as symptoms improve and could be used to monitor response to treatment. Taken together, these findings underscore how phenotypic heterogeneity complicates the identification of replicable associations, while also highlighting the potential value and utility of this heterogeneity.
Low cross-study replicability was also driven by some of the same limitations as DNAm candidate gene studies including small sample sizes, use of multiple testing correction methods, and lack of adjustment for potential psychiatric comorbidities. Most studies used samples of less than 250 participants that were likely underpowered to detect the small DNAm differences expected between cases and controls [42]. Unlike candidate gene studies, all reviewed studies consistently corrected for multiple testing. However, the use of different methods for multiple testing correction may have impacted our ability to detect cross-study replication. The FDR approach used by most studies controls for an expected percentage of false positives among significant associations, typically 5%, instead of trying to eliminate false discoveries entirely. Being more lenient than other methods, using FDR may have introduced spurious associations that are unlikely to replicate across studies. The Bonferroni correction used in the remaining studies significantly reduces the likelihood of false positives in comparison to FDR approaches. Yet, this method may be too conservative for EWAS as correlations between proximally located CpGs mean the number of independent tests is often lower than the number of sites tested. Thus, some relevant associations may not have met stricter Bonferroni-corrected significance thresholds. Replicability was likely further impacted by the presence of psychiatric comorbidities as most studies did not control for psychiatric comorbidities by either excluding participants who had them or by adding comorbidity indicators as covariates. The few that did accounted for specific comorbidities like depression [47, 50, 51]. Other psychiatric disorders, such as PTSD, likely possess a unique DNAm signature. If they are not accounted for, it becomes unclear if anxiety EWAS associated effects reflect the anxiety disorder, the comorbidity or a combined anxiety-comorbidity effect. Thus, the specific effect being examined in each study may have varied, which could have affected replicability.
Three CpGs, cg03019505 of TFIP11, cg25372841 of CYHR1 and cg25947600 of C19orf68/ZSWIM9, met methylome-wide significance in two studies, positively associating with GAD [49] while negatively associating with a composite phenotype of either panic, agoraphobia, SAD or multiple phobic disorders [47]. Though none of these genes have been previously implicated in anxiety, TFIP11, CYHR1 and C19orf68/ZWIM9 are involved in pre MRNA splicing, cell proliferation, and blood cell development [57,58,59], respectively. Our finding suggests these processes may be impacted in multiple anxiety disorders. However, opposing directions of effect imply impacts to these processes could differ between distress-related anxiety disorders (e.g., GAD) and the fear-based anxiety disorders captured in Domschke et al.’s (2022) composite phenotype. Four CpGs, cg05910615 in C19orf65/HSPB6, cg20340149 in CLASP1, cg11029475 in GCAT/H1F0, cg13484946 in KIAA1841, met methylome-wide significance in Shimada-Sugimoto et al. [55] and nominal significance (p < 0.05) in Petersen et al. [54], which both examined DNAm in panic disorder. Though this supports the importance of these CpGs in panic disorder, Petersen et al. [54] conducted a meta-analysis that included the Shimada-Sugimoto study. As such, this overlap reflects shared participants across studies.
Genes implicated in more than one study have primarily been linked to anxiety-like behavior in murine models and other psychiatric phenotypes. Mice deficient in MUTYH, a part of the base excision repair pathway [60], and FAM19A5, a regulator of synaptic structure and function [61], display greater anxiety-like behaviors [62, 63]. Additionally, RASSF1 helps activate cell death to eliminate damaged cells and was found to influence anxiety-like behavior in a murine GWAS of anxiety [64, 65]. It is possible the impaired neural plasticity that accompanies anxiety could partly result from DNAm-related impacts on DNA damage repair and apoptosis processes. However, this link is not definitive and requires further substantiation. Other overlapping genes are also associated with DNAm changes in comorbid psychiatric disorders. For example, TNXB, the only gene annotated to significant CpGs in three studies [23, 49, 55], was hypermethylated in children and adolescents with PTSD in comparison to healthy controls [66]. The promoter of SYNE2 was also hypermethylated in the brains of individuals who committed suicide in comparison to non-psychiatric sudden death controls [67]. Replicating genes may impact mechanisms shared across psychiatric disorders but given high rates of misdiagnosis between anxiety disorders and other psychiatric disorders [68, 69] this offers little insight into their anxiety-specific importance.
Notably, replicating genes suggest DNAm may dysregulate immune and inflammatory processes in anxiety disorders. Examples include SMAD6, a known pro-inflammatory signaling inhibitor [70], and MUTYH, which helps activate the inflammatory response following oxidative stress [71]. However, single-study genes suggest immunity and inflammation may be impacted in a sex-specific manner. For example, a significant association with cg07308824 in the promoter of HECA was found in females, but not males, with panic disorder [51]. Given the posited role of HECA in mediating inflammation [72], some panic-related disruptions to inflammatory processes could be female-specific. As another example, severe GAD was associated with cg12701571 in the cytokine signaling regulator ASB1 in a sample of both sexes, suggesting cytokine production should be similarly altered in males and females [48]. Yet, a significant interaction between the pro-inflammatory cytokine Interleukin 18 (IL-18) and cg12701571 was only found in females, indicating specific inflammatory effects may be sex-dependent. Though preliminary evidence collectively indicates anxiety-related DNAm dysregulates immune and inflammatory processes in both sexes, future studies should explore potential sex differences in this dysregulation.
Considerations for future studies
While we found the ethnic diversity of samples used in EWASs of anxiety disorders was comparable to, or more diverse, than the EWAS literature for other psychiatric disorders, most included studies primarily focused on White participants. DNAm can differ between ethnicities and ancestries due to genetics and unique cultural influences [73, 74]. Methylation quantitative-trait loci (meQTLs) are genetic variants that influence DNAm at certain locations and may be specific to ancestral subgroups, yet only 75% of DNAm variation across ethnicities is explained by genetic variation [75]. Environmental and sociocultural experiences that vary across ethnicities, such as racial discrimination, dietary habits, or parenting styles, may explain remaining differences [76, 77]. CpGs associated with anxiety disorders and their effect sizes may differ between White and non-White individuals, bringing the generalizability of DNAm associated with anxiety disorders in mostly White participants into question. Future studies should make a concerted effort to include more ethnically diverse participants, which would allow the identification of associations relevant to all ethnicities. Such shared associations would enable a better assessment of the clinical relevance of DNAm associated with anxiety disorders, as they would not be specific to certain ethnic subgroups.
As all studies were cross-sectional, we were unable to identify the temporal ordering or direction of effect of DNAm associations with anxiety disorders (i.e., do anxiety disorders precede changes in DNAm or vice versa?). Changes in DNAm can promote their own downstream effects, yet studies have also found DNAm changes can occur downstream of transcription factor binding or alterations in gene expression [78,79,80]. Knowing if DNAm associations with anxiety disorders contribute to disease development or are promoted by it heavily impact their potential clinical relevance. Should DNAm changes precede anxiety disorder onset they may be useful predictive biomarkers or therapeutic targets in preventive or early intervention efforts. If anxiety disorder manifestation instead promotes changes in DNAm, they could be used for diagnostics and monitoring disease course. Carefully designed longitudinal studies that collect multiple phenotypic and DNAm measurements overtime, with measurement beginning before anxiety onset, should be conducted to establish if DNAm drives anxiety disorder development, or results from it.
Limitations of the current review
First, we limited our review to studies examining associations with DNAm assessed on a methylome-wide scale. Thus, we excluded candidate gene studies with their limited focus on DNAm in promoter regions and a high likelihood of false positives. However, we note none of the commonly studied candidate genes (e.g., NR3C1, OXTR) were among significant findings in the EWASs reviewed here (Table S1). We also did not examine the relationship between anxiety disorders and other epigenetic modifications (e.g., histone modifications, non-coding RNAs) for which few to no studies in humans exist despite evidence from animal studies supporting their involvement in anxiety [81]. Third, due to substantial variations in study design, PRISMA guidelines suggested the currently available literature was not sufficiently homogeneous to conduct a meta-analysis. As such, we were unable to further quantify the strength of evidence regarding DNAm associations with anxiety disorders. We also used each study’s definition of methylome-wide significance to assess replicability across studies. Some replicating findings may reflect false positives promoted by using FDR correction, which allows more CpGs to reach specified significance thresholds than other methods. Despite this, cross-study replication was minimal, and it is unlikely adopting a uniform replication threshold would have changed our overall conclusions. Further, for studies that did not report inflation metrics, we were unable to investigate potential inflation issues as most studies did not make the relevant information to do this readily available. In summary, our review was thorough, provides meaningful insight into key themes and limitations of the literature, and offers clear suggestions to enhance our understanding of DNAm associations with anxiety disorders.
Conclusion
The literature reviewed here lays the groundwork for future discoveries by providing initial evidence of DNAm associations with anxiety disorders in humans. Research examining the relationship between DNAm and anxiety disorders continues to expand and evolve, buoyed by advances in technology and a growing acknowledgment of the widespread and detrimental impacts of anxiety disorders. We are optimistic more replicable findings will be identified as sample sizes increase and limitations we have noted are addressed.
References
Craske MG, Rauch SL, Ursano R, Prenoveau J, Pine DS, Zinbarg RE. What is an anxiety disorder?. Depress Anxiety. 2009;26:1066–85.
Javaid SF, Hashim IJ, Hashim MJ, Stip E, Samad MA, Ahbabi AA. Epidemiology of anxiety disorders: global burden and sociodemographic associations. Middle East Current Psychiatry. 2023;30:44.
American Psychiatric Association. Diagnostic and statistical manual of mental disorders (5th ed.), 2013.
Doering S, Lichtenstein P, Gillberg C, Ntr, Middeldorp CM, Bartels M, et al. Anxiety at age 15 predicts psychiatric diagnoses and suicidal ideation in late adolescence and young adulthood: results from two longitudinal studies. BMC Psychiatry. 2019;19:363.
Drzewiecki CM, Fox AS. Understanding the heterogeneity of anxiety using a translational neuroscience approach. Cogn Affect Behav Neurosci. 2024;24:228–45.
Kessler RC, Chiu WT, Demler O, Merikangas KR, Walters EE. Prevalence, severity, and comorbidity of 12-month DSM-IV disorders in the national comorbidity survey replication. Arch Gen Psychiatry. 2005;62:617–27.
Hettema JM, Prescott CA, Myers JM, Neale MC, Kendler KS. The structure of genetic and environmental risk factors for anxiety disorders in men and women. Arch Gen Psychiatry. 2005;62:182–9.
Levey DF, Gelernter J, Polimanti R, Zhou H, Cheng Z, Aslan M, et al. Reproducible genetic risk loci for anxiety: results from approximately 200,000 participants in the million veteran program. Am J Psychiatry. 2020;177:223–32.
Tesfaye M, Jaholkowski P, Shadrin AA, van der Meer D, Hindley GFL, Holen B, et al. Identification of novel genomic loci for anxiety symptoms and extensive genetic overlap with psychiatric disorders. Psychiatry Clin Neurosci. 2024;78:783–91.
Mitchell BL, Skelton M, Wang R, ter Kuile AR, Murphy AE, Morneau-Vaillancourt G et al. Genome-wide meta-analysis identifies genetic risk factors and implicates multiple body systems in panic attacks and disorder. medRxiv [Preprint]. 2025. Available from: https://www.medrxiv.org/content/10.1101/2025.06.15.25329656v1.
Stein MB, Chen CY, Jain S, Jensen KP, He F, Heeringa SG, et al. Genetic risk variants for social anxiety. Am J Med Genet B Neuropsychiatr Genet. 2017;174:120–31.
Gottschalk MG, Domschke K. Genetics of generalized anxiety disorder and related traits. Dialogues Clin Neurosci. 2017;19:159–68.
Purves KL, Coleman JRI, Meier SM, Rayner C, Davis KAS, Cheesman R, et al. A major role for common genetic variation in anxiety disorders. Mol Psychiatry. 2020;25:3292–303.
Koromina M, Ravi A, Panagiotaropoulou G, Schilder BM, Humphrey J, Braun A, et al. Fine-mapping genomic loci refines bipolar disorder risk genes. Nat Neurosci. 2025;28:1393–403.
Owen MJ, Legge SE, Rees E, Walters JTR, O’Donovan MC. Genomic findings in schizophrenia and their implications. Mol Psychiatry. 2023;28:3638–47.
Danielsdottir HB, Aspelund T, Shen Q, Halldorsdottir T, Jakobsdottir J, Song H, et al. Adverse childhood experiences and adult mental health outcomes. JAMA Psychiatry. 2024;81:586–94.
Nelson EC, Heath AC, Madden PA, Cooper ML, Dinwiddie SH, Bucholz KK, et al. Association between self-reported childhood sexual abuse and adverse psychosocial outcomes: results from a twin study. Arch Gen Psychiatry. 2002;59:139–45.
Musci RJ, Augustinavicius JL, Volk H. Gene-Environment interactions in psychiatry: recent evidence and clinical implications. Curr Psychiatry Rep. 2019;21:81.
Ehrlich M, Lacey M. DNA methylation and differentiation: silencing, upregulation and modulation of gene expression. Epigenomics. 2013;5:553–68.
Farlik M, Halbritter F, Muller F, Choudry FA, Ebert P, Klughammer J, et al. DNA methylation dynamics of human hematopoietic stem cell differentiation. Cell Stem Cell. 2016;19:808–22.
Houseman EA, Kim S, Kelsey KT, Wiencke JK. DNA methylation in whole blood: uses and challenges. Curr Environ Health Rep. 2015;2:145–54.
Loyfer N, Magenheim J, Peretz A, Cann G, Bredno J, Klochendler A, et al. A DNA methylation atlas of normal human cell types. Nature. 2023;613:355–64.
Wiegand A, Kreifelts B, Munk MHJ, Geiselhart N, Ramadori KE, MacIsaac JL, et al. DNA methylation differences associated with social anxiety disorder and early life adversity. Transl Psychiatry. 2021;11:104.
Schiele MA, Gottschalk MG, Domschke K. The applied implications of epigenetics in anxiety, affective and stress-related disorders - a review and synthesis on psychosocial stress, psychotherapy and prevention. Clin Psychol Rev. 2020;77:101830.
Li M, D’Arcy C, Li X, Zhang T, Joober R, Meng X. What do DNA methylation studies tell us about depression? a systematic review. Transl Psychiatry. 2019;9:68.
Morrison FG, Miller MW, Logue MW, Assef M, Wolf EJ. DNA methylation correlates of PTSD: Recent findings and technical challenges. Prog Neuropsychopharmacol Biol Psychiatry. 2019;90:223–34.
Tyrka AR, Parade SH, Welch ES, Ridout KK, Price LH, Marsit C, et al. Methylation of the leukocyte glucocorticoid receptor gene promoter in adults: associations with early adversity and depressive, anxiety and substance-use disorders. Transl Psychiatry. 2016;6:e848.
Zou Z, Xiang M, Zhang Y, Huang Y, Wang J, He Y, et al. Associations of DNA methylation of HPA axis-related genes and neuroendocrine abnormalities in panic disorder. Psychoneuroendocrinology. 2022;142:105777.
Hua Y, Huang C, Guo Y, Du X, Guo L, Wang W, et al. Association between academic pressure, NR3C1 gene methylation, and anxiety symptoms among Chinese adolescents: a nested case-control study. BMC Psychiatry. 2023;23:376.
Wang W, Feng J, Ji C, Mu X, Ma Q, Fan Y, et al. Increased methylation of glucocorticoid receptor gene promoter 1(F) in peripheral blood of patients with generalized anxiety disorder. J Psychiatr Res. 2017;91:18–25.
Chagnon YC, Potvin O, Hudon C, Preville M. DNA methylation and single nucleotide variants in the brain-derived neurotrophic factor (BDNF) and oxytocin receptor (OXTR) genes are associated with anxiety/depression in older women. Front Genet. 2015;6:230.
Ziegler C, Dannlowski U, Brauer D, Stevens S, Laeger I, Wittmann H, et al. Oxytocin receptor gene methylation: converging multilevel evidence for a role in social anxiety. Neuropsychopharmacology. 2015;40:1528–38.
Shabalin AA, Aberg KA, van den Oord EJ. Candidate gene methylation studies are at high risk of erroneous conclusions. Epigenomics. 2015;7:13–15.
Qi L, Teschendorff AE. Cell-type heterogeneity: why we should adjust for it in epigenome and biomarker studies. Clin Epigenetics. 2022;14:31.
Border R, Johnson EC, Evans LM, Smolen A, Berley N, Sullivan PF, et al. No support for historical candidate gene or candidate gene-by-interaction hypotheses for major depression across multiple large samples. Am J Psychiatry. 2019;176:376–87.
Howe AS, Buttenschon HN, Bani-Fatemi A, Maron E, Otowa T, Erhardt A, et al. Candidate genes in panic disorder: meta-analyses of 23 common variants in major anxiogenic pathways. Mol Psychiatry. 2016;21:665–79.
Johnson EC, Border R, Melroy-Greif WE, de Leeuw CA, Ehringer MA, Keller MC. No evidence that schizophrenia candidate genes are more associated with schizophrenia than noncandidate genes. Biol Psychiatry. 2017;82:702–8.
Sullivan PF. Spurious genetic associations. Biol Psychiatry. 2007;61:1121–6.
Campagna MP, Xavier A, Lechner-Scott J, Maltby V, Scott RJ, Butzkueven H, et al. Epigenome-wide association studies: current knowledge, strategies and recommendations. Clin Epigenetics. 2021;13:214.
Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. Int J Surg. 2021;88:105906.
Alisch RS, Van Hulle C, Chopra P, Bhattacharyya A, Zhang SC, Davidson RJ, et al. A multi-dimensional characterization of anxiety in monozygotic twin pairs reveals susceptibility loci in humans. Transl Psychiatry. 2017;7:1282.
Mansell G, Gorrie-Stone TJ, Bao Y, Kumari M, Schalkwyk LS, Mill J, et al. Guidance for DNA methylation studies: statistical insights from the Illumina EPIC array. BMC Genomics. 2019;20:366.
Bortoluzzi A, Salum GA, da Rosa ED, Chagas VS, Castro MAA, Manfro GG. DNA methylation in adolescents with anxiety disorder: a longitudinal study. Sci Rep. 2018;8:13800.
Zou Z, Zhang Y, Huang Y, Wang J, Min W, Xiang M, et al. Integrated genome-wide methylation and expression analyses provide predictors of diagnosis and early response to antidepressant in panic disorder. J Affect Disord. 2023;322:146–55.
Zhou Q, Jackson-Cook C, Lyon D, Perera R, Archer KJ. Identifying molecular features associated with psychoneurological symptoms in women with breast cancer using multivariate mixed models. Cancer Inform. 2015;14:139–45.
Ciuculete DM, Bostrom AE, Tuunainen AK, Sohrabi F, Kular L, Jagodic M, et al. Changes in methylation within the STK32B promoter are associated with an increased risk for generalized anxiety disorder in adolescents. J Psychiatr Res. 2018;102:44–51.
Domschke K, Schiele MA, Crespo Salvador O, Zillich L, Lipovsek J, Pittig A, et al. Epigenetic markers of disease risk and psychotherapy response in anxiety disorders - a longitudinal analysis of the DNA methylome. Mol Psychiatry. 2025;30:4529–42.
Emeny RT, Baumert J, Zannas AS, Kunze S, Wahl S, Iurato S, et al. Anxiety associated increased CpG methylation in the promoter of Asb1: a translational approach evidenced by epidemiological and clinical studies and a murine model. Neuropsychopharmacology. 2018;43:342–53.
Guo L, Ni Z, Wei G, Cheng W, Huang X, Yue W. Epigenome-wide DNA methylation analysis of whole blood cells derived from patients with GAD and OCD in the Chinese Han population. Transl Psychiatry. 2022;12:465.
Hettema JM, van den Oord E, Zhao M, Xie LY, Copeland WE, Penninx B, et al. Methylome-wide association study of anxiety disorders. Mol Psychiatry. 2023;28:3484–92.
Iurato S, Carrillo-Roa T, Arloth J, Czamara D, Diener-Holzl L, Lange J, et al. “DNA Methylation signatures in panic disorder”. Transl Psychiatry. 2017;7:1287.
Kwon Y, Blazyte A, Jeon Y, Kim YJ, An K, Jeon S, et al. Identification of 17 novel epigenetic biomarkers associated with anxiety disorders using differential methylation analysis followed by machine learning-based validation. Clin Epigenetics. 2025;17:24.
Ohi K, Fujikane D, Takai K, Kuramitsu A, Muto Y, Sugiyama S, et al. Epigenetic signatures of social anxiety, panic disorders and stress experiences: insights from genome-wide DNA methylation risk scores. Psychiatry Res. 2024;337:115984.
Petersen CL, Chen JQ, Salas LA, Christensen BC. Altered immune phenotype and DNA methylation in panic disorder. Clin Epigenetics. 2020;12:177.
Shimada-Sugimoto M, Otowa T, Miyagawa T, Umekage T, Kawamura Y, Bundo M, et al. Epigenome-wide association study of DNA methylation in panic disorder. Clin Epigenetics. 2017;9:6.
Ziegler C, Grundner-Culemann F, Schiele MA, Schlosser P, Kollert L, Mahr M, et al. The DNA methylome in panic disorder: a case-control and longitudinal psychotherapy-epigenetic study. Transl Psychiatry. 2019;9:314.
Tannukit S, Crabb TL, Hertel KJ, Wen X, Jans DA, Paine ML. Identification of a novel nuclear localization signal and speckle-targeting sequence of tuftelin-interacting protein 11, a splicing factor involved in spliceosome disassembly. Biochem Biophys Res Commun. 2009;390:1044–50.
Desaki R, Sawada G, Okumura H, Ikeda R, Tanabe K, Komatsu H, et al. As a novel prognostic marker, cysteine/histidine-rich 1 (CYHR1) is a therapeutic target in patients with esophageal squamous cell carcinoma. Ann Surg Oncol. 2017;24:586–93.
ZSWIM9 zinc finger SWIM-type containing 9 (Homo sapiens) – Gene (Gene ID: 374920). https://www.ncbi.nlm.nih.gov/gene/374920, 2025, Accessed Date Accessed 2025 Accessed.
Curia MC, Catalano T, Aceto GM. MUTYH: Not just polyposis. World J Clin Oncol. 2020;11:428–49.
Shahapal A, Park S, Yoo S, Ma SX, Lee J, Kwak H, et al. Partial FAM19A5 deficiency in mice leads to disrupted spine maturation, hyperactivity, and an altered fear response. PLoS One. 2025;20:e0327493.
Bjorge MD, Hildrestrand GA, Scheffler K, Suganthan R, Rolseth V, Kusnierczyk A, et al. synergistic actions of Ogg1 and mutyh DNA glycosylases modulate anxiety-like behavior in mice. Cell Rep. 2015;13:2671–8.
Huang S, Zheng C, Xie G, Song Z, Wang P, Bai Y, et al. FAM19A5/TAFA5, a novel neurokine, plays a crucial role in depressive-like and spatial memory-related behaviors in mice. Mol Psychiatry. 2021;26:2363–79.
Hettema JM, Webb BT, Guo AY, Zhao Z, Maher BS, Chen X, et al. Prioritization and association analysis of murine-derived candidate genes in anxiety-spectrum disorders. Biol Psychiatry. 2011;70:888–96.
Dubois F, Bergot E, Zalcman G, Levallet G. RASSF1A, puppeteer of cellular homeostasis, fights tumorigenesis, and metastasis-an updated review. Cell Death Dis. 2019;10:928.
Ensink JBM, Keding TJ, Henneman P, Venema A, Papale LA, Alisch RS, et al. Differential DNA methylation is associated with hippocampal abnormalities in pediatric posttraumatic stress disorder. Biol Psychiatry Cogn Neurosci Neuroimaging. 2021;6:1063–70.
Labonte B, Suderman M, Maussion G, Lopez JP, Navarro-Sanchez L, Yerko V, et al. Genome-wide methylation changes in the brains of suicide completers. Am J Psychiatry. 2013;170:511–20.
Bradford A, Meyer AND, Khan S, Giardina TD, Singh H. Diagnostic error in mental health: a review. BMJ Qual Saf. 2024;33:663–72.
Vermani M, Marcus M, Katzman MA. Rates of detection of mood and anxiety disorders in primary care: a descriptive, cross-sectional study. Prim Care Companion CNS Disord. 2011;13:PCC.10m01013.
Lee YS, Park JS, Jung SM, Kim SD, Kim JH, Lee JY, et al. Inhibition of lethal inflammatory responses through the targeting of membrane-associated Toll-like receptor 4 signaling complexes with a Smad6-derived peptide. EMBO Mol Med. 2015;7:577–92.
Casorelli I, Pannellini T, De Luca G, Degan P, Chiera F, Iavarone I, et al. The Mutyh base excision repair gene influences the inflammatory response in a mouse model of ulcerative colitis. PLoS One. 2010;5:e12070.
Tissue Expression of HECA - summary - The human protein atlas. https://www.proteinatlas.org/ENSG00000112406-HECA/tissue, 2024, Accessed Date Accessed 2024 Accessed.
Elliott HR, Burrows K, Min JL, Tillin T, Mason D, Wright J, et al. Characterisation of ethnic differences in DNA methylation between UK-resident South Asians and Europeans. Clin Epigenetics. 2022;14:130.
Galanter JM, Gignoux CR, Oh SS, Torgerson D, Pino-Yanes M, Thakur N, et al. Differential methylation between ethnic sub-groups reflects the effect of genetic ancestry and environmental exposures. Elife. 2017;6:e20532.
Smith AK, Kilaru V, Kocak M, Almli LM, Mercer KB, Ressler KJ, et al. Methylation quantitative trait loci (meQTLs) are consistently detected across ancestry, developmental stage, and tissue type. BMC Genomics. 2014;15:145.
Chan MH, Merrill SM, Konwar C, Kobor MS. An integrative framework and recommendations for the study of DNA methylation in the context of race and ethnicity. Discov Soc Sci Health. 2023;3:9.
Meloni M, Moll T, Issaka A, Kuzawa CW. A biosocial return to race? a cautionary view for the postgenomic era. Am J Hum Biol. 2022;34:e23742.
Martin-Trujillo A, Patel N, Richter F, Jadhav B, Garg P, Morton SU, et al. Rare genetic variation at transcription factor binding sites modulates local DNA methylation profiles. PLoS Genet. 2020;16:e1009189.
Tseng CC, Wong MC, Liao WT, Chen CJ, Lee SC, Yen JH, et al. Genetic variants in transcription factor binding sites in humans: triggered by natural selection and triggers of diseases. Int J Mol Sci. 2021;22:4187.
Deplancke B, Alpern D, Gardeux V. The genetics of transcription factor DNA binding variation. Cell. 2016;166:538–54.
Ell MA, Schiele MA, Iovino N, Domschke K. Epigenetics of fear, anxiety and stress - focus on histone modifications. Curr Neuropharmacol. 2024;22:843–65.
Funding
This work was supported by the National Institutes of Health (SLC, grant number 5R01HD104297).
Author information
Authors and Affiliations
Consortia
Contributions
Conceptualization: SJI and SLC; Methodology: SJI, SLC; Data curation: SJI, SLC; Writing – original draft: SJI, LZ, SJI, SLC; Writing – review and editing: SJI, LZ, MAS, KD, JMH, SLC; Supervision: KD, JMH, SLC; Project administration; SLC.
Corresponding author
Ethics declarations
Competing interests
The authors declair no competing interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ingram, S.J., Zillich, L., Schiele, M.A. et al. A systematic review of methylome-wide associations with anxiety disorders. Transl Psychiatry 16, 183 (2026). https://doi.org/10.1038/s41398-026-03950-6
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41398-026-03950-6
