
Abstract
Dynamic mechanical cues are crucial for glial neuromodulation and energy metabolism in neural regeneration, yet the mechanisms underlying mechanotransduction and intracellular organelle responses in glia after neurotrauma remain vague. In this study, we develop mechano-bioactive piezoelectric hydrogel bioelectronics (BaTiO3-embedded collagen-1 hydrogel) and investigate mechanotransduction in astrocytes and Schwann cells. Ultrasound-driven piezoelectric hydrogel bioelectronics exerts electrical signals from mechanical stimulation and upregulates PIEZO1 channel in astrocytes and PIEZO2 channel in Schwann cells. This mechanoelectrical conversion increases calcium influx to activate ATP synthase subunit and promote MFN/OPA1 mediated mitochondrial fusion. Consequently, it enhances ATP synthesis by forming an efficient energy network as a central bioenergetic hub to promote glia mediated neural repair. Furthermore, this mechano-bioactive piezoelectric hydrogel bioelectronics exhibits therapeutic efficacy for treating central and peripheral nervous injuries in multiple animal models (mice, rats, Beagle dogs, and Rhesus monkeys), demonstrating its wide adaptivity and significant translational potential. The findings elucidate a multilevel mechanobiological energy transduction (mechanical-electrical-bioenergetic conversion) design in neural repair as a promising clinical treatment mode.
Similar content being viewed by others

Introduction
Nerve injuries present significant medical challenges worldwide due to their high incidence of disability and the inherently limited regenerative capacity of neurons1,2. Accumulating evidence suggests that effective neural regeneration relies heavily on glia, which actively regulate neuronal survival, axon regeneration, and remodeling of the neural microenvironment3,4,5,6. Specifically, astrocytes in the central nervous system (CNS) and Schwann cells in the peripheral nervous system (PNS) are pivotal glial populations to orchestrate neural repair by modulating neuronal activity, energy metabolism, and intercellular communication7,8.
Mechanical stimulation governs a wide range of vital cellular activities in nerve development and regeneration, from macroscopic tissue remodeling to microscopic molecular signaling9,10. For instance, axons are propelled by mechanical tension from the growth cone during neurogenesis11,12. External mechanical stimulation like ultrasonic and electrical signals can modulate neural cell activity and organelle function13,14,15,16,17. It indicates mechanical cues as pivotal contributors of neuromodulation9.
Mechanotransduction involves detecting mechanical stimulation and their subsequent conversion into biochemical signals at cellular and subcellular levels18,19,20. Cells detect mechanical cues from their surroundings via receptor-ligand interactions, cell surface coupling, and mechanically gated ion channels21,22. Rapid transduction of mechanical cues occurs via mechanosensors at the plasma membrane and brings immediate changes in membrane potentials and subsequent intracellular biochemical reactions23,24. Among these mechanosensors, PIEZO channels (PIEZO1 and PIEZO2) emerge as critical mediators of mechanical stress in diverse biological processes such as sensory perception and organ development25,26,27,28,29,30,31. These PIEZO channels are prominently activated and expressed in glia during neurodevelopment. Nevertheless, their role in glia-mediated neural regeneration is underexplored7,32,33.
Generally, mechanical stimulation to cells can be achieved in three ways: (i) applying external cues such as electrical or sonic stimulation; (ii) altering matrix mechanical properties such as substrate stiffness to affect cell interactions; and (iii) imposing mechanical forces such as shear flow21. Mechano-bioactive implants, such as piezoelectric materials, are gaining attention for their ability to transduce external mechanical signals for tissue regeneration19. Integrating piezoelectric nanomaterial-based bioelectronics with advanced tissue engineering techniques provides promising platform for mechanobiology research in glial neuromodulation. While these materials show promise for neural repair, the mechanisms are unclear and likely involve more than just increases in ion flow34,35,36. It remains inadequately understood as to the impact of calcium influx on intracellular signaling transduction and organelle function, including mitochondria, for piezoelectric neural repair.
In this study, we developed mechano-bioactive BaTiO3 hydrogel bioelectronics to investigate glial mechanotransduction and intracellular organelle responses in traumatic central and peripheral nerve injuries (Fig. 1). We used ultrasound to drive piezoelectric materials generating piezoelectric potentials and producing localized microcurrents. The ultrasound-driven piezoelectric bioelectronics exerted electrical signals from external mechanical stimulation and upregulated PIEZO1 channel in astrocytes and PIEZO2 channel in Schwann cells. The mechanoelectrical transduction process increased calcium influx in glia to subsequently activate mitochondrial function and promote ATP synthesis. Furthermore, we validated the efficacy of piezoelectric bioelectronics across diverse animal models (mice, rats, Beagle dogs, and Rhesus monkeys) to evaluate the wide adaptivity and clinical translational potential for neural repair. This study uncovers a novel glial mitochondrial dynamics strategy supported by the mechano-bioactive piezoelectric bioelectronics for developing advanced clinical treatment mode in neural regeneration.

A BaTiO3 piezoelectric hydrogel bioelectronics design and fabrication. The BaTiO3 nanoparticles, synthesized via a solvothermal approach, were optimized for uniform particle size and single-domain structure to enhance piezoelectric properties. Post-synthesis surface modification with hydroxyl (-OH) groups improved the hydrophilicity of BaTiO3 nanoparticles, facilitating their homogeneous dispersion and stability within COL-1 hydrogel matrix. This optimized BaTiO3@COL-1 hydrogel bioelectronics was fabricated as an injectable system for neural repair. Under low-intensity ultrasound, the hydrogel bioelectronics exerted mechanoelectrical conversion. B Mechanical-electrical-bioenergetic energy conversion for energy-consuming neural repair. Ultrasound-driven piezoelectric hydrogel bioelectronics delivered mechanical stimulation to glia, which primarily sensed the stimulation through surface PIEZO ion channel proteins to mediate calcium ion influx. The calcium ion influx via PIEZO channels (PIEZO1 in astrocytes and PIEZO2 in Schwann cells) activated intracellular signaling pathways to promote mitochondrial fusion as a central bioenergetic hub and modulate glial-mediated neural repair in both central and peripheral nervous systems. C The therapeutic efficacy and biosafety of piezoelectric bioelectronics was verified in multiple animal models (mice, rats, Beagle dogs, and Rhesus monkeys) from peripheral and central nervous injuries, demonstrating its wide adaptivity and significant translational potential.
Results
Properties of ultrasound-driven piezoelectric hydrogel bioelectronics
BaTiO3 nanoparticles were synthesized via a solvothermal approach, resulting in predominantly obtuse-angled cuboids with a uniform average diameter of approximately 90.53 nm (Fig. 2A–C). Elemental mapping (Fig. 2D) revealed the homogeneous distribution of O, Ba, and Ti, confirming the composition of the synthesized BaTiO3 nanoparticles.

A Scanning electron microscopy (SEM) image of BaTiO3 nanoparticles (BTNPs). B Particle size distribution by SEM image of BTNPs. C Transmission electron microscopy (TEM) image of BTNPs. D Ba, Ti, and O element mapping of a single BTNP by Energy Dispersive Spectrometer (EDS). A, C and D are representative images from one of three independently prepared samples with similar results. E XRD pattern of BTNPs and enlarged (002) and (200) peaks (the inset). F High-resolution transmission electron microscopy (HRTEM) image of BTNP. G Fast Fourier transform (FFT) pattern of an individual BTNP. H Raman spectra of BTNPs. I The amplitude variation curve of a point on the BTNP as a function of the applied DC bias voltage measured by Piezoresponse Force Microscopy (PFM). J Phase variation of BTNP with DC bias scanning. (K, L) Confocal reflection microscopy images of COL-1 hydrogel K and BaTiO3@COL-1 hydrogel L under 488 nm laser. Representative image from one of three independently prepared samples with similar results. (M-O) Rheological characterization of BaTiO3@COL-1 hydrogel. Data are presented as mean ± SD from n = 3 independent samples.
The piezoelectric properties of BaTiO3 arise from the displacement of Ti ions from the B-site in the tetragonal perovskite structure (ABO3), generating a dipole moment and extending the crystal cells along the c-axis. Lattice parameters were quantified microscopically to confirm the phase structure, with X-ray diffraction (XRD) analysis (Fig. 2E) yielding lattice constants a = 0.399416 nm and c = 0.404559 nm, consistent with the tetragonal P4mm phase (JCPDS No. 96-151-3253). The c/a ratio aligned with literature values37. The presence of distinct (002) and (200) peaked at 45°, with an intensity ratio of approximately 1:2, further supported the existence of a well-defined tetragonal structure. Fast Fourier Transform (FFT) analysis of the high-resolution transmission electron microscopy (HRTEM) images (Fig. 2F) identified the (001) and (100) planes with interplanar spacings of 0.4041 nm and 0.3983 nm in agreement with the XRD results. These observations were corroborated by selected area electron diffraction (SAED) patterns (Fig. 2G). Raman spectroscopy further confirmed the tetragonal phase, peaking at 243.95, 307.981, 515.026, and 715.117 cm−1 corresponding to A1(TO), E, B1(TO + LO), and E, A1(TO) vibrational modes (Fig. 2H), consistent with prior reports of tetragonal BaTiO338. In cubic BaTiO3, the Ti4+ ions are symmetrically located in the TiO6 octahedra, preventing first-order Raman activity38; however, the observed Raman peaks indicated the presence of tetragonal distortions in the TiO6 octahedra. In summary, analysis of XRD, HRTEM, and Raman spectra confirmed that the BaTiO3 nanoparticles synthesized exhibited a tetragonal phase, crucial for their piezoelectric behavior.
Piezoresponse force microscopy (PFM) demonstrated a characteristic butterfly loop under an applied voltage ranging from −15 V to 15 V (Fig. 2I), indicating a mechanical response to electrical stimulation. Additionally, a 180° phase shift in the ferroelectric domain after polarization reversal in the coercive field confirmed the ferroelectric properties of the BaTiO3 nanoparticles (Fig. 2J). Comsol Multiphysics technique verified the piezoelectric response of BaTiO3 under ultrasound stimulation (Supplementary Fig. 1A). Significant electric potential variations were observed with a cubic shell side length of 100 nm (Supplementary Fig. 1B–D), particularly at the cube’s center where charge accumulation and enhanced piezoelectric effects were noted (Supplementary Fig. 1E, F). It should be noted that due to the absence of specific hydrogel parameters, water was selected as the medium in the simulation performed at 100 kHz. These findings showed the potential of BaTiO3 nanoparticles for converting mechanical energy into electrical energy by ultrasound, providing a foundation for their use in electrical stimulation application.
For hydrogel bioelectronics fabrication, BaTiO3 nanoparticles were incorporated into an injectable type 1 collagen (COL-1) hydrogel, the most abundant ECM protein in nerve perineurium and epineurium39. COL-1 fiber network with or without BaTiO3 nanoparticles illustrated the material suitability for further biomedical use (Fig. 2K, L). Further rheological analysis of the BaTiO3@COL-1 hydrogel bioelectronics revealed that under a rotational speed of 1 Hz, this hydrogel retained gel characteristics at a strain of up to 10%, while the elastic modulus started to decrease at 3% strain (Fig. 2M). Additionally, during a frequency sweep at a constant 3% strain, the hydrogel exhibited stable mechanical performance up to 10 Hz, beyond which the gel structure was disrupted (Fig. 2N). Based on these findings, further experiments were conducted with a strain of 3 % and a frequency of 8 Hz for long-term rheological testing. The storage modulus (value of G’) rises quickly and reaches equilibrium in 1 min and over the next 30-minute period, the hydrogel demonstrated sustained mechanical integrity (Fig. 2O). The hydrogel was pre-shaped for structure, gelation in the incubator at 37 °C for 30 min in vitro and implanted to the injury site (Supplementary Movies 1, 2). We were also interested in the behavior of bioactive hydrogel bioelectronics under high frequency. Following ultrasound treatment, we observed that the hydrogel was sufficient to maintain its structure, suggesting minimal degradation under the applied ultrasound regime (Supplementary Movies 3, 4). These results suggested that the mechanical properties of the hydrogel were sufficient to maintain its structure in vivo by providing adequate encapsulation strength.
PIEZO2 mediated mechanotransduction for Schwann cell neuromodulation
Schwann cells are the predominant glial cells of the peripheral nervous system, playing indispensable roles in myelination, axonal regeneration, and neural repair, thereby rendering them highly sensitive to changes in the mechanical and metabolic environment8. Schwann cells were embedded in BaTiO3 hydrogel bioelectronics. The biosafety threshold of barium titanate compounds has not yet been defined by official regulatory bodies. Therefore, in our study, the selection of a safe concentration was based on previously published research40. Compared to fully dissolved BaCl₂ solutions, ICP-MS analysis showed that BaTiO3 nanoparticles released only a small fraction of Ba²⁺ under physiological conditions (Supplementary Fig. 2). To determine the optimal BaTiO3 concentration for cell compatibility, we varied the concentration from 0.05 mg/mL to 0.5 mg/mL (concentration of BaTiO3 in phosphate-buffered saline (PBS) suspension, the final concentration of BaTiO3 in hydrogel varied from 0.005 to 0.05, respectively) according to previous literature41. As shown by cell counting kit-8 (CCK-8) and live/dead cell staining, changes in BaTiO3 concentration altered cell proliferation and apoptosis (Supplementary Fig. 3A–C). There was no statistically significant difference between 0.05 mg/mL and 0.1 mg/mL, while 0.5 mg/mL displayed appreciable cytotoxicity. We next investigated the electrochemical property of BaTiO3 hydrogel bioelectronics with 0.1 mg/mL concentration. When the detecting electrode spacing was 6 cm, the resistance decreased sixfold, from 57.03 ± 4.40 kΩ to 9.06 ± 1.02 kΩ, after BaTiO3 particles addition (Supplementary Fig. 1G). Meanwhile, BaTiO3@COL-1 hydrogel bioelectronics generated microcurrent nearly one time higher than that of non-piezoelectric hydrogel bioelectronics under the same ultrasound stimulation (Supplementary Fig. 1H). As such, we used 0.1 mg/mL for subsequent in vivo and in vitro experiments in this study.
Next, we performed serial cytotoxicity assays to assess cell compatibility of ultrasound-driven BaTiO3 hydrogel bioelectronics. To distinguish the expression patterns of piezoelectric channel proteins (PIEZO1/2), we set different groups as COL-1 hydrogel, BaTiO3@COL-1 hydrogel bioelectronics, ultrasound-driven COL-1 hydrogel, and ultrasound-driven BaTiO3@COL-1 hydrogel bioelectronics and term these groups as different abbreviations: HG, Piezo-HG, HG + US and Piezo+US. The ultrasound stimulation lasted for five minutes per session each day and intensity remained 0.5 W/cm2 as low-intensity application42. The results indicated that Schwann cells viability in Piezo+US group was increased compared with piezoelectric nanoparticles or ultrasound treatment alone (Supplementary Fig. 4A, B). Piezo+US group exhibited increased 5-ethynyl-2’-deoxyuridine (EdU)-labeled proliferative cells compared with the HG group (Supplementary Fig. 4C, D). Moreover, flow cytometric analysis and superoxide content detection showed that ultrasound-driven BaTiO3 hydrogel bioelectronics decreased intracellular ROS levels (Supplementary Fig. 4E, F). It indicated that piezoelectricity via ultrasound was favorable to cell compatibility42,43.
To clarify the influence of ultrasound-driven BaTiO3 hydrogel bioelectronics on mechanotransduction and cell fate, Schwann cells were cultured in 3D piezoelectricity culture system with or without ultrasound for 3 days. It was noteworthy that ultrasound stimulation or piezoelectric materials alone did not increase PIEZO 1/2 gene expression level (Fig. 3A). Although the gene expression of piezo1 showed statistical significance between Piezo+US group and HG-control group, the fold change was less than 1.2 and there was no difference in PIEZO1 protein expression (Fig. 3A, Supplementary Fig. 5A). However, Piezo+US treatment upregulated PIEZO2 expression level nearly 9-fold higher than the control group. The expression level of PIEZO2 was higher in Piezo+US group compared with the piezoelectric hydrogel alone, indicating that ultrasound was required to enhance the PIEZO2 response to mechano-bioactive hydrogel bioelectronics (Fig. 3B, Supplementary Fig. 5B). We propose that PIEZO2 is the major factor for Schwann cells mechanotransduction following mechano-bioactive hydrogel bioelectronics treatment.

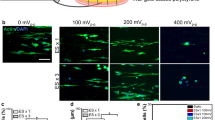
A PIEZO1/2 mRNA expression. n = 3. B Western blot of PIEZO2. C Temporal profile of PIEZO2 mRNA expression in rat Schwann cells underwent single or two rounds ultrasound stimulation, relative to levels in the control group. n = 3. D Analysis of mRNA expression of voltage-gated calcium channels (VGCCs). n = 3 per group. E Illustration of mechanotransduction to cellular biological behaviors by ultrasound. F IF staining of calcium marker F4-AM. Scale bar, 25 μm. G Statistical analysis of mean F4-AM fluorescence intensity (%) compared to Piezo+US group. n = 8 in HG group, n = 13 in Piezo+HG group, n = 14 in HG + US group, and n = 18 in Piezo+US group. H Western blot bands of Calmodulin, cAMP, and JNK with different treatments. I Independent temporal profile of bdnf, gdnf, and S100β mRNA expression (qRT-PCR) in rat Schwann cells. n = 3. J IF staining of outer mitochondrial membrane TOM20. Scale bar, 50 μm. K Statistical analysis of mean TOM20 fluorescence intensity (%) compared to Piezo+US group. n = 12 in HG + US group and n = 14 in the other groups. L Cell energy status was assessed by measuring ATP levels. n = 6. M Western blot bands of Glut1, Glutamine Synthetase, and ATP Synthase beta. N Western blots bands of proteins related to mitochondrial dynamics. O RNA sequencing of rat Schwann cells embedded in ultrasound-driven piezoelectric hydrogel bioelectronics. Upregulated differentially expressed genes (DEGs) related with Schwann cells energy metabolism and signaling pathways (Log2FC > 2), right panel. P Volcano plot of DEGs in rat Schwann cells relative to the ctrl group. Q Activation of Schwann cells mitochondria metabolism and signaling transduction using KEGG pathway analysis of upregulated genes. The statistical tests were two-sided and no adjustments were made for multiple comparisons. Statistical significance was determined by one-way ANOVA with Dunnett’s post hoc test or Student’s t-test as indicated (*p < 0.05, **p < 0.01, ***p < 0.001). Data are mean ± SD. HG: collagen hydrogel; Piezo-HG: BaTiO₃ hydrogel; HG + US: ultrasound-driven hydrogel; Piezo+US: ultrasound-driven BaTiO3 hydrogel.
Gene expression curve showed that PIEZO2 activated upon stimulation and peaked at 30 min after ultrasound treatment. Interestingly, when these cells continued to receive ultrasound stimulation on day 2, it triggered a more intense PIEZO2 gene expression response, with expression levels surpassing the peak of day 1 in a short time (Fig. 3C), endowing Schwann cells with a greater ability for mechanical response. The properties of piezoelectric materials depend on generating electrical charges in response to mechanical pressure. We noticed that mechanical signal transduction subjected to ultrasound could be accompanied with the change of membrane potential, leading to coordinated activation of some voltage-gated channels44. Thus, we also analyzed gene expression of voltage-gated calcium channels (VGCCs) to evaluate whether the calcium influx in Schwann cells involves the other ion channels activated by Piezo+US (Fig. 3D). In this experiment we studied VGCCs families from Cav1 to Cav3. Piezo+US significantly upregulated Cav1.1 (CACNA1S), Cav 1.2 (CACNA1C), Cav 2.2 (CACNA1B), Cav 2.3 (CACNA1E), and Cav 3.1 (CACNA1G). Some VGCCs were reported to regulate neurotransmitter transmission45,46,47. The PIEZO channels primarily mediate calcium influx (Fig. 3E)48. In Piezo+US group, obvious calcium influx was detected by Fluo-4AM compared with the HG and Piezo+HG groups (Fig. 3F, G)49. It indicated that calcium influx was significantly increased only when cells were treated with ultrasound-driven piezoelectric nanoparticles, whereas either ultrasound or nanoparticles only did not change intracellular calcium concentrations obviously19. We next explored the downstream cellular activities responding to external mechanical signals. After calcium influx, the calcium-binding messenger protein calmodulin served as primary intracellular receptors for Ca2+ signal and regulated various target proteins and enzymes50. The cAMP was another important neurotransmitter in calcium signaling51. Ultrasound-driven BaTiO3 hydrogel bioelectronics significantly upregulated calmodulin and cAMP protein levels (Fig. 3H, Supplementary Fig. 5C, D). Notably, pls3 influenced Piezo-mediated mechanotransduction through its effects on the cytoskeleton stabilization. The result revealed the highest pls3 expression level, more than 1.7-fold compared with other groups, in the Piezo+US group (Supplementary Fig.4G). Increased calcium influx regulated Schwann cells neuromodulation activity52,53. We found upregulated JNK, an important factor required for dedifferentiation, in Piezo+US group (Fig. 3H, Supplementary Fig. 5E). In addition, gene expression curves indicated that neurotrophic factor BDNF, GDNF, and Schwann cell marker S100β expression increased and peaked at 30 min after ultrasound treatment, highly consistent with PIEZO2 expression curve (Fig. 3I). The results suggested that these neurotrophic factors had a synergistic relationship with PIEZO2 expression. Taken together, mechanical stimulation from ultrasound-driven BaTiO3 hydrogel bioelectronics upregulated PIEZO2 channel and mediated calcium influx in Schwann cells.
Given that the coordination between Schwann cells and endothelial cells was crucial in neural regeneration, we investigated the effect of Schwann cells conditioned medium on angiogenesis54. The human umbilical vein endothelial cell line (HUVEC) showed increased lumen-like structures cocultured with conditional medium derived from Piezo+US group (Supplementary Fig. 4H, I). Furthermore, we performed chicken chorioallantoic membrane (CAM) assay to evaluate the role of conditional medium in angiogenesis in vivo (Supplementary Fig. 4J, K). The results showed the proangiogenic ability of Schwann cells in the Piezo+US group.
Typically, mitochondria coordinate cellular adaptation and served as the primary organelle for ATP production and energy supply in Schwann cells55,56. We aimed to investigate the relationship between cell behavior with mitochondria activity. In this regard, immunofluorescence (IF) was used to observe mitochondria with the outer membrane protein TOM-20 and inner membrane dye MitoLite. Ultrasound-driven BaTiO3 hydrogel bioelectronics induced an increase of TOM-20 and MitoLite-labeled mitochondria compared to the HG and Piezo+HG groups (Fig. 3J, K, Supplementary Fig. 4L, M). Consistently, more ATP was produced in Piezo+US group than either piezoelectric hydrogel bioelectronics or ultrasound treatment, indicating more active energy metabolism after ultrasound-driven BaTiO3 hydrogel bioelectronics treatment (Fig. 3L). We further analyzed proteins related to ATP production. Upregulation of GLUT1 enhanced glucose uptake and increased ATP production57. Glutamine synthetase catalyzed the conversion of glutamate to glutamine, a process intricately linked to ATP production by amino acid metabolism58. ATP synthase subunit beta was a key component of the ATP synthase complex in the mitochondrial inner membrane, responsible for ATP production59. In Piezo+US group, these proteins were more active than others (Fig. 3M, Supplementary Fig. 5F–H). Mitochondria are adaptive to fluctuations in energy status and nutrient availability through morphological alterations such as fusion and fission60. We next assessed Schwann cell mitochondrial dynamics proteins. The increased expression of Opa1, MFN1, MFN2 and decreased expression of pDrp1 and MFF suggested an improved mitochondrial fusion activity facilitating cellular energy metabolism and resistance to stress (Fig. 3N, Supplementary Fig. 5I–M).
After mechano-bioactive hydrogel bioelectronics treatment, both PIEZO2 channel and VGCCs were upregulated and contributed to calcium influx. To determine which channel played the predominant role, we employed PIEZO2 knockout technique and L-type VGCCs inhibitor verapamil. PIEZO2 knockout Schwann cells displayed weakened neuromodulation ability. Upon mechanical stimulation from piezoelectric hydrogel bioelectronics, calcium influx decreased significantly in cells following PIEZO2 knockout (Supplementary Fig. 6A, B). Given that VGCCs were also activated after mechanical stimulation (Fig. 3D), calcium influx was not fully inhibited after PIEZO2 knockout. This was further supported by the evidence that the fluorescence intensity of Schwann cell marker S100β and mitochondria marker TOM20 were largely reduced in PIEZO2 knockout Schwann cell (Supplementary Fig. 6C-F). Moreover, PIEZO2 knockout attenuated mechanoelectrical conversion-induced calcium signaling and mitochondrial dynamics activity at protein levels (Supplementary Fig. 6G, H). In contrast, immunofluorescence analysis showed that verapamil mildly reduced calcium intensity, suggesting that PIEZO2 was the primary channel mediating calcium influx (Supplementary Fig. 7A, B). Western blot revealed that verapamil had minimal impact on PIEZO2 expression and cAMP signaling pathway activation (Supplementary Fig. 7C, D). Additionally, mitochondrial activation and energy production were not significantly altered (Supplementary Fig. 7E–G), nor did verapamil affect the activation of the Schwann cell repairing phenotype (Supplementary Fig. 7I–K). Collectively, these results confirmed the mechanotransduction effect of PIEZO2 on Schwann cell neuromodulation.
To understand the mechanism underlying PIEZO2-mediated mechanotransduction in Schwann cells, we identified the transcriptional signature by bulk RNA-Seq. Schwann cells cultured in 3D piezoelectric hydrogel bioelectronics under ultrasound stimulation showed global differences in gene expression compared with the control group (Fig. 3O, P). The top differentially upregulated genes could be categorized in extracellular matrix sensing and remodeling (Crnn, Mmp15, Vcan, Mmp10), signal transduction (Mcoln3, Ednrb, Pln, Adgrv1), and amino acid synthesis (Flrt1, Tph1, Ptprv). Extracellular matrix changes collaborate with mechanosensitive membrane proteins to sense and regulate mechanotransduction61. Several of the above genes, including Mcoln3, Ednrb, and Pln, were involved in calcium signaling regulation. In addition, Flrt1, Tph1, and Ptprv were closely related with leucine, tryptophan, and tyrosine which played crucial roles in neuromodulation and synthesis in mitochondria55. Consistently, Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis revealed that these dysregulated genes were enriched in signal transduction pathways represented of calcium signaling and amino acid metabolism pathways (Fig. 3Q). Collectively, ultrasound-driven piezoelectric hydrogel bioelectronics exerted electrical signals from mechanical stimulation and upregulated PIEZO2 channel in Schwann cells. The mechanoelectrical conversion increased calcium influx to activate ATP synthase subunit and promote MFN/OPA1-mediated mitochondrial fusion.
PIEZO1 mediated mechanotransduction for repairing phenotype of astrocytes
Astrocytes, the most abundant resident cells in the CNS, represented central regulator of neuronal function and energy metabolism. To clarify whether astrocytes adapt their function similarly as Schwann cells in response to mechanical stimulation from piezoelectric bioelectronics, we cultured astrocytes in BaTiO3@COL-1 hydrogel bioelectronics with or without ultrasound. Unlike the Schwann cells, astrocytic PIEZO1 rather than PIEZO2 level was significantly upregulated in Piezo+US group (Fig. 4A, B), consistent with previous literature indicating that astrocytic PIEZO1 sensed and mediated mechanotransduction7. Notably, ultrasound was indispensable for the response of astrocytic PIEZO1 to hydrogel bioelectronics. The PCR array indicated that activation of VGCCs in astrocytes may contribute to the complexity of calcium influx induced by mechanical stimulation, although most changes were not statistically significant (Fig. 4C, Supplementary Fig. 8A). After spinal cord injury (SCI), astrocytes undergo phenotypic change known as reactive astrocytes (RAs) and then scar-formed astrocytes (SAs). We identified molecular signatures for RAs and SAs associated with mechanotransduction. Cdh2, csgalnact1, chst11, slit2, nes, axin2, mmp13, and mmp2 genes were selected as characteristics of RAs and SAs62,63. The axin2, mmp13, and mmp2 expression levels in Piezo+US group were higher than those in other groups while expression level of SAs-related genes was the lowest (Fig. 4D). We next investigated genes associated with astrocytic secretion function (A2m), neural supportive function (Ngf, Pdgf, GLP-1R, Slc2a1), and synapses (Thbs2)63. The data indicated that although piezoelectric hydrogel bioelectronics or ultrasound alone upregulated several genes, the synergistic effect of ultrasound-driven piezoelectric bioelectronics had stronger effect on upregulating the expression of these genes (Fig. 4E, Supplementary Fig. 8B).

A, B PIEZO1/2 mRNA expression in astrocytes. n = 3. C PCR array of voltage-gated calcium channels (VGCCs) and Caveolin (Cav) genes expression shown in heatmaps, normalized to HG group. D Marker gene signatures of reactive astrocytes and scar astrocytes, normalized relative to HG group. E Gene signatures of reactive astrocytes were analyzed using PCR array method and indicated in the heat map. The expression data were processed and normalized. F IF staining of outer mitochondrial membrane TOM20. Scale bar, 10 μm. G Co-localization analysis of TOM20 and DAPI. H Statistical analysis of mean TOM20 fluorescence intensity (%). n = 8. I IF staining of inner mitochondrial membrane MitoLite™ Red CMXRos and calcium marker F4-AM. Scale bar, 10 μm. J Statistical analysis of mean MitoLite and F4-AM fluorescence intensity (%) compared to Piezo+US group. n = 5. K Western blot bands of Glutamine Synthetase, ATP Synthase beta, and GAD67 in rat Schwann cells. L Western blots bands of proteins related to mitochondrial dynamics. M HUVEC tube formation assay that investigate the effect of conditional medium from rat brain astrocytes cells on HUVEC tube formation. Scale bar, 200 μm. N Statistical analysis of the HUVEC tube formation assay. n = 5. (O, P) Representative images O and quantitative analysis of blood vessels P of CAM treated with astrocytic conditional medium from ultrasound-driven piezoelectric hydrogel bioelectronics. Axitinib was set as the control inhibitor group. Scale bar, 0.5 cm. n = 3. Q Volcano plot of DEGs in rat brain astrocytes cells relative to ctrl group. R Activation of rat brain astrocytes cells energy metabolism and signaling transduction using KEGG pathway analysis of upregulated genes. Size represents number of genes regulated and the pathways of the colored circles are highlighted by the text. The statistical tests were two-sided and no adjustments were made for multiple comparisons. Statistical significance was determined by one-way ANOVA with Dunnett’s post hoc test or Student’s t-test as indicated (*p < 0.05, **p < 0.01, ***p < 0.001). Data are mean ± SD. HG: collagen hydrogel; Piezo-HG: BaTiO₃ hydrogel; HG + US: ultrasound-driven hydrogel; Piezo+US: ultrasound-driven BaTiO3 hydrogel.
Calcium signaling also regulates mitochondrial functions in astrocytes64. We explored mitochondrial morphology and functions, and found that mitochondrial network was maintained with less point structure in Piezo+US group revealed by TOM20 and MitoLite-labelled immunofluorescence (Fig. 4F–J). We also determined the crucial role of PIEZO1 in mediating mechanical-electrical-bioenergetics conversion. Following PIEZO1 knockout, TOM20 immunofluorescence intensity was reduced significantly in Piezo+US group, indicating decreased mitochondrial activity (Supplementary Fig. 9A, B). After verapamil treatment, piezoelectric material-induced calcium influx decreased by approximately 20% (Supplementary Fig. 9C, D). Downregulation of PIEZO channel and cAMP signaling induced mitochondria activity were more pronounced in astrocytes than in Schwann cells (Supplementary Fig. 9E–H). Piezo+US treatment promoted astrocyte differentiation toward the S100a10+ A2 repairing phenotype while significantly reducing C3+ A1 phenotype. Verapamil not only decreased downstream S100a10 expression but also increased C3 levels (Supplementary Fig. 9K). Astrocytes have essential metabolic functions like glutamatergic neurotransmitter recycling. ATP was crucial for powering these metabolic processes. GAD67 is an enzyme involved in the conversion of glutamate to gamma-aminobutyric acid, primarily in neurons but also expressed in astrocytes65. Therefore, we investigated if these key markers were activated during mechanotransduction. The expression levels of glutamine synthetase, ATP synthase subunit beta and GAD 67 were significantly higher than those in the HG/control group (A fold-change of 5.6, 1.6, and 4, respectively) (Fig. 4K, Supplementary Fig. 10A–C). A similar trend in the expression of mitochondrial dynamics proteins was observed in astrocytes, akin to Schwann cells in PNS, suggesting calcium signaling activated mitochondrial energy metabolism in glia with ultrasound-driven piezoelectric hydrogel bioelectronics (Fig. 4L, Supplementary Fig. 10D-G). In gliovascular units, astrocytes were necessary for micro-vasculature function66. Astrocytic conditioned medium greatly enhanced vascular formation in Piezo+US group (Fig. 4M–P).
We then focused on the transcriptional signature of mechanotransduction in astrocytes. There were 29 downregulated genes and 298 upregulated genes after three-day ultrasound-driven BaTiO3 hydrogel bioelectronics treatment. The top elevated differentially expressed genes (DEGs) involved Krt23, Krt15 (regulation of cellular structure and mechanical stability), Mast4, Pvrl4 (cytoskeletal dynamics), Ppp2r2c, Cacna1s (signal transduction), and Setbp1, Il6 (neural regulation) (Fig. 4Q). Furthermore, KEGG analysis suggested potential changes in signaling pathways associated with mechanotransduction (Fig. 4R)67. Accordingly, mechanical stimulation by ultrasound-driven BaTiO3 hydrogel bioelectronics activated astrocytic PIEZO1 channel. Calcium influx initiated mitochondrial fusion as energetic hub for astrocytic repairing process. It indicated a similar intracellular mechanism in glia of the CNS.
PIEZO-calcium-CREB signaling functioned in glial neuromodulation and energy metabolism
PIEZO1/2 were the main mechano-transducers and mediated calcium influx. Ultrasound-driven mechano-bioactive hydrogel bioelectronics treatment upregulated cAMP and calmodulin in both Schwann cells and astrocytes. KEGG enrichment analysis of both cell types revealed enrichment of the cAMP and calcium signaling pathways, underscoring the pivotal role of calcium signaling in piezoelectric material–mediated glial activation. The present study further investigated how glia expressed PIEZO1/2 to respond to ultrasound-driven mechano-bioactive hydrogel bioelectronics. JASPAR prediction of transcription factors with PIEZO1/2 gene promoters indicated high likelihood of binding by cAMP-responsive element-binding protein 1 (CREB1) (Supplementary Fig. 11, Table S7). Inhibition of calcium influx with verapamil markedly reduced CREB protein expression in both Schwann cells and astrocytes (Supplementary Fig. 7H, 9I, J), suggesting the involvement of the calcium–CREB signaling in glial activation. Furthermore, inhibition of CREB with KG501 downregulated glial PIEZO1 and PIEZO2 protein expression and significantly reduced calcium influx (Supplementary Fig. 12A–E, I–J). KG501 also suppressed mitochondrial fusion proteins OPA1, Mfn1, and Mfn2, indicating that the calcium–CREB axis regulated mitochondrial fusion in activated glia (Supplementary Fig. 12D–M). Moreover, KG501 significantly reduced GDNF secretion from repair-phenotype (S100β) Schwann cells and NGF secretion from A2-type (S100a10) astrocytes, demonstrating that calcium–CREB axis contributed to glial neuromodulation. Collectively, PIEZO–calcium–CREB signaling played a central role in glial neuromodulation and energy metabolism.
Schwann cell responsive mechanical neuroengineering for peripheral nerve regeneration
Based on material characterization and in vitro experiments, we hypothesize this mechanical neuroengineering would be an alternative for neural repair. Rat and rhesus monkey sciatic nerve crush injury models were used to verify this hypothesis.
Sprague Dawley (SD) rats were randomly divided into four groups: (i) PNI only without treatment; (ii) non-Piezo hydrogel; (iii) Piezo hydrogel bioelectronics; (iv) and ultrasound-stimulating Piezo hydrogel bioelectronics at two time points (two and four weeks after surgery). On the surgery day, BaTiO3 hydrogel bioelectronics was implanted and wrapped the injured nerve (Fig. 5A). After closing the wound, ultrasound stimulation was applied (five minutes once a day for three-week period) (Supplementary Movie 5). To assess the in vivo concentration of Ba²⁺ following material implantation, muscles and blood samples were collected one month post-surgery. Given the slow and minimal release rate of Ba²⁺ from BaTiO3 nanoparticles, the Ba²⁺ concentrations detected in vivo are typically well below established toxicological thresholds (Supplementary Tables 1, 2).

A Schematic images of ultrasound-driven piezoelectric hydrogel bioelectronics in rat peripheral nerve crush injury model. B Representative transverse sections of nerves were immunohistochemically stained with PIEZO2 polyclonal antibody. Scale bar in low magnification images, 200 μm. Scale bar in higher magnification images, 100 μm. C Quantification of PIEZO2 positive staining contribution. n = 5 per group. D H&E staining of longitudinal sections of injured nerves two weeks after surgery. Scale bar, 100 μm. E Sirius red staining of longitudinal sections of injured nerves four weeks postoperatively under polarized light microscopy. Scale bar, 100 μm. F Representative IF staining of growth cones marker growth associated protein 43 (GAP43, green), Schwann cell marker S100β (red), and peripheral neurons marker βIII tubulin (orange). Scale bar, 100μm. G Statistical analysis of mean fluorescence intensity (%) of GAP43, S100β, and βIII tubulin. n = 3 per group. H Quantitative analysis (mean ± SD) for nerve conduction velocity (NCV) and compound muscle action potential (CMAP). n = 5. I Representative electromyography curves of rats in different groups. J Transmission electron microscope images of myelin sheath four weeks after PNI. Scale bar, 5 μm. Small bar insert, 2μm. K Toluidine blue (TB) staining of regenerated nerves four weeks after PNI. Scale bar, 40 μm. (L, M) Quantitative analysis (mean ± SD) for axon diameter L and thickness of myelin sheath M. n = 10 per group. N Representative images of gastrocnemius sections stained for basement membrane marker laminin to visualize muscle fiber structure. Scale bar, 100 μm. O Rat hot plate pain model four weeks postoperatively. n = 6 per group. Statistical significance was determined by one-way ANOVA with Dunnett’s post hoc test or two-tailed unpaired Student’s t-test (*p < 0.05, **p < 0.01, ***p < 0.001). Data are mean ± SD. PNI: peripheral nerve injury group; HG: hydrogel group; Piezo+HG: piezoelectric hydrogel group; Piezo+US: ultrasound-driven piezoelectric hydrogel group. Figure 5A was drawn by Adobe Photoshop.
For histological evaluation, we employed (i) immunohistochemistry to visualize PIEZO2 expression in the injury site; (ii) H&E staining and Sirius red staining for nerve structure evaluation; and (iii) immunofluorescence (IF) staining to assess Schwann cell activation and neural regeneration. After two weeks of ultrasound-driven piezoelectric hydrogel bioelectronics implantation, more PIEZO2 positive areas were observed in nerve transverse sections (Fig. 5B, C). Meanwhile, the nerve fibers exhibited clearer and more homogeneous structure in Piezo+US group, while in PNI group nerve fibers were still disorganized with a broken axon network (Fig. 5D). Four weeks post injury, denser and more oriented collagen deposition was found in Piezo+US group to favorably support nerve fiber and reduce scar formation (Fig. 5E)54. Furthermore, the longitudinal nerve sections were labeled with growth cone marker GAP43 (green), microtubule marker βIII-Tubulin (yellow), and Schwann cell marker S100β (red) (Fig. 5F). Despite no statistical difference in GAP43 intensity among all groups, piezoelectric hydrogel bioelectronics preserved nerve fibers and activated Schwann cells regardless of the application of additional ultrasound stimulation (Fig. 5G). The transverse sections suggested activation of more Schwann cells after Piezo+US mechanical stimulation, consistent with in vitro results (Supplementary Fig. 13A, B). CD34 was a common marker for vascular endothelial cells and angiogenesis. The mean fluorescence intensity was stronger in Piezo+US group than the PNI group, indicating active angiogenesis in vivo by ultrasound-driven piezoelectric hydrogel bioelectronics (Supplementary Fig. 13A, C).
Accelerated nerve fiber structure was beneficial for electrical signal transmission and functional recovery. We performed electrophysiological monitoring and found that ultrasound-driven BaTiO3 hydrogel bioelectronics significantly improved nerve conduction velocity (NCV) (57.04 ± 0.30 m/s) and compound motor action potential (CMAP) (16.66 ± 3.54 mV) compared with other groups (Fig. 5H, I). The myelin sheath and axon diameter decide the quality of electrical signal transmission after nerve injury. We therefore performed TEM and toluidine blue (TB) experiments to observe the regenerated sciatic nerve morphology68. The result revealed the thickest myelin sheath (0.49 ± 0.11 μm) and longest axon diameter (3.16 ± 0.23 μm) in Piezo+US group (Fig. 5J–M). Repaired myelin structure alleviated nerve denervation, and induced stronger hindlimb muscles and more coordinated movement in rats of Piezo+US group (Fig. 5N, Supplementary Fig. 13D-F, Supplementary Movie 6). We next evaluated the plantar of injured limb four weeks after injury. Due to self-mutilation behavior following denervation, PNI group exhibited significant plantar ulceration, while non-Piezo hydrogel and Piezo hydrogel bioelectronics group also showed a certain degree of atrophy and degeneration at the tips of the toes (Supplementary Fig. 13G). Furthermore, we investigated sensory recovery and emotional state by hot plate test and tail suspension test, respectively. In the hot plate test, thermal nociception of rats received piezoelectric hydrogel bioelectronics under ultrasound stimulation recovered more quickly at the fourth week and showed a reduced reaction latency compared with other controls (Fig. 5O). When rats were suspended by their tails in an enclosed chamber, a shorter struggling duration (mobility rate) indicates a depressive state69,70. The Piezo+US group exhibited superior mobility rate (90.47 ± 8.15 %) than the PNI group (44.20 ± 21.30 %). It suggested that piezoelectric hydrogel bioelectronics with ultrasound stimulation kept more active emotional state of PNI rats (Supplementary Fig. 13H, I). For in vivo biocompatibility analysis, we performed H&E staining to study the morphology of major organs harvested from rats and observed no notable abnormality in the heart, liver, spleen, lung, and kidney (Supplementary Fig. 14).
The ultrasound-driven piezoelectric hydrogel bioelectronics successfully promoted peripheral nerve regeneration in rat model. We further tested its efficacy and biosafety in rhesus monkeys sciatic nerve crush injury model, to better evaluate the clinical translational potential (Fig. 6A, Supplementary Movie 7). After 4-week implantation, rhesus monkey sciatic nerves were harvested. The piezoelectric hydrogel bioelectronics did not provoke obvious inflammation or cell apoptosis to the major organs as shown by H&E staining, indicating it was non-toxic to primates (Supplementary Fig. 15). Furthermore, blood samples were collected from rhesus monkeys subjected to BaTiO3 hydrogel bioelectronics implantation to analyze the haematology and biochemistry profiles. No obvious abnormal alternations were identified (Supplementary Table 3). Although ultrasound stimulation was not applied to the injured limb, we still observed more PIEZO2 positive immunohistochemistry distributions in the transverse sections of Piezo+HG group, possibly due to more intense muscle contraction activity in rhesus monkeys. It exerted considerable mechanical stimulation to the piezoelectric hydrogel bioelectronics (Fig. 6B, C). In contrast to the control group, nerve fibers were denser and more arranged in the Piezo+HG group (Fig. 6D). Next, we assessed neural regeneration based on multiple-labeling immunofluorescence. Piezo+HG samples had higher neuronal microtubules and growth cones distribution accompanied with accelerated angiogenesis (Fig. 6E, F). Notably, most immunohistochemistry or immunofluorescence antibodies have not been validated in rhesus monkeys, so we conducted preliminary staining trials and selected more specific antibodies for presentation. For innervated organs, we employed histological evaluation to assess target tissue atrophy (Fig. 6G). Piezo+HG treatment promoted higher muscle fiber perimeter and area (Fig. 6H, I). Meanwhile, Piezo+US group displayed more hair follicles in density and quantity of the innervated skin samples (Fig. 6J). These results suggested that piezoelectric hydrogel bioelectronics improved the reinnervation of the target organ. PIEZO2 is directly associated with the touch sensation in sensory neurons and skin cells71. Four weeks after surgery, the affected limb of the rhesus monkey showed significant muscle strength recovery and activity (Supplementary Movie 8). Moreover, we inserted stimulation electrodes at the proximal end of crushed sciatic nerve and applied stimulation at different voltages (Fig. 6K). As the voltage increased, the Piezo+HG group recorded higher amplitudes of electrical signals (Fig. 6L, Supplementary Movie 9). Representative nerves were selected for TEM observation and TB staining of the transverse ultrastructural view four weeks postoperatively. Specimens collected from PNI group (6.26 ± 0.99 μm) showed comparable axon diameter to Piezo+HG group (7.38 ± 1.79 μm) while the latter displayed significantly better myelin sheath thickness (2.52 ± 0.23 μm versus 2.10 ± 0.38 μm) (Fig. 6M-O). Overall, these results demonstrated the therapeutic effect and biosafety of this piezoelectric hydrogel bioelectronics for non-human primate peripheral nerve repair.

A Schematic and surgical images of modeling and piezoelectric hydrogel bioelectronics implantation. (i) Exposure of bifid sciatic nerve. (ii) Establishment of peripheral nerve crush model. The white arrow indicated the injury site. (iii) BaTiO3 piezoelectric hydrogel bioelectronics implantation. The white dashed line indicated wrapped hydrogel. B Representative transverse sections of sciatic nerves were immunohistochemically stained with PIEZO2 polyclonal antibody. Scale bar in low magnification images, 500 μm. Scale bar in higher magnification images, 100 μm. C Quantification of PIEZO2 positive staining contribution. Significant differences were evaluated by two-tailed unpaired Student’s t-test. Data are presented as mean ± SD. Five visual fields in each monkey were randomly selected. D H&E staining of longitudinal sections of injured nerves four weeks after surgery. Scale bar, 200 μm. E Representative longitudinal sections IF staining of vessel marker CD34 (red), growth cones marker growth associated protein 43 (GAP43, pink) and peripheral neurons marker βIII tubulin (green). Scale bar, 400 μm. F Representative transverse sections IF staining of endothelial marker CD31. Scale bar, 200 μm. White arrow denoted vascular lumen-like structure. G Schematic of denervation affecting the corresponding muscles and skin. H Representative H&E staining of harvested gastrocnemius muscles. Scale bar in low magnification images, 200 μm. Scale bar in higher magnification images, 100 μm. I Quantitative analysis of average muscle fiber perimeter and muscle fiber area. J Masson’s trichrome-stained sections of cutaneous innervation. Scale bar, 500 μm. K Images of the electrodes integrated on the peripheral nerve for recording in electrophysiological testing procedure. L Recorded potential during electrical stimulation. M Representative transmission electron microscope images of myelin sheath four weeks after PNI. Scale bar, 10 μm. N Representative toluidine blue staining of myelin sheath four weeks after PNI. Scale bar, 100 μm. O Quantitative analysis for axon diameter and thickness of myelin sheath. Significant differences were evaluated by two-tailed unpaired Student’s t-test; ***p < 0.001. Data are presented as mean ± SD. PNI refers to PNI refers to peripheral nerve injury group without additional treatment, Piezo+HG refers to piezoelectric hydrogel bioelectronics group.
Collectively, piezoelectric hydrogel bioelectronics under external mechanical stimulation effectively enhanced neurogenesis and facilitated angiogenesis in peripheral nerve injury models of rats and rhesus monkeys. Mechanosensitive channel PIEZO2 and Schwann cells were activated after treatment. Desirable biocompatibility showed translational potential of BaTiO3 piezoelectric hydrogel bioelectronics.
Astrocyte-active mechanical neuroengineering for spinal cord repair
In vitro experiments revealed that piezoelectric hydrogel bioelectronics under ultrasound activated the PIEZO1-mediated astrocytic repairing process. We established SCI models in rats and beagle dogs to evaluate this neuromodulation strategy in vivo. In the rat SCI model, we implanted piezoelectric hydrogel bioelectronics into a 2-mm spinal defect gap (Fig. 7A). After closing the wound, ultrasound stimulation was applied (five minutes once a day for a three-week period). The rats in the Piezo+US group exhibited comfort state (Supplementary Movie 10). Compared with the control group receiving sham injury, immunohistochemistry staining was markedly positive for PIEZO1, indicating the activation of this mechanosensitive channel (Fig. 7B, C). To directly measure neuronal calcium channel activity, we separated dorsal root ganglia from the injured core of SCI rats three weeks postoperatively and performed the patch clamp technique (Fig. 7D). The piezoelectric hydrogel bioelectronics, irrespective of ultrasound stimulation, consistently maintained the resting potential of dorsal root ganglion (DRG) at approximately −70 mV. Conversely, the resting potential of cells in both the SCI group and the non-piezoelectric hydrogel bioelectronics group exhibited more positive values (Fig. 7E). This deviation is likely attributable to membrane permeability alteration and dysregulated ion channel activity induced by the injury. We subsequently performed depolarizing stimulation, spanning from −60 mV to +80 mV, and acquired the corresponding voltage-current relationship. The results demonstrated that the Piezo+US group exhibited the highest current density, indicative of significantly elevated electrophysiological activity associated with calcium ion influx through membrane channels (Fig. 7F). This observation aligned with the calcium ion fluorescence data obtained from in vitro experiments.

A Schematic and surgical images of 2-mm spinal cord defect and hydrogel implantation. White and black arrows indicate defect and BaTiO3 hydrogel. (B, C) PIEZO1 IHC staining and quantification. Scale bar, 250 μm in (i) and (ii), 100 μm in (iii) and (iv). d, dorsal. v, vendral. n = 3. D Schematic for whole-mount patch clamp experiments in DRGs. E Analysis of resting membrane potential in DRGs. n = 8 in HG group and n = 7 in the other groups. F Quantification and representative traces of Ca2+ currents. n = 10. G IF staining of GFAP, S100β, Sox9, and Nestin. Scale bar, 500μm. H Statistical analysis of mean fluorescence intensity (%) of GFAP, S100β, Sox9, and Nestin. n = 3 per group. I IF staining of neuronal marker NeuN, sprouted axon marker GAP43, and vessel marker CD34. Scale bar, 1 mm. White circle indicated scar area after injury. J Statistical analysis of mean fluorescence intensity (%) of NeuN, GAP43, and CD34. n = 3 per group. K Representative IF staining of chondroitin sulfate marker CS-56 and glutamine synthetase antibody. Scale bar in left panel, 500 μm. Scale bar in (i) and (ii), 60 μm. L Statistical analysis of mean fluorescence intensity (%) of CS-56 and glutamine. n = 3 per group. M Representative trajectory diagram and analysis of open-field monitoring test. n = 3 per group. Significant differences were assessed by one-way ANOVA with Dunnett’s post hoc test unless otherwise stated (*p < 0.05, **p < 0.01, ***p < 0.001). Data are mean ± SD. Sham sham-operated, SCI spinal cord injury only, HG hydrogel group, Piezo+HG piezoelectric hydrogel group, Piezo+US ultrasound-driven piezoelectric hydrogel group.
Astrocytes are the most prevalent resident cells in the CNS. Following SCI, astrocytes undergo a phenotypic transformation termed reactive astrogliosis62. During this process, reactive astrocytes proliferate and secrete inflammatory cytokines to aid in tissue repair initially. These reactive astrocytes form scar tissue around the lesion core over time and affect SCI prognosis. Excessive astrocytic scars in the injury site at later stages of SCI are recognized as the primary cause of axonal damage. Twelve weeks after surgery, increased GFAP and S100β expression were observed both in RAs and SAs (Fig. 7G, H). Sox9 and Nestin are specific genes of SAs and RAs, respectively62. At twelve weeks post-surgery, the SCI group exhibited the most intense IF staining of GFAP/S100β, characterized by a predominance of Sox9-positive SAs and a minimal presence of Nestin-positive RAs. Conversely, piezoelectric hydrogel bioelectronics under ultrasound stimulation significantly diminished the population of scar-forming astrocytes for favorable regenerative microenvironment. For longitudinal sections, a more axon-conductive “bridge-like” scar structure was found in Piezo+US group while astrocytes and the glial scar in general posed a “wall-like” barrier at the scar border for chronic failure of axon regeneration (Supplementary Fig. 16A, C).
To assess neuronal survival after SCI, we performed IF labeling using NeuN, GAP43, TPH2, PGP9.5, and fibronectin on sagittal sections and Tuj1 on transverse sections. The result showed that piezoelectric hydrogel bioelectronics under ultrasound preserved more axons from the injury and promoted 5-HT neuronal activity, crucial for neural signal conduction and functional recovery after SCI (Fig. 7I, J, Supplementary Fig. 16B–H). Chondroitin sulfate proteoglycan (CSPG) was the predominant extracellular inhibitor of axon regeneration. Its expression was selectively observed in the astrocytic scar region following SCI. To visualize the distribution and expression of CSPG, we employed the CS-56 antibody to label CSPG. Fewer CSPG was observed in Piezo+HG and Piezo+US groups than in the other two groups (Fig. 7K, L). Moreover, glutamine synthesis was activated around the lesion core in Piezo+US group, consistent with in vitro results. SCI also led to lower limb atrophy. Gastrocnemius muscle histological assessment indicated a compact muscle structure with tight fibers in Piezo+US group (Supplementary Fig. 16I, J). For sensory recovery, we carried out hot plate test. Twelve weeks after surgery, the average reaction latency of Piezo+US group (10.83 s) was inferior to that of SCI group (27.88 s) and non-piezoelectric hydrogel bioelectronics group (25.75 s) (Supplementary Fig. 16K). The Basso, Beattie, Bresnahan Locomotor Rating Scale (BBB) Score was used to assess rat motor function recovery following SCI at three, six, and twelve weeks postoperatively. Although BBB scores increased over time in all groups, the Piezo+US group exhibited significantly higher scores than the others, aligning with the treadmill test findings (Supplementary Fig. 16L, Supplementary Movie 11). To evaluate rat emotional state after SCI, we conducted open field test. The number of times a rat crossed grid lines in an enclosed area was an indicator of depressive behaviors. The Piezo+US group exhibited more entries to different grids and more trajectories. It suggested that the treatment maintained a positive emotional state in rats after injury (Fig. 7M). Taken together, these data demonstrated that ultrasound-driven piezoelectric hydrogel bioelectronics promoted rat SCI repair.
Afterwards, we implanted BaTiO3 hydrogel bioelectronics into spinal cord hemisection site of beagle dog to test the translational potential and biosafety (Fig. 8A). Ultrasound stimulation was performed as in the previous procedure (Supplementary Movie 12). At four weeks following surgery, beagle dogs in both groups exhibited full contact of hind limbs with the ground, incapable of supporting the body weight (Fig. 8B, Supplementary Movie 13). By the eighth week postoperatively, the Piezo+US group displayed partial recovery of hind limb muscle strength. However, these dogs still could not fully control the locomotion and stepping frequency (Fig. 8B, Supplementary Movie 14). At twelve weeks post-surgery, Piezo+US group showed marked improvement in hind limb strength and even regained tail wagging activity, whereas SCI group showed no noticeable functional improvement (Fig. 8B, Supplementary Movie 15). Compared with the SCI group, significantly higher PIEZO1 positive immunohistochemistry staining was observed in Piezo+US group three weeks after ultrasound treatment, indicating the activation of PIEZO1 channel (Fig. 8C, D). The biosafety of BaTiO3 hydrogel bioelectronics application in SCI was verified with H&E staining and no notable difference was identified in the main organs, indicating the good biocompatibility of Piezo+US (Fig. 8E, Supplementary Fig. 17). Furthermore, lower limb paralysis after SCI reduced activity and denervation, and led to significant muscle atrophy. We found obvious gastrocnemius muscle fiber atrophy in SCI group with reduced fiber perimeter and area (Fig. 8F, G). Pain, stress, and metabolic changes associated with SCI all accelerated protein breakdown, leading to significant weight loss in animals. However, we demonstrated that piezoelectric hydrogel bioelectronics with ultrasound stimulation considerably improved the Olby Scores at different time points and maintained post-surgery body weight without significant loss (Fig. 8H, I). The injured spinal segments were subjected to MRI examination twelve weeks post-surgery. The results showed that spinal cord continuity in SCI group was still interrupted, while continuity in Piezo+US treated beagle dogs was restored, providing anatomical basis for neural signal transmission (Fig. 8J). Specimens were sectioned and stained for NeuN immunofluorescent to investigate neuronal survival. The mean intensity of NeuN was higher in both transverse and longitudinal sections of Piezo+US group, indicating the neuronal damage were effectively improved (Fig. 8K).

A Schematic and surgical images of ultrasound-driven piezoelectric hydrogel bioelectronics in beagle dog spinal cord injury model. (i) Establishment of 5-mm hemi-transected spinal cord injury model. The white arrow denoted spinal cord hemi-transection injury. (ii) implantation of BaTiO3 hydrogel bioelectronics to the defect. The white lines arrow denoted BaTiO3 hydrogel bioelectronics. B Representative images of hindlimbs recovery at different time points after surgery. C Representative longitudinal sections of spinal cord were immunohistochemically stained with PIEZO1 polyclonal antibody. Scale bar, 100 μm. D Quantification of PIEZO2 positive staining contribution. Significant differences were evaluated by two-tailed unpaired Student’s t-test. n = 3 per group. Data are presented as mean ± SD. E Toxicity evaluation of ultrasound-driven piezoelectric hydrogel bioelectronics in beagle dog spinal cord injury model. H&E staining of heart, liver, spleen, lung, and kidney in different groups. Scale bar, 500 μm. F Representative H&E staining of harvested gastrocnemius muscles. Scale bar, 50 μm. G Analysis of average muscle fiber perimeter and area. Significant differences were evaluated by two-tailed unpaired Student’s t-test.; ***p < 0.001 versus the SCI group. H Gradual recovery of hindlimb movement, strength, and weight-bearing abilities were evaluated using Olby scoring system. n = 3 in each group. I beagle dogs were weighted on the day of surgery and twelve weeks after surgery. n = 3 in each group. Significant differences were evaluated by two-way ANOVA with Turkey’s post-hoc test; **p < 0.01. Data are presented as mean ± SD. J MRI scanning of injured spinal segments twelve weeks after surgery. MRI setting: 0.5 T, SF = 21 MHz, O1 = 232246 Hz, RFA90° = 2.6, RFA180° = 3.9, TR = 300 ms, TE = 20 ms. K Representative IF staining of neuronal marker NeuN (green). Nuclei were counterstained with DAPI (blue). Figures a-d, transverse sections of spinal cord. Figures e, f, longitudinal sections of spinal cord. Scale bar, 500 μm. Yellow dashed lines indicated co-localization analysis of NeuN (green) and DAPI (blue) by ImageJ software. SCI refers to spinal cord injury group without additional treatment and Piezo+US refers to ultrasound-driven piezoelectric hydrogel bioelectronics implantation group.
Overall, in vitro and in vivo experiments suggested that ultrasound-driven piezoelectric hydrogel bioelectronics improved SCI repair in an astrocyte-responsive and PIEZO1 channel activation way. In spinal cord, another mechanosensitive channel PIEZO2 is mainly expressed in neurons such as DRGs72. We also found that PIEZO2 channel involved in SCI mechanotransduction (Supplementary Fig. 18), thereby underlying the intricate mechanism associated with ultrasound-driven piezoelectric materials. Because global knockouts of PIEZO1 or PIEZO2 in mice were lethal, preliminary validation was conducted in heterozygous mice (Supplementary Fig. 19)28,48. Piezoelectric hydrogel bioelectronics was implanted to PIEZO2eKO+/- and WT SCI mice with ultrasound treatment equally. Immunostaining for glutamine and β III-Tubulin showed reduced fluorescent signals in PIEZO2eKO+/- mice. It confirmed knockdown of PIEZO2 also affected the repair function of the ultrasound-driven piezoelectric hydrogel bioelectronics (Supplementary Fig. 20A, B). In addition, PIEZO2 knockdown downregulated serotonin release (Supplementary Fig. 20C). Meanwhile, the NeuN and GAP43 intensity were also reduced in PIEZO2eKO+/- mice while CS56 intensity increased (Supplementary Fig. 20D, E). These results suggested a potential role of the PIEZO channel in the process of mechanical neuroengineering of SCI.
Discussion
Mechanical stimulation, achieved by hardware, soft robots, is increasingly recognized as crucial regulators of cellular behavior (proliferation, differentiation, or phenotype switching), tissue formation and regeneration21,73,74,75. In the context of neural repair, several tissue engineering techniques are used to exert mechanical forces to cells and tissues. Xu et al. proposed multilevel piezoelectric nanofiber conduit with ultrasound-triggered electrical stimulation to accelerate functional recovery and axon regeneration of peripheral nerves36. However, mechanical stimulation (e.g., ultrasonic, magnetic) cannot effectively penetrate the skin and soft tissues to reach and act on the damaged tissues and organs, resulting in suboptimal therapeutic efficacy for relatively deep tissues. Recent research has shifted from using mechanical stimulation directly to focusing on neural repair strategies that employ mechano-bioactive biomaterials to convert external mechanical energy into other forms, like electrical energy76.
Ultrasound-driven piezoelectric bioelectronics are such mechano-bioactive biomaterials and show great potential in regenerative medicine36,42,77,78,79. The present study provided compelling evidence to support the therapeutic potential of piezoelectric hydrogel bioelectronics in neuro-mechanobiology. We demonstrated that ultrasound-driven piezoelectric hydrogel bioelectronics exerted electrical signals from mechanical stimulation, and mediated calcium ion influx primarily via PIEZO1 channel in astrocytes and PIEZO2 channel in Schwann cells. These channels are extensively implicated in mechanotransduction and regulate cellular processes such as calcium influx, cytoskeletal rearrangement, and downstream gene expression, particularly for neuronal survival and axon outgrowth. Piezoelectric materials can be activated through three mechanisms: cellular traction, physiological mechanical movements, and ultrasound, generating electrical stimulation of varying intensities80. The electrical charge generated on piezoelectric material surface under ultrasound can trigger the depolarization of cytomembrane, leading to calcium influx81. Our results indicated that while the piezoelectric hydrogel alone induced mild biological responses, ultrasound activation significantly amplified these effects. Ultrasound-driven mechano-bioactive hydrogel bioelectronics enhanced PIEZO1/2, leading to increased calcium influx and mitochondrial activation. This, in turn, promoted a glial repair phenotype, facilitating neural regeneration and functional improvement in vivo. This suggests that ultrasound was not merely supportive but functionally essential to fully activate the therapeutic potential of mechano-bioactive hydrogel bioelectronics, especially in low-strain tissue environments such as the central and peripheral nerve system.
While PIEZO1/2 were the main mechanotransducers, our study found that VGCCs were also activated by piezoelectric hydrogel+US treatment. Blocking L-type VGCCs with verapamil showed these channels contribute 10% ~ 20% to calcium influx but had little effect on mitochondrial bioenergetics or glial repair compared to PIEZO knockout. PIEZO2 in Schwann cells and PIEZO1 in astrocytes were key in mechanoelectrical conversion, significantly impacting calcium signaling, mitochondrial fusion, phenotypic changes, and neural regeneration. This highlighted that PIEZO channels were central to regenerative processes while VGCCs played an ancillary role in fine-tuning calcium dynamics.
The mechanosensing property and mechanotransduction varied considerably across multiple cells in different tissues82,83,84. This study shed light on the distinct role of glia, specifically Schwann cells and astrocytes, in orchestrating mechanobiological repair processes. Upon BaTiO3 hydrogel bioelectronics and ultrasound treatment, glia exhibited a dynamically shifting biomechanical landscape of complex regulatory network in response to external cues. This study showed that electrical flow from ultrasound-driven piezoelectric bioelectronics upregulated calcium signaling pathway more effectively than ultrasonic stimulation alone, as demonstrated in PIEZO1/2 knockout glia. It indicated that electrical stimulation exerted a more pronounced and predominant role in activating PIEZO channels. These activated mechanosensors (PIEZO1 in astrocytes and PIEZO2 in Schwann cells) initiated intracellular signaling to modulate intracellular functions and drive mitochondrial metabolic reprogramming.
Nerve regeneration orchestrated by astrocytes or Schwann cells is highly energy demanding. Astrocytes or Schwann cells are metabolically demanding cells, particularly in the context of neural injury and repair, where they provide both structural and energetic support to neurons85,86. Mitochondria, the key organelles for cellular energy production, structurally and functionally respond to physical forces and mechanotransduction87,88,89,90. Mechanical cue could be transmitted through PIEZO1-mediated calcium flux and ATP production to regulate organelle function91. This study revealed that mechano-bioactive hydrogel bioelectronics specifically promoted glial mitochondrial fusion in response to electrical stimulation. The ultrasound-driven piezoelectric activation of PIEZO channels (PIEZO1 in astrocytes and PIEZO2 in Schwann cells) induced a substantial intracellular calcium influx. Our findings reveal that the PIEZO–calcium–CREB axis was a pivotal driver of glial activation, orchestrating mitochondrial fusion and neurotrophic support. This pathway likely underpined the regenerative effects of ultrasound-driven mechano-bioactive hydrogel bioelectronics through coordinated regulation of energy metabolism and neuromodulation. Ultrasound-driven mechano-bioactive hydrogel bioelectronics treatment enhanced ATP synthase subunit activity to increase ATP production. Elevated ATP levels not only provided a direct energy supply but also supported the maintenance of mitochondrial membrane potential demonstrated by TOM20 and MitoLite labeling in this study. In this microenvironment, MFN/OPA1-mediated mitochondrial fusion facilitated the formation of a highly efficient mitochondrial network to optimize bioenergetic function and further enhance ATP synthesis and energy production capacities. Mitochondrial adaptations aligned closely with the glial repairing phenotype toward regenerative metabolic programs for neural repair56, including alterations in glutamine metabolism pathways92. We also found upregulation of glutamatergic neurotransmitters (GAD67 and glutamine synthetase) with mitochondria fusion events in ultrasound-driven piezoelectric-activated glia. These findings indicated that mechano-bioactive piezoelectric hydrogel bioelectronics successfully achieved mechanical-electrical-bioenergetic energy conversion for energy-consuming neural repair.
Although this mechano-neuromodulation strategy was focused on glia, it was plausible that other cell types may also be implicated. Astrocytes express PIEZO1 but barely express PIEZO27, consistent with the findings in this study. However, the PIEZO2eKO+/- mice had weakened nerve regenerative ability and neuron activity compared to KOWT mice, suggesting the potential regulation of PIEZO2 on other cell types, such as DRGs. Moreover, immune cells are sensitive to piezoelectric stimulation and play a pivotal role in tissue repair by mediating inflammatory responses and promoting cellular differentiation93,94,95,96. In this study, glia interacted with neighboring cells via cell-cell junctions or modulated cellular responses by recruiting diverse cell populations, such as endothelial cells. The recruitment of these cells was crucial for processes such as angiogenesis, extracellular matrix remodeling, and tissue integration. This underscored the complexity of cellular and tissue-level impacts of ultrasound-activated piezoelectric materials in neural repair.
For in vivo biosafety validation, we assessed histological changes of major organs across four animal models and performed hematological and biochemical analyses. No systemic toxicity or inflammatory responses were observed over periods ranging from 4 to 12 weeks, which correspond to the species-specific time scales of nerve regeneration in different animal models. This design ensured that biosafety assessment was aligned with the critical repair phases of each model, thereby providing stage-appropriate validation of BaTiO₃-based hydrogel bioelectronics. Together with ICP-MS measurements showing minimal Ba²⁺ release and in vitro cytotoxicity assays confirming good tolerance of glial cells, these results collectively support the short- to mid-term biosafety of the material. Nevertheless, long-term stability, potential accumulation, and chronic immune responses remain unresolved and require extended follow-up studies in large animals and standardized toxicological testing to establish the long-term safety and translational reliability of BaTiO₃-based implants.
The limitation of this study is that precise glial subpopulations most sensitive to PIEZO activation were not fully investigated. Definitive identification of responsive glial subtypes requires single-cell RNA sequencing and lineage tracing. In addition, due to current technical constraints, we were unable to non-invasively monitor glial mechanotransduction dynamics across all phases in vivo without altering the mechanical environment. Future advances in real-time, non-destructive imaging or recording technologies will be essential to delineate glial mechanotransduction dynamics. Another future direction is to determine whether mitochondrial bioenergetics provide feedback regulation of PIEZO-mediated mechanosensitivity, because our work mainly focused on how PIEZO activation drives mitochondrial dynamics and bioenergetics.
In conclusion, we developed mechano-bioactive piezoelectric hydrogel bioelectronics to achieve mechanical-electrical-bioenergetic energy conversion for energy-consuming neural repair. Mechanoelectrical conversion-mediated calcium ion influx via PIEZO channels activated intracellular signaling pathways to promote MFN and OPA1-mediated mitochondrial fusion as a central bioenergetic hub. As a result, it enhanced ATP production and formed an efficient energy network to support glia-mediated neural repair. Validated from large animal and non-human primate models, this study highlighted the applicability and biosafety of mechano-bioactive hydrogel bioelectronics for neural regeneration in the central and peripheral nervous systems.
Methods
Solvothermal synthesis of BaTiO3 nanoparticles
Tetragonal BaTiO3 nanoparticles were synthesized using a solvothermal method, starting with Ba(OH)2·H2O (98%, Macklin) and Ti[O(CH2)3CH3]4 (97%, Sigma-Aldrich). For this, 3.4036 g (10 mmol) of Ti[O(CH2)3CH3]4 was dissolved in 4 mL of high-purity ethanol and subsequently mixed with 1.4 mL of NH4OH solution (28%-30% NH3 in H2O, Sigma-Aldrich) under vigorous stirring. Separately, 2.8408 g (15 mmol) of Ba(OH)2·H2O (barium/titanium ratio = 1.5) was dissolved in 5 mL of deionized water and added to the titanium precursor solution. The resulting mixture was transferred into a 50 mL Teflon-lined steel autoclave and heated at 200 °C for 48 h. After the reaction, the product was thoroughly washed with acetic acid, high-purity ethanol, and water, followed by drying at 80 °C. To remove BaO and/or BaCO3, the product was heated in an oven for 2 h, with acetic acid treatment.
ICP-MS analysis of Ba2+ ions
To accurately assess the ionic release profile of Ba²⁺ from BaTiO₃ nanoparticles, we incubated suspensions of 0.1 mg/mL and 0.01 mg/mL BaTiO₃ in PBS (5 mL) at 37 °C. At 1 h, 6 h, 1 day, 3 days, and 7 days, samples were subjected to high-speed centrifugation to ensure complete nanoparticle removal before collecting the supernatants for ICP-MS analysis.
Synthesis of injectable piezoelectric hydrogel bioelectronics
Piezoelectric hydrogel bioelectronics was synthesized as follows (per 1 mL): 530 μL rat tail-derived collagen 1 (Corning Inc., Corning, NY, USA), 100 μL BaTiO3 nanoparticles at different concentrations dissolved in culture medium, 270 μL PBS, 270 μL 0.1 M NaOH. The homogenates were placed on ice and were mixed well by pipetting up and down while trying to avoid generating air bubbles. The mixture was then incubated at 37 °C for 30 minutes to form piezoelectric hydrogel bioelectronics.
Characterization of BaTiO3 nanoparticles
SEM images were captured with a field-emission high-resolution SEM (Zeiss Merlin; 15 kV), and the size of BaTiO3 nanoparticles was quantified using Image J. TEM and HRTEM images were obtained using a field-emission TEM (JEM-2100F, JEOL; 200 kV) with EDS (Be(4)-U(92) and energy resolution better than 127 eV), the measurement and quantification were conducted by RADIUS software. XRD patterns were measured using an X-ray diffractometer (D/max-2550, Rigaku) with Cu Kα radiation (λ = 1.54 Å) at 40 mA and 40 kV. Raman spectra were obtained using a Raman spectrometer (LabRAM HR Evolution, Horiba).
The piezoelectric response test was conducted using PFM (Dimension Icon, Bruker) based on the reverse piezoelectric effect of piezoelectric materials. The potential difference between the voltage-applied probe tip and the surface of the film induced localized piezoelectric displacement on the sample surface. Consequently, the changes in piezoelectric displacement measured under a DC bias reflected the material piezoelectric properties. The DART mode was used to assess the changes in polarization direction at specific nanoparticle locations.
Characterization of BaTiO3@COL-1 hydrogel bioelectronics
The structure of collagen fibers was observed using a reflective model of confocal laser scanning microscope (CLSM, Leica DMI8). COL-1 hydrogel and BaTiO3@COL-1 hydrogel were immersed in 1× PBS and positioned under a 63× oil immersion inverted objective. Reflection microscopy was set up to detect 488-nm light reflected from the sample when illuminated by 488-nm lasers.
The mechanical properties of the BaTiO3@COL-1 hydrogel were tested using a rotational rheometer (Kinexus Pro + , Netzsch, Germany) with a 2 mm rotor. During the test, the sample stage was maintained at 37 °C, and the gap between the two plates was adjusted to 1 mm. A 300 μL sample of the neutralized solution was immediately dispensed onto the preheated rheometer plate. For strain and frequency sweep tests, the sample was allowed to gel for 30 minutes before testing, while time-dependent rheology tests at fixed strain and frequency were conducted without waiting.
Piezoelectric property simulation of BaTiO3 hydrogel bioelectronics by ultrasonic vibration
A 3D model was constructed in Comsol Multiphysics software (version 6.1.0; Comsol Inc., Sweden) to simulate the piezoelectric properties of BaTiO3 nanoparticles under ultrasonic excitation. The simulation domain consisted of a cubic barium titanate particle (side length 100 nm) embedded in a cubic water domain (side length 400 nm). the central cubic domain represented the barium titanate particle with its inherent piezoelectric properties. The ultrasonic wave was applied at the bottom surface of the water domain as a plane wave source, defined as P = P0*sin(2*pi*f0*t). Since the COMSOL database did not include data of BaTiO₃ hydrogels, water was selected as the medium for the simulation. The piezoelectric property of the nanoparticles was investigated by applying continuous ultrasonic vibration to water or and at a frequency of 100 kHz97. A transient study was performed to capture the dynamic response of the system over a time range defined as range (0, T0/8, T0/2). Post-processing of the simulation results included visualization of the electric potential distribution and current density within BaTiO3. The output data were analyzed by plotting 2D surface plots and line graphs at different time points. Figures depicting the spatial distribution of the electric potential and current density were generated to illustrate the transient behavior of the system under ultrasonic excitation.
Cell culture
Rat Schwann cells (cat. #ZQ0154; Zhong Qiao Xin Zhou Biotechnology Co., Ltd, Shanghai, China) were cultured in high glucose Dulbecco’s Modified Eagle Medium (DMEM, cat. #ZQ120; Zhong Qiao Xin Zhou Biotechnology Co., Ltd, Shanghai, China) containing 10% fetal bovine serum and 1% penicillin/streptomycin. Rat primary astrocytes cells (cat. #PRI-RAT-00166; Zhong Qiao Xin Zhou Biotechnology Co., Ltd, Shanghai, China) were cultured in completed astrocyte growth medium (cat. #PCM-R-166; Zhong Qiao Xin Zhou Biotechnology Co., Ltd, Shanghai, China). Rat DRG cells were obtained from Zhong Qiao Xin Zhou Biotechnology Co., Ltd, (cat. # PRI-RAT-00182). Cells were cultured in complete culture medium of rat DRG cells (cat. #PCM-R-182; Zhong Qiao Xin Zhou Biotechnology Co., Ltd, Shanghai, China) on culture plates precoated with polylysine. Human umbilical vein endothelial cells (cat. #ZQ1099; Zhong Qiao Xin Zhou Biotechnology Co., Ltd, Shanghai, China) were cultured in endothelial cell medium (cat. #1001; ScienCell Research Laboratories) containing endothelial cell growth supplement (cat. #1052; ScienCell Research Laboratories) and FBS. Cells were incubated at 37 °C in a humidified incubator with a 5% CO2/95% ambient air atmosphere. Rat Schwann cells and HUVECs from passage numbers 4–10 were used for all experiments. Once confluence reached about 80%, cells were digested and cultured in BaTiO3 piezoelectric hydrogel bioelectronics 3D culture system under ultrasound stimulation. The ultrasound was applied according to previous protocol (setting: five minutes per session each day, power intensity 0.5 W/cm, Sonidel, China)35.
PIEZO2 knockout Schwann cell generation
PIEZO2 knockout RSC96 cell line was generated using the CRISPR/Cas9 system. sgRNA was designed targeting the coding region of exon 21 and exon 22 in gene PIEZO2 (NM_001427318.1) (Supplementary Table 4). Lentiviral vectors were constructed and used to infect cells. Positive mixed clones were counted using a hemocytometer and then serially diluted and seeded into 96-well plates for culture. After 7–10 days, single-cell colonies were observed under a microscope. Based on the size of the single-cell colonies, medium was either replenished or replaced for wells containing single-cell colonies. After 14 days, the cells were collected, and genomic DNA was extracted. PCR amplification was performed, and the PCR products were analyzed via electrophoresis and subsequently verified by sequencing.
PIEZO1 knockout astrocyte generation
PIEZO1 knockout CTX TNA2 cell line was generated using the CRISPR/Cas9 system. sgRNAs specifically designed to target the coding regions of exon 6, exon 7, and exon 8 of the PIEZO1 gene (Supplementary Table 5). These lentiviral constructs were subsequently packaged and utilized to infect CTX TNA2 cell lines. Post-infection, puromycin selection was used to enrich positively infected cells. Subsequently, single-cell clones were isolated, characterized, and expanded, culminating in the establishment of monoclonal cell lines with knockout of the PIEZO1 gene.
L-type calcium channel inhibition
To assess the functional contribution of VGCCs, L-type VGCC blocker verapamil (cat. #HY-14275, MedChemExpress, USA) was applied to evaluated its effect to the neural regeneration with the mechano-bioactive hydrogel bioelectronics. RSC96 cell lines and CTX TNA2 cell lines were cultured in mechano-bioactive hydrogel with or without verapamil treatment (10 μM).
CREB inhibition
To assess the functional contribution of Calcium-CREB signaling, CREB inhibitor KG501 (cat. #HY-103299, MedChemExpress, USA) was applied in hydrogel-embedded RSC96 cell lines and CTX TNA2 cell lines.
Flow cytometric analysis
Schwann cells were treated with the material-conditioned medium for 24 hours. After treatment, cells were digested and collected by centrifugation at 1000 × rpm for 5 minutes according to manufacturer’s protocol (cat. #S0035M; Shanghai Beyotime Biological Co., Ltd.). Cell pellet was then washed twice with PBS to remove residual medium and trypsin. The cells were subsequently incubated with 2’,7’-dichlorofluorescin diacetate (DCFH-DA, 10 μmol/L, Beyotime, China) at 37 °C for 20 minutes in the dark. Flow cytometry analysis was performed using a FACSCanto II flow cytometer (BD Biosciences, USA) to measure the fluorescent intensity of DCF, which reflects intracellular ROS levels. The data were analyzed using FlowJo V10 software (BD Biosciences, USA).
Cell proliferation assay
Cell proliferation was detected using Cell Counting Kit-8 (CCK-8), live/dead assay and Edu assay. For CCK-8 assay, a standard curve was first established which facilitated the comparison of absorbance values obtained from different experimental groups (Supplementary Fig. 3A). Cells were plated at a density of 1.0 × 103 cells per well in 96-well culture plates and incubated for 24 hours at 37 °C in a 5% CO₂ incubator. After incubation, 10 μL of CCK-8 solution (cat. #CK04; Dojindo Molecular Technologies, Inc., Rockville, MD, USA) was added to each well, and the samples were incubated for additional 2 h to allow the formation of the formazan dye. The optimal density was measured at 450 nm using a microplate reader. Cell viability was calculated based on the standard curve generated. The Live/Dead® Viability/Cytotoxicity Kit (cat. #L3224; Invitrogen, Carlsbad, CA, USA) was used to assess cell viability. After co-culturing cells with piezoelectric hydrogel bioelectronics with or without ultrasound stimulation, a staining solution containing 2 μM calcein-AM and 4 μM ethidium homodimer-1 for 20 min at room temperature. Following incubation, the cells were gently washed with PBS to remove excess dye. Fluorescence images were captured using a Leica epifluorescence microscope (Leica Microsystems GmbH, Wetzlar, Germany). Cell proliferation was also examined by Cell-Light EdU Apollo488 In Vitro Kit (cat. # C10310-3; Ribo Biological Co., Ltd., Guangzhou, China)68. Briefly, cell-hydrogel system was seeded onto 24-well plates. At the detection time points, cells were incubated with 200 μL Edu solution for 2 h. Subsequently, cells were fixed using 4% paraformaldehyde (PFA; cat. #G1101, Servicebio) and incubated with 2 mg/mL glycine solution for 5 min with oscillation. The cells were incubated with 200 μL of the penetrant to each well with 10 min of oscillation followed by 200 μL of Apollo staining solution for 30 min. After rinsing with methanol, Hoechst 33342 was used to stain cell nuclei. Immunofluorescence was imaged with a Leica DMI8 fluorescence microscope (Leica, Wetzlar, Germany), and images were acquired for analysis. The intensity of Edu staining was quantified using ImageJ software.
Angiogenesis functional experiment
Tube formation assay and chicken chorioallantoic membrane (CAM) assay was used to determine the role of Schwann cell conditioned medium in angiogenesis. In tube formation assay, HUVECs (104 cells/well) were seeded into µ-Slide Angiogenesis plate (cat. #81506; ibidi LLC, Verona, WI) pre-coated with Matrigel™ Matrix Growth Factor Reduced (cat. #356230; BD Biosciences, Franklin Lakes, NJ, USA). After incubating HUVECs at 37 °C for 6 h, cells were subsequently incubated with calcein-AM (cat. #L3224; Invitrogen, Carlsbad, CA, USA). The pseudo-capillary networks were captured with a Leica DMI8 fluorescence microscope (Leica, Wetzlar, Germany) and analyzed using Angiogenesis Analyzer plugin for ImageJ software98,99. In CAM assay, the fertilized eggs were incubated at 38 ± 0.5 °C with a humidity of 65-70%. After 7 days incubation, a window (approximately 1.5 cm × 1.5 cm) was carefully opened in the shell surface over the air sac using forceps. A sterile silicone ring was placed on the surface of the exposed CAM, and 100 μL of the conditioned medium was added in each group. Axitinib (cat. #AG013736; Selleck) was used as an inhibitor control. After that, the air sac was sealed with a sterile waterproof breathable adhesive patch. After 48 h, the air sac was reopened, and the CAM was observed under a stereomicroscope to evaluate the formation of new blood vessels. Images were captured and analyzed the characteristics (e.g., numbers of nodes and total length).
Cell immunofluorescence
Cells were embedded in piezoelectric hydrogel bioelectronics under ultrasound stimulation on cell climbing slices (14 mm) of 24-well cell culture plates. For cell calcium, cytoskeleton, or superoxide anion imaging, cells were incubated with 0.5 μM Fluo-4 AM solution (cat. #20552; AAT Bioquest, Inc., CA, USA), 100 nM TRITC Phalloidin (cat. #40734ES75; Yeasen Bio Co., Ltd., Shanghai, China) or 0.5 μM dihydroethidium (cat. #S0063; Beyotime Bio Co., Ltd., Shanghai, China) for 30 min. This was subsequently fixed with 4% PFA and DAPI (cat. #D4080; UElandy, Suzhou, China) counterstain. Images were observed using a confocal laser scanning microscope (FV1000, Olympus, Japan). For cell mitochondrial ATP staining, cells were incubated with ATP probe (1 μg/ml, cat. #Mito-AT1.03; Beyotime Bio Co., Ltd., Shanghai, China) for 30 min. This was subsequently fixed with 4% PFA and DAPI (cat. #D4080; UElandy, Suzhou, China) counterstain. Images were observed using fluorescence microscopy. For inner mitochondrial membrane staining, 200 nM MitoLiteTM Red CMXRos (cat. #22698; AAT Bioquest, Inc., CA, USA) staining solution was prepared according to manufacturer’s protocol. Growth media was removed and the staining solution was added to cells. After 30 min incubation, cells were washed gently with PBS and observed using a confocal laser scanning microscope. For outer mitochondrial membrane staining, cells were fixed with 4% PFA, permeabilized with 0.3% Triton X-100 for 15 min, and blocked with 5% BSA. Samples were incubated overnight at 4 °C with TOM20 (cat. #42406; Cell Signaling Technology, CST, Danvers, MA, USA). The next day, cells were washed gently with PBS for three times and incubated with species-appropriate Alexa Fluor® secondary antibodies (Jackson Research) for 1 h at RT. Immunofluorescence was imaged with confocal laser scanning microscope and images were acquired for analysis.
ATP concentration detection
The ATP concentration was detected using the ATP Assay Kit (cat. #S0027; Beyotime Bio Co., Ltd., Shanghai, China). After thorough mixing, cell lysates were centrifuged at 12000 × g for 5 min at 4 °C, and the supernatants were collected for further analysis. For the standard curve, ATP standard solutions were prepared in a series of concentrations (0.01, 0.03, 0.1, 0.3, 1, 3, and 10 μM) using the ATP assay buffer. ATP working solution was prepared according to the manufacturer’s instructions. To measure ATP level, 100 μL of the working solution was added to each well, followed by a 5 min incubation at RT to eliminate background ATP. Luminescence was recorded using a luminometer. Results were expressed as relative fluorescence units (RFU) and ATP concentration was calculated using the standard curve.
Quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA was isolated from cultured cells and homogenized spinal cord using the EZ-press RNA Purification Kit (cat. #B0004D-100; EZBioscience, Roseville, MN, USA). Complementary DNA (cDNA) was synthesized using the cDNA Reverse Transcription Kit (EZBioscience) following the manufacturer’s protocol. Quantitative real-time PCR (qRT-PCR) was conducted using SYBR Green I Master Mix (EZBioscience) on a LightCycler® 480 Real-Time PCR System (Roche, Basel, Switzerland). Gene expression in astrocytes was detected using PCR array by BioTNT© Transitor® qPCR SybrGreen Detection Kit. PCR array data was uploaded to Zendo in an excel file. Primer sequences used for qRT-PCR are listed in the Supplementary Table 6. GAPDH was used as the reference gene for relative quantification100.
Western blotting
Proteins from the cell membrane, cytoplasm, and mitochondria were extracted separately using corresponding extraction kits (cat. #P0033, #C3601; Beyotime Bio Co., Ltd., Shanghai, China). The protein supernatants were boiled with SDS loading buffer, separated by SDS-PAGE (Epizyme Biomedical Technology Co., Ltd, Shanghai, China), and transferred to PVDF membranes (Millipore, MA, USA). The membranes were blocked with 7% milk and incubated with primary antibodies overnight at 4 °C. Subsequently, the membrane was incubated with secondary antibodies and detected using a ChemiDoc Imaging System (#JY-MINI610, JY-Clear ECL, China).
The following primary antibodies were used in this study: PIEZO1 (Rabbit polyclonal to PIEZO1, cat. #PA5-106296, 1:1000, Invitrogen, the antibody was validated by the company refer to the website https://www.thermofisher.cn/cn/zh/antibody/product/PIEZO1-Antibody-Polyclonal/PA5-106296), PIEZO2 (Rabbit polyclonal to PIEZO2, cat. #PA5-72976, 1:1000, Invitrogen, the antibody was validated by the company refer to the website https://www.thermofisher.cn/cn/zh/antibody/product/PIEZO2-Antibody-Polyclonal/PA5-72976), Calmodulin (Rabbit monoclonal to Calmodulin, cat. #ab45689, 1:2000, Abcam, the antibody was validated by the company refer to the website https://www.abcam.cn/products/primary-antibodies/calmodulin-1-2-3-antibody-ep799y-c-terminal-ab45689), cAMP (Rabbit polyclonal to cAMP, cat. #A15653, 1:1000, ABclonol, the antibody was validated by the company refer to the website https://abclonal.com.cn/catalog/A15653), JNK (Rabbit monoclonal to JNK, cat. #A4867, 1:1000, ABclonol, the antibody was validated by the company refer to the website https://abclonal.com.cn/catalog/A4867), GLUT1 (Rabbit monoclonal to GLUT1, cat. #73015S, 1:2000, CST, the antibody was validated by the company refer to the website https://www.cellsignal.cn/products/primary-antibodies/glut1-e4s6i-rabbit-mab/73015), Glutamine Synthetase (Rabbit polyclonal to Glutamine Synthetase, cat. #ab73593, 1:1000, Abcam, the antibody was validated by the company refer to the website https://www.abcam.cn/products/primary-antibodies/glutamine-synthetase-antibody-ab73593), ATP synthase subunit β (Mouse monoclonal to ATP synthase subunit β, cat. #A-21351, 1:1000, Invitrogen, the antibody was validated by the company refer to the website https://www.thermofisher.cn/cn/zh/antibody/product/ATP-Synthase-beta-Antibody-clone-3D5AB1-Monoclonal/A-21351), GAD67 (Rabbit polyclonal to GAD67, cat. #A2938, 1:1000, ABclonol, the antibody was validated by the company refer to the website https://abclonal.com.cn/catalog/A2938), OPA1 (Rabbit monoclonal to OPA1, cat. #80471, 1:1000, CST, the antibody was validated by the company refer to the website https://www.cellsignal.cn/products/primary-antibodies/opa1-d6u6n-rabbit-mab/80471), MFN1 (Rabbit monoclonal to Mitofusin-1, cat. #14739, 1:1000, CST, the antibody was validated by the company refer to the website https://www.cellsignal.cn/products/primary-antibodies/mitofusin-1-d6e2s-rabbit-mab/14739), MFN2 (Rabbit monoclonal to Mitofusin-2, cat. #11925, 1:1000, CST, the antibody was validated by the company refer to the website https://www.cellsignal.cn/products/primary-antibodies/mitofusin-2-d1e9-rabbit-mab/11925), pDRP1 (Rabbit monoclonal to Phospho-DRP1, cat. #6319, 1:1000, CST, the antibody was validated by the company refer to the website https://www.cellsignal.cn/products/primary-antibodies/phospho-drp1-ser637-d3a4-rabbit-mab/6319), MFF (Rabbit monoclonal to MFF, cat. #84580, 1:1000, CST, the antibody was validated by the company refer to the website https://www.cellsignal.cn/products/primary-antibodies/mff-e5w4m-xp-rabbit-mab/84580), β-actin (Mouse monoclonal to β-actin, cat. #Ab8226, 1:10000, Abcam, the antibody was validated by the company refer to the website https://www.abcam.cn/products/primary-antibodies/beta-actin-antibody-mabcam-8226-loading-control-ab8226) was used as the cell membrane and cytoplasm protein loading control. COX IV (Rabbit monoclonal to COX IV, cat. #A11631, 1:10000, ABclonol, the antibody was validated by the company refer to the website https://abclonal.com.cn/catalog/A11631) was used as mitochondria protein loading control.
RNA sequencing
Total RNA was extracted from Schwann cells or astrocytes using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). RNA quality was assessed with an Agilent 5300 Bioanalyzer, and concentrations were measured using a NanoDrop ND-2000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). RNA purification, reverse transcription, library preparation, and sequencing were conducted by Shanghai Biotechnology Co., Ltd. (Shanghai, China) following the manufacturer’s protocols. Differentially expressed genes (DEGs) were identified using DESeq2 analysis with a significance threshold of p < 0.05 and |log2FC | ≥ 1. The RNA-Seq data analysis was performed on the Cloud Platform (http://tools.shbio.com/cloud-tool/drawing-tool/list). Raw RNA-Seq data have been deposited in the GenBank GEO database.
Transcription factors (TF) prediction
To predict upstream transcriptional regulators of PIEZO1 and PIEZO2, 2 kb promoter sequences upstream of the transcription start sites was extracted. TF binding sites within these regions were predicted using the JASPAR database, and TFs with a relative binding score above the defined threshold were selected. In parallel, TFs with ChIP-seq peaks located in the promoter regions were obtained from the GTRD database. The intersection of these datasets was considered as the putative upstream TFs for each gene101. TFs prediction of CREB1 was listed in Supplementary Table 7.
Animal surgery
The mouse and rat surgery were performed in accordance with the guidelines approved by the Animal Ethics Committee of Shanghai Laboratory Animal Research Center (Ethics No. PA2023002021). The beagle dog SCI model and postoperative care were approved by the SJTU Education Service Industry Investment Management (Group) Co., Ltd. (Ethics No. JDLL20231125). The rhesus monkey sciatic nerve crush model and postoperative care were approved by the Animal Welfare Ethics Committee of Kunming Medical University (Ethics No. kmmu20231567). Every animal experiment has been carried out following a protocol approved by an ethical commission. Sample size in each experiment was illustrated in corresponding figure legends.
For the sciatic nerve crush injury model68,102, Sprague-Dawley rats were anesthetized with Zoletil and xylazine hydrochloride (8:1, 0.1 ml/kg, i.m.). As a prophylactic antibiotic treatment, penicillin was injected for three consecutive days after surgery (0.08 ml/kg, i.m, once a day). Rhesus monkeys were anesthetized with atropine (0.03-0.05 mg/kg, i.m.), and then injected with ketamine (40 mg/kg, i.m.). Cephalosporin was injected for five consecutive days after surgery (25 mg/kg/day, i.m, once a day). All surgical procedures were conducted under aseptic conditions. Sprague-Dawley rats or rhesus monkeys underwent shaving procedures before exposure to the right sciatic nerves through dissection of the gluteal muscle. Nerve crush injuries were compressed using straight hemostat. We implanted 1.5 mL of different solid hydrogels into rat injury site (Fig. 5A) and 15 mL of piezoelectric hydrogel bioelectronics into rhesus monkey injury site (Fig. 6A). Subsequently, the muscles and skin were sutured. Amoxicillin was administered in the drinking water for one week after surgery.
For SCI model, Sprague-Dawley rats were anesthetized as described. Beagle dogs were anesthetized with Zoletil and xylazine hydrochloride (8:1, 0.8 ml/kg, i.m.). As a prophylactic antibiotic treatment, penicillin was injected for three consecutive days after surgery (0.06 ml/kg, i.m, once a day). To prevent spinal cord–injured beagle dogs from biting their wounds, protective collars were used. Surgical sites of female Sprague-Dawley rats and beagle dogs were shaved and sterilized, and then a midline incision was made on the animal back at the level of T8-T10 vertebra. The surrounding muscles were carefully retracted to expose the spinal cord by opening the vertebral plate. A 2-mm spinal defect gap was made in rat spinal cord with subsequent 0.3 mL of different solid hydrogels implantation (Fig. 7A). Hemisection SCI model was established in beagle dog with subsequent 10 mL of different solid hydrogels implantation (Fig. 8A). After closing the wound, ultrasound stimulation was applied (five minutes once a day for three-week period). Amoxicillin was administered in the drinking water for one week after surgery. Postoperative assisted urination was performed for three weeks.
PIEZO2 Cas-9-KO+/- mice were purchased from Shanghai Biomodel Organism Science & Technology Development Co.,Ltd. (Ensembl ID: ENSMUSG00000041482). PIEZO2 deletion was confirmed by recombinant PCR (Supplementary Fig. 19). Mice were maintained under specific pathogen–free conditions. Animals were housed in standard polycarbonate cages (3–5 mice per cage) with ad libitum access to food and water. The housing environment was maintained at a constant temperature of 22 ± 2 °C and relative humidity of 50 ± 10 %, under a 12 h light/dark cycle.
Electrophysiology assessment
To assess the electrophysiological properties of rats and rhesus monkeys at four weeks post-surgery, the sciatic nerves on the experimental side were surgically re-exposed under anesthesia. Bipolar hooked stimulating electrodes were then applied to the proximal end of the sciatic nerve trunk. NCV and CMAP during electrical stimulation were recorded.
Whole-cell patch clamp
DRGs were isolated from SCI rats three weeks after surgery and resuspended in neurobasal medium supplemented with 2% B27, 50 ng/mL NGF, and 2 mM L-glutamine. After filtration through a 40 µm cell strainer, the single-cell suspension was seeded onto culture dishes and incubated at 37 °C with 5% CO2. The extracellular solution for Ca²⁺ current recording contained 140 mM TEA-Cl, 4 mM KCl, 1 mM MgCl2, 10 mM HEPES, 10 mM CaCl2, and 5 mM D-glucose (pH 7.4). The intracellular solution consisted of 110 mM CsCl, 1 mM CaCl2, 5 mM HEPES, 10 mM EGTA, 4 mM Na2-ATP, and 4.5 mM Mg-ATP (pH 7.2). The glass pipettes were pulled using a Sutter P-1000 puller. Under a 40x objective lens, pipettes were positioned near the cell surface. Gentle suction was applied to form a giga-ohm seal, followed by further suction to establish the whole-cell configuration. After that, cells were held at −60 mV and depolarized to +80 mV for 500 ms. For action potential stimulation, cells were held at −80 mV, and current steps (10 pA increments from −50 pA to +140 pA, 1000 ms duration) were applied.
TEM detection
At the indicated times, animals were sacrificed. The regenerative nerves were fixed in 2.5 % glutaraldehyde, embedded in EPONTM resin, sectioned at 50 nm, and stained with OsO4/uranyl acetate for contrast enhancement before TEM analysis. Sections were captured by TEM (HITACHI, HT7800 Exalens) with an accelerating voltage of 80 kV. ImageJ was utilized to assess myelination morphology, specifically the average myelinated axon diameter and myelin sheath thickness.
Histological analyses
The regenerated nerve, muscle, skin, and major organ samples of rats, beagle dogs, and rhesus monkeys were submitted to H&E staining, Masson staining, Sirius red staining, toluidine blue staining, immunohistochemistry staining, and multiple labeling IF staining. For paraffin sections of fresh tissue samples, target tissues were harvested, fixed in 4% PFA overnight, cleared in xylene, embedded in paraffin, and sectioned. For frozen sections, tissues were fixed in 4% PFA, passed through a sucrose gradient, frozen, and sectioned on a freezing microtome. The sections of H&E staining, Masson staining, and toluidine blue images were captured under optical microscope. The Sirius red staining was observed under a polarized light microscope. Immunohistochemical staining was performed using primary antibodies against PIEZO1 and PIEZO2, followed by species-appropriate HRP-conjugated secondary antibody. Positive staining contribution was measured using ImageJ software. Multiple labeling IF staining was performed according to the previous protocol. Briefly, histological sections were incubated overnight at 4 °C with the first primary antibody. After thorough washing, sections were treated with an HRP-conjugated secondary antibody (Abcam) for 50 minutes, followed by a 10-minute incubation with TSA working solution (World Advanced Science Co., Ltd, Shanghai, China). Microwave heating was used to strip off the bound antibodies, allowing for subsequent rounds of staining with additional primary antibodies. This process was repeated for the second and third primary antibodies as needed. Cell nuclei were counterstained with DAPI. Fluorescence images were captured using a Leica DMI8 fluorescence microscope (Leica, Wetzlar, Germany). Due to species specificity, the immunofluorescence and immunohistochemistry antibodies used for beagle dogs and rhesus monkeys have not been validated by the respective companies.
MRI scanning
At the indicated times, beagle dogs were sacrificed and the injured spinal segments were subjected to MRI scanning using small animal in vivo imaging system (MesoMR23-060-I, Shanghai Electronic Technology Co., Ltd, Shanghai, China). The resonance frequency was 21 MHz and the magnetic field strength was 0.5 T. The coil diameter was set at 60 mm, and the magnet was maintained at a constant temperature of 32 °C. Imaging parameters were as follows: SF = 21 MHz, O1 = 232246 Hz, RFA90° = 2.6, RFA180° = 3.9, TR = 300 ms, TE = 20 ms, Slice width (mm) =58, Slices=1, Average=4, Read Size=256, Phase Size=256.
Barium ions detection in rat samples
To assess the in vivo concentration of barium ions following material implantation, muscle tissues adjacent to the implantation site were collected one month post-surgery. The analysis was conducted by Mérieux NutriScience Testing Technology (Ningbo) Co., Ltd. using Method I (Inductively Coupled Plasma Mass Spectrometry) in accordance with the National Food Safety Standard of the PR China, GB 5413.21-2010. Blood samples were determined using ICP Mass Spectrometer (NexION 5000 G, PerkinElmer) in accordance with the Association Standard (Determination of multiple elements in whole blood Inductively coupled plasma mass spectrometry), T/STEI 0003-2024.
Whole blood test
At the indicated times, blood harvested from rhesus monkeys was centrifuged to obtain serum samples. An automatic biochemical analyzer (CCIC Medical Diagnostic Testing Center, Shanghai, China) was used to perform blood biochemical analysis.
Behavioral experiment
Animal behavioral experiments were conducted utilizing a series of established methodologies, including the hot plate test, treadmill running exercise, tail suspension test, and open field test. In the hot plate test, rats were positioned within a cylindrical enclosure on a heated metal plate maintained at 55 °C. The response latency was measured as the time elapsed before the rats exhibited paw licking or attempted to jump off the plate. The treadmill running exercise was performed with the following parameters: an electrical current of 0.8 mA, a speed of 10 meters per minute, and an acceleration of 6 meters per second squared. In the tail suspension test, rats were suspended in an enclosure for a duration of 5 minutes, during which the duration of mobility, characterized by body movement, was documented. A reduction in immobility time was interpreted as indicative of antidepressant activity. Additionally, the open field test was conducted within a square arena, where rats were permitted to explore for five minutes. Throughout this period, their movement trajectories and the frequency of entries into distinct grid sections were recorded using a video camera positioned on the ceiling of the enclosure. The overall activity of mice in an open field area was monitored twelve weeks after surgery. The first 1 min of the test were analyzed assess differences in the number of entries to different grids.
Image Copyright
We owned copyright for the figures used in the manuscript. Platform/software were used as followed:
Adobe Illustrator: Fig. 3a, d, e, 1.
Adobe Photoshop: Figs. 5a, 6a, 7a, d, 8a.
Figdraw platform (https://www.figdraw.com): Fig. 6g, Fig. S13-d, Fig. S20-e.
Statistical analyses
All quantitative data are presented as means ± standard deviation (SD). SPSS 22.0 (IBM Corp. Released 2013. IBM SPSS Statistics for Windows, Version 22.0. Armonk, NY: IBM Corp.) was used for statistical analyses. One-way analysis of variance (ANOVA) followed by Dunnett’s test was used for multiple group comparisons, and two-tailed Student’s t-test was used for comparisons between two groups, as indicated in corresponding figure legends. All experiments were repeated at least three times to support reproducibility. p-value < 0.05 was considered statistically significant. The P value level is indicated by asterisks in the figure legends (*, **, *** signifying P value < 0.05, <0.01, <0.001, respectively).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Raw data generated in this study have been deposited in the Zenodo database under accession code ZZ (10.5281/zenodo.17350627). Raw data of RNA-seq have been deposited in the NCBI BioProject database (http://www.ncbi.nlm.nih.gov/bioproject/1334459). All data supporting the findings of this study are available within the article and its supplementary files. Any additional requests for information can be directed to, and will be fulfilled by, the corresponding authors. Source data are provided with this paper.
References
Varadarajan, S. G., Hunyara, J. L., Hamilton, N. R., Kolodkin, A. L. & Huberman, A. D. Central nervous system regeneration. Cell 185, 77–94 (2022).
Serger, E. et al. The gut metabolite indole-3 propionate promotes nerve regeneration and repair. Nature 607, 585–592 (2022).
Lein, E. S. et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature 445, 168–176 (2007).
Li, F. et al. Glial metabolic rewiring promotes axon regeneration and functional recovery in the central nervous system. Cell Metab. 32, 767–785 e767 (2020).
Babetto, E., Wong, K. M. & Beirowski, B. A glycolytic shift in Schwann cells supports injured axons. Nat. Neurosci. 23, 1215–1228 (2020).
Cameron, E. G. et al. A molecular switch for neuroprotective astrocyte reactivity. Nature 626, 574–582 (2024).
Chi, S. et al. Astrocytic Piezo1-mediated mechanotransduction determines adult neurogenesis and cognitive functions. Neuron 110, 2984–2999 e2988 (2022).
Kantarci, H. et al. Schwann cell-secreted PGE(2) promotes sensory neuron excitability during development. Cell 187, 4690–4712 e4630 (2024).
Kasuba, K. C. et al. Mechanical stimulation and electrophysiological monitoring at subcellular resolution reveals differential mechanosensation of neurons within networks. Nat. Nanotechnol. 19, 825–833 (2024).
Vining, K. H. & Mooney, D. J. Mechanical forces direct stem cell behaviour in development and regeneration. Nat. Rev. Mol. Cell Biol. 18, 728–742 (2017).
Cao, R., Tian, H., Tian, Y. & Fu, X. A hierarchical mechanotransduction system: from macro to micro. Adv. Sci. (Weinh.) 11, e2302327 (2024).
Franze, K. Integrating chemistry and mechanics: the forces driving axon growth. Annu Rev. Cell Dev. Biol. 36, 61–83 (2020).
Cadoni, S. et al. Ectopic expression of a mechanosensitive channel confers spatiotemporal resolution to ultrasound stimulations of neurons for visual restoration. Nat. Nanotechnol. 18, 667–676 (2023).
Duque, M. et al. Sonogenetic control of mammalian cells using exogenous Transient Receptor Potential A1 channels. Nat. Commun. 13, 600 (2022).
Lorach, H. et al. Walking naturally after spinal cord injury using a brain-spine interface. Nature 618, 126–133 (2023).
Ino, D. et al. Neuronal Regulation of Schwann Cell Mitochondrial Ca Signaling during Myelination. Cell Rep. 12, 1951–1959 (2015).
Wang, S.T. et al. A Mitochondrion-Targeting Piezoelectric Nanosystem for the Treatment of Erectile Dysfunction via Autophagy Regulation. Adv Mater 37, 10.1002/adma.202413287 (2025).
Iskratsch, T., Wolfenson, H. & Sheetz, M. P. Appreciating force and shape-the rise of mechanotransduction in cell biology. Nat. Rev. Mol. Cell Biol. 15, 825–833 (2014).
Kim, T. et al. Deep brain stimulation by blood-brain-barrier-crossing piezoelectric nanoparticles generating current and nitric oxide under focused ultrasound. Nat. Biomed. Eng. 7, 149–163 (2023).
Xiao, B. Mechanisms of mechanotransduction and physiological roles of PIEZO channels. Nat. Rev. Mol. Cell Biol. 25, 886–903 (2024).
Zhang, Y. & Habibovic, P. Delivering mechanical stimulation to cells: state of the art in materials and devices design. Adv. Mater. 34, e2110267 (2022).
Pillai, E. K. & Franze, K. Mechanics in the nervous system: From development to disease. Neuron 112, 342–361 (2024).
Delmas, P. & Coste, B. Mechano-gated ion channels in sensory systems. Cell 155, 278–284 (2013).
Chalfie, M. Neurosensory mechanotransduction. Nat. Rev. Mol. Cell Biol. 10, 44–52 (2009).
Parpaite, T. & Coste, B. Piezo channels. Curr. Biol. 27, R250–R252 (2017).
Murthy, S. E., Dubin, A. E. & Patapoutian, A. Piezos thrive under pressure: mechanically activated ion channels in health and disease. Nat. Rev. Mol. Cell Biol. 18, 771–783 (2017).
Saotome, K. et al. Structure of the mechanically activated ion channel Piezo1. Nature 554, 481–486 (2018).
Delmas, P., Parpaite, T. & Coste, B. PIEZO channels and newcomers in the mammalian mechanosensitive ion channel family. Neuron 110, 2713–2727 (2022).
Ma, S. et al. A role of PIEZO1 in iron metabolism in mice and humans. Cell 184, 969–982 e913 (2021).
Hill, R. Z., Loud, M. C., Dubin, A. E., Peet, B. & Patapoutian, A. PIEZO1 transduces mechanical itch in mice. Nature 607, 104–110 (2022).
Zeng, W. Z. et al. PIEZOs mediate neuronal sensing of blood pressure and the baroreceptor reflex. Science 362, 464–467 (2018).
Velasco-Estevez, M. et al. Inhibition of Piezo1 attenuates demyelination in the central nervous system. Glia 68, 356–375 (2020).
Abdo, H. et al. Specialized cutaneous Schwann cells initiate pain sensation. Science 365, 695–699 (2019).
Zhang, Y. et al. Magnetoelectric nanoparticles incorporated biomimetic matrix for wireless electrical stimulation and nerve regeneration. Adv. Health. Mater. 10, e2100695 (2021).
Qian, Y. et al. Mechano-informed biomimetic polymer scaffolds by incorporating self-powered zinc oxide nanogenerators enhance motor recovery and neural function. Small 16, e2000796 (2020).
Xu, D. et al. Ultrasound-responsive aligned piezoelectric nanofibers derived hydrogel conduits for peripheral nerve regeneration. Adv. Mater. 36, e2307896 (2024).
Lee, H. W., Moon, S., Choi, C. H. & Kim, D. K. Synthesis and size control of tetragonal barium titanate nanopowders by facile solvothermal method. J. Am. Ceram. Soc. 95, 2429–2434 (2012).
Didomenico, M., Wemple, S. H. & Porto, S. P. S. Raman spectrum of single-domain batio 3. Phys. Rev. 174, 522-+ (1968).
Meizlish, M. L. et al. Mechanosensing regulates tissue repair program in macrophages. Sci. Adv. 10, eadk6906 (2024).
Ricotti, L. et al. Ultrasound stimulation of piezoelectric nanocomposite hydrogels boosts chondrogenic differentiation in vitro, in both a normal and inflammatory milieu. Acs Nano 18, 2047–2065 (2024).
Sood, A. et al. A comprehensive review on barium titanate nanoparticles as a persuasive piezoelectric material for biomedical applications: prospects and challenges. Small 19, 10.1002/smll.202206401 (2023).
Vinikoor, T. et al. Injectable and biodegradable piezoelectric hydrogel for osteoarthritis treatment. Nat. Commun. 14, 6257 (2023).
Wang, A., Hu, M., Zhou, L. & Qiang, X. Self-powered well-aligned p(vdf-trfe) piezoelectric nanofiber nanogenerator for modulating an exact electrical stimulation and enhancing the proliferation of preosteoblasts. Nanomaterials (Basel) 9, 10.3390/nano9030349 (2019).
Marino, A. et al. Piezoelectric Nanoparticle-Assisted Wireless Neuronal Stimulation. ACS Nano 9, 7678–7689 (2015).
Few, A. P. et al. Asynchronous Ca2+ current conducted by voltage-gated Ca2+ (CaV)-2.1 and CaV2.2 channels and its implications for asynchronous neurotransmitter release. Proc. Natl Acad. Sci. USA 109, E452–E460 (2012).
Ronzitti, G. et al. Exogenous alpha-synuclein decreases raft partitioning of Cav2.2 channels inducing dopamine release. J. Neurosci. 34, 10603–10615 (2014).
Zhang, J. et al. Presynaptic Excitation via GABAB Receptors in Habenula Cholinergic Neurons Regulates Fear Memory Expression. Cell 166, 716–728 (2016).
Li, J. et al. Piezo1 integration of vascular architecture with physiological force. Nature 515, 279–282 (2014).
Yao, X.Y. et al. Metal organic framework-modified bioadaptable implant potentiates the reconstruction of nerve microenvironment immunometabolism reprogramming. Nano Today 49, https://doi.org/10.1016/j.nantod.2023.101814 (2023).
Marcelo, K. L., Means, A. R. & York, B. The Ca(2+)/Calmodulin/CaMKK2 Axis: Nature’s Metabolic CaMshaft. Trends Endocrinol. Metab. 27, 706–718 (2016).
Arnsten, A. F., Wang, M. J. & Paspalas, C. D. Neuromodulation of thought: flexibilities and vulnerabilities in prefrontal cortical network synapses. Neuron 76, 223–239 (2012).
Heredia, D. J., De Angeli, C., Fedi, C. & Gould, T. W. Calcium Signaling in Schwann cells. Neurosci. Lett. 729, 134959 (2020).
Rousse, I. & Robitaille, R. Calcium signaling in Schwann cells at synaptic and extra-synaptic sites: active glial modulation of neuronal activity. Glia 54, 691–699 (2006).
Shen, J. et al. In Situ Prevascularization Strategy with Three-Dimensional Porous Conduits for Neural Tissue Engineering. ACS Appl Mater. Interfaces 13, 50785–50801 (2021).
Spinelli, J. B. & Haigis, M. C. The multifaceted contributions of mitochondria to cellular metabolism. Nat. Cell Biol. 20, 745–754 (2018).
Della-Flora Nunes, G. et al. Prohibitin 1 is essential to preserve mitochondria and myelin integrity in Schwann cells. Nat. Commun. 12, 3285 (2021).
Lee, W. C., Guntur, A. R., Long, F. & Rosen, C. J. Energy metabolism of the osteoblast: implications for osteoporosis. Endocr. Rev. 38, 255–266 (2017).
Miller, K. E., Richards, B. A. & Kriebel, R. M. Glutamine-, glutamine synthetase-, glutamate dehydrogenase- and pyruvate carboxylase-immunoreactivities in the rat dorsal root ganglion and peripheral nerve. Brain Res. 945, 202–211 (2002).
Pareek, G., Thomas, R. E., Vincow, E. S., Morris, D. R. & Pallanck, L. J. Lon protease inactivation in Drosophila causes unfolded protein stress and inhibition of mitochondrial translation. Cell Death Discov. 4, 51 (2018).
Timper, K. et al. GLP-1 Receptor Signaling in Astrocytes Regulates Fatty Acid Oxidation, Mitochondrial Integrity, and Function. Cell Metab. 31, 1189–1205 e1113 (2020).
Saitoh, F. & Araki, T. Proteasomal degradation of glutamine synthetase regulates schwann cell differentiation. J. Neurosci. 30, 1204–1212 (2010).
Hara, M. et al. Interaction of reactive astrocytes with type I collagen induces astrocytic scar formation through the integrin-N-cadherin pathway after spinal cord injury. Nat. Med 23, 818–828 (2017).
Endo, F. et al. Molecular basis of astrocyte diversity and morphology across the CNS in health and disease. Science 378, eadc9020 (2022).
Khakh, B. S. & Deneen, B. The emerging nature of astrocyte diversity. Annu Rev. Neurosci. 42, 187–207 (2019).
Zhang, B. H. et al. B cell-derived GABA elicits IL-10<SUP>+</SUP> macrophages to limit anti-tumour immunity. Nature 599, 471−+ (2021).
Abbott, N. J., Ronnback, L. & Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 7, 41–53 (2006).
de Ceglia, R. et al. Specialized astrocytes mediate glutamatergic gliotransmission in the CNS. Nature 622, 120–129 (2023).
Kong, L. et al. Multilevel neurium-mimetic individualized graft via additive manufacturing for efficient tissue repair. Nat. Commun. 15, 6428 (2024).
Mao, Z., Zheng, Y. L. & Zhang, Y. Q. Behavioral impairment and oxidative damage induced by chronic application of nonylphenol. Int J. Mol. Sci. 12, 114–127 (2010).
Wilkowska, A., Wiglusz, M.S., Jakuszkowiak-Wojten, K. & Cubala, W.J. Ketamine and Lamotrigine Combination in Psychopharmacology: Systematic Review. Cells 11, 10.3390/cells11040645 (2022).
Ranade, S. S. et al. Piezo2 is the major transducer of mechanical forces for touch sensation in mice. Nature 516, 121–125 (2014).
Ray, K. A prime role for PIEZO2 in DRG neurons in mechanosensation in the gut. Nat. Rev. Gastro Hepat. 20, 693–693 (2023).
Douguet, D. & Honore, E. Mammalian mechanoelectrical transduction: structure and function of force-gated ion channels. Cell 179, 340–354 (2019).
McMurray, R. J. et al. Nanoscale surfaces for the long-term maintenance of mesenchymal stem cell phenotype and multipotency. Nat. Mater. 10, 637–644 (2011).
Monge, C. et al. Rigidity-patterned polyelectrolyte films to control myoblast cell adhesion and spatial organization. Adv. Funct. Mater. 23, 3432–3442 (2013).
Niu, Q. et al. Fully biodegradable ion-induced silk fibroin-based triboelectric nanogenerators with enhanced performance prevent muscle atrophy. Biomaterials 318, 123185 (2025).
Chen, S. et al. Piezocatalytic medicine: an emerging frontier using piezoelectric materials for biomedical applications. Adv. Mater. 35, e2208256 (2023).
Wang, Z. et al. Stretchable Unsymmetrical Piezoelectric BaTiO Composite Hydrogel for Triboelectric Nanogenerators and Multimodal Sensors. Acs Nano 16, 1661–1670 (2022).
Wu, P. et al. The marriage of immunomodulatory, angiogenic, and osteogenic capabilities in a piezoelectric hydrogel tissue engineering scaffold for military medicine. Mil. Med Res. 10, 35 (2023).
Wang, X., Stefanello, S.T., Shahin, V. & Qian, Y. From mechanoelectric conversion to tissue regeneration: translational progress in piezoelectric materials. Adv Mater, e2417564, 10.1002/adma.202417564 (2025).
Ni, X. et al. Piezoelectric Biomaterials for Bone Regeneration: Roadmap from Dipole to Osteogenesis. Adv. Sci. (Weinh), e14969, 10.1002/advs.202414969 (2025).
Li, Y.W., Wong, I.Y. & Guo, M. Reciprocity of cell mechanics with extracellular stimuli: emerging opportunities for translational medicine. Small 18, 10.1002/smll.202107305 (2022).
Petroll, W.M., Varner, V.D. & Schmidtke, D.W. Keratocyte mechanobiology. Exp. Eye Res. 200, 10.1016/j.exer.2020.108228 (2020).
Baghdadi, M. B. et al. PIEZO-dependent mechanosensing is essential for intestinal stem cell fate decision and maintenance. Science 386, eadj7615 (2024).
Sundaram, V. K. et al. Adipo-glial signaling mediates metabolic adaptation in peripheral nerve regeneration. Cell Metab. 35, 2136–2152 e2139 (2023).
Ruan, Y. et al. Chronic stress hinders sensory axon regeneration via impairing mitochondrial cristae and OXPHOS. Sci. Adv. 9, eadh0183 (2023).
Moore, A. S., Wong, Y. C., Simpson, C. L. & Holzbaur, E. L. Dynamic actin cycling through mitochondrial subpopulations locally regulates the fission-fusion balance within mitochondrial networks. Nat. Commun. 7, 12886 (2016).
Helle, S.C.J. et al. Mechanical force induces mitochondrial fission. Elife 6, 10.7554/eLife.30292 (2017).
Bartolak-Suki, E. & Suki, B. Tuning mitochondrial structure and function to criticality by fluctuation-driven mechanotransduction. Sci. Rep. 10, 407 (2020).
Bartolak-Suki, E. et al. Fluctuation-driven mechanotransduction regulates mitochondrial-network structure and function. Nat. Mater. 14, 1049–1057 (2015).
Phuyal, S., Romani, P., Dupont, S. & Farhan, H. Mechanobiology of organelles: illuminating their roles in mechanosensing and mechanotransduction. Trends Cell Biol. 33, 1049–1061 (2023).
Torrino, S. et al. Mechano-induced cell metabolism promotes microtubule glutamylation to force metastasis. Cell Metab. 33, 1342–1357 e1310 (2021).
Mittelheisser, V. et al. Evidence and therapeutic implications of biomechanically regulated immunosurveillance in cancer and other diseases. Nat. Nanotechnol. 19, 281–297 (2024).
Li, J. et al. Bioelectret Materials and Their Bioelectric Effects for Tissue Repair: A Review. ACS Appl Mater. Interfaces 16, 38852–38879 (2024).
Wu, H. et al. Electrical stimulation of piezoelectric BaTiO3 coated Ti6Al4V scaffolds promotes anti-inflammatory polarization of macrophages and bone repair via MAPK/JNK inhibition and OXPHOS activation. Biomaterials 293, 121990 (2023).
Kong, Y. et al. Wireless localized electrical stimulation generated by an ultrasound-driven piezoelectric discharge regulates proinflammatory macrophage polarization. Adv. Sci. (Weinh.) 8, 2100962 (2021).
Zhao, D. et al. Electromagnetized-nanoparticle-modulated neural plasticity and recovery of degenerative dopaminergic neurons in the mid-brain. Adv. Mater. 32, e2003800 (2020).
Carpentier, G. et al. Angiogenesis Analyzer for ImageJ - A comparative morphometric analysis of “Endothelial Tube Formation Assay” and “Fibrin Bead Assay. Sci. Rep. 10, 11568 (2020).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2012).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408 (2001).
Bardou, P., Mariette, J., Escudie, F., Djemiel, C. & Klopp, C. jvenn: an interactive Venn diagram viewer. BMC Bioinforma. 15, 293 (2014).
Shen, J. et al. Nerve regeneration potential of antioxidant-modified black phosphorus quantum dots in peripheral nerve injury. ACS Nano 18, 23518–23536 (2024).
Acknowledgements
We gratefully acknowledge Dr. Lei Zhan at Key Laboratory of Textile Science and Technology of Donghua University for his help with the piezoelectricity assay. We acknowledge Dr. Yan Shen from Shanghai Qingpu District Center for Disease Control and Prevention for her help with Ba2+ detection in blood samples. This work was supported by the National Key R&D Program of China (No. 2021YFC2400800), National Natural Science Foundation of China (No. 82372409, 52272278, 52072210), Sino-German Mobility Program (No.M-0699), Excellent Youth Cultivation Program of Shanghai Sixth People’s Hospital (No. ynyq202201), Basic Scientific Research Cultivation Project of Shanghai Sixth People’s Hospital (ynqn202404), Laboratory Open Fund of Key Technology and Materials in Minimally Invasive Spine Surgery (2024JZWC-ZDA05), Shanghai Sailing Program (24YF2735800) and Postdoctoral Fellowship Program of China Postdoctoral Science Foundation (2024M752035).
Author information
Authors and Affiliations
Contributions
X.S. and Y.Q. conceived the idea, designed the study and directed the project. J.S. performed all the in vitro and in vivo experiments and analyzed the data. S.W. assisted with Rhesus monkey behavior tests and data analysis. Y.W. performed piezoelectric hydrogel bioelectronics fabrication and characterization analysis. Z.Y. assisted with in vivo electrophysiology assessment. T.L. helped with in vitro fluorescence imaging. X.S. provided insightful advice. J.S. and Y.Q. wrote the manuscript. Y.Q. edited and corrected the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Thanh Nguyen and Lin Dong for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Shen, J., Wu, S., Wang, Y. et al. Mechano-bioactive hydrogel bioelectronics for mechanical-electrical-bioenergetic conversion and glia-modulating neural regeneration. Nat Commun 16, 11582 (2025). https://doi.org/10.1038/s41467-025-66779-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-66779-5
