
Abstract
The kidney maintains fluid homeostasis by reabsorbing essential compounds and excreting waste. Proximal tubule cells, crucial for reabsorbing sugars, ions, and amino acids, are highly susceptible to injury, often leading to pathologies necessitating dialysis or transplants. Human pluripotent stem cell-derived kidney organoids offer a platform to model renal development, function, and disease, but proximal nephron differentiation and maturation in these structures is incomplete. Here, we drive proximal tubule development in pluripotent stem cell-derived kidney organoids by mimicking in vivo proximal differentiation. Transient PI3K inhibition during early nephrogenesis activates Notch signaling, shifting nephron axial differentiation towards epithelial and proximal precursor states that mature to proximal convoluted tubule cells broadly expressing physiology-imparting solute carriers including organic cation and organic anion family members. The “proximal-biased” organoids thus acquire function, and on exposure to nephrotoxic injury, display tubular collapse and DNA damage, and upregulate injury response markers HAVCR1/KIM1 and SOX9 while downregulating proximal transcription factor HNF4A. Here, we show that proximally biased human-derived kidney organoids provide a robust model to study nephron development, injury responses, and a platform for therapeutic discovery.
Similar content being viewed by others


Introduction
Proximal nephron cells are the most abundant cells in the human kidney, are responsible for reabsorbing 65% of the nephron filtrate1, and their pathologies are the primary reason patients require dialysis and kidney transplants2,3. In spite of their clinical importance, proximal tubule disease mechanisms are poorly understood, and human models are needed to scrutinize disease origins and etiology. Stem cell-derived human kidney models recapitulating proximal tubule functions would therefore provide a critical tool to study renal disease and develop new therapeutic approaches.
Directed differentiation protocols coaxing induced pluripotent stem cells (iPSCs) to intermediate mesoderm lineages have led to the development of human kidney-like organoids4,5. These models partially replicate developmental kidney cell profiles, but do not form mature proximal tubule cells, and the proximal precursor-like cells that do develop exhibit low expression of genes that normally impart nephron-specific physiologies6,7,8,9,10,11. Similarly, while current organoid models respond to nephrotoxic compounds by upregulating specific injury markers such as KIM1/HAVCR1 and γH2AX in LTL+ cells12,13,14, the lack of homogenous proximal tubule like cells in organoids expressing nephrotoxin-transporting solute carriers limits their utility in studying acute proximal tubular injury and performing proximal nephron-specific drug screens.
Studies performed in mice and organoids show that proximal tubule development is dependent on expression of Hnf4a/HNF4A, whose transcription factor protein product is required for the normal expression of roughly 300 proximal tubule solute carriers, protein, ion, and substrate transporters, and other proximal nephron functional genes15,16,17. Recently, effort has therefore focused on generating HNF4A+ proximal tubule precursors as these would serve as building blocks for generating functional proximal tubules. One strategy has enriched for this cell population by expanding the pool of nephron progenitors in early organoids and subsequently allowing their differentiation10, but regardless of the protocol, proximal nephron precursors display relatively low expression of HNF4A and HNF4A-dependent genes6,7,8,9,10,11,15.
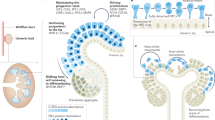
In vivo, proximal tubule precursors emerge in the S-shaped body (SSB) nephron following a stereotyped and deeply conserved developmental program18. In this program, nephron progenitors are gradually recruited from their niche into pretubular aggregates that progressively undergo epithelial-to-mesenchymal transitions, generating epithelial renal vesicles. Complex morphogenetic events form tubular Comma-shaped and thereafter the aforementioned SSB nephrons with distal and proximal gene signatures positioned along the emerging distal-to-proximal axial polarity18,19,20. Transcriptionally distinct HNF4A+ proximal tubule precursors develop in narrow 2–3 cell-wide populations within each medial domain of SSBs, in a field of Notch ligand JAG1+ cells18,19,21. Their development is dependent on Notch signaling, as Notch1 and Notch2 loss-of-function mice fail to normally express transcription factor Hnf1b, which in turn binds to and is necessary for Hnf4a expression22,23,24. In addition to Notch, the development of the proximal-distal nephron axis requires integrated signaling between several pathways as spatial positions in the nascent nephron are known to be driven by Notch, Wnt, BMP, and PI3K signaling, each tuning the formation of precursor populations18,25.
In this study, we address the challenge of generating proximal tubule cells by developing a protocol to expand the HNF4A+ proximal nephron precursor population within kidney organoids and in individual nephrons. We direct organoid cell differentiation along an in vivo-like developmental trajectory proceeding through JAG1+/HNF1B+ fates and culminating in maturing HNF4A+ proximal precursors. Comparative analyses with in vivo development show the organoid proximal precursors resemble HNF4A+ proximal tubule cells in the capillary loop stage (CLSN) nephron with emerging physiologies. Proximal-biased (PB) nephrons display expression of solute carriers and transporters and are capable of selectively transporting albumin and dextran. We leverage the prevalence of proximal structures expressing cisplatin transporters to demonstrate significant upregulation of KIM1/HAVCR1 within HNF4A+ organoid nephron tubules in response to nephrotoxic injury. In line with in vivo injury, this response is mosaic and KIM1/HAVCR1+, HNF4A+ PB organoid nephrons display collapsed apical-basal polarities, DNA damage, and upregulate injury-response marker SOX9. The PB nephron model represents a significant step towards recapitulating kidney development and function in organoid systems and provides a direct strategy to study proximal nephrotoxicity and tubulopathies in a robust human assay.
Results
Identifying abnormal kidney organoid developmental programs
To identify differences between developing nephrons in kidneys and organoids that can explain why organoids do not generate maturing proximal tubule cells, we characterized how proximal precursors form in vivo in developing human kidneys and compared organoids to this framework using single-cell RNA-sequencing and secondary validation for proteins that mark and drive proximal tubule development.
The specification of proximal precursors is consequent to the gradual recruitment of nephron progenitor cells into the forming nephron and signaling pathways that tune differentiation along the progressively emerging proximal-distal axis21,25. This process begins with a cellular domain developing in the distal renal vesicle nephron, formed by early recruited nephron progenitor cells. The domain is marked by membrane-localized JAG1 and nuclear HNF1B and is in direct contact with the ureteric epithelium. It abuts the proximally located WT1+ region where cells are actively recruited from the nephron progenitor cell niche (Fig. 1a). As the renal vesicle develops into a comma-shaped body nephron, the HNF1B+/JAG1+ domain expands proximally. The initial distal domain downregulates JAG1 to form HNF1B+/JAG1LOW cells, while the proximally expanding domain further upregulates JAG1 to become HNF1B+/JAG1HIGH (now considered the medial domain of the comma-shaped body), and the medial domain forms a boundary with the WT1+ proximal-most domain where the last nephron progenitors are recruited (Fig. 1a, b). At the comma-shaped nephron stage, the nephron is subdivided by these markers into three distinguishable domains. As the nephron develops further, HNF1B is upregulated in nuclei positioned at the border between the medial and proximal-most domain, where HNF4A is detected in late comma-shaped/early S-shaped body nephrons (Fig. 1b). Proximal tubule precursor cells (HNF1BHIGH/HNF4A+) then develop and downregulate JAG1 when the S-shaped nephron matures into capillary loop stage nephrons adopting an elongated form with a HNF1BHIGH/HNF4AHIGH/JAG1− proximal tubule cell state (Fig. 1b). These findings are consistent with single-cell RNA-sequencing of the developing human nephron lineage (Supplementary Fig. 1a–c and Supplementary Data 1). Proximal tubule precursor cells diverge from other nephron lineages in an early PAX8+ cell population in the pretubular aggregate, which generates WT1+ podocyte precursors, TFAP2A+ distal precursors, and JAG1+ cells that sequentially upregulate HNF1B and HNF4A (data combined from refs. 8,18), (Supplementary Fig. 1a–c and Supplementary Data 1).

a, b Immunofluorescent antibody stains at defined stages of nephrogenesis. Human kidneys from week 16.1 of development. a represents stages before the onset of HNF4A, while b represents the commencement and elongation of the HNF4A+ domain. White arrowheads indicate earliest detectable HNF4A+ cells. S-shaped body-stage channels are split. Scale bars: 10 microns. c, d Whole-mount immunofluorescent stains of day 10 kidney organoids. Boxed regions are magnified and split into individual channels. Scale bars: 10 microns. e, f Whole-mount immunofluorescent stains of kidney organoid nephrons. Data in (e) match proteins and stages in (a), while (f) corresponds to (b). White arrowheads in (f) indicate earliest detectable HNF4A+ cells (Day 13) and earliest emerging HNF4A+ domain (Day 14). Scale bars: 10 microns. g Hierarchical clustering from bulk RNA-sequencing of kidney organoids at differentiation days 10, 12 (control) and 12 (treated for 48 h with 10 µM LY294002). Genes shown are those that are highlighted in Supplementary Fig. 1j. Legend categorizes genes and timepoints. Timeline of organoid differentiation and samples profiled is shown in the top right. h, i Gene set enrichment analysis plots from Day 12 LY294002-treated / Day 12 control kidney organoids. Plots depict pathway effects from treatment with the PI3K inhibitor LY294002: downregulated (PI3K-Akt signaling pathway, h and upregulated (Notch signaling pathway, (i). Normalized enrichment scores (NES), false discovery rates (FDR), and p-values are shown for each.
Organoid nephrons, however, form differently. At day 10, kidney organoids consist of individual cell-aggregates positive for nephron progenitor markers (WT1, SIX1) and nephron lineage markers (PAX2, PAX8). These cells coalesce around a forming apical epithelial polarity (CDH1, ZO1, JAG1) with an accreting basement membrane (LAMB1). These dynamics are consistent across organoids generated from different human iPSC lines using a modified Takasato protocol4,26 (Fig. 1c, d and Supplementary Fig. 1g–i).
As the aggregates further epithelialize (days 11-12), they display uniform deposition of WT1 in nuclei, and JAG1 as cytoplasmic puncta and weakly at cell membranes around the periphery of the forming nephron (Fig. 1e, f), while HNF1B is upregulated between days 11 and 12. At these stages, there are no indications of WT1, JAG1, and HNF1B being distributed along a gradually forming proximal-distal axis as seen in vivo (Fig. 1a); rather, the organoid nephrons exhibit an initially homogenous HNF1B+/JAG1+/WT1+ triple-positive cell state (Fig. 1e). During further differentiation, organoid nephrons generate HNF1B+/HNF4A+ cells, but these do not transition into a rapidly elongating phase as observed in vivo, and JAG1 remains strongly detectable (Fig. 1b, f). Single-cell data from organoids sampled over time and differentiation protocols indicate that the abnormal early and late triple-positive PAX8+/JAG1+/WT1+ and late HNF1B+/HNF4A+/JAG1+ cell states are a common trend across models (reanalyzed and combined from several sources)4,5,6,7,8,9 (Supplementary Fig. 1d–f and Supplementary Data 2).
Given the importance of Hnf1b and Hnf4a for normal proximal tubule development, we examined the possibility that organoids abnormally regulate functionally important Hnf1b/HNF1B and Hnf4a/HNF4A-mediated transcriptional programs and assessed organoid proximal fate development against the in vivo developmental blueprint. To identify genes that are co-expressed with each transcription factor in vivo (Supplementary Fig. 1a), we performed a Pearson expression correlation analysis for each gene and characterized the expression of HNF1B and HNF4A correlates. The expression of these genes in vivo was ordered along a predicted proximal developmental trajectory, and their expression patterns highlight a transition from HNF1B correlates in early cells (cluster 9: pretubular aggregate/renal vesicle; e.g., JAG1, HES1, KRT8) through to a gradually more HNF4A-correlating signature (clusters 21, 24: S-shaped body; e.g., CLU, DCDC2, ANXA4), and finally genes strongly enriched in the maturing proximal precursors (cluster 20: Capillary loop stage nephron; e.g., ASS1, SLC34A1, SLC22A8; Supplementary Fig. 2a (human)).
Using this progression as a framework for the temporal in vivo sequence of transcriptional events, we compared it to organoid proximal precursor development across models4,5,6,7,8,9 (Supplementary Fig. 2a (organoid) based on Supplementary Fig. 1d). In vitro, HNF1B-correlates such as JAG1, HES1, KIF12, and KRT8 were detected while other genes, for instance transcription factor ELF3, fibroblast growth factor receptor FGFR4, serine protease inhibitor SERPINF2, and extracellular signaling protein CYR61 were not. This partial recapitulation of the transcription profile was further reduced for HNF4A-correlates, with only 25/46 genes detected across in vitro models. Genes coding for a range of protein types were not detected or significantly expressed, for instance, solute carriers SLC22A8, SLC5A8, SLC16A9, and transferases and enzymes such as AGXT2, GLYAT, and ANPEP.
To independently examine whether HNF4A-dependent genes are underrepresented in the organoid proximal program, we intersected genes downregulated upon in vivo Hnf4a loss-of-function16, with transcriptional profiles from detailed expression maps of the adult male and female mouse kidney27. We categorized Hnf4a-dependent genes as those expressed only in development and those that persist into the adult functional nephron. Two hundred and fifty seven genes are significantly downregulated (control vs. mutant kidneys, padj < 0.01, log2FC ≥ 1.5)28 in postnatal day 0 animals on loss of Hnf4a (reanalyzed from ref. 16), and 415 genes are enriched in the adult male and female proximal tubules (Kidney Cell Explorer)27 (Supplementary Fig. 2b). Of the 257 Hnf4a-dependent genes, 124 genes were enriched only in the developing kidney while 133 genes showed persistent expression into functional adult nephrons. To determine whether human orthologs are expressed in the developing human proximal tubule and in organoid models, we intersected our gene lists with single-cell transcriptional data (Supplementary Fig. 1a and Supplementary Fig. 1d). Of the 133 Hnf4a-dependent genes expressed during development and in adult mouse nephrons, 92 human orthologs were detected. 91/92 were also detected in organoids, and 89/92 (96.7%) were detected at greater abundance in vivo (Supplementary Fig. 2c, e and Supplementary Data 3). Similarly, of the 124 Hnf4a-dependent genes expressed during development, 81 human orthologs were detected. Eighty of these were detected in organoids, but 70/81 (86.4%) were again detected at a higher frequency in vivo (Supplementary Fig. 2d, f and Supplementary Data 4).
These data show that correlates of HNF4A and human orthologs of Hnf4a-dependent genes are expressed infrequently in organoids. This is consistent across organoid models, collectively pointing to abnormal regulation of HNF4A-mediated gene expression.
Initiating a proximal-forming cell-state in kidney organoids
We have previously identified a relationship between PI3K signaling and Notch ligand Jag129. In mouse kidneys, WNT/β-catenin and PI3K signaling have opposing effects on Jag1 expression, and pharmacological inhibition of PI3K signaling results in rapid upregulation of Jag1 in nephron progenitors and early nephrons25,29. We therefore tested whether inhibition of PI3K and upregulation of JAG1 could drive a proximal nephron program and provide a tool to generate human proximal tubule precursors in kidney organoids.
We treated organoids with PI3K inhibitor LY294002 (Ly29)30,31 from days 10-12, when JAG1 is normally partially upregulated in vitro and organoid nephrons epithelialize. We transcriptionally profiled them before and after treatment using RNA-sequencing. PI3K inhibitor-treated organoids displayed a 1.78-fold increase in JAG1 expression, and Notch pathway genes LNFG, HES1, and HES4 were upregulated, as were differentiation markers PAX8 and LHX1. Nephron progenitor markers SIX1, MEOX1, OSR1, and EYA1 were downregulated. Wnt/β-catenin targets WNT4 and LGR5 were unchanged (Fig. 1g, Supplementary Fig. 1j and Supplementary Data 5). Gene set enrichment analyses confirmed that the Notch pathway was selectively upregulated, PI3K pathway downregulated, and other unrelated pathways, e.g., Hedgehog, were unaffected (Fig. 1h, i and Supplementary Fig. 1k). Within this 48 h timeframe, HNF1B expression was upregulated 2.88-fold in PI3K inhibited samples, but HNF4A was not detected at day 12 (TPM < 1.7; Fig. 1g, Supplementary Fig. 1j and Supplementary Data 5). These data show that organoids upregulate JAG1 and HNF1B in response to transient PI3K inhibition.
To unbiasedly investigate proximalization in organoid nephron cells, we performed single-cell RNA-sequencing on day 10 untreated, day 12 control, and day 12 Ly29-treated organoids (Fig. 2a). Differential gene expression analyses revealed distinct transcriptional profiles (Fig. 2b and Supplementary Data 6). Day 10 organoid nephrons were enriched for nephron progenitor markers SIX1, CITED1, MEOX1, and CRABP2. By day 12, control organoid nephrons upregulated podocyte markers TARID, an antisense lncRNA that activates podocyte-specific TCF21 expression32, as well as podocyte markers PODXL, CLDN5, KIRREL3, and CLIC5. Ly29-treated organoid nephrons, however, showed increased expression of epithelialization and nephron tubule markers, including CDH4, DCDC2, JAG1, LHX1, and HNF1B, indicating a shift toward an epithelial nephron tubule fate (Fig. 2b–d and Supplementary Data 6).

a UMAP of single-cell RNA-sequencing from day 10 (pre-treatment), day 12 control, and day 12 PI3K inhibitor (LY294002)-treated kidney organoid nephrons. Cells are colored by sample with marker gene annotations. b Heatmap of Z-scores for hierarchically clustered differentially expressed genes across samples. Representative genes are listed, with a Z-score legend. c Feature plots of genes enriched in each sample group from the differentially expressed gene list. d Feature plots of selected genes for each group based on sample characteristics. For c, d human nephron single-cell detection (Supplementary Fig. 1a) is shown on the left, organoid plot on the right. e UMAP of single-cell RNA-sequencing from day 12 control and day 12 LY294002-treated kidney organoid nephrons. Cells match a but exclude day 10 sample. Cells are colored by sample with marker gene annotations. f Heatmap of log2 fold change (log2FC) for differentially expressed genes, calculated as the ratio of LY294002-treated to control aggregate expression. Representative genes are listed with a log2FC legend. g Feature plots of genes enriched in each sample from the differentially expressed gene list. Human nephron single-cell (Supplementary Fig. 1a) detection is on the left, organoid plot on the right. h Split violin plots of genes from the differentially expressed gene list. Top two rows show genes enriched in day 12 control cells; bottom two rows show genes enriched in day 12 LY294002-treated cells. Plot colors match sample colors from (e). i Gene ontology terms for the top 50 differentially expressed genes per sample. Top graph: day 12 control nephron cells; bottom graph: LY294002-treated nephron cells. j–l Whole-mount immunofluorescent stains of day 12 control and LY294002-treated kidney organoids. Boxed regions are magnified. Inset in j shows absence of detectable HNF4A protein (orange) in day 12 samples. Scale bars: 10 microns. m Whole-mount immunofluorescent stain of day 12 control and LY294002-treated kidney organoids. Insets highlight JAG1 and HNF1B protein detection. Scale bars: 500 microns. n Quantification of JAG1+ and HNF1B+ nephron size (µm2) and HNF1B+ intensity (RFU) for n = 3 day 12 organoids each, across all positive segments. SEM error bars shown. Statistical significance determined by two-sided Student’s t-test.
To directly assess the impact of PI3K inhibition on differentiation, we compared day 12 control and Ly29-treated organoid nephron cells (Fig. 2e). Differential gene expression analyses substantiated strong divergent profiles of day 12 control and Ly29-treated samples with control cells enriched for progenitor genes MEOX1 and BMPER, and podocyte markers PODXL and CCBE1, while Ly29-treated cells upregulated epithelial and nephron tubule markers, including CDH4, LHX1, and POU3F3; Notch pathway ligands JAG1 and DLL1; and the HNF4A-binding transcription factor HNF1B23 (Fig. 2e–g and Supplementary Data 7). Most HNF1B+ cells at day 12 originated from Ly29-treated nephrons, with similar patterns observed for other key markers (Fig. 2h). Unbiased gene ontology analyses of the top 50 most differentially expressed genes for each condition showed day 12 control cells being enriched for processes related to podocyte differentiation and glomerulus development, while day 12 Ly29-treated cells were enriched for nephron tubule development, kidney epithelium development, and cell adhesion (Fig. 2i). Together, these findings suggest that day 12 control cells are primed for podocyte development, while PI3K inhibitor treatment shifts cells toward an epithelial nephron tubule fate, with strong upregulation of Notch and pre-proximal tubule markers, reflecting patterns observed in human proximal nephron development.
To validate and extend these findings, we performed whole-mount immunofluorescence in organoids using two iPSC lines to confirm phenotype robustness. Consistent with single-cell RNA-sequencing results, day 12 Ly29-treated cells showed strong upregulation of JAG1 and HNF1B throughout organoid nephrons, while control organoids exhibited mosaic detection of HNF1B and lower levels of JAG1. As expected based on timing, HNF4A was not yet detected in either condition at day 12 (Fig. 2j). The upregulation of JAG1 was accompanied by increased detection of Notch target HES1, which remained low in controls (Fig. 2k). By day 12, a fully formed basement membrane, marked by LAMB1, was observed in both control and Ly29-treated organoids (Fig. 2k). Co-upregulation of epithelialization marker CDH1 and HNF1B was observed, with increased intensity in Ly29-treated samples (Fig. 2l). Quantitative assessment of JAG1 and HNF1B abundance in day 12 organoids showed increases in JAG1 and HNF1B in Ly29-treated organoids. Their upregulation occurred in nephrons alongside elongation (HNF1B 1.64-fold size increase, p-value < 0.0001; JAG1, 2.36-fold size increase, p-value < 0.0001), and the abundance of HNF1B also increased within each nephron (1.22-fold increase, p-value < 0.0001) compared to controls (Fig. 2m, n). Consistent with PI3K inhibition resulting in Notch signaling linked to HNF1B expression, nephrons did not upregulate HNF1B or HES1 when cultured with Notch/gamma-secretase inhibitor DAPT33,34, either with or without Ly29 (Supplementary Fig. 3a, b). The increase in HES1 upon Ly29 treatment persisted through differentiation days 13 and 14, 1 and 2 days after the removal of the inhibitor. Transiently blocking PI3K signaling therefore drives Notch signaling and HES1 protein production throughout organoid nephrons, while control organoid nephrons exhibit low and non-uniform HES1 protein levels (Supplementary Fig. 3c). Importantly, inhibition of PI3K signaling altered the structural dynamics of the organoids, leading to the emergence of JAG1+/HNF1B+ structures distributed across both the periphery and center of the organoid discs, while control organoids predominantly exhibited tubulogenesis biased towards the periphery (Fig. 2j–m and Supplementary Fig. 3c).
To scrutinize how cells differentiate from their day 12 states, we performed single-cell RNA-sequencing at day 14, two days after PI3K inhibitor removal (Fig. 3a). Differential gene expression analyses revealed that control organoid nephron cells largely follow a podocyte differentiation program, marked by enrichment of podocyte-associated genes MAFB, PTPRQ, NPHS2, PODXL, and WT1 (Fig. 3b, c and Supplementary Data 8). In contrast, day 14 Ly29-treated organoid nephron cells exhibited proximal-biased (PB) differentiation—defined here as an increase in the abundance, length, and spatial positioning of HNF4A+ proximal segments—with increased expression of human medial SSB and proximal nephron genes, such as brush-border markers DPP4 and FLRT3, along with sustained expression of HNF1B and JAG1 (Fig. 3b–d and Supplementary Data 8). The distinct expression patterns of these markers between control and treated cells suggest each condition maintains its differentiation program. Notably, HNF4A—previously undetected at day 12— was now observed in both conditions but prominent in day 14 Ly29-treated cells (Fig. 3e). Overall, these results suggest that early PI3K inhibition promotes a population of progenitor-like cells primed for HNF4A+ proximal nephron emergence.

a Single-cell RNA-seq UMAP from day 14 control and PI3K inhibitor-treated (LY294002 from days 10–12, proximal-biased, PB) kidney organoid nephrons. Cells colored by sample with marker gene annotations. b Log2 fold change (log2FC) heatmap for differentially expressed genes, calculated as the ratio of proximal-biased to control aggregate expression. Representative genes listed with a legend. c Feature plots of genes enriched in each sample from the differentially expressed gene list. d Split violin plots of differentially expressed genes. Top two rows: genes enriched in day 14 control cells; bottom two rows: genes enriched in day 14 proximal-biased cells. Plot colors match sample colors from (a). e Feature plots of selected genes for each group based on sample characteristics. For c, e human nephron (Supplementary Fig. 1a) detection is on the left, organoid plot on the right. f Immunofluorescent stains of day 14 control and proximal-biased kidney organoids. Boxed regions split and magnified. White arrowheads mark early HNF4A+ nephron regions, yellow arrowheads indicate autofluorescence. Scale bars: 10 microns. g Bulk RNA-sequencing (TPM) of select genes on days 10, 12, 14, and 18 from n = 2 whole organoids. SEM error bars shown. h, i FACS of day 18 kidney organoids from an HNF4A-YFP iPSC reporter line35 for control (h) and proximal-biased (i) conditions. Brightfield and YFP channels are shown as merged and split panels. FACS plots in duplicates with YFP+ percentages in bottom right. Scale bars: 500 microns. j Immunofluorescent stains of week 16.1 human kidneys during HNF4A+, HNF4G+ proximal tubule elongation. White arrowheads mark endothelial autofluorescence. Scale bars: 10 microns. k Immunofluorescent stain of control and proximal-biased day 18 organoids. Boxed regions magnify HNF4A detection. Scale bars: 200 microns. l Total HNF4A+ nephrons in n = 4 whole day 18 organoids. SEM error bars shown. Statistical significance by two-sided Student’s t-test. m Average area (µm2) of HNF4A+ nephrons from n = 4 whole day 18 organoids, across all positive segments. SEM error bars shown. Statistical significance by two-sided Student’s t-test. n Sum total area (µm2) of HNF4A+ nephron segments from n = 3 whole day 18 organoids.
The transition from a JAG1+/HNF1B+ state to HNF4A+ proximal cells, as we observed in vivo (Fig. 1b), is mirrored in organoids. Two different human iPSC lines confirmed onset of HNF4A expression by day 14, detected as compact, nascent structures not yet elongated into tubules (Fig. 3f and Supplementary Fig. 3d). In control organoids, HNF4A expression was observed primarily at the periphery. In contrast, PI3K inhibitor-treated organoids exhibited extensive HNF4A upregulation throughout the organoid (Fig. 3f and Supplementary Fig. 3d), indicating a broader activation of proximal fate. Bulk RNA-sequencing performed over the differentiation timeline shows that by day 18, 6 days after PI3K inhibitor removal, HNF4A transcript levels were increased 2.31-fold (p-value: 0.0318) in PB organoids compared to controls, indicating that the early upregulation in HNF1B/HNF1B and JAG1/JAG1 seen at days 12 and 14 transition into a sustained HNF4A+ proximal fate (Fig. 3g and Supplementary Data 5). To validate these findings, we used HNF4A-YFP reporter iPSCs to label proximal tubule cells with yellow fluorescent protein (YFP)35. FACS analyses at day 18 revealed a 2.69-fold increased abundance (p-value: 0.0021) of YFP+ cells in treated organoids compared to controls. Among the total dissociated organoid cells, 10.14% of PB organoids were YFP+, compared to 3.78% in controls. Consistent with day 14 observations, YFP+ proximal nephron formation remained localized to the periphery in control organoids, while HNF4A-YFP was upregulated throughout the entire organoid in day 18 PB samples (Fig. 3h, i).
In vivo, the paralog of HNF4A, HNF4G, is co-upregulated when the proximal precursor domain elongates (Fig. 3j). As PB organoid nephrons differentiated, they upregulated HNF4A/HNF4A and HNF4G/HNF4G (1.72-fold increase in transcript levels compared to controls, p-value: 0.0305) (Fig. 3g, k and Supplementary Data 5), while displaying unaltered WT1 RNA and protein levels (marking podocytes) (Fig. 3f–g and Supplementary Fig. 3d). PB organoids increased the total number of HNF4A+ segments per organoid (4.02-fold increase, p-value: 0.0195), the average size of individual HNF4A+ segments (1.30-fold increase, p-value: 0.0009), and therefore the sum area of HNF4A+ organoid nephron segments (2.83-fold increase for n = 3 organoids) (Fig. 3l–n). To verify that HNF4A+ segments in control and PB organoids correspond to proximal tubules, we co-stained for HNF4A and Lotus tetragonolobus lectin (LTL), a broadly used proximal tubule marker, observing co-detection of these markers in both conditions (Supplementary Fig. 4a). Consistent with previous finds, HNF4A+ tubules were labeled with LTL, however LTL is a less-preferred marker for human proximal tubules given broad non-specific labeling of other nephron segments36.
To determine whether HNF4A+ cell differentiation is Notch-signaling dependent, we treated HNF4A-YFP control and PB organoids with DAPT for 48 h between days 12 and 14, after Ly29 withdrawal (Supplementary Fig. 3e, f). Inhibiting Notch signaling after PI3K inhibitor treatment reduced HNF4A-YFP in both control and PB conditions. In line with our previous experiments, PB organoids formed YFP+ structures throughout, while control organoids displayed YFP signal primarily at the periphery. In DAPT conditions, YFP− structures still developed but were HNF1B+/JAG1+/HNF4A− (Supplementary Fig. 3e), indicating Notch signaling is required for the transition from HNF1B+ to HNF4A+ cells, as anticipated. PB organoids contained more HNF4A+ structures than other conditions, and controls formed more HNF4A+ structures than DAPT-treated conditions (p-values < 0.01) (Supplementary Fig. 3g). These data confirm that Notch is required for HNF4A+ proximal cell formation and suggest that PI3K inhibition promotes proximal cell fate through Notch-dependent priming of HNF1B+/JAG1+ cells.
To further test the robustness of proximal biasing and its targeting of nephron cells, we applied the approach to our recently developed induced human nephron progenitor cell (iNPC) propagation system37. iNPCs were differentiated from H1 embryonic stem cells, expanded, aggregated, and differentiated into nephrons. The resulting organoids are derived from SIX2+ iNPCs and highly enriched for nephron structures37. To stage-match iNPC and kidney organoid protocols, iNPC organoids were treated for 48 h with Ly29 between differentiation days 4–6, at which point the organoids show a gene signature characteristic of the pretubular aggregate and renal vesicle-stage nephron (PAX8+/WNT4+/CRYM+)37, equivalent to day 10 iPSC-derived kidney organoids (Supplementary Fig. 4b, c). Six days after the removal of the PI3K inhibitor, day 12 iNPC-derived organoids showed more abundant proximal HNF1BHIGH/HNF4A+/LRP2+/LTL+ segments compared to controls (Supplementary Fig. 4d–f). PB iNPC-derived organoids formed PODXL+/WT1+ podocyte-like structures and tubules displayed expected apical-basal polarities marked by ZO-1 and LAMB1 (Supplementary Fig. 4d–f). Minimal distal TFAP2A+ or loop of Henle-like SLC12A1+ structures were observed in either condition (Supplementary Fig. 4d–f). Collectively, these data show that the proximal-biasing approach is applicable to other kidney systems and confirms that the mechanism acts directly on nephron cells rather than other kidney cell populations.
Given that PI3K signal transduction can act downstream of FGF signaling, and that FGF9 is a component of the organoid differentiation protocol4,26, we tested whether decreasing FGF9 concentrations during organoid development can mimic the effect of PI3K inhibition. We explored reducing FGF9 concentrations from days 10 to 12 (Supplementary Fig. 5a, b) by decreasing concentrations from standard 200 ng/mL FGF94,26 to concentrations between 0 to 200 ng/mL. Complete removal of FGF9 severely impaired nephron differentiation, and few WT1+, HNF1B+, or HNF4A+ nephrons formed. Increasing FGF9 concentrations from 0 ng/mL to 50 ng/mL or 100 ng/mL trend-improved development of nephrons above the 0 ng/mL state, but they nevertheless remained below standard (Supplementary Fig. 5c). This suggests that lowering FGF9 concentrations does not mimic the effects of PI3K inhibition. To further test this, we simultaneously inhibited PI3K in organoids exposed to only 50 ng/mL FGF9. In these conditions, nephrogenesis was rescued to control organoid levels (p-value: 0.8209), but only in the PB condition (200 ng/mL FGF9 + 10 µM Ly29) was there a significant increase in HNF4A+ structures above standard control conditions (Supplementary Fig. 5c, p-value: 0.0270). Collectively, these findings demonstrate the importance of FGF9 for normal differentiation and decouple the proximal-biasing mechanism of PI3K inhibition and FGF9 signaling.
Enriching a functional proximal precursor identity in kidney organoids
To examine the differentiation outcomes of PB cells, we captured and integrated single-cell RNA-sequencing data from control and PB organoids across days 10, 12, 14, and through to 18 (Supplementary Fig. 6a, b and Supplementary Data 9). Consistent with previous views, the data from nephrogenic cells highlight the divergent day 12 signatures with distinct transcriptional states exhibited by controls (primed towards TARID+/OLFM3+ podocytes) and Ly29-treated nephrons biased to generate nephron tubule-forming cells (HNF1B+/LHX1+/DLL1+) (Figs. 4a, b, and S6c Supplementary Data 10). Inferred developmental trajectories from progenitors through to HNF4A+ cells suggests day 10 cells transition from progenitors and post-proximalization, gradually differentiate into proximal nephron lineages by day 18, as evident from a continuum of transcriptional states (Supplementary Fig. 6d–f). While organoid nephrogenic cells partially separated by sample timepoint, overlap was also observed, indicating that early and transient Ly29 treatment coaxed cells towards a PB state, increased the number of these cells, but did not introduce sufficient transcriptional changes to otherwise separate clusters. Highlighting the strong biasing introduced by PI3K inhibition, 70.8% of all HNF4A+ proximal tubule precursors originated from the PB condition, with similar ratios for HNF1B and JAG1 (Fig. 4c).

a Schematic of single-cell RNA-sequencing timepoints with UMAP reduction of kidney organoid nephrogenic cells, colored by samples. b UMAP reduction of nephrogenic cell clusters in kidney organoids, colored by clusters, with identifying marker gene annotations. UMAPs split by sample type of origin (untreated day 10; control days 12, 14, and 18; or proximal-biased (PB) days 12, 14, and 18) are shown at the top. c Feature plots for select nephron marker genes with insets separating control from proximal-biased cells, and scaled percent quantification of total cells expressing the gene. Whole-mount immunofluorescent stains of control (d) and proximal-biased (e) kidney organoid nephrons from differentiation days 11, 12, 14, 15, and 18, with the bottom row highlighting HNF4A protein detection. Scale bars: 10 microns. Differentiation model for control kidney organoids (f) based on (d), and for proximal-biased kidney organoids (g) derived from (e). h Time-lapse video captures of kidney organoids differentiated from a human HNF4A-YFP iPSC reporter line35 from differentiation days 15–18, showing the emergence and progression of HNF4A-YFP detection in control and proximal-biased conditions. Brightfield and YFP channels are merged. Scale bars: 200 microns. Movies are included as supplementary files. i Whole-mount immunofluorescent stains of control and proximal-biased kidney organoids from differentiation days 14 and 21. Boxed regions are magnified. Scale bars: 10 microns. j Whole-mount immunofluorescent stains of control and proximal-biased kidney organoids from differentiation days 14, 18, and 21. Boxed regions are magnified. Scale bars: 10 microns. k Quantification of nephron segments positive for indicated markers per 15X field of view at differentiation days 14 and 21. Data are from n = 3 independent organoids, including those shown in (i, j). SEM error bars shown. Statistical significance is determined using two-sided Student’s t-test. l Quantification of the average segment length (µm) for indicated proteins in day 18 control and proximal-biased kidney organoids. Data are from n = 4 independent organoids (15X), including those shown in Supplementary Fig. 7c, d. SEM error bars are shown. Statistical significance is determined using two-sided Student’s t-test.
To further understand the cellular composition of our control and PB organoid nephron cells, we compared their transcriptional profiles with in vivo nephron cells (Supplementary Figs. 1a and 6g and Supplementary Data 11). As expected, organoid nephrogenic cells largely co-clustered with human nephron cells (Supplementary Fig. 6h–i). Sample contributions for each cluster were quantified (Supplementary Fig. 6i), showing organoid podocytes (clusters 2 and 3: MAFB+, OLFM3+, NPHS2+), early proximal (cluster 12: HNF1B+, JAG1HIGH), proximal (cluster 10: HNF1B+, HNF4A+, SLC3A1+), and proximal and loop of Henle precursors (cluster 9: LRP2+, SLC3A1+, SLC12A1+) clustered with in vivo cells, while in vivo human CITED1+ nephron progenitor states (clusters 0 and 5) were poorly represented in day 10 organoids (cluster 1). This was expected since CITED1+ day 10 organoid cells also express PAX8 and LHX138, thus displaying differences to normal in vivo nephron progenitors (Supplementary Fig. 6g–j and Supplementary Data 9–11). Distinct precursor clusters were observed between day 12 control (clusters 7 and 8: PDGFRB+, PI3KHIGH), day 12 PB (cluster 4: JAG1+, DLL1+, NotchHIGH), and early human nephron (cluster 6: SNAI2+, DAPL1+) cells, pointing to transcriptional differences induced by Ly29 treatment (Supplementary Figs. 6h and 1h–i and Supplementary Data 11). Distal human nephron cells (clusters 20, 19, and 16) separated from organoid nephron cells. Collectively, these data indicate that the PB organoids primarily generate in vitro equivalents of podocyte and proximal cells.
Validating these data in individual organoid nephrons showed consistent views (Fig. 4d–g). Morphologically, PB organoids display expansion of the HNF1B+/HNF4A+ identity throughout the tubular portion of the organoid nephron. Initially, PB organoids display a subtle increase in the WT1+ domain immediately following PI3K inhibition at day 12 (1.08-fold larger, p-value: 0.0015) (Supplementary Fig. 6k–l). However, this effect is not sustained, and by day 18 control nephrons display WT1+ renal corpuscle-like structures larger than those in the PB organoids (1.42-fold larger, p-value: 0.0002) (Fig. 4d–g and S6k–l). The persistent PB organoid nephron phenotype is an elongated HNF4A+ proximal domain (Figs. 3k–n and 4e, g) compared to controls (Fig. 4d, f). HNF4A-YFP reporter35 organoids show YFP is gradually upregulated and increasing in abundance between days 15 and 18, expanding throughout PB organoids, while in control organoids, YFP+ is restricted to the periphery (Fig. 4h and Supplementary Movies 1–2).
To broadly assess nephron differentiation outcomes in PB organoids, we immunostained for proximal and distal segment-specific markers. At day 14, proximal tubule maker LRP2/Megalin (proposed HNF4A target16) was weakly detected. By day 21 LRP2/Megalin+ structures were abundant throughout PB organoids but not in controls (Fig. 4i). WT1+ structures adjacent to LRP2/Megalin+ tubules were smaller in PB organoids, consistent with earlier observations (Supplementary Fig. 6k–l). TFAP2A+ distal-like structures were sparse across both differentiation conditions, with no significant difference between control and PB organoids (Day 14: no change ± 0.149 SEM; Day 21: no change ± 0.197 SEM) (Fig. 4i, k). The spreading of nephron-forming events from periphery to center of each organoid upregulated HNF4A centrally (Supplementary Fig. 7a, b). Nephrons at the organoid periphery increased expression of transcription factors HNF4A and POU3F3 (Supplementary Fig. 7a, b), the latter detected in segment 3 (S3) in the in vivo proximal tubule27, and otherwise distally. The upregulation of POU3F3 was confirmed by our bulk RNA-sequencing (Supplementary Data 5). At day 21, SLC12A1+ loop-of-Henle-like structures were detected in both conditions, with a slight increase in abundance in PB organoids (1.63-fold higher, ±0.464 SEM). Proximal LRP2/Megalin+ and HNF4A+ structures were more abundant (LRP2/Megalin: 3.06-fold higher 0.189 ± SEM; HNF4A: 2.72-fold higher ±0.299 SEM) and elongated in PB organoids compared to controls (Fig. 4j, k). HNF1B+ and HNF4A+ tubules elongated (HNF1B+: 2.44-fold longer, p-value: 0.0002; HNF4A+: 1.95-fold longer, p-value: 0.0013), while POU3F3+ tubules did not (p-value: 0.6069), supporting a role for HNF1BHIGH proximal conditions in promoting HNF4A+ tubule formation and elongation (Figs. 4l and S7c, d).
To assess maturation of the proximal tubule cells in the PB model, we performed single-nucleus RNA-sequencing at days 21 and 27, marking 7 and 13 days after HNF4A first emerges. Integration of day 21 and day 27 PB organoid cells revealed overlap between time points, with cells belonging to either the nephrogenic or interstitial lineage, consistent with the model lacking off-target cell populations and being enriched for only metanephric mesenchyme lineages (Fig. 5a–c and Supplementary Data 12). Scrutiny of the nephrogenic lineage (nephrogenic subset of day 21 and day 27 PB organoids) showed co-clustering (Fig. 5d) and expression of genes involved in organic ion, glucose, and drug transport, suggesting PB organoid nephrons mature and express key proximal nephron solute carriers. Five out of six39 (83.3%) organic cation transporters showed expected expression in HNF4A+ cell clusters of day 21 and day 27 PB organoid nephrons, including SLC22A3/OCT3 and SLC22A2/OCT2 (Fig. 5e), the latter a known Hnf4a target16. Similarly, five out of eight39 (62.5%) organic anion transporters displayed expected regional expression within HNF4A+ clusters of day 21 and day 27 PB organoid nephrons. Of these, SLC22A6/OAT1 and SLC22A8/OAT3 are Hnf4a/Hnf4a-dependent in the mouse16 and enriched in the maturing proximal nephron in the human (Fig. 5e). The glucose and monocarboxylate cotransporter SLC5A12/SMCT2, expressed in the maturing in vivo proximal nephron, was detected, yet glucose cotransporter SLC5A2/SGLT2 only sparsely so (Fig. 5e). Comparison with the developing human nephron indicated that PB organoids recapitulate key solute transporter expression patterns. These transporters emerged within HNF4A+ cell clusters, consistent with their known co-detection with Hnf4a/HNF4A in vivo, showing strong similarity between the PB organoid transcriptional profile and the in vivo proximal nephron (Fig. 5e).

a UMAP reduction of single-nucleus RNA-sequencing of differentiation day 21 proximal-biased and day 27 proximal-biased (PB) kidney organoids. Cells are colored by cluster, with marker gene annotations. UMAPs split by sample timepoint of origin are shown at the top. b Dot plot with identifying genes for each cluster. c Feature plots for select genes, showing a matching comparison to Supplementary Fig. 1c, f. d UMAP reduction of the nephrogenic subset of day 21 proximal-biased and day 27 proximal-biased kidney organoid single-nucleus RNA-sequencing from (a). Cells are colored by sample timepoint of origin, with marker gene annotations. e Feature plots of select proximal nephron organic cation, organic anion, solute, and drug transporters. Detection of the gene in human nephron single-cell RNA-sequencing (Supplementary Fig. 1a) data is shown on the left, with the organoid plot on the right. In vivo proximal tubule segment specificity is listed. f UMAP reduction of single-cell RNA-sequencing of differentiation day 13 + 14 (27) proximal tubule-enhanced kidney organoids, with marker gene annotations, acquired from ref. 10. g Dot plot with identifying genes for each cluster. h Feature plots for select genes, showing a matching comparison to Supplementary Fig. 1cf. i Diagram of differentiation and sampling timelines for day 21 proximal-biased, day 27 proximal-biased, and day 13 + 14 (27) proximal tubule-enhanced kidney organoids. j Schematic of merging single-cell RNA-sequencing datasets: day 21 proximal-biased and day 27 proximal-biased kidney organoids (a) were integrated with differentiation day 13 + 14 (27) proximal tubule-enhanced kidney organoids in (f). UMAP reduction of resulting integration is shown, with cells colored by cluster. Split UMAP reductions showing sample of origin are shown on the right. k Feature plots of select nephron, proximal tubule, and off-target genes, split by sample of origin: proximal-biased cells (left plot) or proximal tubule-enhanced cells (right plot).
To directly assess PB organoids against a leading model of proximal tubule cells, we compared PB transcriptional profiles with data from the proximal tubule-enhanced (PT-E) kidney organoid protocol10 (Fig. 5f–h and Supplementary Data 13). Our single-nucleus RNA-sequencing at day 21 stage-matched the PT-E organoids by their time in 3D culture (14 days post-initiation), and our day 27 data stage-matched by total differentiation time (27 days, equivalent to the combined 13 + 14 days PT-E protocol10; Fig. 5i). Both organoid types contained SLC12A1+ loop of Henle-like cells and WT1+ podocyte-like cells (Fig. 5a–c, f–h, k), but distal nephron markers TFAP2A, AQP2, and GATA3 were strongly detected in PT-E organoids, forming distinct groupings that were absent in PB organoids (Fig. 5k and Supplementary Data 12–13). As expected, neither PB nor PT-E organoids displayed a pronounced muscle differentiation signature (MYOG⁺) (Fig. 5b, g, k).
Extensive co-clustering was observed upon merging day 13 + 14 PT-E organoid cells with day 21 and day 27 PB organoid cells (Fig. 5i, j and Supplementary Data 14). To assess fidelity to normal development, we leveraged HNF1B- and HNF4A-correlated genes to delineate ‘early’ and ‘late’ proximal nephron programs (early: genes detected in human clusters 21, 24, and 20; late: genes absent in human cluster 21 but detected in clusters 24 and 20) (Supplementary Fig. 2a and 8a–c). All early and late proximal genes were present in both models, though detection frequencies varied. Early genes such as zinc transporter SLC39A5/ZIP5, brush-border marker VIL1, and monooxygenase factor FMO1 were frequently detected in both models, while genes such as sodium-sulfate cotransporter SLC13A1/NAS1, Hnf4a-dependent16 transcription factor TFEC, and deamination factor A1CF were more frequently detected in PB organoid cells (Supplementary Fig. 8a, b). Late proximal tubule genes followed a similar trend. PB organoids exhibited higher detection frequencies of key late markers, including organic anion transporter SLC22A8/OAT3 and iodide transporter SLC5A8/AIT (Supplementary Fig. 8c), while BHMT and AFP were detected at similar levels across cells from both organoid origins. Notably, these genes were detected in day 21 PB cells and remained detectable at day 27, indicating that transcriptional states emerge by day 21 and are maintained through to day 27 in the PB model.
Collectively, these data demonstrate that PB organoids establish broad proximal tubule transcriptional programs by differentiation day 21, which are maintained through day 27, while displaying minimal off-target and non-proximal populations.
Modeling proximal tubule physiologies and cisplatin-induced injury in proximal-biased nephrons
Kidney organoid physiologies are often assessed by cells’ ability to absorb and accumulate compounds selectively transported by solute carriers. Uptake of dextran and albumin is controlled by HNF4A-target genes LRP2/Megalin and CUBN16,40,41,42,43. Given that PB nephrons display increased and sustained expression of LRP2 and CUBN compared to controls (LRP2: 1.32-fold higher TPM; CUBN: 1.48-fold higher TPM, Supplementary Data 5; single-nucleus RNA-sequencing at day 21 and day 27, Fig. 5d, k), this raised the possibility of amplified proximal tubule function. To scrutinize this, we performed live dextran and albumin uptake assays using HNF4A-YFP reporter organoids35 co-cultured with fluorescently tagged LRP2/Megalin antibodies to label proximal tubules and apical surfaces. Day 21 PB organoids showed increased uptake of fluorescently tagged dextran and albumin in LRP2+/HNF4A-YFP+ tubules (Fig. 6a, b 1.61-fold increase, p-value < 0.0001; Supplementary Fig. 8d–e, 2.92-fold increase, p-value < 0.0001) compared to controls. Albumin and dextran were only detected within HNF4A-YFP+, LRP2/Megalin+ regions of nephrons with strong apical antibody labeling, indicating apical accessibility in live organoids and suggesting that proximal organoid nephrons are competent to perform uptake in standard assays commonly used to assess proximal identity10,40,43,44,45 (Figs. 6a and S8d).

a Live imaging of control and proximal-biased HNF4A-YFP35 kidney organoids on day 21 co-cultured with Alexa 647-Dextran and Alexa 594-LRP2 antibody. Split channels shown. Arrowheads indicate autofluorescence (yellow) and true signal (white). Scale bars: 100 microns. b Quantification of HNF4A-YFP+ segments co-detected with 594-LRP2 and 647-Dextran. Averaged from 3 images/organoid, n = 3 organoids. SEM error bars shown. Statistical significance by two-sided Student’s t-test. c Whole-mount immunofluorescence of uninjured and cisplatin-injured kidney organoids on day 21, with HNF4A+ nephron regions magnified. Scale bars: 10 microns. d Timeline of kidney organoid injury strategy. e Quantification of HAVCR1+ signal in control and proximal-biased nephrons on day 21 post-injury (n = 3 replicate organoids). SEM error bars shown. Statistical significance by two-sided Student’s t-test. f Quantification of HNF4A+ nephron segments with HAVCR1 expression from (c). Data from 4 images/organoid (20X), n = 2 replicates. SEM error bars shown. Statistical significance by two-sided Student’s t-test. g Immunofluorescence of uninjured and cisplatin-injured proximal-biased kidney organoid nephrons on day 21. Dashed white line marks uninterrupted tubule lumen. Scale bars: 10 microns. h Individual continuous lumen length quantifications from (g), n = 4 organoids per condition. SEM error bars shown. Statistical significance by two-sided Student’s t-test. i Immunofluorescence of uninjured and cisplatin-injured proximal-biased kidney organoid nephrons on day 21. Yellow arrowheads: HNF4A+ tubule cells with low HNF4A and high γH2AX. White arrowheads: cells with high HNF4A and low γH2AX. Split channels for HNF4A and γH2AX shown. Scale bars: 10 microns. j Immunofluorescence of uninjured and cisplatin-injured kidney organoid nephrons on day 21. White arrowheads: SOX9+ interstitial cells. Scale bars: 10 µm. k HNF4A and SOX9 intensities from (j). Intensities from individual cells in duplicates, min-max normalized. l Kidney organoid single-injury timeline for organoids in (m). m Immunofluorescence of uninjured and cisplatin-injured proximal-biased kidney organoid nephrons on days 19, 21, and 25. Arrowheads indicate cells with high HNF4A and low SOX9 (white), and low HNF4A and high SOX9 (yellow). Scale bars: 10 microns. n Schematic model of single-injury data, depicting gradual SOX9 activation, collapsing lumens, and loss of HNF4A in organoid nephrons.
The single-nucleus RNA-sequencing data from days 21 and 27 also revealed that PB cells express physiology-imparting genes, including those responsible for transporting cisplatin (Fig. 5e), a chemotherapeutic with severe nephrotoxic side-effects in the proximal convoluted tubule46,47,48,49,50,51,52,53,54,55,56. Recent studies show that organoids are sensitive to cisplatin and respond by activating expression of early proximal tubule injury marker KIM1/HAVCR1 and display DNA damage akin to that observed in patients12,13,14. Given that the PB model expresses organic ion and copper transporters OCT2/SLC22A2, OAT1/SLC22A6, OAT3/SLC22A8, CTR1/SLC31A1 (responsible for uptake) and MATE1/SLC47A1 and MRP2/ABCC2 (shuttling cisplatin to lumen)57,58,59,60,61,62,63,64 (Fig. 5e), we tested whether PB cells model cisplatin-induced injury by treating PB and control organoids with two low staggered doses of cisplatin (5 µM) and assayed their response 3 days after initial injury (Fig. 6d). Cisplatin-injured PB organoids showed strong detection of KIM1/HAVCR1 in HNF4A+ cells compared to injured and uninjured control, and uninjured PB organoids (all p-values < 0.0001) (Fig. 6c, f) and 1.56-fold higher protein abundance (p-value: 0.0154) (Fig. 6e). Cisplatin-injured nephrons also displayed loss of tubular integrity and apicobasal polarity as evidenced by depolarized distribution of apical and basal cell-surface markers ATP1A1 and LAMB1, respectively (Fig. 6g). Luminal integrity was lost in injured HAVCR1+ organoids, with collapsed apical-basal polarities and shorter lumen surfaces, while uninjured samples preserved tubular architecture and maintained continuous open lumens (Fig. 6h; 2.82-fold longer uninterrupted tubular lumens, p-value < 0.0001).
Following injury, Hnf4a/HNF4A are mosaically downregulated in proximal tubules65,66,67,68,69, and cisplatin-injured PB nephrons displayed HNF4A protein levels anticorrelating with DNA damage response marker γH2AX12,13,14 (r = −0.67, p-value < 0.0001) (Fig. 6i). Interstitial γH2AX was also detected, highlighting broad DNA damage throughout organoids as previously reported14. In the mouse acute kidney injury (AKI) model, injured proximal tubular cells activate Sox968,70,71,72, and under conditions of ischemia-reperfusion injury and rhabdomyolysis-induced AKI, a persistent Sox9+ state is tightly linked to maladaptive repair, tubular degeneration, and fibrosis68. PB kidney organoid nephrons activated SOX9 in response to cisplatin-induced injury with extensive co-detection of HNF4A and SOX9, unlike controls (Fig. 6j, k). Reducing treatment to a single cisplatin dose generated a mosaic response producing HNF4AHIGH/SOX9LOW cells 1 day post-injury, HNF4AMED/SOX9MED cells 3 days post-injury, and a broadly HNF4ALOW/SOX9HIGH state 6 days post-injury, indicating dynamic changes over time (Fig. 6l–n). Collectively, these data highlight the potential of the PB nephron proximal tubule cell model as a sensitive and rapid system to delineate nephrotoxic and acute kidney injury.
The PB organoid nephron, therefore, emerges as a robust method for generating in vivo-mimicking proximal tubule cells that replicate physiological processes and respond to injury-driving mechanisms.
Discussion
We developed an in vivo informed approach to mimic human proximal nephron precursor cells in organoids derived from multiple human iPSC lines by delineating early in vivo human proximal tubule development, comparing it to existing kidney organoid models, and thereafter driving changes in cell states to bias differentiation outcomes. Our new model generates nephron-like structures in thin self-organizing discs that preserve nephron 3D complexities, and we take advantage of the plasticity in nephron axial positional identities25 to drive organoid nephrons towards a proximal-biased (PB) precursor-like state where organoid cells sequentially activate transcription factors and function-imparting proximal tubule genes. Our approach directly targets nephron cells, as iNPC-derived organoids—initially lacking interstitial or other non-nephron cell types—are sensitive to proximal-biasing. Our system offers a reproducible model with improved fidelity to in vivo cells and expression of developmentally appropriate genes, including OCT and OAT family members, and a host of physiology-imparting solute carriers, making it a tool to study development, congenital disease, injury, and physiology.
We demonstrate that in vivo, human proximal precursors develop through a series of cell-state transitions marked by activation of JAG1, followed by HNF1B, and production of HNF4A in the upper bend of the medial S-shaped body nephron. This is consistent with our previous studies showing human nephrons activate HNF4A in the medial nephron segment18, and work by others in the mouse model23. Genetic evidence from mice shows that Notch signaling is required for proximal nephron differentiation, nephron segmentation, and nephron number22,73,74,75,76,77. Inhibition of PI3K resulted in a transient upregulation of Notch target genes and sustained upregulation of HNF1B. Inhibiting PI3K and simultaneously blocking Notch signaling in PB organoids prevented HNF1B expression, consistent with Notch1; Notch2 compound knockout mice failing to upregulate Hnf1b in nephrons22. Blocking Notch at any point between early organoid nephron formation and HNF4A+ cell emergence was sufficient to prevent proximal-biasing and eventual emergence of HNF4A+ cells. In vivo, Hnf1b binds to and is required for expression of Hnf4a23, with a conserved role for human HNF1B indicated by mutations in Hnf1b/HNF1B being linked to congenital kidney anomalies affecting tubulogenesis, as well as studies in kidney organoids24,36,78,79,80,81. In our model, increased HNF1B/HNF1B expression and protein levels preceded detection of HNF4A/HNF4A, concurring with in vivo and in vitro studies showing HNF1B is essential for proximal nephron cell-fate development23,24. Consistent with Notch signaling being required for the proximal nephron program, inhibition of Notch at day 12 reduced JAG1, HES1, and HNF1B protein levels in both control and PB settings. Of note, while the PI3K pathway can act downstream of FGF signaling (KEGG pathway: hsa04151)82,83,84, lowering FGF9 failed to replicate PI3K inhibition and instead broadly impaired nephron development and organoid cell growth, underscoring the known role of FGF9 as a growth factor for early kidney organoid nephrons85.
While Hnf4a/HNF4A and Hnf4g/HNF4G are co-expressed in proximal tubules in vivo8,18,27, evidence suggests a non-redundant relationship, with Hnf4a knockout mice and HNF4A knockout kidney organoids having prominent phenotypes15,16,17. Loss of HNF4G, however, only moderately alters kidney organoid development15, and Hnf4g knockout mice display a phenotype in the intestine only when removed alongside Hnf4a, likely due to redundant roles of Hnf4 factors in enterocyte maturation86,87. As part of our comparative analyzes between in vivo and in vitro, we sought to understand how Hnf4a-dependent genes are expressed during development and in adult nephrons. It is interesting to note that a group of Hnf4a-dependent genes are not expressed in the adult, indicating temporally dynamic roles for Hnf4a, or that gene expression is mediated by indirect mechanisms requiring, for example, co-factors. Given the clear role of Hnf4a in injury and repair mechanisms, this is an area needing further scrutiny. Overall, our analyses point to that human orthologs of Hnf4a-dependent genes are usually expressed at much lower frequencies in organoids, which in turn suggests that most organoid systems at present only partially mirror the in vivo proximal program. However, the onset of HNF4A and HNF4G expression occurs within a short timeframe in the S-shaped body nephron, and PB organoids do express both genes as the nephron tubules elongate, which mirrors the temporal in vivo sequence. Other kidney models generating HNF4A+ proximal cell-states have previously enriched kidney organoid cultures for the nephron progenitor state and subsequently optimized culture conditions to permit proximal-like cell development10. In contrast, the PB system imposes a specific signaling event that shifts cells to a proximal developmental program, thus enabling reproducibility, synchronicity, and application across cell lines, providing control over differentiation outcomes.
In all kidney organoid systems, it is unclear how JAG1 is initially activated in the forming nephron. In vivo, it is thought that Jag1 is bound by and sensitive to β-catenin mediated transcription88,89 and Jag1/JAG1 expression initiates in cells adjacent to the WNT9B secreting collecting duct21,75,90,91. Recent spatial transcriptomic analyses of the pretubular aggregate indicate that JAG1 is activated in a small subset of cells closest to the collecting duct within a 1–2 cell-layer thick domain, suggesting a short-range collecting duct-derived ligand is responsible for JAG1 expression91. However, simultaneous inhibition of PI3K and activation of WNT/β-catenin results in strong and broad upregulation of Jag1 throughout the nephron progenitor population, indicating that Jag1 is controlled via multiple integrated pathways29. In the organoid setting, it remains unclear how the brief dosing with CHIR at day 7 (a GSK3-β antagonist and WNT/β-catenin agonist) can control the emergence of JAG1+ nephron aggregates at day 10; a subject warranting further investigation. In favor of multiple pathways controlling JAG1 transcription during induction and nephron patterning, in vivo JAG1 expression follows an intriguing wave-like pattern where it is initially activated in collecting-duct adjacent pretubular aggregate cells and thereafter moves proximally through the nephron and is downregulated distally, suggesting multiple layers of control over this Notch ligand19,21,91. Identifying the mechanisms that regulate JAG1 expression and activities will therefore be critical for understanding nephron induction and subsequent proximal-distal patterning.
In vivo, proximal tubule cells exhibit a well-defined apical-basal polarity, appropriately positioned solute carriers and transporters, vascularization, intraluminal solute flow, and sensitivity to nephrotoxic compounds1,92,93,94. Kidney organoids display accessibility to apical surfaces, as evidenced by live culture of organoids with fluorescently conjugated LRP2 antibodies that bind to LRP2/Megalin at the apical proximal tubule surface10. Here we replicate this using HNF4A-YFP+ organoids and detect strong labeling of anti-LRP2-594nm and anti-LRP-647nm at the apical surfaces of HNF4A-YFP+ tubules, but not elsewhere, and co-labeling with fluorescently tagged albumin and dextran. These uptake assays, which rely on LRP2/Megalin40,43, are widely used as proxies for proximal tubule function in kidney organoids10,40,43,44,45. In our model, albumin and dextran accumulated apically within HNF4A+ structures, consistent with apical accessibility and reflective of proximal nephron structure and function.
Cisplatin-induced nephron injury has been demonstrated in organoids by several groups10,12,13,14,95 in systems with variable expression of cisplatin transporters and differing abundance of maturing proximal tubules. The PB model demonstrates expression of solute carriers capable of transporting cisplatin46,96,97, including OCT2/SLC22A2, OAT1/SLC22A6, OAT3/SCL22A8, CTR1/SLC31A1, MATE1/SLC47A1, and MRP2/ABCC2 (Fig. 5e). PB nephrons are sensitive to cisplatin early in the differentiation protocol (day 18), at a low concentration (5 µM), in single doses, and in a short timeframe (72 h), making the model suitable for screening experiments requiring short dosing plans and endpoints. While expression of HAVCR1/KIM1 was specific to HNF4A+ cells, as expected, DNA-damage marker γH2AX was detected throughout organoids, corroborating recent findings of potential broad cellular toxicity14. As injured PB nephrons upregulate SOX9, display apicobasal tubular collapse, and resolve into a persistent SOX9+ state 6 days post-injury, this resembles the recently identified Sox9ON-ON switch that marks maladaptive repair in mice68. Closer scrutiny of the transcriptional state and dynamics of SOX9 activation and HNF4A downregulation, coupled with experiments critically assessing the regenerative potential of PB organoids will improve understanding of strengths and limitations of modeling cisplatin-induced injury in organoid-derived proximal tubules. Further, while cisplatin transporters are now detected, it will be important in future experiments to determine how they function in this model.
The current PB model represents a cell-state that we best estimate to resemble a post-capillary loop stage nephron based on the expression of solute carriers during the day 21-27 period. A full complement of mature nephron markers, such as SGLT2/SLC5A2, remains elusive in kidney organoids, and our data suggest that generating a mature proximal tubule will require further modifications, including tuning of culture conditions to drive maturation, elongation, and separation of proximally distinct segments. In comparison to other PT-enriching models10,98, the PB models display minimal development of non-kidney lineages such as cartilage, neuron, and muscle, and addressing why these are generated in some protocols and not others remains an important future goal.
Beyond manipulation of pathways, the PB model currently lacks physiological properties such as intraluminal flow, vascularization, culture conditions optimized for long-term expansion, as well as inclusion of potentially supportive interstitial cell-types. Evidence shows that sorted proximal tubule-like cells from human iPSC-derived kidney organoids can be cultured for months99,100, introducing the possibility to generate PB organoids for long-term tubule culture or scaling for large-scale screens using, for instance, bioreactor approaches36. Additionally, bioengineered chips offer the possibility of highly controlled studies of pharmacological drug uptake101. These chip technologies have further allowed for the introduction of nephrons with vasculature to model renal physiologies102, and collectively these methods and the rapidly developing kidney organoid field contribute to the expanding toolkit available for kidney research and complement our developmental-mimicry approach.
Methods
Human kidney samples
Developing human samples were collected under Institutional Review Board-approved protocols (USC-HS-13-0399 and CHLA-14-2211). Following the patient decision for pregnancy termination, informed consent for donation of the products of conception for research purposes was obtained, and samples were collected without patient identifiers. The person obtaining informed consent was different than the physician performing the pregnancy termination procedure, and the decision to donate tissue did not impact the method of pregnancy termination. Developmental age was determined according to the American College of Obstetrics and Gynecology guidelines. The kidney samples ranged from 14 to 17 weeks of gestation with no sex reported. Intact samples within the kidney capsule were analyzed. Samples were transported on ice at 4 °C in high glucose DMEM (Gibco, 11965-118) supplemented with 10% fetal bovine serum (Genesee Scientific, 25–550) and 25 mM HEPES (Gibco, 15630080).
Human induced pluripotent stem cell lines
An iPSC line #3C15 FiPSC, derived from human male foreskin fibroblasts (BJ fibroblasts, ATCC, CRL-2522), was generously provided by Dr. Amy L. Ryan (University of Iowa; formerly USC). This line has been used in prior studies and protocols to generate human lung airway tissue, including induced pulmonary neuroendocrine cells103. An HNF4A-YFP reporter iPSC line (male) was obtained through the Washington University Kidney Translational Research Center (KTRC) and the (Re)Building a Kidney (RBK) Consortium. Details on the HNF4A-YFP iPSC line can be found elsewhere35,104. An H1 hESC-derived iNPC line37 was generously provided by the lab of Dr. Zhongwei Li and used to make iNPC-derived kidney organoids following an established protocol37.
iPSC maintenance
Matrigel-coated plate preparation
DMEM (Corning, 10-017-CV) was aliquoted into a 15 mL conical vial, and 120 µL of Matrigel (Corning, 354277) was added to make a 1% Matrigel mix. After thorough mixing, 2 mL of 1% Matrigel was pipetted into each well of a 6-well plate (or 1 mL/well for a 12-well plate). Matrigel plates were then incubated at 37 °C/5% CO2 overnight before use.
Biolaminin 521 LN-coated plate preparation
DMEM was aliquoted into a 15 mL conical vial, and 300 µL of biolaminin 521 LN (Biolamina, LN521) was added to make a 5% biolaminin 521 LN mix. After thorough mixing, 0.5 mL of 5% biolaminin 521 LN was pipetted into each well of a 12-well plate. Biolaminin 521 LN plates were then incubated at 4 °C overnight before use.
iPSC expansion and maintenance
iPSCs were thawed in Essential 8 media (Thermo Fisher Scientific, A1517001) on 1% Matrigel-coated plates. Media was initially supplemented with 10 µM Y-27632 (Rho kinase inhibitor from Tocris, 1254) for 24 h after thawing iPSCs. Media was changed every 24 h until cells reached 70–80% confluency (3 days). For freezing, when iPSCs reached 70–80% confluency, cells were collected for passage as described below and resuspended into a mix of 50% Essential 8, 40% KnockOut Serum Replacement (Thermo Fisher Scientific, 10828010), and 10% DMSO (Millipore Sigma, D5879). Cells were then aliquoted into 2 mL cryovials at 500,000 cells/vial, gradually frozen at −80 °C overnight, and then transferred to liquid nitrogen storage.
Directed differentiation to generate kidney organoids
Differentiation protocols were developed based on published protocols26,105 and adapted in our laboratory. Each biological replicate was generated from a distinct frozen vial of iPSCs. On reaching 70–80% confluency (3 days), iPSCs were rinsed with 1xPBS (Thermo Fisher Scientific, 10010049) and then incubated in TrypLE Select Enzyme (Thermo Fisher Scientific, 12563011) for 6 min at 37 °C/5% CO2. The enzymatic reaction was neutralized using a volume of Essential 8 media 2X that of TrypLE. Cells were collected and resuspended in Essential 8 media supplemented with 10 µM Y-27632, with 15,000 cells/well plated onto a 5% biolaminin 521 LN-coated 12-well plate. 6 h after plating, differentiation was initiated by changing culture media to TeSR-E6 (Stem Cell Technologies, 05946) supplemented with CHIR99021 (Tocris, 4423). Briefly, culture medium was supplemented with CHIR99021 for 5 days (exact concentration and duration are dependent on cell line being used), followed by 2 days with 200 ng/mL FGF9 (R&D Systems, 273-F9) and 1 µg/mL Heparin (Millipore Sigma, H4784). At day 7, the cells were detached using TrypLE as described above and resuspended in TeSR-E6 supplemented with 10 µM Y-27632. 200,000 cells were seeded into each well of a round-bottom non-adhesive 96-well plate. Organoids were manually transferred to a 0.4 µm pore culture plate (Corning, 3450; Stem Cell Technologies, 100–1026). Organoids received a pulse of TeSR-E6 supplemented with CHIR99021 for 1 h; media was then switched back to TeSR-E6 supplemented with 200 ng/mL FGF9 and 1 µg/mL Heparin until day 12. Proximal-biased kidney organoids were cultured in TeSR-E6 supplemented with 200 ng/mL FGF9, 1 µg/mL Heparin, and 10 µM LY294002 (Tocris, 1130) for 48 h between differentiation days 10 and 12. Untreated control organoids were cultured with 200 ng/mL FGF9, 1 µg/mL Heparin, and DMSO vehicle. From day 12 onward, organoids were cultured in TeSR-E6 alone.
Small-molecule inhibitor screen and culture optimization
For all small molecule inhibitors used in this study, initial screens were performed on kidney organoids to determine the optimal dosage and duration of culture. To determine optimal dosage of a molecule, kidney organoids were cultured for 48 hours between differentiation days 10 and 12 in TeSR-E6 supplemented with the molecule at varying concentrations. Concentrations tested used insights from prior studies25,29. After an ideal concentration was established, to determine optimal duration of treatment with the small molecule, separate differentiation experiments were set up with the small molecule from differentiation days 10 to 18, as well as differentiation days 12 to 18. For all optimization screens, time-course bright-field imaging was performed, and organoids were collected for whole-mount immunofluorescent analyses on differentiation day 18.
Single-cell preparation of kidney organoids
Approximately 6 organoids from multiple independent wells of both untreated and treated (48 h culture with 10 µM LY294002 between days 10 and 12) conditions were collected at differentiation days 10, 12, 14, and 18 for single-cell RNA-sequencing, from separate differentiation batches. The organoids were dissociated using Accumax (Stem Cell Technologies, 07921) at 37 °C/5% CO2 for 35 min and pipetted twenty times with a P-1000 wide-bore pipette tip every 5 min. For FACS analyses, the same number of organoids from separate differentiations were dissociated as previously described27 using 10 mg/mL Native Bacillus licheniformis Protease (Creative BioMart, NATE-0633), 2.5 mg/mL collagenase type IV (Worthington Biochemical, LS004186), and 50 µg/mL DNase1 (Worthington Biochemical, LS002058) in 1xPBS−/−. FACS data were processed using flowCore (Ellis et al., 2024)106. Dissociations were neutralized by using a 2X volume of AutoMACS Running Buffer (Miltenyi Biotec, 130-091-221). Cells were pelleted by centrifugation at 4 °C for 3 min at 300 × g. After resuspending the pellet in 2 mL of AutoMACS Running Buffer, cell suspensions were strained through 40 µm strainer (VWR, 21008-949), washing the filter with an additional 1 mL AutoMACS. The flow-through was pelleted, again at 300 × g at 4 °C for 3 min. Cells were gently resuspended in AutoMACS Running Buffer supplemented with 14 µM DAPI (Thermo Fisher Scientific, D1306) and 5 µM DRAQ5 (Thermo Fisher Scientific, 62-254) and sorted on an ARIA I FACS at a low flow rate. Viable cells (DRAQ5-positive, DAPI-negative) were collected into a 1.7 mL low binding tube (Corning, 3207).
Single-nucleus preparation of kidney organoids
For single-nucleus RNA-sequencing, the same number of organoids from multiple independent wells were snap-frozen in liquid nitrogen before nuclei isolation as previously described107. Briefly, organoids were transferred to a 1.7 mL low binding tube, centrifuged to form a pellet, and snap-frozen in liquid nitrogen. Flash-frozen organoids were dounced in cold Nuclei EZ Lysis Buffer (Sigma, N3408) supplemented with protease inhibitor (Roche, 05892791001). Homogenization was performed using a loose pestle and tight pestle system (Sigma, P0485) with 15 loose-dounce strokes, followed by filtration through a 200 µm strainer (pluriSelect, 43-50200). The homogenate was then subjected to an additional 5 strokes with the tight pestle and incubated on ice for 5 min. The suspension was filtered through a 40 µm strainer (pluriSelect, 43-50040) and centrifuged at 500 × g for 5 min at 4 °C in a swinging-bucket centrifuge. After removing the supernatant, the nuclei pellet was resuspended in just Nuclei EZ Lysis Buffer, incubated on ice for an additional 5 min, and centrifuged again. The final nuclei pellet was resuspended in chilled 1X Nuclei Buffer (10x Genomics, #2000297) and filtered through a pre-wetted 10 µM filter (pluriSelect, 43-50010). All reagents contained Protector RNAse inhibitor (Promega, N2615) at a final concentration of 1U/µL and DTT (Sigma, 6465663) at a final concentration of 1 mM.
Single-cell and single-nucleus RNA-sequencing data collection and analyses
SPLiT-seq single-cell processing and sequencing
Sorted, single, live cells were immediately processed and fixed using Evercode Cell Fixation kit (Parse Biosciences, ECF2001). Fixed cells were then stored at −80 °C until all samples were ready to be processed with the Evercode WT v2 single-cell RNA-sequencing platform (Parse Biosciences, ECW02030). After recovery of barcoded cells, cDNA was cleaned up and amplified by PCR, as per Evercode WT v2 platform protocols. cDNA quality was examined at multiple points on a 4200 TapeStation (Agilent) for yield and quality assessment. Paired-end sequencing was performed on the NovaSeq X Plus PE150 (Novogene).
Single-nucleus processing and sequencing
Single-nucleus RNA-sequencing libraries were generated using the 10X Genomics Chromium Next GEM Single Cell Multiome reagents (PN-1000283) following the manufacturer’s protocols (10x Genomics; CG000338 Rev E). Briefly, freshly prepared single-nuclei suspensions were counted on a Countess 3 FL cell counter and ~9000 nuclei per condition went through 1 h of transposition at 37 °C followed by GEM generation and barcoding on a Chromium X machine with immediate subsequent RT reaction. cDNA was generated from 7 preAmp cycles and 8 cDNA cycles. GEX libraries were made from fragmented cDNA and 12 SI cycles. Sequencing was performed on a NovaSeqX 10B machine with 100 cycle Illumina flow cells at the Translational Genomics Center, Children’s Hospital Los Angeles (CHLA) Center for Personalized Medicine.
Single-cell RNA-sequencing analyses
From fastq files, demultiplexing, quality control, alignment to reference genome (hg38 Ensembl 105 annotation), and generation of count tables were done using splitpipe (Parse Biosciences). The Seurat 4.0 package was used for single-cell RNA-sequencing analyses108,109,110,111,112. To filter out low-quality cells, we kept cells with between 500 and 13,000 features, between 100 and 200,000 RNA counts, and less than 35% mitochondrial gene content. Sample datasets were integrated using Seurat integration functions across organoid time-points and experimental variables113. 50 PCs were used to calculate cell relationships, cluster assignment, and UMAPs. After quality control, the organoid single-cell dataset included 9686 cells from day 10, 22,229 cells from control samples (days 12, 14, and 18), and 24,405 cells from proximal-biased samples (days 12, 14, and 18). For the percentages shown in Fig. 4c, which represent the proportion of cells expressing each gene that originated from the control and proximal-biased samples, these values were scaled to normalize for differences in the total number of cells post-quality control between the control and proximal-biased samples. Differentially expressed genes of each cluster were identified using the FindAllMarkers function, selectively returning genes with expression in at least 25% of cells within the cluster (min.pct = 0.25) and with a minimum fold-change of 0.25 (logfc.threshold = 0.25), while restricting the output to only positively expressed markers (only.pos = TRUE). Nephrogenic cell-clusters were identified based on expression of nephron lineage and nephron cell-fate markers, and interstitial lineages were similarly separated by their gene enrichment. Nephron-forming cells were selected by serial clustering and subsetting while monitoring cluster-based quality metrics. Single-cell RNA-sequencing data are deposited to the GEO database under accession number GSE264678.
Single-nucleus RNA-sequencing analyses
Single-nucleus RNA-sequencing data were processed using Cell Ranger (v9.0.0) and the Seurat 5.2 R package108,109,110,111,112. Raw FASTQ files were processed using the cellranger count command, aligning to the pre-built human reference genome (GRCh38-2024-A). All minimum QC thresholds were achieved from sequencing with an average GEX saturation of 80% and greater than 3000 median gene expression per nucleus. Feature-barcode matrices were subsequently imported into Seurat using the Read10X function. Quality control was performed in Seurat as described above. 30 PCs were used to calculate cell relationships, cluster alignment, and UMAPs. After quality control, the organoid single-nucleus dataset included 6265 cells from the day 21 proximal-biased sample and 6637 cells from the day 27 proximal-biased sample. Differentially expressed genes for each cluster were identified, and the nephrogenic lineage was subset, each as described above. Single-nucleus RNA-sequencing data are deposited to the GEO database under accession number GSE290931.
In vivo kidney datasets
In vivo datasets of human week 1418 (GSE139280) and human week 178 (GSE124472) were analyzed independently (Supplementary Fig. 1) and were used for comparison to single-cell and single-nucleus RNA-sequencing profiles of the nephrogenic subsets of differentiating organoids. To identify genes that are co-expressed with HNF1B and HNF4A, a Pearson correlation test was performed for each factor, and visualized against clusters 9 (JAG1+, HNF1BLOW, HNF4A-), 21 (JAG1+, HNF1B+, HNF4ALOW), 24, and 20 (JAG1LOW, HNF1B+, HNF4A+), which served as the early medial to proximalizing lineage (Figs. S1 and S2). The expression of these genes in vivo and in corresponding clusters of reanalyzed kidney organoid nephrons (Supplementary Fig. 1; assembled as described below) was analyzed in Supplementary Fig. 2. Lastly, this human single-cell RNA-sequencing dataset was integrated with the proximal-biased organoid dataset using RunCCA108,109,110,111,112.
In vitro organoid datasets
To unbiasedly assess transcriptional profiles of proximalizing nephron cells, we assembled and reanalyzed differentiating kidney organoids generated from multiple sources. Data from day 25 (GSE114802)6; days 7, 15, and 29 (GSE136314)7; days 16 and 28 (GSE124472)8; and day 26 (GSE118184)9 organoids were reanalyzed. Raw data from each dataset were acquired and analyzed individually as above before being integrated. This integrated dataset was subset into nephrogenic cell types for direct comparison to in vivo nephrogenic cells (Supplementary Fig. 1). Here, clusters 7, 6, 14, 12, 8, 18, and 3 served as the proximalizing lineage (Supplementary Fig. 2). Separately, we re-analyzed published single-cell RNA-sequencing data from organoids from a protocol generating proximal tubule-enhanced kidney organoids (GSE184928)10. These data were acquired and re-analyzed independently.
Integration and batch effect correction
Single-nucleus RNA-sequencing data from day 21 proximal-biased and day 27 proximal-biased kidney organoids were merged and integrated with the single-cell RNA-sequencing dataset from days 13 + 14 (27) proximal tubule-enhanced organoids10 using sctransform to mitigate batch effects and integrated using IntegrateLayers with the canonical correlation analysis (CCAIntegration) method in Seurat 5.2108,109,110,111,112.
Pseudotime reconstruction of lineages
Podocytes were subset from the organoid nephron single-cell RNA-sequencing dataset for a more streamlined developmental trajectory analysis from a day 10 equipotent lineage progenitor cell to a day 18 proximal-biased nephron cell. Monocle 3 was used to reconstruct the differentiation trajectory114,115,116,117.
Gene expression heatmaps
Gene expression heatmaps comparing organoids at individual timepoints were generated using the AggregateExpression function in Seurat 5.2108,109,110,111,112 on the list of all differentially expressed genes identified in the dataset. DEGs were calculated as described above and by setting the cluster identifier to the sample, enabling direct comparison of samples rather than cluster numbers. For the heatmap displaying three samples (day 10, day 12 control, and day 12 proximal-biased), Z-scores were calculated from the aggregated expression values and visualized using the pheatmap (Klode, 2018) R package. For the remaining heatmaps, where only two samples were compared at each timepoint (control vs. proximal-biased), aggregated expression values were used to calculate log2 fold changes (log2FC) of the proximal-biased sample relative to the control. These log2FC values were subsequently ordered and plotted as heatmaps using pheatmap (Klode, 2018).
Other datasets referenced
To analyze Hnf4a-dependent gene expression in kidney organoids, we acquired bulk RNA-sequencing data from control and Hnf4a-mutant P0 mouse kidneys (GSE144772)16. Bulk RNA-sequencing fastq files from two control and two mutant (Osr2Cre-driven deletion of Hnf4a) P0 kidneys were acquired. Sample quality was assessed using fastqc. Using STAR version 2.7.10a, reads were aligned to the mouse mm10 genome, Ensembl release 102. Mapping quality was appraised using qualimap. Resulting.bam files were then used to generate a feature counts table using featurecounts. A list of 257 genes that are significantly downregulated in the Hnf4a-mutant kidney (DESeq2 comparing control vs. mutant kidneys, padj < 0.01, log2FC ≥ 1.5) were used for analysis28 (Supplementary Fig. 2). Separately, a list of 415 genes that are highly expressed and enriched in the adult mouse proximal tubule27 (GSE129798; Kidney Cell Explorer; https://cello.shinyapps.io/kidneycellexplorer/; https://github.com/kimpenn/kidneycellexplorer) were used for analyses in Supplementary Fig. 2. To generate the heatmaps presented in Supplementary Fig. 2, three proximalizing human clusters (Supplementary Fig. 1, clusters 20, 21, and 24) were analyzed, alongside the organoid data, which exclusively focused on cluster 3. These clusters were selected based on their relatively elevated expression levels of HNF4A, each surpassing 15% of all cells in the cluster. Human orthologs of mouse genes were retrieved using Ensembl BioMart, and genes expressed in less than three cells of the human dataset were removed.
mRNA-sequencing and data analyses
Whole organoid samples (3 independent biological replicates per timepoint, per condition for differentiation days 10 and 12, and 2 replicates for differentiation days 14 and 18) were prepared and purified according to the RNeasy Mini Kit (Qiagen, 74104). Purified RNA was sent to Novogene for sequencing. Reads were aligned to the human genome hg38 Ensembl 105 annotation using STAR version 2.7.10a118. A noise-reduction filter was applied to keep genes in which the maximum count in at least one sample was greater than or equal to 10. Partek Flow version 10.0 was then used to normalize these raw feature counts following the transcripts per million (TPM) normalization method. After filtering genes with TPM values greater than or equal to 25 in at least one sample, Z-score averages were calculated, and heatmaps were clustered using pheatmap (Klode, 2018). Separately, gene set enrichment analyses were performed specifically on Day 12 replicate samples by first normalizing noise-reduced feature counts using the DESeq2 median ratio method28, as recommended for gene set enrichment analyses. Partek Flow version 10.0 was then subsequently used for gene set enrichment analyses using default parameters. Data are deposited to the GEO database under accession number GSE264217 and will be made publicly available as of the date of publication.
Immunofluorescent imaging and analyses
Kidney organoids
Whole kidney organoids were fixed for 20 min on ice in 4% PFA (Electron Microscopy Sciences, 15710) in 1xPBS. In 1xPBS, organoids were then carefully cut out of the permeable membrane well and placed into 1xPBS with 1.5% SEA Block (Thermo Fisher Scientific, 37527 × 3) and 0.1% TritonX100 (EMD Millipore, 1.08643) (blocking solution) at 4 °C with gentle movement for one hour. Primary antibodies were resuspended in blocking solution, and organoids were incubated in primary antibody and blocking solution overnight. Samples were rinsed and then washed for at least 3 h through several rounds of 1xPBS with 0.1% Triton X-100. Secondary antibodies were resuspended in blocking solution, and organoids were incubated in secondary antibody and blocking solution overnight. Rinsing and washing steps were repeated the next day before a 25 min counterstain incubation in 1xPBS supplemented with 0.1% Triton X-100 and 1 µg/mL Hoechst 33342 (Thermo Fisher Scientific, H3570) to stain nuclei. Organoids were then washed for 1 h in 1xPBS alone. Individual organoids were mounted for imaging on slides with a glass coverslip in Immu-Mount (Thermo Fisher Scientific, 9990402). Slides were stored at 4 °C in the dark. Primary antibodies used in this study were: WT1 (abcam, ab89901, 1:1000), JAG1 (R&D Systems, AF599, 1:300), HNF1B (Thermo Fisher Scientific, MA5-24605, 1:500), HNF4A (R&D Systems, MAB4605, 1:200), CDH1 (BD Biosciences, 610181, 1:300), ZO-1 (Thermo Fisher Scientific, 33-9100, 1:200), PAX2 (R&D Systems, AF3364, 1:50), SIX1 (Cell Signaling Technology, 12891S, 1:300), HES1 (Cell Signaling Technology, 11988, 1:300), POU3F3 (Novus Biologicals, NBP1-49872, 1:500), HNF4G (Thermo Fisher Scientific, PA5-82189, 1:200), LRP2 (My Bio Source, MBS690201, 1:500), HAVCR1 (R&D Systems, AF1750, 1:200), γH2AX (Cell Signaling Technology, 2577), LAMB1 (Santa Cruz Biotechnology, sc-33709, 1:250), ATP1A1 (Abcam, ab7671, 1:200), SOX9 (Abcam, ab185230, 1:300), PODXL (R&D Systems, AF1658, 1:200), PAX8 (Abcam, ab189249, 1:100), LEF1 (Santa Cruz Biotechnology, sc-374412, 1:200), LTL (Vector Laboratories, B-1325-2, 1:300), SLC12A1 (Sigma Aldrich, HPA018107, 1:200), TFAP2A (Santa Cruz Biotechnology, sc-12726, 1:200), Alexa 647-conjugated LRP2 (R&D Systems, FAB9578R, 1:100), and Alexa 594-conjugated LRP2 (R&D Systems, FAB9578T, 1:100). Secondary antibodies conjugated with AlexaFluor 488, 555, 594, and 647 were all diluted to 1:500 in blocking solution.
Human kidney sections
Human kidneys were carefully dissected out from donated tissues and placed in 1xPBS. Kidneys were then placed into 4% PFA to fix overnight (18 h). Kidneys were washed twice in 1xPBS (1 h per wash), and samples were placed in 30% sucrose in 1xPBS rocking at 4 °C overnight (18 h). The next day, kidneys were swirled in Optimal Cutting Temperature (OCT from Sakura Finetech USA Inc, 4583) compound 3 times before embedding in an OCT block. The OCT blocks with tissues were frozen on a dry ice/ethanol slurry. Blocks were stored at -80 °C, and ~12 µm sections were obtained using a cryostat. Slides were stored at -80 °C before being processed for immunofluorescent imaging.
Image acquisition and analyses
Image acquisition of whole-mounted organoids and cryo-sectioned human kidneys was performed using (1) Leica SP8-X confocal fluorescence imaging system (Leica Microsystems, Germany), in 1024 × 1024 pixels using 25X water, 40X oil, and 63X oil objectives, and (2) Leica Stellaris confocal microscope at 20X oil and 93X glycerol objectives. A Leica Thunder microscope was additionally used at 10X dry objective to image entire whole-mounted kidney organoids. Live organoids were captured using (1) Zeiss Axio Zoom at 16X, 32X, and 112X objectives and (2) Olympus APX100 at 4X objective. Image masking was performed and quantified with Imaris 9.7 software (Oxford Instruments) and with Fiji119. Fiji was used for individual cell and nephron segment counting, and for quantifying relative protein signal and organoid nephron size.
In vitro albumin and dextran uptake assays
For the albumin uptake assay, differentiation day 21 control and proximal-biased organoids were incubated at 37 °C/5% CO2 overnight (18 h) in TeSR-E6 supplemented with 50 µg/mL TRITC-albumin (Sigma Aldrich, A2289) and Alexa 647-conjugated LRP2 antibody (R&D Systems, FAB9578R, 1:100). After the incubation, organoids were washed at least 3 times with 1xPBS and live imaged immediately after. For the dextran uptake assay, organoids were treated and processed in the same way, except that TeSR-E6 was supplemented with 20 µg/mL Alexa 647-conjugated Dextran (Thermo Fisher Scientific, D22914) and Alexa 594-conjugated LRP2 (R&D Systems, FAB9578T, 1:100).
In vitro cisplatin injury assay
For the kidney organoid cisplatin injury assay, we adapted an approach undertaken by two recent studies13,14. Differentiation day 18 control and proximal-biased organoids were incubated overnight (18 h) at 37 °C/5% CO2 in TeSR-E6 supplemented with 5 µM cisplatin (Millipore Sigma, 232120). Organoids recovered between days 19 and 20, before another overnight culture (18 h) between days 20 and 21 with 5 µM cisplatin. Uninjured organoids were cultured with DMSO vehicle. Organoids were then processed for immunofluorescent imaging and analyses.
Statistics and reproducibility
All whole-mount immunofluorescent experiments, including those shown as representative images (e.g., Figs. 1a–f; 2j–m; 3f, h–k; 4d, e, h–j and 6c, g, i, j, m), were independently replicated a minimum of three times with equivalent results. Proximal-biasing was replicated in kidney organoids differentiated from two different human iPSC lines and one human iNPC line, and is shown throughout this work. Biological replicates were defined as organoids generated from independent differentiation batches. Representative organoids selected for imaging were chosen from randomized sets of well-formed structures with comparable morphology. Further details of statistical tests, where applicable, are provided in the figure legends and Methods.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The bulk RNA-sequencing data generated in this study have been deposited in the GEO database under accession code GSE264217. The single-cell RNA-sequencing data generated in this study have been deposited in the GEO database under accession code GSE264678. The single-nucleus RNA-sequencing data generated in this study have been deposited in the GEO database under accession code GSE290931. This paper uses referenced sources of code, and no novel code was created for the analyses in this study. The following datasets was accessed for this study: GSE139280 (single-cell RNA-sequencing of week 14 human embryonic kidney cortex cells); GSE124472 (single-cell RNA-sequencing of week 17 human embryonic kidney outer and inner cortical cells); GSE114802 (single-cell RNA-sequencing of day 25 kidney organoids); GSE136314 (single-cell RNA-sequencing of days 7, 15, and 29 kidney organoids); GSE124472 (single-cell RNA-sequencing of days 16 and 28 kidney organoids); GSE118184 (single-cell RNA-sequencing of day 26 kidney organoids); GSE184928 (single-cell RNA-sequencing of PT-enhanced kidney organoids); GSE144772 (bulk RNA-sequencing data from control and Hnf4a-mutant P0 mouse kidneys); GSE129798 (single-cell RNA-sequencing of the adult mouse kidney).
References
Zhuo, J. L. & Li, X. C. Proximal nephron. Compr. Physiol. 3, 1079–1123 (2013).
Chevalier, R. L. The proximal tubule is the primary target of injury and progression of kidney disease: role of the glomerulotubular junction. Am. J. Physiol. Ren. Physiol. 311, F145–F161 (2016).
Nakhoul, N. & Batuman, V. Role of proximal tubules in the pathogenesis of kidney disease. Contrib. Nephrol. 169, 37–50 (2011).
Takasato, M., Er, P. X., Chiu, H. S. & Little, M. H. Generation of kidney organoids from human pluripotent stem cells. Nat. Protoc. 11, 1681–1692 (2016).
Morizane, R. & Bonventre, J. V. Generation of nephron progenitor cells and kidney organoids from human pluripotent stem cells. Nat. Protoc. 12, 195–207 (2017).
Combes, A. N., Zappia, L., Er, P. X., Oshlack, A. & Little, M. H. Single-cell analysis reveals congruence between kidney organoids and human fetal kidney. Genome Med 11, 3 (2019).
Subramanian, A. et al. Single cell census of human kidney organoids shows reproducibility and diminished off-target cells after transplantation. Nat. Commun. 10, 5462 (2019).
Tran, T. et al. In vivo developmental trajectories of human podocyte inform in vitro differentiation of pluripotent stem cell-derived podocytes. Dev. Cell 50, 102–116.e6 (2019).
Wu, H. et al. Comparative analysis and refinement of human PSC-derived kidney organoid differentiation with single-cell transcriptomics. Cell Stem Cell 23, 869–881.e8 (2018).
Vanslambrouck, J. M. et al. Enhanced metanephric specification to functional proximal tubule enables toxicity screening and infectious disease modelling in kidney organoids. Nat. Commun. 13, 5943 (2022).
Yoshimura, Y. et al. A single-cell multiomic analysis of kidney organoid differentiation. Proc. Natl. Acad. Sci. USA120, e2219699120 (2023).
Morizane, R. et al. Nephron organoids derived from human pluripotent stem cells model kidney development and injury. Nat. Biotechnol. 33, 1193–1200 (2015).
Gupta, N. et al. Modeling injury and repair in kidney organoids reveals that homologous recombination governs tubular intrinsic repair. Sci. Transl. Med. 14, eabj4772 (2022).
Digby, J. L. M., Vanichapol, T., Przepiorski, A., Davidson, A. J. & Sander, V. Evaluation of cisplatin-induced injury in human kidney organoids. Am. J. Physiol. Ren. Physiol. 318, F971–F978 (2020).
Yoshimura, Y., Muto, Y., Omachi, K., Miner, J. H. & Humphreys, B. D. Elucidating the proximal tubule HNF4A gene regulatory network in human kidney organoids. J. Am. Soc. Nephrol. JASN https://doi.org/10.1681/ASN.0000000000000197 (2023).
Marable, S. S., Chung, E. & Park, J.-S. Hnf4a is required for the development of Cdh6-expressing progenitors into proximal tubules in the mouse kidney. J. Am. Soc. Nephrol. JASN 31, 2543–2558 (2020).
Marable, S. S., Chung, E., Adam, M., Potter, S. S. & Park, J.-S. Hnf4a deletion in the mouse kidney phenocopies Fanconi renotubular syndrome. JCI Insight 3, e97497 (2018).
Lindström, N. O. et al. Spatial transcriptional mapping of the human nephrogenic program. Dev. Cell 56, 2381–2398.e6 (2021).
Schnell, J., Achieng, M. & Lindström, N. O. Principles of human and mouse nephron development. Nat. Rev. Nephrol. https://doi.org/10.1038/s41581-022-00598-5 (2022).
Georgas, K. et al. Analysis of early nephron patterning reveals a role for distal RV proliferation in fusion to the ureteric tip via a cap mesenchyme-derived connecting segment. Dev. Biol. 332, 273–286 (2009).
Lindström, N. O. et al. Progressive recruitment of mesenchymal progenitors reveals a time-dependent process of cell fate acquisition in mouse and human nephrogenesis. Dev. Cell 45, 651–660.e4 (2018).
Chung, E., Deacon, P. & Park, J.-S. Notch is required for the formation of all nephron segments and primes nephron progenitors for differentiation. Dev. Camb. Engl. 144, 4530–4539 (2017).
Heliot, C. et al. HNF1B controls proximal-intermediate nephron segment identity in vertebrates by regulating Notch signalling components and Irx1/2. Dev. Camb. Engl. 140, 873–885 (2013).
Massa, F. et al. Hepatocyte nuclear factor 1β controls nephron tubular development. Dev. Camb. Engl. 140, 886–896 (2013).
Lindström, N. O. et al. Integrated β-catenin, BMP, PTEN, and Notch signalling patterns the nephron. eLife 3, e04000 (2015).
Takasato, M. & Little, M. H. A strategy for generating kidney organoids: Recapitulating the development in human pluripotent stem cells. Dev. Biol. 420, 210–220 (2016).
Ransick, A. et al. Single-cell profiling reveals sex, lineage, and regional diversity in the mouse kidney. Dev. Cell 51, 399–413.e7 (2019).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Lindström, N. O., Carragher, N. O. & Hohenstein, P. The PI3K pathway balances self-renewal and differentiation of nephron progenitor cells through β-catenin signaling. Stem Cell Rep. 4, 551–560 (2015).
Vlahos, C. J., Matter, W. F., Hui, K. Y. & Brown, R. F. A specific inhibitor of phosphatidylinositol 3-kinase, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one (LY294002). J. Biol. Chem. 269, 5241–5248 (1994).
Wang, J. et al. PI3K-AKT pathway mediates growth and survival signals during development of fetal mouse lung. Tissue Cell 37, 25–35 (2005).
Arab, K. et al. Long noncoding RNA TARID directs demethylation and activation of the tumor suppressor TCF21 via GADD45A. Mol. Cell 55, 604–614 (2014).
Dovey, H. F. et al. Functional gamma-secretase inhibitors reduce beta-amyloid peptide levels in brain. J. Neurochem. 76, 173–181 (2001).
Cheng, H.-T. et al. Gamma-secretase activity is dispensable for mesenchyme-to-epithelium transition but required for podocyte and proximal tubule formation in developing mouse kidney. Dev. Camb. Engl. 130, 5031–5042 (2003).
Vanslambrouck, J. M. et al. A toolbox to characterize human induced pluripotent stem cell-derived kidney cell types and organoids. J. Am. Soc. Nephrol. JASN 30, 1811–1823 (2019).
Przepiorski, A. et al. A simple bioreactor-based method to generate kidney organoids from pluripotent stem cells. Stem Cell Rep. 11, 470–484 (2018).
Huang, B. et al. Long-term expandable mouse and human-induced nephron progenitor cells enable kidney organoid maturation and modeling of plasticity and disease. Cell Stem Cell 31, 921–939.e17 (2024).
Lindström, N. O. et al. Conserved and divergent features of mesenchymal progenitor cell types within the cortical nephrogenic niche of the human and mouse kidney. J. Am. Soc. Nephrol. JASN 29, 806–824 (2018).
Roth, M., Obaidat, A. & Hagenbuch, B. OATPs, OATs and OCTs: the organic anion and cation transporters of the SLCO and SLC22A gene superfamilies. Br. J. Pharmacol. 165, 1260–1287 (2012).
Ren, Q. et al. Distinct functions of megalin and cubilin receptors in recovery of normal and nephrotic levels of filtered albumin. Am. J. Physiol. Ren. Physiol. 318, F1284–F1294 (2020).
Bajaj, P. et al. Human pluripotent stem cell-derived kidney model for nephrotoxicity studies. Drug Metab. Dispos. Biol. Fate Chem. 46, 1703–1711 (2018).
Schuh, C. D. et al. Combined structural and functional imaging of the kidney reveals major axial differences in proximal tubule endocytosis. J. Am. Soc. Nephrol. JASN 29, 2696–2712 (2018).
Molitoris, B. A., Sandoval, R. M., Yadav, S. P. S. & Wagner, M. C. Albumin uptake and processing by the proximal tubule: physiological, pathological, and therapeutic implications. Physiol. Rev. 102, 1625–1667 (2022).
Freedman, B. S. Physiology assays in human kidney organoids. Am. J. Physiol. Ren. Physiol. 322, F625–F638 (2022).
Rizki-Safitri, A. et al. Live functional assays reveal longitudinal maturation of transepithelial transport in kidney organoids. Front. Cell Dev. Biol. 10, 978888 (2022).
Pabla, N., Murphy, R. F., Liu, K. & Dong, Z. The copper transporter Ctr1 contributes to cisplatin uptake by renal tubular cells during cisplatin nephrotoxicity. Am. J. Physiol. Ren. Physiol. 296, F505–511 (2009).
Miller, R. P., Tadagavadi, R. K., Ramesh, G. & Reeves, W. B. Mechanisms of cisplatin nephrotoxicity. Toxins 2, 2490–2518 (2010).
Motwani, S. S., Kaur, S. S. & Kitchlu, A. Cisplatin nephrotoxicity: novel insights into mechanisms and preventative strategies. Semin. Nephrol. 42, 151341 (2022).
Tang, C., Livingston, M. J., Safirstein, R. & Dong, Z. Cisplatin nephrotoxicity: new insights and therapeutic implications. Nat. Rev. Nephrol. 19, 53–72 (2023).
Yamashita, N. et al. Cumulative DNA damage by repeated low-dose cisplatin injection promotes the transition of acute to chronic kidney injury in mice. Sci. Rep. 11, 20920 (2021).
Huang, G., Zhang, Q., Xu, C., Chen, L. & Zhang, H. Mechanism of kidney injury induced by cisplatin. Toxicol. Res. 11, 385–390 (2022).
Singh, G. A possible cellular mechanism of cisplatin-induced nephrotoxicity. Toxicology 58, 71–80 (1989).
Sánchez-González, P. D., López-Hernández, F. J., López-Novoa, J. M. & Morales, A. I. An integrative view of the pathophysiological events leading to cisplatin nephrotoxicity. Crit. Rev. Toxicol. 41, 803–821 (2011).
Choie, D. D., Longnecker, D. S. & del Campo, A. A. Acute and chronic cisplatin nephropathy in rats. Lab. Investig. J. Tech. Methods Pathol. 44, 397–402 (1981).
Cullen, K. J., Yang, Z., Schumaker, L. & Guo, Z. Mitochondria as a critical target of the chemotheraputic agent cisplatin in head and neck cancer. J. Bioenerg. Biomembr. 39, 43–50 (2007).
Mandic, A., Hansson, J., Linder, S. & Shoshan, M. C. Cisplatin induces endoplasmic reticulum stress and nucleus-independent apoptotic signaling. J. Biol. Chem. 278, 9100–9106 (2003).
George, B., You, D., Joy, M. S. & Aleksunes, L. M. Xenobiotic transporters and kidney injury. Adv. Drug Deliv. Rev. 116, 73–91 (2017).
Nakamura, T., Yonezawa, A., Hashimoto, S., Katsura, T. & Inui, K.-I. Disruption of multidrug and toxin extrusion MATE1 potentiates cisplatin-induced nephrotoxicity. Biochem. Pharmacol. 80, 1762–1767 (2010).
Iwata, K. et al. Effects of genetic variants in SLC22A2 organic cation transporter 2 and SLC47A1 multidrug and toxin extrusion 1 transporter on cisplatin-induced adverse events. Clin. Exp. Nephrol. 16, 843–851 (2012).
Cui, Y. et al. Drug resistance and ATP-dependent conjugate transport mediated by the apical multidrug resistance protein, MRP2, permanently expressed in human and canine cells. Mol. Pharmacol. 55, 929–937 (1999).
Kawabe, T. et al. Enhanced transport of anticancer agents and leukotriene C4 by the human canalicular multispecific organic anion transporter (cMOAT/MRP2). FEBS Lett. 456, 327–331 (1999).
Ciarimboli, G. et al. Cisplatin nephrotoxicity is critically mediated via the human organic cation transporter 2. Am. J. Pathol. 167, 1477–1484 (2005).
Yokoo, S. et al. Differential contribution of organic cation transporters, OCT2 and MATE1, in platinum agent-induced nephrotoxicity. Biochem. Pharmacol. 74, 477–487 (2007).
Filipski, K. K., Mathijssen, R. H., Mikkelsen, T. S., Schinkel, A. H. & Sparreboom, A. Contribution of organic cation transporter 2 (OCT2) to cisplatin-induced nephrotoxicity. Clin. Pharmacol. Ther. 86, 396–402 (2009).
Gerhardt, L. M. S., Liu, J., Koppitch, K., Cippà, P. E. & McMahon, A. P. Single-nuclear transcriptomics reveals diversity of proximal tubule cell states in a dynamic response to acute kidney injury. Proc. Natl. Acad. Sci. USA. 118, e2026684118 (2021).
Gerhardt, L. M. S. et al. Lineage tracing and single-nucleus multiomics reveal novel features of adaptive and maladaptive repair after acute kidney injury. J. Am. Soc. Nephrol. JASN https://doi.org/10.1681/ASN.0000000000000057 (2023).
Ledru, N. et al. Predicting proximal tubule failed repair drivers through regularized regression analysis of single cell multiomic sequencing. Nat. Commun. 15, 1291 (2024).
Aggarwal, S. et al. SOX9 switch links regeneration to fibrosis at the single-cell level in mammalian kidneys. Science 383, eadd6371 (2024).
Kha, M. et al. Injured proximal tubular epithelial cells lose hepatocyte nuclear factor 4α expression crucial for brush border formation and transport. Am. J. Pathol. S0002-9440, 00047–1 (2025).
Kumar, S. et al. Sox9 activation highlights a cellular pathway of renal repair in the acutely injured mammalian kidney. Cell Rep. 12, 1325–1338 (2015).
Kang, H. M. et al. Sox9-positive progenitor cells play a key role in renal tubule epithelial regeneration in mice. Cell Rep. 14, 861–871 (2016).
Zhang, K. et al. In vivo two-photon microscopy reveals the contribution of Sox9+ cell to kidney regeneration in a mouse model with extracellular vesicle treatment. J. Biol. Chem. 295, 12203–12213 (2020).
Duvall, K. et al. Revisiting the role of notch in nephron segmentation confirms a role for proximal fate selection during mouse and human nephrogenesis. Dev. Camb. Engl. https://doi.org/10.1242/dev.200446 (2022).
Cheng, H.-T. et al. Notch2, but not Notch1, is required for proximal fate acquisition in the mammalian nephron. Dev. Camb. Engl. 134, 801–811 (2007).
Chen, L. & Al-Awqati, Q. Segmental expression of Notch and Hairy genes in nephrogenesis. Am. J. Physiol. Ren. Physiol. 288, F939–F952 (2005).
Liu, Z. et al. The extracellular domain of Notch2 increases its cell-surface abundance and ligand responsiveness during kidney development. Dev. Cell 25, 585–598 (2013).
Bonegio, R. G. B., Beck, L. H., Kahlon, R. K., Lu, W. & Salant, D. J. The fate of Notch-deficient nephrogenic progenitor cells during metanephric kidney development. Kidney Int. 79, 1099–1112 (2011).
Bohn, S. et al. Distinct molecular and morphogenetic properties of mutations in the human HNF1beta gene that lead to defective kidney development. J. Am. Soc. Nephrol. JASN 14, 2033–2041 (2003).
Nakayama, M. et al. HNF1B alterations associated with congenital anomalies of the kidney and urinary tract. Pediatr. Nephrol. Berl. Ger. 25, 1073–1079 (2010).
Naylor, R. W., Przepiorski, A., Ren, Q., Yu, J. & Davidson, A. J. HNF1β is essential for nephron segmentation during nephrogenesis. J. Am. Soc. Nephrol. JASN 24, 77–87 (2013).
Clissold, R. L., Hamilton, A. J., Hattersley, A. T., Ellard, S. & Bingham, C. HNF1B-associated renal and extra-renal disease-an expanding clinical spectrum. Nat. Rev. Nephrol. 11, 102–112 (2015).
Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. Publ. Protein Soc. 28, 1947–1951 (2019).
Kanehisa, M. & Goto, S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30 (2000).
Kanehisa, M., Furumichi, M., Sato, Y., Kawashima, M. & Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 51, D587–D592 (2023).
Barak, H. et al. FGF9 and FGF20 maintain the stemness of nephron progenitors in mice and man. Dev. Cell 22, 1191–1207 (2012).
Chen, L. et al. A reinforcing HNF4-SMAD4 feed-forward module stabilizes enterocyte identity. Nat. Genet. 51, 777–785 (2019).
Chen, L. et al. HNF4 factors control chromatin accessibility and are redundantly required for maturation of the fetal intestine. Dev. Camb. Engl. 146, dev179432 (2019).
Guo, Q. et al. A β-catenin-driven switch in tcf/lef transcription factor binding to DNA target sites promotes commitment of mammalian nephron progenitor cells. eLife 10, 1–25 (2021).
Park, J.-S. et al. Six2 and Wnt regulate self-renewal and commitment of nephron progenitors through shared gene regulatory networks. Dev. Cell 23, 637–651 (2012).
Lindström, N. O. et al. Conserved and divergent molecular and anatomic features of human and mouse nephron patterning. J. Am. Soc. Nephrol. JASN 29, 825–840 (2018).
Fausto, C. C. et al. Defining and controlling axial nephron patterning in human kidney organoids with synthetic Wnt-secreting organizers. Preprint at https://doi.org/10.1101/2024.11.30.626171 (2024).
Blaine, J., Chonchol, M. & Levi, M. Renal control of calcium, phosphate, and magnesium homeostasis. Clin. J. Am. Soc. Nephrol. CJASN 10, 1257–1272 (2015).
Klootwijk, E. D. et al. Renal Fanconi syndrome: taking a proximal look at the nephron. Nephrol. Dial. Transplant. 30, 1456–1460 (2015).
Baum, M. & Quigley, R. Proximal tubule water transport-lessons from aquaporin knockout mice. Am. J. Physiol. Ren. Physiol. 289, F1193–F1194 (2005).
Takasato, M. et al. Kidney organoids from human iPS cells contain multiple lineages and model human nephrogenesis. Nature 526, 564–568 (2015).
Wilmes, A. et al. Mechanism of cisplatin proximal tubule toxicity revealed by integrating transcriptomics, proteomics, metabolomics and biokinetics. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 30, 117–127 (2015).
Nieskens, T. T. G. et al. Expression of organic anion transporter 1 or 3 in human kidney proximal tubule cells reduces cisplatin sensitivity. Drug Metab. Dispos. Biol. Fate Chem. 46, 592–599 (2018).
Sahara, Y., Fukui, C., Kuniyoshi, Y. & Takasato, M. Proximal tubule cell maturation rate and function are controlled by PPARα signaling in kidney organoids. Commun. Biol. 7, 1532 (2024).
Banan Sadeghian, R. et al. Cells sorted off hiPSC-derived kidney organoids coupled with immortalized cells reliably model the proximal tubule. Commun. Biol. 6, 483 (2023).
Yousef Yengej, F. A. et al. Tubuloid culture enables long-term expansion of functional human kidney tubule epithelium from iPSC-derived organoids. Proc. Natl. Acad. Sci. USA120, e2216836120 (2023).
Aceves, J. O. et al. 3D proximal tubule-on-chip model derived from kidney organoids with improved drug uptake. Sci. Rep. 12, 14997 (2022).
Carracedo, M. et al. 3D vascularised proximal tubules-on-a-multiplexed chip model for enhanced cell phenotypes. Lab. Chip https://doi.org/10.1039/d2lc00723a (2023).
Hor, P., Ichida, J. K., Borok, Z. & Ryan, A. L. Protocol for differentiation of Human iPSCs into pulmonary neuroendocrine cells. STAR Protoc. 1, 100068 (2020).
Howden, S. E. et al. A Cas9 variant for efficient generation of indel-free knockin or gene-corrected human pluripotent stem cells. Stem Cell Rep. 7, 508–517 (2016).
Howden, S. E. & Little, M. H. Generating kidney organoids from human pluripotent stem cells using defined conditions. Methods Mol. Biol. Clifton NJ 2155, 183–192 (2020).
Ellis, B. et al. lowCore: flowCore: Basic structures for flow cytometry data. R package version 2.20.0 https://doi.org/10.18129/B9.bioc.flowCore (2025).
Kim, S. et al. Comparative single-cell analyses identify shared and divergent features of human and mouse kidney development. Dev. Cell S1534-5807, 00450–00457 (2024).
Hao, Y. et al. Dictionary learning for integrative, multimodal and scalable single-cell analysis. Nat. Biotechnol. 42, 293–304 (2024).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587.e29 (2021).
Stuart, T. et al. Comprehensive integration of single-cell data. Cell https://doi.org/10.1016/j.cell.2019.05.031 (2019).
Butler, A., Hoffman, P., Smibert, P., Papalexi, E. & Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 36, 411–420 (2018).
Satija, R., Farrell, J. A., Gennert, D., Schier, A. F. & Regev, A. Spatial reconstruction of single-cell gene expression data. Nat. Biotechnol. 33, 495–502 (2015).
Samuel, M., Maëlle, S. & Paul, H. Samuel-marsh/scCustomize: version 2.1.2. https://doi.org/10.5281/ZENODO.5706430 (2024).
Trapnell, C. et al. The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat. Biotechnol. 32, 381–386 (2014).
Qiu, X. et al. Reversed graph embedding resolves complex single-cell trajectories. Nat. Methods 14, 979–982 (2017).
Qiu, X. et al. Single-cell mRNA quantification and differential analysis with Census. Nat. Methods 14, 309–315 (2017).
Cao, J. et al. The single-cell transcriptional landscape of mammalian organogenesis. Nature 566, 496–502 (2019).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinforma. Oxf. Engl. 29, 15–21 (2013).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Acknowledgements
We thank all past and present members of the Lindström Laboratory and extend our gratitude to Dr. Andrew McMahon for input. We further thank Dr. Alicia McDonough for insights to human physiology, Dr. Seth Ruffins and the Optical Imaging Facility for assistance with microscopy, and Dr. Bernadette Masinsin and the Flow Cytometry Facility for help with cell isolation. Lastly, we thank Dr. Amy L. Ryan (University of Iowa) for providing iPSCs used in this study. J.S. and L.T. were supported by the T32 Training Grant in Development, Stem Cells, and Regenerative Medicine from NICHD (T32HD060549). C.C.F. was supported by the CIRM EDUC4-12756 Training Grant. Sequencing and organoid work was supported by NIH/NIDDK (Re)Building a Kidney Opportunity Pool funding (N.O.L.), NIH/NIDDK 1R01DK136802-01 (N.O.L.), additional funds from ASN/HHS KidneyX (N.O.L.), renalART (N.O.L.), and the University of Southern California (N.O.L.).
Author information
Authors and Affiliations
Contributions
J.S. and N.O.L. planned experiments and analyzed data. J.S. assembled the figures. J.S., Z.M., M.A., C.C.F., K.K., L.T., V.W., and F.D.K. performed the experiments, data collection, and/or data analysis. L.T., B.H., M.S., P.M., and Z.L. assisted with iNPC organoid culture. Z.M. and J.K. planned and conducted data analyses. M.E.T. and B.G. provided human tissue. J.S. and N.O.L. wrote the manuscript and incorporated input from all authors.
Corresponding author
Ethics declarations
Competing interests
J.S. and N.O.L. have applied for intellectual property protection on work presented here (patent pending). The remaining authors declare no competing interests for this study.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Schnell, J., Miao, Z., Achieng, M. et al. Controlling nephron precursor differentiation to generate proximal-biased kidney organoids with emerging maturity. Nat Commun 16, 8136 (2025). https://doi.org/10.1038/s41467-025-63107-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-63107-9
This article is cited by
-
Developmentally inspired synthetic kidney engineering
Nature Biotechnology (2026)
-
Axial nephron fate switching demonstrates a plastic system tunable on demand
Nature Communications (2025)
