
Abstract
The majority of heat associated with climate change has been absorbed in the sunlit surface ocean where phytoplankton carry out half of biospheric net primary production (NPP). The physical entrainment of nutrients from depth into the surface constrains NPP across most of the ocean, therefore it has been widely hypothesized that a warmer and more thermally stratified ocean will diminish NPP. As phytoplankton are the dominant driver of ocean color, the satellite remote sensing record is the best approach to assess global NPP trends. Here we show that statistically significant decreases in NPP have occurred in almost half of the ocean and these changes are dominated by declines in the tropical and subtropical stratified ocean. A deeper analysis confirms that strengthening nutrient limitation is largely driving declining NPP. Climate-mediated shifts in NPP represent a fundamental perturbation to biogeochemical cycles that can further weaken global fisheries.
Similar content being viewed by others

Introduction
The ocean has absorbed more than 90% of the heat generated by anthropogenic climate change, resulting in a warmer and more thermally stratified environment across the vast majority of the ocean1,2. Most of this heat is confined to the upper few hundred meters of the ocean2, and it is in these environments where phytoplankton, a diverse assemblage of microscopic protists and cyanobacteria, carry out approximately half of biospheric photosynthesis3. Phytoplankton net primary production (NPP), the rate at which organic carbon is generated through photosynthesis, sets the foundation for marine food webs and fisheries4, is a key component of global biogeochemical cycles and a primary driver of biogenic carbon sequestration to the deep ocean5. The physical entrainment of vital nutrients from depth into the well-lit surface layer constrains NPP in most of the ocean6,7. Therefore, the prevailing conceptual view is that a warmer ocean with stronger thermal gradients will weaken nutrient entrainment and diminish NPP in much of the ocean8,9. Yet, how climate change is impacting NPP is still unclear. For example, multi-decadal direct measurements in the Atlantic and Pacific Oceans10,11 and earth system model projections show opposing trends12.
The key role of NPP in ecosystem services and the global carbon cycle has given rise to an ongoing pursuit of accurate, synoptic estimates at high resolution in both time and space13,14,15. Satellite remote sensing is the only platform that offers this capability, and past and present ocean color missions collectively provide an unbroken global time. Continual advances algorithms derived from remote sensing reflectance (Rrs), the primary metric of ocean color, have fundamentally improved the way we observe and estimate phytoplankton biomass, physiology, growth rates, and ultimately NPP from space16,17,18.
Here we present a 25-year global assessment across two satellite missions (SeaWiFS and MODIS-Aqua) to investigate how a warming ocean has influenced NPP in conjunction with underlying metrics of nutrient limitation. We use an absorption-based model that derives NPP as the product of photosynthetic active radiation (PAR), the fraction of PAR absorbed by phytoplankton (aϕ/a, where QPAR = PAR x aϕ/a), and the efficiency with which absorbed photosynthetic energy is converted into carbon biomass (ϕμ)19. Direct field measurements20,21 and culture-based experiments22,23 have shown that ϕμ has muted variance across taxonomic and nutrient gradients, consequently the vast majority of NPP variance is governed by ocean color measurements of QPAR. Physiological responses to increased nutrient limitation in phytoplankton are well understood and visible in the ocean color record24. As nutrient limitation increases, phytoplankton manufacture fewer light-harvesting pigments and associated photosynthetic machinery to match the lower energetic demands for growth25,26. This phenomenon is observable as concomitant decreases in the phytoplankton pigment absorption coefficient (aϕ) and phytoplankton growth rates (μ), where μ is derived by normalizing NPP to phytoplankton carbon biomass (CPhyto) as estimated from particulate backscattering27.
Monthly time series of NPP, growth rates, and all other ocean color parameters presented here (n = 303, global 1/12° data) were deseasonalized using monthly climatologies (1997–2022). Spatially explicit trend analysis (Type 1 ordinary least-squares regression and uncertainty analysis accounting for autocorrelation, see Methods), was performed on deseasonalized data and is presented as absolute trends and percent change relative to the 25-year mean. Empirical orthogonal function (EOF) analysis was performed to identify dominant modes of variance in time and space and compared to a suite of climate indices. The specific NPP model variant chosen has no direct dependence on sea surface temperature (SST)19, allowing for an independent assessment of NPP trends and variance in a warming ocean.
Results
Trend analysis
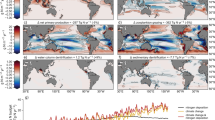
Across the satellite record, NPP is decreasing throughout vast regions of the subtropical and tropical ocean and increasing in polar and subpolar waters (Fig. 1a). Overall, NPP has significantly declined in 48% of the ocean at an average rate of −34 mg C m−2 d−1 decade−1, equivalent to a –7.1% normalized decadal rate. Conversely only 4% of the ocean has experienced a significant increase in NPP but at a higher average rate of 48 mg C m−2 d−1 decade−1, this is equivalent to a + 20% normalized decadal rate reflecting the fact that significant increases in NPP are largely occurring in low productivity regions (Fig. 1b, c). Spatially contiguous trends broadly correspond to major ocean biomes and currents (Supplementary Fig. 1). NPP is declining in all subtropical gyres and along currents that advect warm water poleward (Brazil, Gulf Stream, Kuroshio, Norwegian). Conversely, NPP is increasing along the austral subtropical front and either increasing or unchanged along eastern boundary currents that advect colder water towards the equator (California, Greenland, Malvinas, Oyashio, and Peru). A clear latitudinal anomaly is increasing NPP centered around 15°S in the southeast Pacific Ocean (Peru current). This region, along with the North Atlantic where NPP is also increasing, is one of the few areas globally where SST has cooled across the ocean color record1. The strong correspondence of SST and ΔNPP is further shown by grouping trends into respective 1 °C mean SST (1998–2022) bins and computing the area within each bin where NPP trends are significant (Fig. 1d, e). These data show that the 15 °C isotherm, which has been used to delineate temperate seasonally mixing biomes from warmer permanently stratified biomes24, also represents the transition from increasing to decreasing ΔNPP (Fig. 1d). Above 15 °C most declining trends exceed the 90% confidence interval, whereas below this temperature most of the increasing trends are not significant, and the largest increases in NPP are occurring in polar waters (Fig. 1e).

a NPP anomaly trends (Sep-1997 to Dec-2022) where hatching is used to denote regions where the trend exceeds the 90% confidence interval. b Mean NPP across the same period and c trends expressed as percent change from the mean. d All NPP trends divided into respective 1 °C mean sea-surface temperature (SST) bins (1998–2022 average) shown in the inset where boxes represent the interquartile range and whiskers represent the 10 and 90th percentile. e the areal extent of significant NPP trends (colored) within each SST bin relative to the total ocean area within each SST bin.
Trend analyses of the atmospheric (PAR), bio-optical (aϕ/a), and physiological (ϕμ) components of NPP demonstrates that changes in the fraction of energy absorbed by phytoplankton (aϕ/a) is largely driving NPP trends (Fig. 2). Overall, aϕ/a has significantly decreased in 43% of the ocean at an average normalized decadal rate of −7.6% and increased in 6% of the ocean at an average normalized decadal rate of +18%. Significant trends in PAR and ϕμ are present in only 16 and 22% of the ocean, respectively, and are comparatively muted, with average decadal rates less than 4% (both positive and negative for each parameter). There is strong spatial overlap in regions where both NPP and aϕ/a have significant trends. 86% (81%) of regions where NPP is significantly decreasing (increasing) has seen commensurate significant trends in aϕ/a. This analysis reveals that the dominant driver of NPP is embedded within the ocean color record, and the computationally simple term aϕ/a, derived directly from satellite data, is a robust indicator of shifts in marine productivity.

Average a photosynthetic active radiation (PAR), b fraction of PAR absorbed by phytoplankton (aϕ/a), and c net photosynthetic carbon efficiency (ϕμ), d–f, corresponding decadal trends normalized to their respective mean (1998–2022), where hatching denotes regions where the trend exceeds the 90% confidence interval.
Declining NPP and aϕ/a across most of the stratified ocean is consistent with stronger and more pervasive nutrient limitation. However, these declines could also occur through nutrient-independent increases in the spectral absorption coefficient of dissolved organic matter (adg) or decreases in phytoplankton biomass (Cphyto) without commensurate changes in growth rates (μ). Therefore to better resolve the mechanisms causing changes in NPP embedded in the ocean color record, normalized trends, slopes divided by their respective mean, of aϕ and adg, as well as euphotic zone integrated Cphyto and μ were derived and directly compared. Significant decreases (increases) in aϕ are occurring in 36% (6%) of the ocean (Fig. 3), significant decreases (increases) in adg are occurring in 25% (3%) of the ocean, a direct comparison of normalized decadal trends confirms that declining aϕ is the globally dominant trend in 33% of the ocean (Supplementary Fig. 2). Significant decreases (increases) in μ are occurring in 44% (4%) of the ocean and euphotic zone integrated Cphyto also registers statistically significant decreases (increases) in 17% (6%) of the ocean. Moreover, the normalized decadal Cphyto rates (−2% and +4%) are comparatively muted relative to the normalized decadal μ rates (−11%, +23%, Fig. 3). In this framework, decreases in aϕ, μ and NPP strongly suggest that the warming and more thermally stratified surface ocean is an increasingly nutrient limited environment, and that cellular reductions in photosynthetic pigments rather than changes in biomass is the dominant response observable in the ocean color record. This observation is mathematically consistent with statistically significant changes in Rrs that have emerged in more than half of the global ocean28.

Average a phytoplankton absorption coefficient (aϕ), b phytoplankton biomass integrated over the euphotic zone (Cphyto), and c phytoplankton growth rates (μ), d–f) corresponding decadal trends normalized to their respective mean (1998–2022), where hatching denotes regions where the trend exceeds the 90% confidence interval.
NPP anomaly variability in space and time
Previous analyses of global NPP anomalies (1997–2008) have shown that interannual variability is on the order of ±2 Petagram (Pg) C year−1 and is strongly influenced by SST (r = 0.80), particularly in the permanently stratified surface ocean29. Past analyses should be interpreted with some caution, however, as they often employ NPP models with first-order dependencies on SST30 and, unlike the model used here, their behavior cannot be evaluated independently from changes in SST. Across the 25-year record, NPP averaged 56.8 Pg C year−1 with a standard deviation of 1.9 Pg C year−1 (Fig. 4a). Global NPP declined at an average rate of 1.8 Pg C decade−1, though interannual variability is pervasive. This change represents a net 7.4% decrease over the satellite record, which greatly exceeds the CMIP6 high emissions scenario models (RCP8.5) that project NPP to decrease by 2.99% by the end of the 21st century12.

a Global Annual NPP across the satellite record (solid line), the dashed line is the linear fit and the gray region is the 90% confidence intervals. b The spatial (left panels) and temporal (right panels) components of the four greatest modes of NPP anomaly variance. Modes of variance are labeled as PC1-4 in the left panel and are shown in blue and green shading alongside their respective most correlated climate index (solid line, right axis). The percent explanatory variable of each mode is shown at the top of the panel alongside the correlation coefficient with the respective climate indices. Note that the magnitude of NPP anomalies through time have different scales.
To better elucidate the drivers and global patterns of NPP anomalies, we conducted EOF analysis to identify the dominant modes of NPP anomaly variance in space and time and then compared these modes to a suite of oceanic and atmospheric climate indices, including monthly global SST anomalies over the same time period (Fig. 4b). The first mode of global NPP anomaly variance (PC1) explains 13.3% of total variance, encapsulates the largest magnitude of NPP variance with a total range of 2.8 Pg C, and corresponds to global declines in NPP that become increasingly pronounced towards the equator. Global SST anomalies were the highest correlated climate index (r = -0.63), suggesting that a warming ocean is now the dominant driver of interannual variability in marine photosynthesis. The second mode of variance explains 5.3% of variability, is weakly positively correlated with SST anomalies (r = 0.23) and encapsulates a comparatively smaller range of NPP (0.7 Pg C). This second mode has a strong latitudinal signal, with modest negative and positive NPP anomalies in the subtropical northern and southern hemispheres, respectively. The most striking feature of PC2 is the pseudo-seasonal time component that emerges from deseasonalized data. A more detailed inspection of this mode shows that regions of high PC2 variance occur near the 15 °C isotherm in both hemispheres and these regions have undergone pronounced phenological shifts in NPP (Supplementary Fig. 3). In regions of high PC2 variance, NPP is increasing in the winter and decreasing in the summer in both hemispheres, resulting in diminished seasonality (Supplementary Fig. 3) that gives rise to pseudo-seasonal anomaly variance. That shifting phenology explains more of the global NPP anomaly variance than other established climate indices (e.g., El Niño) is notable. The third mode of variance explains 3.9% of NPP anomaly variability, encapsulates a total range of 0.9 Pg C, and is highly negatively correlated with the Pacific Decadal Oscillation (PDO, r = −0.63). The transition from a positive to strongly negative PDO in part explains the reversal in declining global rates from 2018 to 2021. Finally, the fourth mode of variance explains 2.8% of variability, encapsulates a total range of 0.5 Pg C, and is positively correlated with the El Niño 3.4 index (r = 0.61).
Deepening of NPP
Satellite ocean color measurements are restricted to the upper few meters of the surface mixed layer. In most of the ocean, the euphotic zone (depths where NPP > 0) extends beyond the surface mixed layer, and phytoplankton residing at depths beyond the reach of ocean color sensors often form so-called deep chlorophyll maxima (DCM). A key feature of DCMs is that they often represent a photoacclimative response to low-light, whereby cellular pigmentation increases with no change in biomass31. The NPP model employed here includes a photoacclimative response beneath the surface mixed layer that performs well compared to direct measurements19 and, when applied globally, constitutes more than 25% of global annual NPP (Fig. 5). Declining absorption in the surface deepens euphotic depths, thus permitting greater NPP at depth (Supplementary Fig. 4). The fraction of NPP within the DCM is sensitive to the surface mixed layer depth (MLD) which is regulated by wind stress, buoyancy input, and other physical processes. The MLD is deepening in some regions of the ocean and becoming shallower in other regions (Supplementary Fig. 4), although in general, the density contrast across the base of the surface mixed layer is strengthening32. On aggregate, based on a photoacclimative response alone, NPP beneath the surface mixed layer has increased by almost 3 Pg C across the satellite record and now constitutes more than 30% of global NPP (Supplementary Fig. 4).

a 1997–2022 average NPP beneath the surface mixed layer depth (NPPz>MLD). b The fractional contribution of NPPz>MLD to total NPP through time.
Discussion
Oceanic waters are the predominant sink of anthropogenic heat, and the rate of SST warming is now accelerating33. The analyses presented here definitively show that NPP is decreasing across most of the stratified ocean. By segregating the atmospheric, bio-optical, and physiological components of NPP, our analysis further reveals that phytoplankton are absorbing less solar energy and have diminished growth rates, consistent with a physiological response to strengthening nutrient limitation that is emerging in a warming ocean. Given that this physiological response is primarily manifested as diminished photosynthetic light harvesting capacity, it is also likely that marine photosynthetic oxygen production is declining globally. This climate-mediated trend also has significant implications for global fisheries. Earth system models demonstrate that diminishing NPP in marine environments is amplified at higher trophic levels through the elongation of plankton food-webs caused by an increased dominance of smaller phytoplankton cells as well as declining zooplankton growth efficiency as consumers have limited energy above basal metabolic requirements34,35.
The model and analyses presented here are agnostic to potential long-term changes in nutrient limitation beneath the surface mixed layer, and satellite-based NPP models that account for nutrient enhancement at depth use climatological nutrient data owing to the paucity of direct nutrient observations27. Given that the ocean color demonstrates that the warming and more stratified surface ocean is decreasing NPP in regions where DCMs are prevalent, and declining growth rates in these regions point to increasing nutrient limitation, the declining global NPP presented here may be conservative if nutrient limitation is becoming more prevalent beneath the surface mixed layer. Euphotic zone nutrient concentrations and NPP are declining in the subtropical North Atlantic10, and this decline has been linked to weakening ventilation of nutrient enriched sub-tropical mode water from depth36. Global change models forecast that enhanced stratification will diminish nitrate concentrations and NPP throughout the euphotic zone37, though this may be mitigated by enhanced atmospheric deposition in specific regions like the North Pacific38. In some regions, however, particularly equatorial waters, nutrient enhancement beneath the surface mixed layer can fuel additional NPP39.
Our analysis is observable within and consistent with reported changes in the satellite ocean color record28, emphasizing the critical importance of continued investment in Earth-observing satellites and associated research. There are a number of emerging approaches and capabilities that can improve our ability to monitor changes in NPP at global scales40. For example, the now operational hyperspectral plankton, aerosol, cloud, and ecosystem mission can provide more accurate assessments of phytoplankton community composition and biomass41. NPP model variants that incorporate these features lead to more accurate and detailed assessments40. Moreover, phytoplankton exhibit intrinsic diel periodicity42 that can be observed through geostationary Earth-observing satellites. Regional proof-of-concept studies have exploited this orbital class of satellites to observe diel changes in phytoplankton biomass to infer growth rates43 and model NPP with estimates that compared favorably to in-situ measurements44. Looking beyond satellites, this study has shown that the global fraction of NPP beneath the surface mixed layer and beyond the reach of passive ocean color sensors is increasing. Thus, the growing importance of deep NPP demonstrates that additional measurement platforms (e.g., autonomous profiling floats), large-scale field surveys, and numerical models are essential to fully characterize and understand how climate change is impacting NPP and related global biogeochemical cycles.
Methods
Data sources
Absorption-based NPP is calculated using the CAFE model19. Level 3, monthly, 1/12° ocean color data (2022 reprocessing version) were downloaded from the NASA ocean color webpage for the SeaWiFS (09/1997–12/2007) and MODIS Aqua (07/2002–12/2022) missions. Inherent optical properties are determined via inversion of remote-sensing reflectance using the generalized inherent optical property model18. The CAFE model is spectrally explicit and evaluated at 10 nm increments in the visible spectrum (400–700 nm). Monthly MLD were retrieved from the Oregon state ocean productivity website (https://sites.science.oregonstate.edu/ocean.productivity/). The MLD is calculated as the depth where the density of water is 0.125 kg m−3 greater than the density of near-surface water (z = 10 meters), where density is derived from HYCOM salinity and temperature data. Monthly 1/12° SST data are from the optimum interpolation sea surface temperature advanced very high resolution radiometer version 2.145.
Satellite blending
To account for persistent biases between SeaWiFS and MODIS Aqua missions, we compared NPP from collocated monthly 1/12° data for the period 2002–2007. Overall, NPP showed remarkable correspondence between satellite missions (Supplementary Fig. 5); Type-1 linear regression returned an r value and slope of 0.96 and 0.97, respectively, and a mean absolute error of 1.09. That said, MODIS Aqua generally had higher NPP (Supplementary Fig. 5), therefore to accommodate for persistent spatial biases we followed the general convention of inter-satellite comparisons17,46: SeaWiFS data were adjusted (SeaWiFS*) by the monthly climatology differences derived from overlapping periods between the two missions. This adjustment resulted in an r value and slope of 0.98 and 0.99, respectively, and mean absolute error of 1.04. Satellite data (e.g., NPP, IOPs) were averaged between SeaWiFS* and MODIS-Aqua for the 2002–2007 overlap.
Statistical and spatial analysis
For all time series, monthly anomalies were first calculated by subtracting monthly data from monthly climatology (n = 303). All statistics were performed using the Python package SciPy47. Spatially explicit trends and residuals were computed using ordinary least-squares regression. Serial autocorrelation in anomaly residuals was quantified using a first-order model (i.e., monthly) that reduced the effective sample size of the time series. The standard error of the slope was recomputed using the reduced effective sample size. Uncertainties are expressed as 90% two-tailed confidence intervals derived from the effective sample size and recomputed standard error of the slope. EOF were derived after downsampling the data to an equal area 50 × 50 km grid. The full list of climate and oceanographic indices and their sources that were compared to the principal components are given in Supplementary Table 1. Gap filling of data using monthly climatological data was only performed to compute global NPP totals (Fig. 4a). On average less than 2% of ocean surface area where PAR > 1 mol photons m−2 day−1 required gap filling. All spatial analyses account for areal changes in pixel size and apply a global land mask created for satellite ocean color data48.
Data availability
Monthly global CAFE model output is available at https://doi.org/10.5281/zenodo.12586956. Remotely sensed data are available at https://oceancolor.gsfc.nasa.gov/l3/. Monthly global MLD data are available at http://orca.science.oregonstate.edu/2160.by.4320.monthly.hdf.mld030.hycom.php. Data generated in Figs. 1 through 5 are available at https://doi.org/10.5281/zenodo.15497141. All other data that support the findings of this study are available from the corresponding author upon reasonable request.
Code availability
All major codes are available at doi.org/10.5281/zenodo.15497980. This repository includes Phyton and C code to compute NPP and associated data, and Python scripts for the trend and EOF analysis presented here.
References
Johnson, G. C. & Lyman, J. M. Warming trends increasingly dominate global ocean. Nat. Clim. Change 10, 757–761 (2020).
Li, G. et al. Increasing ocean stratification over the past half-century. Nat. Clim. Change 10, 1116–1123 (2020).
Field, C. B., Behrenfeld, M. J., Randerson, J. T. & Falkowski, P. Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281, 237–240 (1998).
Stock, C. A. et al. Reconciling fisheries catch and ocean productivity. Proc. Natl. Acad. Sci. 114, E1441–E1449 (2017).
Boyd, P. W., Claustre, H., Levy, M., Siegel, D. A. & Weber, T. Multi-faceted particle pumps drive carbon sequestration in the ocean. Nature 568, 327–335 (2019).
Falkowski, P. G., Lin, H. & Gorbunov, M. Y. What limits photosynthetic energy conversion efficiency in nature? Lessons from the oceans. Philos. Trans. R. Soc. B Biol. Sci. 372, 20160376 (2017).
Moore, C. M. et. Al. Processes and patterns of oceanic nutrient limitation. Nat. Geosci. 6, 701–710 (2013).
Doney, S. C. Plankton in a warmer world. Nature 444, 695–696 (2006).
Dutkiewicz, S. et al. Ocean colour signature of climate change. Nat. Commun. 10, 578 (2019).
Lomas, M. W., Bates, N. R., Johnson, R. J., Steinberg, D. K. & Tanioka, T. Adaptive carbon export response to warming in the Sargasso Sea. Nat. Commun. 13, 1211 (2022).
Karl, D. M. et al. Seasonal-to-decadal scale variability in primary production and particulate matter export at Station ALOHA. Prog. Oceanogr. 195, 102563 (2021).
Tagliabue, A. et al. Persistent uncertainties in ocean net primary production climate change projections at regional scales raise challenges for assessing impacts on ecosystem services. Front. Clim. 3, 738224 (2021).
Kulk, G. et al. Primary production, an index of climate change in the ocean: satellite-based estimates over two decades. Remote Sens. 12, 826 (2020).
Lee, Z., Marra, J., Perry, M. J. & Kahru, M. Estimating oceanic primary productivity from ocean color remote sensing: a strategic assessment. J. Mar. Syst. 149, 50–59 (2015).
Graff, J. R. et al. Analytical phytoplankton carbon measurements spanning diverse ecosystems. Deep Sea Res. Part I Oceanograph. Res. Pap. 102, 16–25 (2015).
Behrenfeld, M. J. et al. Satellite-detected fluorescence reveals global physiology of ocean phytoplankton. Biogeosciences 6, 779–794 (2009).
Mouw, C. B., Ciochetto, A. B. & Yoder, J. A. A satellite assessment of environmental controls of phytoplankton community size structure. Glob. Biogeochem. Cycles 33, 540–558 (2019).
Werdell, P. et al. Generalized ocean color inversion model for retrieving marine inherent optical properties. Appl. Opt. 52, 2019–2037 (2013).
Silsbe, G. M., Behrenfeld, M. J., Halsey, K. H., Milligan, A. J. & Westberry, T. K. The CAFE model: A net production model for global ocean phytoplankton. Glob. Biogeochem. Cycles 30, 1756–1777 (2016).
Babin, M. et al. Nitrogen-and irradiance-dependent variations of the maximum quantum yield of carbon fixation in eutrophic, mesotrophic and oligotrophic marine systems. Deep Sea Res. Part I Oceanograph. Res. Pap. 43, 1241–1272 (1996).
Marra, J., Trees, C. C. & O’Reilly, J. E. Phytoplankton pigment absorption: a strong predictor of primary productivity in the surface ocean. Deep Sea Res. Part I Oceanograph. Res. Pap. 54, 155–163 (2007).
Halsey, K. H., O’Malley, R. T., Graff, J. R., Milligan, A. J. & Behrenfeld, M. J. A common partitioning strategy for photosynthetic products in evolutionarily distinct phytoplankton species. N. Phytol. 198, 1030–1038 (2013).
Jakob, T., Wagner, H., Stehfest, K. & Wilhelm, C. A complete energy balance from photons to new biomass reveals a light-and nutrient-dependent variability in the metabolic costs of carbon assimilation. J. Exp. Bot. 58, 2101–2112 (2007).
Behrenfeld, M. J. et al. Revaluating ocean warming impacts on global phytoplankton. Nat. Clim. Change 6, 323–330 (2016).
MacIntyre, H. L., Kana, T. M., Anning, T. & Geider, R. J. Photoacclimation of photosynthesis irradiance response curves and photosynthetic pigments in microalgae and cyanobacteria 1. J. Phycol. 38, 17–38 (2002).
Behrenfeld, M. J., Halsey, K. H. & Milligan, A. J. Evolved physiological responses of phytoplankton to their integrated growth environment. Philos. Trans. R. Soc. B Biol. Sci. 363, 2687–2703 (2008).
Westberry, T., Behrenfeld, M. J., Siegel, D. A. & Boss, E. Carbon-based primary productivity modeling with vertically resolved photoacclimation. Glob. Biogeochem. Cycles 22, GB2024 (2008).
Cael, B. B. et al. Global climate-change trends detected in indicators of ocean ecology. Nature 619, 551–554 (2023).
Chavez, F. P., Messié, M. & Pennington, J. T. Marine primary production in relation to climate variability and change. Annu. Rev. Mar. Sci. 3, 227–260 (2011).
Behrenfeld, M. J. & Falkowski, P. G. Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnol. Oceanogr. 42, 1–20 (1997).
Cullen, J. J. Subsurface chlorophyll maximum layers: enduring enigma or mystery solved? Annu. Rev. Mar. Sci. 7, 207–239 (2015).
Sallée, J. B. et al. Summertime increases in upper-ocean stratification and mixed-layer depth. Nature 591, 592–598 (2021).
Cheng, L. et al. New record ocean temperatures and related climate indicators in 2023. Adv. Atmos. Sci. 41, 1068–1082 (2024).
Kwiatkowski, L., Aumont, O. & Bopp, L. Consistent trophic amplification of marine biomass declines under climate change. Glob. Change Biol. 25, 218–229 (2019).
Stock, C. A., Dunne, J. P. & John, J. G. Drivers of trophic amplification of ocean productivity trends in a changing climate. Biogeosciences 11, 7125–7135 (2014).
Stevens, S. W., Johnson, R. J., Maze, G. & Bates, N. R. A recent decline in North Atlantic subtropical mode water formation. Nat. Clim. Change 10, 335–341 (2020).
Kwiatkowski, L. et al. Twenty-first century ocean warming, acidification, deoxygenation, and upper-ocean nutrient and primary production decline from CMIP6 model projections. Biogeosciences 17, 3439–3470 (2020).
Kim, I. N. et al. Increasing anthropogenic nitrogen in the North Pacific Ocean. Science 346, 1102–1106 (2014).
Cornec, M. et al. Deep chlorophyll maxima in the global ocean: occurrences, drivers and characteristics. Glob. Biogeochemical Cycles 35, e2020GB006759 (2021).
Westberry, T. K., Silsbe, G. M. & Behrenfeld, M. J. Gross and net primary production in the global ocean: an ocean color remote sensing perspective. Earth-Sci. Rev. 237, 104322 (2023).
Dierssen, H. M. et al. Synergies between NASA’s hyperspectral aquatic missions PACE, GLIMR, and SBG: Opportunities for new science and applications. J. Geophys. Res. Biogeosci. 128, e2023JG007574 (2023).
Prézelin, B. B. Diel periodicity in phytoplankton productivity. In: Proc. The Daily Growth Cycle of Phytoplankton: Proceedings of the Fifth International Workshop of the Group for Aquatic Primary Productivity (GAP), held at Breukelen, The Netherlands 20–28 April 1990 (pp. 1-35). (Springer, 2023).
Salisbury, J. E. et al. Assessing net growth of phytoplankton biomass on hourly to annual time scales using the geostationary ocean color instrument. Geophys. Res. Lett. 48, e2021GL095528 (2021).
Wu, J. et al. Estimates of diurnal and daily net primary productivity using the Geostationary Ocean Color Imager (GOCI) data. Remote Sens. Environ. 280, 113183 (2022).
Huang, B. et al. Improvements of the daily optimum interpolation sea surface temperature (DOISST) version 2.1. J. Clim. 34, 2923–2939 (2021).
Sathyendranath, S. et al. An ocean-colour time series for use in climate studies: the experience of the ocean-colour climate change initiative (OC-CCI). Sensors 19, 4285 (2019).
Virtanen, P. et al. SciPy 1.0: fundamental algorithms for scientific computing in Python. Nat. Methods 17, 261–272 (2020).
Mikelsons, K., Wang, M., Wang, X. L. & Jiang, L. Global land mask for satellite ocean color remote sensing. Remote Sens. Environ. 257, 112356 (2021).
Acknowledgements
The authors would like to acknowledge funding from NASA grants 80NSSC21K0421, 80NSSC17K0560.
Author information
Authors and Affiliations
Contributions
G.M.S.: data analysis, visualization, original draft, review, and editing. J.F.: conceptualization, data analysis, review, and editing. T.K.W.: conceptualization, review and editing. K.H.H.: review and editing.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Stephanie Henson and Chih-hao Hsieh for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Silsbe, G.M., Fox, J., Westberry, T.K. et al. Global declines in net primary production in the ocean color era. Nat Commun 16, 5821 (2025). https://doi.org/10.1038/s41467-025-60906-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-60906-y
This article is cited by
-
Quantifying coral reef–ocean interactions is critical for predicting reef futures under climate change
Nature Ecology & Evolution (2025)
-
Enhanced North Pacific biological pump contributed to atmospheric carbon dioxide plateau during the Bølling-Allerød period
Communications Earth & Environment (2025)
