
Abstract
Effective environmental policies for the tropics depend on accurate, representative scientific data. However, there is strong evidence from particular disciplines and regions that existing research is patchily distributed. Here, we show that poor representation of sampling and citation in some biomes and across key environmental gradients from all disciplines for the entire tropics may lead to flawed scientific paradigms and inappropriate policy prescriptions. We map sampling locations and citations from 2 738 published studies in natural terrestrial tropical environments across all disciplines to identify gaps in field sampling effort and research attention. Five ecoregions – all in moist broadleaf forests – generate 22% of the total citations but cover only 3% of the tropical land area. By contrast, drier biomes with low tree cover account collectively for 57% of the tropical area but generate only 20% of total citations. Locations that are drier, colder, with greater plant species richness, lower tree cover and facing greater climate change extremes are under-sampled and under-cited. Our results will help to correct these imbalances to improve the scientific basis for environmental policies across the tropics.
Similar content being viewed by others

Introduction
The terrestrial tropics are highly populated1 and encompass a wide range of valuable yet threatened ecosystems2,3,4. Numerous international initiatives have emerged to mitigate these threats. These initiatives are shaped by broad syntheses of regional knowledge across all disciplines4,5,6, drawing on fieldwork by numerous researchers. However, field research effort across the tropics is uneven, with certain areas disproportionately represented while other regions remain relatively overlooked7,8,9,10,11,12,13,14,15. Previous studies have tracked research activity within particular disciplines, regions or time frames7,8,9,10,11,12,13,14,15, consistently revealing strong geographic and thematic biases in research effort, citations, peer review and publication. Moreover, site-specific findings may be extrapolated far beyond their original contexts15,16, exacerbating the risk of inappropriate policy applications. Still missing is a comprehensive, cross-disciplinary overview of the spatial distribution of tropical field research sampling and study citation, and a robust assessment of how well this distribution represents the full spectrum of environmental variation across the tropics.
Here, we identify 4260 articles featuring primary field data within the tropics. Habitat types with a high degree of anthropogenic influence (urban and agricultural) account for 36% and 32% of sampling locations and citations, respectively. The spatial distributions of sampling and citation across the tropics are dominated by this prevalence of research on heavily impacted environments (Supplementary Fig. 1). While heavily human-impacted environments in the tropics are widespread and important for policy, they are subject to distinct drivers than environments where direct human influence is minimal17,18. To focus on the spatial distribution and drivers of sampling and citation across relatively natural environments, we remove studies featuring urban and agricultural habitats for subsequent analyses.
We map 6370 field measurements from 2738 published articles representing 89,468 citations, across all disciplines in different natural habitats in terrestrial tropical biomes and ecoregions, and relate their spatial distribution to a selection of key environmental conditions across the tropics. We compile an initial list of studies with a minimum of 1 citation from a keyword search for “trop*” in the article title on the Web of Science database19. The search is designed to minimize introduction of spatial biases arising from the search process itself, such that any biases in the identified body of literature likely reflect genuine trends in research effort and attention20,21 (Supplementary Fig. 2). Each article is scanned by a trained human reviewer to extract geographic coordinates of field sampling sites, article citation data and habitat sampled (including aquatic freshwater). Citation data are included as a proxy for the scientific influence exerted by specific studies22. In cases where multiple coordinates for sampling locations are identified in a paper, citations per sampling location are calculated as total paper citations divided by the number of locations identified per paper.
Using this geo-referenced database, we first summarize the distribution of sampling and citation among tropical biomes and ecoregions using a widely held definition of the terrestrial tropics based upon vegetation structure (Table 1)23. Then, we compare the frequency distribution of sampling and citation under different environmental conditions with the actual frequency distribution with which the same conditions occur in nature across the tropics. We select the following eight conditions because of their recognized importance either as ecosystem drivers or as ecosystem attributes controlling major processes or services2,3,24,25,26. (i) current mean annual temperature (MAT) and (ii) precipitation (MAP), (iii) projected changes in future mean annual temperature (∆MAT) and (iv) precipitation (∆MAP) by 2100 compared to recent conditions (1970-2000) under the SSP 245 scenario in the coupled model inter-comparison project27, (v) MODIS derived leaf area index (LAI)28 and (vi) soil organic carbon stock in the upper 2 meters (SOC) from the SoilGrids product29, and (vii) vascular plant species richness30 and (viii) mammal and bird species richness31. Finally, we use statistical multivariate modeling21 to highlight tropical regions with combinations of environmental conditions which are not adequately sampled according to our database.
Results
Uneven global representation of field research across tropical biomes
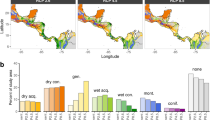
The spatial distribution of sampling locations and citations across the tropics is highly variable (Fig. 1). The moist broadleaf forest biome covers around 29% of the tropics (Supplementary Fig. 3, Table 1) but accounts for 68% and 73% of all sampling locations and citations, respectively (Fig. 1, Table 1). The top five most cited ecoregions (Fig. 2, Supplementary data 1) - all in moist broadleaf forests with major field stations and/or resident population centers - account for 11% and 22% of total locations and citations respectively, but cover only 3% of the tropical biome area. By contrast, drier biomes (dry broadleaf forest, deserts and xeric shrublands, grasslands, savannas and shrublands) account collectively for 57% of the tropical area (Table 1) but feature only 21% and 20% of sampling locations and citations respectively (Fig. 1, Table 1). Deserts and xeric shrublands stand out as poorly sampled and cited both in absolute terms, and after correcting for biome area (Table 1). Mangroves are generally frequently sampled and cited given their limited area (Table 1), although the Guinean mangroves in west Africa is one of the least cited tropical ecoregions (Fig. 2, Supplementary Data 1). Flooded grasslands and savannas, and coniferous forests are cited much less than expected given how often they are sampled (Table 1, citation:sampling ratio).

Density of sampling locations (A) and citations (B) per unit land area across natural terrestrial habitats in the tropics. Spatial resolution is 3° (~ 330 km). Maps were produced from a database of 2738 articles, representing 6370 sampling locations and 89,468 citations. The full extent of tropical biomes is highlighted in dark gray, using widely accepted boundaries23. To account for transition zones between the biomes, we added a buffer of 100 km around the formally defined tropical area. Overall, the study area consisted of 52.9 × 106 km-2 of terrestrial land (ca. 36 % of the global land area). Base map from Natural Earth98.

Values are derived from a database of 2738 articles, representing 6370 sampling locations and 89,468 citations. Values represent probabilities (1 = high, 0 = low) that environmental conditions within a location have been sampled, using statistical multivariate modeling16. A value above 0.5 effectively classifies an environmental condition as one where a sampling location is present. Photographs show the locations of the top five most cited (red outline and arrow) and least cited (black outline and arrow) ecoregions across the terrestrial tropics23. Photo credits in Supplementary Table 1. Data of locations and citation metrics across the full list of ecoregions are presented in Supplementary data 1. Base map from Esri99.
Regional gaps in tropical research
Current sampling efforts capture some tropical habitats and conditions well, while others remain relatively under-sampled (Fig. 2). Specifically, current sampling locations adequately represent environmental conditions from only around 30% of the tropics, corresponding mainly with the moist broadleaf forest biome, particularly in Asia (Fig. 2). Areas with environmental conditions that are poorly represented by the present distribution of sampling correspond mainly with biomes in drier regions with low tree cover, particularly in Africa (Fig. 2).
Representativeness of sampling and citation across the tropical environmental space
The observed distribution of research sampling locations and citations with varying MAT, MAP, ∆MAT, ∆MAP, LAI, SOC, vascular plant, mammal and bird species richness are different from the expected distribution based upon the tropical land area characterized by these same conditions (Fig. 3, Supplementary Fig. 3). Specifically, relatively cold (<20 oC MAT) or dry (<1000 mm MAP) locations with low LAI ( < 3 m2 m-2) and predicted to face more climate extremes (greater future warming, cooling or increased precipitation) are less sampled and cited than expected given their spatial extent (Fig. 3), which corresponds to the following tropical biomes: dry broadleaf forest, coniferous forest, grasslands, savannas and shrublands (Supplementary Fig. 3). These areas tend to occur at relatively high and low latitudes, and at higher elevations, within the tropics (Fig. 2). Conversely, areas with high LAI and high diversity of mammals and birds, corresponding roughly with the moist broadleaf forest biome (Supplementary Fig. 3), are sampled and cited more often than expected from the frequency of their occurrence (Fig. 3). It is important to note, however, that the datasets used to quantify actual biodiversity distributions30,31 are themselves likely spatially biased32,33, for many of the same reasons that drive sampling and citation biases7,8,9,10,11,12,13,14,15,16. Therefore, the extent to which actual biodiversity distributions are accurately represented by existing research should be interpreted with caution, though the present analysis likely overestimates representation (Fig. 3).

Frequency distribution of actual occurrence (A, D, G, J), sampling locations (B, E, H, K) and citations (C, F, I, L) for different combinations of environmental conditions across natural terrestrial habitats in the tropics. Values are derived from a database of 2738 articles, representing 6370 sampling locations and 89,468 citations. The tropics are defined using widely accepted boundaries23. Gray pixels denote the full range of ambient conditions across the entire tropics, from a random sample (n = 100 000) of the total pixels within the study area. To be representative, sampling locations (B, E, H, K) and citations (C, F, I, L) should cover the full range of environmental conditions shown in gray and display a frequency distribution similar to the actual occurrence of environmental conditions (A, D, G, J) observed across the entire tropics.
Discussion
Policy risks from unrepresentative sampling and citation in tropical research
Scientific research depends on finite resources, which necessitates difficult decisions about where to focus field sampling efforts. We document major spatial biases in research foci across the terrestrial tropics, which means that valuable ecoregions and widespread environmental conditions remain largely overlooked. For example, the under-sampled and poorly recognized drier biomes show high floristic diversity34 and play a central role in regulating inter-annual variability in global atmospheric carbon dioxide levels35. Yet, these drier biomes are home to around one-third of the global human population36, their habitats are generally more threatened37 and they receive less formal protection than other biomes38. By contrast, certain geographic areas, biomes, and ecoregions are disproportionately favored in terms of research effort and attention. These relatively well-sampled and cited regions tend to occur in humid forest biomes, particularly in Asia. The vast diversity of tropical environments exacerbates the risks of extrapolating findings from a narrow set of well-researched contexts to broader, ecologically distinct regions15,16.
One possible example of such extrapolation is the widespread advocacy – both within and beyond the scientific community – for afforestation in ecosystems with naturally low tree cover as a climate change mitigation strategy39,40,41. The prevalence and persistence of this narrative16,42,43 may stem, at least in part, from the strong research emphasis we observe in moist broadleaf forest biomes, and the relative scarcity of research in dry forests and open tropical ecosystems such as deserts, grasslands, and shrublands37,38. The scientific inferences and policy prescriptions derived from the limited number of intensively sampled locations often stretches far beyond the wider regions which possess clear climatic or ecosystem analogues to the original locations15,16, hampering the development of effective environmental management actions tailored to suit local conditions.
Drivers of research imbalances and pathways to more representative insights
As science enters an era of “big data”, the urgency to make sense of massive data streams has increased dramatically. One critical challenge is that many large-scale data collection initiatives do not collect representative samples of their variable of interest44, which means both that the effective sample sizes are much lower and that the mean variable estimates from these samples are inaccurate45. The spatial biases we reveal likely emerge from a complex mix of factors: locations of research stations46, article peer review outcomes and citation rates8, evolution of population centers and transport infrastructure1, as well as imbalances among regions in resources available for research47. Further, there may be biome-specific differences in the likelihood that research will be referred to as tropical. Although technically a potential methodological artifact in the current analysis, if true, it would nevertheless contribute to the continued marginalisation of certain tropical biomes from policy discussions. As it is, we believe that the trends identified mostly reflect genuine trends in tropical research effort and attention. First, because they are confirmed by multiple independent sources7,8,9,10,11,12,13,14,15,37,38. Second, because the biome most closely linked to the tropics – mangroves48,49 – where there should be the weakest incentive to specify the tropical origin of the research, is more sampled and cited relative to its extent, not less as would be expected if the search term in our literature review introduced sampling artifacts.
We emphasize that the spatial biases identified are an emergent property resulting from synthesizing many individual research studies, then drawing broad conclusions from them (“external validity” in reviews and meta-analyses50), even though individual studies may not make inferences beyond their immediate study site. As such, our results make no claim about the accuracy and quality of individual articles (“internal validity”50). Nor do our results suggest that intensively studied research sites and field stations are inherently problematic or not deserving of investment. On the contrary, these infrastructures enable in-depth investigations that would be difficult to execute elsewhere and often yield a high return on investment51. Instead, we advocate for complementing the detailed, long-term perspectives provided by intensive research sites with broader pan-tropical perspectives from spatially extensive measurement networks when formulating integrative outputs intended to inform policy. Such networks have already been established to address these challenges, mainly focused on carbon cycling52,53 and species occurrence54,55. While these networks may also be affected by problems associated with unrepresentative spatial sampling56,57, they remain essential tools for broadening scientific perspectives. More networks addressing other biomes38, ecosystem components and processes are developing, and will contribute to a more balanced picture of pan-tropical processes as long as the underlying spatial distribution of sampled sites is explicitly considered when deriving broad principles and metrics of tropical ecosystem functioning.
As larger-scale – but often unrepresentative – data collection initiatives flourish, the need to develop strategies to derive accurate, balanced inferences from these datasets is growing ever more urgent. A range of qualitative and quantitative approaches could be used to account for the spatial distribution of sampling44. Rigorous assessments of the contributions of bias on descriptive inference - so called “risk-of-bias” assessments – are standard in medical research proposals and papers58. Expanding the use of risk-of-bias assessments to other fields could improve scientific transparency and rigor, helping both authors and readers better understand the limitations and generalizability of research findings. Risk-of-bias can be reduced with auxiliary variables which are associated with both the likelihood of a unit being sampled and with the underlying values of the variable of interest, to adjust the overall population-level estimate so that it lies closer to the true value44,59,60. Moreover, such variables can also be used to guide future sampling efforts – to target locations which have been sampled less frequently than would be expected by chance61,62,63.
The spatial distribution of sampling documented in this study could serve as an auxiliary variable to correct current estimates of ecosystem properties and processes and guide future, more balanced sampling, improving biome or pan-tropical estimates of environmental variables. However, even after statistical and sampling corrections, some residual biases are inevitable. These biases and uncertainties should be clearly communicated to readers and data users to aid interpretation64,65. Specifically, the temporal and spatial scope of inference supported by the data should be clearly reported. Where inferences extend beyond the sampled populations or regions, such extrapolations should be explicitly acknowledged and critically assessed. These practices may not currently be incentivized within academia66 but will become increasingly critical to maintain a clear view of knowns and unknowns in a rapidly changing world inundated with data.
Toward broader and more inclusive tropical sciences
Our results highlight biomes and environmental conditions that dominate tropical research, and identify priorities for future sampling to improve assessments of the overall current and potential future state of the tropics as a whole. While global disparities and inequities in science and research have received considerable attention7,8,9,10,11,12,13,14,15,16,47,67,68,69, our study highlights the extent and importance of regional disparities within the tropics70,71,72, particularly between South America and Asia versus Africa, and tropical lowland forests versus other habitats. The underlying causes of these regional disparities likely overlap substantially with those driving global patterns: including unequal access to research resources and infrastructure among tropical countries and regions47,73, variation in social and political stability74, administrative barriers to knowledge transfer across regions and countries75, the preferential channeling of international funding and collaborations through a small subset of tropical institutions and countries46, a bias in research toward forested landscapes relative to other tropical habitat types70, unethical collaborative practices which disproportionately benefit partners from wealthier regions and/or countries often outside the tropics76 and systemic biases in the recognition of scientific knowledge production77,78. Many of these issues lack straightforward solutions, requiring a paradigm shift in global scientific collaborative practices76,79. However, reducing administrative and financial barriers to scientific engagement across different tropical regions and globally – such as the costs of journal subscriptions and the difficulty of securing visas for research visits and study – would represent a major concrete advance68,75.
Based upon our study, we offer three broad, related but distinct, conclusions and suggestions for future action. First, large portions of the tropics representing valuable ecoregions are relatively well-sampled but poorly cited, and therefore may have had limited influence over scientific narratives or environmental policy. Similar bodies of scientific knowledge originating from different locations receive very different levels of recognition77. This issue may be exacerbated by the under-representation of non-English language literature80,81, which we recommend integrating more fully into future reviews. Fast-evolving translation tools make this a realistic vision82. Efforts to increase the diversity of scientific groups – such as journal editors, reviewers, society board members, conference organizers – could help to increase representation from under-recognized tropical regions and countries and reduce systemic bias69,79. Some journals have begun experimenting with tools designed to reduce discriminatory practices in academic publishing83. If adopted more broadly as standard practice, such tools could contribute to the creation of a more equitable scientific landscape.
Second, some tropical areas remain significantly under-sampled despite their broad extent and ecological value. These areas should be prioritized in future research efforts, as an effective means to increase the amount of novel environmental knowledge per unit research investment. Greater recognition of the scientific value of under-sampled regions by governments, research institutions, funding bodies and journal editors or reviewers would help incentivize researchers to undertake the additional costs often inherent to sampling these areas16,42,70. International support for local research infrastructures and field stations could be restructured to begin to counteract the accumulated effects of historical preferences for highly accessible locations near lowland tropical forests.
Finally, we highlight disparities in research attention across a few key axes of environmental variation across the tropics, but there remain many other globally or locally critical drivers of ecosystem processes (e.g., anthropogenic influence, geology, soil type, or plant/animal phylogenetic relatedness), which are likely also not well represented by the current distribution of field research. Further work might highlight new priority areas for future sampling or deserving of greater attention, depending on the process or driver in question. Addressing these data gaps is essential for producing truly integrative, globally relevant ecological insights.
Methods
Inclusion & ethics
The authorship team comprises a diverse range of nationalities and career stages with a reasonably balanced gender composition. There is, however, a distinct overrepresentation of North European and American institutions, although several members of the team are nationals of tropical countries but are now employed outside of the tropics. In large part, this reflects the fact that much of the group was initially established to complete a conceptually similar article focused on Arctic systems20. For the present analysis, considerable effort was made to widen the authorship group, enlisting assistance and inputs from researchers working in tropical countries, with limited success. Therefore, in the present article, we have taken particular care to evaluate and thoroughly describe the diverse perspectives about the patterns and drivers of regional and global variation in knowledge production.
Literature review
On 3 November 2021, we searched the Web of Science database for articles with the term “trop*” in their title. Wider keyword searches of the abstract or main text were not performed since they yielded an intractably large number of articles. The approach was not designed to yield a complete list of tropical research, but to provide as close to an unbiased subset of tropical research as possible. As such, more specific search terms were avoided since they could introduce biases if particular names or terms were more likely to be used in particular locations or by particular disciplines. Non-English language articles were not screened out but represent a minority of the Web of Science database84. Uncited papers were not included because it was assumed that they have not yet exerted much influence over scientific paradigms or policy strategy22. We include all studies irrespective of discipline and all time periods, including social sciences and laboratory studies, as long as the geographic origin of the samples was reported.
The resulting initial list of 11,804 cited papers was then screened to assess their relevance to our objectives (see key steps in the screening process in a PRISMA flow diagram format (Supplementary Fig. 2). Of these papers, 11,713 (99% of initially screened papers) were successfully accessed via university institutional access to the publisher in question or by writing to the corresponding author for a personal copy. After full review, papers were excluded (6625, 56.0% of initially screened papers) if: (1) they featured only measurements in marine environments; tidal estuaries were counted as terrestrial and labeled as river habitats; (2) they did not include primary field measurements because they were broad reviews, modeling analyses, the data presented had already been published elsewhere, or the measurements were from laboratory measurements using samples without a clear provenance; (3) the primary field measurements featured were located outside of the tropics and buffer regions as defined in our study23. Studies that were not field-based (for example, remote sensing, geographical information science, and modeling analyses) were, in some cases, included where they included ‘groundtruthing’ field measurements and/or the spatial extent of the study was relatively limited.
After removing papers that did not fulfill these criteria, 5088 (43% of initially screened papers) papers remained, which were subjected to further analysis. Content analysis was used to: (1) extract geographical coordinates of the field measurements. In cases where coordinates were not explicitly provided, we used place or landform names mentioned in the text to determine the approximate coordinates of the field site(s) on Google Maps; (2) classify the habitats sampled within the paper. The habitats featured were forest, grassland, wetland, desert, rocky area, agricultural, urban, lake, and river. Content analysis inevitably included a degree of subjective judgment on the part of the reviewer. All reviewers were trained at least to a university undergraduate level in environmental sciences and received identical review instructions. Individual papers frequently featured multiple habitats and/or single habitats which represented aspects of multiple habitat categories, in which case a maximum of 3 habitats could be assigned to the same sampling location. The information from the content analysis was then paired with basic paper information derived from Web of Science (authors, journal, title, volume and page numbers, science categories and research areas, citations as of 3 November 2021) to form the central dataset for subsequent analyses.
Mapping study sampling locations and citations
To further define our study domain for spatial analysis, we used the biome boundaries that were classified as tropical in the ecoregions database (i.e. BIOME_NAME field included a word “tropical”)23. To acknowledge that there might be transition zones between the biomes, we added a buffer of 100 km around the tropical area. Overall, our domain consisted of 52.9 × 106 km-2 of terrestrial land (ca. 36 % of the global land area). After removing articles that were outside this tropical domain, the number of articles, sampling locations, and citations decreased to 4260, 9987, and 131,030, respectively. Finally, to focus on terrestrial environments with lower intensity of direct human influence, we excluded sampling locations in urban and agricultural areas based on the habitat description in the literature database, which resulted in a dataset of 2738 articles, 6370 sampling locations and 89,468 citations for the final analyses.
Extraction of environmental conditions variables from study site locations
All the data processing and analyses were conducted in R program version 4.4.185. We used a range of climatic, vegetation, soil and biodiversity data to characterize the tropical region as a whole and to extract data to study site locations. From the biome dataset23, we utilized the variables ecoregion (ECO_NAME) and biome (BIOME_NAME) to broadly classify articles to key ecological domains. We used the 1-km WorldClim v2 mean annual average air temperature (oC) and annual cumulative precipitation data (mm) over 1970-2000 and 2081-2100 based on the SSP 2.45 scenario, produced from an ensemble of 12 downscaled CMIP6 layers27. Climate anomaly layers were calculated based on the difference between 2081-2100 and 1970-2000 layers. We used the MODIS MOD15A2H dataset, which provides 500-meter resolution data on Leaf Area Index (Lai_500m)28. We applied quality filtering to exclude poor-quality pixels (included FparLai_QC bit 0 value 0 data, i.e., good quality) and areas affected by clouds (included FparLai_QC bit 3-4 value 0 data, i.e., no clouds). We then calculated annual means over 2002-2023 and filled gaps in the average MODIS LAI map by applying a moving window analysis (window size: 19) with the focal command in the R package terra86. Soil organic carbon stock data for the uppermost 2 meters were extracted from the SoilGrids product29. We used a dataset of predicted vascular plant species richness (i.e.: alpha diversity) for a plot size of 1000 m2 including forest and non-forest species (ca. 5 km pixel resolution)30. This plot size was chosen as it is commonly used when sampling forests. We further extracted predicted bird and mammal species richness datasets31 and summed them as one animal diversity measure. All the geospatial layers were re-projected to WGS 1984 at 1 km resolution and masked by the climate datasets using the R package terra86.
Spatial analyses
We calculated the total number of articles, sampling locations, and citations across biomes and ecoregions. Then, we examined the distribution of sampling locations and citations across the full range of tropical conditions, to compare with the actual prevalence of the same conditions in reality. To describe the conditions across the entire tropics, we took a random sample (n = 100 000) of the total pixels within our study domain.
We used statistical multivariate modeling to highlight areas lacking sampling locations when considering overall environmental variability21,87. This approach is conceptually grounded in species distribution models (SDMs)88. SDMs define a geographic space based on environmental variables and identify areas where environmental conditions are suitable for a given species. We adapted this framework to evaluate representativeness of sampling locations, aiming to delineate the spatial distribution of environmental conditions across a geographic envelope that reflects the range of environments captured by the current sampling locations21,87.
We used a binomial/categorical response variable for the presence-absence data (1 = sampling location exists, 0 = sampling location is missing), and climate (MAT and MAP), soil (SOC), vegetation (LAI), and biodiversity (plant and animal species richness) as explanatory variables. Since our database contains information about sampling locations only, we needed to artificially create locations with the absence of sampling. To do this, we followed an established methodology89, creating a random sample of terrestrial absence locations with the same number of observations as our presence locations (n = 6447) with the R package sp90. Then, we obtained spatial data in these randomly sampled locations based on coordinate colocation. These were then combined with the literature database, which resulted in a data frame of 12,894 locations. The predictors in the final data set did not suffer from high multi-collinearities, as the correlations between the predictor variables were <0.70.
We used common statistical and machine learning models – generalized additive models91, random forest models92 and generalized boosted regression trees93 – to predict both the presence-absence of sampling locations and the probabilities for the presence. To reduce uncertainties associated with individual models, we calculated the median probability across the three models, which was used to describe the representativeness of sampling locations for each raster pixel across the whole tropics. In the final map, high probabilities indicate a good coverage of current sampling locations in similar conditions (1 = high probability that there is a sampling location in such conditions), and low probabilities suggest lack of locations (0 = no probability for a sampling location). From these probabilities, we also calculated the total area capturing the environmental conditions where sampling sites are covered (>0.5).
The performance of the three models and their ensemble was assessed using cross-validation with 99 permutations from which we calculated the area under the curve (AUC) test statistic94 with the R package ROCR95. In the cross-validation procedure, a random sample of 70% of the data was used to test the model fit, and the remaining 30% were used to assess predictive performance. Test statistics were calculated after each permutation to evaluate the ensemble model. AUC scores varied from 0.76 to 0.9, with the mean AUC being 0.83. An AUC value of 1 represents perfect accuracy and 0.5 indicates that the model is no better than random. All the visualizations from the spatial analyses were created with the R package ggplot296 and maps with ESRI ArcGIS Pro version 3.0.3.
Use of AI
Large language models were used to copy-edit existing text, to check for errors in grammar and syntax, and to suggest alternative sentence formulations.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data generated in this study have been deposited in a Zenodo repository file, with DOI 10.5281/zenodo.15423742 (https://zenodo.org/records/15423743)97. The data can be downloaded from this link by any user, there are no access restrictions. Additional data are presented together with the article in the file Supplementary data 1, which presents sampling and citation data for the full list of ecoregions included within the study area.
Code availability
All code generated in this study have been deposited in a Zenodo repository file, with DOI 10.5281/zenodo.15423742 (https://zenodo.org/records/15423743)97. The code can be downloaded from this link by any user, there are no access restrictions.
References
Liu, L., Cao, X. & Li, S. A. 31-year (1990–2020) global gridded population dataset generated by cluster analysis and statistical learning. Sci. Data 11, 124 (2024).
Jung, M. et al. Areas of global importance for conserving terrestrial biodiversity, carbon and water. Nat. Ecol. Evolution 5, 1499–1509 (2021).
Chaplin-Kramer, R. et al. Mapping the planet’s critical natural assets. Nat. Ecol. Evolution 7, 51–61 (2023).
Olsson L. et al. Land degradation in climate change and land: An IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems, Shukla, P. R., Skea, J., Calvo Buendia, E., Masson-Delmotte, V., Pörtner, H.-O., Roberts, D. C., Zhai, P., Slade, R., Connors, S., van Diemen, R., Ferrat, M., Haughey, E., Luz, S., Neogi, S., Pathak, M., Petzold, J., Portugal Pereira, J., Vyas, P., Huntley, E., Kissick, K., Belkacemi, M., Malley, J., Eds (Cambridge university press, cambridge, UK, 2019),
J. P. Ometto, et al. Cross-chapter paper 7: Tropical Forests. In climate change 2022: Impacts, adaptation and vulnerability. contribution of working group ii to the sixth assessment report of the intergovernmental panel on climate change, Pörtner, H.-O., Roberts, D. C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, Craig, M., Langsdorf, S., Löschke, S., Möller, V., Okem, A., Rama, B., Eds. (Cambridge University Press, Cambridge, UK, 2022).
FAO and UNEP. The state of the world’s forests 2020. Forests, biodiversity and people. (FAO and UNEP, Rome, Italy, 2022).
Martin, L. J., Blossey, B. & Ellis, E. Mapping where ecologists work: biases in the global distribution of terrestrial ecological observations. Front. Ecol. Environ. 10, 195–201 (2012).
Meijaard, E., Cardillo, M., Meijaard, E. M. & Possingham, H. P. Geographic bias in citation rates of conservation research. Conserv. Biol. 29, 920–925 (2015).
Stocks, G., Seales, L., Paniagua, F., Maehr, E. & Bruna, E. M. The geographical and institutional distribution of ecological research in the tropics. Biotropica 40, 397–404 (2008).
Reddy, S. & Dávalos, L. M. Geographical sampling bias and its implications for conservation priorities in Africa. J. Biogeogr. 30, 1719–1727 (2003).
Kindt, R. TreeGOER: A database with globally observed environmental ranges for 48,129 tree species. Glob. Change Biol. 29, 6303–6318 (2023).
Pyšek, P. et al. Geographical and taxonomic biases in invasion ecology. Trends Ecol. Evolution 23, 237–244 (2008).
White, C. R. et al. Geographical bias in physiological data limits predictions of global change impacts. Funct. Ecol. 35, 1572–1578 (2021).
Muenchow, J., Dieker, P., Kluge, J., Kessler, M. & von Wehrden, H. A review of ecological gradient research in the Tropics: identifying research gaps, future directions, and conservation priorities. Biodivers. Conserv. 27, 273–285 (2018).
Gardner, T. A. et al. Prospects for tropical forest biodiversity in a human-modified world. Ecol. Lett. 12, 561–582 (2009).
Silveira, F. A. O. et al. Biome Awareness Disparity is BAD for tropical ecosystem conservation and restoration. J. Appl. Ecol. 59, 1967–1975 (2022).
Bonebrake, T. C. et al. Tropical cities as windows into the ecosystems of our present and future. Biotropica 57, e13369 (2025).
Defries, R. S., Bounoua, L. & Collatz, G. J. Human modification of the landscape and surface climate in the next fifty years. Glob. Change Biol. 8, 438–458 (2002).
Metcalfe, D. B. et al. Patchy field sampling biases understanding of climate change impacts across the Arctic. Nat. Ecol. Evolution 2, 1443–1448 (2018).
Virkkala, A.-M., Abdi, A. M., Luoto, M. & Metcalfe, D. B. Identifying multidisciplinary research gaps across Arctic terrestrial gradients. Environ. Res. Lett. 14, 124061 (2019).
Petersen, A. M. et al. Reputation and impact in academic careers. Proc. Natl. Acad. Sci. USA 111, 15316–15321 (2014).
Dinerstein, E. An ecoregion-based approach to protecting half the terrestrial realm. BioScience 67, 534–545 (2017).
Sheldon, K. S. Climate change in the tropics: ecological and evolutionary responses at low latitudes. Annu. Rev. Ecol., Evolution, Syst. 50, 303–333 (2019).
Fang, H., Baret, F., Plummer, S. & Schaepman-Strub, G. An overview of global leaf area index (LAI): Methods, products, validation, and applications. Rev. Geophysics 57, 739–799 (2019).
Scharlemann, J. P., Tanner, E. V., Hiederer, R. & Kapos, V. Global soil carbon: understanding and managing the largest terrestrial carbon pool. Carbon Manag. 5, 81–91 (2014).
Fick, S. E. & Hijmans, R. J. WorldClim 2: new 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315 (2017).
Myneni, R., Knyazikhin, Y., Park, T. MODIS/Terra Leaf Area Index/FPAR 8-Day L4 Global 500m SIN Grid V061. NASA EOSDIS Land Processes Distributed Active Archive Center (2021).
Hengl, T. et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 12, e0169748 (2017).
Sabatini, F. M. et al. Global patterns of vascular plant alpha diversity. Nat. Commun. 13, 4683 (2002).
Jenkins, C. N., Pimm, S. L. & Joppa, L. N. Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. USA 110, E2602–E2610 (2013).
Hughes, A. C. et al. Sampling biases shape our view of the natural world. Ecography 44, 1259–1269 (2021).
Bowler, D. E. et al. Treating gaps and biases in biodiversity data as a missing data problem. Biol. Rev. 100, 50–67 (2025).
DRYFLOR. Plant diversity patterns in neotropical dry forests and their conservation implications. Science 353, 1383–1387 (2016).
Poulter, B. et al. Contributions of semi-arid ecosystems to interannual variability of the global carbon cycle. Nature 509, 600–603 (2014).
Global Land Project. Science plan and implementation strategy. International Geosphere Biosphere Program, report no. 53 / International Human Dimensions Programme Report No. 19, (IGBP Secretariat, Stockholm, Sweden, 2005).
Janzen, D. H. Tropical dry forests: The most endangered major tropical ecosystem. In Biodiversity, E. O. Wilson, Ed. (National Academy Press, Washington DC, USA, 1988).
Moonlight, P. W. et al. Expanding tropical forest monitoring into dry forests: the DRYFLOR protocol for permanent plots. Plants, people, planet 3, 295–300 (2021).
Hanson, C., Buckingham, K., DeWitt, S. & Laestadius, L. Therestoration diagnostic (world resources institute, washington DC, USA, 2015).
Romijn, E. et al. Land restoration in latin america and the caribbean: An overview of recent, ongoing and planned restoration initiatives and their potential for climate change mitigation. Forests 10, 510 (2019).
Bastin, J.-F. et al. The global tree restoration potential. Science 365, 76–79 (2019).
Bond, W. J. Ancient grasslands at risk. Science 351, 120–122 (2016).
Parr, C. L., Te Beest, M. & Stevens, N. Conflation of reforestation with restoration is widespread. Science 383, 698–701 (2024).
Boyd, R. J., Powney, G. D. & Pescott, O. L. We need to talk about nonprobability samples. Trends Ecol. Evolution 38, 521–531 (2023).
Meng, X. L. Statistical paradises and paradoxes in big data (I): Law of large populations, big data paradox, and the 2016 US presidential election. Ann. Appl. Stat. 12, 685–726 (2018).
Raby, M. The colonial origins of tropical field stations. Scientific American 105, https://doi.org/10.1511/2017.105.4.216 (2017).
Livingston, G. et al. Perspectives on the global disparity in ecological science. BioScience 66, 147–155 (2016).
Worthington, T. A. et al. A global biophysical typology of mangroves and its relevance for ecosystem structure and deforestation. Sci. Rep. 10, 14652 (2020).
Morrisey, D. J. et al. The ecology and management of temperate mangroves. In Oceanography and Marine Biology, R. N. Gibson, R. J. A. Atkinson, J. D. M. Gordon, Eds. (Taylor & Francis group, London, UK, 2010).
Avellar, S. A. et al. External Validity: The next step for systematic reviews?. Evaluation Rev. 41, 283–325 (2016).
Eppley, T. M. et al. Tropical field stations yield high conservation return on investment. Conserv. Lett. 17, e13007 (2024).
ForestPlots.net et al. Taking the pulse of earth’s tropical forests using networks of highly distributed plots. Biol. Conserv. 260, 108849 (2021).
Malhi, Y. et al. The global ecosystems monitoring network: monitoring ecosystem productivity and carbon cycling across the tropics. Biol. Conserv. 253, 108889 (2021).
Davies, S. J. et al. ForestGEO: Understanding forest diversity and dynamics through a global observatory network. Biol. Conserv. 253, 108907 (2021).
GBIF Secretariat. GBIF Science Review. No. 11. (GBIF Secretariat, Copenhangen, Denmark, 2024)
Marvin, D. C. et al. Amazonian landscapes and the bias in field studies of forest structure and biomass. Proc. Natl. Acad. Sci. Usa. 111, E5224–E5232 (2014).
Phillips, O. L. et al. Changes in growth of tropical forests: evaluating potential biases. Ecol. Appl. 12, 576–587 (2002).
Sterne, J. A. C. et al. RoB 2: a revised tool for assessing risk of bias in randomised trials. BMJ 366, 1–8 (2019).
Elliott, M. R. & Valliant, R. Inference for nonprobability samples. Stat. Sci. 32, 249–264 (2017).
Boyd, R. J., Stewart, G. B. & Pescott, O. L. Descriptive inference using large, unrepresentative nonprobability samples: An introduction for ecologists. Ecology 105, e4214 (2023).
Pescott, O. L., Powney, G. D. & Boyd, R. J. Adaptive sampling for ecological monitoring using biased data: a stratum-based approach. Oikos e11115 (2025).
Schouten, B. & Shlomo, N. Selecting adaptive survey design strata with partial R-indicators. Int. Stat. Rev. 85, 143–163 (2017).
Henrys, P. A., Mondain-Monval, T. O. & Jarvis, S. G. Adaptive sampling in ecology: Key challenges and future opportunities. Methods Ecol. Evolution 15, 1483–1496 (2024).
Sutherland, W., Spiegelhalter, D. & Burgman, M. Policy: Twenty tips for interpreting scientific claims. Nature 503, 335–337 (2013).
Blastand, M. et al. Five rules for evidence communication. Nature 587, 81–98 (2020).
Van Calster, B., Wynants, L., Riley, R. D., van Smeden, M. & Collins, G. S. Methodology over metrics: current scientific standards are a disservice to patients and society. J. Clin. Epidemiol. 138, 219–226 (2021).
Graves, J. L., Kearney, M., Barabino, G. & Malcom, S. Inequality in science and the case for a new agenda. Proc. Natl. Acad. Sci. USA 119, e2117831119 (2022).
Nakamura, G., Soares, B. E., Pillar, V. D., Felizola Diniz-Filho, J. A. & Duarte, L. Three pathways to better recognize the expertise of global south researchers. npj Biodivers. 2, 17 (2023).
Hughes, A. C. et al. Who is publishing in ecology and evolution? the underrepresentation of women and the Global South. Front. Environ. Sci. https://doi.org/10.3389/fenvs.2023.1211211 (2023).
Pillar, V. D. & Overbeck, G. E. Nature conservation policies are biased toward forests and neglect grassy ecosystems worldwide. Science 388, eadx7441 (2025).
Jarić, I., Diagne, C. & Chowdhury, S. Moving beyond continents for global and inclusive science. Front. Ecol. Environ. 23, e2851 (2025).
Tanalgo, K. C. Open and FAIR data sharing are building blocks to bolster biodiversity conservation in Southeast Asia. Biol. Conserv. 307, 111192 (2025).
UNESCO. UNESCO Science Report: the race against time for smarter development. https://unesdoc.unesco.org/ark:/48223/pf0000377433 (2021).
Hilario-Husain, B. A. et al. Caught in the crossfire: biodiversity conservation paradox of sociopolitical conflict. npj Biodivers. 3, 10 (2024).
Bamba, G. et al. Strategies for more equitable engagement for african researchers. Lancet Glob. Health 13, e14–e15 (2025).
Armenteras, D. Guidelines for healthy global scientific collaborations. Nat. Ecol. Evolution 5, 1193–1194 (2021).
Gomez, C. J., Herman, A. C. & Parigi, P. Leading countries in global science increasingly receive more citations than other countries doing similar research. Nat. Hum. Behav. 6, 919–929 (2022).
Smith, O. M. et al. Peer review perpetuates barriers for historically excluded groups. Nat. Ecol. Evolution 7, 512–523 (2023).
Nuñez, M. A. et al. Assessing the uneven global distribution of readership, submissions and publications in applied ecology: Obvious problems without obvious solutions. J. Appl. Ecol. 56, 4–9 (2019).
Serrano, F. C. et al. Knowledge from non-English-language studies broadens contributions to conservation policy and helps to tackle bias in biodiversity data. J. Appl. Ecol. 62, 2148–2162 (2025).
Amano, T. et al. The manifold costs of being a non-native English speaker in science. PLoS Biol. 21, e3002184 (2023).
Steigerwald, E. et al. Overcoming language barriers in academia: machine translation tools and a vision for a multilingual future. BioScience 72, 988–998 (2022).
Pettorelli, N. et al. How international journals can support ecology from the Global South. J. Appl. Ecol. 58, 4–8 (2021).
Vera-Baceta, M. A., Thelwall, M. & Kousha, K. Web of Science and Scopus language coverage. Scientometrics 121, 1803–1813 (2019).
R Core Team. R: A language and environment for statistical computing. Rfoundation for statistical computing (vienna, austria, 2021).
Villarreal, S. & Vargas, R. Representativeness of FLUXNET sites across latin america. J. Geophys. Res.: biogeosciences 126, e2020JG006090 (2021).
Guisan, A. & Thuiller, W. Predicting species distribution: offering more than simple habitat models. Ecol. Lett. 8, 993–1009 (2005).
Barbet-Massin, M., Jiguet, F., Albert, C. H. & Thuiller, W. Selecting pseudo-absences for species distribution models: how, where and how many?. Methods Ecol. evolution 3, 327–338 (2012).
Hastie, T. & Tibshirani, R. Generalized additive models: some applications. J. Am. Stat. Assoc. 82, 371–386 (1987).
Friedman, J. H. Greedy function machine: A gradient boosting machine. Statistics 29, 1189–1232 (2001).
Breiman, L. Random Forests. Mach. Learn. 45, 5–32 (2001).
Hanley, J. A. & McNeil, B. J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 14, 29–36 (1982).
Metcalfe, D. B. & Virkkala, A.-M. Data and code for “Gaps in tropical science arising from biased spatial patterns of sampling and citation”. Zenodo https://doi.org/10.5281/zenodo.15423742 (2025).
Esri. World topographic map, https://www.arcgis.com/home/item.html?id=30e5fe3149c34df1ba922e6f5bbf808f- (2024).
Acknowledgements
We thank Maja Sundqvist, David Wardle and Gesche Blume-Werry for insightful comments on the idea and early versions of the manuscript and data. We acknowledge that certain data included in the manuscript are derived from Clarivate™ (Web of Science™). © Clarivate 2021. All rights reserved. We acknowledge the World Climate Research Programme, which, through its Working Group on Coupled Modeling, coordinated and promoted CMIP6. We thank the climate modeling groups for producing and making available their model output, the Earth System Grid Federation (ESGF) for archiving the data and providing access, and the multiple funding agencies who support CMIP6 and ESGF. Funders: Arctic Six Chairs programme (DBM); European research council consolidator grant ECOHERB 682707 (DBM); Swedish research council for sustainable development 2023-00361 (DBM), 2023-00307 (ABT), 2016-20005 (AL); Swedish Research Council 2021-05265 (GW), 2019-01151 (NC), 2022-04565 (PA); Strategic Research Area “Biodiversity and Ecosystem Services in a Changing Climate” (GW, NC); Strategic Research Area “Modeling the Regional and Global Earth system” (NC); US National Science Foundation 1749252 (HC-Q); Villum Young Investigator grant VIL53048 (JT); Smathers Endowment for Tropical Biology (KJF).
Funding
Open access funding provided by Umea University.
Author information
Authors and Affiliations
Contributions
Conceptualization: D.B.M., A.-M.V.; Methodology: D.B.M., A.-M.V.; Investigation: D.B.M., E.A., H.A., E.P.A., A.E.B., D.C.B., H.B., H.C.-Q., N.C., T.C., C.A.L.D., M.E.D., K.J.F., T.C.W., B.C.H., T.D.G.H., M.J., P.K., A.L., D.L., S.L., G.M., M.M., O.J.L.M., N.S., J.S.P., A.B.-T., J.T., O.K.V., M.W., G.W., W.Z., Y.Y., A.-M.V.; Visualization: D.B.M., A.-M.V.; Project administration: DBM; Writing – original draft: DBM; Writing – review & editing: D.B.M., E.A., H.A., E.P.A., A.E.B., D.C.B., H.B., H.C-.Q., N.C., T.C., C.A.L.D., M.E.D., K.J.F., T.C.W., B.C.H., T.D.G.H., M.J., P.K., A.L., D.L., S.L., G.M., M.M., O.J.L.M., N.S., J.S.P., A.B.-T., J.T., O.K.V., M.W., G.W., W.Z., Y.Y., A.-M.V.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Rob James Boyd, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Metcalfe, D.B., Anders, E., Axén, H. et al. Gaps in tropical science from unrepresentative distribution of sampling and citation across natural terrestrial environments. Nat Commun 16, 11378 (2025). https://doi.org/10.1038/s41467-025-67617-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-67617-4
