
Abstract
The Hanging Coffin represents a distinctive mortuary tradition practiced across southern China, Southeast Asia (e.g., the Log Coffin), and the Pacific approximately 3,000 years. Historical records attribute this funerary practice to the Bo people, a group that largely disappeared from documented history by the end of the Ming Dynasty (1,368–1,644 AD). Here we report eleven ancient genomes from four Hanging Coffin sites in China, alongside thirty whole genomes from the extant Bo people in Southwest China. We also sequence four ancient genomes from Log Coffin sites in northwestern Thailand. Our findings indicate that present-day Bo people derive a substantial proportion of their ancestry from practitioners of the Hanging Coffin mortuary tradition. Both ancient and modern groups exhibit elevated genetic affinity with coastal Neolithic populations from southern East Asia, who are ancestral to Tai-Kadai and Austronesian speakers. Unexpectedly, we also find evidence of long-range interactions and cultural inclusivity between Northeast Asian and Yellow River farmers and Hanging Coffin communities over 1,200 years ago. Finally, shared genetic components between Hanging (Log) Coffin populations in China and Thailand point to a common origin and a broader genetic and cultural network underlying this distinctive mortuary tradition across southern China and Southeast Asia.
Similar content being viewed by others

Introduction
Anatomically modern humans first occupied in the southern regions of East Asia at least 65,000 to 50,000 years ago, giving rise to deeply divergent hunter-gatherer lineages such as those associated with the Hòabìnhian culture complex1 and extant groups including the Onge and Jarawa2. Around 4500 to 4000 years ago, Neolithic farming populations, bearing East Asian ancestry and likely speakers of proto-Austroasiatic and proto-Austronesian languages, migrated southward from southern China, introducing rice and millet agriculture into Mainland and Island Southeast Asia3,4. These major demographic and cultural transitions coincided with the emergence and transformation of regional mortuary practices, including the development of the Hanging Coffin custom.
Human mortuary practices have undergone profound transformations throughout human history, reflecting a wide range of cultural, religious, and ecological contexts. From prehistory to the present, burial customs serve as critical windows into cultural identity, spiritual beliefs, and the relationship between humans and their environment. The Bo people’s Hanging Coffin tradition, adorning the cliffs along the Yangtze River, is a striking example. The Hanging Coffin burial is a funerary practice in which wooden coffins containing the deceased are placed in elevated locations, such as cliffs, caves, or rock crevices, typically along riverbanks, coastlines, or mountainous regions5. In the past 30 years, archaeological surveys and cultural relic conservation efforts in China have documented hundreds of Hanging Coffins across more than 20 counties in Yunnan, Sichuan, Chongqing, Hunan, Jiangxi, Zhejiang, Guangxi, Fujian, and Taiwan5. According to the archaeological record, the earliest securely dated evidence of this tradition comes from the Wuyi Mountains in Fujian Province, located near the southeastern coast below the lower Yangtze River, with radiocarbon dates ranging from 3445 ± 150 to 3370 ± 80BP. From this region, the practice spread to other parts of China, eventually moving southward and westward across various cultural zones5.
While archaeological data provide evidence of the practice itself, the cultural meanings and identities associated with the Hanging Coffin custom are also reflected in historical texts and oral traditions. The Bo people, a historically obscure ethnic group inhabiting the rugged regions of Southwest China5, are frequently mentioned in this context. Their funerary practices, especially Hanging Coffins, have long captured the imagination of historians and local communities. Where historical clarity is lacking, mythology often fills the void. This is exemplified in the case of the Bo burial customs. A central question remains: why were coffins suspended at such great heights, and what significance this practice holds? Jing Li (李靖), in his Brief Chronicles of Yunnan during the Yuan Dynasty (1279–1368 AD), offers one of the earliest interpretations: “Coffins set high are considered auspicious. The higher they are, the more propitious they are for the dead. Furthermore, those whose coffins fell to the ground sooner were considered more fortunate” (seen at www.china.org.cn). The Bo people is an ancient ethnic group in China, with limited historical documentation concerning their origins and language. It is known that they thrived up to the Ming Dynasty (1368–1644 AD), but very little is recorded about them in official Chinese documents. Over time, the Bo were referenced in regional folklore with names such as “Subjugators of the Sky” and “Sons of the Cliffs, and even described as being capable of flight6, reflecting how cultural narratives have evolved around them.
Although references to the Bo are limited in historical records, a small Bo community continues to reside in Qiubei County today (丘北县, 23 °45’–24 °28’N, 103 °34’–104 °45’E) in Wenshan Zhuang and Miao Autonomous Prefecture of southeastern Yunnan Province. As of 2003, the Bo people comprised 1087 households and 5084 individuals distributed across 42 natural villages in six townships. Situated about 700 km south of the Hanging Coffin sites in Zhaotong City (a border region between Yunnan and Sichuan Provinces), this community has managed to preserve remarkable cultural continuity due to geographic isolation. In 1956, local officials classified the Bo people as part of the Bai Yi branch of the Yi ethnic group. They speak a Loloish language, which belongs to the Tibeto-Burman branch of the Sino-Tibetan family, though their dialect, clothing, and traditional customs differ significantly from those of the Yi group. Historically, the Bo, sometimes referred to as the “Haiba” people by neighbouring groups, have lived in remote karst landscapes characterised by rugged topography and limited infrastructure, factors that have helped maintain their unique cultural practices. Particularly noteworthy is their distinctive funerary tradition known as “cave burial”, a ritual diverging from conventional coffin interment. This tradition involves the ceremonial placement of the spiritual essence of the deceased, rather than their physical remains, within ancestral caves, marking an important aspect of their intangible cultural heritage7 (Supplementary Note 2).
In recent years, several studies have investigated on ancient human DNA from Southwest China8,9,10,11,12,13,14,15, with notable findings regarding cliff tombs12 and cave burials11. Ancient human genome-wide data from archaeological sites have revealed important genetic insights into late Neolithic to Bronze Age farmers10 and late Pleistocene population9. Specifically, ancient genomic data from Yunnan and Sichuan suggest the demographic diffusion of Neolithic millet farmers from the Yellow River region into Southwest China10; Similarly, the genetic influence of Hòabìnhian hunter-gatherers has been traced back to approximately 6000–9000 years ago in Guangxi, and historical populations in this region were closely associated with Tai-Kadai and Hmong-Mien speakers15. Previously, analyses of uniparental genetic markers suggested that the Bo people in Qiubei County were genetically closer to southern populations than northern populations in China16. Moreover, our recent mitochondrial DNA analysis of human remains from Hanging Coffins in southern China and northwestern Thailand (Log Coffin) revealed genetic connections between these populations across these regions, we propose that the Hanging Coffin custom spread in southern China through substantial human migration. In contrast, the major dispersal pattern in Southeast Asia has undergone cultural assimilation over the past 2000 years8. However, the demographic and genetic history of the Hanging Coffin tradition and its relationship to the present-day Bo population remains largely unknown. Whole-genome sequencing (WGS) of both ancient and present-day populations offers a deeper scientific understanding of their genetic connections.
Overall, the Hanging Coffin burial custom was one of the most widespread practices in southern China in ancient times. However, the practice gradually declined, coinciding with the broader reduction in references to the Bo people in historical sources. Historical records suggest that during the Ming Dynasty (1368–1644 AD), the Bo people faced persecution, leading many survivors to flee to neighbouring regions, change their identities, and merge with surrounding populations (Biography of Liu Xian” in the History of Ming《明史·刘显传》; Chronicle of Military Campaigns in the Wanli Period《万历武功录》). This raises the crucial question: to what extent are the present-day Bo people genetically and culturally linked to the ancient practitioners of the Hanging Coffin tradition? What are the genomic profiles of both the ancient and contemporary populations? To explore these questions, we sequenced the genomes of eleven ancient Hanging Coffin individuals from four archaeological sites in Yunnan and Guangxi, dating from 660 to 2490 years ago. In addition, we conducted WGS of thirty genomes of the remnant Bo population in She De village (舍得村: 24 °14'26.43“N, 103 °52'29.29“E), Qiubei County, Yunnan Province, China (Fig. 1a). To further strengthen our comparison, we successfully sequenced four ancient genomes from three Log Coffin sites in the Pang Mapha region of northwestern Thailand, including one sample from the Bor Krai site not previously analysed in a recent study17.

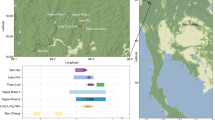
a Geographic location of the seven ancient Hanging (Log) Coffin archaeological sites and the contemporary Bo village (Qiubei, QBBR) analysed in this study. b Principal component analysis (PCA) of genome-wide SNP data, illustrating genetic affinities among the ancient individuals examined here (red) and a broader panel of published ancient and present-day East Asian populations. The Bo individuals from Qiubei (QBBR) are shown in blue. Ancient individuals are projected onto the genetic variation of present-day East Asians, as defined by the first and second principal components (PC1 and PC2). Population abbreviations: N Neolithic; EN Early Neolithic; MN Middle Neolithic; LN Late Neolithic; BA Bronze Age; LBIA Late Bronze to Iron Age; IA Iron Age. See also Supplementary Fig. 3. Source data are provided in this paper.
Results
Overview of sequenced genomic data in this study
We screened 31 human remains from twelve Hanging (Log) Coffin sites across Yunnan, Guangxi, and northwestern Thailand8, as well as two additional samples (WXLM1 from Long Ma and BK_737 from Bor Krai). After removing individuals with fewer than 30,000 single-nucleotide polymorphisms (SNPs) in the 1240 K SNP panel, we recovered genome-wide data from 15 genomes across seven archaeological sites, all of which exhibited characteristic patterns of post-mortem damage (Supplementary Fig. 1). Based on the estimated contamination level and terminal damage patterns, four genomes were analysed using only deaminated DNA fragments. The final dataset comprised between 40,280 and 1,202,746 SNPs per individual in the 1240 K SNP panel (Supplementary Data 1). Among these, samples LP_37_397 and LP_47_398 from the Lahu Pot site in northwestern Thailand showed a first-degree kinship relationship (Supplementary Fig. 2a and Supplementary Data 2). Consequently, we excluded LP_37_397 from further analyses, retaining 14 ancient genomes for downstream population genetic investigation. In addition, for the 32 Bo samples from Qiubei County previously reported16, we performed high coverage WGS on 30 samples of them, achieving an average sequencing depth of 37-fold (Supplementary Data 3). Although the Bo volunteers reported no close kinship, our analysis revealed up to third-degree kinship in 9 samples, which were excluded from further analysis. We retained the remaining 21 samples for downstream genomic investigation (Supplementary Fig. 2b and Supplementary Data 4).
Genetic diversity and affinities of ancient Hanging Coffin populations
The 14 ancient Hanging (Log) Coffin individuals were categorised into three groups based on their geographical origins: Yunnan, Guangxi, and Thailand. Our analysis revealed that individuals from Guangxi (HangCoff_Guangxi hereafter) and Thailand (LogCoff_Thailand hereafter) exhibited genetic homogeneity within their respective groups, while individuals from Yunnan (HangCoff_Yunnan hereafter) showed significant divergence (Figs. 1b and 2a). In general, almost all Hanging Coffin individuals clustered with typical southern populations in East Asia, but they showed substructure according to their geographical sites (Fig. 1b). Most individuals from Yunnan (except DSM_484 and DSM_1067) and the two individuals from Guangxi were genetically similar, clustering with coastal Neolithic southern East Asians and ancient populations from Mainland Southeast Asia (MSEA). Although DSG_4 did not differ from the other Hanging Coffin individuals in the PCA, it was slightly different from other Yunnan ancient individuals in pairwise-qpWave analysis (Fig. 2a). Furthermore, HangCoff_Yunnan (notably individual DSG_4) and HangCoff_Guangxi shared substantial drift with each other, ranking among the top 10 populations in mutual affinity, though both also showed high drift sharing with ancient individuals from Guangxi and Taiwan (Supplementary Fig. 4a and Supplementary Data 6), and we found that except for Guangxi_Yiyang (together with HangCoff_Guangxi are located in Baise city region, Z = − 3.09), no other Z score of f4(Mbuti, HangCoff_Yunnan; ancient East Asians, HangCoff_Guangxi) ≤ − 3 (Supplementary Data 7a). Specifically, the f4(Mbuti, ancient East Asians; HangCoff_Guangxi, HangCoff_Yunnan) test suggests that HangCoff_Yunnan may carry slightly more genetic ancestry related to Yellow River populations than HangCoff_Guangxi, as indicated by the f4(Mbuti, YR-related ancestry; HangCoff_Guangxi, HangCoff_Yunnan) statistic, where 2 < Z < 3 (Supplementary Data 11b). Furthermore, the f4(Mbuti, ancient East Asians; DSG_4, HangCoff_Yunnan) test also suggests that HangCoff_Yunnan harbours only marginally more Yellow River farmer-related ancestry than DSG_4, with f4(Mbuti, YR_LBIA; DSG_4, HangCoff_Yunnan) yielding a Z-score of 2.391 (Supplementary Data 8b). All Z-scores fall below the commonly accepted threshold for statistical significance ( | Z | ≤ 3). Moreover, using qpWave analysis, we verified the genetic homogeneity between HangCoff_Yunnan and HangCoff_Guangxi populations is even higher than that between HangCoff_Guangxi and the published Guangxi cave burial population (Supplementary Fig. 5). These results together indicate a clear genetic affinity between Hanging Coffin populations from Yunnan and Guangxi, and a secondary close affinity to the coastal Neolithic southern East Asians in China (Supplementary Fig. 4a), which is in line with the proposed Mount Wuyi origin of Hanging Coffin custom5,18 and supports our findings from a matrilineal perspective8.

a Pairwise qpWave modelling of ancient individuals from Hanging Coffin sites in Yunnan, Guangxi, and Thailand, based on 1240 K SNP data. Individuals DSM_1067 and DSM_484 (Wa Shi site, Yunnan; ~ 1200 BP) show significant genetic divergence (highlighted in red). p-values indicate whether individual pairs share the same number of ancestry streams, computed using qpWave with a block jackknife; values are two-sided and unadjusted for multiple testing (outgroups: Mbuti71, Russia_EHG72, Tianyuan73, Dushan15, DevilsCave_N21, Mongolia_N_North20, Haimenkou_BA10, YR_MN23, Liangdao219, Vietnam_BA1, Hanben_Taiwan20 and BaBanQinCen15). b qpWave results for Bo individuals from Qiubei. QBBR20, QBBR21, and QBBR28 are significantly differentiated (highlighted in blue). Statistical parameters and outgroups are identical to (a). c Unsupervised ADMIXTURE analysis (K = 7) of ancient and modern East Asian populations, including two Native American groups. Individuals from this study are highlighted. Most Hanging Coffin and Bo individuals exhibit predominant southern East Asian ancestry, with varying northern and Tibeto-Burman contributions. DSM_484 shows strong affinity with the upper Yellow River and Himalayan groups, while DSM_1067 aligns with populations from the Mongolian Plateau and Northeast Asia. ADMIXTURE details are shown in Supplementary Fig. 6. Abbreviations: MSEA, Mainland Southeast Asia, ISEA, Islands of Southeast Asia. Source data are provided in this paper.
Unexpectedly, two individuals (DSM_484 and DSM_1067) from the Wa Shi site (瓦石) in Yunnan, dated to ~ 1200 years ago, exhibited pronounced genetic divergence (Figs. 1b, 2a, c). DSM_484 displayed close genetic affinities with Upper Yellow River (YR) farming populations and Tibetan-related ancient groups. It also clustered closely with previously reported ancient individuals from southwestern China, including those from Gaoshan (4500 BP, Sichuan) and Haimenkou (2500 BP, Yunnan)10, with whom it shared substantial allele drift (Fig. 1b, Supplementary Fig. 5 and Supplementary Data 6). In contrast, DSM_1067 was genetically aligned with ancient Northeast Asian (ANA) populations, especially those from the Mongolian Plateau, with which it shared the greatest drift affinity (Fig. 1b, Supplementary Fig. 5 and Supplementary Data 6). The degree of genetic differentiation between these two individuals is notable when compared to other members of the Hanging Coffin cultural group (Fig. 2a and Supplementary Fig. 5), suggesting complex population structure within the Hanging Coffin tradition and possible episodes of long-distance population movement. These findings raise intriguing questions about interregional contact and the incorporation of non-local individuals into the Hanging Coffin communities. Notably, the genetic affinities of DSM_484 and DSM_1067 to Yellow River–Tibetan-related and Mongolian Plateau populations, respectively, were markedly distinct from other ancient and modern populations (Supplementary Data 9b and 10b), indicating distinct ancestral sources and potential gene flow events into Southwest China during the late first millennium CE.
Consistent with the above findings, ADMIXTURE analysis revealed that most Chinese Hanging Coffin individuals shared predominant ancestry components with Tai-Kadai-speaking populations from southeastern coastal China and MSEA, as well as with their associated ancient counterparts (e.g., Liangdao219, BaBanQinCen15, and Hanben_Taiwan20). However, DSM_484 harboured predominant ancestry components (represented in blue) mainly found in Tibeto-Burman speakers from the Himalayas and Southwest China (and their relevant ancient population Haimenkou_BA10), as well as ancient YR-related populations. DSM_1067 exhibited a dominant ancestry component (depicted in purple) identical to ANA-related populations (e.g., DevilsCave_N21, Mongolia_N_North20 and Ulaanzuukh_SlabGrave22) (Fig. 2c). These findings suggest a degree of gene flow from Northeast Asia and northern China into the Hanging Coffin communities in Southwest China, possibly coinciding with broader demographic movements during the Tang Dynasty (618–907 AD).
Using qpAdm modelling, we estimated ancestry proportions in the three primary Hanging Coffin groups (HangCoff_Yunnan, HangCoff_Guangxi, and LogCoff_Thailand). The qpAdm modelling revealed that the primary ancestry sources for these populations could be traced back to three main genetic groups: Tanshishan19 (late Neolithic population from southeastern coastal China), Upper_YR_LN23 (Upper Yellow River Late Neolithic), and Laos_Hòabìnhian1 (Southeast Asian hunter-gatherers) (Fig. 3a and Supplementary Data 17). The results indicated that the majority of Yunnan and Guangxi Hanging Coffin populations (HangCoff_Yunnan and HangCoff_Guangxi) harboured significant ancestry from the ancient eastern coastal East Asians (approximately 61–63%), with a notable contribution from YR-related ancestry (approximately 27–30%) and a smaller proportion of Hòabìnhian ancestry (around 6–11%). In contrast, the LogCoff_Thailand population exhibited a higher proportion of Hòabìnhian ancestry (20.1%), indicating that interactions with local Southeast Asian populations likely influenced the formation of the Log Coffin tradition. Furthermore, DSM_484 exhibited a predominantly Yellow River-related genetic profile (approximately 88.5%), with a minor contribution from Hòabìnhian-associated ancestry (11.5%). In contrast, DSM_1067 showed a stronger genetic affinity with ancient Northeast Asian populations, particularly resembling individuals from the Mongolian Plateau (Fig. 3a and Supplementary Data 17). While this pattern may indicate some degree of genetic input from Northeast Asia and northern China into the Hanging Coffin populations around 1200 years ago, the limited sample size and localised nature of the evidence suggest that such influence was likely sporadic or peripheral rather than a dominant demographic force. These results may reflect individual episodes of gene flow or limited-scale mobility during a period of increased population movement, possibly associated with the broader socio-political dynamics of the Tang Dynasty (618–907 AD), rather than sustained migratory influxes.

a qpAdm-estimated ancestry proportions for ancient individuals associated with the Hanging (Log) Coffin tradition (Yunnan, Guangxi, Thailand) and three genetically differentiated Bo sample sets (QBBR20, QBBR21, QBBR28). Ancestry components are modelled using Tanshishan (Late Neolithic), Upper Yellow River, and Hòabìnhian-related sources. Points represent point estimates; error bars denote ± 1 standard error, calculated via block jackknife. Detailed modelling parameters are provided in Supplementary Data 17 and 18. b Schematic map depicting approximate dispersal scenarios of the Hanging Coffin mortuary tradition and associated population movements. Coloured regions represent major ancestry source areas. Scenario I aligns with archaeological evidence placing the origin of the Hanging Coffin custom in southern China5. Scenario III indicates contributions from Hòabìnhian-related ancestry, possibly introduced locally within southern China, consistent with archaeological findings of Hòabìnhian-like tools and ancestry in earlier populations across the region10,15,37,38. Source data are provided in this paper.
Although geographically more southern, the three Log Coffin individuals from Thailand exhibited a genetic shift toward ancient Yellow River farmers and Han Chinese populations (Fig. 1b), paralleling genetic signals observed in previously published ancient Thai populations (Thailand_1700BP17 and Thailand_IA1). This supports the above inference of elevated Yellow River–related ancestry in LogCoff_Thailand (Fig. 3a). Consistent with a recent report17, Log Coffin samples from northwestern Thailand share ancestry components with local Tai-Kadai, Austroasiatic and Sino-Tibetan language speakers, indicating complex admixture dynamics in northwestern Thailand (Fig. 2c). As expected, LogCoff_Thailand shares the greatest number of drift alleles with recently published Log Coffin individuals from the surrounding region17, followed by HangCoff_Yunnan and HangCoff_Guangxi, suggesting genetic affinities among these groups (Supplementary Fig. 4a). However, multiple f₄-statistics indicate that these groups do not form a simple clade, and qpAdm modelling reveals notable differences in ancestry components, highlighting genetic heterogeneity across Log and Hanging Coffin-associated individuals. These results likely reflect regional genetic interactions or shared ancestry rather than unidirectional or large-scale migration. Importantly, methods such as f₄-statistics, qpAdm, and ADMIXTURE do not allow inference of directionality, and therefore cannot substantiate claims of a continuous demographic influx from the Yellow River basin into northwestern Thailand. Interestingly, previously published cave burial-associated individuals (GaoHuaHua) from Guangxi15 also shared a high level of drift alleles with LogCoff_Thailand (Supplementary Fig. 4a), highlighting the potential demographic history relationship of mortuary traditions in these regions. In addition, the elevated Hòabìnhian-related ancestry in Log Coffin individuals likely reflects recent admixture with Austroasiatic-speaking agriculturalists who interbred with local Hòabìnhian groups in MSEA24.
Characterising genetic profile of the Bo people
The present-day Bo people formed three distinct genetic clusters, with the majority (18 individuals), along with QBBR20_QBBR21, clustering with southern East Asian populations (Figs. 1b and 2b). Consistently, ADMIXTURE analysis revealed genetic components in the Bo people resembling those of southern Chinese ethnic groups, including Tai-Kadai, Hmong, Miao, Lahu and She populations. Their genetic makeup primarily reflected a southern East Asian component (red), with a smaller proportion of Yellow River-related and Tibeto-Burman ancestry (blue) (Fig. 2c). Compared to other ancient East Asian populations, the Bo people (except QBBR28) shared the highest level of allele drift with HangCoff_Yunnan, followed by Hanben_Taiwan (which was relevant to the Ami ethnic group)20 (Supplementary Fig. 4b and Supplementary Data 6). Furthermore, nearly all f₄(Mbuti, QBBR; ancient East Asians, HangCoff_Yunnan) tests yielded Z-scores > 3, indicating a statistically significant genetic affinity between the QBBR group and HangCoff_Yunnan. A few ancient Guangxi/Taiwan/Vietnam populations shared similar alleles ( | Z | ≤ 2) with the QBBR compared with HangCoff_Yunnan, reflecting the situation of QBBR sharing the highest drift alleles with present-day living ethnic groups from the same regions, likely suggesting their common ancestry (Supplementary Figs. 4c, 7b and Supplementary Data 13). These observations demonstrate that the present-day Bo people likely inherited their primary ancestry from ancient Hanging Coffin practitioners of Yunnan (or relevant ancient coastal southern East Asians). In addition, a couple of YR-related populations (Shimao_LN, Upper_YR_IA, Nyingchi0.1k and YR_LBIA) also showed comparable allele sharing with QBBR ( | Z | ≤ 2) relative to HangCoff_Yunnan, reflecting the Bo people’s Tibeto-Burman linguistic affiliation and suggesting recent northern genetic influence. Unexpectedly, compared to present-day living populations, the Bo people shared more drift alleles with the PhuLa and Lahu ethnic groups, who are Tibeto-Burman speakers from northern Vietnam and southern China (Supplementary Fig. 7b). The affinity degree of Bo people with PhuLa and Lahu is significantly different from that with other populations, underscores the role of localised gene flow and cultural contact with surrounding Tibeto-Burman-speaking populations over time. (Supplementary Fig. 4d and Supplementary Data 16). In addition, in f4(Mbuti, ancient East Asians; QBBR/QBBR20_21, HangCoff_Yunnan), we observed several significant differences (Z < − 3), including those from ancient Tibetans, YR-related farmers and ANA-related populations (Supplementary Data 13), suggesting that, like other southern groups (e.g., Tai-Kadai and Hmong-Mien speakers25,26, the Bo have undergone recent northern genetic admixture. Among the Bo individuals, the QBBR set harbours more northern ancestry than the QBBR20_21 set does (f4(Mbuti, ancient northern East Asians; QBBR, QBBR20_21), Z < − 3 for Russia_Shamanka_EBA, Nepal_Suila_LateC, Nepal_Kyang_LIA, Liaoning_WLR_MN and Tibet_Zongri5.1k) (Supplementary Data 13 and 14). In contrast to these observations, we identified an outlier, QBBR28, which stands apart from other Bo individuals but clusters with southern Sino-Tibetan speakers (such as Lahu) in the PCA (Fig. 1b). QBBR28 displayed higher drift affinity with Yellow River–related ancient populations (Supplementary Fig. 4b). The elevated proportion of northern ancestry in the Bo individuals may reflect historical demographic interactions during the Tang Dynasty (618–907 AD), aligning with broader patterns of movement and admixture in this period.
Using qpAdm modelling, we modelled the Bo people’s ancestry proportions, testing alternative source populations by sequentially incorporating diverse outgroups (Fig. 3a). We identified two-way models incorporating HangCoff_Yunnan ancestry and Haimenkou_BA10 (one of the earliest Bronze Age sites on the Yunnan-Guizhou Plateau of Southwest China) ancestries. The three QBBR sample sets could be modelled as a mixture of 43.4%–79.3% of HangCoff_Yunnan-related ancestry and from 20.7% to 56.6% of Haimenkou_BA-related ancestry (Fig. 3a). Taken together with previous results, these findings provide strong evidence that the Bo people most likely inherited their primary ancestries from the ancient practitioners of the Hanging Coffin tradition in China. It suggests shared ancestry that spans thousands of years, and aligns with the cultural continuity observed among the Bo people, as their funerary practices have evolved from the ancient Hanging Coffin burial custom to the more recent soul cave burial practice. Variation in Haimenkou_BA ancestry across QBBR subsets may reflect both deep shared ancestry with other ancient southwestern populations and more recent genetic and cultural (e.g., linguistic) interactions with neighbouring groups. This observation supports the notion that, although the Bo people share the majority of their genetic components with southern East Asians (Figs. 1b, 2c), they speak a Tibeto-Burman language belonging to the Sino-Tibetan family, which is thought to have originated in northern China27. To investigate the degree of genetic isolation among the Bo people, we analysed runs of homozygosity (ROH), linkage disequilibrium (LD), inbreeding coefficients, and population dynamics, comparing them with those of other East Asian populations and well-documented isolated groups such as the Andamanese2 (Supplementary Figs. 8, 9). In terms of ROH patterns, the QBBR individuals show a similar landscape to certain East Asian populations, such as the Lahu and Yakut, though with subtle differences compared to other East Asians. Notably, we identified four Bo individuals as outliers (Supplementary Fig. 8a), designated as QBBR_o, who exhibit extremely long ROH segments ( > 10 Mb), indicative of probable consanguineous unions (Supplementary Fig. 8b). Excluding these outliers, the Bo showed LD and inbreeding patterns comparable to those of other East Asian populations (Supplementary Fig. 8b–e). Consistently, similar to several other populations (e.g., CHS and BEB) and the Arras culture-associated individuals from the Yorkshire region in England28, the Bo population shows evidence of a modest recent population contraction, and the same is true for Lahu, Naxi and Hezhen populations (Supplementary Fig. 9). This demographic trend may reflect geographic localisation, prolonged isolation, or distinct ancestral origins29. Alternatively, recent admixture between diverged ancestral lineages mimic the signature of recent demographic contraction28. In summary, although the Bo inhabit remote and mountainous terrain, they do not exhibit genomic features indicative of long-term extreme isolation (Supplementary Figs. 8, 9).
Discussion
Throughout human history, diverse and creative burial practices have evolved, reflecting the cultural and spiritual beliefs of different societies. Monumental structures like the Great Pyramid of Giza, the Mayan tombs at Chichen Itza, and the Qin Shi Huang Mausoleum, or polarising rituals like Tibetan and Mongolian sky burials, all serve as cultural markers that reveal much about the people who built them. The Bo people’s Hanging Coffin custom is unique in that it stands in stark contrast to the surrounding cultures, many of which traditionally buried their dead. The Bo people placed their deceased in a wooden Coffin carved from a single tree and then suspended it from sheer cliff faces dozens or hundreds of metres in the air. In this study, in terms of ancient and modern comparative genomics exploration, we traced the genetic ancestries, admixture profiles and population histories of ancient Hanging Coffin practitioners and remnant living Bo people in Southwest China.
Our findings provide valuable insights into the genetic, cultural, and historical roots of this burial custom. Through comprehensive comparative genomics, we traced the genetic ancestries and population histories of the ancient Hanging Coffin practitioners and the modern Bo people. We revealed that most ancient Hanging Coffin individuals from Southwest China exhibited significant genetic similarity with each other and that their mainstream ancestries may initially originated from southeastern coastal China. These ancestries are associated with the late Neolithic predecessors of present-day living Tai-Kadai and Austronesian speakers (Fig. 2c). This observation reinforces the idea that serving as the earliest Hanging Coffin sites distribution region (~3600 years ago), Mount Wuyi in southeastern coastal China was the cradle of this custom5,30. The dispersal of the Hanging Coffin culture from southern China to Mainland Southeast Asia (MSEA) is supported by a convergence of archaeological, historical, and genetic evidence. Similar wooden coffin burial practices, typically involving log coffins placed in caves or on cliff faces, have been documented across southern China (e.g., Fujian, Jiangxi, Guangxi) and into Sichuan, Yunnan, northern Thailand, Laos, and Vietnam, all dating to the same Iron Age timeframe (ca. 2300–1000 BP)5,31. These sites exhibit shared construction methods and mortuary traditions, suggesting cultural continuity or the diffusion of funerary practices across regions. Historical Chinese sources describe the mortuary customs of the southern “Baiyue” populations, many of whom were likely Tai-Kadai speakers, as involving elevated burials5,32. Craniological evidence further supports the association of Tai-Kadai-speaking groups with the Hanging Coffin tradition33. Dendrochronological and stylistic analyses reveal strong parallels between coffins from Yunnan and those from northwestern Thailand, pointing to a possible transmission route along the Salween River corridor34. Recent archaeogenomic studies have also identified northern East Asian ancestry in ancient individuals associated with the Log Coffin culture in Thailand8,17, consistent with a population movement from southern China. This mortuary tradition likely spread alongside or through interactions with southward-expanding Sino-Tibetan-speaking groups17. Together, these lines of evidence support a model of cultural transmission and demographic movement of the Hanging Coffin custom from southeastern coastal China into MSEA.
Genetic affinities observed among ancient Hanging Coffin-associated individuals from southern China and northwestern Thailand highlight the wide-reaching scope of this cultural and genetic tradition. A recent comprehensive study of the Log Coffin populations from highland Pang Mapha in northwestern Thailand revealed that these Iron Age communities harboured a three-way admixture of Hòabìnhian hunter-gatherer, Yangtze River farmer, and Yellow River-related ancestries, components not observed in contemporaneous Iron Age individuals from northeastern Thailand17. These findings suggest that highland Pang Mapha functioned as a crucial corridor for north-south gene flow, potentially linked to the spread of Sino-Tibetan-speaking populations17.
Although the Hanging Coffin practice ceased to appear in historical records, the genetic traces left behind provide compelling evidence of a shared origin and cultural continuity that transcends modern national boundaries. Unexpectedly, we identified two ancient outliers, DSM_1067 and DSM_484, from the Wa Shi Hanging Coffin site in Yunnan Province, dating to approximately 1200 years ago. The two individuals clustered with ancient Northeast Asians from the Mongolian Plateau and Yellow River farmers from northern China and could be modelled as dominant genetic ancestries from these two resource populations (Fig. 3a). This finding suggests a probable demic diffusion from Northeast Asia and northern China into Hanging Coffin communities in Southwest China at least ~ 1200 years ago, during the demographic heyday period of the Tang Dynasty (618–907 AD) which could have been triggered by the strength and prosperity of the Tang Dynasty. This event also further implies the likely cultural inclusiveness property of the Hanging Coffin community in China. Furthermore, it is likely an independent event with the Mongol Empire’s southwards conquest of the Dali kingdom (located in Yunnan) in the 12th century during the Yuan Dynasty (1206–1368 AD)35. In addition to sharing most drift alleles with the relevant published Log Coffin populations in the region17, LogCoff_Thailand shares secondly most drift alleles with HangCoff_Yunnan and HangCoff_Guangxi, indicating not only cultural but also genetic connections among them (Supplementary Fig. 4a). Despite this, the Hanging (Log) Coffin populations from China and Thailand commonly shared only 27.4% to 33.9% YR-related ancestries, and very few mtDNA lineages (1 out of 14), with the overwhelming majority of matrilineal lineages in LogCoff_Thailand being local Indigenous lineages8. This suggests that the Hanging Coffin custom transmission is likely involved in predominant male-mediated sex-biased admixture processes that contribute from southern China to Southeast Asia. This phenomenon is akin to the spread of the Austroasiatic language in these regions36, although the timeframes between the two events greatly differ. Our study sheds light on the complex dynamics of human interaction within and between regions, and the results contribute to a deeper understanding of the sociopolitical and environmental factors that influence population mobility and intermarriage. Further analyses of paternal lineages with the addition more male ancient samples from Hanging (Log) Coffins could support this speculation.
The most striking discovery of our work is that by sharing most drift alleles with and being modelled as having overwhelming source ancestry from HangCoff_Yunnan, we found compelling evidence that the remnant Bo people most likely inherited their primary ancestry from ancient Hanging Coffin practitioners. Interestingly, oxidation degree analysis of copper pieces in ancestral Coffins of cave burials by living Bo people inferred that these copper pieces had been in use as early as 400 years ago, approaching the late Ming Dynasty (http://www.ynqb.gov.cn/info/1097/11512.htm). This timing corresponds with the period when both the Hanging Coffin practice and references to the Bo people declined in historical documentation. Moreover, over time, the funeral practices of the Bo people have undergone significant changes, evolving from the past Hanging Coffin burial custom to the soul cave burial custom today. Collectively, our study confirms the longstanding oral traditions and legends about the Bo people’s Hanging Coffin custom, which have been transmitted from generation to generation over the past several thousand years in China. In addition, our findings underscore the cultural and genetic persistence of the Bo people, despite historical disruptions such as the Ming Dynasty’s persecution of their community. This genetic continuity provides valuable insights into the resilience of Indigenous cultures in the face of external pressures. Unexpectedly, comparing to modern populations, the Bo people show a significant high degree of genetic affinity to the PhuLa and Lahu ethnic groups, which do not live in the vicinity of the Bo people (Supplementary Fig. 7b), with the exception of commonly speaking Loloish language (a branch of the Tibeto-Burman language), for which we cannot know more about the potentially complex history among these populations.
Finally, through comprehensive comparative genomic dissection, we present a panorama of the population history of the Asian Hanging Coffin burial custom and the remnant living Bo people, shedding light on its previously unclear origins. By integrating our molecular anthropology findings with multidisciplinary perspectives, we note that this once-flourishing and unique burial custom was likely initially created by descendant populations of late Neolithic southern East Asians in southeastern coastal China ~ 3000 years ago5,18,30. These ancestors were closely related to present-day Tai-Kadai and Austronesian language speakers in local and Southeast Asia. The Yangtze River and its tributary valleys served as corridors for the cultural and population movements of this burial custom and its practitioners from east to west over the last millennium. This human and cultural transmission process was accompanied by the incorporation of more genetic components that originated from the Yellow River valley as well as Northeast Asia (e.g., the Mongolian Plateau), and some of them were even involved in demic diffusion into Hanging Coffin communities. Approximately 600 years after the custom vanished from historical records, we found that the Bo people are the direct descendants of the Hanging Coffin custom’s practitioners. In addition, we discovered that the further southward expansion of Yellow River valley farmers profoundly influenced Log Coffin populations in northwestern Thailand as the custom spread to Southeast Asia. This influence probably involved Yellow River farmer males intermarrying with local Hòabìnhian cultural females, a process accompanied by cultural acculturation (Fig. 3b). Alternatively, however, regarding the presence of Hòabìnhian population-related ancestry in Hanging (Log) Coffin populations, we cannot rule out other possible scenarios, such as local absorption from southern China. This is because southern Chinese archaeological sites yielded very early Hòabìnhian lithic37,38, and evidence of Hòabìnhian population-related ancestry in older populations10,15.
Although our study has made significant strides in understanding the genetic and cultural history of the Hanging Coffin custom and Bo people, we could not present historical profiles for the earliest Hanging Coffin communities from the Mount Wuyi region, as well as other relevant sites in island Southeast Asia and the Pacific. Further exploration with additional human remains and archaeological content from these regions, incorporating interdisciplinary scientific perspectives, could contribute to a more comprehensive understanding of the history of suspended wooden coffin burial customs in the future.
Methods
Ethical statement and research permission
The archaeological excavations and ancient DNA analyses of the Hanging Coffin samples from China were authorised by the Yunnan Institute of Cultural Relics and Archaeology, Kunming, China. The genome sequencing and analysis of present-day Bo individuals were approved by the Institutional Review Board of the Kunming Institute of Zoology, Chinese Academy of Sciences (approval number: SWYX-2012008). Sample collection and open access to the Bo samples genomic data were granted by the Chinese Human Genetic Resources Administration (CHGRA) under permit number 2025SQXB000149. Archaeological research, including surveys, excavations, and ancient DNA analyses for the four individuals from northwestern Thailand, were granted by Thailand Research Fund (RDG01/0001/2544 (R.S.) and RTA6080001 (R.S.)) and Thailand Science Research and Innovation (RDG55H0006 (R.S., W.K.)), which was carried out by the Faculty of Archaeology, Silpakorn University, Thailand. Sample analyses, permission and exportation were permitted under No. WTh-0417-236, which issued by the Fine Arts Department of Thailand, Ministry of Culture, Thailand. The Hanging Coffin human remains samples from China are curated at Yunnan Institute of Cultural Relics and Archaeology, Kunming, China, and can be accessed upon request to Xue-Ping Ji and Yun Wu. The Log Coffin human remains samples from Thailand are housed at the Department of Archaeology, Faculty of Archaeology, Silpakorn University, Bangkok, Thailand, and can be accessed upon request to Rasmi Shoocongdej.
Ancient DNA laboratory work, sequencing, data processing and quality control
From our previous study8, together with one additional sample (WXLM1) from the Long Ma site and another (BK_737) from Bor Krai, we extracted 50–80 mg of bone powder per sample. Radiocarbon dating analyses were previously conducted on both Hanging Coffin sites in China and Log Coffin sites in northwestern Thailand. For the Chinese sites, analyses used either human skeletal material (Yunnan Province sites) or wooden coffin fragments (Guangxi Province site). The results yielded calibrated ages as follows: 660 ± 30 yBP for DSG site8, 1246 ± 65 yBP for WXWS site33, and 1070 ± 60 yBP for WXLM site33, all located in Yunnan. The Guangxi BS site was dated to 2490 ± 30 yBP8. For the northwestern Thailand Log Coffin sites studied herein, analyses employed wooden coffin fragments, yielding calibrated age ranges of 1240 ± 90 to 1450 ± 110 yBP for the BK site, 1540 ± 120 to 2080 ± 60 yBP for the LP site, and 1450 ± 110 to 1240 ± 90 yBP for the TLC site39.
All laboratory procedures were conducted in dedicated ancient DNA clean-room facilities40. Skeletal remains were decontaminated by wiping with 75% ethanol and 10% sodium hypochlorite (NaClO), followed by ultraviolet (UV) irradiation for 30 min per side. Outer surfaces were mechanically removed, and inner dense bone or tooth powder (50–100 mg) was collected using a dental drill or sandblaster. Ancient DNA was extracted using a modified large silica spin column method41 in the newly established dedicated ancient DNA laboratory of the Molecular Palaeontology Research Group (MPG) at the Kunming Institute of Zoology (KIZ), Chinese Academy of Sciences (CAS), Kunming, China. Briefly, bone powder was digested in 1 ml of lysis buffer containing 0.5 mM EDTA and 0.25 mg/mL Proteinase K at 37 °C with shaking for 18–24 h. After centrifugation, the supernatant was combined with chaotropic binding buffer (guanidine hydrochloride, isopropanol), passed through silica spin columns, washed with ethanol-based buffer, and eluted in TE buffer with 0.05% Tween. DNA was further purified using the MinElute system (Qiagen, Germany). Extraction blanks and positive controls were processed in parallel. Library preparation and in-solution DNA hybridisation capture were performed at the ancient DNA laboratories of MPG-KIZ and the clean room facility at the Institute of Anthropology, Xiamen University. We constructed single-stranded libraries (SSLs) for all samples using a 10 μL DNA aliquot, following an updated protocol42 omitting uracil-DNA-glycosylase (UDG) treatment. Libraries were enriched using in-solution hybridisation capture targeting both mitochondrial and nuclear DNA, including the human 1240 K SNP panel using a custom probe set produced by Twist Biosciences43. Captured products were purified, quantified, and quality-checked prior to sequencing. Final enriched libraries were sequenced on both the Element Biosciences AVITI™ platform (2 × 75 cycles) and the MGI-DNBSEQ™-T7 platform (2 × 75 cycles).
Raw FASTQ data were processed by merging paired-end reads with a minimum of 11 bp overlap and removing adaptors using LeeHom (v1.2.15)44 with the “--ancientdna” parameter. Reads were aligned to the hs37d5 human reference genome (GRCh37 + decoy sequences) using BWA (v0.7.18)45 with parameters “-l 16500 -n 0.01.” PCR duplicates were removed using DeDup (v0.12.8)46. Reads with mapping quality < 30 or length < 30 bp were filtered out using SAMtools (v1.20)47.
Terminal damage patterns were visualised using mapDamage2 (v2.2.2)48, and contamination was estimated using Schmutzi (v1.5.7)49 for mtDNA, ANGSD (v0.940)50 (for males) and ContamLD (version 0.1)51 for nuclear DNA (Supplementary Data 1 and Supplementary Fig. 1). To assess potential contamination that could affect population analyses, we performed an f₄(Mbuti, ancient East Asians; all set, deamination set) test across all 15 samples (Supplementary Data 5). Based on results from all contamination assessments, we applied the following criteria: (1) Sample TL_356: Due to > 5% nuclear DNA contamination, we restricted downstream analysis to deaminated sequences with typical ancient DNA damage, filtered via pmdtools (v0.60)52 (also used for other necessary samples). (2) Samples GXBS1 and GXBS2: Showed > 5% X chromosome contamination, possibly due to < 100 Y-SNPs. mtDNA contamination estimates were anomalous, but f₄-test results showed no significant differences, so all sequences were retained. (3) Samples DSG_4 and WXLM1: Showed 5 and 11 significant negative f₄-values, respectively; thus, only deaminated sequences were used. (4) Samples DSM_484, DSM_1067, and WXWS2: Despite having 1–3 significant negative f₄-values, low mtDNA and nuclear DNA contamination allowed the use of all sequences. (5) Sample DSG_9: Showed low terminal damage; therefore, only deaminated sequences were used (Supplementary Data 1 and Supplementary Data 5). To minimise the impact of DNA damage, 6 bp were trimmed from both ends of each read using BamUtil (v1.0.15)53. We called pseudohaploid genotypes for 1,240 K SNPs using the pileupCaller algorithm in sequenceTools (v1.5.4.0, https://github.com/stschiff/sequenceTools), resulting the recovery of 40,280 to 1,202,746 SNPs per individual across the 15 Hanging Coffin individuals, with 0.83 × to 74.49 × fold average coverage in the 1240 K SNP panel (Supplementary Data 1).
Biological sex determination, kinship detection, and uniparental haplogroup assignment of Hanging Coffin individuals
Using the captured 1240 K SNP panel, we determined the biological sex of individuals by calculating the ratio of Y chromosome coverage (YCov) to autosomal coverage (autoCov)54. Males exhibit a YCov/autoCov ratio of ~0.5, while females show ~ 0. Individuals with ratios > 0.2 were classified as male, and those < 0.05 as female. Based on this, we identified 8 males and 7 females (Supplementary Data 1).
Pairwise genetic relationships were estimated using READ (v1.01) software55 with default parameters. LP_37_397 and LP_47_398 (from Lahu Pot coffin cave) were found to be first-degree relatives; LP_37_397 was therefore excluded from downstream population analyses. Other pairs showed third-degree kinship or were unrelated (Supplementary Fig. 2a and Supplementary Data 2).
Mitochondrial consensus sequences were generated using the log2fasta tool in Schmutzi49. Mitochondrial haplogroups were assigned via Haplogrep (v3)56 and Y chromosome haplogroups via Yleaf (v3.2.1)57 (Supplementary Data 1). Mitochondrial haplogroup results were consistent with our previously reported findings8. We described the manual check for the Y chromosomal haplogroup assignment for 8 males (Supplementary Methods).
Bo People whole-genome sequencing and data processing
In 2004, in She De Village (舍得村: 24 °14'26.43“N, 103 °52'29.29“E), Qiubei County, Yunnan Province, with the consent and assistance of the Cultural Relics Management Office of Qiubei County, peripheral blood samples were collected from 32 adult male Bo people volunteers with informed consent. Pedigree and ethnic background were assessed via questionnaires, ensuring that participants were unrelated within three generations16.
Whole-genome sequencing (WGS) was performed on 30 individuals using the MGI DNBSEQ™-T7 platform (2 × 150 cycles), generating ~100 GB ( ~ 30x coverage) of raw data per sample. Personal identifiers were anonymized to ensure privacy, and all procedures complied with Chinese genetic resource regulations.
Adaptor trimming was performed with SOAPnuke (v2.1.9)58, and quality control was conducted using FastQC (v0.12.1) (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Clean reads were aligned to the GRCh37 (hs37d5) reference genome using BWA (v0.7.18)45. PCR duplicates were marked with Picard MarkDuplicates (v2.1.0)59, and sorting was performed using SAMtools47. Clean BAM files were realigned and recalibrated; variants were called using the HaplotypeCaller module in GATK4 (v4.2.6.1)60, followed by joint genotyping and variant quality score recalibration (VQSR), yielding 8,284,447 biallelic SNPs.
Because genome-wide data capture patterns of genetic relatedness that are independent of biological sex, we did not perform analyses stratified by sex. Both male and female individuals contribute equivalently to the reconstruction of population structure and demographic history when using genome-wide markers. Accordingly, the genetic information obtained from male individuals is sufficient to represent the broader ancestry and population history of the group under study.
Despite self-reported unrelatedness among Bo volunteers, PLINK (v2.0)61 detected first-, second-, and third-degree kinship in 9 of 30 samples (Supplementary Fig. 2b and Supplementary Data 4). To minimise the influence of linked SNPs prior to kinship estimation, data were pruned using PLINK with the “-indep-pairwise 200 50 0.4” parameter, and 21 unrelated individuals were retained for downstream analysis. Genetic sex was confirmed using PLINK (v1.90) (www.cog-genomics.org/plink/1.9). All were male, consistent with records. The mitochondrial and Y chromosome haplogroups were inferred and assigned via Haplogrep (v3)56 and Y-SNP Haplogroup via Hierarchy Finder62 (Supplementary Data 3).
To align with ancient DNA reference datasets, we re-called variants only for 1240 K SNPs using GATK4 (v4.2.6.1)60 HaplotypeCaller module, excluding related individuals and SNPs with > 5% missing data and Hardy-Weinberg p < 1e − 10. This resulted in 1,201,142 SNPs for analysis, used for all population genetic analyses unless otherwise noted.
Bioinformatics for population genetic analysis
To compare ancient Hanging Coffin individuals and modern Bo people to other East Asians, we merged our data with the Allen Ancient DNA Resource (AADR)63 using the EIGENSOFT (v7.2.1) mergeit (v2450) tool64, which includes both 1240 K and Human Origins (HO) datasets.
The merged HO dataset (maximising modern samples) was used for Principal components analysis (PCA)65 and ADMIXTURE (v1.3.0)66 analyses, while the merged 1,240 K dataset (maximising SNPs in ancient individuals) was used for f-statistics, qpWave, and qpAdm. PCA was conducted by projecting ancient individuals onto the first two PCs of modern samples using smartpca (v16000)65 from EIGENSOFT64 with parameters “lsqproject: YES” and “numoutlieriter: 0.” Ancestry proportions were estimated using ADMIXTURE (v1.3.0)66 with 10 parallel threads, random seeds, and cross-validation. K values from 3 to 9 were tested, with K = 7 having the lowest error. Prior to ADMIXTURE, LD pruning was performed using PLINK (v1.9) with “-indep-pairwise 200 25 0.4”.
For quantitative analysis, ADMIXTOOLS (v1.0)67 was used to compute outgroup-f3 statistics (qp3pop, v650, inbreed: YES) and f₄ statistics (qpDstat, v980, f4-mode: YES). qpWave (v410) assessed ancestry wave correlation, and qpAdm (v810) modelled ancestry proportions with “details: YES, inbreed: YES, allsnps: YES.”
Runs of homozygosity, linkage disequilibrium decay, inbreeding, and recent demographic history of the Bo people
To assess potential genetic isolation in the Bo population, we filtered the original dataset of 8,284,447 biallelic SNPs by excluding nine related individuals, SNPs with > 5% missing data, deviations from Hardy-Weinberg equilibrium (p < 1 × 10⁻¹⁰), and SNPs with a minor allele frequency (MAF) < 0.03. This resulted in a final dataset comprising 6,217,778 high-quality SNPs. The Bo dataset was then merged with publicly available genotypic data from the Human Genome Diversity Project (HGDP)68, Tibetans69, and the Andamanese2. To minimise the effects of linkage disequilibrium (LD) on the identification of runs of homozygosity (ROH), SNPs were pruned using the parameter “--indep-pairwise 200 50 0.4”, and only autosomal SNPs were retained. ROH segments were subsequently identified using PLINK (v1.9) with default settings. The decay of linkage disequilibrium (LD) in the Bo people and other groups was evaluated using PopLDdecay (v3.42)70 under default parameters. On the basis of biallelic autosomal SNPs, inbreeding was calculated via PLINK (v1.9). Finally, we estimated the recent effective population sizes (within the past 100 generations) of the Bo people, representative East Asians, and well-documented isolated groups such as the Andamanese using HapNe (v1.20240807)28. To contextualise the Bo population’s demographic history, we contrasted its inferred effective population size (Nₑ) with those of other East Asian groups and isolated Andamanese populations. Because the HGDP genotypes are mapped to GRCh38, we first converted them to the GRCh37 assembly via the UCSC LiftOver utility (https://genome-euro.ucsc.edu/cgi-bin/hgLiftOver), thereby reducing reference-allele bias and ensuring compatibility with HapNe28. The resulting VCFs were then analysed with both HapNe-LD and HapNe-IBD to obtain independent Nₑ trajectories. Because HapNe-IBD requires phased haplotypes and pre-identified identity-by-descent segments, we followed the recommended HapNe pipeline (https://github.com/PalamaraLab/HapNe) for genotype phasing and IBD detection. All analyses employed default HapNe parameter settings. Consistent with published benchmarks, HapNe-IBD produced systematically higher recent Nₑ estimates than HapNe-LD. Given that each population comprises fewer than forty individuals, some divergence between the LD- and IBD-based inferences is expected. In particular, HapNe-LD estimates may be downwardly biased in populations exhibiting pronounced cross-chromosomal LD—indicative of recent bottlenecks or admixture—thereby reducing the inferred Nₑ in those groups.
Creation of maps
The map in Fig. 1a was plotted using the ggplot2 tool (https://cran.r-project.org/web/packages/ggplot2/) in R. The base maps for Fig. 3a, b were generated using QGIS (http://qgis.org/, v3.22.x LTR, QGIS Development Team, 2022). All maps are for illustrative purposes only and do not imply any official endorsement of boundaries.
Limitations of this study
While hundreds of Hanging Coffins have been found in southern China, and we do not know how many human remains are preserved there, as a whole, the number of sampled hanging coffins, as well as the number of human remains, is limited in this study.
Inclusion and Ethics statement of Bo people samples
In June 2004, with official approval from the Cultural Heritage Administration of Qiubei County, Wenshan Zhuang and Miao Autonomous Prefecture, Yunnan Province, and under the coordination of local personnel, our research team initiated negotiations with the Shede Village Committee of Shede Yi Ethnic Township regarding the collection and genetic analysis of biological samples from individuals of the Bo people. Upon receiving formal consent from the village committee, the team was guided to Shede Natural Village by a designated local coordinator. Under the facilitation of the village head, 32 adult male volunteers of Bo descent were recruited for participation in the study. As the research focused on patrilineal inheritance patterns based on Y-chromosomal markers, only male individuals were included. All participants were thoroughly informed of the study’s aims and significance, and each provided written informed consent prior to sample collection. Peripheral blood samples were subsequently obtained from each participant for downstream genetic analyses.
Data availability
All data generated in this study have been deposited in public repositories. Whole-genome sequencing data are available in the Genome Sequence Archive at the National Genomics Data Centre under accession numbers HRA008615 (QBBR-bam) and HRA009608 (HangCoff-bam). Genome assemblies are deposited in the Genome Warehouse under accession PRJCA030009, and additional Bo people’s variant data are accessible via the Genome Variation Map under accession number: GVM000922. The Hanging (Log) Coffin’s pseudohaploid genotype data (Eigenstrat format) are available through OMIX, from the China National Centre for Bioinformation/Beijing Institute of Genomics, Chinese Academy of Sciences, under accession number OMIX008124. To protect participant confidentiality, access to raw sequencing data of the Bo people is subject to controlled access. The raw sequencing data will be made available to the scientific community for general research purposes. Researchers may request access by submitting a formal application, which must include a detailed research proposal and Institutional Review Board (IRB) approval from the applicant’s affiliated institution. Applications should be directed to the Data Access Committee of the Kunming Institute of Zoology, Chinese Academy of Sciences (KIZ, CAS), via email at: aqkaqssaq24670@163.com. Source data are provided in this paper.
References
McColl, H. et al. The prehistoric peopling of Southeast Asia. Science 361, 88–92 (2018).
Mondal, M. et al. Genomic analysis of Andamanese provides insights into ancient human migration into Asia and adaptation. Nat. Genet. 48, 1066–1070 (2016).
Lipson, M. et al. Ancient genomes document multiple waves of migration in Southeast Asian prehistory. Science 361, 92–95 (2018).
Blust, R. The prehistory of the Austronesian-speaking peoples: A view from language. J. World Prehist. 9, 453–510 (1995).
Chen, M. F. CLIFF-BURIAL IN CHINA. (Chongqing Press, 1992).
Jiang, W. X. Investigation report on hanging coffin burials in the Yibin area. Kaogu. 5, 18 (1981).
Guan, P. The mysterious cave burials of the Bo people. Jinri Minzu. 8, 2 (2004).
Zhang, X. et al. A matrilineal genetic perspective of hanging coffin custom in Southern China and Northern Thailand. iScience 23, 101032 (2020).
Zhang, X. et al. A Late Pleistocene human genome from Southwest China. Curr. Biol. 32, 3095–3109 (2022).
Tao, L. et al. Ancient genomes reveal millet farming-related demic diffusion from the Yellow River into southwest China. Curr. Biol. 33, 4995–5002 (2023).
Guo, J. et al. Genetic affinity of cave burial and Hmong-Mien populations in Guangxi inferred from ancient genomes. Archaeol. Anthropol. Sci. 16, 121 (2024).
Zhang, F. et al. Ancient genome analyses shed light on the origin and kinship among humans of a cliff tomb from southwestern China. J. Archaeol. Sci. Rep. 53, 104333 (2024).
Zhang, F. et al. Ancient genomes provide insights into the genetic history in the historical era of southwest China. Archaeol. Anthropol. Sci. 16, 120 (2024).
Wei, X. et al. Neolithic to Bronze Age human maternal genetic history in Yunnan, China. J. Genet. Genom. 52, 483–493 (2025).
Wang, T. et al. Human population history at the crossroads of East and Southeast Asia since 11,000 years ago. Cell 184, 3829–3841. e3821 (2021).
Shi, H., Li, Y., Luo, H., Ji, X. & Su, B. The genetic study on the patrilineal and matrilineal origin of the Bo people in Yunnan. Zool. Res. 27, 449–455 (2006).
Carlhoff, S. et al. Genomic portrait and relatedness patterns of the Iron Age Log Coffin culture in northwestern Thailand. Nat. Commun. 14, 8527 (2023).
Lu, J. Y. & Liu, D. S. A comprehensive study of hanging coffins in China. Tongji Univ. J. Soc. Sci. Ed. 1, 10 (1990).
Yang, M. A. et al. Ancient DNA indicates human population shifts and admixture in northern and southern China. Science 369, 282–288 (2020).
Wang, C.-C. et al. Genomic insights into the formation of human populations in East Asia. Nature 591, 413–419 (2021).
Sikora, M. et al. The population history of northeastern Siberia since the Pleistocene. Nature 570, 182–188 (2019).
Jeong, C. et al. A dynamic 6000-year genetic history of Eurasia’s Eastern Steppe. Cell 183, 890–904 (2020).
Ning, C. et al. Ancient genomes from northern China suggest links between subsistence changes and human migration. Nat. Commun. 11, 2700 (2020).
Yu, J. et al. The origin and dispersal of Austroasiatic languages from the perspectives of linguistics, archeology, and genetics. Hum. Biol. 94, 141–154 (2022).
Xie, M. X. et al. Genetic formation of Sui populations in southwest China. J. Syst. Evol. 62, 1054–1067 (2024).
Luo, T., Wang, R. & Wang, C.-C. Inferring the population structure and admixture history of three Hmong-Mien-speaking Miao tribes from southwest China based on genome-wide SNP genotyping. Ann. Hum. Biol. 48, 418–429 (2021).
Zhang, M., Yan, S., Pan, W. & Jin, L. Phylogenetic evidence for Sino-Tibetan origin in northern China in the Late Neolithic. Nature 569, 112–115 (2019).
Fournier, R., Tsangalidou, Z., Reich, D. & Palamara, P. F. Haplotype-based inference of recent effective population size in modern and ancient DNA samples. Nat. Commun. 14, 7945 (2023).
Patterson, N. et al. Large-scale migration into Britain during the Middle to Late Bronze Age. Nature 601, 588–594 (2022).
Zeng, F., Yang, Q. C. & Fu, S. J. A survey and preliminary study on the boat-shaped coffin burials in the Wuyi Mountains. Wenwu. 6, 11 (1980).
Shoocongdej, R. Log Coffin in Thailand in Southeast Asian Context. (Charnsanitwong Press, 2016).
Bellwood, P. First Farmers: The Origins of Agricultural Societies. (Wiley, 2004).
Ji, X. P. et al. Unique biological affinity of the hanging coffin people in ancient China based on craniometry of two skulls from Yunnan province. Anthropol. Sci. 113, 259–271 (2005).
Samrit, C. & Shoocongdej, R. Social networks in log coffin culture, in highland Pang Mapha District, Mae Hong Son Province. Damrong J. Fac. Archaeol. Silpakorn Univ. 2, 120–1250 (2023).
Daniels, C. Buddhist kingship and governance in the Dali Kingdom, 1140s to 1200. J. Southeast Asian Stud. 54, 590–620 (2024).
Zhang, X. et al. Y-chromosome diversity suggests southern origin and Paleolithic backwave migration of Austro-Asiatic speakers from eastern Asia to the Indian subcontinent. Sci. Rep. 5, 15486 (2015).
Wu, Y. et al. Dedan Cave: Extending the evidence of the Hoabinhian technocomplex in southwest China. J. Archaeol. Sci. Rep. 44, 103524 (2022).
Ji, X. et al. The oldest Hoabinhian technocomplex in Asia (43.5 ka) at Xiaodong rockshelter, Yunnan Province, southwest China. Quat. Int. 400, 166–174 (2016).
Hotchkis, M. A. C. et al. 14C analyses at the ANTARES AMS Centre: Dating the log coffins of northwest Thailand. Nucl. Instrum. Methods Phys. Res. B Beam Interact. Mater. 92, 27–30 (1994).
Knapp, M., Clarke, A. C., Horsburgh, K. A. & Matisoo-Smith, E. A. Setting the stage - building and working in an ancient DNA laboratory. Ann. Anat. 194, 3–6 (2012).
Rohland, N., Glocke, I., Aximu-Petri, A. & Meyer, M. Extraction of highly degraded DNA from ancient bones, teeth and sediments for high-throughput sequencing. Nat. Protoc. 13, 2447–2461 (2018).
Gansauge, M.-T., Aximu-Petri, A., Nagel, S. & Meyer, M. Manual and automated preparation of single-stranded DNA libraries for the sequencing of DNA from ancient biological remains and other sources of highly degraded DNA. Nat. Protoc. 15, 2279–2300 (2020).
Rohland, N. et al. Three assays for in-solution enrichment of ancient human DNA at more than a million SNPs. Genome Res. 32, 2068–2078 (2022).
Renaud, G., Stenzel, U. & Kelso, J. leeHom: adaptor trimming and merging for Illumina sequencing reads. Nucleic Acids Res. 42, e141 (2014).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bmc Bioinform. 25, 1754–1760 (2009).
Peltzer, A. et al. EAGER: efficient ancient genome reconstruction. Genome Biol. 17, 1–14 (2016).
Li, H. et al. The sequence alignment/map format and SAMtools. Bmc. Bioinform. 25, 2078–2079 (2009).
Jónsson, H., Ginolhac, A., Schubert, M., Johnson, P. L. & Orlando, L. mapDamage2. 0: fast approximate Bayesian estimates of ancient DNA damage parameters. Bmc Bioinform. 29, 1682–1684 (2013).
Renaud, G., Slon, V., Duggan, A. T. & Kelso, J. Schmutzi: estimation of contamination and endogenous mitochondrial consensus calling for ancient DNA. Genome Biol. 16, 1–18 (2015).
Korneliussen, T. S., Albrechtsen, A. & Nielsen, R. ANGSD: analysis of next generation sequencing data. Bmc Bioinform. 15, 356–368 (2014).
Nakatsuka, N. et al. ContamLD: estimation of ancient nuclear DNA contamination using breakdown of linkage disequilibrium. Genome Biol. 21, 199–221 (2020).
Skoglund, P. et al. Separating endogenous ancient DNA from modern day contamination in a Siberian Neandertal. Proc. Natl Acad. Sci. USA 111, 2229–2234 (2014).
Jun, G., Wing, M. K., Abecasis, G. R. & Kang, H. M. An efficient and scalable analysis framework for variant extraction and refinement from population-scale DNA sequence data. Genome Res. 25, 918–925 (2015).
Fu, Q. et al. The genetic history of ice age Europe. Nature 534, 200–205 (2016).
Monroy Kuhn, J. M., Jakobsson, M. & Günther, T. Estimating genetic kin relationships in prehistoric populations. PLoS ONE 13, e0195491 (2018).
Schönherr, S., Weissensteiner, H., Kronenberg, F. & Forer, L. Haplogrep 3-an interactive haplogroup classification and analysis platform. Nucleic Acids Res. 51, W263–W268 (2023).
Ralf, A., Montiel González, D., Zhong, K. & Kayser, M. Yleaf: software for human Y-chromosomal haplogroup inference from next-generation sequencing data. Mol. Biol. Evol. 35, 1820 (2018).
Chen, Y. et al. SOAPnuke: a MapReduce acceleration-supported software for integrated quality control and preprocessing of high-throughput sequencing data. GigaScience 7, 1–6 (2018).
Baroud, G. & Steffen, T. A new cannula to ease cement injection during vertebroplasty. Eur. Spine J. 14, 474–479 (2005).
DePristo, M. A. et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 43, 491–498 (2011).
Chang, C. C. et al. Second-generation PLINK: rising to the challenge of larger and richer datasets. Gigascience 4, 7 (2015).
Tseng, B. et al. Y-SNP Haplogroup Hierarchy Finder: a web tool for Y-SNP haplogroup assignment. J. Hum. Genet. 67, 487–493 (2022).
Mallick, S. et al. The Allen Ancient DNA Resource (AADR) a curated compendium of ancient human genomes. Sci. Data 11, 182 (2023).
Patterson, N., Price, A. L. & Reich, D. Population structure and eigenanalysis. PLoS Genet. 2, e190 (2006).
Price, A. L. et al. Principal components analysis corrects for stratification in genome-wide association studies. Nat. Genet. 38, 904–909 (2006).
Alexander, D. H., Novembre, J. & Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 19, 1655–1664 (2009).
Patterson, N. et al. Ancient admixture in human history. Genetics 192, 1065–1093 (2012).
Bergstrom, A. et al. Insights into human genetic variation and population history from 929 diverse genomes. Science 367, eaay5012 (2020).
Lu, D. et al. Ancestral origins and genetic history of Tibetan highlanders. Am. J. Hum. Genet. 99, 580–594 (2016).
Zhang, C., Dong, S. S., Xu, J. Y., He, W. M. & Yang, T. L. PopLDdecay: a fast and effective tool for linkage disequilibrium decay analysis based on variant call format files. Bioinformatics 35, 1786–1788 (2019).
Mallick, S. et al. The Simons Genome Diversity Project: 300 genomes from 142 diverse populations. Nature 538, 201–206 (2016).
Mittnik, A. et al. The genetic prehistory of the Baltic Sea region. Nat. Commun. 9, 442 (2018).
Yang, M. A. et al. 40,000-Year-Old Individual from Asia Provides Insight into Early Population Structure in Eurasia. Curr. Biol. 27, 3202–3208 (2017).
Acknowledgements
We would like to express our sincere gratitude to all thirty-two anonymous Bo volunteers from She De village, Qiubei County, Yunnan Province, for their voluntary participation and contribution of biological samples to this study. This study was supported by the National Natural Science Foundation of China (grants T2222030, T2425014, U23A20161, and 32270667), Yunnan Province “Thriving Yunnan Talent Support Programme” (2019), Thailand Science Research Fund (grants RDG01/0001/2544), the Natural Science Foundation of Fujian Province of China (2023J06013), the Major Project of the National Social Science Foundation of China (21&ZD285). This study also funded by the Institute of Suvanabhumi Studies, Thailand Academy of Social Sciences, Humanities and Arts (TASSHA), the Ministry of Higher Education, Science, Research and Innovation, and Naresuan University under the Global and Frontier Research University Fund (R2566C051). We also acknowledge the financial support from the Open Research Fund of State Key Laboratory of Genetic Engineering at Fudan University (SKLGE-2310), the Open Research Fund of Forensic Genetics Key Laboratory of the Ministry of Public Security (2023FGKFKT07), and the National Key Research and Development Programme of China (2023YFC3303701-02). The Exploration Society of China in Hong Kong and Zhaotong Institute of Cultural Relics Protection and Archaeology organised and financially supported field surveys of Hanging Coffin sites.
Author information
Authors and Affiliations
Contributions
X.Z., X.J. and C.C. conceived and designed the study. X.J. led archaeological excavations and surveys for Hanging Coffin sites in China. R. S. and W. K. collected human remains from Log Coffins in Thailand. X.J., H.S., X.Z., T.Y., Y.G., H.L., Y.W., J.C., Z.K., S.W., F.L. and S.H. contributed to the collection of additional human samples from the Hanging Coffin sites. B.S., H.S., X.J., Y.Y. and L.X. organised the collection of Bo samples. Y.H. was involved in preparing the Bo samples for whole-genome sequencing. H.Z., L.T. and Y.Z. conducted laboratory work on the samples. X.W. and W.L. participated in laboratory preparation. H.Z., L.T., Y.Z., H.H., X.Y., Y.L., K.Z., Y.X., R.W. and B.W. performed the data analysis. X.Z., X.J., C.C., H.Z. and L.T. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks David Lambert, who co-reviewed with Manoharan Kumar; Massimo Mezzavilla, Charles Higham and Selina Carlhoff for their contribution to the peer review of this work. [A peer review file is available.]
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhou, H., Tao, L., Zhao, Y. et al. Exploration of hanging coffin customs and the bo people in China through comparative genomics. Nat Commun 16, 10230 (2025). https://doi.org/10.1038/s41467-025-65264-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-65264-3
