
Abstract
In the pre-synapse, vesicle fusion is a crucial process for neurotransmitter release in the nervous system. Under physiological conditions, synaptotagmin-1 (Syt1) locks synaptic vesicles in a priming state, allowing them to undergo synchronized neurotransmitter release upon Ca2+ activation. Elevation of intracellular ions during diseases or injuries may lead to uncontrolled neurotransmitter release independent on Ca2+ influx. However, its underlying molecular mechanism remains elusive. Here we show that elevation of the intracellular Zn2+ concentration leads to the increased frequency of spontaneous neurotransmitter release in hippocampal neurons. In the reconstituted system with neuronal SNAREs and Syt1, Zn2+ markedly enhances the fusion efficiency of liposomes via its binding to the interface between tandem C2 domains of Syt1. Moreover, Syt1 exhibits an elevated capacity to bind to anionic vesicles in the context of interaction with Zn2+, which leads to an augmentation in vesicles docking within the synaptic active zone. Finally, the mutation of the tentative Zn2+-binding site (Syt1_3M) results in a loss of activation function in spontaneous release by Zn2+, while disruption of the primary interface and the polybasic region show negligible impact on the modulatory action of Zn2+. Thus, these findings suggest that with Zn2+, Syt1 adopts an alternative regulatory mode to drive spontaneous neurotransmitter release.
Similar content being viewed by others
Introduction
Neurons engage in communications through the release of neurotransmitters into the synaptic gap, a process that is achieved by the highly specific task of orchestrating synaptic vesicle fusion with the plasma membrane. Proteins that control synaptic vesicular exocytosis include SNAREs (soluble N-ethylmaleimide-sensitive factor attachment protein receptors), synaptotagmin-1 (Syt1) and other regulators, which composite the key determinants of the specificity of synaptic neurotransmission1,2,3,4.
At the synapses, the predominant release occurs during the synchronous phase, a critical phenomenon orchestrated by Syt1 in response to Ca2+ influx5. Syt1 comprises of two cytoplasmic C2 domains—C2A and C2B. Both domains exhibit analogous structures and bind three and two Ca2+ ions, respectively, through the negatively charged residues situated at the apexes of the structures5,6,7. Mutations in the Ca2+-binding sites significantly impede the process of synchronous release in synapses8,9. Upon Ca2+ binding, hydrophobic residues in the loops facilitate Syt1 insertion into the anionic membrane10, culminating in the synchronized release of neurotransmitters. Moreover, Syt1 establishes connections with syntaxin-1A (Stx1A) and SNAP-25 in the SNARE complex, through a conserved primary interface (encompassing residues R281, E295, Y338, R398/399) located on its C2B domain11, which is indispensable for the triggering function of Syt1. Meanwhile, the polybasic region (specifically Lys326 and Lys327) of the C2B domain binds to acidic lipids, including PS and PI(4,5)P2, forming a Syt1-SNARE-membrane super-structure12,13 and its conformational change upon Ca2+ binding facilitates the initiation of synchronized release in response to Ca2+.
The binding of Syt1 to the membrane in the presence of Ca2+ aligns more cogently with the temporal dynamics of membrane fusion between synaptic vesicles and the plasma membrane, occurring in < 1 ms post Ca2+ influx2,14,15. Nonetheless, the explorations into the role of metal ions in triggering exocytosis have extended beyond the confines of Ca2+. An assessment of the total energy required to dissociate Syt1 from the membrane in the presence of Mg2+ and Ca2+ revealed that the binding lifetime of Syt1 with the Mg2+-bound membrane is comparatively short, allowing for the separation of Syt1 oligomers and subsequent binding of Ca2+. This, in turn, promotes a more protracted binding life of Ca2+16. Other divalent metal ions were also postulated to partake in the intricate process of neurotransmitter release. Sr2+, for instance, exhibits sufficient affinity for the C2B domain, leading to the initiation of rapid release, albeit with diminished efficacy17. In addition, Ba2+ demonstrates the capability to induce acetylcholine secretion, yet its effectiveness in instigating exocytosis in bag cell neurons is notably limited18.
Prior to Ca2+ influx, spontaneous release is prevented and maintained at a basal level by Syt1 via several super-structures: the SNARE/complexin1 (Cpx1)/Syt1 tripartite complex19 and the Syt1 oligomers20,21. Upon Ca2+ binding, these superstructures may undergo a conformational change and release the spontaneous fusion clamp13,19,22.
In the central nervous system, Zn2+, as a mediator in cell–cell signaling, plays an important role in cognition and long-term synaptic plasticity. Synaptic Zn2+ concentrations, maintained within the nanomolar range at rest, undergo a substantial increase to potentially reach sub-micromolar intraneuronal toxic levels during sustained synaptic activity23,24. The pathophysiological relevance is underscored by the selective accumulation of chelatable endogenous Zn2+ in damaged neurons, particularly rising in neuronal cell bodies subsequent to events such as seizures25,26, transient forebrain ischemia25,27,28, or trauma28,29,30. Moreover, emerging evidences indicate that Zn2+ has possesses the potential to intricately modulate synaptic transmission by influencing neurotransmitter release, vesicle recycling, and the structural aspects of the presynaptic environment in temporal lobe epilepsy31,32. Alteration in the Zn2+ levels is postulated to have relevance to the function of neuronal networks where synapses enriched with Zn2+8. These findings strongly imply that the accumulation of Zn2+ may substantively contribute to the causal mechanisms underlying irregular neurotransmission. Despite these insights, the detailed mechanism underlying the neurotoxicity of Zn2+ remains elusive.
In this work, using the patch-clamp techniques to record neurotransmitter release from hippocampal neurons, we observed that elevation of intracellular Zn2+ concentration significantly increased the frequency of miniature excitatory postsynaptic currents (mEPSCs) without affecting their amplitude. In in vitro reconstituted fusion systems with neuronal SNAREs and Syt1, Zn2+ significantly improved the fusion probability in both the lipid and single vesicle content-mixing assays. Moreover, Zn2+ could bind to the putative Zn2+-binding pocket composed of the residues of D261, E386, and H389 within the interface between tandem C2 domains of Syt1 with an affinity of hundreds nanomole. Upon binding to Zn2+, Syt1 exhibits an enhanced anionic membrane-binding ability, which in turn resulted in an enhancement effect on vesicle docking at synaptic active regions. Finally, mutations at the Zn2+-binding sites (Syt1_3M) led to the loss of the activation function in spontaneous release and Zn2+-dependent vesicle fusion, while the disruption of the primary interface and the polybasic region of Syt1 only had a negligible effect on the fusion probability in the reconstituted fusion assay in the presence of Zn2+. Thus, these outcomes suggest the distinctive regulatory mechanism of Syt1/Zn2+ in neurotransmitter release under pathological conditions, providing an inception for a deeper exploration into the repercussions of Zn2+ on neurological disorders.
Results
Zn2+ promotes mEPSC in cultured neurons
SNARE/Syt1-mediated synaptic vesicle fusion with the plasma membrane is an essential process for neurotransmitter release in the nervous system. To investigate the impact of Zn2+ on neurotransmitter release, we first examined spontaneous excitatory postsynaptic currents (sEPSC) in hippocampal neurons. Upon adding 0–200 µM Zn2+ and 2 mM Ca2+ into the extracellular fluid of hippocampal cells (Fig. 1a), we found that the including of Zn2+ (50–200 µM) in the standard 2 mM Ca2+-containing bath solution increased the frequency of sEPSC (Supplementary Fig. S1a, b) in a dose-dependent manner, but showed no effect on the amplitude of these signals (Supplementary Fig. S1c). Although a modest increase of extracellular Zn2+ concentration to 50 µM led to elevated spontaneous release, the excessive of extracellular Zn2+ to 500 µM, which might cause neuronal toxicity, in turn resulted in the loss of neuronal activity (Supplementary Fig. S1b, c). To verify if the abnormal spontaneous transmission was due to the altered neural excitability, we further recorded the miniature excitatory postsynaptic currents (mEPSC) in hippocampal neurons (Fig. 1) and hippocampal slices (Supplementary Fig. S2). Zn2+ also led to a dose-dependent increase in the frequency (Fig. 1b, c and Supplementary Fig. S2a, b) but not amplitude (Fig. 1d and Supplementary Fig. S2c) of mEPSC, suggesting an increase in the release probability of synaptic vesicles during resting conditions. In contrast, the presence of Zn2+ showed no effect on the amplitude of EPSC (Fig. 1e, f). Note that, when 2 mM Ca2+ was removed from the standard bath solution, although Zn2+ still increased the frequency but not the amplitude of mEPSC (Supplementary Fig. S3a–c), it could not mediate EPSC in the absence of Ca2+ (Supplementary Fig. S3d, e). Thus, these results show that elevation of intracellular Zn2+ leads to a specific increase in the frequency of spontaneous release during the synaptic transmission of hippocampal neurons.

a Experimental scheme: different concentrations of Zn2+ were added to the culture of hippocampal cells, and the changes of postsynaptic current signals were recorded by the patch-clamp technique. b–f Cultured hippocampal neurons with Syt1_WT in the electrophysiological experiments. b Representative traces of mEPSC in the presence of 0, 50, 100, 200 μM Zn2+. c Corresponding mEPSC frequency of spontaneous release in panel (b). d Corresponding amplitude of spontaneous release in panel (b). e Representative traces of action potential-evoked EPSCs in the presence of 0, 50, 100, 200 μM Zn2+. f Corresponding amplitude of evoked release in panel (e). g–i Cultured hippocampal neurons infected with lentiviruses encoding cre-EGFP in the electrophysiological experiments. g Representative traces of mEPSC in the presence of 0, 50, 100, 200 μM Zn2+. h Corresponding mEPSC frequency of spontaneous release in panel (g). i Corresponding amplitude of spontaneous release in panel (g). All traces were recorded in 1 μM tetrodotoxin (TTX) and 0.1 mM PTX. In panels (c, d, f, h, i), bargraphs and data points are shown for n (indicated below each bargraph) independent repeat cells. Statistical analysis was performed using one-way ANOVA followed by Tukey’s multiple comparisons test. Source data are provided as a Source Data file.
Syt1 is an important regulator for controlling the frequency of spontaneous release8,33. To test if Syt1 serves as a primary sensor of Zn2+ to mediate the modulation of spontaneous release, we next performed Syt1 knockout (KO) by infecting hippocampal neurons from homozygous floxed Syt1-null mice with synapsin-driven Cre recombinase-expressing lentivirus. Consistent with the previous report8, Syt1 cKO resulted in an increase in the frequency of mEPSCs (Fig. 1g) and sEPSC (compare the conditions of “0 µM” in Supplementary Figs. S1 and S4). However, the augmented effect of Zn2+ in spontaneous release was impeded (Fig. 1h, i). Consistently, Zn2+ had little effect on the frequency of sEPSC in Syt1 cKO hippocampal neurons (Supplementary Fig. S4a–c). These findings demonstrate that Syt1 plays an essential role in mediating Zn2+-induced spontaneous release in hippocampal neurons.
Zn2+ activates Syt1/SNARE-mediated vesicle fusion via a different molecular mechanism from Ca2+
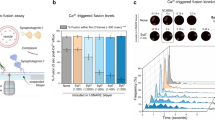
In our next pursuit to explore the molecular mechanism of Zn2+ in regulating SNARE/Syt1-mediated neurotransmitter release, we utilized the single vesicle content-mixing assay with reconstituted full-length neuronal SNAREs and Syt134. In this assay, we reconstituted two types of vesicles in-vitro: the plasma membrane-mimicking vesicles reconstituted with Stx1A and SNAP-25 (referred as PM-vesicle), and synaptic vesicle-mimicking vesicle containing sulforhodamine B (SRB) reconstituted with VAMP2 and Syt1 (referred as SV-vesicle) (Fig. 2a). As described previously34,35, PM-vesicles were immobilized on the PEG passive surface via the biotin-neutravidin (NTV) interaction, then SV-vesicles were added and associated with PM-vesicles (Fig. 2a). Fusion probabilities were calculated by the total fusion events divided by the corresponding number of associated SV-PM vesicle pairs. When different concentrations of Zn2+ were loaded into this system, the fusion probability exhibited a notable augmentation trend with the increased Zn2+ concentration (Fig. 2d, e). As controls, other divalent cations such as Mg2+ and Sr2+ did not induce an increase in the fusion probability, indicating a specificity of Zn2+-dependent enhancement on SNARE/Syt1-mediated membrane fusion (Fig. 2b, c). When either SNAP-25 or VAMP2 was left out, fusion did not occur even in the presence of 500 µM Zn2+. Similarly, when Syt1 was removed, Zn2+ was incapable of driving membrane fusion with SNARE proteins alone (Supplementary Fig. S5a, b), excluding the possibility of the enhancement effect in the fusion assay attributing to the non-specific binding of Zn2+.

a Single-vesicle content-mixing assay using two different types of vesicles (SV-vesicle and PM-vesicle). Fusion was monitored by measuring the fluorescence intensity of SRB encapsulated in SV-vesicle. b Normalized histograms of the fusion probability in the presence of 500 μM ions. c Corresponding cumulative probability of per minute upon ion injection in panel (b). Statistical significance was assessed by a two-tailed Student’s t test, compared to the case with Sr2+. d Normalized histograms of the probability of Zn2+-dependent fusion at various concentrations. e Corresponding cumulative probability per min upon Zn2+ injection in panel (d). Statistical significance was assessed by a two-tailed Student’s t test, compared to the case with 0 μM Zn2+. f Normalized histograms of Zn2+-dependent fusion probability with Syt1_WT and its mutants (Syt1-C2A*B*, Syt1-4A and Syt1-4W). g Corresponding cumulative probability per min upon Zn2+ injection in panel (f). In panels (c, e, and g), box plots and data points are shown for n (indicated below each box plot) independent repeat experiments. Statistical significance was assessed by a two-tailed Student’s t test, compared to the case with Syt1_WT. Box plots are defined as follows: the whiskers show the minimum and maximum values (excluding outliers), the box limits show the 25% and 75% percentiles, the center line denotes the median. Source data are provided as a Source Data file.
Ensemble lipid-mixing assays with reconstituted SNARE proteins were used to further assess the probability of vesicle fusion with the FRET efficiency calculated from a lipidic donor–acceptor fluorescence pair (Supplementary Fig. S6a). In this assay, PM-vesicles and SV-vesicles were mixed and subjected to continuous monitoring for 1000 s, followed by the injection of Zn2+, Ca2+, Mg2+, or Sr2+ at 200 s. The results validated that both Ca2+ and Zn2+ ions could stimulate lipid-mixing that mediated by SNARE and Syt1, while Mg2+ or Sr2+ not (Supplementary Fig. S6b, c). Moreover, sequential loading of Ca2+ and Zn2+ revealed an additional enhancement by Zn2+ following the stimulation by Ca2+. This observation remained consistent when the order of adding Ca2+ and Zn2+ is reversed (Supplementary Fig. S7a, b). These results underscored that the regulatory mode of Zn2+ on Syt1 might be distinct from that of Ca2+.
To validate this hypothesis, we examined the impact of Zn2+ on vesicle fusion by mutating the Ca2+-binding sites of Syt1 (Syt1_C2A*B*) and four membrane-inserting residues upon binding to Ca2+ (Syt1_4A and Syt1_4W, corresponding residues were substituted with alanine or tryptophan, respectively, to change its membrane binding ability10) in a single-vesicle content-mixing assay. As expecting, Syt1_C2A*B* disrupted the stimulating effect of Ca2+ on the probability of vesicle fusion, while the fusion probabilities that mediated by Syt1_4A and Syt1_4W showed a parallel correlation with its membrane binding ability in the presence of Ca2+ (Supplementary Fig. S8a, b). In contrast, these Syt1 mutants, compared with Syt1_WT, did not alter the effect of Zn2+ on promoting the probability of vesicle fusion (Fig. 2g, f). Thus, the result suggests that Zn2+ has its unique binding site on Syt1, which may be mutually exclusive with the binding sites of Ca2+.
Zn2+ may bind to the interface between C2A and C2B domains with an affinity of hundreds of nanomole
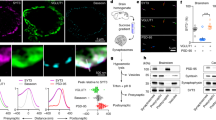
Next, we investigated the binding mode of Syt1 and Zn2+ using an in-vitro binding assay. In this experiment, the emission spectrum of the Zn2+ indicator FluoZin™-1 is proportional to the concentration of Zn2+ in solution. Upon the formation of complexes of Zn2+ and C2AB (a cytosolic region of Syt1), the concentration of Zn2+ in solution was reduced, which in turn resulted in a proportional attenuation in the fluorescence intensity of FluoZin™-1 (Fig. 3a). This result elucidated a consequential reduction in fluorescence intensity when the C2AB was present in the solution, but this attenuation diminished when employing C2AB pre-soaked with Zn2+ (Fig. 3b), indicating a propensity of C2AB to interact with Zn2+. Notably, the fluorescence intensity of the emission spectrum remained unchanged when C2A or C2B was added separately (Fig. 3c). This delineates a crucial dependence on the existence of both C2A and C2B domains for the binding of Zn2+.

a Schematic representation of the Zn2+ binding assay. In this experiment, 8 μM Zn2+ and 400 nM Zn2+ fluorescent indicator FluoZin™-1 were included in the buffer, unless specified. FluoZin™-1 was excited when free Zn2+ bound. The emission spectrum (500–600 nm) of FluoZin™-1 was obtained by using an excitation wavelength at 488 nm. b The emission spectrum of FluoZin™-1 in the absence or the presence of 2 μM C2AB. C2AB(Zn2+): C2AB was pre-soaked with Zn2+ solution. The data are shown as mean ± SEM for n independent repeat experiments (None, n = 3; C2AB, n = 3; C2AB(Zn2+), n = 3). c The emission spectrum of FluoZin™-1 in the presence of 2 μM C2AB fragment and its variants C2A and C2B. The data are shown as mean ± SEM for n independent repeat experiments (None, n = 3; C2AB, n = 3; C2A, n = 4; C2B, n = 3). d The sequence alignment of Syt3 and Syt1 at the region of the C2A-C2B interface. The residues involved in the binding of Zn2+ was colored in orange. Black highlights: absolutely conserved region, gray highlights: similar region. e Structure alignment of Syt1 (yellow) with its homolog protein Syt3 (gray) in the presence of Zn2+. The crystal structure of the C2AB fragment of Syt3 was derived from PDB IDs 3HN8. Zn2+ and Ca2+ were shown as blue and green sphere, respectively. f The emission spectrum of FluoZin™-1 in the presence of 2 μM C2AB fragment and its variants C2AB_3M and C2A*B*. The data are shown as mean ± SEM (None, n = 3; C2AB, n = 3; C2A*B*, n = 3; C2AB_3M, n = 4). g The averaged intensity of FluoZin™-1 with 2 μM Zn2+ at different concentrations of C2AB or C2AB_3M. In panel (g), box plots and data points are shown for n (indicated below each box plot) independent repeat experiments. The binding affinity between C2AB/C2AB_3M and Zn2+ was obtained by fitting the corresponding plots with Hill1 functions. Source data are provided as a Source Data file.
Previous results indicated that Syt3, a homolog protein of Syt1, bound to Zn2+ via an interface formed by C2A and C2B domains36. However, the predicted binding mode of Zn2+ to Syt1 by AlphaFold3 proposed that Zn2+ bound to the residues of D178, D232, D309, D363 that buried within the Ca2+-binding site of both C2 domains, and similar prediction results were also applied to Syt3 (Supplementary Fig. S9). We suspected that such a contradiction may result from the incapability of predicting the intramolecular conformation of C2A and C2B that linked by a flexible region in the presence of Zn2+. In addition, our results with the mutants of Syt1 have excluded the mode of Zn2+ binding to either the separated C2 domain or the Ca2+-binding sites of both C2 domains (Figs. 3c, f, 2f). Through a comparative analysis of the sequences and structure of Syt1 and Syt3, a putative binding site for Zn2+ in Syt1 was postulated as D261 on C2A, and E386 and H389 on C2B (Fig. 3d, e). Mutations of these residues (C2AB_3M) maintained the structure of C2AB in CD spectroscopy (Supplementary Fig. S10), but disrupted the binding between Zn2+ and Syt1 (Fig. 3f). As the control, the mutations on the Ca2+-binding sites (C2A*B*) did not interfere the binding of Zn2+ (Fig. 3f). Via a serial titration of C2AB or C2AB_3M in the fluorescence-based binding assay, we found that the binding affinity (~ 6.91 ± 1.08 µM) of C2AB_3M to Zn2+ exhibited > 25-folds increase compared to that (~ 259.78 ± 36.14 nM) of C2AB (Fig. 3g), validating that the Zn2+-binding pocket is distinct from that of Ca2+, and composed of three critical residues contributed by both C2A and C2B domains of Syt1.
The enhanced spontaneous release requires the binding of Zn2+ to Syt1
To further investigate the functional implications of Zn2+ binding to Syt1, we neutralized the Zn2+-binding sites on Syt1 (Syt1_3M) and reconstituted it onto SV-vesicles. In the single-vesicle content-mixing assays, the probability of Syt1_3M-mediated vesicle fusion was substantially decreased in the presence of 500 µM Zn2+, in comparison to that of Syt1_WT (Fig. 4a, b). At a lower concentration of 500 nM Zn2+, Syt1_WT was still capable of inducing a notable number of vesicle fusion events, while Syt1_3M failed to do that (Fig. 4a, b). In addition, in the ensemble lipid-mixing assay, Syt1_3M reduced the fusion percentage significantly at a serial of Zn2+ concentrations (ranging from 10 nM to 100 µM) (Supplementary Fig. S11a, b), but it did not impede the fusion percentage in the presence of Ca2+ (Supplementary Fig. S12a, b). Consistent with the results of the single-vesicle content-mixing assay, Syt1_WT elicited a robust enhancement of vesicle fusion in the presence of 1 µM Zn2+ (Supplementary Fig. S11a, b), suggesting that Zn2+ has the capacity to augment vesicle fusion at the physiological/pathological relevant concentrations23. To further characterize the specificity of the interaction between Syt1 and Zn2+, we included 2 mM Mg2+ in the lipid-mixing assay, which has an ionic shielding effect on certain cellular processes37,38. We found that the Zn2+-dependent enhancement of fusion percentage of Syt1-mediated lipid-mixing remained in the presence of 2 mM Mg2+ (Supplementary Fig. S11c, d), suggesting that the physiological bindings between Syt1 and Zn2+ are not due to non-specific electrostatic interactions. To validate that Syt1 serves as a primary Zn2+ sensor in mediating spontaneous neurotransmitter release, we next performed rescue experiments with Syt1_WT or Syt1_3M mutant in Syt1 cKO hippocampal neurons. As expected, 100 µM Zn2+ restored its ability to increase the frequency of mEPSC in Syt1_WT rescued hippocampal neurons without exerting an impact on the amplitude of the postsynaptic current (Fig. 4c–e). However, when the Syt1_3M mutant was used, Zn2+ had no discernable impact on either the frequency or the amplitude of mEPSC (Fig. 4f–h), and EPSC in Syt1_3M-expressing hippocampal neurons remained unchanged in response to the increased concentration of Zn2+ (Supplementary Fig. S13). Similarly, Zn2+ dose-dependently increased the frequency of sEPSC in Syt1_WT, but not Syt1_3M, rescued hippocampal neurons (Supplementary Fig. S14). Together, these findings support the notion that the binding of Zn2+ to Syt1 is required for the enhancement of synaptic spontaneous neurotransmitter release.

a Normalized histograms of the fusion probability with reconstituted neuronal SNAREs and Syt1_WT or its mutant Syt1_3M in the presence of 500 μM or 500 nM Zn2+. b Corresponding cumulative fusion probability per min upon ion injection in panel (a). In panel b, box plots and data points are shown for n (indicated below each box plot) independent repeat experiments. Statistical significance was assessed by a two-tailed Student’s t test, compared to the case with Syt1_WT. Box plots are defined as follows: the whiskers show the minimum and maximum values (excluding outliers), the box limits show the 25% and 75% percentiles, and the center line denotes the median. c–h The spontaneous release of cultured Syt1 cKO hippocampal neurons rescued by Syt1_WT or Syt1_3M in the electrophysiological experiments. Cultured hippocampal neurons infected with lentiviruses both encoding cre-EGFP and Syt1_WT-mCherry, or Syt1_3M-mCherry. c Representative traces of mEPSC with cKO Syt1_WT expressed neurons in the presence of 0, 50, 100, 200 μM Zn2+. d Corresponding mEPSC frequency of spontaneous release in panel (c). e Corresponding amplitude of spontaneous release in panel (c). f Representative traces of mEPSC with cKO Syt1_3M expressed neurons in the presence of 0, 50, 100, 200 μM Zn2+. g Corresponding mEPSC frequency of spontaneous release in panel (f). h Corresponding amplitude of spontaneous release in panel (f). All traces were recorded in 1 μM TTX and 0.1 mM PTX. In panels (d, e, g, h), bargraphs and data points are shown for n (indicated below each bargraph) independent repeat cells. Statistical significance was assessed by one-way ANOVA followed by Tukey’s multiple comparisons test. Source data are provided as a Source Data file.
The binding of Zn2+ to Syt1 facilitates its membrane-binding ability
Intensive studies have revealed that Syt1 binding to negatively charged lipids has a functional consequence on SNARE-mediated synaptic vesicle fusion2,9,12,13. The positively charged residues surrounding the Zn2+-binding pocket endow Syt1 an ability to bind the anionic membrane (Fig. 5a, b). To test our hypothesis, we assessed the membrane binding between vesicles and Syt1, or its mutants through a co-sedimentation assay in the presence of a serial concentrations of Zn2+ (Fig. 5c). In this assay, C2AB and vesicle membrane in the fractions of precipitant and supernatant were quantified by SDS-PAGE analysis and the emission spectrum analysis of the fluorphore DiI, respectively (Fig. 5c).

a The electrostatic potential map of the Zn2+-binding pocket of C2AB. b Model of C2AB binding to membrane. The dash line indicates negative charged membrane. c The diagram shows the vesicle sedimentation assay. Details were shown in Methods. d SDS-PAGE gel analysis of C2AB or C2AB_3M in the fraction of supernatant (S) and precipitant (P) at different conditions of lipid components and Zn2+ concentrations as indicated. e Quantification the P/S ratio of C2AB or C2AB_3M in panel (d). The P/S ratio of protein at 0 μM Zn2+ was set as 1. f The fluorescence intensity of DiI-containing vesicles in the fraction of precipitant. Each data point represents the intensity of the emission spectrum of DiI at 570 nm from one independent experiment, as shown at the top-right corner of each plot. In panels (e and f), box plots and data points are shown for n (indicated below each box plot) independent repeat experiments. Statistical significance was assessed by one-way ANOVA followed by Tukey’s multiple comparisons test. Box plots are defined as follows: the whiskers show the minimum and maximum values (excluding outliers), the box limits show the 25% and 75% percentiles, and the center line denotes the median. Source data are provided as a Source Data file.
By changing the concentration of PS on the vesicle membrane from 0 to 15%, we observed a robust increase of C2AB fragment in the fraction of precipitant in the presence of Zn2+ (Fig. 5d, e), as well as the fluorescence intensity of DiI in the vesicle membrane (Fig. 5f). Specifically, Zn2+ dose-dependently promotes the binding between C2AB and vesicles that exhibited a significant increase when the Zn2+ concentration reached to 100 μM (Fig. 5d–f). Moreover, when an additional 1% PIP2, whose headgroup is highly negatively charged, was included in the vesicle membrane, further increase of C2AB and lipids in the precipitant fraction was observed (Fig. 5d–f). And the significant increases were achieved in response to both 10 μM and 100 μM Zn2+ (Fig. 5d–f). However, the mutations on the Zn2+-binding sites of C2AB (C2AB_3M) led to the loss function of membrane binding in the presence of Zn2+(Fig. 5d–f), suggesting that Zn2+ binding to the pocket of the C2A-C2B interface is indispensable for the Zn2+-dependent membrane binding of Syt1. As controls, the mutants of C2A*B* that disrupt the Ca2+-binding site of C2AB diminished the precipitant fraction of C2AB and lipids that induced by Ca2+ (Supplementary Fig. S15a–c), but not Zn2+ (Supplementary Fig. S16a–c). The mutants of C2AB_QQQ that disrupted the polybasic region of C2AB showed little effect on either the Ca2+-dependent (Supplementary Fig. S15a–c) or Zn2+-dependent precipitant of C2AB and lipids (Supplementary Fig. S16a–c). In summary, our results revealed that the binding of Zn2+ enables Syt1 with a strong capability to bind to anionic membranes in the absence of Ca2+.
Zn2+/Syt1 promotes synaptic vesicle docking via an alternative mode
To further investigate the mechanism underlying the Zn2+-stimulated Syt1/SNARE-dependent vesicle fusion, and the functional consequence of the Zn2+-dependent membrane binding by Syt1, we next performed the real-time total internal reflection fluorescence microscopy (TIRF) imaging of synaptophysin–pHluorin (Syp–pH) to visualize the docked vesicles in hippocampal nerve terminals (Fig. 6a). Docked presynaptic vesicles were identified as fluorescent puncta that were de-quenched by the 50 mM NH4Cl neutralization and illuminated within the evanescent field of TIRF microscopy (within 100 nm nearby the plasma membrane). The intensity of fluorescent puncta of synaptic vesicles during NH4Cl perfusion was referred as I0, while that obtained in the presence of 200 µM Zn2+ was defined as IZn2+ (Fig. 6b, c). When Zn2+ was perfused into the standard extracellular solution, a notable amount of Zn2+ was translocated into pre-synapse as indicated by Zinpyr-1 (Supplementary Fig. S17), and an increase of the ratio of the fluorescence intensity (IZn2+/I0) was observed (Fig. 6d), suggesting the increased docking of synaptic vesicles in individual presynaptic boutons. Importantly, the addition of Zn2+ was incapable of promoting vesicle docking in Syt1 cKO hippocampal synapses (Fig. 4b, c), suggesting that the increased occurrence of SV docking upon the influx of Zn2+ was strictly relied on Syt1.

a The scheme of the synaptic vesicle docking experiment by TIRF microscopy. Hippocampal cells were transiently transfected with a synaptophysin–pHluorin construct. The fluorescence intensity within the evanescent field was detected in the presence or the absence of 200 μM Zn2+. b The representative images of hippocampal axons expressing synaptophysin–pHluorin in the presence or the absence of 200 μM Zn2+. Upper: the hippocampal cells expressing Syt1_WT; bottom: the hippocampal cells with Syt1 knockout (Syt1 cKO). The red circles indicate the area of axons applied for the fluorescence intensity analysis. Scale bar: 2 μm. c Time-lapse of the intensity at sthe pecified area of hippocampal neurons as indicated in panel (b). The maximal fluorescence intensity in ROI during NH4Cl perfusion was defined as I0, after injection of 200 μM Zn2+, the maximal fluorescence intensity during NH4Cl perfusion was defined as IZn2+. d Corresponding box plots of the ratio of fluorescence intensities (IZn2+/I0) in panel (b). In panel (d), box plots and data points are shown for n (indicated below each box plot) ROIs, which were selected from 4 images/3 cultures. Statistical significance was assessed by a two-tailed Student’s t test, compared to the case with Syt1_WT. Box plots are defined as follows: the whiskers show the minimum and maximum values (excluding outliers), the box limits show the 25% and 75% percentiles, the center line denotes the median. e Representative TIRF microscopic images of docked SV-vesicle reconstituted with VAMP2 and Syt1_WT or its mutants in the absence or the presence of 500 μM Zn2+. f The bargraph shows the number of docked vesicles from more than 10 randomly chosen fields of view in the sample chamber, as shown in panel (e). In panel (f), bargraphs and data points are shown for n (indicated below each bargraph) independent repeat experiments. Statistical significance was assessed by a two-tailed Student’s t test, compared to the case with 0 μM Zn2+. Source data are provided as a Source Data file.
To validate critical roles of Syt1/Zn2+ in mediating the docking of synaptic vesicles, we next conducted a single-vesicle docking assay, in which unlabeled PM-vesicles were immobilized on the biotin-PEG-coated surface, and the SRB-containing SV-vesicles reconstituted with Syt1_WT or its mutants were load into the flow chambers, allowing vesicle docking in the absence or the presence of Zn2+. The results revealed that Zn2+ significantly promotes the docking of Syt1_WT vesicles by 3-fold, but the Syt1_3M mutant that disrupts the Zn2+-binding sites diminished Zn2+-dependent vesicle docking (Fig. 6e, f). In contrast, the mutants of Syt1_QM (a variant that disrupting the primary interface with the SNARE complex) and Syt1_QQQ both impaired SNARE/Syt1-mediated synaptic vesicle docking under Zn2+-free conditions, yet retained the ability of promoting vesicle docking upon Zn2+ binding (Fig. 6e, f). Consistent with this, Syt1_WT and its mutants Syt1_QM, Syt1_QQQ exhibited enhancement effects on the fusion probability in the presence of Zn2+ (Supplementary Fig. S18), while Syt1_3M was incapable of promoting the Zn2+-dependent vesicle fusion (Supplementary Fig. S18a, b). Taken together, these results suggest that the binding between Syt1 and Zn2+ facilitates synaptic vesicle docking and thus leads to the enhancement of spontaneous neurotransmitter release during synaptic transmission.
Discussion
Previous studies in situ have demonstrated that the raise of the endogenous Zn2+ concentration in damaged neurons caused by seizures, transient forebrain ischemia, or trauma, may lead to abnormal neurotransmitter release39,40,41, which is considered to be crucial to Zn2+ toxicity. Syt1, as a key fusogenic protein, plays a central role in mediating neuronal transmission under physiological conditions. Dysfunction of Syt1 results in abnormality of neurotransmitter release, including the impaired Ca2+-evoked release and the over-activation of spontaneous release8,42. However, the lack of detailed structural information of Syt1 under pathological conditions, in combination with the severity of pathological phenotype, has hindered a detailed description of the molecular mechanism of Syt1 in abnormal neurotransmitter release. Here, by combining cell-based electrophysiological recordings and the single-vesicle fusion assay with reconstituted proteins, we have defined Syt1 as a primary Zn2+ sensor, which can Zn2+-dependently mediate spontaneous synaptic transmission. Zn2+ dose-dependently promotes both the fusion efficiency of liposomes and spontaneous neurotransmitter release in hippocampal neurons via its binding to the interface of the C2A and C2B domains. The binding of Zn2+ facilitates the membrane-binding ability of Syt1 in a Ca2+-independent manner, and thus leads to the enhanced spontaneous neurotransmission by facilitating the docking of synaptic vesicles to the plasma membrane.
Our data not only extend the function of Syt1 beyond its critical roles in Ca2+-dependent neurotransmitter release, but also in Zn2+-induced abnormal neurotransmitter release under pathological conditions. Our data reconcile previously observed defects in abnormal neurotransmission in damaged neurons that cannot be fully explained by the existing working model that Syt1 serves as a primary Ca2+ sensor to mediate synaptic vesicle fusion8.
In this work, our data revealed a specific, evolutionarily conserved and Zn2+-dependent interface between the C2A and C2B domain. Tests of this interface by mutagenesis showed that this region is essential for the Zn2+-dependent vesicle fusion in a reconstituted system in-vitro. Moreover, this region is essential for abnormal spontaneous neurotransmitter release with elevated Zn2+.
Several mechanisms and models had been proposed on how Syt1 and Ca2+ cooperate to regulate the neurotransmission8,9,33,43. Most of these models, however, were established based on its physiological function in response to the Ca2+ influx. In these modes, Syt1 Ca2+-dependently binds to SNARE complex and acidic lipids on the plasma membrane via its primary interface and polybasic region on the C2B domain to initiate the synaptic vesicle docking and priming11,12,13,43,44. In the absence of Ca2+, Syt1 may lock synaptic vesicles at a pre-fusion state by forming a super-complex with SNAREs and Cpx44 Upon Ca2+ binding to both C2 domains, the loops insert into the plasma membrane and generate a positive membrane curvature, which is critical for Syt1 to stimulate Ca2+-triggered fast synchronized release under physiological conditions2,8,9,10 (Fig. 7 left).

Under physiological conditions, spontaneous release is prevented at a baseline level until Ca2+ arrival, via a cooperative action by SNAREs and Syt1. While under pathological conditions with an elevation of intracellular Zn2+ concentration, Zn2+ may bind to Syt1 at the pocket of the C2A-C2B interface, which results in an increased capacity of anionic membrane binding, and an increase in synaptic vesicle docking, thereby leading to the abnormal spontaneous release at the presynapse.
In the present work, our model of the Syt1-Zn2+ interaction suggests a new, parsimonious mechanism (Fig. 7 right) that involves an interface consisting of both C2A and C2B domains (Fig. 3e). The putative Zn2+-binding pocket contains an acidic center surrounded by several positively-charged residues, which provide a potential cation binding site (Fig. 5a). This specific conformation may only be induced when endogenous Zn2+ is elevated under pathological conditions. Zn2+ likely binds to this pocket and stabilizes this hypothetical conformation with an affinity of hundred nanomole (Fig. 3c). This interaction between Syt1 and Zn2+ in turn empower Syt1 with a stronger membrane-binding capacity (Fig. 5d). We suspected that upon Syt1/Zn2+ binding to the anionic membrane, it may induce conformational changes in the super-structures of the Syt1 oligomers and the tripartite complex composed of Syt1, SNAREs and Cpx, resulting in the relief of the clamping effect on the spontaneous release. Also, the enhancement of membrane binding ability of Syt1 may directly facilitate Syt1-mediated vesicle docking (Fig. 6), thereby significantly increasing the Zn2+-dependent fusion probability in the single vesicle fusion assay (Fig. 2), as well as increasing the frequency of spontaneous synaptic transmission in hippocampal neurons (Fig. 1).
Because intracellular Zn2+ is maintained below subnanomolar concentrations under physiological conditions23, it is unlikely that Zn2+ binds to Syt1 at a sufficient level to induce abnormal synaptic vesicle fusion. The accumulation of Zn2+ in distinctive locations is associated with certain nervous system diseases or injuries, such as the observation of enriched Zn2+ in amyloid plaques in Alzheimer’s disease (AD)45 and transient global ischemia25,27,28. Under pathological conditions with elevated Zn2+ concentration caused by neuron damage, Zn2+ could bind to the interface of the C2A and C2B domains, and induce abnormal neurotransmitter release. However, the interaction between Syt1 and Zn2+ seems to have a negligible effect on the Ca2+-triggered synchronous release (Fig. 1), indicating Ca2+ binding may disrupt the transient conformation of C2AB with Zn2+. Thus, we propose that such conformation of the Syt1-Zn2+ complex contributes to the pathological function of Syt1 in inducing the abnormal spontaneous neurotransmitter release prior to the action potential.
Moreover, in this network of interactions, C2AB domains are not required to interact with SNARE complexes. Although the mutation on the primary interface severely impaired the Ca2+-triggered fast fusion11, it does not affect the Zn2+-dependent membrane binding and vesicle fusion. Similar results were observed by the mutation on the polybasic region and the Ca2+-binding loop. Taken together, these results suggest that under pathological conditions, Syt1 could adopt a new cooperative conformation upon Zn2+-binding while preserving SNARE and Ca2+-binding modes, which is required for neurotransmission under physiological conditions. The hypothetical alternative activation model could represent a molecular mechanism underlying the excessive excitatory discharge of neurotransmitters in brain neurons under pathological conditions.
Methods
Animals
The floxed Syt1-null transgenic mice were generously provided by Peng Cao (National Institute of Biological Sciences) with the permission of Thomas C. Sudhof (Stanford University School of Medicine). Hippocampal neuron cultures were established using floxed Syt1-null mice/C57 from postnatal day 0 to 1. All animals were accommodated in an animal facility, maintaining a 12 h light/12 h dark cycle at 22 ± 2 °C, with unrestricted access to food and water. Ethical considerations for the use and care of animals were obtained through approval from the Animal Care and Use Committee of Xi’an Jiaotong University and compliance with the standards set by the Association for Assessment and Accreditation of Laboratory Animal Care International.
Hippocampal neuronal cultures and Lentivirus infection
Hippocampal neurons were cultured following established protocols46. Hippocampi obtained from floxed Syt1-null mice at postnatal day 0 to 1 underwent treatment with 0.25% trypsin at 37 °C for 15 min. Subsequently, cells were plated on polyethyleneimine-coated glass coverslips in 35 mm dishes and sustained in DMEM (Gibco) supplemented with 10% fetal bovine serum (FBS). After 3 h, the medium transitioned to Neurobasal (Gibco) supplemented with 2% serum-freemedium Supplement (Gibco), 0.5 mM L-glutamine (Sigma), and 5 µM cytosine arabinoside (Sigma). Cre-expressing Lentivirus (0.6 μL virus per well in a 24-well plate) was introduced at 5 days in-vitro (DIV) for a 72 h incubation period. Infected neurons were cultured until 13–15 DIV for subsequent biochemical, morphological, and electrophysiological analyses.
Whole-cell electrophysiological analysis of postnatal hippocampal cultures
Whole-cell patch recordings from cultured hippocampal neurons at 13–15 days in-vitro (DIV) were conducted at room temperature using an EPC-10-USB-Double amplifier (HEKA Electronic) with patchMaster acquisition software. The data were digitized at 10 kHz with a 2 kHz low-pass filter. Evoked EPSCs were selectively isolated by introducing 100 μM PTX (Sigma) to the bath solution (0.2 ms, 72 V). In mEPSC recordings, PTX (100 μM) and TTX (1 μM) (Sigma) were added to the bath to block GABA receptors and evoked synaptic responses, respectively. In sEPSC recordings, PTX (100 μM) was included to block GABA receptors. Whole-cell patches were excised from positively transfected cells, identified by epifluorescence microscopy, using 3–5 MΩ borosilicate glass pipettes (World Precision Instruments).
Synaptic transmission was triggered with a concentric focal stimulus electrode, and EPSCs were recorded with a patch electrode (3–5 MΩ) in whole-cell recording mode, filtered at 0.2 kHz. The pipette solution comprised: 130 mM K+-Gluconate, 8 mM NaCl, 10 mM HEPES, 0.6 mM EGTA, 1 mM MgATP, and 1.25 mM Na2GTP (adjusted to pH 7.4 with KOH). The extracellular solution was a HEPES-buffered saline containing: 144 mM NaCl, 10 mM KCl, 10 mM HEPES, 2 mM CaCl2, 1 mM MgCl2, 10 mM D-glucose (pH 7.4). Glutamate-induced currents were recorded by perfusing different concentrations of Zn2+ (0–200 µM) into the bath solution at 2 mL/min.
Protein expression and purification
VAMP2, Stx1A, Syt1 and its mutants (Syt1_C2A*B*(D178A D230A D232A/D309A D363A D365A), Syt1_4A(M173A F234A V304A T367A) and Syt1_4W(M173W F234W V304W T367W), Syt1_QM(R281 E295 Y338 R398/399), Syt1_QQQ(K325Q K327Q K331Q)) were expressed in Escherichia coli BL21 (DE3) cells and purified as 6×his-tagged or GST-tagged proteins, as described previously with modifications (Tucker et al., 2004; Bao et al., 2018). In brief, VAMP2, Stx1A, Syt1 and its mutants were expressed at 20 °C in Escherichia coli BL21 (DE3) cells (Tsingke Biotechnology). Bacterial pellets were resuspended (~ 10 mL per liter of culture) in resuspension buffer (25 mM HEPES-NaOH [pH 7.4], 500 mM NaCl and 1 mM TCEP). Protease inhibitor (1 mM PMSF) (Sangon Biotech) and DNase I (Sigma) were then lysed by three passes through the ATS AH-1400 homogenizer at 1000 psi. n-dodecyl-β-D-maltoside (DDM) (LABLEAD) was added to 2% (w/v) and incubated 1 h with rotation at 4 °C before centrifugation of the cell lysate at 135,900 x g for 30 min in a T-647.5 rotor (Thermo Scientific). The supernatant was then incubated for 1 h at 4 °C with Ni-NTA agarose (Qiagen) or glutathione–agarose (Smart-lifesciences) for Stx1A equilibrated in resuspension buffer. For VAMP2, Syt1 and its mutants, agarose were washed with buffer I (25 mM HEPES-NaOH [pH 7.4], 500 mM NaCl, 20 mM imidazole, 10% glycerol, 1 mM TCEP, 0.8% n-Octyl-β-D-glucoside (OG)). Proteins were eluted with elution buffer (25 mM HEPES-NaOH [pH 7.4], 400 mM NaCl, 500 mM imidazole, 10% glycerol, 1 mM TCEP, 0.8% OG). Then 6×his-tag on VAMP2 was cleavage by TEV at room temperature for 3 h, while 6×his-tag on Syt1 and its mutants remained uncleaved. For Stx1A, agarose was washed with buffer II (25 mM HEPES-NaOH [pH 7.4], 400 mM NaCl, 10% glycerol, 1 mM TCEP, 0.8% OG) and protein was cleaved by Thrombin (Sigma) on agarose. Then, proteins were loaded on a Superdex 200 Increase 10/300 GL column (GE Healthcare) for size exclusion.
For the purification of SNAP-25, C2AB, C2A, C2B, C2AB_QQQ (K325Q K327Q K331Q), and C2AB_3M (D261A E386A H389A), a similar procedure was used except all detergents were omitted from the buffers. The purified proteins were dialyzed against 25 mM HEPES-NaOH [pH 7.4], 500 mM NaCl, 10% glycerol, and 1 mM dithiothreitol (DTT), followed by a Source 15Q column (GE Healthcare) for ion exchange.
All protein sample concentrations were measured by UV absorption at 280 nm, aliquots were flash frozen in liquid nitrogen and stored at − 80 °C.
Vesicle reconstitution
For the ensemble lipid-mixing assay, the lipid composition of the SV-vesicle was 50.5 mol% 1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC), 20 mol% 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine (DOPE), 7.5 mol% 1,2-dioleoyl-sn-glycero-3-phospho-L-serine (DOPS), 20 mol% cholesterol and 1 mol% 1,1’-dioctadecyl-3,3,3’,3’-tetramethylindodicarbocyanine perchlorate (DiD) (Invitrogen); for the PM-vesicle, the lipid composition was 50.5 mol% POPC, 20 mol% DOPE, 15 mol% DOPS, 20 mol% cholesterol and 1 mol% 1,1’-dioctadecyl-3,3,3’,3’-tetramethylindocarbocyanine perchlorate (DiI) (Invitrogen) supplemented 1 mol% PIP2. All the lipids are from Avanti Polar Lipids.
For the single-vesicle content-mixing assay, the lipid composition of the SV-vesicle was 52.5 mol% POPC, 20 mol% DOPE, 7.5 mol% DOPS, and 20 mol% cholesterol; for the PM-vesicle, the lipid composition was 50.5 mol% POPC, 20 mol% DOPE, 15 mol% DOPS, and 20 mol% cholesterol supplemented 1 mol% PIP2 and 0.1 mol% biotinylated PE.
The reconstitution method for PM- and SV-vesicles is described in detail ref. 34. Dried lipid films were dissolved in 110 mM OG buffer containing purified proteins at protein-to-lipid ratios of 1:200 for VAMP2 and Stx1A in SV- and PM-vesicles, and 1:800 for Syt1 in SV-vesicle.
Ensemble lipid-mixing assay
For the lipid-mixing assay, SV-vesicle containing 1 mol% DiD were mixed with PM-vesicle containing 1 mol% DiI in the presence of 100 µM cation ions, or different concentrations of Zn2+ (as indicated in the figures) in a total volume of 100 μL. The intensities of fluorescence emission at 570 nm and 665 nm (excitation 540 nm) were monitored by DuettaTM (Horiba). All experiments were performed at 30 °C for at least 3 times.
For lipid-mixing assays, ΔFRET efficiency was calculated according to [IAccepter/(IAccepter + IDonor)]-FRET0, FRET0 represents the FRET value at the zero time point. The fusion percentage was normalized by the equation Fobs/0.95, where Fobs is the observed fluorescence intensity and 0.95 is the maximum ΔFRET efficiency obtained by using vesicles containing 1 mol% DiD and 1 mol% DiI.
Single-vesicle content-mixing assay
The surface of the quartz slides was passivated by coating the surface with polyethylene glycol (PEG) to alleviate non-specific binding of vesicles. The same protocol and quality controls (surface coverage and non-specific binding) were used as described previously34,35.
In brief, the PM-vesicles were introduced into the chamber and anchored to the PEG-coated surface through biotin-neutravidin (NTV) lipid conjugation. Following a 30 min incubation at room temperature (~ 25 °C), any unbound PM-vesicles were effectively eliminated through two rounds of washing with 200 μL of buffer V. Subsequently, the SV-vesicles reconstituted with Syt1_WT or its mutants, either with or without Zn2+, were loaded into the chamber, and the sample underwent a 1 min incubation at room temperature (~ 25 °C). To ensure the removal of unbound SV-vesicles, the chamber underwent two additional rounds of washing with 200 μL of buffer V. Images were captured with a 200 ms exposure time, selecting ten random imaging areas for quantitative analysis for the single vesicle docking assay. Association of SV-vesicles with PM-vesicles was monitored for 1 min. Ion-dependent fusion events were monitored within the same field of view upon injection for a 1 min period. The injection was performed at a speed of 66 μL/s by a motorized syringe pump (RWD, China). All experiments were carried out at ambient temperature (25 °C).
TIRF imaging
Neurons underwent transient transfection with the synaptophysin–pHluorin construct at 5 days of cultivation in-vitro (DIV 5) and were subsequently imaged at DIV 12–13. Image acquisition spanned 2 min with a 200 ms exposure time, utilizing fluorescence imaging through a TIRF microscope. A solution perfusion protocol was implemented, following the specified order: a 5 s baseline in extracellular solution, 1 s in 45 mM NH4Cl in extracellular solution, 30 s wash in extracellular solution, 30 s in interval, 30 s in 200 μM Zn2+, and 1 s in 45 mM NH4Cl in extracellular solution. The subsequent analysis of fluorescence intensity changes was performed by using the TIRF microscope and ImageJ software for image analysis. Regions of interest (ROIs) were selected based on a consistent fluorescence intensity threshold (Syp-pH for synaptic areas) across different samples. The average intensity of all ROIs, exceeding ten cell regions in each image stack, was exported.
Zn2+ binding assay
8 μM Zn2+, 400 nM FluoZin™-1 (Thermo Fisher Scientific) and 2 μM C2AB or its variants as specified were mixed and incubated for 30 min at room temperature. The emission spectrum (500–600 nm) of FluoZin™-1 in a mixture was obtained by using an excitation wavelength at 488 nm.
In the titration experiment, 2 μM Zn2+ and 400 nM FluoZin™-1 were mixed with different concentrations of C2AB (from 3 nM to 6.25 μM) or C2AB_3M (from 48 nM to 25 μM). The fluorescence intensity of FluoZin™-1 was measured with DuettaTM fluorimeter using an excitation wavelength of 488 nm and emission wavelength of 520 nm at room temperature.
CD spectroscopy
CD spectra were acquired employing the AVIV stop-flow CD spectropolarimeter within the wavelength range of 200–260 nm, utilizing a cell featuring a 1 mm path length. The averaged values of 15 measurements were recorded. Samples consisting of 0.3 mg/mL of C2AB or C2AB mutant were measured in PBS buffer at pH 7.0 at 20 °C.
Lipid co-sedimentation assay
Liposomes containing 43–59 mol% POPC, 20 mol% POPE, 20 mol% cholesterol, 2 mol% DiI, 0–15 mol% DOPS, and 1 mol% PIP2 were prepared following established procedures. Subsequently, 10 µM of C2AB or C2AB mutant were incubated with 0.1 mM liposomes in the presence of various concentrations of Zn2+ at room temperature for 30 min in the protection from light. The mixtures underwent centrifugation at 21,300 x g for 15 min at 4 °C. The resulting supernatant was collected, while the precipitant fraction was resuspended in an equal volume of buffer (25 mM HEPES, 300 mM NaCl, pH 7.4). Subsequent examination of protein and lipids was achieved through SDS-PAGE analysis, and the fluorescence emission in the range of 540–600 nm (with excitation at 540 nm) using a DuettaTM instrument. Each experiment was replicated at least three times, and data analysis was performed using ImageJ and Prism 8.0.
Software and statistical analysis
All experiments were independently replicated at least three times. Data are shown as the mean ± SEM. Softwares used for data collection include PatchMaster v2×90.2, ECLIPSE Ti2 and EzSpecUpgradeV1.3. Statistical data analysis software includes GraphPad Prism 8.0, ImageJ, smCamera and OriginPro 2024.
Statistical comparisons were made with the two-tailed unpaired t test, one-way ANOVA as indicated. The number of experiments, sample size and statistical tests are reported in the respective figure captions. P-values < 0.05 were considered significant, n.s. was considered to indicate non-significance at the 95% confidence level.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The crystal structure of the C2AB fragment of Syt3 used in this study were obtained from the Protein Data Bank (PDB) with accession code 3HN8. The complex of Syt1/Syt3 and Zn2+ data generated in this study have been deposited in the ModelArchive database under accession codes ma-sle84 [https://www.modelarchive.org/doi/10.5452/ma-sle84], ma-36fbl [https://www.modelarchive.org/doi/10.5452/ma-36fbl]. The Syt1 protein sequences used in this study were retrieved from Uniprot with accession ID: P21707. All other data supporting the results of this paper are available in the article, supplementary information or source data file. Source data are provided in this paper.
References
Weber, T. et al. SNAREpins: minimal machinery for membrane fusion. Cell 92, 759–772 (1998).
Südhof, T. C. Neurotransmitter release: the last millisecond in the life of a synaptic vesicle. Neuron 80, 675–690 (2013).
Chapman, E. R. How does synaptotagmin trigger neurotransmitter release?. Annu. Rev. Biochem. 77, 615–641 (2008).
Wickner, W. & Schekman, R. Membrane fusion. Nat. Struct. Mol. Biol. 15, 658–664 (2008).
Fernández-Chacón, R. et al. Synaptotagmin I functions as a calcium regulator of release probability. Nature 410, 41–49 (2001).
Ubach, J., Zhang, X., Shao, X., Südhof, T. C. & Rizo, J. Ca2+ binding to synaptotagmin: how many Ca2+ ions bind to the tip of a C2-domain?. EMBO J. 17, 3921–3930 (1998).
Fernandez, I. et al. Three-dimensional structure of the synaptotagmin 1 C2B-domain: synaptotagmin 1 as a phospholipid binding machine. Neuron 32, 1057–1069 (2001).
Xu, J., Pang, Z. P., Shin, O.-H. & Südhof, T. C. Synaptotagmin-1 functions as a Ca2+ sensor for spontaneous release. Nat. Neurosci. 12, 759–766 (2009).
Shin, O.-H., Xu, J., Rizo, J. & Südhof, T. C. Differential but convergent functions of Ca2+ binding to synaptotagmin-1 C2 domains mediate neurotransmitter release. Proc. Natl. Acad. Sci. USA 106, 16469–16474 (2009).
Bai, J., Tucker, W. C. & Chapman, E. R. PIP2 increases the speed of response of synaptotagmin and steers its membrane-penetration activity toward the plasma membrane. Nat. Struct. Mol. Biol. 11, 36–44 (2004).
Zhou, Q. et al. Architecture of the synaptotagmin-SNARE machinery for neuronal exocytosis. Nature 525, 62–67 (2015).
Honigmann, A. et al. Phosphatidylinositol 4,5-bisphosphate clusters act as molecular beacons for vesicle recruitment. Nat. Struct. Mol. Biol. 20, 679–686 (2013).
van den Bogaart, G. et al. Synaptotagmin-1 may be a distance regulator acting upstream of SNARE nucleation. Nat. Struct. Mol. Biol. 18, 805–812 (2011).
Brunger, A. T., Choi, U. B., Lai, Y., Leitz, J. & Zhou, Q. Molecular Mechanisms of Fast Neurotransmitter Release. Annu. Rev. Biophys. 47, 469–497 (2018).
Martens, S., Kozlov, M. M. & McMahon, H. T. How synaptotagmin promotes membrane fusion. Science 316, 1205–1208 (2007).
Gruget, C. et al. Rearrangements under confinement lead to increased binding energy of Synaptotagmin-1 with anionic membranes in Mg(2+) and Ca(2). FEBS Lett. 592, 1497–1506 (2018). at.
Shin, O.-H. et al. Sr2+ binding to the Ca2+ binding site of the synaptotagmin 1 C2B domain triggers fast exocytosis without stimulating SNARE interactions. Neuron 37, 99–108 (2003).
Silinsky, E. M. On the role of barium in supporting the asynchronous release of acetylcholine quanta by motor nerve impulses. J. Physiol. 274, 157–171 (1978).
Zhou, Q. et al. The primed SNARE-complexin-synaptotagmin complex for neuronal exocytosis. Nature 548, 420–425 (2017).
Tagliatti, E. et al. Synaptotagmin 1 oligomers clamp and regulate different modes of neurotransmitter release. Proc. Natl Acad. Sci. USA 117, 3819–3827 (2020).
Wang, J. et al. Circular oligomerization is an intrinsic property of synaptotagmin. Elife 6, https://doi.org/10.7554/elife.27441 (2017).
Choi, U. B., Zhao, M., Zhang, Y., Lai, Y. & Brunger, A. T. Complexin induces a conformational change at the membrane-proximal C-terminal end of the SNARE complex. Elife 5, https://doi.org/10.7554/elife.16886 (2016).
Frederickson, C. J., Koh, J.-Y. & Bush, A. I. The neurobiology of zinc in health and disease. Nat. Rev. Neurosci. 6, 449–462 (2005).
Sensi, S. L., Paoletti, P., Bush, A. I. & Sekler, I. Zinc in the physiology and pathology of the CNS. Nat. Rev. Neurosci. 10, 780–791 (2009).
Sensi, S. L. et al. Measurement of intracellular free zinc in living cortical neurons: routes of entry. J. Neurosci.17, 9554–9564 (1997).
Lee, J. Y., Cole, T. B., Palmiter, R. D. & Koh, J. Y. Accumulation of zinc in degenerating hippocampal neurons of ZnT3-null mice after seizures: evidence against synaptic vesicle origin. J. Neurosci.20, RC79 (2000).
Koh, J. Y. et al. The role of zinc in selective neuronal death after transient global cerebral ischemia. Science 272, 1013–1016 (1996).
Aguilar-Alonso, P. et al. The increase in zinc levels and upregulation of zinc transporters are mediated by nitric oxide in the cerebral cortex after transient ischemia in the rat. Brain Res. 1200, 89–98 (2008).
Levenson, C. W. Zinc and Traumatic Brain Injury: From Chelation to Supplementation. Med. Sci. 8, https://doi.org/10.3390/medsci8030036 (2020).
Doering, P. et al. Changes in the vesicular zinc pattern following traumatic brain injury. Neuroscience 150, 93–103 (2007).
Coulter, D. A. Mossy fiber zinc and temporal lobe epilepsy: pathological association with altered ‘epileptic’ gamma-aminobutyric acid A receptors in dentate granule cells. Epilepsia 41, S96–S99 (2000).
Upmanyu, N. et al. Colocalization of different neurotransmitter transporters on synaptic vesicles is sparse except for VGLUT1 and ZnT3. Neuron 110, 1483–1497 (2022).
Courtney, N. A., Bao, H., Briguglio, J. S. & Chapman, E. R. Synaptotagmin 1 clamps synaptic vesicle fusion in mammalian neurons independent of complexin. Nat. Commun. 10, 4076 (2019).
Lai, Y. et al. Complexin inhibits spontaneous release and synchronizes Ca2+-triggered synaptic vesicle fusion by distinct mechanisms. Elife 3, e03756 (2014).
Kyoung, M., Zhang, Y., Diao, J., Chu, S. & Brunger, A. T. Studying calcium-triggered vesicle fusion in a single vesicle-vesicle content and lipid-mixing system. Nat. Protoc. 8, 1–16 (2013).
Vrljic, M. et al. Molecular mechanism of the synaptotagmin-SNARE interaction in Ca2+-triggered vesicle fusion. Nat. Struct. Mol. Biol. 17, 325–331 (2010).
Suh, B.-C. & Hille, B. Electrostatic interaction of internal Mg2+ with membrane PIP2 Seen with KCNQ K+ channels. J. Gen. Physiol. 130, 241–256 (2007).
Park, Y. et al. Controlling synaptotagmin activity by electrostatic screening. Nat. Struct. Mol. Biol. 19, 991–997 (2012).
Qian, J. & Noebels, J. L. Exocytosis of vesicular zinc reveals persistent depression of neurotransmitter release during metabotropic glutamate receptor long-term depression at the hippocampal CA3-CA1 synapse. J. Neurosci.26, 6089–6095 (2006).
Hirzel, K. et al. Hyperekplexia phenotype of glycine receptor alpha1 subunit mutant mice identifies Zn(2+) as an essential endogenous modulator of glycinergic neurotransmission. Neuron 52, 679–690 (2006).
Kay, A. R., Neyton, J. & Paoletti, P. A startling role for synaptic zinc. Neuron 52, 572–574 (2006).
Yoshihara, M., Guan, Z. & Littleton, J. T. Differential regulation of synchronous versus asynchronous neurotransmitter release by the C2 domains of synaptotagmin 1. Proc. Natl. Acad. Sci. USA 107, 14869–14874 (2010).
Chang, S., Trimbuch, T. & Rosenmund, C. Synaptotagmin-1 drives synchronous Ca(2+)-triggered fusion by C(2)B-domain-mediated synaptic-vesicle-membrane attachment. Nat. Neurosci. 21, 33–40 (2018).
Krishnakumar, S. S. et al. A conformational switch in complexin is required for synaptotagmin to trigger synaptic fusion. Nat. Struct. Mol. Biol. 18, 934–940 (2011).
Bush, A. I. et al. Rapid induction of Alzheimer A beta amyloid formation by zinc. Science 265, 1464–1467 (1994).
Kaech, S. & Banker, G. Culturing hippocampal neurons. Nat. Protoc. 1, 2406–2415 (2006).
Acknowledgements
This study was supported by Sichuan University West China Hospital (ZYYC24003), National Natural Science Foundation of China (32170686 and 31900688 to Y.L., 32171233 and 31670843 to C.W.), Transformation Foundation of Tianfu Jincheng Laboratory (2025ZH020), and Natural Science Foundation of Sichuan Province (2024NSFSC0385).
Author information
Authors and Affiliations
Contributions
YJ.X., LL.C., JY.Y., and XC.L. contributed equally to this work. CH.W. and Y.L. conceived the study and designed the experiments. YJ.X., LL.C., JY.Y., XC.L., MD.W., JX.H., JQ.F., H.L., KY.L., XD.W., YK.S., XF.Y. YJ.X., and JY.Y. performed the experiments and analyzed these data. YJ.X., CH.W., and Y.L. drafted the manuscript. All authors reviewed the manuscript and approved its submission.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Xiang, Y., Cui, L., Yao, J. et al. Synaptotagmin-1 serves as a primary Zn2+ sensor to mediate spontaneous neurotransmitter release under pathological conditions. Nat Commun 16, 7113 (2025). https://doi.org/10.1038/s41467-025-62496-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-62496-1
