
Abstract
Division of labor in reproduction in eusocial insects is governed by delicate and context- dependent mechanisms, yet the regulatory processes remain largely unclear. Here, we investigate social cues involved in the establishment of reproductive dominance in bumble bee (Bombus terrestris) workers. We reveal the roles of two alkenes, (Z)-9-nonacosene (Z9- C29) and (Z)-9-hentriacontene (Z9- C31) in the activation of ovarian development. These alkenes are found throughout the queenright bumble bee colony, including wax, workers, drones, and gynes, and their levels are positively correlated with colony size. They act as quorum-sensing pheromones that facilitate ovary maturation in a threshold-dependent manner under queenright conditions. A synergistic interaction between the perception of these two alkenes and the occurrence of dominance contests is essential for the ovary development of worker bumble bees, independent of temporal sequence. Artificial addition of these two alkenes into colonies confirms their stimulatory effects on the ovarian developmental potential of worker bees, even in the presence of a queen. Our findings highlight a quorum-sensing-like mechanism that primes the initiation of worker ovary development, providing insights into the complex regulation of context-dependent reproductive plasticity in social insects.
Similar content being viewed by others

Introduction
Reproductive altruism—where workers forgo their own reproduction to rear siblings—is a hallmark of eusociality1,2. This trait varies in degree across insect species3,4. In most highly eusocial insects, such as stingless bees and honeybees, workers show extreme sterility, with atrophied or vestigial ovaries5,6. By contrast, in primitively eusocial species like polistine wasps, bumblebees, and certain ants, workers retain functional ovaries and can reproduce under specific conditions, typically producing viable haploid males via arrhenotokous parthenogenesis7,8. This reproductive capacity often leads to potential conflicts between queens and workers over male offspring parentage, creating a tension between individual and colony-level fitness interests9,10. Kin selection and inclusive fitness theory explain the evolutionary basis of worker altruism11,12. Yet worker reproduction presents a dual impact on colonies: diminish colony productivity by diverting workers from cooperative tasks to selfish egg-laying, creating a 'tragedy of the commons' scenario where individual reproductive interests undermine the collective resource of the colony's workforce13,14, or benefit the colony by altering offspring sex ratio, providing alternative reproductive strategies after the queen's death, and increasing genetic diversity4,15. Therefore, understanding the proximate mechanisms behind worker sterility or fertility in response to social contexts will deepen our insight into hymenopteran eusociality.
Worker reproduction is shaped by a complex interplay of social and environmental cues16. Traditionally, workers are viewed as sterile, with divergent reproductive interests from queens, leading to extensive research on the inhibitory effects of breeding queens, reproductive dominant workers, or brood on worker fertility17,18,19,20. However, reproductive activation can still occur in presence of these inhibitory agents, indicating that additional regulatory mechanisms are at play15,21. Evidence points to self-organized regulation of worker reproduction. For example, in Harpegnathos saltator ants, continuous antennal dueling establishes hierarchies, enabling dominant workers to transition into reproductive gamergates under queen-less conditions22,23. Furthermore, physical attributes contribute significantly to reproductive division of labor among workers in some social insects such as wasp, carpenter bees and bumble bee24. Morphological variation predicts reproductive outcomes—larger-bodied workers consistently achieve higher positions in dominance hierarchies and consequently secure greater reproductive opportunities25. Beyond these factors, the Allee effect has also been observed in social insects26, where group size influences worker reproduction: larger groups accelerate ovarian development and increase egg production27,28. Collectively, these findings suggest that ovarian activation in workers involves complex sensory integration mechanisms finely tuned to the colony's social environment. However, the specific pathways through which these factors interact to regulate reproduction remain incompletely understood.
B. terrestris provides an excellent model system for studying how social cues regulate worker reproduction. Worker reproduction commonly occurs after queen loss and can persist even in queenright colonies during the late-stage "competition phase" (CPh), where up to 40% of workers lay eggs and contributes to a substantial proportion of male offspring despite the queen’s presence29,30. While queen-controlled inhibition of worker reproduction has been emphasized31, evidence suggests its diminishing role in colonies during the CPh32,33, with other social cues becoming more significant. Studies show that factors such as colony size and social interactions among workers promote reproduction34,35, with dominance hierarchies forming among competing workers in queenless colonies36,37. In small queenless groups, dominant workers (α-workers) rapidly activate their ovaries and inhibit the ovarian activity in subordinate workers38, highlighting the importance of dominance contest. Subordinate workers (β-workers) in queenright colonies or queenless groups, as well as workers isolated and separated from others fail to fully activate their ovaries, underscoring the importance of physical interactions39,40,41. Chemical signals, particularly from colony wax, also play a role, with specific compounds linked to worker reproduction42. Evidently, the regulation of worker reproduction in bumble bees is a multifaceted process influenced by a variety of factors, suggesting the involvement of intricate and interconnected mechanisms that respond to social contexts43.
The aim of the current project is to unveil the social mechanisms underlying the promotion of worker reproduction in bumble bees. As a first step towards this goal, we establish a paradigm to assess the influence of social cues on ovarian development in bumble bee workers using pairwise dyadic assays37. Next, we identify the critical components of chemical cues that stimulate ovarian development in α-queenless workers using GC-MS/MS and electrophysiological experiments. Finally, we determine the effect of these key compounds on ovarian development under different social contexts. This study reveals a novel regulatory mechanism governing worker reproduction, in which chemical signals stimulate worker fertility within a social context through a quorum sensing-based process.
Results
Ovarian development of α-workers requires contact cues from β-workers or substrates
To investigate the influence of group size on worker reproductive development (Experiment 1a), we housed workers in varying social configurations (isolated individuals, pairs, trios, and groups of 10 or 30 workers) and measured their Terminal Oocyte Size (TOS) after 7 days. Our results revealed a clear social facilitation effect on ovarian development. Workers in dyadic (mean = 1.247 mm) and triadic groups (mean = 1.337 mm) exhibited ~2.5-fold higher of the TOS of isolated individuals (mean = 0.4929 mm), while workers in larger groups (10 workers: mean = 1.893 mm, 30 workers: mean = 1.884 mm) showed fourfold increase in TOS compared to solitary workers (Supplementary Fig. 1a, one-way ANOVA with Tukey post-hoc test, F = 73.31, df = 4, P < 0.0001). Given that the dyadic arrangement provided both clear evidence of socially facilitated reproduction and a simplified context for studying social interactions44, we established this configuration as our experimental paradigm for subsequent investigations. In dyadic groups, rearing in pairs resulted in the acquisition of reproductive dominance in one of the paired bumble bee workers (α-workers), whereas ovaries of individually isolated workers failed to activate (Fig. 1a, Kruskal-Wallis with Dunn post-hoc analysis, H = 38.98, df = 2, P < 0.0001). For paired workers, we examined the 7 day dynamics of aggressive events and that of average TOS and found dramatic ovarian development in α-workers by the last 2 days when aggressive events almost disappeared (Experiment 1b, Fig. 1b). To test whether aggression is sufficient for ovarian development of α-workers, we established a paradigm to assess the influence of social cues sensed by workers during the last 2 days of pairing on their ovarian development. Firstly, we separately transferred paired workers into new cages for 2 days after pairing for 5 days (Experiment 1c, Fig. 1c, two-way ANOVA with Bonferroni correction, F (treatment) = 5.091, df = 1, P = 0.0289). Compared with continuously paring, the absolute social isolation at the last 2 days significantly reduced ovarian development of α-workers (Fig. 1c). Furthermore, we found that the access to either filter paper in the original cage (original filter paper herein and in the following text) or to β-workers significantly increased ovarian development in α-workers compared with the absolutely isolated ones (Experiment 1d, Fig. 1d, two-way ANOVA with Bonferroni correction, F (treatment) = 111.5, df = 1, P < 0.0001; Experiment 1e, Fig. 1e, two-way ANOVA with Bonferroni correction, F (treatment) = 42.61, df = 1, P < 0.0001). These results suggested that the ovarian development of α-workers required cues from substrates or β-workers in the last 2 days. We further clarified whether sounds or volatile chemicals or sight from β-workers might be effective using a double mesh treatment (Experiment 1f). The result showed that α-workers in the double mesh group failed to develop their ovaries, compared with those that had full access to β-workers indicating the ineffectiveness of non-tactile cues from β-workers (Fig. 1f, two-way ANOVA with Bonferroni correction, F (treatment) = 32.02, df = 1, P < 0.0001). In addition, ovarian development of workers with last 2 days’ access to original filter paper (Average TOS: mean = 2.115 mm) is similar with those under continuously paring (Average TOS: mean = 2.182 mm), demonstrating contact cues from original filter paper fully recapitulated the stimulatory effects of direct social contact cues on ovarian development (Experiment 1e, Supplementary Fig. 1b, two-way ANOVA with Bonferroni correction, F (treatment) = 0.7574, df = 1, P = 0.9824). Therefore, in addition to aggressive events, contact cues from β-workers or substrates (original filter paper) are required for reproductive development of α-workers in B. terrestris.

a Average Terminal Oocyte Size (TOS) of singly isolate (n = 29) or paired workers (n = 21) housed in Petri dishes for 7 days. For each pair, the bee with the greater or lesser ovarian development was classified as an α-worker or a β-worker, respectively. b Aggression index (n = 10 for Days 1, 2, 3, 4, 5, 6 and 7, respectively) and average TOS of α-workers over 7 days in dyadic assays (n = 10, 8, 9, 10, 9, 9, 10 for Days 1, 2, 3, 4, 5, 6 and 7, respectively). Dots represent means, and the shaded areas represent the 95% CI. c Average TOS (right panel) of workers isolated in new cage (n = 12) and paired in original cage (n = 13) in Experiment 1b (left panel). The yellow or white indicates original or new filter paper in Petri dishes, respectively. d Average TOS (right panel) of workers exposed to new filter paper (n = 19) or original filter paper (n = 16) in Experiment 1c (left panel). e Average TOS (right panel) of workers isolated (n = 8) and paired (n = 8) in new cage in Experiment 1d (left panel). f Average TOS (right panel) of workers paired (n = 9) and paired under double mesh treatment (DM paired, n = 8) in new cage in Experiment 1e (left panel). All data are shown as mean ± SEM (standard mean error). Data in (a) was analyzed using Kruskal–Wallis test with Dunn’s post-hoc analysis (two-tailed) for multi-group comparisons. Data in (c−f) were analyzed by two-way ANOVA with Bonferroni’s correction (two-tailed) for multiple comparisons (adjusted P-values are shown in the respective columns). Detailed statistical test results and adjusted P-values for multiple comparison were shown in Supplementary Data 1. Source data are provided as a Source Data file.
Chemical cues involved in worker ovarian activation are ubiquitous with a dosage-dependent effect
To verify whether chemical compounds are responsible for the effect of contact cues on ovarian development of α-workers, we separately transferred workers paired for 5 days into new cages with pentane-washed filter papers or filter papers with different chemical additives for 2 days and measured the TOS of workers after this two-day period (Fig. 2a). The ovaries of α-workers transferred to cages with pentane-washed filter papers failed to develop (Experiment 2a, Fig. 2b, two-way ANOVA with Bonferroni correction, F (treatment) = 0.08598, df = 1, P = 0.7702). Pentane extracts of original filter paper led to significantly accelerating ovarian development of α-workers compared to no-adding or pentane-adding filter papers (In Experiment 2b, Fig.2c, two-way ANOVA with Tukey post-hoc test, F (treatment) = 11.09, df = 2, P < 0.0001). In all treatments, the ovaries of β-workers failed to develop, implying the necessary role of dominance effect. Therefore, the chemical compounds on the original filter paper were both necessary and sufficient, serving as primer pheromone(s) for ovarian development of α-workers in the last 2 days.

a Schematic diagram of dyadic assays to assess the effect of chemicals. FP: filter paper. b Average TOS of workers on original FP (n = 16) and odorless original FP (n = 20). c Average TOS of workers under three chemical add-back treatments (n = 16, 20 and 10 for new FP, new FP + pentane and new FP + original FP extracts). d Average TOS of workers exposed to new FP (n = 13), FP of paired workers (n = 10) and FP of two isolated workers (n = 10). e Average TOS of workers exposed to different doses of FP extracts. S5-FP: FP on which a single worker was reared for 5 days (n = 10, 11, 11 or 10 for 0.5, 1, 2 and 4 FP). f Average TOS of workers after exposure to different doses of the pentane extract of worker bees (n = 15, 9 and 10 for new FP, 1 worker equivalent and 10 worker equivalents). g Average TOS of workers provided with new FP (n = 15), FP of drones (n = 12) or gynes (n = 14). h Average TOS of workers exposed to pentane (n = 13) or crude pentane extracts of drones (n = 10) or gynes (n = 8). i Average TOS of workers exposed new FP (n = 9) or FP of honeybee workers (n = 9). j Average TOS of workers exposed to pentane (n = 13) or pentane extracts of honeybee workers (n = 9). All data are presented as mean ± SEM. Data in (b) and (j) were analyzed using two-way ANOVA with Bonferroni correction (two-tailed). Data in (c−h) were analyzed by two-way ANOVA (two-tailed) with Tukey's post-hoc test. In (j), TOS of worker α were compared by unpaired t-test (two-tailed) while those of worker β groups were analyzed by Mann–Whitney U test (two-tailed). Detailed statistical results and adjusted P-values for multiple comparison were shown in Supplementary Data 1. Source data are provided as a Source Data file.
To further test whether contest-induced chemicals39,45 were involved in the ovarian development of α-workers, we determined the 7 day TOS of workers provided with the filter paper on which paired workers or isolated workers had been reared for 5 days (Experiment 2c), following the standard paradigm (Fig. 2a). α-Workers subjected to both treatments had activated ovaries (Fig. 2d, two-way ANOVA with Tukey post-hoc test, F (treatment) = 12.89, df = 2, P < 0.0001), indicating that effective chemicals were spontaneously produced by workers but not induced via social interaction. We conducted parallel detections on the cuticular extracts of isolated workers and α-workers and β-workers. We found that there was no difference in the total amount of cuticular compounds among isolated workers (mean = 0.6927) and α-workers (mean = 0.6206) and β-workers (mean = 0.5830) (Supplementary Fig. 2a, Kruskal-Wallis with Dunn post-hoc analysis, H = 3.020, df = 2, P = 0.2209), indicating that interaction does not affect the total amount of cuticular compounds on the surface of workers. We further examined the effect of the crude pentane extracts of filter paper on which an isolated worker had been reared for 5 days (S5-filter paper) on ovarian development with different levels of dosages including 0.5, 1, 2 and 4 equivalents of one S5-filter paper (Experiment 2d). The result showed that only two and four equivalents had a significantly stimulatory effect (Fig. 2e, two-way ANOVA with Tukey post-hoc test, F (treatment) = 11.08, df = 3, P < 0.0001). In parallel with this result, the addition of ten equivalents of cuticular extract from a singly reared worker, which contained similar quantity of compounds as two equivalents of S5-filter paper or one equivalent of original filter paper (Supplementary Fig. 2b, Kruskal-Wallis with Dunn post-hoc analysis, H = 20.16, df = 2, P < 0.0001), also significantly promoted ovarian development in α-workers (Fig. 2f, two-way ANOVA with Tukey post-hoc test, F (treatment) = 3.195, df = 2, P = 0.0478). Thus, these results indicated that chemicals from the cuticles of bumble bee workers are deposited on the substrates (filter paper in this study), and these chemicals stimulate ovarian development of α-workers when they might reach a threshold dosage.
We further investigated whether the biologically active chemical compounds are produced by other castes of bumble bees and honeybee workers46,47 shown to stimulate the reproduction of bumble bees, following the standard paradigm (Fig. 2a). We found that both drone- or virgin-contacted filter paper, and drone or virgin cuticle extracts, successfully promoted ovarian activation in α-workers (Experiment 2f, Fig. 2g, two-way ANOVA with Tukey post-hoc test, F (treatment) = 7.636, df = 2, P = 0.0010; Experiment 2g, Fig. 2h, two-way ANOVA with Tukey post-hoc test, F (treatment) = 6.136, df = 2, P = 0.0039). Interestingly, filter papers on which honeybee workers had been reared or cuticle extracts from honeybee workers had similar stimulatory effects on the ovaries of α-workers (Experiment 2h, Fig. 2i, two-way ANOVA with Bonferroni correction, F (treatment) = 3.436, df = 1, P = 0.0730; Experiment 2i, unpaired t-test for worker α, t = 5.237, df = 20, P < 0.0001, U test for worker β, U = 30, df = 19, P = 0.0955). These results implied that the stimulatory compounds should be ubiquitous across castes and at least some other bee species.
Given the ubiquitous presence of queens within natural colonies, we explored the interaction between queen inhibitory signals and worker stimulatory compounds. Workers were exposed to three distinct conditions: new filter paper, founder queen-contacted filter paper, and filter paper contacted by paired workers. The results revealed that founder queen-contacted filter paper failed to activate ovarian development in workers (Supplementary Fig. 3a, two-way ANOVA with Tukey post-hoc test, F (treatment) = 5.668, df = 2, P = 0.0049). We then designed two complementary experiments to directly examine the interaction between queen and worker compounds. Results showed that even when supplemented with worker-derived extracts known to stimulate reproduction, queen-contacted filter paper exhibited its inhibitory effect on worker ovarian development (Supplementary Fig. 3b, two-way ANOVA with Tukey post-hoc test, F (treatment) = 1.525, df = 2, P = 0.2271). Similarly, the addition of queen-derived compounds to worker-contacted filter paper effectively prevented worker reproductive activation (Supplementary Fig. 3c, two-way ANOVA with Tukey post-hoc test, F (treatment) = 6.997, df = 2, P = 0.0018). Collectively, these findings demonstrate that in bumble bee colonies where both pheromonal signals coexist, the inhibitory queen signals dominate the interaction, effectively overriding the stimulatory effects of worker pheromones.
The priming effect of chemical cues depends on the synergism of dominance effect, but not on their timing sequence
To further explore the coordinated action of facilitating chemical cues with dominance effect in the dyadic assays, we firstly quantified the amounts of chemical compounds deposited on the filter papers at day 1, 3, 5, 7 after pairing newly-eclosed workers. The amounts of compounds deposited on the filter papers increased dramatically from day 1 to day 5 and reached a peak on day 7 (0.38 mg/cm2 on day 5, i.e. 2.4 times higher than day 3, Fig. 3a, Kruskal-Wallis with Dunn post-hoc analysis, H = 18.25, df = 2, P = 0.0004). Then, to test whether chemical compounds alone can lead to ovarian development in α-workers, we individually reared newly eclosed workers on filter papers previously exposed to bees for 5 days and on new filter papers, and then measured TOS after 7 days. Filter papers from cages with pair of workers failed to promote ovarian activation in individually isolated workers (Experiment 3a and Fig. 3b). Furthermore, we tested if the perception timing of chemical compounds, particularly at time points associated with dominance contests, has an effect on the ovarian development of α-workers (Experiment 2c). Interestingly, ovaries of α-workers were developed when exposed to original filter paper extracts for either the first 5 days or the last 2 days after pairing (Fig. 3c, two-way ANOVA with Tukey post-hoc test, F (treatment) = 9.807, df = 2, P = 0.0002). This result suggested that α-workers activate their ovaries irrespective of whether they contact critical chemical cues, relative to the time dominance interactions.

a Amount of compounds deposited on filter paper after rearing pairs of workers for 7 days (n = 6, 6, 8, and 6 for Days 1, 3, 5, and 7, respectively). b Average TOS for individually isolated workers housed in cages provided with a new filter paper (n = 29) or a filter paper from a cage housing pair of workers (n = 28) in Experiment 3a (left panel). c Average TOS of workers reared in pairs in cages with filter papers subjected to chemical additions at different time points in Experiment 3b (left panel): no exposure (n = 13), early exposure (n = 12), and late exposure (n = 10). Yellow or white indicates original filter papers with heavy chemical deposition or new filter papers with no chemical deposition. All data are shown as mean ± SEM. Data in (a) was analyzed using Kruskal-Wallis test (two-tailed) with Dunn’s post-hoc analysis. Data in (b) was analyzed by Mann-Whitney U test (two-tailed). Data in (c) was analyzed by two-way ANOVA with Tukey post-hoc test (two-tailed) using log10-transformed data. Detailed statistical test results and adjusted P-values for multiple comparison were shown in Supplementary Data 1. Source data are provided as a Source Data file.
Two cuticular hydrocarbons, Z9-C29 and Z9-C31, promote ovarian development in α-workers
Using GC-MS/MS, we analyzed cuticle extracts from bumble bee workers, gynes, drones and honeybee workers, and found substantial overlap in the composition of cuticular chemicals across all four groups, primarily consisting of alkanes and alkenes with carbon chain lengths ranging from 21−31 (Supplementary Fig. 4). To identify functional component(s) in the complex mixture of cuticular compounds, we utilized column chromatography to fractionate the crude extracts, and evaluated the effects of different fractions following the standard assay paradigm (Fig. 4a and Fig. 2a). Initially, we fractionated the cuticular extracts into non-polar hydrocarbons and polar compounds, using silica gel column chromatography. The cuticular hydrocarbon (CHC) fraction significantly promoted ovarian development of α-workers (Experiment 4a and Fig. 4b, two-way ANOVA with Tukey post-hoc test, F (treatment) = 9.835, df = 2, P = 0.0002), whereas the polar fraction did not. We further fractionated the CHCs into alkanes and alkenes ranged from 21−31 carbons using silver nitrate-silica gel column chromatography (Fig. 4c). The alkenes, rather than the alkanes promoted ovarian development in α-workers (Experiment 4b and Fig. 4d, two-way ANOVA with Tukey post-hoc test, F (treatment) = 21.79, df = 2, P < 0.0001). In addition, to exclude possible dosage effect of alkanes, we exposed workers to blends of alkanes from 23 to 29 carbon with different levels of dosages including 1 and 10 equivalents of one original filter paper. No significant effect on ovarian development was found for the two dosage treatments (Experiment 4Sa and Supplementary Fig. 5a, Kruskal-Wallis with Dunn post-hoc analysis, H (worker α) = 13.67, df (worker α) = 3, P (worker α) = 0.0034, H (worker β) = 13.02, df (worker β) = 3, P (worker β) = 0.0046). These results suggest that alkenes in the cuticular extracts of workers are crucial for ovarian development of α-workers. The positions of the double bonds in the alkenes were further determined by analyzing their corresponding dimethyl disulfide adducts (Supplementary Fig. 5b). We identified 17 alkene isomers in bumble bee workers, with Z9-C29 consistently being the most abundant, followed by Z9-C31, and these were 7 to 10 times more abundant than the other compounds (Supplementary Fig. 5c). Given the similar stimulatory effect of cuticle extracts of both bumble bee workers and honeybee workers (Fig. 2j), we deduced that the candidate functional alkenes might be shared by bumble bee workers and honey bee workers. On the basis of the reported chemical profiles of honey bee workers48, we selected eight alkenes shared with bumble bee workers as candidate compounds, including Z9-C23 ((Z)-9-tricosene), Z9-C25 ((Z)-9-pentacosene), Z7-C27 ((Z)-7-heptacosene), Z9-C27 ((Z)-9-heptacosene), Z7-C29 ((Z)-7-nonacosene), Z9-C29 ((Z)-9-nonacosene), Z9-C31 ((Z)-9-hentriacontene), and Z11-C31 ((Z)-11-hentriacontene) (Fig. 4e).

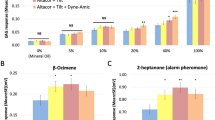
a Schematic diagram of separation of CHCs and non-CHCs extracted from filter paper using column chromatography. b Average TOS of workers exposed to non-CHCs (n = 10), CHCs (n = 12) and pentane (n = 12). c Representative traces of GC-MS/MS of alkanes and alkenes in the filter paper extracts of single workers separated using AgNO₃-silica gel chromatography. d Average TOS of workers exposed to alkanes (n = 10) and alkenes (n = 13) and pentane (n = 12). e Venn diagram of alkenes in cuticular extracts of honeybee and bumble bee workers. f GC-EAD responses of bumble bee workers to eight alkenes in two synthetic blends (n = 17 for both blends). g Average TOS of α-workers exposed to a blend of eight synthetic alkenes (n = 12) and that of three synthetic alkenes (n = 13) and pentane (n = 20). h Average TOS of workers exposed to: Z9-C23 (n = 8, pentane: n = 7); Z7- C27 (n = 15, pentane: n = 13), Z7-C29 (n = 10, pentane: n = 9), Z9-C29 (n = 13, pentane: n = 11), Z9- C31 (n = 12, pentane: n = 12). i Average TOS of workers exposed to a blend of Z9-C29 and Z9- C31 at three different doses (n = 25, 10, 7, 13 for pentane, 0.026 mg/cm3, 0.051 mg/cm3 and 0.103 mg/cm3, respectively). Data are shown as mean ± SEM. Data in (b−g) were analyzed by two-way ANOVA with Tukey post-hoc test (two-tailed). Data in (f) was analyzed by Friedman test with Dunn’s post-hoc analysis (two-tailed). Data in (h) was analyzed using unpaired t-test (two-tailed). In (i), TOS of worker α was compared by one-way ANOVA with Tukey post-hoc test (two-tailed) while TOS of worker β were analyzed by Kruskal-Wallis (two-tailed) with Dunn’s post-hoc analysis. Detailed statistical results and adjusted P-values for multiple comparison were shown in Supplementary Data 1. Source data are provided as a Source Data file.
We used GC-EAD to measure the antennal responses to candidate alkenes. Because of the trivial differences in retention time among alkene isomers during gas chromatography, we separately tested the electrophysiological activity of the blend of five synthesized Z-9 alkenes and that of the three other synthesized alkenes. All eight alkenes consistently elicited responses from worker antennae (Fig. 4f, Friedman test with Dunn post -hoc test, FM (right panel) = 39.95, df (right panel) = 5, P (right panel) < 0.0001, FM (left panel) = 34.98, df (left panel) = 3, P (left panel) < 0.0001). Specifically, worker antennae were more sensitive to Z9-C23, Z9-C29, Z9-C31, Z7-C27, and Z7-C29 than to Z9-C25, Z9-C27, and Z11-C31 (Fig. 4f). We evaluated the effect of these alkenes by exposing workers to mixed or single compounds following the standard paradigm. The full blend of eight compounds, but not the blend of the three compounds with lower electrophysiological activity promoted ovarian development in α-workers (Experiment 4c and Fig. 4g, two-way ANOVA with Tukey post-hoc test, F (treatment) = 15.30, df = 2, P < 0.0001). This result suggested that key effective component(s) should be among the five alkenes with higher electrophysiological activity. The bioassays conducted to evaluate the individual effects of the five alkenes revealed that Z9-C29 and Z9-C31 specifically promoted the ovarian development of α-workers (Experiment 4d and Fig. 4h, unpaired t-test for Z9-C29, t = 2.840, df = 22, P = 0.0095, unpaired t-test for Z9-C31, t = 2.387, df = 22, P = 0.0260). Further, we performed a dose-response experiment to assess the stimulatory effect of the blend of Z9-C29 and Z9-C31 on ovarian development in α-workers. Our results revealed that the stimulation followed a dose-threshold pattern, with a minimum effective dose of 0.051 mg/cm² (Experiment 4e and Fig. 4i, one-way ANOVA with Tukey post-hoc test for worker α, F = 6.241, df = 3, P = 0.0011, Kruskal–Wallis with Dunn post-hoc analysis for worker β, H = 6.527, df = 3, P = 0.0886).
Z9-C29 and Z9-C31 display a colony-context-dependent stimulatory effect on worker reproductive potential under queenright condition
To investigate whether Z9-C29 and Z9-C31 have an effect on worker reproduction in bumble bee colonies, we first established a paradigm to assess reproductive potential of workers. This procedure started by introducing newly emerged workers for 1 day into queenright colonies with 5-or 60-workers, subsequently pairing workers for 4 days and finally separating the pair for 2 additional days (Fig. 5a). The measurement of TOS at day 7 revealed that 1-day exposure to large colonies led to an increase in reproductive potential in α-workers compared to those exposed to small colonies (Fig. 5b, unpaired t-test for worker α, t = 2.840, df = 22, P = 0.0095, U test for worker β, U = 39, n = 24, P = 0.0629). We further compared the amount of the two alkenes deposited in wax, which serves as the major substrate in colonies of both sizes (Fig. 5c). The quantitative analysis revealed significantly higher amounts of Z9-C31 and Z9-C29 in larger queenright colonies (mean = 0.001957) compared to smaller colonies (mean = 0.001225), with the concentrations in larger colonies being ~1.6-fold greater than those measured in smaller colonies (Fig. 5d, unpaired t-test, t = 3.034, df = 40, P = 0.0042). Exposure to small colonies sprayed with pentane extracts of wax from large colonies significantly enhanced reproductive potential in α-workers, compared with those sprayed with pentane (Fig. 5e). In addition, the effect of wax extracts was dose-dependent; extracts of 10 g and 20 g wax led to 2.16-fold and 2.53-fold increases in TOS respectively, whereas extracts of 5 g wax only caused a 1.28-fold increase in TOS (Fig. 5f, two-way ANOVA with Tukey post-hoc test, F (treatment) = 6.763, df = 3, P = 0.0003).

a Schematic diagram of the colony experiment. b Average TOS of workers introduced to a small (n = 13) or a large colony (n = 11) for 24 h post eclosion. c Schematic diagram of quantification of CHCs in wax from colonies of different sizes. d Relative amount of Z9-C29 and Z9-C31 in wax of small (10−15 workers: n = 20) and large (60−80 workers: n = 22) queenright colonies. e Schematic diagram of chemical additions to the colony. f Average TOS of workers exposed to pentane extracts of colony wax at three different doses (n = 25, 8, 16 and 14 for pentane, 5g, 10g and 20g, respectively). g Average TOS of worker exposed to a blend of Z9-C29 and Z9-C31 in Experiment 5c (left panel). Gray: three colonies with pentane addition, n = 18 paired workers; Red: three colonies with Z9-C29 and Z9-C31 addition, n = 23 paired workers. Data are shown as mean '±SEM. Bars labelled with different letters indicate statistically significant differences across different groups. In (b), average TOS of worker α were compared by unpaired t-test (two-tailed) while that of worker β groups were analyzed by Mann–Whitney U test (two-tailed). Data in (d) was analyzed using unpaired t-test (two-tailed). Data in (f) was analyzed by two-way ANOVA with Tukey post-hoc test (two-tailed) using log10-transformed data. Data in (g) was determined by linear mixed-effects models with treatment as a fixed effect and colony identity as a random effect. Detailed statistical test results and adjusted P-values for multiple comparison were shown in Supplementary Data 1. Source data are provided as a Source Data file.
Based on these results, we hypothesized that higher quantities of Z9-C29 and Z9-C31 in the wax led to higher reproductive potential in large colonies. To test the stimulatory role of Z9-C29 and Z9-C31, we introduced workers to small colonies sprayed with the two alkenes for 24 h and assessed their ovarian development following the abovementioned paradigm (Fig. 5a). The addition of Z9-C29 and Z9-C31 significantly enhanced ovarian development in the α-workers, resulting in TOS two times higher than those in small colonies sprayed with pentane solvents (Fig. 5g, Linear Mixed-Effects Model, t (worker α) = 3.413, P (worker α) = 0.0006427; t (worker β) = 0.941, P (worker β) = 0.3468). Taken together, the amounts of Z9-C29 and Z9-C31 which are correlated with colony population, increase worker reproductive potential in queenright colonies in a dose dependent manner.
Discussion
We identified two cuticular alkenes, Z9-C29 and Z9-C31, which function as primer pheromones promoting ovarian development in bumble bee workers under queenless conditions. The amounts of these two alkenes are correlated with colony population size and their dose regulated worker reproductive potential in a quorum-sensing manner in queenright B. terrestris colony (Fig. 6). This finding adds a new mechanism to the complex regulation of worker reproduction in insect societies.

In small under queenright condition colonies with low amounts of these two chemicals, both the ovaries of α- and β- workers fail to activate. As these chemicals accumulate beyond a dose threshold as colony population size increases, the ovaries of α-workers are activated.
To account for potential confounds including pheromone signaling and behavioral dynamics, we employed pairwise dyadic assays using newly emerged workers under queenless conditions. To our knowledge, this approach revealed a previously underexplored role of chemical signals in regulating bumblebee worker fertility. Whereas many studies have emphasized inhibitory pheromonal effects on worker fertility16,17,20, the current study revealed the existence of fertility-stimulating social cues in social insects. Exposure to chemical cues alone was insufficient to induce ovarian activation, suggesting that while aggression primarily acts as an arousal stimulus at the level of bipartite interactions40,49, pheromones also play a key role in triggering ovarian activation in workers. Our findings are consistent with multimodal reproductive regulation observed in territorial mammals and fish50,51, implying the universality of the intricately fine-tuned modulation of reproduction in social animals through the integration of pheromones and behavior.
Our findings suggested that Z9-C29 and Z9-C31 serve as primer pheromones that stimulate worker reproduction in a quorum-sensing manner under queenright conditions. One significant aspect of these pheromones is their widespread distribution, as they were found in bee cuticular lipids, wax, and the colony environment, where they were likely deposited (actively or passively) by isolated bumble bee workers, gynes, and drones. In addition, our research further revealed that these two crucial components can serve as the indicatives of colony size, as evidenced by the difference in the total amount of Z9-C29 and Z9-C31 between the wax of small and large colonies. And, the two alkenes were found to be effective only when their amount exceeded a certain concentration threshold (in the dyadic assay: Z9-C29, 37 μg; Z9-C31, 21 μg; in queenright colony conditions: Z9-C29, 16.5 mg; Z9-C31, 22 mg). Notably, these primer pheromones exerted comparable stimulatory effects on reproduction at both intermediate and high amounts, demonstrating a distinct all-or-none response pattern. This threshold-dependent binary activation contrasts with the dose-dependent linear effects or habituation typically observed in insect pheromone signaling52,53, where physiological responses usually scale with concentration or are suppressed at high doses. Based on the three key characteristics of these two pheromones including self-release, high ubiquity, and activation pattern, Z9-C29 and Z9-C31 could function as quorum-sensing signals in bumble bees, facilitating the activation of worker reproduction. Quorum sensing is recognized as a self-organizing behavior used by social insects to mediate decisions such as nest site selection or nest emigration without the need for central control54,55,56. This mechanism has been extensively studied in bacteria, where gene expression is triggered in response to chemical signals secreted by individual bacteria to assess population density57,58. Similarly with that of other social species, the accumulation of chemical cues released by each individual worker guides decision-making for reproduction activation at the group level59,60, although they elicit physiological changes rather than immediate behavioral responses61.
Quorum sensing likely functions as an adaptive mechanism regulating worker reproduction according to colony developmental stage, with worker population density serving as a critical signal for timing reproductive decisions. Studies demonstrate that increased worker numbers trigger earlier transitions to the competition phase32,62. This colony-size dependent regulation confers significant fitness benefits. In small colonies, lower reproductive potential among workers helps maintain colony productivity regardless of queen presence. Workers benefit from this reproductive restraint as maximizing gyne production is the optimal strategy to enhance their inclusive fitness in the early stage of colony development when colony mortality is high. Conversely, in larger, established colonies, workers can produce drones without compromising gyne production once the colony reaches maximum workforce capacity. This timing mechanism ensures worker reproduction occurs when it minimally impacts colony fitness, suggesting quorum sensing optimizes the balance between individual reproductive interests and colony-level productivity across developmental stages.
Our results show that the synergy between pheromones and dominance contest is independent of the sequence in which they occur. By manipulating wax and wax-borne primer pheromones at the colony level, we observed that the detection of primer pheromones deposited into wax of queenright colonies could influence the reproductive potential of workers during future dominance contests under queenless conditions. Exposure to alkene pheromones equivalent to those in wax from larger colonies for 24 h led to ovarian development in workers following dyadic interactions. This suggested the rapid effect of these alkene pheromones in workers within a day, even under the inhibitory influence of young queens in Pre- Competition Phase (Pre-CPh) colonies. The change of reproductive potential in response to alkenes and their associated colony status may elucidate the earlier onset of the competition phase when additional workers are introduced into Pre -CPh colonies62. The flexibility in the order of sensory input enabled rapid ovarian development when both sufficient primer pheromones and tactile cues from contests were experienced. It is likely that workers in Pre -CPh colonies integrate sources of both stimulatory and inhibitory cues that eventually determine their reproductive state. Despite the unobservable effect of pheromones on ovaries in the absence of contests, early pheromonal stimulation might be stored and affect reproduction at the molecular level, especially including epigenetic regulation of sensory neuroplasticity through histone acetylation or DNA methylation63,64.
This study identified two alkenes as key active components of primer pheromones increasing reproductive potential in bumble bee workers. It is notable that these cuticular hydrocarbons (CHCs) had also been detected in cuticular extracts of honeybees and ants in previous research65,66. However, in the jewelry wasp, Nasonia vitripennis, Z9-C31 is involved in in sexual communication by preventing male-male courtship67. In ants and honeybees, the levels of these alkenes are associated with fertility, with queens or dominant workers displaying higher concentrations of these compounds66,68. This finding contrasts with our results in B. terrestris, where the quantity of alkenes is not correlated with fecundity. Interestingly, in ants, the compound Z9-C29 does not stimulate worker reproduction but instead functions as a dominance signal, inducing submissive behaviors among workers69. These differences in the correlation between alkenes and fertility, as well as their distinct roles in reproductive behaviors across taxa, suggest that the function of alkenes as fertility stimulants may be unique to bumblebees. This specialized role could have evolved in response to the high propensity for worker reproduction in bumblebee colonies. Given these complexities, further investigation into the underlying factors—beyond these two alkenes—of conspecific-induced reproductive stimulation discovered in other social Hymenoptera is warranted70,71,72.
While subsets of CHCs have been recognized as semiochemicals with diverse effects, such as the maintenance of water balance and the encode of specific information about the identity of species, sex, or nestmates48,73,74, these distinct roles of CHCs underscore the significance of individual-specific cues that enable reliable identity differentiation among conspecifics. In addition to these established functions, our study identified alkenes Z9-C29 and Z9-C31 that operate independently of individual identity and serve as ' cues of common '—chemical signals ubiquitous across colony members including workers, gynes, and drones. These compounds are particularly notable for their abundance, occurring at concentrations 7−10 times higher than other isomers, suggesting their potential significance in colony-wide communication systems. The mechanism underlying the stimulatory effect of these two alkenes remained an intriguing question. Odorant receptors (ORs) in the antennae appear to be responsible for the peripheral sensing of alkenes, particularly given the expansion of OR gene families in social Hymenoptera, which is likely associated with the complex chemical communication demands of social life. In support of this, in ants, HsOr263—a member of the expanded nine-exon OR family—has been shown to be tuned to candidate queen pheromone hydrocarbons75. Regarding physiological pathways, juvenile hormone (JH) plays a key role in the regulation of reproduction in B. terrestris by enhancing the transcription of metabolism-related genes and vitellogenin in both the fat body and hemolymph76,77. It is therefore plausible that the detection of alkenes may stimulate JH production in workers. However, whether this occurs and the underlying mechanisms remain to be investigated.
Collectively, this study demonstrates that two alkenes function as sexual maturation-accelerating pheromones with an elegant quorum-sensing mechanism through which workers adjust their reproductive strategies in a self-organized manner. By signaling the presence of conspecifics, the two alkenes scale colony size and enhance worker reproductive potential once their quantity exceeds a threshold. Therefore, worker reproduction in bumble bees is controlled by an elaborate orchestration of multi-source cues associated with social context.
Methods
Collection and rearing of bees
The laboratory B. terrestris colonies were reared in plastic nestboxes (22 cm x 23 cm and 16 cm high), which were placed on shelves in a constant darkness room with a temperature of 28 °C and relative humidity of 50−60%. The colonies were supplied with 50% sugar syrup ad libitum and fresh pollen mixed with sugar syrup. Honeybee hives were purchased from Beijing Nongzhiyi apiary. Callow bumble bee workers emerging from pupae were collected from young, queenright, pre-competition colonies with about 30 workers. Bumble bee drones and gynes were collected upon eclosion from late-stage colonies with about 100 workers. Honeybee adult workers of unknown age were collected from bee hives after treatment with CO2.
The collected bees were reared randomly in pairs or individually in Petri dishes (radius = 4.5 cm, height = 2.5 cm), with paired workers having similar body mass (0.24 ± 0.02 g) and all extremely large or small individuals being excluded from the study. A filter paper was attached to the bottom of the Petri dishes. Pollen and syrup were supplied after workers were introduced into the containers and were replaced every 3 days. The Petri dishes were placed in an incubator (dark, 28°C; 50 ± 10% RH).
Assessment of the states of ovarian development
Workers were dissected 7 days post eclosion unless otherwise mentioned. The workers were frozen in liquid nitrogen and stored at -20 °C until dissection. They were fixed on a wax-filled dissecting plate under a stereomicroscope (Leica DFC450, Leica Microsystems Ltd., Wetzlar, Germany) to assess the ovarian development of focal workers. The ventral plates (sternites) of each worker's gaster were dissected away to expose the ovaries. The ovaries were gently removed into a drop of double distilled water on a microscope slide. The length of the terminal oocyte was measured using the ruler tool of LAS Macro Editor (Leica Microsystems Ltd.) and the mean value of the all terminal oocytes (at least two) was used as the index of ovarian development.
Chemical compounds
Z9-C23 (Z-9-tricosene, CAS: 27519-02-4) and Z9-C25 (Z-9-pentacosene, CAS: 51865-00-0) were purchased from Merck (Darmstadt, Germany). The following compounds were custom-synthesized by Prof. Jocelyn G. Millar: Z7-C27 (Z-7-heptacosene, CAS: 63623-50-7), Z9-C27 (Z-9-heptacosene, CAS: 36258-12-5), Z7-C29 (Z-7-nonacosene, CAS: 63623-51-8), Z11-C31 (Z-11-hentriacontene, CAS: 66648-68-8), Z9-C29 (Z-9-Nonacosene, CAS: 36258-10-3), Z9-C31 (Z-9-hentriacontene, CAS: 56987-72-5).
Experiment 1: The effect of contact cues on worker reproduction in dyadic assay
Analysis of aggression dynamics
Ten pairs of workers were constantly recorded by an infrared camera for 7 days after eclosion (Sony FDR-150, Japan). The videos were analyzed using Observer XT 11 (Noldus Information Technology Co., Ltd., Wageningen, Netherlands). Aggressive events including pushing, dragging, clamping, buzzing, grappling and butting were identified and coded. The aggression index was defined as the average frequency of total aggressive events per pair of bees per hour.
Reproductive bioassays
In Experiment 1a, we tested the effect of group size on ovarian development of workers, newly emerged workers from thirty colonies were divided into queen-less groups of different sizes: single workers, dyads (2 workers), trios (3 workers), groups of 10 workers, and large groups of 30 workers. After maintaining these five distinct group sizes for 7 days, all workers were dissected for ovarian examination. We used the average of terminal oocytes size of all workers within each group as the index of ovarian development. In Experiment 1b, to examine the dynamics of reproduction in a dyadic assay, 70 callow workers from five colonies were randomly paired, and a subset of workers were sacrificed every 24 h to determine the size of their terminal oocytes (TOS). In Experiment 1c, to assess the role of contact social cues in reproduction, newly eclosed workers were paired for 5 days, then transferred either individually to new cage with new Petri dishes and filter paper for 2 days, or continuously reared in their original cage. In Experiment 1d, to examine the role of context in the reproduction of α-workers, workers paired for 5 days were individually transferred into new Petri dishes for 2 days, with either new filter paper or original filter paper (prepared as a living substrate in the pairwise rearing of workers for 5 days). In Experiment 1e, to examine the role of β-workers in the reproduction of α-workers, workers paired for 5 days were transferred to new cages for two additional days under isolated or paired conditions. Additionally, to examine the effect of acoustic signals or volatiles or sight from β-workers on α-workers’ reproduction, workers paired for 5 days were transferred to new cages for 2 days under paired or double-mesh-paired conditions (with a 1 cm double mesh separation to prevent direct contact but allow volatiles to pass through). In Experiment 1f, newly eclosed worker bees were divided into two treatment groups: (1) newly eclosed workers were paired for 7 days. (2) newly eclosed workers were paired for 5 days, then transferred either individually to old cage for 2 days. The ovaries of all workers were dissected.
Experiment 2: The effect of chemical cues from diverse sources on worker reproduction in dyadic assays
Chemical extraction of filter paper and bee cuticle
Pieces of filter paper used for rearing bees for 5 days were shredded and soaked in a 50 ml centrifuge tube with 20 ml of pentane (Innochem, Beijing) for 30 min (min). After being vortex twice, tubes were kept at room temperature for 30 min to allow sedimentation of insoluble material. The supernatant was pipetted into a 40 ml glass sample vial (Agilent, USA). The filter paper extracts used for both bioassay and GC-MS analysis were prepared by concentrating the samples to a final concentration of one filter paper equivalent per milliliter using high-purity nitrogen gas. Six groups of filter paper were collected: (1) filter paper used to rear paired workers for 5 days. (2) filter paper used to rear single workers for 5 days. (3) filter paper used to rear two drones for 5 days. (4) filter paper used to rear two virgins for 5 days. (5) filter paper used to rear founder for a week. (6) filter paper used to rear 20 honeybee workers for 2 weeks. For each sample collected, we set up a blank control group to exclude contaminants.
All bees were killed by immersion in liquid nitrogen and held frozen at −20 °C until extraction. When handling each sample, we include chromatographic blank controls to rule out potential contaminants. Cuticular extracts were obtained by extracting an individual whole body for 2 min in pentane (worker = 2.0 ml; drone & virgin = 4.0 ml). The supernatant was pipetted into a 4 ml glass sample vial (Agilent, USA) and concentrated to 1 ml using high-purity nitrogen gas. Seven groups of cuticle extracts of bees were collected: (1) isolated workers reared for 1 day and 7 day. (2) paired workers reared for 7 days are classified as α-workers (with developed ovaries) or β-workers (with undeveloped ovaries) based on their ovarian development status. (3) drones reared for 5 days. (4) virgin reared for 5 days. (5) honeybee workers.
All sample extractions were filtered using Pasteur pipettes with a pad of diatomaceous earth. Dodecane (50 μl, 100 ng/μl) was added as an internal standard to all samples to quantify the absolute amounts of compounds extracted from filter paper, cuticle surfaces and wax. The filtration method and the quantitative method with C12 as the internal standard for all chemical samples in this paper were the same as above unless mentioned otherwise.
GC-MS/MS of crude extracts from filter paper and cuticle
Two μl of each extract was injected splitless into an HP 7890B Series II gas chromatograph coupled to a 7000C MSD, equipped with an HP-5 capillary column (30 m x 0.25 mm inner diameter, Agilent, Santa Clara CA, USA), using hydrogen as the carrier gas (constant flow, 2.0 ml/min). The chromatograph operated at 50 °C for 1 min, following which the split valve was opened and the oven temperature was programmed as follows: (1) 3 °C/min to 210 °C with a 1 min hold; (2) 3 °C/min to 286 °C with a 1 min hold; (3) 20 °C/min to 310 °C with a 15 min hold. The MS was operated in (EI) mode with a 70 eV ionization energy. A PC-based interface and software package (MassHunter qualitative analysis version B.07, Agilent) was used for data storage and processing. Compound structures were tentatively identified by comparing their mass spectra with reference spectra from the NIST02 library (Scientific Instrument Services, Inc., Ringoes, NJ, USA) and confirmed by matches of GC retention times and mass spectra with those of authentic standard samples (primary method).
Reproductive bioassays
Workers were paired from the eclosion for 5 days, and were individually transferred to filter paper in new Petri dishes with different chemical additives for 2 days. The filter paper with chemical additives was dried for 30 min to allow the solvent to evaporate, then bees were introduced. This standard paradigm was applied in subsequent reproductive bioassays unless mentioned otherwise.
In Experiment 2a, we tested the role of chemical cues from bee-treated filter paper in worker reproduction by exposing workers to either filter paper from Petri dishes used to hold bees, or odorless filter paper (extracted with pentane). In Experiment 2b, we tested the role of these chemical cues by exposing workers to new filter paper with either nothing, 1 ml pentane, or 1 ml pentane extracts of original filter paper. In Experiment 2c, we examined whether the production of reproductive stimulatory chemicals was elicited by contests, by introducing workers to new filter paper, original filter paper, and filter paper used to rear two isolated workers for 5 days (with an opaque barrier). In Experiments 2d and 2e, workers were introduced to filter paper with extracts of either filter paper or whole bodies in varying dosages (0.5, 1, 2, and 4 equivalents of one filter paper, or 1 and 10 equivalents of body cuticle extracts from single workers). In Experiment 2f, workers were introduced to filter papers used to rear two drones or two virgin queens. In Experiment 2g, workers were introduced to clean filter paper treated with cuticle extracts equivalent to ten drones or ten virgins. In Experiment 2h, workers were introduced to filter paper used to rear honeybee workers (20 honeybee workers reared for 2 weeks). In Experiment 2i, workers were introduced to new Petri dishes with cuticle extracts equivalent to twenty honeybee workers.
In Experiment 2j, 2k, 2l, we tested the interaction between inhibitory effect of queen and facilitating effect of chemical compounds. In experiment 2j, workers were introduced to new filter paper, filter paper of founder queen- contacted for 5 days, filter paper of paired workers contacted for 5 days. In Experiment 2k, workers were exposed to three different treatment: (1) clean filter paper with pentane, (2) filter paper used to rear a founder queen for a week, and (3) founder queen- contacted filter paper with extracts of original filter paper. In Experiment 2l, workers were exposed to three different treatments: (1) clean filter paper with pentane, (2) original filter paper, (3) original filter paper with extracts of founder queen- contacted filter paper.
Experiment 3: The effect of chemical cues on worker reproduction in relation to cues from contests
Chemical extraction of filter paper
To quantify the temporal changes of compounds on filter papers, filter paper used to rear paired workers for 1 day, 3 days, 5 days, or 7 days were extracted using pentane. Each piece of filter paper was immersed completely in a glass Petri dish with 4 ml pentane for 30 min, the supernatant was pipetted into a 4 ml glass sample vial (Agilent, USA) and concentrated to 1 ml using high-purity nitrogen gas.
Filter paper crude extracts used in reproductive bioassays were extracted using the same procedure described in Experiment 1.
GC-MS/MS of crude extracts from filter paper
The chromatograph operated at 60 °C for 1 min, following which the split valve was opened and the temperature was increased as follows: (1) 10 °C/min to 240 °C; (2) 5 °C/min to 320 °C with a 10 min hold. For the details of mass spectrometry, see Experiment 2.
Reproductive bioassays
In Experiment 3a, to test the effects of chemical compounds on single worker reproduction, we introduced single workers into new Petri dishes with either new or original filter paper for 7 days. In Experiment 3b, to test if early perception of adequate compounds activates reproduction in α-workers, we set up three groups: (1) Nothing added: workers paired for 5 days were individually introduced into a new Petri dish for 2 days. (2) Early addition: callow workers paired for 5 days with original filter paper extracts added on the first day, then introduced individually into a new Petri dish for 2 days. (3) Late addition: workers paired for 5 days were introduced individually into a new Petri dish for 2 additional days with original filter paper extracts added.
Experiment 4: The effect of candidate compounds on worker reproduction
Chemical extraction of filter paper
Filter paper crude extracts used in column chromatography were extracted using the same procedure described in Experiment 1.
Silica gel column chromatography
The glassware, including the chromatography column and beakers, was preheated in an oven at 180 °C for 3 h. Subsequently, 12 g of silica gel (100-200 mesh, Beijing Chemical Industry, Beijing) were packed into the chromatography column (radius = 2 cm, length 30 cm). Pentane was added and the column was eluted 2-3 times to remove air bubbles in the silica gel packing. Following this, the filter paper extraction (70 filter paper in 35 ml pentane) was introduced and allowed to elute until the liquid level reached the top of the silica gel bed, at which point 35 ml of pentane was added to ensure thorough washing of the non-polar components from the eluate, glass beakers were used to collecting the non-polar eluate. Once the pentane had completely drained, 40−50 ml of a methanol (InnoChem, Beijing): pentane (95:5) solution was added to the column to ensure thorough elution of the polar components, collection switches back to glass beakers to gather the polar components. This procedure was designed for chromatography using silica gel as the stationary phase to separate compounds based on their polarity.
Silver nitrate silica gel chromatography
Preparation of silver nitrate silica gel78: 40 grams of chromatography-grade silica gel was added a solution of 8 grams of silver nitrate in 100 ml 80% methanol solution (Beijing Chemical Industry, Beijing). After mixing, the mixture was allowed to stand undisturbed for 2 h, shielded from light. The solvent was then evaporated by immersing the foil-covered flask in a 70 °C water bath for ~1 h. The residue was then placed in a vacuum drying oven at 120 °C for 12−15 h to remove residual solvent.
Subsequently, 15 g of silver nitrate silica gel were packed into a chromatography column (radius = 2 cm, length = 30 cm), and the column was rinsed several times with pentane to pack the bed. The hydrocarbon fraction from above was then loaded onto the column, which was eluted with 100 ml pentane to collect the saturated hydrocarbons. The unsaturated hydrocarbons were then eluted with 100 ml 10% ether solution. Double bond positions in alkenes were determined by formation of the corresponding dimethyl disulfide (DMDS) adducts followed by analysis by GC-MS79.
GC-MS/MS
For alkane and alkene compounds, the chromatograph operated at 60 °C for 3 min, following which the split valve was opened and the temperature increased. The temperature program consisted of 2 steps: (1) 20 °C/min to 240°C; (2) 5 °C/min to 320 °C with a 5-min hold. For DMDS compounds, the procedure of GC-MS/MS was the same with the procedure of filter paper crudes in Experiment 2. For the mass spectrometry details, see Experiment 2.
GC-EAD of synthesized compounds
For GC-EAD recordings (n = 15), an antenna was carefully dislodged from the head of one individual worker of bumble bees. The tip of the antenna was cut off. Then, the clipped antenna was suspended between two metal electrodes with the conducting gel. Signals were passed through a high-impedance DC amplifier (UN-06, Syntech, Hilversum, Netherlands) into a signal connection interface box (Auto Spike, IDAC 2/3, Syntech). A PC-based interface and software package (EAD version 2.3, Syntech) was used for data storage and processing. We injected 4 μl of two mixed solutions (mixture 1 contain five compounds: Z9-C23, Z9-C25, Z9-C27, Z9-C29, Z9-C31, and mixture 2 contain 3 compounds: Z7-C27, Z7-C29, Z11-C31, 1000 ng/μl) into a modified Agilent 7890 A gas chromatograph equipped with an HP-5 column (30 m × 0.25 mm × 0.25 µm, Agilent Technologies, Inc.). The end of the column was equipped with a column splitter (OSS-2, SGE, Ringwood, Victoria, Australia) and an additional outlet (temperature controller, TC-02, Syntech), allowing simultaneous flame ionization detection (FID) and electroantennographic detection (EAD). The effluent from the column was split into two streams: one (50%) was directed to the FID, and the other (50%) was directed to the EAD. The temperatures of the injector, detector, and EAD transfer line were set at 300, 320 °C, and 280 °C, respectively. The gas chromatograph oven temperature was held at 100 °C for 1 min, then ramped to 240 °C at a rate of 30 °C/min, increased to 320 °C at a rate of 10 °C/min, and held at 320 °C for 2 min.
Reproductive bioassays
Workers were treated following the standard paradigm in Experiment 2 but in round Teflon containers (r = 1.5 cm, h = 3 cm) instead of Petri dishes. In Experiments 4a and 4b, workers were exposed to non-polar and polar compounds separated through column chromatography (with a surface density equivalent to one piece of original filter paper), and to saturated and unsaturated compounds separated through silver nitrate silica gel chromatography (with a surface density equivalent to one piece of original filter paper), each for 2 days. In Experiment 4c, workers were exposed to (1) full blends of eight candidate compounds, (2) blends of the first subset of compounds (Z9-C23, Z7-C27, Z7-C29, Z9-C29, Z9-C31), or (3) blends of the second subset of compounds (Z9-C25, Z9-C27, Z11-C31). The eight synthesized compound mixtures were as follows, with respective surface densities equivalent to one piece of original filter paper: Z9-C23: 0.026 mg/cm², Z9-C25: 0.015 mg/cm², Z7-C27: 0.0111 mg/cm², Z9-C27: 0.037 mg/cm², Z7-C29: 0.018 mg/cm², Z9-C29: 0.163 mg/cm², Z9-C31: 0.093 mg/cm², Z11-C31: 0.037 mg/cm². In Experiment 4d, candidate compounds were tested individually at surface densities equivalent to ten pieces of original filter paper. In Experiment 4e, workers were exposed to mixture of Z9-C29 and Z9-C31 at different dosage, Z9-C29 and Z9-C31 were mixed at surface densities of 0.026 mg/cm², 0.051 mg/cm², and 0.103 mg/cm², the equivalents of one, two, and four pieces of original filter paper, respectively.
In Experiment 4Sa, workers were treated following the standard paradigm in Experiment 2. Workers were exposed to blends of alkanes from 23−29 carbon with different levels of dosages. The four alkane compound mixtures were as follows, with respective equivalents to one and ten original filter paper: C23: 0.08 mg and 0.8 mg, C25: 0.08 mg and 0.8 mg, C27: 0.15 mg and 1.5 mg, C29: 0.04 mg and 0.4 mg.
Experiment 5: The effect of Z9-C29 and Z9-C31 on worker reproductive potential in colony experiments
Chemical extraction of wax
Wax samples from four B. terrestris colonies (n = 10 wax samples from each colony at least) at two stages of colony development (colony size was given in the number of workers present in the colony). Small colonies (n = 2): a colony size of 10–15 workers; Large colonies (n = 2): a colony size of 60−80 workers. Wax pieces were weighed (0.1 g - 0.2 g) and transferred to glass vials (4 ml) and extracted in 2 ml pentane at room temperature for 30 min, while being vortexed twice. The resulting solutions were held at minus 20 °C for at least 1 h to allow sedimentation of insoluble, nonvolatile wax components. The supernatant was aspirated and concentrated to 1 ml with N2. Double bond positions in alkene compounds were determined by GC-MS analysis of the corresponding dimethyl disulfide (DMDS) adducts81 as described above.
We also extracted 5, 10, or 20 g wax samples from large colonies (60−80 workers), in 35−50 ml pentane at room temperature for 30 min, samples were kept frozen (minus 20 °C) overnight. The supernatant was concentrated to 5 ml with N2, transferred to a glass watering can bottle, and stored at minus 20 °C until used in experiments.
GC-MS/MS for wax
For DMDS products of wax extracts, the chromatograph was operated at 60 °C for 3 mi, following which the split valve was opened and the temperature increased. The temperature program consisted of 2 steps: (1) 20 °C/min to 240 °C with a 1 min hold; (2) 5 °C/min to 320 °C with a 15 min hold. For the details of mass spectrometry, see Experiment 2.
Reproductive bioassays
We used a paradigm to test the reproductive potential of workers in queenright colonies. Newly emerged workers were marked with color markers and introduced to queenright colonies for 1 day. Subsequently, marked workers were collected from colonies and pairwise reared in Petri dishes for 4 days and then isolated in new Petri dishes with new filter paper for 2 days.
In Experiment 5a, workers were introduced to either small colonies (with 6 workers) or large colonies (with 50−60 workers) to assess the effect of colony-size-related cues on reproductive potential. In Experiment 5b, we tested the effect of wax from large colonies on worker reproduction. Before introducing workers, we sprayed the pentane extracts of 5 g, 10 g, or 20 g wax from large colonies onto the recipient colonies. The bees in the colony were removed before spraying the wax extracts, and the colony was allowed to dry for 30 min. After the colony dried, the removed worker bees were transferred back, and 4-6 newly marked workers were introduced into the recipient colonies. Each treatment included three colonies. In Experiment 5c, we tested the effect of Z9-C29 and Z9-C31 on worker reproduction in colonies. We sprayed a mixture of 33 mg synthesized Z9-C29 and 44 mg Z9-C31 (equivalent to the amounts of compounds in 20 g wax of large colonies) or pentane into recipient colonies. The workers in the recipient colonies were manipulated as described in Experiment 5b. Four to six newly marked workers were introduced into the recipient colonies. Each treatment included three colonies.
Statistical analysis
Statistical analysis was conducted using GraphPad Prism (9.5.0) unless otherwise mentioned. All data are presented as the mean ± SEM.
For t-test, We evaluated data normality using Shapiro-Wilk tests (α = 0.05) and verified variance homogeneity using F-tests (α = 0.05) to determine appropriate analyses. Normally distributed data with equal variance were analyzed using t-tests for two-group comparisons; otherwise, Mann-Whitney U test was used.
For one-way ANOVA, normality of residuals was assessed using Shapiro-Wilk tests (α = 0.05), and homogeneity of variance was evaluated with Brown-Forsythe tests (α = 0.05). When both assumptions were satisfied, one-way ANOVA with Tukey's post-hoc test was applied; otherwise, the nonparametric Kruskal-Wallis with Dunn’s post-hoc analysis was used.
For two-way ANOVA, the normality assumption was verified by Shapiro-Wilk tests (α = 0.05) on model residuals, with variance homogeneity evaluated via residual plots. Data satisfying both assumptions underwent two-way ANOVA with Bonferroni correction (each factors containing exactly two levels) or with Tukey's post-hoc test (any factors containing three or more levels). When assumptions were violated, data were log10-transformed and re-assessed. Data satisfying parametric assumptions after transformation were analyzed by two-way ANOVA with Bonferroni correction or with Tukey post-hoc test, while persistently non-compliant data were analyzed using one-way ANOVA with Tukey post-hoc test (or Kruskal-Wallis with Dunn’s post-hoc analysis) and unpaired t-test (or Mann–Whitney U test).
For Experiment 5g, linear mixed-effect models in R version 4.2.1 (The R Foundation for Statistical Computing, Vienna, Austria, http://www.r-project.org) were used to examine the effect of the alkenes on reproductive potential in colonies. Chemical treatment was added as a fixed effect and colony identity was added as a random effect.
Additional information on statistics that is relevant to only a certain experiment, is reported in the figure legend and Supplementary Data 1.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The raw mass spectrometry data and other source data generated in this study have been deposited in the Figshare database under https://doi.org/10.6084/m9.figshare.29391479. All other data supporting the findings of this study are available within the article and its Supplementary Information. Source data are provided with this paper.
References
Michener, C. D. The Social Behavior of the Bees. (Harvard University Press, Cambridge, 1974).
Wilson, E. O. The Insect Societies Illustrated edition, Vol. 562 (Harvard University Press, Cambridge, 1971).
Barron, A., Oldroyd, B. & Ratnieks, F. L. W. Worker reproduction in honey-bees (Apis) and the anarchic syndrome: a review. Behav. Ecol. Sociobiol. 50, 199–208 (2001).
Bourke, A. F. G. Worker reproduction in the higher eusocial hymenoptera. Q. Rev. Biol. 63, 3 (1988).
Cruz-Landim, C. Ovarian development in meliponine bees (hymenoptera: Apidae): the effect of queen presence and food on worker ovary development and egg production. Genet. Mol. Biol. 23, 83–88 (2000).
Visscher, P. K. A quantitative study of worker reproduction in honey bee colonies. Behav. Ecol. Sociobiol. 25, 247–254 (1989).
Ratnieks, F. L. W. Reproductive harmony via mutual policing by workers in eusocial hymenoptera. Am. Nat. 132, 217–236 (1988).
Van Honk, C. G. J., Röseler, P. F., Velthuis, H. H. W. & Hoogeveen, J. C. Factors influencing the egg laying of workers in a captive Bombus terrestris colony. Behav. Ecol. Sociobiol. 9, 9–14 (1981).
Trivers, R. L. & Hare, H. Haploidploidy and the evolution of the social insect: the unusual traits of the social insects are uniquely explained by Hamilton’s kinship theory. Science 191, 249–263 (1976).
Wenseleers, T. & Ratnieks, F. L. W. Comparative analysis of worker reproduction and policing in eusocial hymenoptera supports relatedness Theory. Am. Nat. 168, E163–E179 (2006).
Hamilton, W. D. Altruism and related phenomena, mainly in social insects. Annu. Rev. Ecol. Syst. 3, 193–232 (1972).
Hamilton, W. D. The genetical evolution of social behaviour. I. J. Theor. Biol. 7, 1–16 (1964).
Alpedrinha, J., West, S. A. & Gardner, A. Haplodiploidy and the evolution of eusociality: worker reproduction. Am. Nat. 182, 421–438 (2013).
Dampney, J. R., Barron, A. B. & Oldroyd, B. P. Measuring the cost of worker reproduction in honeybees: work tempo in an ‘anarchic’ line. Apidologie 35, 83–88 (2004).
Carmona-Aldana, F., Yong, L. W., Reinberg, D. & Desplan, C. Phenomenon of reproductive plasticity in ants. Curr. Opin. Insect Sci. 63, 101197 (2024).
Ge, J., Ge, Z., Zhu, D. & Wang, X. Pheromonal regulation of the reproductive division of labor in social insects. Front. Cell Dev. Biol. 8, 837 (2020).
Hefetz, A. The critical role of primer pheromones in maintaining insect sociality. Z. Naturforsch. C. Biosci. 74, 221–231 (2019).
Kocher, S. D. & Grozinger, C. M. Cooperation, conflict, and the evolution of queen pheromones. J. Chem. Ecol. 37, 1263–1275 (2011).
Schultner, E., Oettler, J. & Helanterä, H. The role of brood in eusocial hymenoptera. Q. Rev. Biol. 92, 39–78 (2017).
Villalta, I., Abril, S., Cerdá, X. & Boulay, R. Queen control or queen signal in ants: What remains of the controversy 25 years after Keller and Nonacs’ seminal paper? J. Chem. Ecol. 44, 805–817 (2018).
Van Zweden, J. S., Fürst, M. A., Heinze, J. & D’Ettorre, P. Specialization in policing behaviour among workers in the ant Pachycondyla inversa. Proc. R. Soc. B. 274, 1421–1428 (2007).
Liebig, J., Peeters, C. & Hölldobler, B. Worker policing limits the number of reproductives in a ponerine ant. Proc. R. Soc. Lond. B 266, 1865–1870 (1999).
Sasaki, T. et al. A simple behavioral model predicts the emergence of complex animal hierarchies. Am. Nat. 187, 765–775 (2016).
Chole, H., Woodard, S. H. & Bloch, G. Body size variation in bees: regulation, mechanisms, and relationship to social organization. Curr. Opin. Insect Sci. 35, 77–87 (2019).
Hogendoorn, K. & Velthuis, H. H. W. Task allocation and reproductive skew in social mass provisioning carpenter bees in relation to age and size. Insectes soc. 46, 198–207 (1999).
Angulo, E. et al. Allee effects in social species. J. Anim. Ecol. 87, 47–58 (2018).
Brahma, A., Mandal, S. & Gadagkar, R. Emergence of cooperation and division of labor in the primitively eusocial wasp Ropalidia marginata. Proc. Natl Acad. Sci. Usa. 115, 756–761 (2018).
Luque, G. M., Giraud, T. & Courchamp, F. Allee effects in ants. J. Anim. Ecol. 82, 956–965 (2013).
Free, J. B. The effect of social facilitation on the ovarial development of bumble-bee workers. Proc. R. Ent. Soc. Lon. 32, 182–184 (1957).
Alaux, C., Savarit, F., Jaisson, P. & Hefetz, A. Does the queen win it all? Queen-worker conflict over male production in the bumblebee, Bombus terrestris. Naturwissenschaften 91, 400–403 (2004).
Van Oystaeyen, A. et al. Conserved class of queen pheromones stops social insect workers from reproducing. Science 343, 287–290 (2014).
Duchateau, M. J. & Velthuis, H. H. W. Development and reproductive strategies in Bombus Terrestris colonies. Behaviour 107, 186–207 (1988).
Bloch, G. et al. A Juvenile hormone titers, juvenile hormone biosynthesis, ovarian development and social environment in Bombus terrestris. J. Insect Physiol. 46, 47–57 (2000).
Amsalem, E. & Hefetz, A. The Effect of Group Size on the Interplay between Dominance and Reproduction in Bombus terrestris. PLoS ONE 6, e18238 (2011).
Duchateau, M. J. & Velthuis, H. H. W. Ovarian development and egg laying in workers of Bombus terrestris. Entomol. Exp. Appl. 51, 199–213 (1989).
Van Doorn, D. Factors influencing dominance behaviour in queenless bumblebee workers (Bombus terrestris). Physiol. Entomol. 14, 211–221 (1989).
Ge, J. et al. Phased contests allow rapid hierarchy formation in paired bumble bee workers. Anim. Behav. 179, 125–138 (2021).
Bloch, G. & Hefetz, A. Regulation of reproduction by dominant workers in bumblebee (Bombus terrestris) queenright colonies. Behav. Ecol. Sociobiol. 45, 125–135 (1999).
Amsalem, E. & Hefetz, A. The appeasement effect of sterility signaling in dominance contests among Bombus terrestris workers. Behav. Ecol. Sociobiol. 64, 1685–1694 (2010).
Cnaani, J., Schmid-Hempel, R. & Schmidt, J. O. Colony development, larval development and worker reproduction in Bombus impatiens cresson. Insectes Soc. 49, 164–170 (2002).
Cnaani, J., Wong, A. & Thomson, J. D. Effect of group size on ovarian development in bumble bee workers (Hymenoptera: Apidae: Bombus). Entomol. Gen. 29, 305–314 (2007).
Rottler-Hoermann, A.-M., Schulz, S. & Ayasse, M. Nest wax triggers worker reproduction in the bumblebee Bombus terrestris. R. Soc. open sci. 3, 150599 (2016).
Ge, J. et al. Integration of information from multiple sources drives and maintains the division of labor in bumble bee colonies. Curr. Opin. Insect Sci. 60, 101115 (2023).
Amsalem, E., Grozinger, C. M., Padilla, M. & Hefetz, A. The physiological and genomic bases of bumble bee social behaviour. Adv. Insect Physiol. 48, 37–93 (2015).
Sramkova, A. et al. Fertility signals in the bumblebee Bombus terrestris (Hymenoptera: Apidae). Naturwissenschaften 95, 515–522 (2008).
Ptácek, V. Trials of reare bumble bees. Acta Hortic. 288, 144–148 (1991).
Velthuis, H. H. W. The Historical Background Of The Domestication Of The Bumble-bee, Bombus Terrestris. https://www.semanticscholar.org/paper/THE-HISTORICAL-BACKGROUND-OF-THE-DOMESTICATION-OF-Velthuis/5a933c348e4fbebcea88be93a85a5815df935416 (2002).
Vernier, C. L. et al. The cuticular hydrocarbon profiles of honey bee workers develop via a socially-modulated innate process. eLife 8, e41855 (2019).
Amsalem, E., Shamia, D. & Hefetz, A. Aggression or ovarian development as determinants of reproductive dominance in Bombus terrestris: interpretation using a simulation model. Insect Soc. 60, 213–222 (2013).
Fuxjager, M. J., Mast, G., Becker, E. A. & Marler, C. A. The ‘home advantage’ is necessary for a full winner effect and changes in post-encounter testosterone. Horm. Behav. 56, 214–219 (2009).
Fuxjager, M. J. & Marler, C. A. How and why the winner effect forms: influences of contest environment and species differences. Behav. Ecol. 21, 37–45 (2010).
Afify, A. & Potter, C. J. Insect repellents mediate species-specific olfactory behaviours in mosquitoes. Malar. J. 19, 127 (2020).
Liu, W. et al. Social regulation of aggression by pheromonal activation of Or65a olfactory neurons in. Drosoph. Nat. Neurosci. 14, 896–902 (2011).
Franks, N. R., Hooper, J., Webb, C. & Dornhaus, A. Tomb evaders: house-hunting hygiene in ants. Biol. Lett. 1, 190–192 (2005).
Franks, N. R. et al. Not everything that counts can be counted: ants use multiple metrics for a single nest trait. Proc. R. Soc. B. 273, 165–169 (2006).
Seeley, T. D. & Kirk Visscher, P. Group decision making in nest-site selection by honey bees. Apidologie 35, 101–116 (2004).
Lupp, C., Urbanowski, M., Greenberg, E. P. & Ruby, E. G. The Vibrio fischeri quorum-sensing systems ain and lux sequentially induce luminescence gene expression and are important for persistence in the squid host. Mol. Microbiol. 50, 319–331 (2003).
Rutherford, S. T. & Bassler, B. L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2, a012427–a012427 (2012).
Mallon, E. B., Pratt, S. C. & Franks, N. R. Individual and collective decision-making during nest site selection by the ant Leptothorax albipennis. Behav. Ecol. Sociobiol. 50, 352–359 (2001).
Pratt, S. C. Quorum sensing by encounter rates in the ant Temnothorax albipennis. Behav. Ecol. 16, 488–496 (2005).
Funaro, C. F., Böröczky, K., Vargo, E. L. & Schal, C. Identification of a queen and king recognition pheromone in the subterranean termite Reticulitermes flavipes. Proc. Natl Acad. Sci. 115, 3888–3893 (2018).
Princen, S. A., Van Oystaeyen, A., Van Zweden, J. S. & Wenseleers, T. Worker dominance and reproduction in the bumblebee Bombus terrestris: When does it pay to bare one’s mandibles? Anim. Behav. 166, 41–50 (2020).
Haga-Yamanaka, S. et al. Plasticity of gene expression in the nervous system by exposure to environmental odorants that inhibit HDACs. eLife 12, RP86823 (2024).
Shang, A. & Bieszczad, K. M. Epigenetic mechanisms regulate cue memory underlying discriminative behavior. Neurosci. Biobehav. Rev. 141, 104811 (2022).
Falcón, T. et al. Exoskeleton formation in Apis mellifera: cuticular hydrocarbons profiles and expression of desaturase and elongase genes during pupal and adult development. Insect Biochem. Mol. Biol. 50, 68–81 (2014).
Liebig, J. et al. Are variations in cuticular hydrocarbons of queens and workers a reliable signal of fertility in the ant Harpegnathos saltator? Proc. Natl Acad. Sci. Usa. 97, 4124–4131 (2000).
Wang, Y. et al. Silencing doublesex expression triggers three-level pheromonal feminization in nasonia vitripennis males. Proc. Biol. Sci. 289, 20212002 (2022).
Caliari Oliveira, R. et al. The origin and evolution of queen and fertility signals in corbiculate bees. BMC Evol. Biol. 15, 254 (2015).
Smith, A. A., Millar, J. G. & Suarez, A. V. A social insect fertility signal is dependent on chemical context. Biol. Lett. 11, 20140947 (2015).
Nozaki, T. & Matsuura, K. Oocyte resorption in termite queens: seasonal dynamics and controlling factors. J. Insect Physiol. 131, 104242 (2021).
Ozan, M., Helanterä, H., d’Ettorre, P. & Sundström, L. Queen fecundity, worker entourage and cuticular chemistry in the ant Formica fusca. Proc. R. Soc. B. 290, 20230861 (2023).
Sarro, E. et al. An organizing feature of bumble bee life history: worker emergence promotes queen reproduction and survival in young nests. Conserv. Physiol. 9, coab047 (2021).
Jallon, J. M. & Wicker-Thomas, C. Genetic studies on pheromone production in Drosophila. In Insect Pheromone Biochemistry and Molecular Biology (eds. Blomquist, G. & Richard Vogt, R.) 253−281 (Elsevier Academic Press, 2003)
Steitz, I., Paxton, R. J., Schulz, S. & Ayasse, M. Chemical variation among castes, female life stages and populations of the facultative eusocial sweat bee Halictus rubicundus (Hymenoptera: Halictidae). J. Chem. Ecol. 47, 406–419 (2021).
Pask, G. M. et al. Specialized odorant receptors in social insects that detect cuticular hydrocarbon cues and candidate pheromones. Nat. Commun. 8, 297 (2017).
Shpigler, H. et al. Gonadotropic and physiological functions of juvenile hormone in bumblebee (Bombus terrestris) workers. PLoS ONE 9, e100650 (2014).
Shpigler, H. et al. Juvenile hormone functions as a metabolic rate accelerator in bumble bees (Bombus terrestris). Horm. Behav. 136, 105073 (2021).
Ghosh, A., Hoqui, M. & Dutta, J. Separation of polyunsaturated fatty acid esters by argentation column chromatography on silicic acid. J. Chromatogr. A. 69, 207–208 (1972).
Buser, H. R. udolf Arn, Heinrich., Guerin, Patrick. & Rauscher, Stefan. Determination of double bond position in mono-unsaturated acetates by mass spectrometry of dimethyl disulfide adducts. Anal. Chem. 55, 818–822 (1983).
Acknowledgements
We would like to express our utmost gratitude to Professor Le Kang from the Institute of Zoology, Chinese Academy of Sciences, for his tremendous support for this research. We sincerely thank Professor Ping Wen from Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, and Professor Li Chen from College of Life Sciences, HeBei University, for their valuable comments in compound identification and synthesis, and Dr. Jan Buellesbach for comments of manuscripts. We appreciated the assistance of Dr. Xian Zhou from the Institute of Zoology, Chinese Academy of Sciences, in behavioral analysis and Ziying Liu in chemical analysis. This work was supported by the National Natural Science Foundation of China (NSFC number: 32200392, J.G., and 32061143002, X. W.), the Israel Science Foundation (ISF grant number: 3391/20 supporting Israel-China collaboration, G.B.) and Hatch Act fund (number: CA-R*ENT-5181-H, J.M.).
Author information
Authors and Affiliations
Contributions
Z.G., J.G., D.Z. and X.W. designed the research. Z.G. and X.N. conducted the experiments. J.M. synthesized the alkenes for experiments. Z.G. and J.G. analyzed the data. Z.G., J.G. and X.W. wrote the paper. X.W., J.G., Z.G. and G.B. discussed the manuscript. X.W., J.G., Z.G., J.M. and G.B. revised the manuscript. X.W., J.G., D.Z. and G.B. acquired grants.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Alison McAfee and the other anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ge, Z., Ge, J., Zhu, D. et al. The alkenes Z9-C29 and Z9-C31 function as quorum-sensing pheromones stimulating worker reproduction in the bumble bee Bombus terrestris. Nat Commun 16, 10770 (2025). https://doi.org/10.1038/s41467-025-65812-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-65812-x
