
Abstract
Steroidal C12β/15α-hydroxylation are pivotal in synthesizing steroid drugs but remain challenging via chemical and biological methods. To address this, structure-guided divergent evolution is applied to the fungal P450 monooxygenase CYP68J5_fg. Two optimized variants, W12M5 (F107S/Q112R/N295T/V299T/R368K) and W15M4 (Q112C/D126V/V299L/A362M) are created, achieving high selectivity (97.7% for C12β- and 99.6% for C15α-hydroxylation of progesterone) alongside enhanced catalytic efficiency, effectively overcoming the classic activity-selectivity trade-off. Molecular dynamics simulations reveal that key mutations reorient the substrate by reshaping the binding pocket’s polarity and hydrogen-bonding network, enabling hydroxylation at distinct positions. High-density fermentation with engineered Pichia pastoris yields titers of 4.6 g/L 12β-OH progesterone, 10.9 g/L 15α-OH progesterone and 14.1 g/L 15α-OH androstenedione. These products serve as key intermediates for streamlined synthesis of C12-/C15-functionalized steroids such as drospirenone and C-nor-D-homo derivatives. Collectively, this study demonstrates the successful divergent evolution of a fungal P450, a strategy which has so far not been reported in the literature, highlights its broad applicability for the scalable synthesis of complex bioactive molecules.
Similar content being viewed by others

Introduction
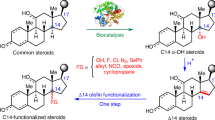
Steroids represent a therapeutically essential class of molecules, accounting for over 17% of FDA-approved drugs, with an annual production exceeding 1 million tons1,2,3. Selective C-H oxidation is pivotal for synthesizing bioactive derivatives, particularly at the C12 and C15 positions, which are critical for a number of pharmaceuticals4,5. For instance, C12α-hydroxylated deoxycholic acid is FDA-approved for fat reduction6, while 12β-hydroxy steroids serve as precursors for bioactive C-nor-D-homo steroids like cyclopamine and veratramine7,8. Similarly, C15α-hydroxylated steroids are as key intermediates for contraceptives (e.g., estetrol/drospirenone)9,10 and anti-inflammatory drugs11,12,13 (Fig. 1a, b). Despite their pharmaceutical value, these hydroxylated steroids are naturally scarce, necessitating synthetic access. Moreover, their near-identical C(sp3)-H bonds pose significant regio- and stereoselective challenges1. Existing chemical methods for C12β and C15α hydroxylation often require prefunctionalization, harsh conditions, and multi-step routes, leading to low efficiency and yield (Fig. 1c, d)14,15,16,17. This highlights the urgent need for direct and selective C12/C15-hydroxylation strategies to streamline steroid functionalization.

a Representative bioactive steroid drugs containing C12β-, C15α-hydroxyl groups. b C12β/C15α-hydroxylated steroid intermediates as key precursors for steroid drug synthesis. c, d Chemical approaches to C12β (c) and C15α (d) hydroxylation of steroids. e The present study: Divergent evolution of P450 for simultaneous access to C12β- and C15α-hydroxylated steroids.
On the other hand, the biocatalytic strategies employing cytochrome P450s (CYPs) for direct steroid hydroxylation in a highly selective fashion, has gained more and more attention due to its high catalytic efficiency and atom economy4,18,19,20. For example, microbial P450-catalyzed C11α-hydroxylation of progesterone (1, PG) streamlined cortisone production by eliminating 10–12 steps from the conventional chemical synthesis route, while reducing the cost from $200 to $1 per gram21. However, the direct C12β/C15α hydroxylation of steroids by microbial (bacterial/fungal) P450s remains underexplored. For bacterial P450s, CYP107D1 (OleP)22 from Streptomyces antibioticus exhibited only 5–10% conversion in the C12β-hydroxylation of 100 μM testosterone (5, TS). The engineered P450 BM323 from Bacillus megaterium showed 82% conversion for 2 mM 5, but with C15α-selectivity of only 73%. In contrast, fungal P450 enzymes demonstrate superior suitability for industrial-scale production, as exemplified in C11 steroid hydroxylation with fungi whole cell as catalyst21,24. Nevertheless, in terms of C12β/C15α hydroxylation, the reported C15 hydroxylases CYP5150AP2 from Thanatephorus cucumeris expressed in Mycolicibacterium neoaurum and CYP-N2 from Nigrospora sphaerica expressed in Pichia pastoris (P. pastoris) exhibited only 70% and 58.8% selectivity toward 200 mg/L and 500 mg/L substrate 1, respectively25,26. Our prior work identified an unique fungal P450 CYP68J5_fg from Fusarium graminearum with both C12β and C15α selectivity, but it exhibited poor selectivity, yielding a product ratio of ~3:5:2 (C12β, C15α- monohydroxylated and C15α-/C12β- dihydroxylated)6.
Enzyme directed evolution is an effective approach to enhance their activity and/or selectivity20,27,28,29,30,31,32. But current fungal CYP engineering efforts remain sparse since most fungal cytochrome P450s are typically endoplasmic reticulum (ER) localized with limited mechanistic insights29,33,34,35,36, which hinders their heterologous expression as well as further investigations on improving their activity, selectivity and practical applications37. Although semi-rational design improved activity of Absidia orchidis CYP5311B2 (11β-hydroxylase), yielding a 3-fold increase in hydrocortisone productivity over the wild type (WT), while maintaining unchanged stereoselectivity36. The rational engineering of Cochliobolus lunatus CYP14A improved C-14 specificity and activity towards various steroids (90–95% site selectivity, 72–78% conversion rate), but mechanistic insights and broader regioselectivity modulation require exploration29. Additionally, no studies demonstrate the divergent evolution of a fungal P450 to achieve two variants with distinct selectivity. Therefore, more research work needs to be done in this aspect, especially for P450-catalyzed direct steroid C12β/C15α hydroxylation.
Herein, our previously identified CYP68J5_fg was selected as the template, the combined structure-guided site-saturation mutagenesis (SSM) and focused rational iterative site-specific mutagenesis (FRISM) were employed to achieve the divergent evolution of CYP68J5_fg for simultaneous access to C12β/15α-hydroxylated steroids in highly selective manner (Fig. 1e). Computational analysis was performed to reveal the key residues that are responsible for the improved activity and selectivity. The substrate scope was explored for both C12β and 15α-selective variants to document their generality. High-density fermentation of recombinant P. pastoris containing the best variants was employed to produce the C12β/15α-hydroxylated steroids to demonstrate their practical applications. Finally, the chemoenzymatic process was proposed based on the isolated C12-/C15-functionalized steroids for streamlined synthesis of steroid drugs such as drospirenone and C-nor-D-homo derivatives, demonstrating a tailored fungal P450 engineering strategy broadly applicable to complex bioactive molecule synthesis.
Results
Mutational scanning on the key residues of CYP68J5_fg
CYP68J5_fg from F. graminearum was heterologously expressed in S. cerevisiae, but unfortunately resulted in poor selectivity and catalytic activity for the hydroxylation of 1, yielding a product ratio of ~3:5:2 (C12β, C15α- and C15α-/C12β- mono and dihydroxylated) (Fig. 2a)6. Due to CYP68J5_fg’s membrane-bound nature and the absence of high-identity structural templates in the Protein Data Bank (PDB), its tertiary structure was predicted using AlphaFold2 and subsequently docked with substrate 1. Substrate binding analysis revealed 15 key residues (F107, A111, Q112, S115, F125, D126, A127, L131, F212, N295, V299, A362, L363, R368, I488) located within 4 Å of the two distinct binding cavities in the C12- or C15-docked conformations of 1 (Fig. 2b). These residues, which encapsulate the 1 molecule, are likely involved in stabilizing its orientation for hydroxylation at the heme iron center. The investigation of their sequence-function relationships can guide the further engineering process. Thus, mutational scanning at all selected 15 residues was conducted by performing site saturation mutagenesis (SSM) using the 22c-trick strategy38. Variants were screened in 96-deep-well plates via high-performance liquid chromatography (HPLC) to assess hydroxylation selectivity and activity38,39.

a The result of hydroxylation of 1 by the wild-type CYP68J5_fg. b Docking of 1 (yellow) in the CYP68J5_fg heme pocket (white). The 15 residues forming interactions within a range of 4 Å near 1 in the heme center are shown, and the 7 key amino acids that affecting the activity and selectivity are shown in magenta. c Optimal variants obtained by SSM of 7 amino acid residues using the CYP68J5_fg gene as a template. Reaction conditions: S. cerevisiae cells were used for fermentation, the final concentration of 1 was 1 mM, and the reaction time was 24 h. The data shown in c are presented as the mean ± s.d. of three biological replicates. Source data are provided as a Source Data file.
After screening the library (~1395 variants), the results revealed that most variants exhibited reduced activity or selectivity. However, seven residual sites (F107, Q112, D126, N295, V299, A362, R368) emerged as exceptions, showing enhanced activity or selectivity (Fig. 2b). Notably, variants with improved activity with moderate-to-high selectivity for 1 at each site were presented in Fig. 2c. Strikingly, selectivity diverged between two pathways: C12β selectivity primarily correlated with five residues (F107, Q112, N295, V299, R368), while C15α selectivity involved four residues (Q112, D126, V299, A362). Mutations at Q112 and V299 were pivotal for switching regioselectivity in CYP68J5_fg, toggling between C12β and C15α hydroxylation of 1 when Q112 alternated long-chain (R) and short-chain (A/C/S/T/M) residues or V299 switched between polar (T) and nonpolar (I/L) amino acids (Fig. 2c). However, these results suggest that single mutations at key residues can modulate activity or selectivity but are insufficient to fully invert selectivity. Thus, we hypothesized that combinatorial mutations at multiple sites might synergistically enhance pathway-specific selectivity with the aid of revealed sequence-function relationships. To test this, we implemented focused rational iterative site-specific mutagenesis (FRISM)40,41,42 since it avoided laborious mutant library screening with minimal screening effort for selectivity and activity evaluation (Fig. 3a). Two distinct mutagenesis routes were designed to optimize selectivity for either C12β or C15α, as detailed in the subsequent study.

a SSM and FRISM engineering strategies for generating a library of CYP68J5_fg variants. b Selectivity and conversion rate results for C12β-related variants in the transformation of 1, where pink bars indicate conversion rate and blue squares indicate selectivity. c Selectivity and conversion rate results for C15α-related variants in the transformation of 1, where blue bars indicate conversion rate and pink squares indicate selectivity. d, e Typical HPLC chromatograms of the C12β (d) and C15α (e)-related variants of CYP68J5_fg against 1 transformation. Reaction conditions: S. cerevisiae cells were used for fermentation, the final concentration of 1 was 1 mM, and the reaction time was 24 h. The data shown in (b, c) are presented as the mean ± s.d. of three biological replicates. Source data are provided as a Source Data file.
FRISM applied on CYP68J5_fg for improved selectivity and activity
Next, based on the revealed sequence-function relationships by mutational scanning, FRISM was carried out by combining the beneficial mutations from single variants that showed positive effects on activity (conversion rate of substrate 1) and/or selectivity. Given that most single mutations in CYP68J5_fg related to C12β selectivity of 1 are accompanied by reduced activity, the variant N295T (W12M1), which exhibited the highest activity, was chosen as the starting template for further mutagenesis. Next, four double variants were developed by combining N295T with mutations at four additional sites (F107S, Q112R, V299T, R368K). After screening the resulting variants, the optimal double variant Q112R/N295T (W12M2) was obtained, which demonstrated a 20% increase in activity and a 46% enhancement in selectivity (relative to WT). Subsequently, based on W12M2, three more triple mutants were generated by individually introducing mutations F107S, V299T and R368K. The best-performing triple mutant, Q112R/N295T/V299T (W12M3) boosted the selectivity to 97% with 51% conversion. Finally, to further enhance the activity, quadruple (W12M4 (F107S/Q112R/N295T/V299T)) and quintuple (W12M5 (F107S/Q112R/N295T/V299T/R368K)) mutants were correspondingly constructed by integrating mutations F107S and R368K. Consequently, variant W12M5 showed the best catalytic performance, achieving 82.3% conversion towards 1 while maintaining its selectivity at 97.7% (Fig. 3b, d).
Similarly, to enhance both selectivity and activity of CYP68J5_fg-catalyzed C15α-hydroxylation of 1, the identified single variant V299L (W15M1), which demonstrated a 39% increase in selectivity over the WT, was selected as the initial template. Then, based on W15M1, double variants were created by individually introducing mutations at sites Q112, D126, and A362 (Q112A/M/T/S/C, D126T/V, A362M). Among them, the double variant D126V/V299L (W15M2) demonstrated the most significant improvement, achieving 17% increased activity and 8% enhanced C15α selectivity relative to W15M1. Subsequently, based on W15M2, tertiary and quaternary variants were generated by introducing mutations of Q112A/M/T/S/C and A362M, respectively. As a result, the quadruple mutant Q112C/D126V/V299L/A362M (W15M4) emerged as the optimal variant, exhibiting 99.6% C15α selectivity with 72.7% substrate conversion (Fig. 3c, e).
It is noteworthy that, while engineering P450 enzymes often results in a “trade-off” between selectivity and catalytic activity, where high selectivity typically diminishes at elevated substrate conversions43. However, our combined SSM and FRISM strategies for P450 engineering successfully overcame this classical limitation by just screening 29 variants. Consequently, divergent evolution of CYP68J5_fg was achieved, leading to two distinct CYP68J5_fg variants W12M5 and W15M4 for highly active and selective C12β and C15α-hydroxylation, respectively. Variant W12M5 boosted the C12β selectivity from 37.1% (for WT) to 97.7% with 82.3% substrate conversion (only 28.1% for WT). Parallel to this achievement, variant W15M4 enhanced the C15α-selectivity to 99.6% from 49.3% with 72.7% substrate conversion.
Molecular mechanism of CYP68J5_fg variants for selective steroid hydroxylation
To elucidate the mechanisms underlying the enhanced catalytic efficiency and selectivity of the CYP68J5_fg variants W12M5 (F107S/Q112R/N295T/V299T/R368K) and W15M4 (Q112C/D126V/V299L/A362M), comprehensive structural and computational analyses were performed by using molecular dynamics (MD) simulations.
The W12M5 variant achieved high selectivity for the C12β-hydroxylation of 1. MD simulation results showed that the F107S mutation created a larger hydrophilic space, which forces the nonpolar methyl groups of 1 to relocate from the polar environment toward more compatible hydrophobic regions. Meanwhile, the Q112R mutation introduced a positively charged guanidinium group, while the N295T substitution reduced local hydrophilicity by removing a polar side chain, collectively altering the electrostatic landscape around the C3 keto group and driving it toward the BC loop through polarity-mediated spatial rearrangement (cyan arrows, Fig. 4a). In addition, the R368K and V299T mutations, which reduce side-chain polarity and enhance hydrophilicity respectively, flank the C20 carbonyl group of 1 and generate asymmetric polarity gradients. These, together with a weak hydrogen bond between H302 and the substrate ((H302)N···O(1) = 3.86 ± 0.74 Å, Supplementary Fig. 5a), collectively guide the C20 carbonyl group of 1 towards the I helix (magenta arrows, Fig. 4a). These concerted modifications establish an optimized catalytic microenvironment favoring C12-specific oxidation, with a mean 12 C···O=Fe(heme) distance of 3.56 ± 0.22 Å and a mean angle of 134 ± 13° formed by O(Fe=O)-(1)-H(12 C)-(1)-C(12) (Fig. 4b, c). The close agreement between these structural parameters and the QM-optimized transition state (TS) for C12β-H abstraction (4TSH,C12, Supplementary Fig. 4) is consistent with the geometric requirements essential for efficient TS formation and reaction feasibility.

a, d Representative snapshots of the MD simulation revealing the substrate-enzyme binding mode in the active sites of W12M5 (a) and W15M4 (d). Mutated residues are shown in red. b, e Plots of the C12, C15(1)···O=Fe distances (y primary axis) and the O(Fe=O)-(1)-H(12 C, 15 C)-(1)-C(12, 15) angles (y secondary axis) along the simulation time (x axis) for one of the replicas of W12M5 (b) and W15M4 (e) (see Supplementary Figs. 1, 2 for replicas 2 and 3). c, f Conformational population analysis of the prereaction states of W12M5 (c) and W15M4 (f) in the 100 ns MD simulation trajectories (three joint MD replicas). Quantum mechanics (QM)-predicted distances (distQM) and angles (angQM) of transition state geometry (Supplementary Figs. 3, 4) for C12β-H/C15α-H abstraction are indicated by dashed lines (b, e) and black dots (c, f), respectively. The data shown in (b, c, e, f) are presented as the mean ± s.d. of three biological replicates. Source data are provided as a Source Data file.
In contrast to W12M5-1 in pose 12 (positioning C12 proximal to the heme for β-hydroxylation), the V299L mutation in W15M4 introduces a steric hindrance effect through the elongated hydrophobic side chain of leucine, thereby driving a reverse orientation of the substrate’s nonpolar methyl groups to minimize hydrophobic packing conflicts. The Q112C and D126V mutations collectively reduce local polarity, while the N295 residue forms a strong hydrogen bond with the substrate ((N295)N···O(1) = 2.84 ± 0.12 Å, Supplementary Fig. 5b), orienting the C3 keto group of 1 toward the I-helix (cyan arrows, Fig. 4d). The C20 carbonyl group of 1 is sandwiched between the hydrophobic side chain of M364 and the polar residue R368, forming a polarity-asymmetric cleft that drives the functional group to migrate toward the polar side. Moreover, R368 further anchors the C20 carbonyl oxygen via hydrogen bonding ((R368)N···O(1) = 3.12 ± 0.39 Å, Supplementary Fig. 5c), guiding the carbonyl group toward the β2 sheet (magenta arrows, Fig. 4d). Consequently, pose 15 (positioning C15 proximal to the heme for α-hydroxylation) emerges as the dominant active conformation in the W15M4-1 complex, characterized by a mean C15···O=Fe(heme) distance of 3.92 ± 0.33 Å and a mean angle formed by O(Fe=O)-(1)-H(15 C)-(1)-C(15) of 144 ± 9° (Fig. 4e, f), both of which are consistent with QM-predicted values for C15α-H abstraction (4TSH,C15, see Supplementary Fig. 4). Moreover, MD simulations of CYP68J5_fg with substrate 1 in poses 12 and 15 (Supplementary Fig. 6a–d), variant W12M5 with 1 in pose 15 (Supplementary Fig. 6e, f), and variant W15M4 with 1 in pose 12 (Supplementary Fig. 6g, h) revealed that the majority of substrate-binding conformations sampled during the trajectory proved to be non-productive, as expected. Interestingly, in variant W12M5, substrate 1 flipped ~90° at 20 ns and ~180° by 25 ns relative to its initial pose, ultimately adopting a C12-binding mode (Supplementary Fig. 7) This spontaneous rearrangement further confirms the instability of C15-binding and strongly supports the C12-hydroxylation specificity of W12M5.
In summary, our MD simulation results offer valuable molecular insights into the binding of 1 in CYP68J5_fg variants. The introduced mutations reshaped the polar distribution of substrate binding pocket, altering the key residue network involved in hydrogen bonding with the polar functional groups (C3 keto group and C20 carbonyl group) of 1. These findings elucidate how polarity remodeling at the binding pocket governs regioselective hydroxylation at C12β or C15α.
Exploring substrate scope of variants of CYP68J5_fg
The catalytic capacity of CYP68J5_fg variants to catalyze regio- and stereoselective hydroxylations at the C15α and C12β of steroids was evaluated. We tested the activity and selectivity of different variants of CYP68J5_fg on a range of steroids: canrenone (2), bisnoralcohol (3), 17α-hydroxyprogesterone (4), testosterone (5), androstenedione (6), followed by scaling up of top-performing mutants (Supplementary Fig. 8). Variants favoring C15α hydroxylation on substrate 1 also selectively hydroxylated substrates 2, 3, 5, and 6, with substrate conversion ranging from 46.7% to > 99.0% and selectivity ranging from 93.8% to > 99.0% (Table 1). Due to the endogenous C17 reductase in S. cerevisiae44, 6 can be reduced to 5, the selectivity of variant W15M4 towards 6 was confounded by its conversion to 5. The capability of this reductase for converting 6 to 5 was further confirmed by its heterologous expression in E. coli (Supplementary Fig. 9). Future work on optimization of the yeast chassis (e.g., reductase gene knock out) could potentially eliminate this reduction activity. However, substrate 4 was not accepted by this variant. MD simulations revealed that the C17 hydroxyl group of 4 forms a stable hydrogen bond with the heme Fe=O center (d4, Supplementary Fig. 10), enforcing a near-planar conformation and resulting in a suboptimal O(Fe=O)-(4)-H(15 C)-(4)-C(15) attack angle (θ = 120 ± 13° vs. ideal 170°), which sterically hinders hydrogen abstraction, leading to the loss of the activity. On the other hand, the variants that prefer C12β hydroxylation at position 1 also selectively act on substrates 2, 3, and 4, exhibiting a preference for C12β hydroxylation. These mutants, exhibiting 72.6% to > 99.0% substrate conversions with the selectivity all being above 94.0% (Table 1). Compared to the acetyl side chain of 1, the computational analysis showed that the enhanced polarity at the C17 position in 5 and 6 promotes strong hydrogen bonding with residue K368 (d5 and d6, Supplementary Fig. 10) in W12M5, inducing a conformational shift that displaces C12 from the Fe=O center and blocks oxidation. Significantly, structural characterization revealed three previously unreported steroidal hydroxylated products: the C12β-hydroxylated derivatives of 2 and 3, as well as the 15α-hydroxylated derivative of 3. Notably, differences in substrate chemical properties influence hydrogen bonding and conformational changes, which in turn affect the catalytic performance of variants and lead to substrate-specific inactivation. Thus, leveraging sequence-function relationship, these variants can be further optimized to enhance their selectivity or/and activity towards these steroidal substrates. Given the special synthetic value of C15 and C12 positional functionalization of steroids, as exemplified in the production of glucocorticoids or deoxycholic acid4,6. Consequently, the discovery of CYP68J5_fg and its variants further diversifies the enzyme toolkit and provides excellent biocatalysts for industrial steroid drug production.
Scale-up reactions with generated variants W12M5 and W15M4
Compared to S. cerevisiae, which shows limited recombinant protein production despite robust fermentation, P. pastoris achieves higher yields due to its Crabtree-negative metabolism that prevents carbon loss to ethanol under respiration, increasing biomass and protein production45,46. We finally evaluated the performance of the variants W12M5 and W15M4 for hydroxylated steroid production under fed-batch fermentation by using a P. pastoris strain GS115 (Supplementary Fig. 11). For the recombinant strain GS115-W12M5, methanol induction began after 49 h of glycerol growth. After 24 h of induction, 10 g of 1 and cyclodextrin was added at a molar ratio of 1:1. As in Fig. 5a, after 144 h of fermentation, the yield of product 1a reached 4.6 g/L, with an isolation yield of 45.1% (purity > 95%). This non-linear reaction profile suggested product inhibition, a hypothesis we validated by adding varying concentrations of 1a to a fixed concentration of substrate 1 (Supplementary Fig. 12). Worth noting is that, the conversion of 1 to 1a catalyzed by GS115-W12M5 represents the highest yield of 12β-hydroxylation of 1 achieved by any fungal-derived P450 mutant in yeast to date. Moreover, the rearrangement of 12β-hydroxysteroids could yield highly bioactive C-nor-D-homo-steroids featuring an unusual 6-6-5-6 ring system, significantly simplifying synthetic routes to natural products containing this scaffold. As exemplified by the previous study7, it had been demonstrated that 1c undergoes rapid skeletal rearrangement within 20 min. Therefore, 1a could potentially undergo skeletal rearrangement via this approach to afford corresponding C-nor-D-homo-steroids (Fig. 5a).

a High-density fermentation of 1 produces 1a by GS115-W12M5 for chemoenzymatic synthesis of 7. b High-density fermentation of 6 produces 6a by GS115-W15M4 for chemoenzymatic synthesis of 8. The data shown in (a, b) are presented as the mean ± s.d. of three biological replicates. Source data are provided as a Source Data file.
For the recombinant strain GS115-W15M4, we also tested the strain’s ability to transform compounds 1 and 6. As shown in Supplementary Fig. 13, for the transformation of 1, cyclodextrin-solubilized 1 was added in batches (10 g and 8 g) after 24 h of induction, and after 148 h of fermentation, the production of 1b finally increased to 10.9 g/L, with an isolation yield of 71.6% (purity > 95%). Furthermore, as the 15α-hydroxylated derivative of 6 (15α-OH-AD) is a key intermediate in the synthesis of the contraceptive drug drospirenone47,48. Using the engineered strain GS115-W15M4, we achieved a 15α-OH-AD (6a) titer of 14.1 g/L in fed-batch fermentation with 20 g/L AD loading, which represent the highest reported yield for steroid hydroxylation by a fungal-derived P450 in yeast (Fig. 5b). Conventional fungi (e.g., Fusarium culmorum47, Penicillium griseopurpureum49, or Fusarium equiseti48) catalyze AD 15α-hydroxylation, but generally yield off-target byproducts (6β-, 7α-, or 11α-hydroxylated derivatives). In contrast, W15M4 exhibits exceptional regioselectivity for C15 hydroxylation. Coupled with the high-density fermentation capacity of P. pastoris, this system offers a superior alternative route for efficient drospirenone production47,48.
These results indicate that P. pastoris serves as a suitable host organism for engineering a promising cell factory for high-yield steroid production, advancing the feasibility of industrial-scale steroidal drug manufacturing. Notably, as shown in Fig. 1a, b, C12-hydroxylated and C15-hydroxylated sterols possess critical pharmacological activities. By achieving high-density fermentation of CYP68J5_fg and its mutants in P. pastoris, optimization of the synthetic pathway of these steroidal drugs offers significant synthetic possibilities.
Discussion
Traditionally, despite extensive engineering of bacterial P450s for steroidal transformations, fungal P450s are not well suited for optimizing enhanced selectivity or activity. Key limitations include inefficient heterologous expression, ER-restricted localization, and the absence of both high-resolution structures and homologous templates for structure-guided design. Furthermore, current engineering of fungal P450s primarily focuses on enhancing single-position selectivity in steroid transformations. To overcome these limitations, we developed a combined structure-guided SSM with FRISM mutagenesis to achieve the divergent evolution of CYP68J5_fg in S. cerevisiae. This approach yielded two variants: W12M5 (F107S/Q112R/N295T/V299T/R368K) and W15M4 (Q112C/D126V/V299L/A362M), which selectively hydroxylate the steroidal substrate 1 to produce 1a and 1b with high selectivity (>97%), respectively. It is noteworthy that, both variants overcome the traditional selectivity-activity trade-off. It demonstrates that the integrated approach of SSM coupled with FRISM strategy effectively modulates selectivity and enhances activity in CYP68J5_fg, with potential applicability to other fungal-derived P450 enzymes. Simultaneously, MD simulations of the optimal mutants W12M5 and W15M4 reveal how key amino acids direct 1 to the desired C12 or C15 orientation. These findings offer valuable insights into the precise control of regio- and stereoselectivity by evolved CYP68J5_fg mutants and highlight the utility of computer simulations in the engineering of non-structure-dependent P450 enzymes derived from fungi. Furthermore, it marks a significant advancement in the field of engineering fungal-derived P450s within S. cerevisiae.
In addition, we further employed the C12- and C15-targeting variants to transform other substrates (2–6), enhanced regiospecificity and conversion efficiency with great surprising for most substrates while maintaining selectivity profiles comparable to those observed for substrate 1. Notably, the variants proved most effective at enforcing C12 or C15 hydroxylation specificity, leading to the identification of three previously unreported hydroxylated products (Supplementary Figs. 14–58). Subsequently, we achieved the production of 1a and 1b up to 4.6 g/L and 10.9 g/L respectively, with isolation yields of 45.1% and 71.6% (purity > 95%) by high-density fermentation of variants W12M5 and W15M4 in P. pastoris, which represent the highest yields of hydroxylation at the C12 position of steroids by fungal-derived P450s in yeast. Significantly, 1a may undergo a potential C/D-ring skeletal rearrangement of the steroid to afford C-nor-D-homo steroids. Moreover, high-density fermentation of the strain GS115-W15M4 yielded 14.1 g/L of the drospirenone intermediate 15α-OH-AD, representing the highest reported titer for fungal P450-mediated steroid hydroxylation in yeast. These results demonstrate the industrial viability of our engineered variants, showcasing a streamlined chemoenzymatic approach for synthesizing C12- and C15-functionalized steroids. Along a different line, we point out that our study contains numerous examples of synergistic (cooperative) and antagonistic non-additive mutational effects uncovered by so-called deconvolution50.
In conclusion, the robust activity and selectivity of the well-characterized variants towards various substrates pave the way for practical industrial applications. By developing a systematic approach that integrates the engineering of fungal P450 enzymes in S. cerevisiae with high-density fermentation in P. pastoris, this work offers a blueprint for future and industrial applications of fungal P450 enzymes. Additionally, we believe that our proposed chemoenzymatic approach will provide a robust platform for subsequent medicinal chemistry exploration of different steroidal drugs. In other words, our study highlights the great potential of tailoring fungal P450 enzymes, offering important opportunities for the synthesis of complex steroidal drugs through the combination of computational simulations and synthetic biology.
Methods
Functional expression of fungal P450 in S. cerevisiae
S. cerevisiae strain INVSc1 cells were prepared for transformation with pYES2 expression plasmid harboring CYP68J5_fg or mutants through a standard protocol for lithium acetate transformations51. Transformed cells were plated on uracil dropout plates containing 2% (w/v) glucose. After 2–3 d of growth at 30 °C, colonies were inoculated in 2 mL uracil dropout minimal medium containing 2% (w/v) glucose and grown for 24 h at 30 °C with shaking at 220 rpm. The seed solution was then transferred to 20 mL uracil dropout minimal medium containing 2% (w/v) glucose for 2 days. Cultures were induced for expression by resuspension (2:1) in uracil dropout minimal media containing 4% (w/v) galactose. Expression cultures were grown at 30 °C with shaking at 220 rpm. For biotransformations, substrates were dissolved in DMF or 2-hydroxypropyl-β-cyclodextrin solution and spiked into the expression culture at the point of induction. After reaction, ethyl acetate was added with an equal volume for product extraction, the organic phase was then obtained by centrifugation, followed by evaporation to dryness before resuspended in methanol for HPLC analysis.
Site saturation mutagenesis (SSM)
Site saturation mutagenesis (SSM) was performed by two-step PCR approach with non-overlapping primers. Additionally, degenerate 22-condon trick primers were designed for each of the 15 active site residues required for SSM (gene sequences and associated primers are provided in the Supplementary Data 1)38. In the first step, the primers were to be mixed in a molar ratio of 12 (NDT): 9 (VHG): 1 (TGG) before mixing with a non-mutagenic (silent) primer in a 1:1 ratio. The mixture was then subjected to first PCR for 28 cycles to obtain short DNA fragments containing mutagenic sites with KOD -Plus- Neo Polymerase. The recovered DNA fragments then serve as a megaprimer to perform the second PCR with KOD -Plus- Neo Polymerase for whole plasmid amplification with 28 cycles, and the resulting PCR products were digested with DpnI (NEB) overnight at 37 °C to eliminate the parental template. And the PCR products were transformed into E. coli DH5α by electroporation. Subsequently, cells were recovered for 45 min at 37 °C in LB medium, aliquots were plated on LB agar plates supplemented with 100 μg/mL ampicillin (LB-Amp plates), and plates were incubated at 37 °C overnight. Resulting colonies were grown up and miniprepped to give a plasmid DNA library. S. cerevisiae strain INVSc1 cells were prepared for transformation with the plasmid DNA library through a standard protocol for lithium acetate transformations. Transformed cells were plated on uracil dropout plates containing 2% (w/v) glucose and incubated at 25 °C for 3 d. Resulting colonies were used directly in library screening.
Site-directed mutagenesis (SDM)
Site-directed mutagenesis (SDM) was performed by one-step PCR approach with non-overlapping primers like SSM. For SDM, mutagenic primer was added in a 1:1 ratio with non-mutagenic (silent) primer and then 28 cycles of PCR were performed using Primer STAR Max DNA Polymerase. Finally, the clones developed on agar plate were picked and subjected to DNA sequencing for confirmation.
High throughput biotransformations
Uracil dropout minimal media containing 2% (w/v) glucose was added to sterile 96-well culture plates to a volume of 500 μL39. S. cerevisiae library colonies were inoculated into each well of the plate except for row A1, A2 and A3, which were reserved for S. cerevisiae control colony, CYP68J5_fg wild type and no colony control. Cultures were grown for 36 h at 30 °C with shaking at 800 rpm. Typically, an aliquot of 100 μL was transferred to a glycerol stock plate and stored at −80 °C for culture collection. Culture plates were centrifuged at 2820 × g for 10 min to pellet cells and the growth media was removed. Cells were resuspended with 200 μL of uracil dropout minimal media containing 4% (w/v) galactose and 300 μM progesterone (PG, 1). The biotransformations were incubated at 30 °C with shaking at 800 rpm for 2–3 days. Reactions were quenched with the addition of 3 equivalents of ethyl acetate and centrifugation at 2820 × g for 10 min. The organic phase was transferred into a 96-wells shallow plate and placed in the fume cupboard until the solvent complete evaporation. The dried samples were resuspended in 200 μL methanol and passed through a PTSF 96-well plate filter to remove solid particles (Pall, VWR, Germany) into a new 96-wells shallow plate. The plates were closed using silicon lids and subjected to HPLC analysis.
Hit validation and sequencing
Functional validation of selected hit variants was performed by culturing from glycerol stocks and conducting triplicate biotransformation assays followed by HPLC analysis. Variants demonstrating enhanced activity relative to the parental template were selected for sequencing to identify mutations in active sites. For this purpose, plasmid DNA was extracted from S. cerevisiae using the TIANprep Yeast Plasmid DNA Kit, transformed into DH5α E. coli competent cells, and subsequently purified using a Plasmid Miniprep Kit. The purified plasmids were submitted to Sangon Biotech (Shanghai, China) for Sanger sequencing.
Screening of CYP68J5_fg mutants for hydroxylation of diverse steroids
Sterile 96-well plates were filled with 500 μL of uracil dropout minimal medium containing 2% (w/v) glucose. Each well was inoculated with a distinct CYP68J5_fg mutant colony, with each mutant replicated three times per substrate. The plates were incubated at 30 °C for 36 h with shaking at 800 rpm. Subsequently, the cultures were centrifuged at 2820 × g for 10 min to pellet the cells, and the supernatant was carefully removed. Cells were resuspended in 200 μL of uracil dropout minimal medium supplemented with 4% (w/v) galactose and 300 μM of the respective substrate: canrenone (2), bisnoralcohol (3), 17α-hydroxyprogesterone (4), testosterone (5), or androstenedione (6). The biotransformations were carried out at 30 °C with shaking at 800 rpm for 48–72 h. Reactions were terminated by adding three volumes of ethyl acetate relative to the reaction mixture, followed by centrifugation at 2820 × g for 10 min. The organic extracts were transferred to a shallow 96-well plate and evaporated to dryness in a fume hood. The residues were reconstituted in 200 μL methanol and filtered through a PTSF 96-well filter plate (Pall, VWR, Germany) into a new shallow 96-well plate. Plates were sealed with silicone lids prior to HPLC analysis.
Scale-up reaction for product preparation with mutants of CYP68J5_fg
For large-scale product preparation, a preculture of S. cerevisiae INVSc1-CYP68J5_fg mutant was used as the inoculum. A single colony was inoculated into 100 mL of SC-Ura medium (2% (w/v) glucose) in a 1 L shake flask and cultivated at 30 °C with shaking at 250 rpm for 48 h. Cells were harvested by centrifugation and resuspended in 50 mL of fresh SC-Ura medium (4% (w/v) galactose). To the culture, we added a steroid substrate dissolved in 2-hydroxypropyl-β-cyclodextrin to a final concentration of 1–2 mM, along with 5-aminolevulinic acid hydrochloride (5-ALA) at a final concentration of 0.5 mM. The biotransformation was continued for 3 days under the same temperature and shaking conditions. This process can be readily repeated and scaled as required.
Functional expression in P. pastoris
The expression plasmids harboring CYP68J5_fg mutants was linearized with PmeⅠ restriction enzyme (NEB), and 1-2 μg of linearized DNA was transferred into GS115 electrocompetent cells by electroporation. Electroporated cells were grown on YPD plates containing 100 μg/mL zeocin at 30°C for 2–3 days. For screening transformants, colonies were inoculated in BMGY medium and grown for 2 days at 30 °C with shaking at 220 rpm, cultures were induced for expression by resuspension (2:1) in BMMY medium at 25 °C with shaking at 220 rpm and supplemented daily with methanol to maintain 1% (v/v) methanol in the cultures. For biotransformations, substrate was spiked into the expression culture at the point of induction.
Fed-batch fermentation in P. pastoris
Fermentation control was adapted from the Pichia Fermentation Process Guidelines (PFPG, Invitrogen) and the fermentation process included three phases. First, the batch phase (phase Ⅰ), the pH of BSM medium was maintained at 5.5 by adding aqueous ammonia, and the temperature was maintained at 28 °C, and the aeration rate of 2–3 vvm. A single colony was first inoculated into a 1 L Erlenmeyer flask containing 150 mL of YPD medium at 30 °C with shaking at 220 rpm for ~24 h. Then, 10% (v/v) of the seed culture was inoculated into 1.5 L BSM medium with 2 g/L histidine in a 5 L fermenter for fed-batch fermentation. After glycerol was exhausted (~24–28 h) (phase Ⅱ), the fed-batch phase was initiated by feeding 50% (w/v) glycerol with 2 g/L histidine and PTM1 at a speed of 4–5 mL/min to biomass increasing, and the dissolved oxygen (DO) was maintained at ~30%. After the OD600 reached ~150 (~24–30 h), glycerol feeding was stopped to allow cells to deplete the glycerol and shifted to methanol feeding (phase III), the methanol feeding rate at 4–7 mL/min until the end of fermentation, the temperature was set to 25 °C and the DO was maintained at ~20%. The substrate 1 and cyclodextrin are added in a molar ratio of 1:1.
Quantum Mechanical (QM) calculations
The P450 enzyme, coordinated with an iron-bound porphyrin complex, features an Fe=O double bond denoted Compound I (Cpd I). To simplify the model of Cpd I, the porphyrin macrocycle was represented with methyl groups for all substituents, and the axial Cys449 ligand was modeled as ethylthiolate anion. To preserve the protein scaffolding, geometry optimizations were performed with the atoms at the truncation sites constrained to their crystallographic positions. All calculations were carried out using unrestricted density functional theory (UDFT) with the hybrid functional B3LYP, as implemented in the Gaussian 16 program package52. Geometry optimizations were performed with the LANL2DZ basis set for the Fe atom, the 6–31 G + (d,p) basis set for all other atoms. Based on the optimized geometries, more accurate single-point energies were obtained using larger basis sets, namely LANL08+ for Fe and cc-pVTZ(-f)+ for nonmetallic elements. To incorporate the solvation effects of the protein environment, single-point calculations at the same level of theory were conducted based on the conductor-like polarizable continuum model (CPCM)53,54. The dielectric constant (ε) is set to 4, which is a conventional value for modeling protein environments55,56. Frequency calculations were performed at the same level of theory as the geometry optimizations to characterize the stationary points and provide zero-point energy (ZPE) corrections. Following geometry optimization of the enzyme-substrate complex in various electronic states, the quartet state was identified as the ground state (Supplementary Tables 1 and 2). This finding is consistent with previous research57, which establish that the quartet C-H abstraction transition state (TS) consistently exhibits lower energy. Thus, all subsequent calculations were conducted exclusively in the quartet state. P450-mediated hydroxylation involves the hydrogen atom abstraction from the C12/C15 position of PG by Cpd I, followed by a hydroxyl radical rebound. With computational results confirming hydrogen abstraction as the rate-determining step (Supplementary Fig. 3), its TS structure was used as a key reference to evaluate the prereactive conformations sampled in MD simulations.
Molecular docking
Molecular docking was conducted employing LeDock software58, recognized for its high performance and accuracy. Initial ligand preparation involved converting the PG substrate structure (retrieved in SDF format from PubChem) to mol2 format. Concurrently, the receptor structures, including the predicted CYP68J5_fg and its variants, were prepared in PDB format. Docking poses were clustered based on a Root Mean Square Deviation (RMSD) cutoff of 1.0 Å. A docking box was defined as a 5 Å-radius sphere centered on the heme using PyMOL’s GetBox plugin59, delineating the expected three-dimensional ligand-binding region. The simulation was set to produce 20 distinct binding poses, which were ranked by their predicted binding energy scores. From these energy-ranked poses, catalytically competent conformations were identified by evaluating both the distance and orientation of the target C12-H or C15-H bonds relative to the ferryl oxygen of Compound I. Thus, the final selected conformation represents the pose that is both thermodynamically stable (most negative binding energy) and mechanistically viable for C12/C15 hydroxylation.
Molecular Dynamics (MD) simulations
Molecular dynamics (MD) simulations were performed using AMBER 2260 based on the CYP68J5_fg (or its variants)/PG complex obtained by molecular docking. Parameters for the substrate (PG) were generated with the Antechamber module in AmberTools 23, employing the Generalized AMBER Force Field (GAFF)61. Atomic partial charges were obtained by fitting the electrostatic potential at the HF/6-31 G(d) level using the restrained electrostatic potential (RESP) approach. Parameters for heme compound I and axial cysteine residue were taken from established computational protocols for heme-protein modeling62. The remaining protein residues were described by the Amber ff14SB force field63, and the protonation states of titratable residues were predicted using the PROPKA3 server64. The complex structures were solvated in a cubic TIP3P65 water box, maintaining a minimum distance of 16 Å between the protein surface and the box edges. Each system was neutralized with Na+ counterions.
The complex system underwent initial energy minimization through 5000 steps of steepest descent followed by 5000 steps of conjugate gradient optimization, with positional restraints applied to the protein using a force constant of 500 kcal·mol−1Å−2. An additional 5000 steps of steepest descent and 5000 steps of conjugate gradient minimization were then performed to fully optimize the system without restraints. Subsequently, the system was heated from 0 to 300 K over 500 ps under the NVT ensemble, with backbone atoms constrained by a force constant of 25 kcal·mol−1Å−2. Next, density equilibration was conducted for 1.0 ns using the NPT ensemble to achieve uniform density.
The Langevin thermostat66 was used to control the temperature at 300 K with a collision frequency of 1 ps−1, while the pressure was kept at 1 bar with a relaxation time of 2 ps. Covalent bonds involving hydrogen atoms were constrained with the SHAKE method67, employing a 2 fs integration time step. A cutoff radius of 12 Å was set for nonbonded interactions. Following 10 ns of equilibration under the NPT ensemble, a 100-ns productive MD simulation was performed in the NPT ensemble. Trajectory analysis was conducted using the AmberTools23 CPPTRAJ module.
HPLC analysis
The conversion analysis was performed via reversed phase HPLC technique using a Shimadzu LC2030C system with an Ultimate® XB-C18 column (4.6 × 250 mm, 5 μm, Welch Materials (Shanghai) Co., Ltd.). For steroids (1, 3 and 4), the mobile phase consisted of acetonitrile (CH3CN) and water with a gradient program of CH3CN: H2O = 30: 70–80: 20 for 0–12 min, 80: 20–70: 30 for 12–15 min at 1.5 mL/min flow rate and 40 °C oven temperature. The injection volume was 10 μL and UV detection was accomplished at 254 nm. For steroids (2, 5 and 6), the mobile phase consisted of CH3CN and water with a gradient program of CH3CN: H2O = 10: 90 for 0–2 min, 10: 90-40: 60 for 2–10 min, 40: 60–70: 30 for 10–15 min, 70: 30–90: 10 for 15–16 min, 90: 10 for 16–17 min at 1.5 mL/min flow rate and 40 °C oven temperature. The injection volume was 10 μL and UV detection was accomplished at 254 nm expect for 2, for which 310 nm was used.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data that support the findings of this study are available within the main text and its Supplementary Information or from the authors upon request. Source data are provided with this paper.
References
Zhao, L., Bo, Z. & Yang, Y. Enzymes on steroids. Nat. Synth. 2, 699–700 (2023).
Peng, H. et al. A dual role reductase from phytosterols catabolism enables the efficient production of valuable steroid precursors. Angew. Chem. Int. Ed. 60, 5414–5420 (2021).
Sun, C. et al. Establishing an efficient electron transfer system for p450 enzyme OleP to improve the biosynthesis of murideoxycholic acid by redox partner engineering. Angew. Chem. Int. Ed. 64, e202423209 (2025).
Zhang, X. et al. Rationally controlling selective steroid hydroxylation via scaffold sampling of a P450 family. ACS Catal. 13, 1280–1289 (2023).
Zhou, X. J. et al. Enantiodivergent synthesis of halohydrins by engineering P450DA monooxygenases. ACS Catal. 13, 15948–15955 (2023).
Wang, L. et al. A fungal P450 enzyme from fusarium graminearum with unique 12β-steroid hydroxylation activity. Appl. Environ. Microbiol. 89, e01963–01922 (2023).
Heretsch, P., Rabe, S. & Giannis, A. A biomimetic approach to C-nor-D-homo-steroids. J. Am. Chem. Soc. 132, 9968–9969 (2010).
Hou, W. et al. Divergent and gram-scale syntheses of (-)-veratramine and (-)-cyclopamine. Nat. Commun. 15, 5332 (2024).
Lee, A. & Syed, Y. Y. Estetrol/Drospirenone: A review in oral contraception. Drugs 82, 1117–1125 (2022).
Deng, Q. et al. Rationally designing P450BM3-H to excavate a novel channel for product exit and enhance overall performance. Int. J. Biol. Macromol. 307, 142162 (2025).
Jia, L. et al. Identification and characterization of the steroid 15α-hydroxylase gene from Penicillium raistrickii. Appl. Microbiol. Biotechnol. 101, 6409–6418 (2017).
Petzoldt, K., Laurent, H. & Wiechert, R. 3β, 7β, 15α-Trihydroxy-5-androsten-17-one and its 3, 15-dipivalate. EP75189 A1 (1983).
Petzoldt, K., Laurent, H. & Wiechert, R. 3β, 7β, 15α-Trihydroxy-5-androsten-17-one, its 3,15-dipivalate, and their preparation. US Patent 4435327 A (1984).
Offei, S. D. et al. Chemical synthesis of 7-oxygenated 12α-hydroxy steroid derivatives to enable the biochemical characterization of cytochrome P450 8B1, the oxysterol 12α-hydroxylase enzyme implicated in cardiovascular health and obesity. Steroids 140, 185–195 (2018).
See, Y. Y., Herrmann, A. T., Aihara, Y. & Baran, P. S. Scalable C-H oxidation with copper: synthesis of polyoxypregnanes. J. Am. Chem. Soc. 137, 13776–13779 (2015).
Williams, J. R., Gong, H., Hoff, N., Olubodun, O. I. & Carroll, P. J. α-Hydroxylation at C-15 and C-16 in cholesterol: synthesis of (25R)−5α-Cholesta-3β,15α,26-triol and (25R)−5α-Cholesta-3β,16α,26-triol from diosgenin. Org. Lett. 6, 269–271 (2004).
Abas, H., Blencowe, P., Brookfield, J. L. & Harwood, L. A. Selective hydroxylation of C(sp3)-H bonds in steroids. Chem. Eur. J. 29, e202301066 (2023).
Zhang, Z. et al. A designed chemoenzymatic route for efficient synthesis of 6-dehydronandrolone acetate: a key precursor in the synthesis of C7-functionalized steroidal drugs. ACS Catal. 13, 13111–13116 (2023).
Peng, Y. et al. A chemoenzymatic strategy for the synthesis of steroid drugs enabled by P450 monooxygenase-mediated steroidal core modification. ACS Catal. 12, 2907–2914 (2022).
Li, A. et al. Regio- and stereoselective steroid hydroxylation at C7 by cytochrome P450 monooxygenase mutants. Angew. Chem. Int. Ed. 59, 12499–12505 (2020).
Carballeira, J. D. et al. Microbial cells as catalysts for stereoselective red-ox reactions. Biotechnol. Adv. 27, 686–714 (2009).
Agematu, H. et al. Hydroxylation of testosterone by bacterial cytochromes P450 using the Escherichia coli expression system. Biosci. Biotech. Bioch. 70, 307–311 (2006).
Chen, W., Fisher, M. J., Leung, A., Cao, Y. & Wong, L. L. Oxidative diversification of steroids by nature-inspired scanning glycine mutagenesis of P450BM3 (CYP102A1). ACS Catal. 10, 8334–8343 (2020).
Petzoldt, K., Annen, K. & Laurent, H. Process for the preparation of 11-beta-hydroxy steroids. EP19800103478 (1980).
Sui, L., Chang, F., Wang, Q., Chang, Z. & Xia, H. Functional reconstitution of a steroidal hydroxylase from the fungus Thanatephorus cucumeris in Mycolicibacterium neoaurum for 15α-hydroxylation of progesterone. Biochem. Eng. J. 193, 108859 (2023).
Li, S., Chang, Y., Liu, Y., Tian, W. & Chang, Z. A novel steroid hydroxylase from Nigrospora sphaerica with various hydroxylation capabilities to different steroid substrates. J. Steroid Biochem. Mol. Biol. 227, 106236 (2023).
Qu, G., Li, A., Acevedo-Rocha, C. G., Sun, Z. & Reetz, M. T. The crucial role of methodology development in directed evolution of selective enzymes. Angew. Chem. Int. Ed. 59, 13204–13231 (2020).
Kille, S., Zilly, F. E., Acevedo, J. P. & Reetz, M. T. Regio- and stereoselectivity of P450-catalysed hydroxylation of steroids controlled by laboratory evolution. Nat. Chem. 3, 738–743 (2011).
Song, F. et al. Chemoenzymatic synthesis of C14-functionalized steroids. Nat. Synth. 2, 729–739 (2023).
Sun, C. et al. Engineering the substrate specificity of a P450 dimerase enables the collective biosynthesis of heterodimeric tryptophan-containing diketopiperazines. Angew. Chem. Int. Ed. 62, e202304994 (2023).
Gao, H. et al. Mechanistic insights into the selective C-S bond formation by P450 TleB. ACS Catal. 14, 10658–10669 (2024).
Deng, Q. et al. Modification strategy of two-step ordered selective hydroxylation of DHEA catalyzed by CYP-cl3. J. Agric. Food Chem. 73, 8062–8072 (2025).
Zhang, X. et al. Bacterial cytochrome P450-catalyzed regio- and stereoselective steroid hydroxylation enabled by directed evolution and rational design. Bioresour. Bioprocess. 7, 2 (2020).
Klingenberg, M. Pigments of rat liver microsomes. Arch. Biochem. Biophys. 75, 376–386 (1958).
Chen, J. et al. Production of 14α-hydroxysteroids by a recombinant Saccharomyces cerevisiae biocatalyst expressing of a fungal steroid 14α-hydroxylation system. Appl. Microbiol. Biotechnol. 103, 8363–8374 (2019).
Chen, J. et al. Identification of Absidia orchidis steroid 11β-hydroxylation system and its application in engineering Saccharomyces cerevisiae for one-step biotransformation to produce hydrocortisone. Metab. Eng. 57, 31–42 (2020).
van den Brink, H. M., van Gorcom, R. F. M., van den Hondel, C. A. M. J. J. & Punt, P. J. Cytochrome P450 enzyme systems in fungi. Fungal Genet. Biol. 23, 1–17 (1998).
Kille, S. et al. Reducing codon redundancy and screening effort of combinatorial protein libraries created by saturation mutagenesis. ACS Synth. Biol. 2, 83–92 (2013).
Zetzsche, L. E. et al. Biocatalytic oxidative cross-coupling reactions for biaryl bond formation. Nature 603, 79–85 (2022).
Li, D., Wu, Q. & Reetz, M. T. Methods in Enzymology. (ed Dan S. Tawfik) Vol. 643, 225–242 (Academic Press, 2020).
Xu, J. et al. Stereodivergent protein engineering of a lipase to access all possible stereoisomers of chiral esters with two stereocenters. J. Am. Chem. Soc. 141, 7934–7945 (2019).
Xu, K. et al. (+)−3,6-Epoxymaaliane: a novel derivative of (+)-bicyclogermacrene oxidation catalyzed by CYP450 BM3-139-3 and its variants. ChemBioChem 25, e202400410 (2024).
Acevedo-Rocha, C. G. et al. P450-catalyzed regio- and diastereoselective steroid hydroxylation: efficient directed evolution enabled by mutability landscaping. ACS Catal. 8, 3395–3410 (2018).
Singer, Y., Shity, H. & Bar, R. Microbial transformations in a cyclodextrin medium. Part 2. reduction of androstenedione to testosterone by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 35, 731–737 (1991).
Vieira Gomes, A. M., Souza Carmo, T., Silva Carvalho, L., Mendonça Bahia, F. & Parachin, N. S. Comparison of yeasts as hosts for recombinant protein production. Microorganisms 6, 38 (2018).
Porro, D., Sauer, M., Branduardi, P. & Mattanovich, D. Recombinant protein production in yeasts. Mol. Biotechnol. 31, 245–259 (2005).
Soeroes, B. et al. Industrial process for the preparation of 17-hydroxy-6β,7β;15β,16β-bismethylene-3-oxo-17α-pregn-4-ene-21-carboxylic acid γ-lactone and key intermediates for this process. WO2006059167 A1 (2006).
Galik, G. et al. A process for the preparation of 17-hydroxy-6β,7β,15β,16β-bismethylene-17α-pregn-4-ene-3-one-21-carboxylic acid γ-lactone and key-intermediates for this process. WO2006059168 A1 (2006).
Huang, L.-H. et al. Biotransformation of dehydroepiandrosterone (DHEA) with Penicillium griseopurpureum Smith and Penicillium glabrum (Wehmer) Westling. Steroids 75, 1039–1046 (2010).
Hollmann, F., Sanchis, J. & Reetz, M. T. Learning from protein engineering by deconvolution of multi-mutational variants. Angew. Chem. Int. Ed. 63, e202404880 (2024).
Daniel Gietz, R. & Woods, R. A. Methods in Enzymology Vol. 350 (eds Christine Guthrie & Gerald R. Fink) 87-96 (Academic Press, 2002).
Frisch, M. J. et al. Gaussian 16, Revision B.01 (Gaussian, Inc., Wallingford, CT, 2016).
Cammi, R., Mennucci, B. & Tomasi, J. Second-Order Møller-Plesset analytical derivatives for the polarizable continuum model using the relaxed density approach. J. Phys. Chem. A 103, 9100–9108 (1999).
Tomasi, J., Mennucci, B. & Cammi, R. Quantum mechanical continuum solvation models. Chem. Rev. 105, 2999–3094 (2005).
Blomberg, M. R. A., Borowski, T., Himo, F., Liao, R.-Z. & Siegbahn, P. E. M. Quantum chemical studies of mechanisms for metalloenzymes. Chem. Rev. 114, 3601–3658 (2014).
Siegbahn, P. E. M. Mechanisms of metalloenzymes studied by quantum chemical methods. Q. Rev. Biophys. 36, 91–145 (2003).
Narayan, A. R. H. et al. Enzymatic hydroxylation of an unactivated methylene C-H bond guided by molecular dynamics simulations. Nat. Chem. 7, 653–660 (2015).
Wang, Z. et al. Comprehensive evaluation of ten docking programs on a diverse set of protein–ligand complexes: the prediction accuracy of sampling power and scoring power. Phys. Chem. Chem. Phys. 18, 12964–12975 (2016).
Schrödinger, L. L. C. The PyMOL Molecular Graphics System, Version 1.8. Schrödinger, LLC (2015).
Case, D. A. et al. AmberTools. J. Chem. Inf. Model. 63, 6183–6191 (2023).
Wang, J., Wolf, R. M., Caldwell, J. W., Kollman, P. A. & Case, D. A. Development and testing of a general amber force field. J. Comput. Chem. 25, 1157–1174 (2004).
Shahrokh, K., Orendt, A., Yost, G. S. & Cheatham, T. E. 3rd Quantum mechanically derived AMBER-compatible heme parameters for various states of the cytochrome P450 catalytic cycle. J. Comput. Chem. 33, 119–133 (2012).
Maier, J. A. et al. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 11, 3696–3713 (2015).
Olsson, M. H. M., Søndergaard, C. R., Rostkowski, M. & Jensen, J. H. PROPKA3: Consistent treatment of internal and surface residues in empirical pKa predictions. J. Chem. Theory Comput. 7, 525–537 (2011).
Jorgensen, W. L., Chandrasekhar, J., Madura, J. D., Impey, R. W. & Klein, M. L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 79, 926–935 (1983).
Izaguirre, J. A., Catarello, D. P., Wozniak, J. M. & Skeel, R. D. Langevin stabilization of molecular dynamics. J. Chem. Phys. 114, 2090–2098 (2001).
Kräutler, V., van Gunsteren, W. F. & Hünenberger, P. H. J. J. O. C. C. A fast SHAKE algorithm to solve distance constraint equations for small molecules in molecular dynamics simulations. J. Comput. Chem. 22, 501–508 (2001).
Acknowledgements
This study was supported by the National Key Research and Development Program of China (grant number 2024YFA0917800 (A.L.)), the National Natural Science Foundation of China (grant number No. 32371552 (A.L.) & grant number No. 32501127 (Q.L.)), the Outstanding Science and Technology Innovation Team Program for Young and Middle-aged Scholars in Higher Education Institutions of Hubei Province (grant number T2024001 (A.L.)), the Natural Science Foundation of Wuhan City (grant number 2024040701010046 (Q.L.)), the Innovation Base for Introducing Talents of Discipline of Hubei Province (grant number No. 2019BJH021) (A.L.), and Research Program of State Key Laboratory of Biocatalysis and Enzyme Engineering.
Author information
Authors and Affiliations
Contributions
A.L. conceived the project; A.L. and Q.L. supervised the project; A.L., Q.L., C.G. and L.W. designed the experiments; L.W., Y.W. and X.P. performed the experiments and collected the data; A.L., Q.L., L.W., Y.W., X.P., C.G., X.W., P.C. and P.X. analyzed the data; A.L., Q.L.,. L.W. and Y.W. wrote the manuscript; all authors contributed review and editing to the manuscript and gave final approval for the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Zhanfeng Wang and the other anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, L., Wang, Y., Peng, X. et al. Divergent evolution of fungal P450 monooxygenase unlocks simultaneous access to C12β and C15α oxyfunctionalization of steroids. Nat Commun 16, 10713 (2025). https://doi.org/10.1038/s41467-025-65736-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-65736-6
