
Abstract
The role of micro/nanoplastics (M/NPs) in the dissemination of antimicrobial resistance (AMR) remains insufficiently understood. Here, we examine how polystyrene (PS) M/NPs of varying sizes and concentrations affect AMR gene (ARG) transfer in model systems with gram-negative (Escherichia coli) and gram-positive (Enterococcus faecalis) donors. In these systems, the ARG transfer frequency is higher for intrageneric pairs than for intergeneric pairs. The 20- and 120-nm-sized PS broadly facilitate conjugation, whereas the 1-μm-sized PS selectively promotes ARG transfer to E. coli recipients, in addition to altering the expression of conjugation- and pili-associated genes. Notably, an environmentally relevant (0.1 mg/L) concentration of PS M/NPs facilitates AMR transfer in the tested systems, which correlates with increased reactive oxygen species levels, ATP levels, and cell membrane permeability in both donors and recipients. Collectively, our findings underscore the role of M/NPs in facilitating AMR spread in specific bacterial systems, providing valuable insights for understanding their potential ecological risk in water environments.
Similar content being viewed by others

Introduction
Plastic pollution is a global environmental issue that has attracted considerable attention worldwide1. Microplastics (MPs), which are derived from the degradation of larger plastic materials, are defined as particles with diameters less than 5 mm. MPs can be fragmented into nanoplastics (NPs), which typically have particle sizes less than 1 μm1 in diameter. Micro/nanoplastics (M/NPs), which are recognised as emerging contaminants of significant international concern, are widespread, long-lasting and nearly irreversible in various ecosystems2,3, including aquatic environments4,5,6 and the human gut7. Their high longevity8, with half-lives ranging from decades to more than a thousand years, makes them far more durable than most natural particles9. This persistence allows M/NPs to continuously transport and accumulate contaminants, including harmful chemicals and pathogens, across various ecosystems, thus becoming a serious and enduring environmental concern10. Thus, the ecological risks M/NPs pose and the mechanisms through which they mediate these risks are in urgent need of investigation11. Analogously, water systems can act as sources of antibiotics, facilitating the rapid development and dissemination of antimicrobial resistance (AMR)12,13. The transfer of antimicrobial resistance genes (ARGs) can lead to the acquisition of multidrug resistance by pathogens14,15, causing approximately 700,000 deaths and increased healthcare costs every year, thereby increasing the challenge of preventing and treating bacterial infections16. Hence, AMR and ARGs have become significant concerns for human health17. Various nonantibiotic substances, such as disinfectants, nonantibiotic pharmaceuticals and nanoparticles18,19,20,21, have been shown to impact the horizontal gene transfer (HGT) of plasmid-borne ARGs22. The aquatic environment serves as a reservoir for both M/NPs and ARGs, acting as a link with other ecosystems and posing potential risks to public health and the environment. Thus, investigating the ways in which ubiquitous M/NPs impact the dissemination and development of AMR is highly important and can help promote water security23.
ARGs and M/NPs can be emitted or circulated into natural and artificial environments via similar sources and pathways24. The microscale mechanisms underlying the influence of M/NPs as environmental factors on the transfer of ARGs remain to be explored. Most studies have shown that PS M/NPs promote ARG transfer through transformation25 and intrageneric conjugation26. Liu et al.27 reported that PS can inhibit the transfer of ARGs. However, to date, whether M/NPs can facilitate intergeneric conjugation, one of the primary mechanisms for the emergence of new multidrug-resistant pathogens, has not been reported26. Moreover, additional bacterial strains and plasmids should be tested to assess the generalisability of the observed M/NP-induced promotion of ARG transfer. It has been argued that particle size can affect the dissemination of AMR, despite its crucial role in governing the interaction between plastics and bacteria. However, existing research presents varying results regarding the impact of M/NP particle size on the transfer of ARGs. Additionally, the impact of environmentally relevant concentrations of M/NPs on the dissemination of AMR has been largely overlooked, despite its high ecological significance25. Thus, exploring the effects of varying sizes and environmentally relevant concentrations28 of PS M/NPs on both intragenus27 and intergenus AMR transfer through conjugation is environmentally significant. Determining whether environmentally relevant concentrations and representative sizes of M/NPs affect the HGT of ARGs via conjugation is highly important.
To fill this knowledge gap, we investigated the effects of bacterial evolutionary distance, particle size (20 nm, 120 nm, and 1 μm), and subminimum inhibitory concentration (0.1, 1, 10, 50, and 100 mg/L; Table S1) on the transfer of ARGs through conjugation between bacteria and explored the underlying mechanisms. PS pellets were chosen as representative plastic pellets because they are produced in large quantities and are often environmentally tested25. Among the three particle sizes, 20 nm PS can enter bacteria directly, 120 nm PS can enter cells through endocytosis, and 1 μm PS cannot enter cells29 (Fig. S1). PS was detected via pyrolysis‒gas chromatography time-of-flight mass spectrometry (Py-GCToF), with a semi-quantifiable result of 0.2 mg/L in river water30. PS concentrations of 0.1 and 1 mg/L are environmentally relevant28,30,31. In this study, we established an intrageneric (donor E. coli K-12 LE392 and recipient E. coli BL21(DE3)) and an intergeneric (donor E. coli K-12 LE392 and recipient P. putida KT2440) conjugative model of RP4, which encodes resistance to ampicillin, tetracycline, and kanamycin. Using gram-positive Enterococcus faecalis OG1RF carrying the pCF10 plasmid as the donor, plasmid-free E. faecalis OG1RS as the intrageneric recipient, and E. coli DH5α as the intergeneric recipient, we further tested the ability of M/NPs to promote AMR dissemination. Detailed information on the strains and plasmids can be found in Text S1 and Table S2. The underlying mechanisms promoting conjugative frequencies were revealed by phenotypic and genotypic analyses, which included a culture-based conjugation assay and the detection of reactive oxygen species (ROS), cell membrane permeability, ATP production and transcriptome responses. These findings provide evidence that PS M/NPs can promote the propagation of AMR, which is dependent on particle size, the concentration of PS and the evolutionary distance between bacteria. This study highlights the role of M/NPs as active facilitators of HGT, potentially enhancing the spread of ARGs in microbial communities, with profound ecological and human health implications, and emphasises the need for targeted strategies to mitigate M/NP pollution and AMR risk.
Results
The intrageneric conjugation frequency exceeded the intergeneric transfer frequency in the tested model bacterial systems
Plasmid transfer frequencies depend on host genotypes and plasmid‒host interactions32, which favour conjugation among phylogenetically close microorganisms33. This study further verified that conjugative transfer is more likely to occur among microorganisms with a relatively close phylogenetic distance. We found that regardless of changes in the concentration and particle size of PS, microorganisms with closer evolutionary relationships were more likely to undergo conjugation (Fig. 1). The spontaneous intrageneric transfer frequency for RP4 of ARGs was 5.20 × 10−6, whereas the spontaneous intergeneric transfer frequency was 4.71 × 10−8 (Table S3, 4). This observation also held for pCF10 (0.87×10-1 and 5.63 × 10−5 for spontaneous intrageneric and intergeneric transfer frequency, respectively; Table S5). Taking 20 nm PS as an example, the intrageneric conjugation transfer frequency was significantly greater than that of intergeneric conjugation at all concentrations used (Fig. 1a, b).

a Conjugative transfer frequency within and across genera under 20 nm PS exposure for RP4. b Conjugative transfer frequency within and across genera under 20 nm PS exposure for pCF10. c ROS production of bacterial strains in the RP4-related conjugation system without PS. d ROS production in the intrageneric and intergeneric conjugation system for RP4 without PS. e Cell membrane permeability of bacterial strains in the RP4-related conjugation system without PS. f Cell membrane permeability in the intrageneric (donor E. coli K-12 LE392 and recipient E. coli BL21(DE3)) and intergeneric (donor E. coli K-12 LE392 and recipient P. putida KT2440) conjugation systems for RP4 without PS. g ROS production of bacterial strains in the pCF10-related conjugation system without PS. h ROS production in the intrageneric and intergeneric conjugation systems for pCF10 without PS. i Cell membrane permeability of bacterial strains in the pCF10-related conjugation system without PS. j Cell membrane permeability in the intrageneric and intergeneric conjugation systems for pCF10 without PS. Statistically significant differences between intra- and intergeneric conjugation were tested with a two-sided independent-sample t test with Benjamini–Hochberg correction: *p < 0.05 and **p < 0.01(n = 6 for intergeneric system of E. faecalis, while n = 3 for the other 3 systems). Error bars represent the standard deviation (SD).
There was no significant difference in ROS production (Fig. 1c, d) or cell membrane permeability (Fig. 1e, f) between donors and recipients for RP4-related systems. However, compared with its intergeneric counterpart, the intrageneric conjugation system resulted in increased ROS production (Fig. 1g, h) and cell membrane permeability (Fig. 1i, j) in pCF10-related systems. Specific gene expression profiles are provided in Tables S6–S43. There was substantial enrichment of genes associated with the cell membrane in the donor, indicating that when strains with similar phylogenetic distances came into contact, the permeability of the donor cell membrane increased. With respect to the recipients, genes related to redox states were more enriched in the intrageneric recipients. These findings suggest that the ROS generated in the intrageneric conjugation system have a more pronounced effect on the recipient. In contrast, intergeneric conjugation exhibited the opposite pattern, with a more pronounced oxidative stress response in the donor and greater changes in the permeability of the recipient cell membrane. We speculate that the increased permeability of the donor cell membrane is more conducive to the occurrence of conjugation reactions. Although we failed to determine the conjugation frequency using P. putida KT2440 with RP4 as the donor, which may have occurred because of the detection baseline, the increased intrageneric reverse transfer ratio indicated that PS M/NPs could also promote the transfer of newly acquired ARGs to other strains, facilitating the spread of AMR (Fig. S2).
Twenty-nanometre PS broadly promoted ARG dissemination in the tested systems, whereas 120 nm PS had intermediate effects, and 1 μm PS significantly enhanced ARG transfer in E. coli recipient systems
Conjugative transfer experiments of RP4 involved exposing bacteria to various concentrations of three sizes (20 nm, 120 nm, and 1 μm) of PS microspheres for 8 or 2 h. Notably, 20 nm PS can directly enter cells29; 120 nm PS requires endocytosis for uptake, whereas 1 μm PS cannot penetrate cells, as shown by the TEM images in Fig. S1. TEM images of the conjugation system upon exposure to 120 nm PS are shown in Fig. S3. After plasmids from the donors and transconjugants were extracted, PCR amplification of the blaTEM-1 and tetA genes located on the plasmids was performed, followed by electrophoresis of both the plasmids and the PCR amplicons to confirm that the RP4 plasmids had been successfully transferred to the transconjugants (Fig. S4). The details of the independent-sample t test analysis for comparing RP4-relevant conjugative transfer to the control in gram-negative bacteria can be found in Tables S3–S4. To assess the broader applicability of PS-facilitated ARG transfer, we tested an additional model conjugation system. We used the clinically relevant gram-positive strain E. faecalis OG1RF (carrying plasmid pCF10) as the donor, with plasmid-free E. faecalis OG1RS as the intrageneric recipient and E. coli DH5α as the intergeneric recipient. These strains were exposed to polystyrene (PS) particles of three sizes (20 nm, 120 nm, and 1 μm) at four concentrations (0, 0.1, 1, and 10 mg/L). The results indicated increased conjugative transfer frequencies under PS exposure across all tested sizes and concentrations (Fig. S5). To determine whether it is the size or the chemical properties of substances that play a critical role in promoting HGT, we conducted conjugation experiments with exposure to nano-clay montmorillonite, which is a moderate chemical to bacteria, at 0, 0.1, 1, and 10 mg/L. Across all tested concentrations, nano-clay did not produce a statistically significant increase in conjugative transfer frequency compared to the control (Fig. S6, p > 0.05). By contrast, the literature reports significant increases in transfer frequency upon exposure to metal nanoparticles, such as nano-TiO234, nano-Ag/Ag20, as well as Nano-CeO₂35. Correspondingly, these nanoparticles markedly promote ROS production in all donor-recipient systems.
The influence of PS particle size on ARG transfer showed distinct and concentration-dependent patterns across different conjugation systems. At 0.1 mg/L (Fig. 2a), a nonmonotonic trend was observed: the transfer frequency increased with 20 nm PS, decreased with 120 nm PS, and increased again with 1 μm PS in E. coli-based intra- and intergeneric systems, as well as in intergeneric E. faecalis. In contrast, intrageneric E. faecalis conjugation was similarly promoted by 20 and 120 nm PS, but this effect declined with 1 μm particles. At 1 mg/L (Fig. 2b), similar patterns persisted, particularly in systems involving E. coli recipients. Under 10 mg/L (Fig. 2c), the size effect diverged further; e.g., the abundance of the intragenus E. faecalis increased with increasing size, whereas in the intergeneric system, the trend reversed. Notably, in E. coli systems, 120 nm PS began to suppress ARG transfer, whereas 1 μm PS maintained a promoting role, albeit weaker than that of 20 nm. These observations indicate a nonuniform and concentration-sensitive size effect, suggesting that both PS size and exposure level modulate the frequency of horizontal gene transfer in a system-specific manner. To rule out the possibility that the tested M/NPs affect cell viability, we calculated the total number of recipient cells as well as the cell membrane permeability and found that PS M/NPs did not significantly affect the number of recipients during the conjugation process (independent-sample t test, p > 0.05; Fig. S7). PS M/NPs have a limited inhibitory effect on the growth of recipient cells, especially at the early log phase, when we collected the cells for conjugation experiments (Fig. S8).

a Fold changes in conjugative ARG transfer frequencies under 0.1 mg/L PS with different particle sizes. b Fold changes in conjugative ARG transfer frequencies under 1 mg/L PS with different particle sizes. c Fold changes in conjugative ARG transfer frequencies under 10 mg/L PS with different particle sizes. d Expression of core genes in the RP4 plasmid in conjugation within genera. e Expression of core genes in the RP4 plasmid in conjugation across genera. f Expression of core genes in the pCF10 plasmid in conjugation within genera. g Expression of core genes in the pCF10 plasmid in conjugation across genera. Statistically significant differences between the PS M/NP-treated groups and the control (without PS M/NPs) were tested with a two-sided independent-sample t test with Benjamini–Hochberg correction: *p < 0.05 and **p < 0.01 (n = 6 for intergeneric system of E. faecalis, while n = 3 for the other 3 systems). All genes shown in the figure were significantly (p < 0.05) up- or downregulated under exposure to the tested PS M/NPs. Error bars represent the standard deviation (SD).
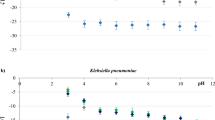
In addition to enhancing ARG transfer frequency, PS particles of different sizes exhibited distinct effects on cellular processes associated with conjugation. Importantly, we further investigated the physiological responses potentially associated with PS-facilitated ARG transfer. Among the tested parameters, ROS generation showed the strongest correlation with transfer frequency, particularly in E. faecalis-based systems, where the trends in the levels of ROS corresponded to increases in ARG transfer frequency (Fig. 3a–c). In E. coli-based systems, both ROS and membrane permeability appeared to contribute, with 1 μm PS particles notably inducing significant changes in membrane permeability (Fig. 3d–f). This finding implies a recipient-specific mechanism, possibly involving physical interactions between large PS particles and bacterial surfaces or membrane destabilization under stress. To further validate the role of ROS, we conducted ROS scavenger assays, which revealed that quenching ROS significantly reduced the PS-induced increase in conjugation frequency (Fig. S9). These findings support a model in which oxidative stress is correlated with enhanced ARG transfer under PS exposure, suggesting that ROS may contribute to the observed phenotype in the tested systems. In support of this interpretation, transcriptomic profiling revealed the upregulation of stress-responsive pathways. Excessive ROS production can drive the development and propagation of multidrug resistance36. PS M/NPs induce ROS production, DNA damage, and oxidative stress37. Notably, the addition of PS to the culture medium increased ROS production (Fig. 3a–c and S10-S11). PS M/NPs increased ROS production in both the intrageneric and the intergeneric conjugative systems, suggesting that they induced an overproduction of ROS, enhanced ROS detoxification, and stimulated the oxidative stress response (SOS response) (Fig. 3g, h). This effect correlated with increased intra- and intergeneric conjugative transfer frequencies in the tested systems. Specifically, in E. faecalis systems, the size-dependent variation in ROS production closely matched the observed changes in conjugative transfer frequency. For example, at 0.1 mg/L PS (Fig. 3a), intrageneric transfer increased, but it then decreased with increasing particle size, peaking at 120 nm, a trend mirrored by ROS levels, which reached 1.27-fold that of the control under the same conditions. These findings suggest that ROS may play a primary role in regulating conjugation in gram-positive bacteria. Conversely, in E. coli systems, the observed transfer frequencies appeared to be shaped by both ROS production and membrane permeability. While 20 nm and 120 nm PS induced significant increases in both indicators, the effect of 1 μm PS was reflected mainly in enhanced membrane permeability rather than elevated ROS. Specifically, compared with the control, 1 μm PS increased permeability by 1.26-fold in intrageneric systems. This may be attributed to the physical size of the particles, which is comparable to that of bacterial cells and could promote membrane disruption or surface interaction-based mechanisms, thereby facilitating ARG transfer via a distinct pathway. This change coincided with changes in transfer frequency, where donor- and recipient-generated ROS collectively promoted the transfer of ARGs. To determine the primary ROS involved, four types of ROS were measured in this study. Among them, singlet oxygen (1O2) and hydroxyl radicals (•OH) had the most substantial effects (Fig. S11)38. Concentrations of 0.1 and 10 mg/L 20 nm PS significantly increased the intrageneric conjugation transfer frequency, with a notable increase in singlet oxygen production at these concentrations. This suggests that singlet oxygen plays a significant role in promoting intrageneric conjugation transfer. PS with a diameter of 120 nm had a less pronounced effect on donor ROS production, with only 20 nm PS significantly increasing hydroxyl radical production in intergeneric recipients. However, as the concentration of PS increased, the concentration of hydroxyl radicals significantly increased, potentially inhibiting conjugation. This excessive hydroxyl radical production might lead to microbial death39, causing a decrease in transfer frequency.

a ROS production under 0.1 mg/L PS with different particle sizes. b ROS production under 1 mg/L PS with different particle sizes. c ROS production under 10 mg/L PS with different particle sizes. d Cell membrane permeability under 0.1 mg/L PS with different particle sizes. e Cell membrane permeability under 1 mg/L PS with different particle sizes. f Cell membrane permeability under 10 mg/L PS with different particle sizes. g Genes related to ROS production and SOS responses in intrageneric recipient E. coli BL21 (DE3). h Genes related to ROS production and SOS responses in intergeneric recipient P. putida KT2440. Statistically significant differences between the PS M/NP-treated groups and the control group were tested with a two-sided independent-sample t test with Benjamini–Hochberg correction and are indicated by *p < 0.05 and **p < 0.01 (n = 3). All genes shown in the figure were significantly (p < 0.05) up- or downregulated under exposure to the tested PS M/NPs. Error bars represent the standard deviation (SD).
In intrageneric donors, genes involved in type IV conjugative transfer (e.g., traD, traI, and traX) were upregulated under 1 μm PS treatment, except for sfmA, whose expression was downregulated by 120 nm PS (Table S31)40. With respect to intergeneric conjugation, larger PS particles inhibited donor pili production, whereas the expression of csgA, which is related to adhesion41, was suppressed by both 120 nm and 1 μm PS. Conjugative transfer via the RP4 plasmid involves the control of global regulatory genes and conjugation-related genes42. Overall, the expression of genes related to plasmid conjugation increased (Fig. 2d–g). traL, trbH, and trbM were upregulated by 20 nm PS, whereas traL was suppressed by 120 nm PS. When exposed to 0.1 and 10 mg/L PS particles, during intrageneric conjugation, 1 μm PS inhibited htrE, ppdD, and sfmH, which are related to enabling fimbrial usher porin activity (Table S16). Notably, type IV pili genes (hofN, ppdD, sfmF, and yadN) were consistently upregulated across all PS particle sizes43. In the case of intergeneric conjugation donors, the expression of genes associated with oxidative stress (Table S10), such as rclA, trxC, and ytfE, significantly increased under the influence of all the tested PS particle sizes. However, 120 nm PS notably inhibited the expression of yfcG, a gene related to oxidative detoxification, while 1 μm PS significantly downregulated the expression of yafO, a gene associated with the SOS response. This difference could contribute to the weaker promotion of intergeneric conjugation by 1 μm PS compared to its promotion of intrageneric conjugation.
When exposed to 20 nm PS at 1 mg/L, the intrageneric ARG transfer frequency increased 2.86-fold compared with that of the controls (p = 0.0004). In the intragenus gram-positive model system (E. faecalis), 20 nm PS exposure at 1 mg/L was associated with a 2.86-fold increase in intrageneric ARG transfer frequency compared with that of the controls (p = 0.0004). In contrast, 1 μm PS promoted even greater transmission from E. faecalis OG1RF to E. coli DH5α. At 1 mg/L, 1 μm PS increased the transfer frequency to 2.97 times that of the control group. Mechanistically, the suppression of prgX expression correlated with PS-enhanced conjugation (Tables S42–S43). prgX is a key transcriptional repressor in the pheromone-responsive system of E. faecalis, and its downregulation alleviates the repression of downstream genes essential for conjugation, thereby facilitating the transfer of ARGs44. With respect to the transmission of AMR between Enterococcus species, PS notably promoted this process by increasing the expression of genes related to conjugation. Specifically, in the E. faecalis intraspecific system, 20 nm and 120 nm PS exposure correlated with the upregulation of type IV secretion system-related genes, such as mobC, pcfB, pcfJ, prgB, prgM, and prgO45 and increased ARG transfer frequency. In contrast, for interspecies transfer, PS exposure correlated with reduced expression of prgX, potentially alleviating its inhibitory role in conjugation. This effect coincided with increased ARG transfer frequency (Fig. 2a–c).
PS M/NPs also promoted ROS production in pCF10-related conjugation systems. PS (20 nm and 120 nm) significantly promoted ROS production in both the intrageneric and the intergeneric conjugation systems (Fig. 2a–b). The 1 µm PS particles significantly increased ROS production in the intergeneric conjugation system but had little effect on ROS production in the Enterococcus intrageneric conjugation system. Specifically, 1 mg/L of 20 nm and 0.1 mg/L of 120 nm PS increased ROS production in the intraspecies conjugation system by 1.6-fold (p = 0.0194) and 1.28-fold (p = 0.0459) that of the control group, respectively (Fig. 3c). In the intergeneric conjugation system, all concentrations of 1 µm PS particles significantly promoted ROS production, with the greatest increase reaching 1.34-fold (p = 0.0146) of the original value (Fig. 3d). With respect to the effect of PS on individual bacteria, PS significantly affected ROS production in the three types of bacteria, but the promotion factor was relatively low for the donor (Figure. S12a). Compared with the control, the 20 nm PS particles significantly promoted ROS production in the intraspecies recipient E. faecalis OG1RS (Fig. S12b), with a maximum increase of 2.31-fold (p = 0.0367). On the other hand, 1 µm PS particles increased ROS production in the interspecies recipient E. coli DH5α by 1.40- to 1.50-fold (p = 0.0015–0.0301) the original level (Fig. S12). Specifically, 20 nm PS significantly increased ROS production (Fig. 3a-c), cell membrane permeability (Fig. 3d-f and S13-15), and ATP levels (Fig. S16) across conjugative systems, suggesting a comprehensive increase in cellular conditions that favour gene transfer. In contrast, 120 nm PS induced moderate increases in membrane permeability and ATP production (Fig. S18), with no marked difference between the intra- and intergeneric systems, indicating a more uniform but less potent biological response. Notably, 1 μm PS specifically enhanced membrane permeability (Fig. 3d–f) in systems involving E. coli, particularly when E. coli served as the recipient, which may represent a distinct mechanism by which larger particles promote ARG dissemination. This size-specific effect warrants further mechanistic investigation to determine whether physical interactions or particle uptake pathways differ depending on the recipient cell type and particle size.
Environmentally relevant concentrations of PS M/NPs increased the ARG transfer frequency in the tested systems
The impact of PS M/NPs on the conjugative transfer of ARGs correlated with fluctuations in concentrations, all of which remained below the minimum inhibitory concentration (Table S1). Despite varying fold changes in transfer frequency across different treatments, we observed a nonlinear concentration-dependent effect of PS particles on the conjugative transfer of ARGs (Fig. 4a, b). In E. coli-donor systems, the transfer frequency initially increased with increasing PS concentration, peaked at moderate concentrations, and then decreased at higher concentrations. For instance, compared with the control, 100 mg/L 20 nm and 120 nm PS significantly suppressed ARG transfer, indicating a threshold beyond which PS may exert inhibitory effects. In contrast, when E. faecalis served as the donor, 0.1 mg/L PS of all the tested sizes significantly enhanced ARG transfer, but the promoting effect diminished or plateaued with increasing concentrations. These findings suggest that low concentrations of PS facilitate ARG dissemination, whereas high concentrations may interfere with this process, possibly because of particle aggregation, toxicity, or saturation of cellular responses. Moreover, the concentration sensitivity appeared to differ between gram-negative and gram-positive donor systems, implying that concentration effects are both nonlinear and species specific. The most substantial increase in conjugative transfer was achieved at environmentally relevant concentrations (0.1 and 1 mg/L) of PS. For instance, 0.1 mg/L PS at 20 nm, 120 nm and 1 μm significantly increased the intrageneric transfer frequency by 6.52- (p = 0.0002), 3.36- (p = 0.0006), and 10.81-fold (p = 0.0005), respectively (Fig. 4a). Conversely, 100 mg/L PS at 20 nm and 120 nm reduced the transfer frequency by 0.43-fold (p = 0.0217) and 0.79-fold (p = 0.0470), respectively.

a Fold changes in conjugative ARG transfer frequencies from E. coli K-12 LE392 to E. coli BL21 (DE3) under varying PS concentrations. b Fold changes in conjugative ARG transfer frequencies from E. coli K-12 LE392 to P. putida KT2440 under varying PS concentrations. c ROS production of the intrageneric recipient E. coli BL21 (DE3). d ROS production of the intrageneric recipient P. putida KT2440. e Cell membrane permeability of the intrageneric recipient E. coli BL21 (DE3). f Cell membrane permeability of the intrageneric recipient P. putida KT2440. g Genes related to cell membrane permeability in the intrageneric recipient E. coli BL21 (DE3). h Genes related to cell membrane permeability in the intergeneric recipient P. putida KT2440. Statistically significant differences between the PS M/NP-treated groups and the control group were tested with a two-sided independent-sample t test with Benjamini–Hochberg correction and are indicated by *p < 0.05 and **p < 0.01. All genes shown in the figure were significantly (p < 0.05) up- or downregulated under exposure to the tested PS M/NPs. Error bars represent the standard deviation (SD).
Bacterial cell membranes serve as a barrier for horizontal material transfer, with their permeability affecting the movement of substances46. We found that the addition of thiourea had no significant effect on cell membrane permeability (Fig. S13), which suggests that M/NPs influence membrane permeability primarily through ROS-related mechanisms and that the role of thiourea is to specifically scavenge ROS activity without introducing any independent effect on cell membranes. Environmentally relevant concentrations of PS (0.1 and 1 mg/L) significantly promoted an increase in cell membrane permeability. (Fig. 4e,f and Fig. S14, S15). Treatment with 1 mg/L 20 nm PS M/NPs increased donor cell membrane permeability by up to 1.29-fold (p = 0.0013) (Fig. S14). This effect extended to recipients, with up to 1.35- and 1.32-fold increases (p = 0.0002–0.0075) (Fig. 4e, f). At 0.1 mg/L, the three sizes of PS increased the donor permeability by 1.38-fold (p = 0.0419), 1.30-fold (p = 0.0490), and 1.33-fold (p = 0.0381), in ascending order of particle size. However, 10 mg/L PS resulted in minor increases of 1.17-fold (20 nm), 1.13-fold (120 nm), and 1.05-fold (1 μm). The impact of higher concentrations—50 mg/L and 100 mg/L—of PS of different sizes on the permeability of the donor varied between the intrageneric and intergenic models. For intrageneric recipients (Fig. 4e), the effects of 0.1 mg/L PS of all sizes were similar. For the intergeneric recipients (Figs. 4f), 1 mg/L 20 nm PS had the greatest effect, with a 1.61-fold increase in permeability. These results suggest that the cell membrane permeability of recipient cells plays a crucial role in the conjugation process. However, 0.1 mg/L 120 nm PS downregulated the expression of aaeX and sanA (Fig. 4g). The expression levels of the phoE gene and the yghE gene differed minimally, suggesting that 10 mg/L 120 nm PS may prevent the intergeneric transfer of RP4 plasmids (Fig. 4g). In the presence of 0.1 mg/L of 1 μm PS, the expression of aaeX, pdeG (related to periplasmic enzyme activity), and the efflux pump-related gene emrY increased significantly, potentially promoting intrageneric conjugation. In intergeneric conjugation donors (Fig. 4h), 0.1 mg/L 20 nm PS had a more pronounced effect on cell membrane permeability, where genes such as bhsA, gltF, pdeG, and yjbF were significantly upregulated under the influence of all three particle sizes (Tables S14–S21).
PS could increase ATP generation in both gram-positive and gram-negative bacteria (Fig. S16 and S17). Compared with the control, 0.1–50 mg/L 1 μm PS increased the production of ATP in the donor by 2.24-fold (p = 0.0046), 3.67-fold (p = 0.0007), 2.74-fold (p = 0.0029), and 1.38-fold (p = 0.0373) (Fig. S14). The highest levels of ATP production in intrageneric recipients were observed at a concentration of 0.1 mg/L for all three particle sizes of PS, where ATP production increased to 1.33-fold, 1.50-fold, and 1.34-fold, respectively, compared with that in the control group (Fig. S16b). With respect to the intergeneric recipients (Fig. S16c), 0.1 mg/L 20 nm PS promoted the highest ATP production; however, with increasing concentrations of PS, ATP production decreased. These results confirmed the influence of PS on ATP production and highlighted that donor ATP production has a greater effect on donor transfer frequency than on that of recipients (Fig. S16). With respect to genes related to ATP generation and utilisation, most genes were upregulated under 0.1 and 10 mg/L PS within the same genus (Tables S22–29). M/NPs upregulated the expression of the citC gene, which encodes citrate lyase synthetase, in E. coli, thereby affecting the tricarboxylic acid cycle and promoting ATP production in bacterial cells (Tables S22 and S24).
In summary, environmentally relevant concentrations of PS M/NPs promoted the conjugation frequency through enhancing ROS production, promoting cell membrane permeability and facilitating ATP generation. Intragenus conjugation systems of RP4 were characterised by substantial changes in recipient cell membrane permeability (Figs. S14, S15), accompanied by oxidative state alterations and increased ATP generation (Fig. S16, S17) in donors, which correlated with increased ARG transfer frequencies. In contrast, intergenus conjugation involved oxidative stress-induced changes and ATP production (Fig. S16–S18) both in donors and recipients, whereas recipients exhibited more favourable cell membrane (Fig. S14) conditions for plasmid uptake. In pCF10-related conjugation, PS M/NPs exhibited a greater promoting effect on recipients.
Discussion
The dissemination of AMR is a serious global threat to public health and environmental ecology12. Prior research has underscored HGT as a primary mechanism by which ARGs are transferred from one microorganism to another in water environments15. In this study, we demonstrated that PS M/NPs, at environmentally relevant concentrations and varying sizes, can significantly promote the conjugative transfer of ARGs both within and across bacterial genera in the tested model systems. This enhancement was associated with cellular stress responses in recipient bacteria, including elevated levels of ROS, increased levels of intracellular ATP, and increased membrane permeability. Additionally, our results highlight that the promotion of HGT by nanoparticles is primarily driven by their chemical properties. Specifically, their ability to induce oxidative stress and disrupt bacterial membranes (Table S48). PS micro/nanoplastics and metal nanoparticles (e.g., nano-TiO234) showed significant effects, while nano-clay montmorillonite (Figure. S6), a less reactive material, did not. These findings reveal a previously underrecognized pathway through which plastic pollution may contribute to the spread of AMR in the environment, emphasising the importance of considering the chemical properties of materials in evaluating their ecological impact.
In our study, 20 nm PS broadly promoted ARG dissemination in the tested systems, whereas 120 nm PS had intermediate effects, and 1 μm PS significantly enhanced the transfer in E. coli recipient systems. Given the widespread contamination of M/NPs and ARGs in aquatic and gut environments47 and their coexistence with complex microbial communities, understanding M/NP-mediated AMR dissemination is critical. Current research suggests that M/NPs promote both transformation25 and intragenus conjugation27,48. Among the various modes of HGT, conjugation is the most prevalent natural means through which ARGs can disseminate22; this allows ARGs to be exchanged within or across genera, thereby creating new ecological niches for microorganisms49. Most studies have focused on plastics with particle sizes exceeding 75 nm25,50. Importantly, environmental shear forces continuously fragment plastics into smaller, higher-risk nanoparticles51, which is consistent with findings in which PS with particle sizes smaller than 100 nm enhanced exogenous ARG uptake25. Our study reveals a strong size-dependent hierarchy in PS-mediated conjugative transfer. We found that 20 nm PS, which is capable of entering bacterial cells through membrane pores, facilitated the spread of ARGs both within and across genera among the tested gram-negative and gram-positive bacterial strains, thereby increasing the likelihood of the emergence of multidrug-resistant microorganisms. These findings align with those of studies showing that NPs can recruit pathogenic ARG hosts in wetlands52, alter membrane transport to potentiate resistance53 and increase soil ARG abundance to a greater extent than microplastics are capable of54. PS (120 nm) had intermediate effects, mirroring size-dependent risks in constructed wetlands52. Unlike transformation, where large plastics ( > 1 μm) have minimal effects25, our study demonstrated that 1 μm PS, which is similar in size to microorganisms, remains consequential for the dissemination of AMR to E. coli. However, all the experiments were performed under simplified laboratory conditions. The effects of aged or weathered M/NPs, multispecies biofilms, and natural aquatic matrices remain unknown and warrant further laboratory and in situ validation55.
For a specific particle size, the impact of PS N/MPs on AMR is dose dependent. Previous studies have typically employed experimental concentrations starting at 5 mg/L27, which are significantly higher than environmental concentrations5. Here, we demonstrated that environmentally relevant concentrations (0.1–1 mg/L) were sufficient to induce significant effects. Indeed, exposure to as low as 0.1 mg/L of 20 nm PS nanoparticles increased the transfer frequency by up to 8.9-fold56. Field evidence also supports this trend, with the abundance of microplastics (14.9 items/g faeces) positively correlated with that of ARGs (6.24 × 10 8 copies/g) and laboratory exposure, increasing conjugation by 1.4–1.7-fold57. Notably, the relationship is nonmonotonic: small NPs can first stimulate but then suppress transfer at higher doses44, and extreme concentrations of microplastics (2000 mg/L) reduce the conjugation frequency by an order of magnitude27. In systems with E. coli carrying RP4, PS M/NPs at higher concentrations (10, 50, and 100 mg/L) may hinder cell-to-cell contact or block conjugative pili and toxicity58, thereby reducing the conjugation efficiency and negatively affecting AMR transmission. For the E. faecalis system, we specifically examined environmentally relevant concentrations but did not evaluate higher concentration exposures. The effects of environmentally relevant concentrations of PS on promoting the dissemination of AMR are not negligible and may have adverse implications for water safety, which underscores the ecological plausibility of our findings and suggests that sublethal exposure to M/NPs may drive nonmonotonic stress responses, with important implications for environmental AMR proliferation59.
This study explored the interplay of the dissemination of ARGs among microorganisms at different evolutionary distances in model systems using gram-negative (E. coli) and gram-positive (E. faecalis) donors (Fig. 5a). Owing to the potential transfer of ARGs between different bacterial genera, originally non-resistant pathogens may acquire resistance to antibiotics, consequently increasing the difficulty of treatment. Therefore, the intergeneric spread of AMR is highly worthy of investigation. While most previous studies have focused on intrageneric conjugation events, particularly within model organisms such as E. coli25,48,50, the role of M/NPs in facilitating intergeneric ARG transfer has remained largely unexamined. We found that intrageneric conjugation transfer frequencies far exceeded those of their intergeneric counterparts in the tested conjugation systems. This observation aligns with the notion that plasmids are more likely to propagate between microorganisms with closer evolutionary relationships. Such a phenomenon is advantageous for mitigating the emergence of novel multidrug-resistant pathogens32. Moreover, our results reveal that PS particles can enhance conjugative gene transfer not only within species but also across species, including between gram-positive and gram-negative bacteria. These findings contrast with those of single-cell studies by Liu et al.27, who reported inhibitory effects of PS on the intrageneric conjugation of E. coli, highlighting the context-dependent and potentially species-specific nature of interactions between M/NPs and microbes. By demonstrating the PS-induced increase in cross-genus gene transfer, our study advances the current understanding of HGT dynamics in plastic-polluted environments. Despite these novel insights, we should acknowledge that the intergeneric conjugation results were derived from a limited number of donor–recipient pairs and only 2 plasmids (RP4 and pCF10), which may not fully capture the diversity of conjugation mechanisms present in environmental microbiomes. Plasmid compatibility, host restriction systems, and recipient recognition vary substantially across taxa; thus, broader validation with diverse bacterial hosts and mobile genetic elements is needed.

a Enhancement of RP4 plasmid conjugative transfer by PS M/NPs. b Enhancement of pCF10 plasmid conjugative transfer by PS M/NPs. c Mechanistic basis of PS M/NP-induced conjugative transfer: The introduction of PS M/NPs increases ROS production, enhances detoxification processes, and increases cell membrane permeability in both donor and recipient bacterial strains. This physiological state promotes close proximity between donor and recipient cells, facilitated by mating pair assembly and pilus channel formation. Consequently, PS M/NPs trigger the activation of replication and transfer processes of the RP4 plasmid through the pilin channel, enabling efficient plasmid transfer from the donor to the recipient. G +: gram-positive; G -: gram-negative; D: donor; R: recipient. Intra-R: intrageneric recipient; Inter-R: intergeneric recipient; Intra-T: intrageneric transconjugant; Inter-T: intergeneric transconjugant.
Mechanistically, PS M/NPs facilitate ARG transfer through size-, concentration-, and strain-specific pathways involving oxidative stress, membrane perturbation, and energy metabolism42,60. Our data revealed that 20 nm PS induced the greatest increase in the production of ROS, particularly •OH61, and increased membrane permeability and ATP levels, whereas 1 μm PS increased E. coli membrane permeability without increasing the corresponding level of ROS, suggesting a ROS-independent route distinct from that reported for metal nanoparticles20. The bacterial outer membrane, acting as a semipermeable barrier, facilitates the entry of plasmids when permeability is increased46. Exposure to environmentally relevant concentrations of PS increased cell membrane permeability, which is associated with an elevated transfer frequency. At the molecular level, PS exposure upregulated genes in the tested systems involved in the mating pair formation (Mpf) system (traL, trbH, trbM)58,62,63; in efflux pumps53,64; and, in gram-positive E. faecalis donors, the phosphotransferase system (PTS) and teichoic acid biosynthesis (Tables S44–47), potentially contributing to species-specific conjugation responses. These stress- and metabolism-linked pathways may collectively promote plasmid mobilisation, natural transformation25, and both intra- and intergenus conjugation, underscoring the multifactorial nature of PS-mediated AMR propagation. While the associations between physiological stress and enhanced transfer were consistent, the partial restoration of conjugation frequency by ROS scavengers supports a partial causal role for oxidative stress, while the direct contributions of ATP production and membrane destabilization remain correlative and should be further clarified through genetic, chemical, or imaging-based approaches.
In light of increased pollution levels from plastics globally11, M/NPs act as a nexus for AMR dissemination across humans, animals, and ecosystems, underscoring the necessity of integrating One Health principles into research and mitigation strategies for AMR. The recalcitrance of PS M/NPs contributes to their significant accumulation in the environment for many years, causing serious pollution. Considering that PS is a versatile plastic used to make a wide variety of consumer products65, our findings constitute a wake-up call to start evaluating the potential antibiotic-like effects induced by PS M/NPs. Collectively, the findings of the present study provide evidence that the widely distributed PS M/NPs can contribute to the HGT of ARGs via conjugation within and across genera in water environments (Fig. 5b). Given the complexity of microbial communities in natural aquatic environments, the transmission of AMR within relatively large bacterial communities is intricate and multifaceted22. Future work should investigate whether other M/NP types and aged M/NPs exhibit similar or stronger ARG transfer potential, confirm the direct causal mechanisms driving enhanced transfer, incorporate integrative multiomics to elucidate the underlying molecular pathways, and assess species-specific responses in complex habitats such as soil, freshwater, marine ecosystems, wastewater, and gastrointestinal environments. Given the persistence and ubiquity of plastics, even low-level pollution may accelerate microbial evolution by promoting the horizontal dissemination of ARGs. These findings underscore the urgent need for interdisciplinary research to quantify public health risks and to inform mitigation strategies in the context of a One Health framework aimed at curbing the environmental spread of AMR. In conclusion, our study provides a more comprehensive understanding of the effects of plastic particles on the dissemination of ARGs and provides a theoretical basis for controlling the risks posed by the spread of ARGs mediated by plastic pollution, which has now become ubiquitous in the environment.
Methods
Bacterial strains, growth conditions, and nano-clay montmorillonite
Detailed information on the strains and plasmids can be found in Text S1 and Table S2. For RP4-related conjugation, the donor strains used in this study were E. coli K-12 LE392—carrying RP4 plasmids containing resistance genes for ampicillin (Amp), kanamycin (Kan), and tetracycline (Tet)—and E. faecalis OG1RF carrying pCF10 (TetR). The recipient strains E. coli BL21 (DE3) (ChlR), E. faecalis OG1RS (StrR), P. putida KT2440 (ChlR), and E. coli DH5α (EryR) were selected for intrageneric and intergeneric conjugation experiments. E. coli K-12 LE392 and P. putida KT2440 were sourced from the Institute of Urban Environment, Chinese Academy of Sciences. E. faecalis OG1RF and E. faecalis OG1RS were kindly provided by the Tianjin Institute of Environmental and Operational Medicine66. E. coli BL21 (DE3) and E. coli DH5α were obtained from Beijing BIOBW Company. The culture conditions for the donor and recipient bacteria are shown in Text S1. PS particles of three different sizes (120 nm, 1 μm, and 100 μm) were obtained from Huge Biotechnology (Shanghai, China). Nano-clay montmorillonite (50 nm) was purchased from Sigma (Germany).
Conjugation experiments in the presence of PS M/NPs or nano-clay montmorillonite
This study established two mating models to explore whether PS M/NPs could enhance ARG transfer. In the environmentally relevant model from E. coli K-12 LE391 with RP4 to E. coli BL21 (DE3) or P. putida KT2440, donors and recipients at 108 cfu/mL were mixed 1:1 in PBS, for a total volume of 20 mL. Various concentrations of PS at different sizes (20 nm, 120 nm, and 1 μm) were added to the mating system, including environmentally relevant and sub-MIC concentrations, i.e., 0, 0.1, 1, 10, 50, and 100 mg/L28. For nano-clay montmorillonite, we did the conjugation experiments under 4 different concentration (0, 0.1, 1 and 10 mg/L). After 8 h of incubation at 25 °C without shaking, 100 μL of the mixture was spread onto LB selection plates containing antibiotics to enumerate transconjugants and recipients. The transfer frequency was calculated by dividing the number of transconjugants by the total number of recipients. To investigate the role of ROS, an ROS scavenger that can remove ROS from the system, thiourea67, was added at a final concentration of 100 μM in some experiments. Detailed information is described in Text S2.
For conjugation from E. faecalis OG1RF with pCF10 to E. faecalis OG1RS or E. coli DH5α, the mating systems were mixed 1:1 using brain heart infusion (BHI) medium and mated for only 2 h to avoid vertical gene transfer. Twenty microlitres of the mixture was spread onto BHI selection plates containing antibiotics to enumerate transconjugants and recipients, with detailed information described in Text S2.
Reverse conjugation experiments under exposure to PS M/NPs
Transconjugant bacteria obtained from the conjugation experiment (E. coli BL21(DE3) and P. putida KT2440 with RP4 plasmid) were used as the donors, and E. coli NK5449, with resistance to Rif, was used as the recipient. The conjugation mating systems were set up as described above and included different sizes of PS M/NPs at environmentally relevant concentrations. The mating systems were then mixed by vortexing and incubated at 25 °C for 8 h without shaking. Afterwards, the mating systems were mixed and used to inoculate selection plates containing antibiotics, which were subsequently incubated as described above. Finally, the colonies of the transconjugants and recipients were counted, and the transfer frequencies were determined as described above.
Transmission electron microscopy (TEM)
TEM (Tecnai G2 Spirit, USA) at 120 kV was used to observe the effect of PS M/NPs on bacterial cells. TEM samples were collected from conjugation systems supplemented with 1 mg/L 120 nm PS or E. coli K-12 LE392 after 2 h of mating with either 1 mg/L 20 nm, 120 nm or 1 μm PS. Details are provided in Text S3.
Measurement of reactive oxygen species (ROS)
The bacterial strains were subsequently washed twice with PBS and resuspended in PBS to 106 cfu/mL. To detect total ROS, the bacterial strains were incubated with 2’,7’-dichlorofluorescein diacetate (DCFDA; final concentration, 20 μM; Abcam®) for 30 min at 37 °C in the dark. Afterwards, 100 μL of bacteria stained with DCFDA were treated with different concentrations of PS M/NPs. For the controls, 1.5% H2O2 was used as the positive control, and ethanol was used as the negative control. After complete mixing, the mixture was incubated at 25 °C in the dark for 2 h, after which the absorbance was measured at 488 nm using a microplate reader (Spark 10 M, Tecan). All the detections were conducted in triplicate (Text S4).
Detection of cell membrane permeability
For cell membrane permeability detection, 100 μL of the strain was exposed to different concentrations of PS M/NPs and incubated at 25 °C for 2 h. The strains were then stained with 1 μL of propidium iodide (PI; 2 mM; Life Technologies) and incubated in the dark for 15 min before being measured at 561 nm. To rule out the possibility that thiourea may affect cell membrane permeability and thus affect conjugation frequency, the cell membrane permeability of RP4-relevant bacterial strains was detected after the addition of thiourea and PS M/NPs as described above. All the detections were conducted in triplicate (Text S5).
Detection of ATP generation
To measure ATP content (Text S5), cells were collected by centrifugation in a centrifuge tube, and the supernatant was discarded. The cells were gently resuspended and then lysed. Chemiluminescence intensity was measured using a microplate reader to determine the changes in ATP production under the influence of different particle sizes and concentrations of PS M/NPs. All the detections were conducted in triplicate (Text S6).
RNA extraction, genome-wide RNA sequencing, and transcriptomic analysis
RNA extraction was carried out after the conjugation systems were treated with three different sizes of PS (0, 0.1, and 10 mg/L) for 2 h. Each system was replicated in biological triplicates. After being subjected to quality control, the pellets were sent to MAGIGENE Co. (Shenzhen, China) for RNA extraction, construction of strand-specific cDNA libraries, and Illumina sequencing of paired-end genomes. An Illumina HiSeq PE150 was used. An NEBNextő Ultra II Directional RNA Library Prep Kit for Illumina was used for library construction. Sequence reads consisting of at least 85% bases were then progressively trimmed at the 3’-ends until a quality value ≥ 20 was maintained. Downstream analyses were performed using the generated clean reads that were no shorter than 75 bp. The clean reads of each sample were aligned to the E. coli reference genome (NC_000913), P. putida reference genome (NC_002947), and IncPα plasmid reference genome (L27758) using Bowtie2. RSeQC was used to calculate the strand-specific coverage for each gene and to analyse the differential expression in triplicate bacterial cell cultures. The statistical analyses and visualisation were conducted using edgeR and the CummeRbund package in R. Gene expression was calculated as fragments per kilobase of a gene per million mapped reads (FPKM, a normalised value generated from the frequency of detection and the length of a given gene). The difference in gene expression between the control group (without the addition of PS M/NPs) and the PS-treated groups is represented as the log2-fold change (LFC), which is defined as the base-2 logarithm of the ratio of expression levels in the treatment group relative to the control group. The significance of the transcriptome data was determined by a false discovery rate (FDR)-adjusted p value of less than 0.05. All samples were prepared in triplicate.
Plasmid verification
To verify that the plasmids had entered the cells, the plasmids of the donors and transconjugants were extracted using the Invitrogen PureLink Quick Plasmid Miniprep Kit (Life Technologies, USA). The blaTEM-1 and tetA genes of the RP4 plasmid were amplified by PCR, and the resulting amplicons as well as the RP4 plasmid were checked using 1% agarose gel electrophoresis. The PCR primers and conditions are described in Text S7.
Statistical analysis
All experiments were conducted independently at least in biological triplicate. All phenotypic data are expressed as the mean ± SD and were analysed with GraphPad Prism 10 and SPSS 25.0 (SPSS, Chicago, USA). The results were analysed by analysis of variance and two-sided independent-sample t test methods with Benjamini–Hochberg correction. A corrected p value less than 0.05 was considered to indicate statistical significance.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data generated or analysed during this study are included in this published article, its Supplementary Information and the accompanying Source Data file. All RNA sequencing data have been deposited in the NCBI Gene Expression Omnibus under accession codes GSE248909 and GSE297944. Source data are provided with this paper.
References
Dawson, A. L. et al. Turning microplastics into nanoplastics through digestive fragmentation by Antarctic krill. Nat. Commun. 9, 1001 (2018).
Koelmans, A. A. et al. Microplastics in freshwaters and drinking water: Critical review and assessment of data quality. Water Res 155, 410–422 (2019).
Rochman, C. M. Microplastics research-from sink to source. Science 360, 28–29 (2018).
Wu, N. Tracing microplastic footprints through the plastisphere. Nat. Rev. Earth Environ. 3, 498–498 (2022).
Materic, D. et al. Micro- and nanoplastics in alpine snow: a new method for chemical identification and (semi)quantification in the nanogram range. Environ. Sci. Technol. 54, 2353–2359 (2020).
Seeley, M. E., Song, B., Passie, R. & Hale, R. C. Microplastics affect sedimentary microbial communities and nitrogen cycling. Nat. Commun. 11, 2372 (2020).
Yan, Z. et al. Analysis of microplastics in human feces reveals a correlation between fecal microplastics and inflammatory bowel disease status. Environ. Sci. Technol. 56, 414–421 (2022).
Koelmans, A. A. et al. Risk assessment of microplastic particles. Nat. Rev. Mater. 7, 138–152 (2022).
Allen, D. et al. Microplastics and nanoplastics in the marine-atmosphere environment. Nat. Rev. Earth Environ. 3, 393–405 (2022).
Kumar, M. et al. Current research trends on micro- and nano-plastics as an emerging threat to global environment: a review. J. Hazard Mater. 409, 124967 (2021).
MacLeo, M., Arp, H. P. H., Tekman, M. B. & Jahnke, A. The global threat from plastic pollution. Science 373, 61–65 (2021).
Andersson, D. I. & Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Microbiol. 12, 465–478 (2014).
Zainab, S. M., Junaid, M., Xu, N. & Malik, R. N. Antibiotics and antibiotic resistant genes (ARGs) in groundwater: a global review on dissemination, sources, interactions, environmental and human health risks. Water Res. 187, 116455 (2020).
Ellabaan, M. M. H., Munck, C., Porse, A., Imamovic, L. & Sommer, M. O. A. Forecasting the dissemination of antibiotic resistance genes across bacterial genomes. Nat. Commun. 12, 2435 (2021).
Jiang, X. et al. Dissemination of antibiotic resistance genes from antibiotic producers to pathogens. Nat. Commun. 8, 15784 (2017).
May, M. Tomorrow’s biggest microbial threats. Nat. Med. 27, 358–359 (2021).
Zhang, Z. Y. et al. Assessment of global health risk of antibiotic resistance genes. Nat. Commun. 13, 1553 (2022).
Lu, J. & Guo, J. H. Disinfection spreads antimicrobial resistance. Science 371, 474–474 (2021).
Wang, Y. et al. Non-antibiotic pharmaceuticals promote the transmission of multidrug resistance plasmids through intra- and intergenera conjugation. ISME J. 15, 2493–2508 (2021).
Lu, J. et al. Both silver ions and silver nanoparticles facilitate the horizontal transfer of plasmid-mediated antibiotic resistance genes. Water Res. 169, 115229 (2020).
Zhang, Y., Gu, A. Z., He, M., Li, D. & Chen, J. Subinhibitory concentrations of disinfectants promote the horizontal transfer of multidrug resistance genes within and across genera. Environ. Sci. Technol. 51, 570–580 (2017).
Shi, X., Xia, Y., Wei, W. & Ni, B. J. Accelerated spread of antibiotic resistance genes (ARGs) induced by non-antibiotic conditions: Roles and mechanisms. Water Res. 224, 119060 (2022).
Redondo-Hasselerharm, P. E. Gort, G. Peeters, E. & Koelmans, A. A. Nano- and microplastics affect the composition of freshwater benthic communities in the long term. Sci. Adv. 6 eaay4054 (2020).
Li, B. et al. Metagenomic and network analysis reveal wide distribution and co-occurrence of environmental antibiotic resistance genes. ISME J. 9, 2490–2502 (2015).
Hu, X., Waigi, M. G., Yang, B. & Gao, Y. Impact of plastic particles on the horizontal transfer of antibiotic resistance genes to bacterium: dependent on particle sizes and antibiotic resistance gene vector replication capacities. Environ. Sci. Technol. 56, 14948–14959 (2022).
Brito, I. L. Examining horizontal gene transfer in microbial communities. Nat. Rev. Microbiol. 19, 442–453 (2021).
Liu, L., Zhang, Q.-H. & Li, R.-T. In situ and individual-based analysis of the influence of polystyrene microplastics on Escherichia coli conjugative gene transfer at the single-cell level. Environ. Sci. Technol. 57, 15936–15944 (2023).
Xu, Y., Ou, Q., Jiao, M., Liu, G. & van der Hoek, J. P. Identification and quantification of nanoplastics in surface water and groundwater by pyrolysis gas chromatography–mass spectrometry. Environ. Sci. Technol. 56, 4988–4997 (2022).
Della Torre, C. et al. Accumulation and embryotoxicity of polystyrene nanoparticles at early stage of development of sea urchin embryos Paracentrotus lividus. Environ. Sci. Technol. 48, 12302–12311 (2014).
Ivleva, N. P. Chemical analysis of microplastics and nanoplastics: challenges, advanced methods, and perspectives. Chem. Rev. 121, 11886–11936 (2021).
Enfrin, M. et al. Release of hazardous nanoplastic contaminants due to microplastics fragmentation under shear stress forces. J. Hazard. Mater. 384, 121393 (2020).
Redondo-Salvo, S. et al. Pathways for horizontal gene transfer in bacteria revealed by a global map of their plasmids. Nat. Commun. 11, 3602 (2020).
Rodríguez-Beltrán, J. et al. Beyond horizontal gene transfer: the role of plasmids in bacterial evolution. Nat. Rev. Microbiol. 19, 347–359 (2021).
Cheng, P. et al. Fe-doped TiO2 nanosheet exposure accelerates the spread of antibiotic resistance genes by promoting plasmid-mediated conjugative transfer. J. Hazard. Mater. 490, 137715 (2025).
Yu, K. et al. CeO2 Nanoparticles regulate the propagation of antibiotic resistance genes by altering cellular contact and plasmid transfer. Environ. Sci. Technol. 54, 10012–10021 (2020).
Kohanski, M. A., DePristo, M. A. & Collins, J. J. Sublethal antibiotic treatment leads to multidrug resistance via radical-induced mutagenesis. Mol. Cell 37, 311–320 (2010).
Huang, S. et al. Metagenomic analysis reveals the responses of microbial communities and nitrogen metabolic pathways to polystyrene micro(nano)plastics in activated sludge systems. Water Res. 241, 120161 (2023).
Duan, J. J. et al. ROS-mediated photoaging pathways of nano- and micro-plastic particles under UV irradiation. Water Res. 216, 118320 (2022).
Kohanski, M. A., Dwyer, D. J. & Collins, J. J. How antibiotics kill bacteria: from targets to networks. Nat. Rev. Microbiol. 8, 423–435 (2010).
Ho, B. T., Basler, M. & Mekalanos, J. J. Type 6 secretion system-mediated immunity to type 4 secretion system-mediated gene transfer. Science 342, 250–253 (2013).
DeBenedictis, E. P., Liu, J., & Keten, S. Adhesion mechanisms of curli subunit CsgA to abiotic surfaces. Sci. Adv. 2 (2016).
Yu, Z. G., Wang, Y., Lu, J., Bond, P. L. & Guo, J. H. Nonnutritive sweeteners can promote the dissemination of antibiotic resistance through conjugative gene transfer. ISME J. 15, 2117–2130 (2021).
Maier, B. & Wong, G. C. L. How bacteria use type IV pili machinery on surfaces. Trends Microbiol. 23, 775–788 (2015).
Dunny, G. M. & Berntsson, R. P. Enterococcal sex pheromones: evolutionary pathways to complex, two-signal systems. J. Bacteriol. 198, 1556–1562 (2016).
Kohler, V., Keller, W. & Grohmann, E. Enterococcus adhesin PrgB facilitates type IV secretion by condensation of extracellular DNA. Mol. Microbiol. 109, 263–267 (2018).
Thomas, C. M. & Nielsen, K. M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 3, 711–721 (2005).
Alimi, O. S., Budarz, J. F., Hernandez, L. M. & Tufenkji, N. Microplastics and nanoplastics in aquatic environments: aggregation, deposition, and enhanced contaminant transport. Environ. Sci. Technol. 52, 1704–1724 (2018).
Zha, Y. Y. et al. Size-dependent enhancement on conjugative transfer of antibiotic resistance genes by micro/nanoplastics. J. Hazard. Mater. 431, 128561 (2022).
von Wintersdorff, C. J. et al. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 7, 173 (2016).
Wang, X. X. et al. A neglected risk of nanoplastics as revealed by the promoted transformation of plasmid-borne ampicillin resistance gene by. Environ. Microbiol. 24, 4946–4959 (2022).
Besseling, E., Redondo-Hasselerharm, P., Foekema, E. M. & Koelmans, A. A. Quantifying ecological risks of aquatic micro- and nanoplastic. Crit. Rev. Environ. Sci. Technol. 49, 32–80 (2019).
Zhao, Y. et al. Size-dependent promotion of micro(nano)plastics on the horizontal gene transfer of antibiotic resistance genes in constructed wetlands. Water Res. 244, 120520 (2023).
Xu, Y., Li, H., Ding, Y., Zhang, D. & Liu, W. How nanoscale plastics facilitate the evolution of antibiotic resistance?. J. Hazard. Mater. 480, 136157 (2024).
Liu, L., Sun, Y., Du, S., Li, Y. & Wang, J. Nanoplastics promote the dissemination of antibiotic resistance genes and diversify their bacterial hosts in soil. Eco-Environ. Health 3, 1–10 (2024).
Bottery, M. J., Wood, A. J. & Brockhurst, M. A. Temporal dynamics of bacteria-plasmid coevolution under antibiotic selection. ISME J. 13, 559–562 (2019).
Liu, X. et al. Nano- and microplastics aided by extracellular polymeric substances facilitate the conjugative transfer of antibiotic resistance genes in bacteria. ACS ES&T Water 2, 2528–2537 (2022).
Yu, X. et al. Microplastics exacerbate co-occurrence and horizontal transfer of antibiotic resistance genes. J. Hazard Mater. 451, 131130 (2023).
Chen, M.-M., Zhang, Y.-Q., Cheng, L.-C., Zhao, F.-J. & Wang, P. Photoaged nanoplastics with multienzyme-like activities significantly shape the horizontal transfer of antibiotic resistance genes. J. Hazard. Mater. 475, 134884 (2024).
Zhou, C. S. et al. Micro) nanoplastics promote the risk of antibiotic resistance gene propagation in biological phosphorus removal system. J. Hazard. Mater. 431, 128547 (2022).
Wu, Y. et al. Biochar effectively inhibits the horizontal transfer of antibiotic resistance genes via restraining the energy supply for conjugative plasmid transfer. Environ. Sci. Technol. 423, 127150 (2022).
Klümper, U. et al. Metal stressors consistently modulate bacterial conjugal plasmid uptake potential in a phylogenetically conserved manner. ISME J. 11, 152–165 (2017).
Gigault, J. et al. Nanoplastics are neither microplastics nor engineered nanoparticles. Nat. Nanotechnol. 16, 501–507 (2021).
Huang, H., Liao, J., Zheng, X., Chen, Y. & Ren, H. Low-level free nitrous acid efficiently inhibits the conjugative transfer of antibiotic resistance by altering intracellular ions and disabling transfer apparatus. Water Res. 158, 383–391 (2019).
Maier, L. et al. Extensive impact of non-antibiotic drugs on human gut bacteria. Nature 555, 623–628 (2018).
Mitrano, D. M. & Wohlleben, W. Microplastic regulation should be more precise to incentivize both innovation and environmental safety. Nat. Commun. 11, 5324 (2020).
Yang, Y. et al. Bisphenols promote the pheromone-responsive plasmid-mediated conjugative transfer of antibiotic resistance genes in Enterococcus faecalis. Environ. Sci. Technol. 56, 17653–17662 (2022).
Fang, F. C. Antibiotic and ROS linkage questioned. Nat. Biotechnol. 31, 415–416 (2013).
Acknowledgements
This work was supported by the National Natural Science Foundation of China (grant nos. 52321005 awarded to A.W., 52293441 awarded to S.-H.G., 52293443 awarded to A.W., and 52070060 awarded to S.-H.G.), the Natural Science Foundation of Guangdong Basic and Applied Basic Research Foundation (Grant No. 2024A1515010085 awarded to S.-H.G.), the Shenzhen Overseas High-level Talents Research Startup Program (No. 20200518750C awarded to S.-H.G.), the Shenzhen Overseas High-Level Talent Innovation and Entrepreneurship Special Fund (No. KQTD20190929172630447 awarded to S.-H.G.) and Shenzhen Science and Technology Program (Nos. GXWD20231127195344001 awarded to A.W. and S.-H.G. and JCYJ20241202123735045 awarded to S.-H.G.), and the State Key Laboratory of Urban-rural Water Resource and Environment (Harbin Institute of Technology) (No.2025TS39 awarded to S.-H.G.). We would like to thank Prof. Zhigang Qiu from the Tianjin Institute of Environmental and Operational Medicine for donating the bacterial strains E. faecalis OG1RF and E. faecalis OG1RS, and Dr. Casey Huang and Dr. Lyman Tze Kin Ngiam from the Australian Centre for Water and Environmental Biotechnology for proofreading the paper. Figure 5 was designed, composed, and edited using BioRender (Kang, Y. (2025) https://BioRender.com/ofu614f), ChemDraw, and Adobe Illustrator.
Author information
Authors and Affiliations
Contributions
Y.K. and S.-H.G. designed the overall experiments for this study. Y.K. performed all analyses, conducted the RP4-relevant conjugation experiments within and across genera, measured ROS levels, and detected changes in cell membrane permeability and ATP generation in RP4-relevant strains. S.-H.G. supervised and managed the project and contributed to the writing and revision of the manuscript. Y.K. and S.-H.G. wrote the full manuscript and illustrated all the figures provided. Y.P., T.L. and R.G. performed the pCF10-relevant conjugation experiments and corresponding measurements of ROS production, changes in cell membrane permeability and ATP generation; Y.S. and W.Z. performed the experiments, analysed the data, and revised the manuscript. J.G. and Z.Y. contributed to the initial planning for this study, provided guidance on the research significance of this study, and contributed to revising the manuscript. L.F. and B.L. assisted in analysing the mechanisms, ecological significance, and potential application scenarios of this study. J.S. and Y.L. provided the donor and recipient strains and provided feedback on the conjugation experiments. Y.W. analysed the transcriptomic data and determined the changes in the expression of related genes under different concentrations and particle size PS treatments. A.W. provided guidance on the concentration and particle size of PS in the study and contributed to revising the manuscript. All the authors provided feedback and discussed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Lorenzo Brusetti, Raffaella Sabatino, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Kang, Y., Gao, SH., Pan, Y. et al. Roles of micro/nanoplastics in the spread of antimicrobial resistance through conjugative gene transfer. Nat Commun 17, 1118 (2026). https://doi.org/10.1038/s41467-025-67879-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-67879-y
