
Abstract
Metastatic cancer cells, originating from cancer stem cells with metastatic capacity, utilize nutrient flexibility to navigate the challenges of the metastatic cascade. However, the nutrient required to maintain the stemness potentials of metastatic cancer cells remains unclear. Here, we reveal that metastatic breast cancer cells sustain stemness and initiate metastasis upon detachment by taking up and oxidizing lactate. In detached metastasizing breast cancer cells, lactate is incorporated into the tricarboxylic acid cycle, boosting oxidative phosphorylation, and promoting the stemness potentials via α-KG-DNMT3B-mediated SOX2 hypomethylation. Moreover, lactate is taken up and oxidized in mitochondria by the CD147/MCT1/LDHB complex, which correlates with stemness potentials and tumor metastasis in patients with breast cancer. An intracellularly expressed single-chain variable fragment targeting mitochondrial CD147 (mito-CD147 scFv) effectively disrupts the mitochondrial CD147/MCT1/LDHB complex, inhibits lactate-induced stemness potential, depletes circulating breast cancer cells, and reduces metastatic burden, suggesting promising clinical applications in reducing lactate-fueled metastasis.
Similar content being viewed by others
Introduction
Breast cancer is the most prevalent cancer and the leading cause of death in women worldwide, with 2.3 million diagnosed new cases and 670,000 deaths in 20221. In patients with breast cancer, metastasis contributes to over 90% of cancer-associated deaths2,3, although the process of metastasis is inefficient. During the metastatic cascade, millions of tumor cells detach from the primary tumor, but very few of them (less than 0.02%) can survive and effectively form growing metastatic foci4. This small proportion of cancer cells is a subgroup of cancer stem cells (CSCs) that are capable of initiating metastasis and setting up metastatic colonies in secondary organs, and are termed metastasis-initiating cells (MICs)5,6,7. Therefore, understanding the origin and maintenance mechanisms of MICs helps to specifically and effectively eradicate MICs and reduce metastasis.
Being different from CSCs that are identified with cancer-driving mutations, no metastasis-specific mutations occurred in MICs8. In fact, the origins of MICs are multifaceted. Besides originating from CSC subclones with pre-existing high metastatic potential and an elevated epithelial–mesenchymal transition (EMT) status9, MICs can also arise from non-CSCs that acquire stemness potentials during metastasis, such as detachment, manifesting phenotypic plasticity with dynamic changes in stemness potentials6. For example, in recurrent colorectal cancer, metastatic outgrowth is driven by Lgr5+ stem cells directly or by Lgr5+ cells that have transformed from Lgr5− non-stem cells via phenotypic conversion10,11. Similarly, our previous study demonstrated that detached ALDH− non-stem breast cancer cells gained the stemness potentials via resistance to detachment-induced anoikis12, the first major survival challenge for MICs and a critical barrier to forming metastasis13. Consequently, phenotypic plasticity ensures MICs bear the adaptability to counteract various stresses and survival pressures during the metastatic cascade and is considered an emerging hallmark of cancer stemness and tumor metastasis9,14,15.
Recently, metastatic cancer cells have been found to have metabolic adaptability and exhibit stage-dependent metabolic traits at every step of the metastatic cascade, which leads to phenotypic plasticity15. Evidence from single-cell RNA sequencing in human breast cancer showed that MICs switched their metabolic phenotype from glycolysis in primary tumors to oxidative phosphorylation (OXPHOS) in established micro-metastases16. Moreover, metabolites like pyruvate, lactate, glutamine, oleic acid, and cystine in the circulation all potentially help detached cancer cells defend against oxidative stress8 and further contribute to anoikis resistance.
These facts suggest that detached cancer cells possess metabolic flexibility, allowing them to adapt to various bioenergetic substrates according to their bioavailability. However, whether this metabolic flexibility contributes to the stemness potentials of MICs and the initiation of tumor metastasis in breast cancer remains unclear. It is well established that glucose uptake decreases upon detachment17,18,19, but the main energy substrates and corresponding metabolic pathways necessary for maintaining MIC stemness under stress and initiating metastasis still need to be determined.
In the present study, we investigated the energy substrates of MICs and their contributions to stemness properties in metastatic breast cancer from the perspective of metabolic adaptation. We found that lactate, taken up and oxidized by the mitochondrial cluster of differentiation 147 (CD147)/monocarboxylate transporter 1 (MCT1)/lactate dehydrogenase B (LDHB) complex, was converted into the tricarboxylic acid (TCA) cycle intermediate α-ketoglutaric acid (α-KG), which subsequently induced MIC stemness potentials via DNA methyltransferase 3 beta (DNMT3B)-mediated SOX2 hypomethylation. By developing an intracellularly expressed single-chain variable fragment (scFv) that specifically targets mitochondrial CD147, we demonstrated that mito-CD147 scFv effectively inhibited metastasis in breast cancer. Therefore, we propose that lactate serves as a fuel for the TCA cycle in detached metastatic breast cancer cells and promotes the stemness potentials of MICs, while blocking of lactate utilization could be used to eliminate such cells.
Results
Lactate serves as a fuel for the TCA cycle in detached breast cancer cells
To determine the energy substrates of detached breast cancer cells, we examined their utilization of major carbon sources. We noticed that the uptake of glucose and the expression of glucose transporter GLUT1 were significantly decreased in both ALDH− and total anoikis-resistant (AR) cells compared to adherent (Ad) cells (Fig. S1A–C). We also observed a significantly decreased uptake of fatty acid and glutamine in AR cells (Fig. S1D, E), but the reduction in fatty acid uptake (28.16 ± 1.52%) and intracellular glutamine levels (19.71 ± 0.71%) was significantly lower than that of glucose (50.62 ± 3.79%). By analyzing the single-cell RNA sequencing data of breast cancer PDXs (GSE123837), however, we found significant enrichment of several metabolic pathways and organic acid metabolic processes in lung metastases compared to primary tumors (Fig. S1F). Given our previous findings showing that detached breast cancer cells exhibit significantly prolonged doubling times, these data suggest that their growth retardation may lead to the reduced utilization of three major nutrients while shifting toward the consumption of alternative energy sources.
To explore potential alternative energy sources for detached breast cancer cells, we conducted a non-targeted metabolomics analysis. The top five reduced metabolites in the supernatant of AR cells were valine, 2-Hydroxy-3-methylpentanoic acid, alanine, lactic acid (lactate), and phosphate (Fig. 1A, B). As glucose utilization was primarily decreased among the three major nutrients, we then focused on lactate, an organic acid and metabolic reservoir for glucose20. As expected, we found that lactate levels in AR cells were significantly decreased in the supernatant but increased inside the cells compared to those in Ad cells (Fig. 1C). Additionally, we observed that plasma lactate levels progressively declined during tumor progression and metastasis, while tumor tissue lactate concentrations increased in 4T1-bearing mice (Fig. 1D). These data suggested that lactate utilization is enhanced in AR cells and correlates with tumor metastasis.

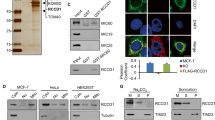
A Non-targeted metabolomics analysis of the supernatant from anoikis-resistant (AR) and adherent (Ad) MCF7 cells. n = 6 biologically independent samples. B Quantification of lactic acid levels from (A). n = 6 biologically independent samples. **P = 0.0022; two-tailed t test. C Extracellular and intracellular lactate (lac) levels in AR and Ad MCF7/T47D/4T1 cells. n = 3 independent experiments. Left, MCF7, ****P < 0.0001; T47D, ****P < 0.0001; 4T1, **P = 0.0012. Right, MCF7, **P = 0.0025; T47D, **P = 0.0028; 4T1, **P = 0.0039; two-tailed t test. D Representative image of tumor burden (left) and measurements of plasma (middle) and tumor (right) lactate levels in 4T1-luciferase-bearing mice (n = 7 mice). E Schematic diagram of 13C3-lactate tracing into TCA cycle intermediates and TCA cycle-derived metabolites. F Differentially M + 3 (left) and M + 2 (right) labeled metabolites in AR and Ad MCF7 cells. n = 6 biologically independent samples. Two-tailed t test. G Heatmap of differentially expressed proteins between AR and Ad MCF7 cells. n = 3 biologically independent samples. H MCT1 and LDHB protein levels in AR and Ad MCF7/T47D/4T1 ALDH−, total, and ALDH+ cells (α-Tubulin, loading control). Representative of 3 independent experiments. All bar graphs are presented as mean ± SEM. Source data are provided as a Source Data file.
To further investigate lactate utilization in AR cells with decreased glucose uptake, we cultured cells in glucose-free medium supplemented with lactate. We found that lactate levels were time-dependently and progressively decreased in the supernatant while increasing inside the cells in lactate-treated ALDH−, total, and ALDH+ cells (Fig. S1H). Lactate has been reported to be imported into cells, oxidized to pyruvate, and incorporated into the TCA cycle21,22. Thus, we analyzed the 13C3-lactate-derived metabolites using stable isotope labeling techniques. Compared to Ad cells, significantly increased 13C3-lactate-derived metabolites in AR cells included pyruvate, acetyl-CoA, TCA cycle intermediates (fold change from high to low: oxaloacetate, succinate, α-ketoglutarate, malate, fumarate, isocitrate, and citrate), and TCA cycle-derived metabolites (glutamate, aspartate, and ribose-5-phosphate), indicating enhanced lactate uptake and oxidation as well as increased TCA metabolism (Fig. 1E, F). Moreover, proteomics screening revealed that monocarboxylate transporter 1 (MCT1, protein for the SLC16A1 gene) and lactate dehydrogenase B (LDHB), responsible for lactate uptake and oxidation, respectively, were among the top three significantly upregulated proteins in AR cells (Fig. 1G). Their successively upregulation was also verified in AR ALDH−, total, and ALDH+ cells (Figs. 1H and S1I) as well as in lung metastases of breast cancer PDXs (Fig. S1G). Taken together, these results suggested that lactate is preferentially taken up by detached ER+ breast cancer cells (may be dependent on ALDH status) and incorporated into the TCA cycle as a fuel.
Lactate induces stemness potentials in detached breast cancer cells
We previously demonstrated that detachment enhances the stemness potentials of AR breast cancer cells12. Given that lactate acts as a fuel for detached breast cancer cells, we wondered whether lactate contributes to their stemness potentials. Incubation of breast cancer cells in glucose-free medium supplemented with lactate induced a dose-dependent increase in ALDH+ and CD44highCD24−/low CSC subpopulations, as well as tumorsphere formation, with the most pronounced effects observed at 20 mM lactate (Fig. S2A, B). In contrast, these effects were attenuated in glucose-containing conditions. PCR array analysis revealed that 30 stem cell transcription factors were significantly altered in AR MCF7 ALDH− cells compared to those in their Ad counterparts, with SOX2 being the only factor changed in both AR and lactate-treated ALDH− and total breast cancer cells (Fig. S2C–E and Supplementary Data 1). Lactate also increased the expression of MCT1/LDHB (Fig. S2F, G), while the inhibition of lactate uptake or oxidation using MCT1/LDHB siRNAs or inhibitors significantly suppressed the stemness potential, specifically the ratios of ALDH+ and CD44highCD24−/low CSC subpopulation, expression of SOX2 protein, and tumorsphere numbers in both AR (Fig. S3A–D) and lactate-treated (Fig. S3E, F) cells. All these data indicate that lactate promotes the stemness potentials in detached ER+ breast cancer cells.
Moreover, lactate administration significantly increased the number of lung metastatic nodules, the ratios of ALDH+ CSC subpopulation, and the expression of SOX2/MCT1/LDHB in lung metastatic lesions from PDX1 (Figs. 2A–D and S3G), PDX2, and MDA-MB-231 human breast cancer cells (Fig. S3H–J), as well as in 4T1 mouse breast cancer cell-derived xenografts (Fig. 2B, C). In contrast, lactate administration had no obvious effect on the tumor growth (Fig. S3K, L) and stemness potentials of primary tumors in all the models mentioned above, although the median survival time in 4T1 xenograft-bearing mice was significantly shortened (Fig. 2E). Lactate administration also significantly increased the numbers (Fig. 2F) and the SOX2 expression (Fig. 2G–I) in CD45−CK18+ CTCs from PDX1 or 4T1 xenografts. However, inhibiting lactate uptake/oxidation by CHC/GNE significantly reduced the number of lung metastatic nodules (Fig. 2J, K), the ALDH+ and CD44highCD24−/low CSC ratios (Figs. 2L and S3M), the SOX2 expression in lung metastatic lesions (Fig. S3N) and in CK18+ CTCs (Fig. 2M, N). Taken together, these findings suggest that lactate increases the stemness potentials in detached triple-negative breast cancer (TNBC) cells, including lung metastatic cells and CTCs, and enhances the metastatic capacity of MICs. Interestingly, within tumor tissues of 4T1-bearing mice, we observed spatially coexistence of lactate-producing glycolytic tumor cells and lactate-importing oxidative tumor cells. Notably, oxidative tumor cells displayed significantly higher MCT1 and SOX2 expression (Fig. S3O, P), implying that these oxidative cells might enhance their stemness potential through lactate consumption.

A–C Experimental design (A), lung nodule numbers (B, n = 10 mice/group) and ALDH+ ratios in primary and lung metastatic tumors (C; PDX1, n = 5 mice/group; 4T1, n = 4 mice/group) from lactate-treated PDX1 or 4T1 mice. B ****P < 0.0001; C **P = 0.0011; ***P = 0.0005; two-tailed t test. A created in BioRender. Song, C. (2025) https://BioRender.com/cphxog5. D SOX2 IHC quantification in primary and metastatic tumors from lactate-treated PDX1 mice (n = 20 mice/group). ****P < 0.0001, two-tailed t test. E Survival curves for lactate-treated 4T1 mice (n = 10 mice/group). ***P = 0.0005, Kaplan–Meier and log-rank test. F Circulating tumor cell (CTC) counts in lactate-treated PDX1 or 4T1 mice (n = 5 mice/group). *P = 0.0254; **P = 0.0017; two-tailed t test. G SOX2 fluorescence intensity in CTCs from lactate-treated PDX1 (lac 0 mg/kg, n = 22 CTCs from 4 mice; lac 800 mg/kg, n = 36 CTCs from 5 mice) and 4T1 mice (lac 0 mg/kg, n = 60 CTCs from 5 mice; lac 800 mg/kg, n = 80 CTCs from 5 mice). ****P < 0.0001; two-tailed t test. H, I Immunofluorescence images of CTCs from lactate-treated PDX1 (H; scale bars, 10 μm) and 4T1 mice (I; scale bars, 5 μm). DAPI (blue), CK18 (green), SOX2 (red). J–L Experimental design (J), lung nodule numbers (K), and ALDH+ ratios in lung metastases (L) from 0/800 mg/kg lactate-treated 4T1 mice (n = 4 mice/group) with NS or CHC/GNE. K ***P = 0.0004, Lac 800 mg/kg NS vs CHC; ***P = 0.0005, Lac 800 mg/kg NS vs GNE; ****P < 0.0001; L ****P < 0.0001; one-way ANOVA with Bonferroni correction. J Created in BioRender. Song, C. (2025) https://BioRender.com/pfhp7j6. M, N Immunofluorescence images of CTCs (M; scale bars, 10 μm), CTC counts (N, left; n = 5 mice/group), and SOX2 intensity in CTCs (N, right; NS, n = 100 cells from 5 mice; CHC, n = 50 cells from 5 mice; GNE, n = 50 cells from 5 mice) from lactate-treated 4T1 mice with NS or CHC/GNE. M DAPI (blue), CK18 (green), SOX2 (red). N left, ***P = 0.0001, ****P < 0.0001; N right, ****P < 0.0001; one-way ANOVA with Bonferroni correction. All bar graphs are presented as mean ± SEM. Source data are provided as a Source Data file.
We previously showed that detachment induces the stemness potentials in non-stem breast cancer cells and reprograms non-CSCs into CSCs12. We then further analyzed the stemness potentials in ALDH− non-stem breast cancer cells cultivated with or without lactate treatment and found that lactate indeed increased the stemness potentials in these ER+ cells (Fig. 3A–C). Similarly, in 4T1 ALDH− cell-bearing mice, lactate administration significantly promoted lung metastasis (Fig. 3D) and augmented the stemness potentials in metastatic lesions (Fig. 3E, F), but did not alter primary tumor growth (Fig. 3G). By establishing tumor-bearing models with mCherry-labelled 4T1 ALDH− cells (Fig. 3H), we discovered that the administration of lactate significantly increased both the proportion of mCherry⁺ALDH+ cells in metastatic lung lesions and the expression level of SOX2 in mCherry⁺ cells isolated from these lesions (Fig. 3I, J). Besides, lactate increased the expression of MCT1/LDHB in ALDH− cells (Fig. 3C) and in lung metastatic lesions from ALDH− xenografts (Fig. 3F), while MCT1/LDHB siRNA or CHC/GNE significantly reduced the lactate- or AR-induced stemness potentials in ALDH− cells (Fig. 3K–N). CHC/GNE also significantly reduced the lactate-enhanced lung metastasis and stemness potentials in metastatic lesions from TNBC 4T1 ALDH− xenografts (Fig. 3O–Q). These results reveal that lactate induces the stemness potentials and facilitates metastatic capacity in non-stem breast cancer cells.

A–C ALDH+ and CD44highCD24−/low ratios (A), tumorsphere numbers (B), and SOX2/MCT1/LDHB levels (C, α-Tubulin, loading control) in cells as indicated. n = 3 independent experiments (A, B); representative of 3 independent experiments (C). A left, ***P = 0.0005, ****P < 0.0001; right, MCF7, *P = 0.0192, T47D, *P = 0.0193; B, *P = 0.0423, **P = 0.0015; two-tailed t test. D Lung nodule numbers in 4T1 ALDH− cell-bearing mice (n = 10 mice/group) as indicated. ****P < 0.0001, two-tailed t test. E, F ALDH+ ratios (E, n = 5 mice/group) and SOX2/MCT1/LDHB levels (F, n = 4 mice/group; α-Tubulin, loading control) in 4T1 ALDH− cell-bearing mice as indicated. E ***P = 0.0005, two-tailed t test. G Tumor growth curves of 4T1 ALDH− cell-bearing mice (n = 8 mice/group) as indicated. Two-way ANOVA with Bonferroni correction. H, I Experimental flowchart (H) and mCherry+ALDH+ ratios in mCherry-4T1 ALDH− cell-bearing mice (I, n = 5 mice/group) as indicated. ns, P = 0.2567, ****P < 0.0001, two-tailed t test. H Created in BioRender. Song, C. (2025) https://BioRender.com/5why6l0. J SOX2/MCT1/LDHB levels of mCherry+ cells from mCherry-4T1 ALDH− cell-bearing mice (n = 4 mice/group; α-Tubulin, loading control). K, L SOX2 levels (K, representative of 3 independent experiments; α-Tubulin, loading control) and ALDH+ ratios (L, n = 3 independent experiments) in cells as indicated. L MCF7, ***P = 0.0004, siNC vs siMCT1, ***P = 0.0005, siNC vs siMCT1; T47D, ***P = 0.0003; one-way ANOVA with Bonferroni correction. M, N ALDH+ and CD44highCD24−/low ratios (M), and tumorsphere numbers (N) in cells as indicated. n = 3 independent experiments. M left, MCF7, **P = 0.0015, NC vs CHC, **P = 0.0032, NC vs GNE; T47D, ****P < 0.0001; right, MCF7, ***P = 0.0005, NC vs CHC, ***P = 0.0006, NC vs GNE; T47D, **P = 0.0024, NC vs CHC, ***P = 0.0006, NC vs GNE; N MCF7, **P = 0.0075, NC vs CHC, **P = 0.0015, NC vs GNE; T47D, *P = 0.0451, NC vs CHC, *P = 0.0232, NC vs GNE; one-way ANOVA with Bonferroni correction. O–Q Lung nodule numbers (O, n = 10 mice/group), ALDH+ and CD44highCD24−/low ratios (P, n = 5 mice/group), and SOX2 levels (Q, n = 3 mice/group; α-Tubulin, loading control) in 4T1 ALDH− cell-bearing mice as indicated. O ****P < 0.0001, ***P = 0.0003, Lac vs Lac+CHC, ***P = 0.0004, Lac vs Lac+GNE; P left, ****P < 0.0001, right, ***P = 0.0002, ****P < 0.0001; one-way ANOVA with Bonferroni correction. Data are presented as mean ± SEM. Source data are provided as a Source Data file.
Lactate induces stemness potentials in detached breast cancer cells by fueling the TCA cycle and promoting OXPHOS
We next focused on how lactate induces stemness potentials in detached breast cancer cells. In addition to being a major circulating carbohydrate fuel, lactate can induce histone lactylation via the histone acetyltransferase P30023,24 or act as a ligand for the G-protein-coupled receptor GPR8125,26. By pre-incubating the lactate-treated cells with the P300 inhibitor A485 or GPR81 siRNA, our findings revealed that, except for the downregulation of SOX2 expression in T47D cells, neither P300 (Fig. S4A–D) nor GPR81 (Fig. S4E–I) significantly affected lactate-induced stemness potential.
To further explore the detailed role of lactate in stemness potential, we performed a proteomics analysis. In both AR and lactate-treated cells, the top two significantly enriched pathways were carbon metabolism and the citrate cycle (TCA cycle) (Figs. 4A and S4J). The TCA cycle is coupled to OXPHOS to provide cells with ATP. Indeed, we observed that ATP levels and oxygen consumption rates (OCRs) were significantly enhanced in both AR and lactate-treated ALDH− or total cells (Fig. 4B–D). Additionally, the activities of the electron transport chain complexes, especially mitochondrial complex I, were significantly increased in AR and lactate-treated ALDH− or total cells (Figs. 4E and S4K). Notably, while complex I assembly and mitochondrial mass remained unchanged, we observed significant upregulation of pan-Kla (lysine lactylation) on the complex I subunit NDUFS1 (Fig. S4L–N). In contrast, the suppression of lactate utilization by MCT1/LDHB inhibitors or siRNAs significantly weakened ATP production, OCR, and complex I activity in lactate-treated cells (Figs. 4F–H and S4O, P). Moreover, the ATP synthase inhibitor oligomycin and the complex I inhibitor rotenone both significantly decreased the ratios of ALDH+ or CD44highCD24−/low CSC subpopulations (Fig. 4I, J), tumorsphere numbers (Fig. 4K), and SOX2 expression in AR and lactate-treated ALDH− or total cells (Fig. 4L). Taken together, these data suggest that lactate induces stemness potentials in detached ER+ breast cancer cells by enhancing the Complex I activity and lactylation of its critical subunit NDUFS1.

A KEGG pathway enrichment of differentially expressed proteins between AR and Ad MCF7/T47D cells. B–E ATP levels (B), oxygen consumption rates (OCRs, C, D), and complex I activities (E) in AR or lactate-treated ALDH− or total cells. n = 3 independent experiments. B left, ****P < 0.0001, B right, MCF7, **P = 0.0013, T47D, **P = 0.0059; E left, *P = 0.0244, **P = 0.0017, E right, MCF7, **P = 0.0032, T47D, **P = 0.0024; two-tailed t test. F–H ATP levels (F), complex I activities (G), and OCRs (H) in cells incubated with 0/20 mM lactate after pretreatment with PBS (NC) or 1 mM CHC/GNE for 2 h. n = 3 independent experiments. F ***P = 0.0002; G **P = 0.002, Lac 20 mM NC vs CHC, **P = 0.0016, Lac 20 mM NC vs CHC; one-way ANOVA with Bonferroni correction. I–L ALDH+ (I) or CD44highCD24−/low (J) ratios, tumorsphere numbers (K), and SOX2 protein levels (L, α-Tubulin, loading control) in AR or lactate-incubated ALDH− or total cells as indicated. n = 3 independent experiments (I–K); representative of 3 independent experiments (L). I left, ***P = 0.0001, ****P < 0.0001, I right, *P = 0.0466, **P = 0.0042; J ***P = 0.0007, **P = 0.0083; K AR MCF7, **P = 0.0042, NC vs Rotenone, *P = 0.0105, NC vs Oligomycin, AR T47D, *P = 0.0124, NC vs Rotenone, **P = 0.0069, NC vs Oligomycin, Lac 20 mM MCF7, **P = 0.0046, NC vs Rotenone, ***P = 0.0010, NC vs Oligomycin, Lac 20 mM T47D, **P = 0.0015, NC vs Rotenone, *P = 0.0188, NC vs Oligomycin; I right and (J), two-tailed t test; I left and (K), one-way ANOVA with Bonferroni correction. All bar graphs are presented as mean ± SEM. Source data are provided as a Source data file.
Lactate induces the stemness potentials of detached breast cancer cells by promoting α-KG-DNMT3B-mediated hypomethylation of SOX2
To clarify the metabolic-epigenetic mechanism of lactate-induced stemness potentials in detached breast cancer cells via fueling the TCA cycle, we next focused on α-ketoglutarate (α-KG), a vital signaling molecule regulating stemness gene expression and one of the top three significantly increased TCA cycle intermediates (oxaloacetate, succinate, α-KG) in 13C3-lactate treated AR cells (Fig. 1E, F). We found that the levels of α-KG were significantly increased in AR and lactate-treated cells (Fig. S5A), but were abolished by the MCT1/LDHB inhibitor CHC/GNE (Fig. S5B). Moreover, the AR- and lactate-boosted stemness potentials (Fig. 5A–C) were significantly facilitated by the α-KG analogue DM-αKG but were suppressed by the IDH inhibitor Enasidenib, which markedly decreased α-KG levels (Fig. S5C). These data indicated that lactate induces the stemness potentials of detached ER+ breast cancer cells through TCA cycle intermediates α-KG.

A–C ALDH+ ratios (A), tumorsphere numbers (B), and SOX2 levels (C, α-Tubulin, loading control) in cells as indicated. n = 3 independent experiments (A, B); representative of 3 independent experiments (C). A ***P = 0.0005, ****P < 0.0001; B MCF7, *P = 0.0411, **P = 0.0066, 0.003 (AR, Lac 20 mM), ***P = 0.0005; T47D, **P = 0.0051, 0.0026 (NC vs DM-αKG, Enasidenib), ***P = 0.0008, 0.0007 (NC vs DM-αKG, Enasidenib); one-way ANOVA with Bonferroni correction. D–F 5-methylcytosine (5-mC) levels detected by dot blot (D, representative of 3 independent experiments), immunofluorescence staining (E; scale bars, 20 μm), and intensity quantification (F, n = 20 typical fields from 3 biologically independent samples) in AR cells. E DAPI (blue), 5-mC (green); F ****P < 0.0001; two-tailed t test. G QPCR assay of methylation levels in 181720250-181720500 region of SOX2 promoter in cells as indicated. n = 3 independent experiments. MCF7, **P = 0.002, ****P < 0.0001, T47D, **P = 0.0021, 0.0091 (NC vs DM-αKG, AZA), ****P < 0.0001; one-way ANOVA with Bonferroni correction. AZA Azacytidine. H, I DNMT3B levels (H, representative of 3 independent experiments; α-Tubulin, loading control) and enzyme activities (I, n = 3 independent experiments) in cells as indicated. I MCF7, ***P = 0.00087, 0.0001 (Ad vs AR, Lac 0 vs Lac 20 mM); T47D, **P = 0.0013, ***P = 0.00059; two-tailed t test. J DNMT3B levels in cells as indicated. Representative of 3 independent experiments. α-Tubulin, loading control. K ChIP-qPCR of DNMT3B binding to the 181720250-181720500 region of SOX2 promoter in cells as indicated. H3, positive control; IgG, negative control. n = 3 independent experiments. MCF7, *P = 0.0176, 0.011 (AR, Lac 20 mM), **P = 0.0016, 0.0013 (AR, Lac 20 mM); T47D, *P = 0.0157, ***P = 0.0001, 0.0003 (NC vs DM-αKG, Enasidenib), ****P < 0.0001; one-way ANOVA with Bonferroni correction. L–N ALDH+ ratios (L), tumorsphere numbers (M), and SOX2 levels (N, α-Tubulin, loading control) in cells as indicated. n = 3 independent experiments (L, M); representative of 3 independent experiments (N). L MCF7, *P = 0.0419, 0.0286 (NC vs DM-αKG, Enasidenib), **P = 0.0057; T47D VC, **P = 0.0011, 0.0042 (NC vs DM-αKG, Enasidenib); T47D DNMT3B, **P = 0.0021, 0.0034 (NC vs DM-αKG, Enasidenib); M, MCF7, **P = 0.0021, ***P = 0.0002, ****P < 0.0001; T47D, *P = 0.0427, **P = 0.0012, ***P = 0.0004, ****P < 0.0001; one-way ANOVA with Bonferroni correction. DM-αKG, dimethyl α-ketoglutarate; DNMT3B, DNA Methyltransferase 3 beta. Data are presented as mean ± SEM. Source data are provided as a Source Data file.
We then turned to determine whether the lactate-induced stemness potential was due to the α-KG-mediated methylation modification of DNA or histones, especially H3K27m3, a critical histone modification responsible for α-KG-regulated gene expression. We found that total DNA methylation levels were obviously decreased in AR (Fig. 5D–F) and lactate-treated (Fig. S5D, E) cells, while H3K27m3 protein levels remained unchanged (Fig. S5F). By predicting the possible regions in the SOX2 promoter where DNA methylation likely occurs via the Ensembl database (Fig. S5G) and designing corresponding primers, we found that the methylation levels in the SOX2 promoter region 181720250-181720500, identified by primer 2 and predicted to be a CpG island by MethPrimer 2.0, were significantly decreased in AR or lactate-treated cells (Fig. S5H). Moreover, DM-αKG significantly decreased, while Enasidenib increased, the methylation levels of the SOX2 promoter region 181720250-181720500 in both AR (Figs. 5G and S5J) and lactate-treated (Fig. S5I, J) cells. Notably, the DNMT inhibitor azacitidine (AZA) significantly counteracted the Enasidenib-induced hypermethylation of the SOX2 promoter region. These data demonstrated that α-KG upregulated SOX2 expression in detached ER+ cancer cells by reducing the methylation level in its promoter region 181720250-181720500.
Finally, we aimed to determine the enzyme involved in α-KG-suppressed methylation of the SOX2 promoter. As DNA methyltransferase DNMT3B was the top significantly downregulated protein upon detachment in our proteomics data (Fig. 1G), the expression and enzymatic activity of DNMT3B were downregulated in AR or lactate-treated cells (Fig. 5H, I), and further decreased by DM-αKG but abolished by Enasidenib (Fig. 5J). We then focused on DNMT3B. In AR and lactate-treated cells, we found that the binding of DNMT3B to the SOX2 promoter region (181720250-181720500) was negatively correlated with α-KG activities (Fig. 5K), and DNMT3B overexpression significantly abolished the α-KG-induced stemness potentials (Figs. 5L–N and S5K). These observations suggested that the lactate-converted α-KG induces stemness potentials in detached ER+ breast cancer cells by reducing DNMT3B-mediated SOX2 methylation.
Mitochondrial CD147/MCT1/LDHB complex responsible for lactate uptake and oxidation correlates with tumor metastasis and CTC stemness potentials in patients with breast cancer
Then, how was the uptake and oxidation of lactate enhanced in detached breast cancer cells? CD147, a chaperone protein that anchors MCTs to specific membrane locations and is listed as one of the top three significantly upregulated proteins (along with MCT1 and LDHB) in AR cells (Fig. 1G), has caught our attention. We noticed that CD147 expression was significantly upregulated in lactate-treated cells and lactate-administered metastatic tumors (Fig. S6A, B). Additionally, CD147 overexpression or knockdown12 correspondingly facilitated or suppressed lactate uptake (Fig. S6C), and the lactate-induced stemness potentials (Fig. S6D–G), as well as MCT1/LDHB expression (Fig. S6F) and lung metastatic nodules (Fig. S6H). These results indicated that CD147 promoted MCT1/LDHB-mediated lactate uptake and oxidation, facilitating lactate-induced stemness potentials in detached breast cancer cells.
The specific subcellular compartment for lactate uptake and oxidation, whether in the cytoplasm or mitochondria, remains controversial. By employing an isotope labeling scheme using 2-2H-lactate27,28, we found that more than 95% of lactate in the mitochondria of AR MCF7 cells was 2H-labeled, and that the 2H-labeled lactate in the mitochondria of AR MCF7 cells was significantly higher than that in Ad cells (Fig. 6A, B), indicating that lactate could be imported into the mitochondria of detached ER+ breast cancer cell.

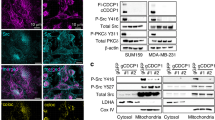
A, B Schematic of 2-2H-lactate metabolism (A) and 2H-labeled M + 1 metabolites (B, n = 3 biologically independent samples). **P = 0.0018, ****P < 0.0001; two-tailed t test. C, D CD147/MCT1/LDHB protein levels in mitochondria (C) and sub-compartments (D) from AR cells. K proteinase K, T triton X-100, O/I outer membrane and intermembrane space, I/M inner membrane and matrix. COX IV, Na+-K+ ATPase, α-Tubulin, TOMM20 mark of inner membrane, plasma membrane, cytoplasm, outer membrane. Data are representative of 3 independent experiments. E Transmission electron microscopy shows CD147/MCT1/LDHB co-localization (orange arrows) in the mitochondrial inner membrane of AR cells. Scale bars: 1 μm (left), 500 nm (right). Red asterisks, mitochondria. F Proximity ligation assay (PLA) between CD147 and LDHB in lactate-treated MCF7 cells. Scale bars, 5 μm. n = 6 typical fields from 3 biologically independent samples. DAPI (blue), PLA signal (red). ****P < 0.0001; two-tailed t test. G Pull-down assay between GST-CD147 N-terminal and His-LDHB. Representative of 3 independent experiments. H CD147 (red)/LDHB (blue)/MCT1 (purple) co-localization (orange arrows) in COX IV (green) labeled mitochondria from PDX1 lung metastatic tumors. Scale bars: 20 μm. I CD147 (green)/LDHB (yellow)/MCT1 (orange) expressions in CK18 (red) labeled cells from breast cancer patients with or without metastasis. Scale bars: 40 μm. J Correlation between CD147 and MCT1/LDHB in 11 metastatic patients with breast cancer. R2, Pearson correlation coefficient. Spearman correlation analysis. K Kaplan–Meier survival (up) and recurrence (down) analysis for 131 patients stratified by CD147/MCT1/LDHB co-expression (high n = 9 patients, low n = 31 patients). L Representative images of HE (left: scale bars, 50 μm) and SOX2 IHC staining (middle: scale bars, 50 μm; right: enlarged image, scale bars, 10 μm) in CTCs from No. 9 patient in Fig. S7M. M Co-localization (orange arrows) of CD147 (red)/LDHB (blue)/MCT1 (purple) in COX IV (green) labeled mitochondria of CTCs from No. 9 patient in Fig. S7M. Scale bars: 20 μm. N SOX2 IHC intensity in CTCs from 10 patients stratified by mitochondrial CD147/MCT1/LDHB complex presence (without, n = 4 patients; with, n = 6 patients). ***P = 0.0004, two-tailed t test. Data are mean ± SEM. Source data are provided as a Source Data file.
As CD147 subcellular localization29 and lactate uptake/oxidation both occur in mitochondria, and PPI network analysis suggested a possible interaction between CD147 and MCT1 or LDHB (Fig. S6I), we presumed that CD147 might regulate lactate uptake/oxidation by interacting with MCT1 or LDHB in the mitochondria. By isolating high-purity mitochondria (Fig. S6J), we indeed found that the protein levels of CD147, MCT1, and LDHB were increased (Fig. S6K), their co-localization was enhanced (Fig. S6L), and the interactions between CD147 and MCT1 or LDHB were augmented (Figs. S6M and S7A) in AR or lactate-treated cells. By conducting protease K protection and mitochondria sub-fractionation isolation assays, we found that the increased expression of these three molecules occurred in the mitochondrial inner membrane and/or matrix of AR (Fig. 6C, D) or lactate-treated (Fig. S7B, C) cells. Excitingly, transmission electron microscopy (TEM) confirmed the existence of a CD147/MCT1/LDHB complex in the mitochondrial inner membrane of AR cells (Fig. 6E). Moreover, molecular docking simulations predicted the binding conformation of three molecules: CD147 structurally bridges between MCT1 and LDHB. Specifically, its transmembrane region near the C-terminus interacts with MCT1 while its N-terminus binds to LDHB (Fig. S7D). Building upon the well-established CD147-MCT1 interaction30, we further investigated the potential CD147-LDHB interaction. Using proximity ligation assay (PLA), we confirmed a direct interaction between CD147 and LDHB, suggesting their functional coupling. Notably, lactate treatment significantly enhanced this interaction (Fig. 6F). To further validate this finding, we conducted GST pull-down assays combined with site-directed mutagenesis using purified GST-tagged CD147 (N-terminal) and His-tagged LDHB proteins, supplemented by molecular docking analyses. These experiments confirmed a direct physical interaction between CD147 and LDHB, mediated by key residues: Glu168 and Arg106 in CD147, and Arg99 and Asp56 in LDHB (Figs. 6G and S7E, F). These results suggested that there is a CD147/MCT1/LDHB complex in the inner membrane, which facilitates mitochondrial lactate uptake and oxidation in ER+ breast cancer cells.
Consistently, CD147 was co-localized and interacted with MCT1 or LDHB in the mitochondria of lung metastases from lactate-administered TNBC PDX1/2 (Figs. 6H and S7G–I) and 4T1 ALDH− (Fig. S7J) bearing mice, unlike in primary tumors. In the primary breast cancer tissues of metastatic patients, the expression of CD147 showed a strong positive correlation with that of MCT1 or LDHB. In contrast, a moderate or low positive correlation was observed in non-metastatic patients (Figs. 6I, J and S7K). Patients with higher co-expression of CD147/MCT1/LDHB had significantly worse overall survival and higher tumor metastasis/recurrence, and their co-expression was an independent predictor of poor overall survival (Figs. 6K and S7L and Table S2–4). Moreover, by analyzing the CTCs in blood vessels of breast cancer patient tissues, we found that mitochondrial CD147/MCT1/LDHB complexes occurred in CTCs of 6 out of 10 patients (Fig. S7M), and patients whose CTCs bore the mitochondrial CD147/MCT1/LDHB complex exhibited high SOX2 expression (Fig. 6L–N and S7N, O) and more advanced stages (Fig. S7M). Altogether, these results uncovered a mitochondrial CD147/MCT1/LDHB complex for lactate uptake/oxidation, which positively correlated with tumor metastasis and CTC stemness potentials in patients with breast cancer.
Mitochondrial CD147 promotes the lactate-induced stemness potentials in detached breast cancer cells
Due to the key role of CD147 in the mitochondrial CD147/MCT1/LDHB complex, we investigated the influence of mitochondrial CD147 on lactate-induced stemness potentials and metastatic capacity by constructing cells with CD147 knocked out (KO) or expressed exclusively in mitochondria (mito147, Fig. S8A, B). In detached MCF7 ALDH− cells with different CD147 levels, the mitochondrial uptake of 2-2H-lactate was completely blocked in CD147 KO cells but was efficiently rescued in mito147 cells to a comparable level to wild type (WT) cells (Fig. 7A, B), which was verified by the corresponding alteration of lactate levels in the supernatant (Fig. 7C), further confirming the role of mitochondrial CD147 in lactate uptake. Upon detachment or lactate treatment, mitochondrial-expressed CD147 efficiently reversed the reduced mitochondrial respiration (Figs. 7D and S8C), stemness potentials (Figs. 7E and S8D), and MCT1/LDHB expression (Fig. S8D) in CD147 KO cells to a degree comparable to WT cells. Likewise, lactate-administered mito147 cell-bearing mice exhibited significantly recovered primary tumor growth, facilitated lung metastasis (Figs. 7F–G and S8E–G), enhanced stemness potentials in metastatic lesions (Figs. 7H, I and S8H), as well as increased CTC numbers and SOX2 expression in CTCs (Figs. 7J, K and S8I, J), compared to lactate-administered CD147 KO cell-bearing mice.

A, B Typical peak maps of 2-2H-lactate metabolic flux analysis (A) and 2H-labeled M + 1 metabolites (B, n = 3 biologically independent samples). WT, wild type; KO, CD147 knockout; mito147, mitochondrial CD147-expression; mito147 + scFv, mito147 cells+anti-mitochondrial CD147 scFv (single-chain variable fragment). C Lactate (Lac) levels in supernatants of AR MCF7/T47D WT/KO/mito147 cells (n = 3 independent experiments). ****P < 0.0001, one-way ANOVA with Bonferroni correction. D, E OCRs (D), ALDH+ ratios (E, left), and tumorsphere numbers (E, right) in AR MCF7/T47D WT/KO/mito147 cells (n = 3 independent experiments). E left, *P = 0.023, 0.0118 (MCF7, T47D), **P = 0.0011, ***P = 0.0003; E right, **P = 0.0033, ***P = 0.0003, ****P < 0.0001, one-way ANOVA with Bonferroni correction. F Tumor growth curves of MCF7 WT/KO/mito147 cell-bearing mice (n = 5 mice/group) with 800 mg/kg lactate administration. WT vs KO, **P = 0.0044, ****P < 0.0001; mito147 vs KO, & &P = 0.0087, 0.0012 (12, 15 day), & & & &P < 0.0001; two-way ANOVA with Bonferroni correction. G–I Lung nodule (G, n = 10 mice/group), ALDH+ and CD44highCD24−/low ratios (H, n = 5 mice/group), and SOX2/MCT1/LDHB protein levels (I, n = 4 mice/group, α-Tubulin, loading control) in tumors from indicated cell-bearing mice with 800 mg/kg lactate administration. **P = 0.0056, ****P < 0.0001, one-way ANOVA with Bonferroni correction. J, K Plasma CTCs numbers (J, n = 10 mice/group) in 800 mg/kg lactate-administered indicated cell-bearing mice, and SOX2 immunofluorescence intensity (K; WT, n = 120 cells from 6 mice; KO, n = 67 cells from 5 mice; mito147, n = 142 cells from 6 mice) in CTCs. ***P = 0.0001, ****P < 0.0001, one-way ANOVA with Bonferroni correction. CTC circulating tumor cell. L DNMT3B protein levels in MCF7/T47D WT/KO/mito147 cells as indicated. Representative of 3 independent experiments. α-Tubulin, loading control. DNMT3B DNA methyltransferase 3 beta. Data are mean ± SEM. Source data are provided as a Source Data file.
For the epigenetic mechanism by which mitochondrial CD147 promotes lactate-induced stemness potential, we found that mito147 cells had significantly higher α-KG accumulation (Fig. S8K), lower DNMT3B expression (Fig. 7L), and weaker binding of DNMT3B to the SOX2 promoter (Fig. S8L) than CD147 KO cells, whether under AR or lactate-treated conditions. Therefore, mitochondrial CD147 is essential for maintaining stemness potentials and promoting metastasis through enhanced lactate uptake/oxidation and the α-KG-DNMT3B mediated SOX2 hypomethylation.
Intracellularly expressed scFv targeting mitochondrial CD147 inhibits lactate-induced stemness potentials in detached breast cancer cells
To block the mitochondrial CD147-facilitated lactate uptake/oxidation and the induced stemness potentials in detached breast cancer cells, we intracellularly expressed a scFv specifically targeting mitochondrial CD147 (mito-CD147 scFv) by fusing a scFv sequence that targets CD147 to a matrix-targeting sequence that guides the scFv into mitochondria. The mito-CD147 scFv expressed in the pGV469 adenoviral vector specifically bound to the mitochondrial localized CD147 in both mito147 and wild-type cells (Figs. 8A and S9A, B), and restrained the formation of the mitochondrial CD147/MCT1/LDHB complex (Figs. 8B and S9C); more importantly, it blocked the uptake of 2-2H-lactate into the mitochondria of MCF7 ALDH− cells (Fig. 7A, B). In lactate-treated WT or mito147 cells, mito-CD147 scFv significantly inhibited mitochondrial respiration (Fig. S9D), ALDH+ and CD44highCD24−/low ratios (Fig. S9E), tumorsphere formation (Fig. S9F up), and SOX2/MCT1/LDHB expression (Fig. S9G). Moreover, mito-CD147 scFv reduced the α-KG levels (Fig. S9F down) and recovered the binding of DNMT3B to the SOX2 promoter (Fig. S9H). These data indicated that mito-CD147 scFv functioned as a brake to the lactate-induced stemness potentials by inhibiting α-KG-DNMT3B mediated SOX2 hypomethylation.

A Immunoactivity of EGFP-fused mito-CD147 scFv to CD147 in MCF7/T47D cell subfractions: C (plasma membrane and cytoplasm), M (mitochondria), COX IV, α-Tubulin mark mitochondrial inner membrane, cytoplasm. NC: pGV469 vector; scFv: pGV469-mito-CD147 scFv. Representative of 3 independent experiments. B Co-IP assays showing CD147 interaction with MCT1/LDHB in mitochondria from lactate-treated MCF7 WT/KO/mito147 cells as indicated. Groups: 1. WT; 2. WT+mito-CD147 scFv; 3. KO; 4. KO+mito-CD147 scFv; 5. mito147; 6. mito147+mito-CD147 scFv. Representative of 3 independent experiments. C–F Experimental design (C), lung nodules (D, n = 10 mice/group), ALDH+ and CD44highCD24−/low ratios (E, n = 5 mice/group), and SOX2 levels (F, n = 4 mice/group, α-Tubulin, loading control) in tumors from MMTV-PyMT mice as indicated. ***P = 0.0001 (E left), ***P = 0.0004, 0.0003 (E right, NS vs Lac, Lac vs Lac + scFv), ****P < 0.0001, one-way ANOVA with Bonferroni correction. C created in BioRender. Song, C. (2025) https://BioRender.com/37z58my. G, H CTC counts (G, left, n = 8 mice/group), SOX2 intensity (G, right; NS, n = 148 cells from 7 mice; Lac, n = 342 cells from 7 mice; Lac+scFv, n = 149 cells from 7 mice), and representative images of CTCs (H; scale bars, 40 μm), from MMTV-PyMT mice as indicated. H DAPI (blue), CK18 (green), SOX2 (purple). ****P < 0.0001, one-way ANOVA with Bonferroni correction. I, J Lung nodules (I, n = 8 mice/group), ALDH+ and CD44highCD24−/low ratios (J, n = 5 mice/group) in tumors from PDX1-bearing mice as indicated. ns, P = 0.9818, 0.869 (J left, right), ***P = 0.0001, ****P < 0.0001, one-way ANOVA with Bonferroni correction. K–M CTC counts (K, left, n = 8 mice/group), SOX2 intensity (K, right; NS, n = 80 cells from 8 mice; scFv, n = 80 cells from 8 mice; Lac, n = 120 cells from 6 mice; Lac+scFv, n = 80 cells from 8 mice), representative CTC images (L), and mito-CD147 scFv binding to mitochondrial CD147 (M) in CTCs from PDX1-bearing mice as indicated. Scale bars: 10 μm. L DAPI (blue), CK18 (green), SOX2 (purple). M DAPI (blue), CD147 (red), scFv (green), COX IV (purple). CTC circulating tumor cell, scFv single-chain variable fragment. Data are presented as mean ± SEM. Source data are provided as a Source Data file.
Being injected into lactate-administered MCF7 mito147 cell xenografts (Fig. S10A), pGV469-mito-CD147 scFv adenovirus markedly abolished the lung metastasis facilitated by mitochondrial CD147 (Fig. S10Bleft), reduced CSC ratios (Fig. S10C), and decreased SOX2 expression in metastatic lesions (Fig. S10D), while not affecting stemness potentials or tumor growth in primary tumors (Fig. S10D, E). Meanwhile, mito-CD147 scFv significantly diminished the numbers of CTCs boosted by mitochondrial CD147 and its SOX2 levels in lactate-administered mice (Fig. S10Bright, Fig. S10F, G).
To investigate the therapeutic potential on CTCs, mito-CD147 scFv adenovirus was administered via the tail vein to lactate-treated MMTV-PyMT spontaneous tumorigenic and PDX1-bearing mice that express native CD147 (Figs. 8C and S10H). We found that mito-CD147 scFv significantly decreased lactate-promoted lung metastasis (Fig. 8D and I), the CSC ratios, and SOX2 expression in lung metastatic lesions (Figs. 8E, F and J and S10I), as well as CTC numbers and stemness (Fig. 8G, H and K–M). Similar effects were also observed in MDA-MB-231 ALDH− cell-bearing mice (Fig. S10J–L). Altogether, mito-CD147 scFv effectively suppressed tumor metastasis and CTC stemness in breast cancer by blocking the formation of the mitochondrial CD147/MCT1/LDHB complex and inhibiting lactate uptake/oxidation.
Discussion
The ability to adopt nutrients flexibly and exhibit plastic stemness potentials enables MICs to survive stressful environments and successfully seed metastasis. However, the energy substrate governing stemness potentials in MICs has not been explored. In this study, we identified a previously undescribed role for lactate as an alternative carbon source in detached breast cancer cells, maintaining MIC stemness. Firstly, we found that lactate was taken up and oxidized in mitochondria, fueling the TCA cycle and enhancing α-KG-DNMT3B-SOX2-mediated stemness potentials. We also observed the CD147/MCT1/LDHB complex on the mitochondrial inner membrane, which facilitates the uptake and oxidation of lactate, thereby enhancing stemness potentials in disseminated breast cancer cells. Finally, we established the crucial role of mitochondrial CD147 in lactate uptake, oxidation, and stemness maintenance, and developed an intracellularly expressed scFv targeting mitochondrial CD147 that effectively inhibited stemness potentials and metastasis. Together, we demonstrated that the mitochondrial CD147/MCT1/LDHB complex contributes to lactate-mediated MIC stemness potentials, and targeting mitochondrial CD147 may be a potential strategy for treating lactate-fueled metastasis.
Growing evidence suggests that lactate plays important roles in tumor metastasis, with efficient metastasizers taking up more lactate28,31. Moreover, lactate has been reported to influence immune cells in the tumor environment, such as macrophages32, NK cells33, regulatory T cells34,35, and CD8+ T cells36,37,38. Nevertheless, the influence of lactate on the stemness potentials of MICs remains unclear. In this study, we demonstrated that lactate facilitated lung metastasis by inducing and maintaining the stemness potentials of detached breast cancer cells. This indicates that the key roles of lactate in tumor metastasis may also be linked to its regulation of MICs, in addition to its effects on host immunity, as previously believed. It is worth noting that lactate had a negligible impact on the growth of primary tumor, a phenomenon that has been previously reported39,40,41. This disparity can likely be ascribed to different metabolic dependencies. Specifically, primary tumor cells retain unimpaired GLUT1 expression, and thus preferentially uptake and utilize glucose rather than lactate for energy production42. Conversely, detached or metastatic tumor cells exhibit downregulated GLUT1 expression43, which impairs their glucose uptake and causes them to rely predominantly on lactate oxidation to fuel oxidative phosphorylation40. Notably, lactate exerts context-dependent regulatory effects on stemness, either suppressing or promoting it according to glucose availability (Fig. S2A). These findings highlight the metabolic plasticity of tumor-initiating cells, particularly during metastasis, a process characterized by detachment-induced glucose limitation43. Intriguingly, under such glucose-deprived conditions, lactate may shift from an inhibitory to a supportive role in maintaining stemness during metastasis.
Lactate is regarded as a central signaling molecule through protein lactylation, interacting with the GPR81 receptor44,45,46, or as a circulating substrate for the TCA cycle21,22. Herein, we found that lactate increased the stemness potentials of MICs by fueling the TCA cycle, which either facilitated OXPHOS and/or generated the TCA intermediate metabolite α-KG. Our study further demonstrated that lactate-derived α-KG inhibited DNMT3B-mediated SOX2 methylation, thereby enhancing the expression of the pluripotency transcription factor SOX2 in detached breast cancer cells. This scenario explains how lactate regulates the stemness potentials of MICs. Of note, Baek et al. revealed that under chemotherapy-induced stress, lactate increasingly enters the TCA cycle of TNBC cells, accompanied by enhanced mitochondrial OXPHOS47. In fact, there are substantial similarities between therapeutic stress and metastasis stress. First, tumor cells encounter comparable survival challenges during both therapeutic stress and metastasis, which may trigger metabolic plasticity that drives a shift from glycolytic metabolism to OXPHOS to sustain cell survival48,49. Thus, our observation that lactate fuels OXPHOS metabolism in anoikis-resistant breast cancer cells via TCA cycle entry further strongly support the findings by Baek et al. Second, CSCs play a role in both therapy resistance and metastatic dissemination50,51. The research by Baek et al. on therapy stress-induced metabolic reprogramming offers a mechanistic basis for understanding how does lactate regulate stemness potential of breast cancer cells during metastasis. Notably, in T47D cells, P300 inhibition or GPR81 knockdown resulted in reduced expression of the stemness transcription factor SOX2. This suggests that, in certain cellular contexts, lactate may regulate stemness not only through metabolic modulation but also via lactylation-dependent mechanisms or GPR81-mediated signaling, a possibility that merits further exploration.
The sequential uptake and oxidation of lactate depend on MCT1 and LDHB, respectively46,52. However, the transport function of MCT1 relies on its chaperone CD147, which regulates the expression, correct folding, and membrane localization of MCT1 via molecular interactions30,53,54. In the mitochondria of M6 muscle cells, for example, CD147 was found to interact with MCT129. Researchers had hypothesized the existence of a mitochondrial CD147/MCT1/LDHB complex mediating the uptake and oxidation of lactate, but no evidence has yet been found. Herein, we uncovered the presence of the CD147/MCT1/LDHB complex in the mitochondria, and this complex was correlated with stemness potentials in lung metastatic tumors and CTCs of patients with breast cancer, contributing to tumor metastasis and poor clinical outcomes for patients.
Lactate can be oxidized in either the cytoplasm or the mitochondria52. In our study, 95% of lactate in the mitochondria in metabolic flux analysis was 2H-labeled, suggesting that lactate primarily enters the mitochondria for oxidation. To determine the exact location of lactate oxidation within the mitochondrion, proteinase K protection and mitochondrial sub-fractionation assays revealed that the CD147/MCT1/LDHB complex is located in the mitochondrial inner membrane or the matrix. Additionally, TEM confirmed the anchoring of the CD147/MCT1/LDHB complex to the mitochondrial inner membrane, pointing to the matrix. These findings support that lactate is oxidized in the matrix side of the inner membrane of detached breast cancer cells.
We further revealed that the CD147/MCT1/LDHB complex adopts a conformation in which CD147 acts as a bridging molecule. Specifically, its C-terminal transmembrane region interacts with MCT1, while its N-terminal domain associates with LDHB. This structural configuration highlights CD147 as the central organizer within the lactate uptake and oxidation complex, underscoring its critical role in regulating both lactate transport and oxidation processes. The central role of mitochondrial CD147 in the CD147/MCT1/LDHB complex inspired us to explore the therapeutic potential of targeting mitochondrial CD147 to eradicate lactate-induced MIC stemness and tumor metastasis. Previously, our lab developed strategies targeting membranous CD147, such as the anti-CD147 metuximab to prevent tumor recurrence55 and enhance the chemosensitivity of pancreatic CSCs56, as well as anti-CD147 scFv-derived chimeric antigen receptor T (CAR-T) cells for inhibiting T cell acute lymphoblastic leukemia57. However, both anti-CD147 metuximab and anti-CD147 scFv cannot differentiate membranous CD147 from mitochondrial CD147. We therefore expressed an intrabody targeting mitochondrial CD147 by infusing anti-CD147 scFv with a mitochondrial presequence (Chinese invention patent 202410075009.2). This mito-CD147 scFv demonstrated significant efficacy in inhibiting lactate-induced stemness and metastasis by disrupting the CD147/MCT1/LDHB complex, thus holding potential for future translational studies.
In conclusion, the mitochondrial CD147/MCT1/LDHB complex enables detached breast cancer cells to import and oxidize lactate, which facilitates MIC stemness potentials and promotes metastasis by serving as a carbon source for the TCA cycle. This metabolic adaptation mechanism of lactate-induced stemness maintenance via the mitochondrial CD147/MCT1/LDHB complex opens an avenue for developing molecular-targeting therapy against mitochondrial lactate metabolism. The mito-CD147 scFv, a mitochondria-targeting intrabody, represents a promising strategy.
Methods
Human tumor tissues
The human tissue study protocol was approved by the Ethics Committee of Fourth Military Medical University (Approval number KY20183305-l) and informed consents have been obtained from the participants. A tissue microarray (TMA) constructed from paraffin-embedded tissue blocks of 131 female patients with breast cancer was purchased from Shanghai Outdo Biotech Company. Clinicopathological data were collected from the patients’ medical records, which included age at diagnosis, tumor type and grade, TNM stage, AJCC stage, overall survival, recurrence, etc. (Tables S2–S4). Additionally, tissue slides from 10 female patients with breast cancer with CTCs in blood vessels were obtained from Bioaitech Co., Ltd (Fig. S7M).
Animals models and experiments
The animal study protocols were approved by the Institutional Animal Care and Use Committee of the National Translational Science Center for Molecular Medicine (2023-NTSCMM-ID003). Six- to eight-week-old female BALB/c mice were purchased from Vital River Laboratory Animal Technologies (Beijing, China). Six- to eight-week-old female BALB/c nude mice, NCG mice, and MMTV-PyMT mice were obtained from Gempharmatech (Nanjing, China). Mice were maintained in a specific pathogen-free (SPF) mouse facility (room temperature, 18–23 °C; room humidity, 40–60%) with free access to food (XTI01ZJ-009, Xietong Shengwu, China) and water under a 12 h light/dark cycle.
The maximal permitted tumor burden was defined as a tumor volume not exceeding 1500 mm³. Tumorigenesis was monitored by measuring tumor dimensions every 3 days using a digital caliper. Tumor volume was calculated using the formula: Volume = (Length × Width²)/2. Mice were humanely euthanized before the tumor burden reached the maximum permitted size if they met any of the following predefined humane endpoint criteria: (1) tumor volume exceeding 1500 mm³; (2) ulceration, necrosis, or infection of the tumor that impaired the animal’s well-being; (3) significant weight loss (> 20% of baseline body weight); (4) difficulty in breathing, locomotion, or feeding; (5) signs of prolonged pain or distress that were not alleviated by analgesia. These endpoints were strictly followed for all survival studies to minimize suffering and to generate the survival curves presented in this study. We confirm that the maximal tumor size/burden permitted by our ethics committee was never exceeded during this study. In the instances where animals reached the humane endpoint criteria described above, they were euthanized promptly.
For lactate administration experiments (Fig. 2A), patient-derived xenograft (PDX) 1/2 tumor masses (2 × 1 × 1 mm3), 1 × 106 MDA-MB-231 cells, 1 × 105 4T1-luciferase cells, or 1 × 107 4T1 ALDH− cells were inoculated into the mammary adipose tissue pad of mice. When tumor volume reached 100 mm3, mice were randomized into two groups and injected intraperitoneally (i.p.) with or without lactate (Lac 800 mg/kg: sodium lactate, 800 mg/kg, 100 μL; Lac 0 mg/kg: 0.9% sterile saline/NS, 100 μL) for five consecutive days. On day 25, mice with 4T1-luciferase xenografts were given 150 mg/kg D-Luciferin potassium salt (ST196, Beyotime) i.p. for bioluminescence imaging, and images were recorded using an IVIS Lumina II (PerkinElmer, Waltham, MA, USA). On day 28, whole-body blood from mice was obtained for circulating tumor cell (CTC) isolation, counting, and staining; then mice were euthanized, and lungs and tumors were collected for lung nodule counting and histological staining as well as for stemness potential analysis via flow cytometry and western blot.
For lactate uptake/oxidation inhibition experiments (Fig. 2J), mice bearing xenografts of 100 mm3 were randomly assigned into four groups and injected i.p. without or with inhibitors for two consecutive days; three days later, mice were treated without or with lactate for five consecutive days. On days 25 and 28, mice were imaged, and the lungs and tumors were collected for the same analysis as the above procedures. For mice bearing 1 × 105 4T1-luciferase xenografts, the three groups were: NS+Lac, CHC+Lac, and GNE+Lac; for mice bearing 1 × 107 4T1 ALDH− xenografts, the four groups were: NS, Lac, Lac + CHC, and Lac+GNE. CHC (5 mg/kg, 100 μL) and GNE (5 mg/kg, 100 μL) were used as inhibitors for MCT1 and LDHB, respectively, with 0.9% sterile saline (NS) as the vehicle control.
For experiment investigating the function of mitochondrial localized CD147 (Fig. S8E), nude mice of similar weights were inoculated with 1 × 106 MCF7 or 1 × 107 T47D derived WT, KO, and mito147 cells. On day 14, all mice received lactate for five consecutive days. On day 28, whole-body blood, as well as the lungs and tumors, were collected from the mice for further analysis.
For experiment involving therapeutic mito-CD147 scFv, we first adopted a mouse xenograft model inoculated with cells expressing CD147 exclusively in mitochondria (Fig. S10A). Mice of similar weights were inoculated with 1 × 106 MCF7-derived WT/KO/mito147 cells; on day 14, MCF7-mito147 cells were randomly assigned into two groups and intratumorally injected with pGV469-mito‑CD147 scFv or pGV469 adenovirus (109 pfu, 30 μL) at multiple sites for three consecutive days; two days later, all mice were treated with lactate and evaluated on day 28. For the mouse model with native CD147 expression, 10-week-old MMTV-PyMT mice and mice bearing 1 × 107 MDA-MB-231 ALDH− cell xenografts or PDX1 derived tumors (100 mm3) were first injected with Lac or NS for five consecutive days; then, two days later, they were treated with pGV469-mito‑CD147 scFv or pGV469 adenovirus (108 pfu, 100 μL, i.v.) for three consecutive days; at 13 weeks old or on day 28, mice were evaluated (Figs. 8C and S10H). In MMTV-PyMT mice or mice with MDA-MB-231 ALDH− xenografts, three groups were assigned: NS, Lac, and Lac + mito‑CD147 scFv. In mice bearing PDX1 xenografts, four groups were assigned: NS, mito‑CD147 scFv, Lac, and Lac+ mito-CD147 scFv.
Cell lines and treatments
MCF7, T47D, MDA-MB-231, and 4T1 cells were cultured in RPMI 1640 medium with 3.15 g/L-glucose. All cells were routinely supplemented with 10% fetal bovine serum (FBS), 1% penicillin/streptomycin, and 2% L-glutamine. The ER+ MCF7 and T47D cells were untreated with charcoal stripping and without exogenous E2 supplementation. They were regularly tested for mycoplasma contamination and authenticated by short tandem repeat (STR) DNA profiling to ensure their identity and purity.
As depicted in the Fig. S11, there are two distinct cellular models in our study. One is the Low-Attachment Model. Cells surviving for 7 days in glucose-containing medium (3.15 g/L) under detached suspension culture conditions in ultra-low attachment plates were considered anoikis-resistant (AR) cells, while cells cultured in uncoated common plates under attached conditions with glucose-containing medium (3.15 g/L) served as control adherent (Ad) cells12. The other is Glucose-Free Model with Lactate. Due to the reduced glucose uptake in detached cells, adherent cells cultured in glucose-free medium, with or without exogenous lactate incubation, were designated as “Lac 20 mM” and “Lac 0 mM”, which were designed to mimic the states of AR and Ad cells, respectively.
Cell lines stably overexpressing the luciferase gene (4T1-luciferase) were generated from 4T1 cells using the LV-17 luciferase lentivirus. DNMT3B overexpressing (DNMT3B) or control (VC) cells were established by transfecting pcDNA3.1(+)–DNMT3B or pcDNA3.1(+) vector control into MCF7/T47D cells.
CD147 knock-in (MCF7/T47D 147) or knockdown (MDA-MB-231 A6) cells were generated by transfecting pGV341 lentiviral CD147 cDNA or pGIPZ lentiviral CD147 shRNA (pGIPZ-A6) into MCF7/T47D or MDA-MB-231 cells, respectively12. The corresponding pGV341 lentiviral expression vector or pGIPZ lentiviral non-target control shRNA (pGIPZ-NTC) transfected cells, MCF7/T47D VC or MDA-MB-231 NTC, were used as control cells12.
CD147 knockout (MCF7/T47D KO) cells were generated by pGV392-U6-sgRNA-EF1a-Cas9-FLAG (pGV392-KO) lentivirus transfection. Cells with CD147 exclusively expressed in mitochondria (MCF7/T47D mito147) were obtained by introducing the mitochondrial CD147 overexpressing pGV371-mito147 lentivirus into MCF7/ T47D KO cells. The mitochondrial CD147 exclusively expressed pGV371-U6-sgRNA-SV40 vector was constructed by cloning the matrix-targeting sequence (MTS, 5′-ATGTCCGTCCTGACGCCGCTGCTGCTGCGGGGCTTGACAGGCTCG GCCCGGCGGCTCCCAGTGCCGCGCGCCAAGATCCATTCGTTG-3′) and the DNA sequence encoding CD147 (NM_198589) into the pGV371 expression vector. pGV392 transfected wild-type cells (MCF7/T47D WT) were used as controls (Table S5).
Rationale for lactate concentration selection
To systematically evaluate the impact of lactate on the stemness potential of detached breast cancer cells, we employed an in vitro concentration range of 0–40 mM, with a primary experimental focus at 20 mM. This range was selected to encompass both physiological serum lactate levels (1.5–3 mM in healthy individuals)39,58 and pathological elevated concentrations observed in human serum (5–8 mM)59,60 and tumor tissues (10–50 mM)59.
For in vivo studies, we administered lactate via intraperitoneal injection at a dose of 800 mg/kg, a concentration previously demonstrated to be non-lethal, without inducing acidosis or severe inflammatory side effects61,62,63. These in vitro and in vivo conditions ensured biologically relevant lactate exposure while maintaining experimental feasibility and reproducibility.
Patient-derived xenograft (PDX) models and tumor cell suspension separation
Breast cancer PDX1/2 were obtained from Suling Liu’s lab64. Both PDX models were derived from triple-negative breast cancer patients. To ensure genetic fidelity and exclude contamination, the identities of the PDX models were rigorously monitored through the establishment a backup library, control of passaging number, and performing short tandem repeat (STR) profiling specificity. Tumor tissue was cut into small pieces (2 × 1 × 1 mm3), and inoculated into the mammary adipose tissue pad of NCG mice. The mice were euthanized when the maximum tumor diameter reached 1 cm. The tumor was removed, minced, and digested with 10 mL/g cell dissociation buffer (DMEM supplemented with 10% FBS, 1% mycillin, 200 U/mL collagenase type II, and 0.6 U/mL dispase) in a 50 mL conical tube at 37 °C for 2 h with vertexing for 1 min every 20 min. The cell suspension was then filtered through a 70 µm strainer (Corning, San Jose, CA, USA) for subsequent experiments.
pGV469-mito-CD147 scFv adenoviral expression vector and virus
Adenoviral constructs targeting and antagonizing mitochondrial localized CD147 were generated by Genechem (Shanghai, China) using the pGV469 plasmid, wherein the gene of interest is expressed under an SV40 promoter. As detailed in Fig. S12, the pGV469-mito-CD147 scFv adenoviral expression vector was constructed by cloning the matrix-targeting sequence (MTS, 5′-ATGTCCGTCCTGACGCCGCTGCTGCTG CGGGGCTTGACAGGCTCGGCCCGGCGGCTCCCAGTGCCGCGCGCCAAGATCCATTCGTTG-3′) of COX VIII and the scFv sequence of HAb1857, a monoclonal antibody targeting CD147, into the GV469 plasmid using Mlu I/Hind III, with EGFP fused at the C terminus; the construct was applied for a Chinese invention patent (202410075009.2).
For in vitro cellular analysis, the pGV469-mito-CD147 scFv expression vector was transfected into cells (6 × 105) at 2.5 μg per well in a six-well plate. For in vivo animal experiments, the pGV469-mito-CD147 scFv expression vector was packaged into adenovirus and injected intratumorally at multiple sites (109 pfu, 30 μL) or intravenously into the tail of mice (108 pfu, 100 μL).
Metabolomics analysis
For metabolomics analysis, supernatants from 1 × 106 MCF7 AR and Ad cells (n = 6 biologically independent samples) were collected and centrifuged at 3000 × g for 10 min at 4 °C to remove cell debris and the lipid layer, followed by untargeted metabolic profiling by Metabo-Profile Inc (Shanghai, China). Briefly, each aliquot of 50 μL of sample was mixed with 10 μL of the internal standard 4-chlorophenylalanine, and then 175 μL of pre-chilled methanol/chloroform (v/v = 3/1) was added. After 20 min of centrifugation at 14,000 × g and 4 °C, 200 μL of supernatant from each sample was evaporated and lyophilized using a FreeZone freeze dryer (Labconco, Kansas City, MI, USA), while the remaining supernatant from each sample was pooled as quality control. A time-of-flight mass spectrometry (GC-TOF/MS) system (Pegasus HT, Kennett Square, PA, USA) with an Agilent 7890B gas chromatograph (Agilent Technologies, Santa Clara, CA, USA) and a Gerstel multipurpose sample MPS2 with dual heads (Gerstel, Germany) was used to investigate the metabolite profiling, with an Rxi-5 ms capillary column (30 m × 250 μm i.d., 0.25-μm film thickness; Restek Corporation, Bellefonte, PA, USA) employed for the separation of metabolites. Metabolite annotation was performed by comparing the retention indices and mass spectral data with those previously generated from reference standards of known structures using ChromaTOF software. The resulting datasets were analyzed using the SIMCA-P 14.1 software package or MetaboAnalyst 5.0 online software (https://www.metaboanalyst.ca/) at the univariate level with an unpaired two-sided nonparametric t-test65. The cut-off for significantly different metabolites between AR and Ad was set to VIP 1.0 and FDR 0.05.
Glucose and fatty acid uptake assay
The uptake of glucose or fatty acids in AR and Ad cells was measured using the fluorescent deoxyglucose analog 2-NBDG (ab287845, Abcam, Cambridge, UK) or TF2-C12 fatty acid (36385, AAT Bioquest, Pleasanton, CA, USA), respectively. Briefly, cells seeded in six-well plates at a density of 8 × 105/2 mL per well were cultivated overnight and then transferred into serum-free medium for 1 h. Subsequently, cells were incubated with 100 μg/mL 2-NBDG in glucose-free medium or with 100 μL/well of the fatty acid dye-loading solution for 1 h. Then, 1 × 106 cells were collected, and the fluorescence signal was measured using a flow cytometer (BD Fortessa, BD Biosciences, Franklin Lakes, NJ, USA) at excitation/emission (Ex/Em) wavelengths of 485/535 nm or 485/515 nm, respectively. The data (mean fluorescence intensity, MFI) were standardized with respect to a reference of 1 × 106 cells.
Extracellular or intracellular glutamine and lactate measurement
Glutamine levels in culture supernatants and cell lysates were detected using a glutamine assay kit (G0429F, Grace Biotechnology, Bend, OR, USA) according to the manufacturer’s protocols. Briefly, AR and Ad cells were cultivated overnight at a density of 8 × 105/2 mL per well, and the corresponding culture supernatants were collected. The cells were disrupted by ultrasound in 200 μL of extraction solution, with the supernatant after centrifugation used for detection, while the culture supernatant was taken for direct detection after centrifugation. After reacting with glutaminase, glutamate dehydrogenase, and color-developing reagents, the absorbance at 450 nm was measured, and the glutamine content was calculated according to the standard curve. The glutamine levels in culture supernatants were normalized to 1 × 106 cells, while those in cell lysates were standardized against total cell protein levels.
Lactate levels in mouse serum and culture supernatants were tested using a lactic acid assay kit (A019-2-1, Jiancheng Bio, Nanjing, China), with lactate levels in cellular supernatants normalized to 1 × 106 cells. In cell lysates, lactate levels were assayed using an L-lactate assay kit (ab65331, Abcam), with total cell protein levels used for data normalization. Serum from 1 × 105 4T1-luciferase cell-bearing Balb/c mice was collected from 100 μL of blood every 4 days starting from the 5th day after inoculation. Cells and the corresponding culture supernatants from AR/Ad cells were collected similarly to the procedures for glutamine measurement, while samples from lactate-treated cells were collected by culturing cells in glucose-free medium with 20 mM lactate incubation for 1, 2, 4, 8, 12, 24, 36, and 48 h.
Proteomics analysis by bottom-up mass spectrometry
1 × 106 AR/Ad or with/without lactate-treated MCF7/T47D cells (n = 3 biologically independent samples) were lysed with RIPA lysis buffer, and the protein lysates were electrophoresed on 10% SDS-PAGE gels. After Coomassie Brilliant Blue staining, five equal parts of the gels, divided by molecular weight, were evaporated using acetonitrile and reconstituted in 1% (v/v) formic acid, then digested with trypsin and injected into the mass spectrometer (Model LTQ, Thermo Fisher Scientific, Waltham, MA, USA). The mass spectrometer (MS) was calibrated using standard compounds and operated in data-dependent mode, cycling between full MS scans (m/z 300–2000), while the MS data were collected by targeting MS scans on the ten most abundant ions detected in the MS scan.
MS raw data were processed using Proteome Discoverer software (version 2.5.0.4) for database searching, quantification, and false discovery rate (FDR) estimation. Spectra were queried against the UniProt Homo sapiens reference proteome database (release 2023_08, containing 20349 sequences). The search parameters were set as follows: Trypsin/P was specified as the protease with up to 2 missed cleavages permitted. Precursor mass tolerance was set to 10 ppm, while the fragment mass tolerance was defined as 0.02 Da. The minimum required peptide length was set to 6 amino acids. Methionine oxidation and protein N-terminal acetylation and lysine methylation were specified as variable modifications, while cysteine carbamidomethylation was set as the stable modification.
Site possibility threshold for peptide-to-spectrum matching (PSM) was set to minimum 75 for PTM site localization. To ensure high-confidence identifications, the false discovery rate (FDR) was controlled at <1% at both the peptide-spectrum match level and the protein level. This was determined by searching against a reversed decoy database. Finally, protein identification required at least two unique peptides per protein group. Three biologically independent samples were conducted for each cell, and proteins with variable importance in projection (VIP) of ≥1 and P < 0.05 were considered differentially expressed proteins.
Western blot
Tumor tissues and cells were lysed in RIPA lysis buffer supplemented with protease inhibitors (4693159001, Roche, Basel, Switzerland). Protein concentration was determined using a BCA protein assay kit (Thermo Fisher Scientific), and 20 μg of protein per sample (the default amount if not specified otherwise) were subjected to SDS-PAGE, transferred onto PVDF membranes, and incubated with primary antibodies summarized in the key resources table, including GLUT1 (1:1000), MCT1 (1:1000), LDHB (1:1000), SOX2 (1:1000), GPR81 (1:1000), DNMT3B (1:1000), H3 (1:1000), H3K27m3 (1:1000), COX IV (1:1000), TOMM20 (1:1000), Na+-K+ ATPase (1:1000), LAMP1 (1:1000), GM130 (1:1000), GRP78 (1:1000), α-Tubulin (1:5000), and anti-CD147 (HAb18, 1:3000) produced in our laboratory54,66. After incubation with horseradish peroxidase (HRP)-conjugated secondary antibodies (Thermo Fisher Scientific, 1:5000), the proteins were visualized using chemiluminescence reagents (29050, Engreen, San Jose, CA, USA) with a Gel Doc EZ Imager (Bio-Rad Laboratories, Hercules, CA, USA) and analyzed using the Image Lab software (Bio-Rad Laboratories).
RT-PCR and PCR array
Total RNA was isolated using an E.Z.N.A. Total RNA Kit II (OMEGA, BioTek, Norcross, GA, USA), and 1 μg of RNA was subjected to reverse transcription with the PrimerScriptTM RT Master Mix kit (RR036A, TaKaRa, Otsu, Japan). Subsequently, RT-PCR was performed with a SYBR Green PCR kit (TaKaRa) on the QuantStudioTM 7 Flex Real-Time PCR system (Life Technologies, Thermo Fisher Scientific) to detect the expression levels of the target genes. Gene expression was quantified using the ΔCt method, normalized to 18sRNA. All primers were synthesized by Tsingke Biotech (Beijing, China), and the primer sequences are shown in Table S1.
For the human stem cell transcription factors PCR array (PAHS-501Z, Qiagen, Duesseldorf, Germany), the gene expression of interest was normalized relative to housekeeping genes (ACTB, B2M, HPRT1, RPLP0; Supplementary data 1). Fold changes in gene expression, scatterplots, and heatmaps were analyzed and generated according to the RT2 PCR array data analysis website (https://geneglobe.qiagen.com/us/analyze).
13C3-lactate metabolic flux analysis
1 × 107 MCF7 AR/Ad cells (n = 6 biologically independent samples) cultivated in regular complete RPMI 1640 medium containing 10 mM 13C3-lactate for 12 h were pelleted, stored at −80 °C, and then sent to Metabo-Profile Inc (Shanghai, China) for metabolic flux assay using ultra-performance liquid chromatography triple-quadrupole mass spectrometry (UPLC-TQ-MS), with unlabeled AR and Ad cells collected at 0 h being used to correct for the natural isotopic abundance that could potentially contribute to false positives.
For the disruption of cell pellets, 400 μL of 80% methanol solution was added, followed by ultrasonic disruption and centrifugation at 4 °C at 18,000 × g for 15 min. The supernatant was concentrated (Labconco), dissolved in 100 μL of 80% methanol solution, and then prepared for injection and analysis. Metabolites were separated using an Acquity amide and an Acquity HSS C18 column (2.1 mm × 100 mm, 1.7 μm, Waters Corp) equipped with an ACQUITY UPLC VanGuard Pre-Column (Waters Corp, Milford, MA, USA). A 5 μL aliquot of the sample was injected into the column, with the temperature maintained at 40 °C and the flow rate set constantly at 0.4 mL/min. The raw UPLC-MS data were analyzed using TargetLynx Application Manager software version 4.1 (Waters Corp). Quantification of each metabolite was performed using linear regression analysis of the peak area of the metabolite versus concentration.
The mass distribution vector (MDV) of 13C-labeled metabolites, which describes the fractional abundance of each isotopologue normalized to the sum of all possible isotopologues, was measured by correcting for naturally occurring isotopes according to a previous publication67. First, the proportion (P0) of natural 13C-labeled metabolite relative to non-13C-labeled metabolite in MCF7 cells collected at 0 h was calculated as: P0 = abundance of natural 13C-labeled metabolite0 h/abundance of non-13C-labeled metabolite0 h; while the abundance of the natural 13C-labeled metabolite in MCF7 cells cultured with 13C3-lactate for 12 h was calculated using the following formula: abundance of natural 13C-labeled metabolite = abundance of non-13C-labeled metabolite×P0. Then, the abundance of the natural 13C-labeled metabolite was subtracted from the total 13C-labeled metabolite in MCF7 cells cultured with 13C3-lactate. Finally, the MDVs of 13C-labeled metabolites were obtained using the following formula: MDVs = (abundances of 13C-labeled metabolites—abundances of natural 13C-labeled metabolites)/(abundances of total metabolites—abundances of natural 13C-labeled metabolites).
Flow cytometry
The analysis of cancer stem cell (CSC) ratios was performed using the ALDEFLUOR assay and CD44highCD24–/low staining. In the ALDEFLUOR assay (01700, STEMCELL Technologies, Vancouver, CA), functional stem cells were analyzed and isolated based on the fact that aldehyde dehydrogenase (ALDH) is a widely recognized key marker for breast CSCs68. Specifically, cells with ALDH enzymatic activity (ALDH+) are considered CSCs, while those without ALDH enzymatic activity (ALDH−) are non-CSCs. The detailed procedures for sorting ALDH− non-CSCs are described in our paper12. Immediately following sorting, these ALDH− cells were directly utilized in the subsequent experiments without undergoing any additional passaging.
For in vivo analysis of CSC ratios in tumors, PE anti-mouse EPCAM and APC anti-mouse CD45 staining were used to distinguish tumor cells from other cell populations. The CSC ratio analysis was performed on a BD LSRFortessaTM cell analyzer, and cell sorting was conducted on a BD FACS AriaTM III. Data collection and analyses were carried out using BD FACSDiVaTM and FlowJo 7.6 software. Corresponding flow cytometry gating plots were provided in the supplementary information file.
siRNA knockdown
The siRNAs targeting MCT1, LDHB, and GPR81, along with the universal negative control siRNA (Table S1), were synthesized by Sangon Biotech (Shanghai, China) and transfected into cells using Lipofectamine 2000™ reagent according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA, USA). After 48 h, total RNA and whole cell lysates were extracted to assess the efficiency of siRNA interference using RT-PCR and western blot, respectively.
Tumorsphere formation assay
The tumorsphere formation assay was performed as described in our previous paper12. Briefly, 2 × 105 cells were cultured in ultra-low attachment six-well plates using serum-free DMEM/F12 medium supplemented with 1% N2, 2% B27, 15% glucose, 1 µg/mL hydrocortisone, 5 µg/mL insulin, 10 ng/mL epidermal growth factor (EGF), 10 ng/mL basic fibroblast growth factor (bFGF), and 0.2% heparin (Table S5). The number of spheres with a diameter greater than 50 μm was monitored and quantified under an inverted microscope (Olympus, Tokyo, Japan) on days 10–14. In lactate-treated cells, the DMEM/F12 medium was replaced with glucose-free DMEM/F12 medium (Procell, Canton, MA, USA), and the addition of 15% glucose was omitted.
Immunofluorescence (IF) staining
Ad cells cultured on glass-bottom dishes (Cellvis, Mountain View, CA, USA) or AR suspended cells mounted on adhesive slide were fixed with 4% paraformaldehyde, permeabilized for 5 min with 0.2% Triton X-100, blocked with 10% goat serum for 30 min, and then incubated overnight at 4 °C with primary antibodies against 5-mC (1:1000; 28692, Cell Signaling Technology, Danvers, MA, USA) for methylation detection, or against CD147 (1:200) and COX IV (a mitochondrial marker; 1:200; 11242-1-AP, Proteintech, Rosemont, IL, USA) to observe the mitochondrial localization of CD147 and the binding of mito-CD147 scFv to mitochondrial CD147. After incubation with fluorescently labeled secondary antibodies, nuclei were counterstained with DAPI (Invitrogen). All images were captured using laser scanning confocal microscopy (LSCM, Olympus FV3000) and collected using FV31S-SW 2.4.1 with consistent settings across all samples, and fluorescence intensity was quantified using Image J 1.50i (NIH, Bethesda, MD, USA).
For cellular co-localization analysis among CD147, MCT1, LDHB, and COX IV, cells were initially stained with primary antibodies against CD147 (1:200), MCT1 (1:200; 20139-1-AP, Proteintech), and LDHB (1:100; ab195964, Abcam) overnight at 4 °C, followed by incubation with Alexa Fluor 555-, 647- or 405-conjugated antibodies, respectively, at room temperature for 1 h. This was followed by incubation with an Alexa Fluor 488-conjugated COX IV antibody (1:200; CL488-60251, Proteintech) at room temperature for 3–4 h. Finally, nuclei were counterstained, and images were captured following the same procedures as described above. For human or mouse tumor samples, tissues were fixed, embedded, sectioned, and antigen retrieval was performed using the same procedures as outlined in the Histological Staining and Analysis section below, and stained following the same protocols as for cells.
Histological staining and analysis
Human or mouse breast primary or lung metastatic tumor tissues were fixed, embedded in paraffin, and cut into 4-µm-thick sections. Sections were deparaffinized, dehydrated, and cleared in xylene and gradient alcohol, then used for hematoxylin-eosin (H&E) or immunohistochemical (IHC) staining.
For H&E staining, the slides were counterstained with hematoxylin for 2 min and eosin for 3 min.
For IHC staining, the slides underwent antigen retrieval with 10 μM citrate buffer (pH 6.0) in pressure cookers at 100 °C for 2 min, followed by blocking with 5% goat serum and staining with anti-SOX2 antibody (1:100; 11064-1-AP, Proteintech) for 2 h at room temperature. Next, the slides were stained with immunoperoxidase using a General SP kit (SP-9000, ZSGB-BIO, Beijing, China), developed with 3,3′-diaminobenzidine (ZLI-9019, ZSGB-BIO), and counterstained with hematoxylin. To evaluate the expression of SOX2, IHC scores were calculated by two independent pathologists according to the following equation: score = 3 (strong positive) × percentage value + 2 (moderate positive × percentage value + 1 (weak positive) × percentage value + 0 (negative) × percentage value. The percentage index 0–4 was defined using the following criteria: 0 (< 10% positive cells), 1 (10–25% positive cells), 2 (25–50% positive cells), 3 (50–75% positive cells), and 4 (75–100% positive cells).
For multiplex immunohistochemical (mIHC) analysis, the deparaffinized, dehydrated, and antigen-retrieved tissues were permeabilized with 0.5% Triton X-100, stained with primary antibodies, and detected using a TSA-indirect kit (PerkinElmer) according to the manufacturer’s instructions. Nuclei were stained with DAPI. The image was scored using HALO software (Indica Labs, USA). During data analysis, the immunostained sections were divided into high- and low-expression groups, with the mean score value serving as the cut-off. High expression was defined as scores of CD147 ≥ 29.5, scores of MCT1 ≥ 17.5, and scores of LDHB ≥ 4.5; scores below these thresholds were considered as low expression.
Circulating tumor cells isolation, counting, and IF staining
Circulating tumor cells (CTCs) were isolated using EPCAM+ magnetic beads (130-105-958, Miltenyi Biotec, Bergisch Gladbach, Germany). Briefly, 800 μL of blood collected from mice was lysed to remove red blood cells and incubated with EPCAM+ magnetic beads at 4 °C for 15 min. Subsequently, EPCAM+ cells that adhere to the LS column in a magnetic field were eluted. The identification and counting of CK18+CD45− CTCs, as well as the examination of SOX2 levels in CTCs, were conducted through IF staining using the following primary antibodies: anti-CD45 (1:200, 60287-1-Ig, Proteintech), CoraLite® Plus 488-conjugated anti-CK18 (1:200, CL488-66187, Proteintech), and CoraLite® Plus 647-conjugated anti-SOX2 (1:200, CL647-66411, Proteintech). Additionally, Alexa Fluor 555 donkey anti-mouse IgG (H + L) (1:200, A31570, Invitrogen) was used to visualize CD45 expression. Finally, IF signals were quantified using Image J 1.50i (NIH).
ATP measurement
A quantity of 1 × 106 cells was lysed to detect cellular ATP levels using an ATP bioluminescence assay kit (S0026, Beyotime) according to the manufacturer’s instructions. The bioluminescence signal generated by luciferase was measured as relative light units (RLU) in a luminometer (Promega, USA Gloma), and the ATP levels were calculated using a standard curve.
Oxygen consumption rate measurement
Cellular oxygen consumption rate (OCR) was measured using the XF96 Flux Analyzer (Agilent Technologies). Briefly, Ad cells were seeded into pre-hydrated 96-well plates (1.5 × 104 cells/well) and cultivated overnight, while AR cells at the same density were seeded directly into poly-L-lysine precoated wells just before the measurement. For lactate-treated cells, they were first cultured in glucose-free media without or with 20 mM lactate for 48 h. Subsequently, an equal number of cells (1.5 × 104 cells per well) were seeded into pre-hydrated 96-well plates and allowed to adhere for 6 h. Then, AR/Ad cells were incubated at 37 °C for 45 min without CO2 in seahorse assay medium containing 10 mM glucose, 2 mM L-glutamine, and 1 mM sodium pyruvate, while the lactate-treated cells were incubated at 37 °C for 45 min without CO2 in a substrate-limited medium containing 0.5 mM glucose, 0.5 mM L-glutamine, and 0.2 mM sodium pyruvate. OCR was measured by adding inhibitors at the following times: oligomycin (14 min, 1 μM), FCCP (33 min, 1 μM), and rotenone/antimycin A (53 min, 0.5 μM). Three readings were taken after each injection. Cells in each well were subsequently trypsinized and counted, and OCR values were normalized to the corresponding cell number per well. Data were collected using WAVE 2.6.1 (Alignment) and final results were presented as pmoles/min for OCR (y-axis) versus time (x-axis).
Additionally, OCR was detected using a commercial kit (600800, Cayman Chemical, Ann Arbor, MI, USA) following the manufacturer’s protocols. Briefly, cells at a density of 5 × 105/2 mL per well were cultivated in a six-well plate for 48 h. Subsequently, MitoXpress-Xtra solution was added, and the plate was read immediately at 650 nm for 2 h using a spectrophotometer (BioTek Instruments, Winooski, VT, USA). The OCR was calculated as (mOD1-mOD2)/T, where mOD1 is the initial absorbance, mOD2 is the final absorbance, and T is time in minutes.
Mitochondrial complex activity assay
The activities of mitochondrial complexes were measured using the enzyme activity microplate assay kit (Abcam). Briefly, 1 × 107 cells were homogenized, and total protein concentrations were adjusted to 5.5 mg/mL with PBS. To extract the proteins, 1/10 volume of detergent solution was added; then the supernatant from lysed cells was incubated at room temperature for 2–3 h in a 96-well microplate precoated with monoclonal antibodies to complexes I, II, or IV. After adding the specific substrates, absorbances were measured continuously using a spectrophotometer (BioTek Instruments) as follows: complex I (450 nm, 30 min), complex II (600 nm, 2 h), and complex IV (550 nm, 2 h). The enzyme activity of each complex (mOD/min), expressed as the reaction rate, was calculated by (mOD1- mOD2)/T, similar to the calculation in the OCR assay mentioned above.
α-ketoglutarate (α-KG) measurement
α-KG levels were measured using an α-ketoglutarate assay kit (ab83431, Abcam). Briefly, lysates of 1 × 106 cells were deproteinized and incubated with the enzyme reaction mixture for 30 min. The α-KG level was calculated by measuring optical density at 570 nm in a spectrophotometer (Bio-Rad Laboratories) using a standard curve.
5-methelcytosine (5-mC) DNA dot blot
To detect global DNA methylation levels, genomic DNA was extracted and purified from cells using a DNA extraction kit (DP304, TIANGEN BIOTECH, Beijing, China), followed by quantification, sonication, and denaturation. Next, 1 μg of DNA was transferred to a PVDF membrane and incubated successively with a 5-mC primary antibody (1:1000; 28692, Cell Signaling Technology) and an HRP-conjugated secondary antibody. The chemiluminescence signals were then visualized with a Gel Doc EZ Imager (Bio-Rad Laboratories) and analyzed using Image Lab 5.2.1 (Bio-Rad Laboratories).
Mitochondrial isolation
The mitochondria were isolated from 2 × 107 cells using the Minute™ Mitochondria Isolation Kit (MP-007, Invent Biotechnologies, Plymouth, MN, USA), which employs protein extraction filter cartridges and differential centrifugation. The isolated mitochondria were stored in Minute™ denatured protein solution for western blot analysis or in Minute™ nondenatured protein solution for co-immunoprecipitation. The purity of the mitochondrial fractions was assessed by performing western blot analysis in a concentration-dependent manner (2, 10, 20, and 50 μg) using antibodies against COX IV, α-tubulin, Na+/K+-ATPase, LAMP1, GM130, and GRP78 to indicate the fractions of mitochondria, cytoplasm, plasma membrane, lysosome, Golgi apparatus, and endoplasmic reticulum, respectively.
Mitochondrial sub-fractionation isolation and protease protection assay
The sub-fractionation of mitochondria was performed according to Huang’s report69. Briefly, 1 mg of mitochondria (measured by protein content) suspended in 40 μL of T10E20 buffer (10 mM Tris-HCl, 1 mM EDTA, pH 7.6) was incubated with 0.1 mg of digitonin for 20 min. After adding 120 μL of 250 mM sucrose, the mitochondria were centrifuged at 10,000 × g for 15 min. The fraction of the mitochondrial inner membrane and matrix (I/M) was collected from the pellet, while the outer membrane and intermembrane (O/I) were obtained from the supernatant.
For further mitochondrial fractionation experiments, protease protection assays were conducted by incubating intact mitochondria with 0.5 μg/mL of proteinase K (K) for 10 min, either alone or following a 5 min pretreatment with 0.2% Triton X-100, and then subjected to western blotting. TOMM20 or COX IV were used to indicate proteins located in the outer or inner mitochondrial membrane, respectively.
Blue native PAGE analysis
Blue native PAGE analysis was performed as described previously62,70. In brief, mitochondrial proteins were isolated using the procedures detailed in the Mitochondrial Isolation section. After isolation, 30 μg of mitochondrial proteins were loaded onto a 4–13% Bis-Tris Native PAGE gel (P0545S, Beyotime) for electrophoresis-based separation. Then the gel was stained with Coomassie Brilliant Blue to visualize the protein bands.
Mitochondrial mass analysis
Mitochondrial mass was assessed via flow cytometric analysis of cells stained with MitoTracker Green FM (M46750, Thermo Fisher Scientific). Briefly, cells were incubated with 100 nM Mitotracker Green FM for 30 min at 37 °C. Subsequently, the cells were rinsed with PBS, detached using trypsin, and then centrifuged at 130 × g for 4 min. The resulting cell pellets were resuspended in PBS supplemented with 1% FBS for FACS. Given that MitoTracker Green FM has an excitation wavelength of 490 nm and an emission wavelength of 516 nm, the FITC channel, with an excitation at 494 nm and emission at 520 nm, was used to detect the signal. Data analyses were performed using FlowJo 7.6 software (Becton, Dickinson and Company, Franklin Lakes, NJ, USA).
Pyruvate assay
Pyruvate levels were measured using a pyruvate colorimetric assay kit (E-BC-K-130-S, Elabscience). Briefly, 20 µL of cytoplasm or mitochondrial subfraction, along with standard and distilled water (blank control), were incubated with 100 µL of chromogenic agent at 37 °C for 10 min. Subsequently, 500 µL of alkali reagents were added to develop color for 5 min at room temperature. The absorbance values for the cytoplasm or mitochondrial subfraction and the standard at 505 nm were obtained by subtracting the absorbance value of the blank control. Pyruvate content was then calculated based on the standard curve and normalized to the protein concentration.
Methylation assay of the SOX2 promoter region
The methylation level in the SOX2 promoter region was assessed using a fast bisulfite conversion kit (ab117127, Abcam) according to the manufacturer’s instructions. Briefly, genomic DNA from 1 × 106 cells was extracted (DP304, TIANGEN), purified, and quantified. Two hundred ng of DNA was denatured and bisulfite converted at 95 °C for 20 min, followed by methylation-specific real-time PCR using three pairs of primers designed with MethPrimer that targeted the CpG islands in the SOX2 promoter region (Table S1). The relative methylation level was calculated as the ratio of the Ct value for methylated DNA to the sum of the Ct values for both methylated and unmethylated DNA.
DNMT3B enzyme activity
The enzyme activity of DNMT3B was measured using a DNMT3B direct activity assay kit (ab113471, Abcam, Cambridge, UK). Briefly, lysates of 1 × 106 cells were incubated with S-adenosylmethionine donor at 37 °C for 2 h in a plate pre-coated with DNA substrate. This was followed by successive incubations with anti-5-mC primary antibody, HRP-labeled secondary antibody, and chemiluminescent substrate. Relative DNMT3B enzyme activity was calculated as the ratio of luminescence values measured using a Bio-Tek fluorescent microplate reader.
Chromatin immunoprecipitation (ChIP)
ChIP assays were performed using a SimpleChIP® Plus Enzymatic Chromatin IP Kit (Magnetic Beads, 9005s, Cell Signaling Technology). Briefly, 4 × 106 cells were crosslinked in 1% formaldehyde at room temperature for 10 min, with the reaction quenched by 2.5 M glycine. Next, cells were lysed, and chromatin was extracted by incubating with micrococcal nuclease at 37 °C for 20 min, followed by brief sonication. The resulting cell lysates (input) were immunoprecipitated at 4 °C for 12–16 h with 3 µg of anti-5-mC (28692, Cell Signaling Technology) and anti-DNMT3B (57868, Cell Signaling Technology). The immunocomplexes were then rotationally incubated with 30 µL of ChIP-Grade Protein G Magnetic Beads at 4 °C for 2 h, and the ChIP DNA was eluted using ChIP elution buffer at 65 °C for 30 min. After reverse cross-linking and proteinase K/RNase treatment, ChIP DNA was purified, amplified, and quantified by RT-PCR. The PCR results for DNA methylation were presented as the percentage of ChIP DNA relative to total input DNA (2 (Ct input–Ct sample)), with the Ct values detected by the indicated antibodies normalized to a Ct value obtained from control ChIP performed with nonspecific rabbit IgGs. The specific primer pair for the SOX2 promoter region is listed in Table S1.
2-2H-lactate metabolic flux analysis
Overall, 2 × 107 MCF7 AR and Ad cells (n = 3 biologically independent samples) were cultured in medium containing 10 mM 2-2H-lactate (693987, ≥ 98% (CP), Sigma) for 6 h. The mitochondria were isolated, verified for purity, and then sent to Applied Protein Technology (Shanghai, China) for targeted LC/MS analysis of 2H-labeled metabolites.
2H-labeled metabolites in the mitochondria were extracted by adding 500 μL of cold extraction buffer (methanol: acetonitrile: water = 2:2:1, v/v/v), followed by sonication for 2 min and centrifugation at 14,000 × g for 5 min at 4 °C. The supernatants were thoroughly lyophilized and reconstituted in 50 μL of methanol-water (1:1, v/v) just prior to measurement. A mixture of equal amounts of supernatant from individual samples, analyzed using the same procedure for experimental samples, was used as quality control.
2H-labeled metabolites in the mitochondria were analyzed with LC-HRMS using Thermo Fisher Q-Exactive mass spectrometry, equipped with a Vanquish UPLC system (Thermo Fisher, MA, US), employing a SeQuant ZIC-HILIC column (100 mm × 2.1 mm, i.d. 3.5 μm, Merck, Germany). A 1 μL aliquot of supernatant extracted from the mitochondria was injected into the column, with the temperature maintained at 35 °C and the flow rate set at 0.4 mL/min. The mass spectrometry operated in both negative and positive modes with a capillary voltage of 2.0 kV. The source temperature and desolvation gas temperature were set at 120 and 500 °C, respectively.
Raw peak data were processed using TraceFinder 5.0 and Xcalibur 4.0 software (Thermo Fisher Scientific), with peak alignment and detection performed according to the manufacturer’s protocol. The method “peak alignment and frame extraction” was applied for targeted metabolite analysis. An input file containing theoretical m/z values and detected retention times was used for targeted metabolite analysis, with the m/z width set to 5 ppm. An output file was generated after data processing, which included detected m/z values and relative intensities across different samples. Metabolite mass isotopomer distribution (MID, also known as MDV) was determined based on the ratio of the integrated peak areas of the chosen isotopomer to the sum of all integrated peak areas of the possible isotopomers for the given metabolites. Correction for natural abundance is necessary for meaningful interpretation. The natural isotopic correction was performed by treating a control group of unlabeled samples (mitochondria isolated from cells collected at 0 h) using both a generally applicable correction matrix and an isotopic correction matrix, as previously reported71,72.
Co-immunoprecipitation (Co-IP) assay
Mitochondria were isolated following the protocol described in the Methods Section under Mitochondrial Isolation. Approximately 500 μg of the mitochondrial pellet was resuspended in 200 μL of nondenatured lysis buffer (WA-010, Invent Biotechnologies), a mild formulation designed to preserve native protein-protein interactions within the mitochondria while solubilizing mitochondrial membranes. The suspension was gently pipetted to ensure homogeneity, followed by centrifugation at 12,000 × g for 2 min to remove insoluble components. The resulting supernatant was then subjected to a Co-IP experiment according to standard procedures. Briefly, the supernatant was incubated with 30 μg of primary antibodies to CD147, MCT1 (20139-1-AP, Proteintech), or LDHB (14824-1-AP, Proteintech) at 4 °C overnight, with irrelevant mouse IgG (P5000-Y, GENIA) or rabbit IgG (I8140, Sigma) used as negative controls. The following day, Protein A&G beads (Invitrogen) were added and incubated at 4 °C for 4 h. The immuno-precipitates were then eluted and identified by western blot, with the isolated mitochondria used as an input control.
Transmission electron microscope
For colloidal gold staining, 2 × 106 cells were fixed with PLP Fixing Solution (G2220, Solarbio, Beijing, China) at 4 °C for 24 h and then subjected to dehydration and embedding. Ultrathin sections of the cells were transferred onto grids, blocked with goat serum for 30 min at room temperature, and incubated overnight at 4 °C with primary antibodies against CD147 (mouse, 1:100), MCT1 (rabbit, 20139-1-AP, Proteintech, 1:100), and LDHB (goat, ab195964, Abcam, 1:100). This was followed by a 1-h incubation at room temperature with 10 nm gold-conjugated anti-mouse IgG (ab39593, Abcam, 1:200), 20 nm gold-conjugated anti-rabbit IgG (ab270563, Abcam, 1:200), and 35 nm gold-conjugated anti-goat IgG (K0038D-Gold, Solarbio, 1:200). Finally, the sections were consecutively counterstained with 5% uranium acetate and lead acetate and observed under an EM-1230 transmission electron microscope (JEOL Ltd., Tokyo, Japan).
Proximity ligation assay (PLA)
The Duolink® in situ assay kit (DUO92008, Sigma) was utilized according to the manufacturer’s instructions. In Brief, MCF7 cells were treated with either 0 or 20 mM lactate and then cultured on small round cover glasses. Subsequently, the cells were fixed with 4% paraformaldehyde and permeabilized with 0.2% Triton X-100. After being blocked with the Duolink® blocking solution, the cells were incubated overnight at 4 °C with mouse monoclonal anti-CD147 (produced in our laboratory, dilution 1:200) and rabbit polyclonal anti-LDHB (14824-1-AP, Proteintech, dilution 1:100). Next, the PLA probe solution was added, and the cells were incubated in a pre-heated humidity chamber at 37 °C for 1 h. After thorough washing, the ligation solution was applied for 30 min at 37 °C, followed by the addition of the amplification solution for 100 min at 37 °C. Afterward, DAPI was used to stain the cell nuclei for 15 min at RT. Images were captured via laser scanning confocal microscopy (LSCM, Olympus FV3000) and analyzed using the ImageJ software.
Molecular docking simulations
The molecular docking simulations were performed using AutoDock 4.273. Briefly, PDBQT files were generated for CD147, MCT1, and LDHB with Gasteiger partial charge assigned, and the AutoGrid algorithm was employed to precalculate the atomic affinity grid for docking calculations. Candidate complexes were selected based on binding energy scores and geometric complementarity, the latter evaluated through cluster analysis. The detailed molecular interactions were analyzed using LIGPLOT version 4.4.274, with hydrophobic interactions defined as ligand-protein residue hydrophobic atom distances <3.9 Å, while hydrogen bonds were identified when (i) the donor-acceptor pair matched predefined criteria and (ii) bond angles and distances met default thresholds.
Expression plasmids Construction, protein expression, and purification
The cDNA sequences encoding the extracellular domain of CD147 (1–205 aa) and the full-length LDHB were cloned into the pGEX-6P-1 and pET-28a plasmids, respectively. Subsequently, GST-CD147 N-terminal and His-LDHB proteins were expressed using BL21 (DE3) bacteria cells. Specifically, bacteria cells carrying the plasmid encoding these proteins were initially incubated at 37 °C for 6 h, then the expression was induced with 0.5 mmol/L isopropyl-β-d-thiogalacto-pyranoside (IPTG) at 16 °C overnight. After that, cell lysates were collected by centrifugation at 13,500 × g for 30 min at 4 °C. Prior to use, appropriate beads were pre-washed with PBS and equilibrated in lysis buffer. Glutathione Sepharose Beads (SMART Lifesciences, Changzhou, China) and His60 Ni Superflow Resin (TaKaRa, Japan) were incubated with cell lysates at 4 °C for 4 h with continuous rotation. Following incubation, the beads were washed with lysis buffer, and the proteins were eluted with GST elution buffer (75 mmol/L Tris-HCl pH 8.0, 20 mmol/L-Glutathione) and His elution buffer (50 mmol/L NaH2PO4, 300 mmol/L NaCl, 250 mmol/L imidazole, adjust pH to 8.0), respectively.
For the expression of proteins with specific-sites mutations, the codons in the cDNA sequence corresponding to the target sites were replaced with alanine-encoding codons. Subsequently, the same experimental procedure as described above was followed.
GST pull-down assay
GST pull-down assays were performed following the instructions provided by the manufacturer of the Pierce GST Protein Interaction Pull-Down Kit (21516, Thermo Fisher Scientific). First, the glutathione agarose was equilibrated. Then, 100 μg of the purified GST-CD147 N-terminal protein or 100 μg of the GST protein alone, each in a 300 μL volume, were incubated with glutathione-agarose beads at 4 °C for 1 h. After the incubation, the beads were washed with the wash buffer. Subsequently, 100 μg of the purified His-LDHB were added to the column and incubated at 4 °C for 2 h. After another round of washing with the wash buffer, the eluted protein was subjected to immunoblotting analysis using HRP-conjugated rabbit polyclonal anti-GST (1006-3, Huabio, dilution 1:500) or mouse monoclonal anti-His (66005-I-Ig, Proteintech, dilution 1:1000) antibodies, respectively.
Statistics and reproducibility
All data were presented as mean ± SEM from at least three independent experiments, and statistical analyses were performed using GraphPad Prism 8.0 software. To determine statistical significance, Student’s t-test or one-way ANOVA was conducted to compare continuous variables between two groups or more than two groups, while the χ2 test was used for categorical variables. Spearman correlation analysis was employed to investigate the correlation between the expressions of CD147 and MCT1 or LDHB. The Kaplan–Meier method and the log-rank tests were applied to compare survival and recurrence times between patients with high or low CD147/MCT1/LDHB co-expression. The multivariate Cox proportional hazards model was used to explore independent prognostic predictors for the overall survival of patients with breast cancer. For all statistical analyses, p-values less than 0.05 were considered significant.
Data availability
The protein mass spectrometry data generated in this study have been deposited in ProteomeXchange under the accession codes PXD059617. The metabolomics data generated in this study have been deposited in Metabolomics Workbench under the accession codes PR002721 [https://doi.org/10.21228/M8XV8X]. Source data are provided with this paper.
References
Breast cancer. https://www.who.int/news-room/fact-sheets/detail/breast-cancer. (2024).
Lambert, A. W., Pattabiraman, D. R. & Weinberg, R. A. Emerging biological principles of metastasis. Cell 168, 670–691 (2017).
Liang, Y. et al. Metastatic heterogeneity of breast cancer: molecular mechanism and potential therapeutic targets. Semin. Cancer Biol. 60, 14–27 (2020).
Chambers, A. F., Groom, A. C. & MacDonald, I. C. Dissemination and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2, 563–572 (2002).
Celià-Terrassa, T. & Kang, Y. Distinctive properties of metastasis-initiating cells. Genes Dev. 30, 892–908 (2016).
Perez-Gonzalez, A., Bevant, K. & Blanpain, C. Cancer cell plasticity during tumor progression, metastasis and response to therapy. Nat. Cancer 4, 1063–1082 (2023).
Oskarsson, T., Batlle, E. & Massague, J. Metastatic stem cells: sources, niches, and vital pathways. Cell Stem Cell 14, 306–321 (2014).
Bergers, G. & Fendt, S. M. The metabolism of cancer cells during metastasis. Nat. Rev. Cancer 21, 162–180 (2021).
Loh, J. J. & Ma, S. Hallmarks of cancer stemness. Cell Stem Cell 31, 617–639 (2024).
Canellas-Socias, A. et al. Metastatic recurrence in colorectal cancer arises from residual EMP1(+) cells. Nature 611, 603–613 (2022).
Fumagalli, A. et al. Plasticity of Lgr5-negative cancer cells drives metastasis in colorectal cancer. Cell Stem Cell 26, 569–578.e7 (2020).
Meng, Y. et al. Detachment activated CyPA/CD147 induces cancer stem cell potential in non-stem breast cancer cells. Front. Cell Dev. Biol. 8, 543856 (2020).
Taddei, M. L. et al. Anoikis: an emerging hallmark in health and diseases. J. Pathol. 226, 380–393 (2012).
Ganesh, K. & Massague, J. Targeting metastatic cancer. Nat. Med. 27, 34–44 (2021).
Massagué, J. & Ganesh, K. Metastasis-initiating cells and ecosystems. Cancer Discov. 11, 971–994 (2021).
Davis, R. T. et al. Transcriptional diversity and bioenergetic shift in human breast cancer metastasis revealed by single-cell RNA sequencing. Nat. Cell Biol. 22, 310–320 (2020).
Schafer, Z. T. et al. Antioxidant and oncogene rescue of metabolic defects caused by loss of matrix attachment. Nature 461, 109–113 (2009).
Jiang, L. et al. Reductive carboxylation supports redox homeostasis during anchorage-independent growth. Nature 532, 255–258 (2016).
Grassian, A. R. et al. Erk regulation of pyruvate dehydrogenase flux through PDK4 modulates cell proliferation. Genes Dev. 25, 1716–1733 (2011).
Zhang, C., Quinones, A. & Le, A. Metabolic reservoir cycles in cancer. Semin. Cancer Biol. 86, 180–188 (2022).
Faubert, B. et al. Lactate metabolism in human lung tumors. Cell 171, 358–371.e9 (2017).
Hui, S. et al. Glucose feeds the TCA cycle via circulating lactate. Nature 551, 115–118 (2017).
Yang, L. et al. Nucleolin lactylation contributes to intrahepatic cholangiocarcinoma pathogenesis via RNA splicing regulation of MADD. J. Hepatol. 81, 651–666 (2024).
Li, F. et al. Positive feedback regulation between glycolysis and histone lactylation drives oncogenesis in pancreatic ductal adenocarcinoma. Mol. Cancer 23, 90 (2024).
Qian, Y. et al. MCT4-dependent lactate secretion suppresses antitumor immunity in LKB1-deficient lung adenocarcinoma. Cancer Cell 41, 1363–1380.e7 (2023).
Liu, X. et al. Activation of GPR81 by lactate drives tumour-induced cachexia. Nat. Metab. 6, 708–723 (2024).
Chen, Y. J. et al. Lactate metabolism is associated with mammalian mitochondria. Nat. Chem. Biol. 12, 937–943 (2016).
Tasdogan, A. et al. Metabolic heterogeneity confers differences in melanoma metastatic potential. Nature 577, 115–120 (2020).
Hashimoto, T., Hussien, R. & Brooks, G. A. Colocalization of MCT1, CD147, and LDH in mitochondrial inner membrane of L6 muscle cells: evidence of a mitochondrial lactate oxidation complex. Am. J. Physiol. Endocrinol. Metab. 290, E1237–E1244 (2006).
Wang, N. et al. Structural basis of human monocarboxylate transporter 1 inhibition by anti-cancer drug candidates. Cell 184, 370–383.e13 (2021).
Li, X. et al. Lactate metabolism in human health and disease. Signal Transduct. Target Ther. 7, 305 (2022).
Morrissey, S. M. et al. Tumor-derived exosomes drive immunosuppressive macrophages in a pre-metastatic niche through glycolytic dominant metabolic reprogramming. Cell Metab. 33, 2040–2058.e10 (2021).
Parida, P. K. et al. Metabolic diversity within breast cancer brain-tropic cells determines metastatic fitness. Cell Metab. 34, 90–105.e7 (2022).
Watson, M. J. et al. Metabolic support of tumour-infiltrating regulatory T cells by lactic acid. Nature 591, 645–651 (2021).
Kumagai, S. et al. Lactic acid promotes PD-1 expression in regulatory T cells in highly glycolytic tumor microenvironments. Cancer Cell 40, 201–218.e9 (2022).
Elia, I. et al. Tumor cells dictate anti-tumor immune responses by altering pyruvate utilization and succinate signaling in CD8( + ) T cells. Cell Metab. 34, 1137–1150.e6 (2022).
Feng, Q. et al. Lactate increases stemness of CD8 + T cells to augment anti-tumor immunity. Nat. Commun. 13, 4981 (2022).
Wenes, M. et al. The mitochondrial pyruvate carrier regulates memory T cell differentiation and antitumor function. Cell Metab. 34, 731–746.e9 (2022).
Walenta, S. et al. High lactate levels predict likelihood of metastases, tumor recurrence, and restricted patient survival in human cervical cancers. Cancer Res. 60, 916–921 (2000).
Liu, S. et al. Lactate promotes metastasis of normoxic colorectal cancer stem cells through PGC-1α-mediated oxidative phosphorylation. Cell Death Dis. 13, 651 (2022).
Bonuccelli, G. et al. Ketones and lactate “fuel” tumor growth and metastasis: evidence that epithelial cancer cells use oxidative mitochondrial metabolism. Cell Cycle 9, 3506–3514 (2010).
Liberti, M. V. & Locasale, J. W. The Warburg effect: how does it benefit cancer cells? Trends Biochem. Sci. 41, 211–218 (2016).
Uen, W. et al. Detachment stress mediated bioenergetic switch of malignant melanoma cells into anti-Warburg phenotype. Aging (Albany NY) 14, 5511–5522 (2022).
Chen, A. N. et al. Lactylation, a novel metabolic reprogramming code: current status and prospects. Front. Immunol. 12, 688910 (2021).
Brown, T. P. & Ganapathy, V. Lactate/GPR81 signaling and proton motive force in cancer: role in angiogenesis, immune escape, nutrition, and Warburg phenomenon. Pharm. Ther. 206, 107451 (2020).
Byun, J. K. Tumor lactic acid: a potential target for cancer therapy. Arch. Pharm. Res. 46, 90–110 (2023).
Baek, M. L. et al. Mitochondrial structure and function adaptation in residual triple negative breast cancer cells surviving chemotherapy treatment. Oncogene 42, 1117–1131 (2023).
Faubert, B., Solmonson, A. & DeBerardinis, R. J. Metabolic reprogramming and cancer progression. Science 368, eaaw5473 (2020).
Liu, S. et al. Metabolic reprogramming and therapeutic resistance in primary and metastatic breast cancer. Mol. Cancer 23, 261 (2024).
Gerstberger, S., Jiang, Q. & Ganesh, K. Metastasis. Cell 186, 1564–1579 (2023).
Shibue, T. & Weinberg, R. A. EMT, CSCs, and drug resistance: the mechanistic link and clinical implications. Nat. Rev. Clin. Oncol. 14, 611–629 (2017).
Kocianova, E., Piatrikova, V. & Golias, T. Revisiting the Warburg effect with focus on lactate. Cancers (Basel) 14, 6028 (2022).
Forero-Quintero, L. S. et al. Membrane-anchored carbonic anhydrase IV interacts with monocarboxylate transporters via their chaperones CD147 and GP70. J. Biol. Chem. 294, 593–607 (2019).
Wang, K. et al. Di-methylation of CD147-K234 promotes the progression of NSCLC by enhancing lactate export. Cell Metab. 33, 160–173.e6 (2021).
Chen, H. et al. Hepatic artery injection of (131)I-metuximab combined with transcatheter arterial chemoembolization for unresectable hepatocellular carcinoma: a prospective nonrandomized, multicenter clinical trial. J. Nucl. Med. 63, 556–559 (2022).
Fan, X. Y. et al. Therapeutic anti-CD147 antibody sensitizes cells to chemoradiotherapy via targeting pancreatic cancer stem cells. Am. J. Transl. Res. 11, 3543–3554 (2019).
Zheng, N. S. et al. CD147-specific chimeric antigen receptor T cells effectively inhibit T cell acute lymphoblastic leukemia. Cancer Lett. 542, 215762 (2022).
Watson, M. J. & Delgoffe, G. M. Fighting in a wasteland: deleterious metabolites and antitumor immunity. J. Clin. Invest. 132, e148549 (2022).
Kerslake, R. et al. Elevated circulating lactate levels and widespread expression of its cognate receptor, hydroxycarboxylic acid receptor 1 (HCAR1), in ovarian cancer. J. Clin. Med. 12, 217 (2022).
Fischer, K. et al. Inhibitory effect of tumor cell-derived lactic acid on human T cells. Blood 109, 3812–3819 (2007).
Cerda-Kohler, H. et al. Lactate administration activates the ERK1/2, mTORC1, and AMPK pathways differentially according to skeletal muscle type in mouse. Physiol. Rep. 6, e13800 (2018).
Chen, D. et al. Deletion of Gtpbp3 in zebrafish revealed the hypertrophic cardiomyopathy manifested by aberrant mitochondrial tRNA metabolism. Nucleic Acids Res. 47, 5341–5355 (2019).
Brizel, D. M. et al. Elevated tumor lactate concentrations predict for an increased risk of metastases in head-and-neck cancer. Int. J. Radiat. Oncol. Biol. Phys. 51, 349–353 (2001).
Wu, Y. et al. PRMT5 regulates RNA m6A demethylation for doxorubicin sensitivity in breast cancer. Mol. Ther. 30, 2603–2617 (2022).
Pang, Z. et al. MetaboAnalyst 5.0: narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 49, W388–w396 (2021).
Cui, J. et al. N-glycosylation by N-acetylglucosaminyltransferase V enhances the interaction of CD147/basigin with integrin β1 and promotes HCC metastasis. J. Pathol. 245, 41–52 (2018).
Trefely, S., Ashwell, P. & Snyder, N. W. FluxFix: automatic isotopologue normalization for metabolic tracer analysis. BMC Bioinforma. 17, 485 (2016).
Bu, J. et al. KK-LC-1 as a therapeutic target to eliminate ALDH(+) stem cells in triple negative breast cancer. Nat. Commun. 14, 2602 (2023).
Huang, N. et al. An upstream open reading frame in phosphatase and tensin homolog encodes a circuit breaker of lactate metabolism. Cell Metab. 33, 128–144.e9 (2021).
Jha, P., Wang, X. & Auwerx, J. Analysis of mitochondrial respiratory chain supercomplexes using blue native polyacrylamide gel electrophoresis (BN-PAGE). Curr. Protoc. Mouse Biol. 6, 1–14 (2016).
Buescher, J. M. et al. A roadmap for interpreting (13)C metabolite labeling patterns from cells. Curr. Opin. Biotechnol. 34, 189–201 (2015).
Millard, P. et al. IsoCor: isotope correction for high-resolution MS labeling experiments. Bioinformatics 35, 4484–4487 (2019).
Morris, G. M. et al. AutoDock4 and AutoDockTools4: automated docking with selective receptor flexibility. J. Comput Chem. 30, 2785–2791 (2009).
Wallace, A. C., Laskowski, R. A. & Thornton, J. M. LIGPLOT: a program to generate schematic diagrams of protein-ligand interactions. Protein Eng. 8, 127–134 (1995).
Acknowledgements
PDXs were generous gifts from Prof. Suling Liu of Fudan University. This study was supported by the State Key Laboratory of Holistic Integrative Management of Gastrointestinal Cancers (grant number: CBSKL2022ZZ09, L.L.), the National Natural Science Foundation of China (grant number: 81872349, L.L.; 82401702, C.-G.S.), and the Major Research Plan of the National Science Foundation of China (grant number: 92169211, Z.-N.C.).
Author information
Authors and Affiliations
Contributions
J.-M.Z., X.-M.Y., Z.-N.C., and L.L. conceived the original idea and designed the study. J.-J.Z., R.-F.T., C.-G.S., R.L., D.H., and G.-Q.Z. performed most experiments. X.-Y.F., Z.-C.D., and K.Z. performed tissue histology. T.-J.Z., Y.-T.C., and J.Z. assisted with the animal model. K.W. performed flow cytometry and assisted with microscope. J.-J.Z., R.-F.T., C.-G.S. wrote the paper. All authors reviewed the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Carlos Sebastian, who co-reviewed with Lisa Pöltl, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhang, JJ., Tian, RF., Song, CG. et al. Lactate mitochondrial oxidation drives stemness potential in metastatic breast cancer. Nat Commun 17, 399 (2026). https://doi.org/10.1038/s41467-025-67091-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-67091-y
