
Abstract
Sortilin is a ubiquitous membrane receptor mediating trafficking of protein cargoes. In the thyroid, sortilin binds thyroglobulin (TG) during its endocytosis, a key process in thyroid homeostasis. Although sortilin has been proposed to recognise highly iodinated TG, the molecular details of this interaction remain unknown. In this work, using an integrative structural biology approach, we reveal that sortilin binds an unstructured TG C-terminal peptide and exhibits a strong preference for the monomeric TG over the commonly known dimeric form. We find that sortilin-TG interaction is independent of the iodination state of TG and instead relies on the conversion to its monomeric state, presumably promoted by extracellular degradation. Furthermore, using AlphaPulldown and sequence analysis, we show that recognition of other reported ligands by sortilin likely relies on similar unstructured peptide motifs, which are not constrained to a single binding orientation within the receptor cavity. Overall, this study reveals the TG-sortilin binding interface and provides insights into the recognition mechanism of other cargoes by sortilin.
Similar content being viewed by others


Introduction
In vertebrates the expression of most metabolic genes is controlled by thyroid hormones (TH), essential iodinated compounds synthesised and secreted by the thyroid gland into the bloodstream, thereby reaching other tissues1,2. The synthesis of TH is mediated by iodination of the large glycoprotein thyroglobulin (TG)3,4,5,6 (dimer ~660 kDa). The release of TH as modified amino acids, embedded in the TG polypeptide chain, occurs via TG endocytosis followed by proteolysis in the lysosome, before the TH selective transport outside the thyroid follicle7,8,9,10,11,12,13. Sortilin has been proposed as a TG post-endocytic receptor and to preferentially recognise highly iodinated TG14, thus probably playing a role in TH release and thyroid homoeostasis.
Sortilin (SORT1, monomer ~90 kDa) is a ubiquitous type-1 receptor belonging to the vacuolar protein sorting 10 protein family, bearing a conserved N-terminal β-propeller domain (the ectodomain, ECD), a single-pass transmembrane helix anchor and a cytosolic domain15,16. Previous crystallographic studies indicate that the peptide cargo neurotensin (NT) binds within the central cavity of the sortilin β-propeller and that its dimerisation on the same binding site, occurring at acidic pH, may promote cargo release in endocytic compartments16,17. Currently, sortilin has been shown to mediate trafficking and lysosomal degradation of various cargoes implicated in cancer, cardiovascular and neurodegenerative disease18,19,20,21. Notably, several protein tags mediating lysosomal degradation via sortilin binding have been devised to control the degradation of extracellular proteins22,23,24. However it remains unclear at a structural level how sortilin can recognise such a large variety of natural cargoes of different size—and, within the thyroid, how it discriminates between iodinated and uniodinated TG25.
In this study we elucidate the molecular recognition of TG by sortilin using an integrative structural biology approach, combining cryo-EM, biophysics, cross-linking mass spectrometry (crosslinking MS) and AlphaFold predictions. Our structural findings are corroborated by cellular endocytosis assays in human thyroid cell lines. Our results indicate that sortilin binds the C-terminal peptide of TG in the same binding pocket of NT, unexpectedly in an antiparallel fashion. We found that binding of sortilin to the TG dimer is prevented by steric clash, suggesting that molecular recognition by sortilin relies on TG adopting a monomeric state, rather than being determined by its iodine content. We hypothesise that the TG transition from dimer to monomer could arise from extracellular degradation, a well-known process in the thyroid9,26. In addition leveraging structural data and AlphaFold predictions, we derived a ligand peptide binding motif, which seems independent on the peptide orientation within the sortilin cavity. Taken together our work provides insights into the role of sortilin-mediated TG trafficking in the thyroid and it lays the basis for the structural understanding of general ligand recognition by the sortilin ECD.
Results
Sortilin preferentially interacts with monomeric TG, independently of iodination
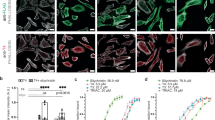
To investigate sortilin-TG interaction and its variation upon iodination, we performed in vitro complex reconstitution of the human his-tagged sortilin ECD (from now on named simply sortilin or sort) with endogenously extracted human TG (eTG). To avoid bias towards a specific TG state, we chose to work with eTG because it naturally contains a mixture of non-iodinated and naturally iodinated protein14 (Fig. 1a).

a Schematic representation of thyroglobulin (TG) and sortilin (sort) forms used in our experiments. b SDS-PAGE showing binding of his-tagged sortilin ectodomain to endogenous human untagged TG (eTG) by pull-down on Ni-NTA beads. c Size exclusion chromatography (SEC) profile and corresponding SDS-PAGE of his-tagged sortilin preincubated with TG. Sortilin-TG coelution as a monomer-monomer complex (SorTG) is highlighted in yellow d Mass photometry (MP) profile of sortilin preincubated with either mTG (orange) or dTG (grey). The mass of SorTG confirms the interaction of monomeric sortilin with mTG. e Representative confocal images of three independent immunofluorescence experiments: intracellular TG was detectable after 120 min in Nthy-ori 3–1 cells overexpressing full-length sortilin-V5 either in the absence or in the presence of 0.1 mg/mL TG in the cellular medium. TG internalisation depends on the presence of sortilin and it is largely increased when TG is enriched in mTG. Scale bar: 20 µm. f Quantification of TG cellular internalisation. P-values are based on the Kruskal–Wallis test (****p < 0.0001). Mean values ± SD (number of cells = 159 at 0 min, number of cells = 76 for eTG at 120 min, number of cells = 87 for dTG at 120 min, number of cells = 113 for mTG at 120 min). Experiments were performed in triplicate n = 3. A.U.: arb units g MP profiles of recombinant TG (rTG, purple) and iodinated rTG (i-rTG, black), showing that iodination does not affect TG oligomerisation state. h Percentage of SorTG complex formed in vitro (circles) and of TG internalised in cells (squares) plotted against the content of TG monomer present in each sample. The plot suggests a correlation between these two parameters. TG internalised by cells was detected by immunofluorescence assays, measuring the average of TG signal mean intensity for each cell, normalised at t = 0 min (background). t = 0 corresponds to the time point in which the cells are not exposed to TG (right before adding it in the medium). The TG signal detected at this time is considered as unspecific background. All the following TG internalisation signal values were normalised with respect to this time point.
The formation of the sortilin-TG complex (SorTG) was tested by sortilin pull-down assays (Fig. 1b). A band corresponding to the molecular weight of TG was detected in the elution fraction but not in the absence of sortilin, confirming this interaction to be specific. To further characterise the SorTG complex and to determine its stoichiometry, the two proteins were pre-incubated together and the mixture was subjected to analytical size exclusion chromatography (SEC). A significant portion of protein remained unbound, showing the interaction to be weak. Unexpectedly, the SorTG complex eluted in SEC at a lower hydrodynamic radius with respect to the dimeric TG (dTG) (Fig. 1c). From this result, we hypothesised that sortilin could bind monomeric TG (mTG) rather than the commonly found dimeric TG form (dTG). Given the variable amount of mTG in extracted batches, we decided to generate samples of higher monomeric content via gentle heat shock of the stable recombinant TG, which is fully dimeric (Supplementary Fig. 1a–d, e). Indeed, in our hands, other additives could not allow conversion of dTG to mTG (Supplementary Fig. 2a). The heat shock procedure was optimised to produce the maximum amount of monomer with minimal thermal exposure, as measured by mass photometry (MP) (Supplementary Fig. 1b). We additionally analysed the mTG sample obtained via heat shock by negative staining, verifying that it bears a similar architecture of the extracted monomer and that it is not unfolded (Supplementary Fig. 1e). MP analysis of sortilin, preincubated with samples either enriched in mTG or dTG, confirmed that the complex forms in solution as a species of ~405 kDa, closely matching the sum of the masses of mTG (~330 kDa) and monomeric sortilin ECD (~80 kDa) (Fig. 1d). The complex formation was not observed by incubating sortilin with dTG. To confirm this finding in a cellular context, we performed TG internalisation assays in human thyroid cell lines Nthy-ori 3–1 overexpressing the full-length sortilin bearing a V5 tag at its C-terminus (Fig. 1e, f). Indeed, we found that mTG is preferentially uptaken over dTG. Based on previous studies reporting stronger interaction of sortilin with iodinated TG14 we surmised that the transition of TG from a dimeric to a monomeric form could be induced by its iodination, thereby enhancing its binding to sortilin. However, recombinant dTG subjected to in vitro iodination4 neither showed formation of mTG (Fig. 1g) nor was able to bind sortilin, as measured by MP. Binding of iodinated TG to sortilin was detected only when TG was further processed to induce the formation of mTG (Supplementary Fig. 1f). Indeed, we observe a correlation between the amount of TG cellular intake or SorTG formation and the mTG content of the sample (Fig. 1h). Taken together, these results suggest that the interaction of sortilin with TG is determined by the TG monomeric content rather than its iodination state. If not iodination, we asked what could drive the conversion from the stable TG dimer to a more relaxed or monomeric form and how this could be connected via a signalling mechanism to thyroid hormone homoeostasis. We hypothesised that this conversion could be promoted by extracellular proteolysis, known to be driven by cathepsins in the thyroid10. We subjected recombinant dTG to cathepsin treatment, but this did not lead to monomer formation, probably due to some suboptimal in vitro proteolysis conditions. Nevertheless, limited proteolysis with chymotrypsin of the stable recombinant dTG resulted in an increase in mTG content (Supplementary Fig. 2b, c), supporting the hypothesis of mTG formation via TG degradation. Yet the obtained amounts of mTG from proteolysed dTG were too little to perform further studies, hence we decided to obtain larger yields of mTG via our established heat shock protocol.
Sortilin interacts with the TG C-terminal peptide
To gain structural insights into the SorTG interaction we first characterised the isolated monomer-monomer SorTG complex by negative staining electron microscopy (NS-EM). Our low-resolution NS-EM map resembled the elongated mTG volume decorated with an extra density similar in size to sortilin (Supplementary Fig. 3a). Yet from our fit, we could not unambiguously infer the relative orientation of the two protein moieties. To improve the stability of the complex and increase the occupancy of sortilin, we then set to use either bis(sulfosuccinimidyl)suberate (BS3) cross-linking (XL) or GraFix27 (Fig. 2a and Supplementary Fig. 3b, c, e). This crosslinking procedure helped to enrich the sample with SorTG complex; however it did not allow its complete separation from free mTG and dTG (Supplementary Fig. 3f). Cryo-EM analysis of the GraFix-derived sample enriched for the SorTG complex resulted in an EM density clearly corresponding to sortilin. This volume faces a poorly resolved extra density pointing towards the NT binding site, opposite to the C-terminal cysteine-rich tail of sortilin16 (Supplementary Fig. 3d). Likely, this low-resolution density represents mTG, but despite signal subtraction attempts, we could not further resolve it, probably due to the high flexibility of the elongated mTG. The high degree of flexibility and solvent exposure is also supported by the high amount of long-range internal crosslinks within TG as measured by crosslinking MS experiments (Supplementary Data 1 and Supplementary Fig. 4h). To identify the SorTG interface, we attempted using AlphaFold328 prediction of the full-length mTG in complex with the sortilin ECD. The proteins appear to have a low chance of interacting, with an interface predicted template modelling (ipTM) score of 0.31 and a predicted aligned error (PAE) between the two protein chains greater than 30 Å (Supplementary Fig. 4a, b). Hence, to experimentally define a region of interaction between sortilin and the large TG sequence, we pursued crosslinking MS experiments with the photo-crosslinker sulfo-NHS diazirine (sulfo-SDA) on the SorTG complex. Our data suggest that the SorTG binding interface may be restricted to the TG choline esterase-like (ChEL) domain, a C-terminal region deeply involved in TG dimerisation3,29 (Fig. 2b; Supplementary Figs. 4e, f and 5b). In particular, TG lysine residues 2677, 2698, 2712 are crosslinked to several sortilin residues of the β-propeller (TG K2677 is crosslinked to sortilin residues E136, G138, I141, T152, A153, V155, S156, S159 and H360; TG K2698 is crosslinked to sortilin residues V155 and G157; TG K2712 is crosslinked to sortilin residues P533 and Y535) (Supplementary Data 1; Supplementary Fig. 4e–h and 6a–d). Notably, the region around residue 2677 is buried in the TG dimer but exposed in the monomer (Fig. 2b and Supplementary Fig. 4e, f), explaining the undetectable binding of sortilin to dTG. Based on the interface suggested by crosslinking MS, we restricted the AlphaFold3 prediction to sortilin with the ChEL domain of TG. The resulting prediction was overall more reliable and indicated a high-confidence interaction between sortilin and the unstructured C-terminal peptide of TG, where at least two residues exhibited a PAE of less than 6 Å (Supplementary Fig. 4a, c). AlphaFold3 predictions of the sortilin binding to other macrodomains of TG did not show a reliable interaction score (Supplementary Fig. 7). The crosslinking MS experiment was performed with multiple injections on a single sample prepared in large amount. Despite we could not assign a reproducibility error to each crosslink, this experiment was key to help us narrowing down the sortilin binding region to the C-terminal domain of TG. Some statistics on the collected data are provided in Supplementary Fig. 8.

a Western blot analysis of sortilin and TG samples cross-linked with BS3. The yellow bands show co-localisation of sortilin and TG (SorTG complex). b Crosslinking MS analysis (performed with sulfo-SDA) of the SorTG complex and relative structural mapping to the C-terminal ChEL domain. The interaction of sortilin with mTG (model on the left) is sterically allowed, while it is prevented by steric clash with the dTG (model on the right). c Cryo-EM map of sortilin (green) in complex with the TG C-terminal peptide (orange), σ = 11.84. d Cryo-EM map of ligand-free sortilin, σ = 11.73. e Close-up view of the sortilin-TG binding site, supported by the relative cryo-EM density map. The TG peptide presents two interaction sites within the sortilin β-propeller: site 1 (f) and site 2 (g).
To experimentally validate the interaction of sortilin with the TG C-terminal peptide (TGpep), we first verified binding of the TG synthetic peptide to sortilin by differential scanning fluorimetry (DSF), which showed a clear dose-dependent response (Supplementary Fig. 9a). Despite this experiment did not lead us to determine a reliable Kd, it prompted us to undertake cryo-EM analysis of sortilin in complex with TGpep. We analysed both ligand-free sortilin and sortilin in the presence of the synthetic TGpep (SorTGpep), by obtaining cryo-EM maps at 3.10 and 3.36 Å, respectively (Fig. 2c, d; Supplementary Fig. 10 and Supplementary Table 1). Our sortilin cryo-EM model within the SorTGpep model closely matched previous crystallographic structures16,17,30 (RMSD 0.922 Å with respect to PDBID 4PO7). Indeed, the sortilin model features a main N-terminal ten-bladed β-propeller (residues 45–576) followed by two small cysteine-rich domains (10CC-a and 10CC-b, residues 577–633 and 634–716) interacting with the β-propeller. Additionally glycosylation sites Asn129, Asn373 and Asn549 could be built in our density map (Supplementary Fig. 5a).
While the ligand-free and TGpep-bound models show a similar overall fold for the receptor (RMSD 0.396 Å), in the SorTGpep complex map we clearly observe an extra density within the β-propeller cavity (Fig. 2c, d), which we could build as the TGpep polypeptide chain (Fig. 2e–g).
TGpep binds sortilin in the same region of the well-characterised cargo NT16 in two main sites within the β-propeller cavity, by analogy, site 1 and site 216,25. The site 1 is occupied by the N-terminal portion of TGpep (Fig. 2e, f), where the carboxyl group of D6 (2754 in the full-length TG) forms a salt bridge with the guanidinium group of the conserved sortilin R29216. Additionally D6 establishes hydrogen bonds with the side chain of sortilin S283 as well as the main chain of Y318. Moreover, TGpep L8 docks into a hydrophobic pocket near sortilin I294 and F281 (Fig. 2f). Overall the DLL motif (residues 6–8 in TGpep) fits into a hydrophobic pocket between blades 5 and 6 of the sortilin β-propeller. The site 2 of sortilin, on the opposite side of the β-propeller, is occupied by the C-terminal residues of TGpep (15–19), forming a β-sheet with the blade 1 of sortilin. Additionally, TGpep residue Y18 (Y2766 in the full-length TG) fits into a hydrophobic pocket, where it establishes an interaction with W553 (Fig. 2g).
TG, neurotensin and the inhibitor AF38469 bind the same sortilin region
The SorTGpep structural analysis showed that a short TG peptide and NT occupy the same binding pocket within the sortilin β-propeller, engaging with both site 1 and site 2. However unexpectedly, the two peptides bind in an anti-parallel orientation with respect to each other and only by performing an inverted sequence alignment we could identify similarities between the two polypeptide stretches (Fig. 3d). In the high-affinity site 1, the conserved sortilin R292 interacts with an acidic residue at the N-terminal side of the TGpep while it binds the C-terminus carboxylate group of NT in the reported crystal structure16,29 (Fig. 3a, d). A similar electrostatic interaction in site 1 is established by the sortilin inhibitor AF38469 (AF) 31 (Fig. 3a). In the lower affinity site 216,29, TG engages through its C-terminal portion SKTYS (residues 15–19; corresponding to 2763–2767 in the full-length TG) while NT binds via its N-terminal segment LYEN (residues 2–5). Both peptide portions exhibit a propensity to form β-strands and primarily interact with sortilin through backbone hydrogen bonding surrounding the tyr residue, by forming a β-sheet with blade 1 of the sortilin propeller (residues 78–83) (Fig. 2g).

a Comparison of three ligands bound to the sortilin (green) R292 in site 1: neurotensin (NT, cyan) (PDBID 4PO7), the inhibitor AF38469 (AF, magenta) (PDBID 4N7E) and the TG C-terminal peptide (TGpep, orange). b Analysis of SorTG complex in the absence or in the presence of either NT, AF or TGpep. The dotted line indicates the background noise baseline. The normalisation was performed on the SorTG complex sample. Data are presented as mean values ± SD (number of biological replicates n = 9 with SorTG, n = 5 with SorTG + AF, n = 4 with SorTG + NT, n = 8 with SorTG + TGpep). P-values are based on ordinary one-way ANOVA (****p < 0.0001). c Quantification of TG cellular internalisation (prior sortilin overexpression) in the presence of the AF inhibitor (left) or TGpep (right). The dotted line indicates the average of TG mean intensity at t = 0 (background). The normalisation was performed without TG (at t = 0) in DMSO for AF and in buffer A for TGpep. P-values are based on the Kruskal–Wallis test (**p = 0.0034, ****p < 0.0001). Data are presented as mean values ± SD (left: AF competition. Number of cells n = 107 at 0 min (background), n = 185 for mTG at 120 min, n = 156 for mTG at 120 min + AF; right: peptide competition. n = 112 at 0 min (background), n = 164 for mTG at 120 min, n = 142 for mTG at 120 min + peptide); number of biological replicates n = 3. d Superimposition of the SorTGpep complex and the NT-sortilin structure, showing that the two cargoes bind the same sites but with polypeptide chains running in opposite directions. e Consensus motif for known cargoes binding sortilin, derived by sequence alignment and AlphaPulldown predictions. Grey scheme: secondary structure prediction for the consensus peptide. f Ligand binding curves and relative affinities of TGpep (dotted, black), TGpep in DMSO (orange), TGpep-T4 (pink), TGpep mutated (grey), NT (cyan) and sortilin propeptide (green) for purified sortilin, measured in vitro by MST. Data presented as mean values ± SD (number of biological replicates, n = 3). g Peptide sequences, with chain direction indicated by the arrow on the right.
To confirm whether NT, AF and the isolated TGpep could compete with full-length TG binding, we ran SorTG in vitro binding assays in the presence of the three compounds and analysed the results via MP (Fig. 3b). In addition, we performed similar competitive TG cellular internalisation experiments in human thyroid cell lines (Fig. 3c). In all cases we observed a significant reduction either in SorTG formation or in the TG cellular uptake. In addition, we performed a titration of the TGpep in the presence of the SorTG complex (Supplementary Fig. 9b) revealing a dose-dependent inhibition by the isolated peptide. Altogether, these results not only validate that NT, AF and TGpep bind to the same pocket, but also suggest that existing sortilin ligand binding inhibitors could interfere with sortilin-mediated TG trafficking in the thyroid.
AlphaFold predictions of sortilin bound to its cargoes
Several sortilin-binding partners involved in critical biological processes are reported in the literature18. When we initiated this study, only the interaction with NT was well characterised and progranulin was reported to bind in a similar fashion and peptide direction, with their C-terminus within sortilin site 1 (PDB ID: 8T8R, 8T8S). Given the different orientation of the TGpep within the sortilin cavity, we set out to investigate which pose is adopted by other protein ligands and whether a consensus binding motif could be identified for other sortilin partners. Leveraging high-confidence TGpep prediction from AlphaFold3 (ipTM of 0.78, with an RMSD of ~2 Å with our experimental structure) (Supplementary Fig. 4a, d, g), we combined AlphaPulldown32,33 and a sequence alignment pipeline to predict the structure of sortilin in complex with the 55 reported binders18 (Supplementary Fig. 11). First, we predicted their interaction with sortilin using AlphaPulldown run with AlphaFold2.3 weights32,33, considering both the full-length proteins and their reported physiological proteolytic fragments. The predictions were then filtered based on PAE scores (cutoff = 8 Å inter-chain) and the models were visually inspected to select the cargo binding within the sortilin β-propeller. We excluded partners binding via their cytosolic region as they presumably would not be in contact with the sortilin β-propeller within a cellular context. If the models presented minor steric clashes, they were improved using AlphaFold3 (Supplementary Fig. 11).
Structural comparison of the best ten selected models relative to cargoes of different size shows that they consistently bind the sortilin β-propeller via an unstructured peptide of approximately 20 residues in either an NT-like or TG-like direction (Supplementary Fig. 11 and Supplementary Table 2).
From a visual inspection in the sortilin site 1 the interaction with the partner peptides is established via either the protein C-terminus (NT-like) or an acidic moiety. Conversely in site 2 interactions with sortilin are primarily mediated by backbone and side-chain contacts, involving a conserved large aromatic residue (Supplementary Fig. 11 and Supplementary Table 2). These two sortilin-binding elements are separated by a proline or proline-rich region in the peptide. Among the predicted models, we found that the sortilin propeptide, crucial for the receptor function, binds in the same direction as TG. Other partners follow the NT-like main chain orientation, yet two do not interact via their C-terminus, but via an internal acidic residue (for example the amyloid beta precursor-like protein 2, APLP2)34. After this preliminary model analysis, we pursued exclusive sequence alignment of the sortilin binding peptides, according to the pose adopted in the cavity. We added an aspartic acid to the sequences where the binders interacted via the C-terminus in sortilin site 1, in order to mimic the carboxylate group moiety in the alignment (Supplementary Table 2). Using the PSSMSearch server35 we then analysed the aligned sequences and derived a 20-amino acid sortilin binding motif (Fig. 3e), highlighting similar interactions of different cargo peptides at site 1 and 2, spaced by a central proline-rich region. Taken together, this computational structural analysis provided insights into a possible general recognition mechanism of cargo peptides by sortilin.
Binding assays of sortilin to cargo peptides
This study highlighted that sortilin ECD may bind a set of cargoes via a peptide recognition motif and that the specific binding to TG is independent of its iodination. To validate these findings, we measured the binding affinity of various peptides to sortilin using microscale thermophoresis (MST) (Fig. 3f, g). As expected, NT binds sortilin with a Kd of 140 nM, in agreement with previous reports16. The sortilin propeptide sequence selected via our sequence alignment shows a Kd of about 1 µM, while the TGpep shows a Kd of about 7 µM. Conversely, we could not detect binding of sortilin to the TGpep where key residues involved in the sortilin interaction site 1 and site 2 (5-EDL-7 and Y18) were mutated.
Furthermore, we noticed that the TGpep bears an hormonogenic tyrosine (at Y18 corresponding to Y2766 in TG, also named site C) and we inquired whether the presence of an iodo-thyronine at this position could affect the binding to sortilin. Hence, we measured the binding of the receptor to the TGpep where Y18 was replaced with the thyroxine hormone (T4), but the derived Kd of 7 µM was comparable to the unmodified peptide. It must be noted that the T4-containing peptide is much less water-soluble, so we had to run the comparative measurement of TGpep in the presence of DMSO (Fig. 3f, g). In addition to further investigate the effect of hormonogenesis on the SorTG complex, we modelled the T4 moiety within the sortilin cavity using Coot, based on our experimental structure36 (Supplementary Fig. 12). In this computational model the T4 extra iodo-phenol ring is exposed to the solvent and the hormone seems establishing similar interactions of a tyrosine side chain with sortilin site 2, without steric clashes.
Taken together, these results and analysis again confirm our hypothesis that the molecular recognition of TG by sortilin is specific and it is not affected by the iodination of the thyroid hormone precursor. In addition these results corroborate the mechanism proposed for the interaction between the sortilin β-propeller and cargo peptides.
Discussion
Sortilin (or NT receptor 3)15,37 is a widely expressed membrane receptor mediating protein sorting and cell signalling in various cell types38,39. Sortilin binds a diversity of ligands, including neurotrophins, lipoproteins, cytokines and amyloid-related proteins and is also implicated in severe human disorders18,19,20,21,40. Among its multiple trafficking roles41,42,43,44, a key sortilin function is to mediate lysosomal delivery via transport along the trans-Golgi (TGN) and endosomal network38,45,46. The importance of sortilin is highlighted by recent protein design efforts and drug repurposing to modulate its activity in neurodegenerative disease or to mediate lysosomal degradation of specific protein targets22,23,24,47.
Structural information on its ligand recognition has been available only for the NT peptide (PDB ID: 3F6K, 4PO7) and for progranulin (PDB ID: 8T8R, 8T8S), where a flexible peptide docks in the β-propeller of the receptor, via interaction with the C-terminus (sortilin site 1)16,30. Despite the growing list of identified sortilin partners involved in pathogenesis18,48 the molecular basis of general recognition by sortilin of many other targets remains unclear25.
In the thyroid, TG is secreted in large amounts in the lumen of the gland follicles, where iodination of its tyrosine residue generates TH embedded in the protein main chain and provides iodine storage for the organism1,5. Only trafficking this iodine and hormone-rich TG to the lysosome is productive for hormone release and iodine recycling8,13; however this critical process remains poorly characterised at a structural and molecular level. In this context, sortilin has been reported to be a TG post-endocytic receptor and to preferably interact with the highly iodinated form of TG14. Here we set out to investigate sortilin specificity for different TG forms to reveal implications in TG cellular sorting.
A combination of structural biology methods revealed that sortilin ECD recognises TG cargo via a C-terminal flexible peptide (TGpep), which was not previously resolved in cryo-EM TG structures3,49,50,51. We surmise that while an initial TG recognition by sortilin might occur via this flexible TGpep, equally exposed in the TG monomer and dimer, a more stable interaction is established in a TG conformation where the ChEL residues around K2677 are exposed, as suggested by our crosslinking MS data (Fig. 2b). Such interaction with sortilin would be hindered in the TG dimer but may presumably occur, as supported by our data, either in the mTG form (already reported to exist in the thyroid) or in an intermediate dimeric relaxed form52,53,54,55,56. These findings are strongly corroborated by cellular assays in which we tested sortilin-mediated internalisation of different forms of TG, featuring a strong preference for mTG uptake (Fig. 1e–h).
Previous studies suggested that more iodinated TG is preferentially recognised by sortilin relying on interaction studies with TG batches containing different amounts of iodine14. However in our hands, iodinating TG in vitro did not enhance its interaction with sortilin, nor induced a transition from dTG to mTG, more prone to sortilin binding (Fig. 1g and Supplementary Fig. 1f). Moreover the affinity of sortilin for the unmodified TGpep and for a TGpep enclosing the T4 hormone is very similar (Fig. 3f, g). This evidence is further supported by modelling the T4-bearing peptide in complex with sortilin (Supplementary Fig. 12), where the hormone does not show either steric clashes or additional interactions with respect to the unmodified tyrosine (Y2766 in the FL TG). Therefore we hypothesise that the previously observed higher affinity of sortilin for TG batches with increased iodine content reported by Marinò and coauthors could have resulted from the concurrent presence of higher amounts of mTG in endogenous samples, not measured at that time14. Taken together, we propose that TG iodination and sortilin-mediated endocytosis are independent events and that the interaction of sortilin with TG is determined by the presence of a monomeric (or a relaxed) TG form (Fig. 4).

a Both TG and sortilin exist in monomeric and dimeric forms. The SorTG complex is established primarily between the monomeric forms of both proteins. Acidic pH conditions promote sortilin dimerisation and ligand release. b Secreted dimeric TG (dTG) can exist in both iodinated and uniodinated forms. Extracellular degradation of both forms is likely generating either a relaxed or a monomeric TG state (mTG). In these species the exposed TG C-terminal peptide (orange) is recognised by sortilin. Hence, sortilin-mediated TG trafficking to the lysosome leads to the release of thyroid hormones (TH) from iodinated TG.
In the thyroid TG is secreted as a dimer and is progressively cross-linked into an extracellular colloid in various iodination states9,26. TG must be disentangled from the colloid to become available for endocytosis, thereby allowing intracellular thyroid hormone release26,57,58 (Fig. 4). Several reports highlight that extracellular cathepsins are involved in this process, targeting TG for proteolysis. Indeed, recombinant TG (secreted by HEK cells) is a more stable and a less degraded dimer than the endogenous thyroid TG (exposed to cathepsins). This is in line with previous cryo-EM studies in which the recombinant TG shows a higher occupancy for the N-terminal domain compared to the extracted sample3. Hence we propose that while TG is secreted as a dimer, which more efficiently forms hormones than a monomer59, over time its extracellular proteolysis may lead to a monomeric or relaxed state, more prone to bind sortilin. Independently of its iodine content, mTG is then recruited by sortilin within the endocytic pathway, likely culminating into hormone release (Fig. 4) and iodine recycling. This hypothesis is in agreement with the observation that both cathepsin and sortilin expression are upregulated by the thyroid hormone receptor (TSH), responsible to increase TG internalisation and the levels of thyroid hormone in the bloodstream. In conclusion, we suggest that, rather than iodination, sortilin-mediated trafficking may be linked to TG proteolysis9,14,60. Despite this assumption we could not fully recapitulate mTG formation in vitro by proteolysis with cathepsins, but we could observe an increase of mTG content upon limited proteolysis with chymotrypsin (Supplementary Fig. 2c). Likely our cathepsin proteolysis setup may be quite different compared to in vivo conditions.
Although sortilin is confirmed to be a receptor for TG, Sort1 knockout in mice61 does not significantly impair hormonogenesis, but it may partially affect male individuals. Given our work performed on human proteins, we believe that further studies in humans are needed to confirm the impact of sortilin in TG trafficking and thyroid homoeostasis. Indeed, sort1 knock out could lead to compensatory mechanisms involving other membrane trafficking factors known to be upregulated by TSH61,62,63,64.
Based on the identified SorTG contacts we also inspected whether any mutation identified in patients could potentially impair the binding of the two proteins and thus be related to pathogenesis65,66. Within the most confident interaction region with sortilin (the TGpep), the Y2766N mutation (affecting the hormonogenic Y2766) has been related to congenital hypothyroidism65. From this study we infer that this mutation could potentially impair not only thyroid hormone synthesis itself but also TG endocytosis mediated by sortilin in site 2 (Fig. 2e–g). Yet it is arduous to deconvolute the two effects. A large number of reported pathogenic mutations are located within the TG ChEL domain a region that our crosslinking MS experiments suggest to be close to the interaction interface with sortilin. We infer that these mutations could affect not only the folding of this crucial TG internal chaperone domain (leading to ER stress, goitre and hypothyroidism) but also possibly impact TG trafficking, influencing the thyroid phenotype8,67.
Overall despite the role of sortilin in the thyroid still remains to be fully elucidated in vivo, the molecular recognition of TG by sortilin described in this work may guide more informed in vivo and in cellulo experiments on TG trafficking to evaluate the impact of such interacton on thyroid physiopathology. In this context our structural data indicate that existing sortilin inhibitors and binding peptides may interfere with TG trafficking, endocytosis and thyroid homoeostasis31.
While previous sortilin-cargo complexes were characterised by X-ray crystallography, in this work we provide a cryo-EM workflow to readily determine the structure of the fully glycosylated sortilin ECD. It must be noted that the identification of the SorTG interface posed some challenges due to the monomer flexibility and the low affinity between the two proteins, sample heterogeneity as well as the inability of predicting a likely interface with AlphaFold. Only the integration of multiple structural information together with educated AlphaFold predictions successfully led us to the identification of the sortilin-interacting TGpep. The common binding site but different orientation of TGpep with respect to NT and progranulin peptides prompted us to derive a common recognition signature in sortilin cargoes, which is compatible with two polypeptide chain directions.
Hence, leveraging restricted alignments and high score AlphaFold predictions, we could extend the search of the interface between sortilin and other reported cargoes, revealing that likely a ‘sortilin tag’ motif exists in disordered regions of proteins to be captured by the receptor. Within our list of selected cargoes, we can identify two classes of peptides: one establishing with the main site 1 R292 a stronger interaction via the C-terminus (e.g. NT) and others via acidic moieties (e.g. the TGpep and the sortilin propeptide). This different affinity may rely on the lower pKa of the C-terminus with respect to the acidic side chains, affectingthe electrostatic interaction with R292. We indeed observed for NT an affinity of about 10-fold higher than the sortilin propeptide and this could be reconducted to functional roles in the cell, which are not yet understood. As for TGpep, we explain the lower binding affinity (7 µM) with respect to the analogous sortilin propeptide with the large local concentration of TG in the thyroid follicular lumen26,58, granting, even at this Kd, an efficient binding. Overall a comprehensive cargo recognition rule for full-length sortilin is yet to be fully determined, as other protein binding modes outside the β-propeller have been reported68.
In conclusion the structural features identified for TG-sortilin complex update the current knowledge on TG trafficking and suggest broad cargo binding principles to the sortilin β-propeller. This knowledge may be crucial for identifying new sortilin-binding proteins and to improve protein engineering strategies aimed at controlling sortilin-mediated lysosomal degradation in several tissues, including in the thyroid22,23,24.
Methods
DNA constructs
The gene for human recombinant thyroglobulin (rTG) was prepared as described before3. The gene encoding for human sortilin luminal segment, residues 1–755, modified by the addition of a 10x histidine tag at the C-terminus preceded by a TEV cleavage site, was codon-optimised for mammalian expression and purchased from GenScript. The gene was cloned in a pLEXm vector with EcoRI/XhoI restriction sites (enzymes purchased from NEB). C-terminal residues 756–831 followed by V5 tag were inserted by Gibson assembly from fragments purchased from GenScript. All translated sequences of recombinant proteins used in this work are listed in Supplementary Fig. 13.
Protein expression and purification
Extracted TG (eTG) from human thyroid glands was purchased from Millipore (609312-1 mg). This eTG is partially degraded and contains some monomer in a batch-dependent measure (Supplementary Fig. 1c). To enrich for monomer content, eTG at 2 µM was heated at 60 °C for 120 s and the monomer was purified by size-exclusion chromatography (SEC) using a Superose 6 Increase 3.2/300 column (GE Healthcare), equilibrated with buffer A (20 mM HEPES, 150 mM NaCl, pH 7.5).
To produce both recombinant TG and sortilin Expi293F™ cells (Thermo Fisher Scientific) were cultured in Dynamis™ AGT™ Medium (Thermo Fisher Scientific), supplemented with 1% (v/v) Pen/Strep (Euroclone) and 4 mM stable Glutamine (Euroclone) at 130 rpm, 8% CO2, 37 °C, 85% humidity. Cells were grown to a concentration of 2–3 × 106 cells/ml with a cell viability greater than 95%. Transfection was performed without previous media change. Per liter of culture, 1.1 mg of plasmid and 3 mg of polyethyleneimine (PEI Prime, Sigma-Aldrich) were pre-diluted separately in 25 mL of pre-warmed media each before combining them. After an incubation for 20 min at room temperature the 50 mL of plasmid-PEI mixtures were added dropwise to the cells. Protein expression was carried out for 4 days. Cells were removed by centrifugation at 1000 × g for 10 min, then the supernatant was cleared by a second centrifugation at 22,000 × g for 45 min and filtration (0.22 µm). The supernatant was diluted 1:1 with buffer A and supplemented with 1x cOmpleteTM EDTA-free protease inhibitor cocktail (Roche) before loading it on the HisTrap Excel 5 ml column with Äkta Pure system (Cytiva) for affinity chromatography at 4 °C (1 ml/min). The column was washed in two steps of 5 column volumes (CV) using buffer A supplemented with 50 mM imidazole and 10 CV with buffer A supplemented with 100 mM imidazole. Hence, the protein was eluted with 10 CV of buffer A supplemented with 500 mM imidazole (all buffers adjusted to pH 7.5). Chromatographic fractions were analyzed by NuPAGE™ 3–8%, tris-acetate SDS-PAGE (Thermo Fisher Scientific) and the ones containing his-protein were pooled and concentrated before purification by SEC using a Superose 6 Increase 10/300 column (GE Healthcare). Fractions containing the protein of interest were joined, concentrated and stored at −80 °C in flash-frozen aliquots at about 2 mg/ml. The final yields of proteins were about 2–3 mg/L of cells for rTG and 7 mg/L of cells for sortilin. Freshly purified rTG was always dimeric, while the monomeric form was achieved by heating at most 2 µM TG at 60 °C for 120 s. TG oligomeric states were verified by SEC and MP. All the peptides used in this study were purchased from Genscript.
In vitro iodination
Enzymatic iodination of rTG was performed as previously described by Coscia and coauthors3. Briefly, TG at a concentration of 0.1 μM was combined with 1 mM KI, 24 mM glucose, 6 μg/ml glucose oxidase and 3 μg/ml lactoperoxidase. All commercial reagents were purchased from Sigma Aldrich. The iodination reaction was carried out at 37 °C for 60 min, then iodinated TG (iTG) was purified by SEC using a Superose 6 Increase 3.2/300 column (GE Healthcare).
Complex reconstitution and purification
eTG and his-tagged sortilin were mixed in 1:1 ratio and co-concentrated with a 30 MWCO concentrator (Millipore) in buffer A to reach a final concentration of 3 µM for each protein in a volume of approximately 100 µL. The sample was added to 50 µL of Ni-SepharoseTM High-Performance resin (Cytiva) equilibrated with buffer A and incubated for 2 h at 4 °C under gentle stirring. Pulldown was carried out using manufacturer protocol for spin cup method purification. Briefly, beads were washed by adding 5 × 4 CV of buffer A followed by a second wash of 5 × 4 CV of buffer A supplemented with 20 mM imidazole to remove unbound eTG. The sample was incubated for 5 min with 4 × 1 CV of buffer A supplemented with 500 mM imidazole and eluted by spinning at 1000 × g for 1 min. All the fractions are analysed by SDS-PAGE.
To study the stoichiometry of the complex, 5 µM sortilin, 1 µM rTG and their mixtures (5:1 molar ratio) in buffer A were injected onto a Superose 6 increase 3.2/300 column (GE Healthcare) on an ÄKTA micro (Cytiva) at 4 °C. Co-elution was examined using NuPAGE™ 3–8% tris-acetate SDS-PAGE (Thermo Fisher Scientific) and the elution profiles were compared using Prism 9 (GraphPad).
To stabilise the complex with crosslinkers, the mTG fraction was isolated through SEC and then incubated with Sortilin in a 1:1 molar ratio at a final concentration of 1.2 µM for each protein. Sortilin, mTG (both at 1.2 µM) and the complex SorTG were incubated with 1.5 mM BS3 for 30 min at room temperature, before quenching the reaction with ammonium bicarbonate (ABC, Sigma, 50 mM final concentration). Crosslinked and non-crosslinked samples were analysed by NuPAGE™ 3–8%, tris-acetate SDS-PAGE (Thermo Fisher Scientific) and transferred to nitrocellulose membrane using the trans-blot turbo mini nitrocellulose transfer kit (Bio-Rad). The membrane was blocked with 5% milk in TBST (TBS + 0.1% tween-20) at room temperature for 45 min, followed by overnight incubation at 4 °C with primary antibodies: anti-sortilin antibody (clone F11, from mouse Sigma-Aldrich diluted 1:250 in 5% TBST milk) and anti-thyroglobulin antibody (ab156008 from rabbit, Abcam diluted 1:10,000 in 5% TBST milk). After washing in TBST, membranes were incubated with secondary antibody for 1 h at room temperature: IRDye 800CW goat anti-rabbit IgG (Abcam) and IRDye 680RD goat anti-mouse IgG (Abcam), both diluted 1:10,000 in 5% TBST milk. Finally, the membrane was washed with TBST and visualised using the Odyssey CLx infra-red imaging system.
To isolate the stabilised complex, we used GraFix27. Sortilin and mTG were mixed at 2:1 molar ratio (2–1 µM) and incubated at 4 °C for at least 2 h. A continuous gradient containing 20 mM HEPES (pH 7.5), 150 mM NaCl, 10–30% glycerol, 0–0.1% glutaraldehyde was generated using a Gradient Master 108 (BioComp) in 13.2 ml thinwall ultra-clear ultra centrifuge tubes (Beckman, 344059). Samples were placed in a SW41 Ti ultracentrifuge rotor and centrifuged at 16,100 × g for 17 h at 4 °C using an Optima-XPN 100H. Samples were manually fractionated and the crosslinking was terminated by adding tris-HCl (pH 7.5) to a final concentration of 65 mM. GraFix fractions were analysed by NuPAGE™ 3–8%, tris-acetate SDS-PAGE (Thermo Fisher Scientific) and transferred to nitrocellulose membrane using the trans-blot turbo mini nitrocellulose transfer kit (Bio-Rad). Western blot anti-sortilin and anti-TG was performed as described above.
Mass photometry analysis
MP experiments were performed using a Refeyn OneMP mass photometer (Refeyn Ltd). Data was acquired using AcquireMP (Refeyn Ltd, version 2024 R1) and analysed with DiscoverMP software (Refeyn Ltd, version 2024 R1). Sample carrier slides (Refeyn) were assembled with CultureWell™ gasket (GBL103250, Sigma-Aldrich). The system was first autofocused to 15 µL of freshly prepared and filtered buffer A before 2 µL of protein sample was quickly mixed in reaching a final concentration of around 20 nM for single molecule measurements. Sixty-second movies were acquired with medium acquisition area field. Raw data were analysed with DiscoverMP software and masses were assigned to binding events by using a calibration curve generated with NativeMark standard (LC0725, Thermo Scientific) diluted in acquisition buffer.
For SorTG measurements, 6 µM sortilin was pre-incubated with 2 µM TG, mTG or iTG for 3 h at 4 °C. After finding the focus on 15 µL of buffer A in the gasket-well complex sample was quickly diluted 1:20 in buffer A, then 2 µL of diluted sample were added to the well for the measurement. For competition assays, 300 nM sortilin and 100 nM mTG were incubated for 3 h at 4 °C. The complex was then perturbated with the addition of NT (MedChemExpress), AF38469 (AF) (Medchemexpress; HY-12802) or TGpep to a final concentration in solution of 30 µM for NT and AF38469 and 1 mM for TGpep (a titration of peptide 30–100 µM–1 mM was also performed) (Supplementary Fig. 9b). Compounds were incubated 45 min at 4 °C and then 2 µL of the protein sample were used for measurement in Refeyn, to a final concentration of ~10 and 30 nM in the drop. For competition assays, all measurements were performed three times independently.
For the panel in Fig. 1h, the percentage of mTG was calculated based on the normalisation of mTG counts with respect to the sum of mTG and dTG counts in the peaks detected by Refeyn Discovery software. The percentage of complex was determined as the counts of SorTG normalised on the total amount of counts belonging to mTG, dTG and SorTG peaks as they are detected by Refeyn Discovery Software. In panel Fig. 3b, since the peaks detected by Refeyn software for mTG and SorTG partially overlap, the amount of complex was calculated as the percentual increase of the counts belonging to SorTG+mTG compared to mTG:
The background noise was evaluated by manual integration of the counts detected between the tail of the mTG peak (400 kDa) and 500 kDa and calculated in the same way:
The effect of sortilin interactors (AF, NT, or TGpep) on SorTG formation was established as the percentual reduction of complex compared to the non-inhibited control. All measurements were done in triplicate and for each independent experimental condition, all the data were normalised on the average of the value found for SorTG.
Miniaturised differential scanning fluorimetry (NanoDSF)
Thermal unfolding profiles of recombinant TG (rTG) and endogenous TG (eTG) were monitored using a Prometheus NT.48 instrument (NanoTemper Technologies). Protein samples purified and diluted in their respective storage buffers to a final concentration of 0.5 mg/mL were loaded into standard-grade capillaries (NanoTemper Technologies). Samples were then subjected to a controlled temperature increase from 20 to 95 °C at a rate of 1 °C per min. The melting temperature (Tm, °C) was calculated from the inflection point of the fluorescence intensity ratio (F350/F330), identified as the maximum of the first derivative curve.
To test the interactions between sortilin and the TGpep, the Tm shift was assessed by incubating 0.5 mg/ml (6 µM) with a 100-fold molar excess of TG peptide at 4 °C for 30 min prior to analysis.
Dose-dependent binding of the TG peptide to sortilin was determined by performing a titration series, using peptide concentrations ranging from 4.7 to 2400 µM against a constant sortilin concentration (6 µM). The Tm was calculated from the inflection point of the fluorescence intensity at both F330 nm and F350 nm, identified as the maximum of the first derivative curve. Tm of sortilin was plotted as a function of peptide concentration using GraphPad Prism. Each titration point was tested in triplicate and the resulting data were analyzed using the PR.ThermControl software (NanoTemper Technologies).
Cross-linking mass spectrometry
The crosslinker sulfo-SDA (sulfosuccinimidyl 4,4′-azipentanoate, ThermoFisher Scientific) was dissolved in water just before use, to a final concentration of 20 mM. For the labelling reaction, 109 µg of extracted thyroglobulin (eTG) and 31 µg of sortilin were pre-incubated for 2 h, yielding a final solution containing 1 µM TG and 2 µM sortilin. Sulfo-SDA was added in different sample:crosslinker weight ratios (1:0.13, 1:0.38, 1:0.5 - corresponding to 300, 880 and 1200 µM sulfo-SDA with constant complex concentration). After incubating the solution for 30 min at RT in the dark, the reaction of photo-crosslinking was carried under a UVP CL-1000 (Analytic Jena) at 365 nm on ice for 10 min. The sample was placed at ~5 cm from the UV lamp. The reaction was quenched, adding ammonium bicarbonate (ABC, Sigma) at a final 50 mM concentration and the crosslinked complex was deglycosylated by incubation at 37 °C for 1 h with PNGase F (NEB, following provider protocol for reaction under denaturing condition). The complex was separated on a NuPAGE™ 3–8%, Tris-Acetate SDS-PAGE (Thermo Fisher Scientific) and the band corresponding to the crosslinked complex excised. The final sample:crosslinker ratios used were 1:0.38 and 1:0.5 weight:weight69. Samples were then processed by in-gel digestion, with reduction with dithiotreitol (DTT) and alkylation with iodoacetamide (IAA) followed by digestion with trypsin (Thermo Fisher Scientific Pierce). The resulting peptides were extracted, desalted and pooled using C18 StageTips.
For crosslinking MS acquisition 1 μg of peptides were injected for each liquid chromatography-mass spectrometry (LC-MS) acquisition. The LC-MS platform consisted of a Vanquish Neo system (ThermoFisher Scientific) connected to an Orbitrap Eclipse Tribrid mass spectrometer (ThermoFisher Scientific). Mobile phases consisted of 0.1% v/v formic acid in water (mobile phase A, Fisher Scientific) and 0.1% formic acid in 80% acetonitrile/water v/v (mobile phase B, Fisher Scientific). Samples were dissolved into 4% mobile phase B. The samples were separated on an EASY-Spray PepMap Neo column (75 μm × 50 cm) (ThermoFisher Scientific) using a 120 min method from 2% B to 35% B (77 min), 35% B to 45% B (5 min), then 45% B to 95% B (22 min) or a 150 min method from 2% B to 35% B (95 min), 35% B to 50% B (5 min), 50% B to 55% B (1 min) and 55% B to 95% B (11 min) prior to column equilibration. MS1 spectra were acquired with a resolution of 120,000 and automated gain control target set to standard and 50 ms maximum injection time. Source RF lens was set to 35%. Dynamic exclusion was set to single count for 30 or 10 s and a precursor charge filter was set to z = 3–7 (two independent acquisitions). Additionally a charge filter set to z = 4–7 was used for a third acquisition. Precursors were selected based on a data-dependent decision tree strategy70 prioritising charge states 4–7 and subjected to stepped HCD fragmentation with normalised collision energies of 20,27,30. MS2 scans were acquired with a normalised gain control target of 750% for sulfo-SDA. Maximum injection time was set to 250 ms and orbitrap resolution to 60,000. Fluoranthene was used for lock-mass correction of MS runs.
For Crosslinking MS data analysis raw files were converted to mgf format using ProteoWizard MSconvert (version 3.0.24184)71. Crosslinking MS database search was performed in xiSEARCH (version 1.8.5)72 on a database comprising TG, sortilin and common mass spec contaminants derived from MaxQuant73 searched with 4 missed cleavages. MS1 precursor mass error tolerance was set to 3 ppm and MS2 error tolerance to 5 ppm. Carbamidomethylation of cysteine was defined as a fixed modification. The search included methionine oxidation, SDA-loop link (+82.04186484 Da) and SDA-OH (+100.0524 Da) as variable modifications. The SDA crosslinker was defined as cleavable74 and reactive from K or protein N-termini to any amino acid. The search was set to account for noncovalent gas-phase associations. Results were filtered to 5% FDR at the residue pair level using xiFDR and the ‘boost’ feature was enabled.
AlphaFold predictions
The structures of the complex between mature sortilin (residues 78–766) and either mTG, TG ChEL domain (TG residues 2185–2768) or TG peptide (TG residues 2749–2768) were predicted in the AlphaFold Server powered by AlphaFold328. The PAE plots were rendered with ChimeraX75.
The flowchart describing the generation of protein-peptide complexes using AlphaPulldown and the selection criteria is provided in Supplementary Fig. 11. For both sortilin full-length and sortilin ectodomain screens, we retrieved an initial list of 55 sortilin ligands reported in the literature18. The full-length protein sequences and their post-translational modification (PTM) derivative fragments were used to generate a FASTA file containing 142 candidates. These were then predicted against two baits (sortilin full-length and mature sortilin residues 78–755) using AlphaPulldown 1.0.433 running AlphaFold 2.3.1. AlphaPulldown was run with cycles = 3, models = 5 and predictions per model = 1.
The models were evaluated based on their inter-chain PAE scores (AlphaPulldown PAE cutoff = 8)32, resulting in a shortlist of 82 high-confidence hits. Candidates with identical predictions for ectodomain or full-length baits were selected and the one with the lower iptm_ptm score was discarded. The remaining structures were visually inspected for binding in the canonical sortilin binding pocket. If steric clashes were detected, the interaction was re-assessed using the AlphaFold3 web server. Physiological relevance of the interaction was inferred by checking the topology of the predicted interacting region of the candidates. Predictions of interactions between the sortilin ECD and intracellular regions of target proteins were discarded. For the 10 selected hits, the residues interacting with the sortilin canonical binding site were identified based on visual inspection in UCSF ChimeraX75 and confirmed based on the PAE plot.
The identified sequences were analysed taking into account their directionality and mode of interaction with sortilin R292. If the interaction occurred via the acidic moiety of the protein C-terminus, the sequence was supplemented with a final aspartate to facilitate sequence alignment. If the interaction proceeded from C-terminus to N-terminus the sequence was reversed for downstream sequence alignments.
To search for putative common binding motifs in sortilin ligands the 10 sequences were aligned using Clustal Omega76 and the alignment was manually adjusted based on structural superposition. The aligned sequences were then analysed using PSSMSearch server35 to identify common features. The most frequently represented residues at each position were used to infer a putative consensus peptide, which was subsequently analyzed in SOPMA77 to predict secondary structure propensity. The plots in Supplementary Fig. 4h were generated using PAE viewer78.
Electron microscopy sample preparation and data collection
For negative stain (NS) EM, the SorTG complex was diluted to final concentrations of ~0.01 mg/mL. A 4 μL sample was applied to glow-discharged (30 mA, 45 s; Pelco EasyGlow) carbon-coated 400 mesh grids (Ted Pella #120821), incubated for 1 minute, and blotted. Grids were stained with a 2% aqueous uranyl acetate solution for 45 seconds, followed by side blotting for 2–3 seconds and air drying. Imaging was performed on a Talos L120C microscope (Thermo Fisher) equipped with a Ceta 16M camera, operating at 120 kV and 45,000x nominal magnification (3.18 Å pixel size), with defocus values ranging from –1.5 to –2.5 µm. The collected data were processed in RELION. For cryo-electron microscopy (cryo-EM), following GraFix, fractions enriched for SorTG complex were combined and buffer exchanged to buffer A to remove glycerol and concentrated to approximately 0.06 mg/ml. To prepare sortilin and sortilin-peptide samples for cryo-EM, the proteins were concentrated to 2.5 and 3.5 mg/ml, respectively and supplemented with 0.5% OG. An excess of TG peptide was incubated with the latter at a final concentration of 1 mM for 3 h at 4 °C. Holey carbon Quantifoil 1.2/1.3 300-mesh grids were glow discharged at 30 mA for 60 s and used as supports for sortilin and sort-peptide samples, while for SorTG a graphene oxide layer was deposited on the grids according to a published procedure79.
3.5 µL of each sample was applied to the grids with different incubation time (30 s for SorTG and 20 s for sortilin and SorTGpep) and plunge-frozen in liquid ethane using a Vitrobot Mark IV (Thermo Fisher) at 4 °C under 100% humidity conditions. Images were acquired on a Falcon 4i camera using a Glacios (Thermo Fisher) electron microscope at 200 kV for SorTG and on a Titan Krios (Thermo Fisher −300 kV) for sortilin and SortTGpep. A SelectrisX energy filter (Thermo Fisher) with a slit width of 20 eV was used to remove inelastically scattered electrons. For SorTG, 7466 movies were acquired at a pixel size of 0.91 Å with a nominal dose of ~50 e/Ų distributed over 44 frames. For sortilin and SorTGpep, respectively 15781 and 14574 movies were aquired at a pixel size of 0.748 Å with a nominal dose of ~50 e/Ų distributed over 25 frames. The target defocus range for all samples was from −1 to −2 µm. All datasets were collected at Human Technopole in Milan using microscopes operated by EPU software v3.7.1.6940.
Electron microscopy data processing
All cryo-EM data processing was performed using the software RELION (v.5.0)80 and cryoSPARC (v.4.5.1). Detailed process is schematized in Supplementary Fig. 10. For SorTG dataset motion correction was performed using Patch MotionCorrection and the CTF parameters were estimated using PatchCTF estimation to remove the micrographs with poor statistics. 1304504 particles were picked with Blob Picking and after two rounds of 2D classification, a final batch of 131283 particles were used to generate 4 ab-initio models. 30545 particles belonging to the best class were subjected to 3D homogeneous refinement. The signal belonging to TG was subtracted with particle subtraction and after 2D classification, a final subset of 21777 particles was used to generate two ab-initio volumes that were analyzed in ChimeraX 1.6.1 by comparing them with sortilin crystal structure (PDB: 4PO7). For sortilin and SorTGpep datasets motion correction was performed using MotCor2 and the CTF parameters were estimated using CTFFIND-4.181. For sortilin 5185162 particles were picked with a template derived by the crystallography structure of sortilin (PDB: 4PO7) and analysed with two rounds of 2D classification that provided a batch of 983088 particles that were subjected to repetitive cycles of ab-Initio model generation followed by non-homogeneous82 and then local refinement. A final batch of 479743 particles converged to a map of 3.10 A resolution that was used for model building at a contour level of 0.716. For sort-peptide 4379021 particles were picked using a template generated from the final volume obtained from sortilin processing. The same pipeline as sortilin dataset was then followed to process SorTGpep. Since the micrographs were noisier due to the high amount of peptide in solution and the map was suffering from anisotropy, a final batch of 320127 particles was exported in RELION to further process. Several step of 3D Refinement with Blush and 3D Classification led to a final set of 47561 particles that were subjected to CTF refinement and Bayesian polishing, leading to a final map of 3.36 Å resolution that was used for model building at a contour level of 0.00659.
Model building
Model building for the cryo-EM data of ligand-free sortilin and SorTGpep was performed using the software Coot (v.0.9.8.95)36, with the initial models based on either the crystal structure of sortilin (PDB:4PO7) or the AlphaFold3 prediction of the SorTGpep complex, respectively. The starting models were iteratively adjusted and refined using the real-space refinement module in the Phenix package (v.1.21)83, as well as Coot and iSOLDE84. The final models were deposited in the Protein Data Bank under the accession codes PDB:9I0N for ligand-free sortilin and PDB:9I0O for SorTGpep. Residue numbering of sortilin begins after the signal peptide, consistent with other sortilin structures in the PDB, while TG peptide is numbered from 1 to 20 as a synthetic ligand.
Microscale termophoresis
The binding affinity of sortilin for different peptides was determined using MST with a Monolith NT.115 instrument (NanoTemper Technologies, Germany). Protein (1 µM) was labelled with RED-tris-NTA dye (Nanotemper Technologies) according to the manufacturer’s instructions in 20 mM HEPES pH 7.4, 150 mM NaCl, 0.1% Tween-20.
Lyophilised TGpep, TGpep mutated NT and sortilin propeptide (all purchased from GeneScript) were resuspended in buffer A to final working concentrations of 8 mM, 5 mM, 400 nM and 2 mM, respectively. Lyophilised TGpepT4 was resuspended in buffer A supplemented with 25% dimethyl sulfoxide (DMSO) to a stock solution of 5 mM and further diluted with buffer A without DMSO to a working concentration of 500 nM. For each ligand a 16-point dilution series was prepared with working concentrations as starting points. Additional 16-point dilution series of TGpep with 2.5% DMSO in buffer A was prepared as a control.
For each concentration point 10 µl of 400 nM labelled protein was mixed with 10 µl of peptide and incubated for 30 min at room temperature. Upon incubation, 10 µl of protein–peptide mixture was loaded into Monolith NT.115 Premium Capillaries (Nanotemper Technologies) and fluorescence intensity was measured at 25 °C using a nano-RED detector with 100% excitation power and 40% MST power. MST data from three independent measurements were analysed and equilibrium dissociation constant (Kd) were calculated using GraphPad Prism. All experiments were performed with the support of Marko Nedeljkovic from the Human Technopole National Facility of Structural Biology, who also performed data analysis in GraphPad Prism.
Internalisation assay
Nthy-ori 3–1 Normal human primary thyroid follicular epithelial cells were a gift from Luca Persani’s laboratory (Istituto Auxologico in Milan, Italy), EACC catalogue no. 90011609. These cells were cultured in RPMI1640 (Euroclone, ECB9006LX10) supplemented with 10% (v/v) heat-inactivated foetal bovine serum (Life Technologies, 10500064), 100 IU ml−1 penicillin-streptomycin (Euroclone, ECB3001D), 2 mM L-glutamine (Euroclone, ECB3004D) at 37 °C and at atmospheric O2 and 5% CO2. Cell lines were repeatedly tested for mycoplasma by PCR. Nthy-ori 3-1 cells were plated in 24-well plates at a density of 50,000 cells per well and left for at least 24 h to adhere. Cells were then lipofected with sortilin-V5 vector using Lipofectamine 2000 (Invitrogen, 11668019). Medium containing transfection reagents was replaced with fresh medium 5 h after transfection. Next day TG was added in the medium for 120 min. The wells were washed three times with PBS. Three types of TG were tested: eTG (extracted TG (eTG) from millipore 609312-1 mg, final concentration of 0.5 or 0.1 mg/ml), mTG (monomeric TG from eTG final concentration of 0.1 mg/ml) and dTG (dimeric TG from eTG final concentration of 0.1 mg/ml). For competition assay we add in the medium 30 uM of AF3846931 or 1 mM of TGpep.
Confocal image acquisition and analysis
After the cellular internalisation assay, the cells were fixed by 4% PFA for 10 min at room temperature. Cells were then washed three times for at least 5 min in PBS before blocking for 1 h at room temperature with PBS supplemented with 0.1% (v/v) triton X-100 and 1% (v/v) donkey serum and later incubating 2 h at room temperature with primary antibodies diluted in staining solution (PBS supplemented with 0.1% (v/v) triton and 1% (v/v) donkey serum). Primary antibodies used: mouse monoclonal anti-sortilin (Millipore MABN1792, ICC 1:100), rabbit monoclonal anti-TG (Abcam, ab15600, ICC 1:100). The cells were then washed with PBS three times for 5 min. The cells were incubated for 30 min at room temperature with Alexa Fluor™ Plus 647 Phalloidin (Thermo Scientific, A30107 ICC 1:400) and Hoechst 33342 (Sigma-Aldrich, B2261) 0.2 μg ml−1 diluted in PBS. The cells were then washed with PBS three times for 5 min and incubated for 1 h at room temperature in secondary antibodies diluted in staining solution. Fluorescence-conjugated secondary antibodies used: donkey anti-rabbit IgG (H + L) Secondary Antibody, Alexa Fluor 488, Invitrogen (Thermo Scientific, A21206 ICC 1:500) and goat anti-mouse IgG (H + L) highly cross-adsorbed secondary antibody, Alexa Fluor Plus 555 (Thermo Scientific, A32727, ICC 1:500). From then on, the plate was protected from light. Cells were washed twice for 5 min with PBS. Coverslips were mounted using DAKO mounting solution (Agilent DAKO, S302380-2). Confocal microscopy analysis was performed using a Zeiss LSM980 point-scanning confocal microscope based on a Zeiss Observer7 inverted microscope. The images were acquired using a PlanApo 40X/1.4NA oil immersion objective with 488 and 561 nm laser lines. Fields of view were randomly selected looking at the sortilin channel. All image acquisition was performed using Zen Blue 3.7 software (Zeiss). The image analysis and quantification were carried out with the support of the Human Technopole Image Analysis Facility using Python pipelines. Each experiment was conducted at least in triplicate. For each replicate, fluorescence intensity was measured in grayscale levels from at least 75 selected cells. An automatic custom Python pipeline was used to analyse the confocal images. Cells and nuclei were segmented with Cellpose pretrained models using phalloidin and DAPI channels. The mean intensity of TG and sortilin signals were calculated for single cells on the whole cell area, excluding the nucleus. Cells not expressing sortilin were excluded from the dataset based on intensity threshold. The threshold was defined by looking at the overall mean intensity distribution.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The cryo-EM data generated in this study have been deposited in the EMDB and PDB database under accession codes EMD-52562 and 9I0N (human sortilin ectodomain); EMD-52563 and 9I0O (human sortilin ectodomain in complex with a TG C-terminal peptide). The mass spectrometry data have been deposited to the ProteomeXchange85 Consortium via the PRIDE partner repository with the dataset identifier PXD06246286. The AlphaFold models are available in ModelArchive (www.modelarchive.org) with the accession codes ma-rurc1, ma-28qa6, ma-daza9, ma-5qqfo, ma-7cor9, ma-nuz4c, ma-8xgtz, ma-76avm, ma-b2lln, ma-mw3nn, ma-f3u4a. All the data generated in this study are provided in the Supplementary Information/Source data file. Source data are provided with this paper.
References
Carvalho, D. P. & Dupuy, C. Thyroid hormone biosynthesis and release. Mol. Cell. Endocrinol. 458, 6–15 (2017).
Mullur, R., Liu, Y.-Y. & Brent, G. A. Thyroid hormone regulation of metabolism. Physiol. Rev. 94, 355–382 (2014).
Coscia, F. et al. The structure of human thyroglobulin. Nature 578, 627–630 (2020).
Stejskalova, C. et al. A conserved acidic residue drives thyroxine synthesis within thyroglobulin and other protein precursors. J. Biol. Chem. 301, 108026 (2025).
Citterio, C. E., Targovnik, H. M. & Arvan, P. The role of thyroglobulin in thyroid hormonogenesis. Nat. Rev. Endocrinol. 15, 323–338 (2019).
Gavaret, J. M., Cahnmann, H. J. & Nunez, J. Thyroid hormone synthesis in thyroglobulin. The mechanism of the coupling reaction. J. Biol. Chem. 256, 9167–9173 (1981).
Luo, Y., Ishido, Y., Hiroi, N., Ishii, N. & Suzuki, K. The emerging roles of thyroglobulin. Adv. Endocrinol. 2014, 189194 (2014).
Di Jeso, B. & Arvan, P. Thyroglobulin from molecular and cellular biology to clinical endocrinology. Endocr. Rev. 37, 2–36 (2016).
Jordans, S. et al. Monitoring compartment-specific substrate cleavage by cathepsins B, K, L, and S at physiological pH and redox conditions. BMC Biochem. 10, 23 (2009).
Dunn, A. D., Crutchfield, H. E. & Dunn, J. T. Thyroglobulin processing by thyroidal proteases. Major sites of cleavage by cathepsins B, D, and L. J. Biol. Chem. 266, 20198–20204 (1991).
Tassinari, M. et al. Molecular mechanism of thyroxine transport by monocarboxylate transporters. Nat. Commun. 16, 4493 (2025).
Friesema, E. C. H. et al. Identification of monocarboxylate transporter 8 as a specific thyroid hormone transporter. J. Biol. Chem. 278, 40128–40135 (2003).
Marinò, M. & McCluskey, R. T. Role of thyroglobulin endocytic pathways in the control of thyroid hormone release. Am. J. Physiol. Cell Physiol. 279, C1295–C1306 (2000).
Botta, R. et al. Sortilin is a putative postendocytic receptor of thyroglobulin. Endocrinology 150, 509–518 (2009).
Petersen, C. M. et al. Molecular identification of a novel candidate sorting receptor purified from human brain by receptor-associated protein affinity chromatography*. J. Biol. Chem. 272, 3599–3605 (1997).
Quistgaard, E. M. et al. Ligands bind to sortilin in the tunnel of a ten-bladed β-propeller domain. Nat. Struct. Mol. Biol. 16, 96–98 (2009).
Leloup, N. et al. Low pH-induced conformational change and dimerization of sortilin triggers endocytosed ligand release. Nat. Commun. 8, 1708 (2017).
Mitok, K. A., Keller, M. P. & Attie, A. D. Sorting through the extensive and confusing roles of sortilin in metabolic disease. J. Lipid Res. 63, 100243 (2022).
Carlo, A.-S., Nykjaer, A. & Willnow, T. E. Sorting receptor sortilin—a culprit in cardiovascular and neurological diseases. J. Mol. Med. 92, 905–911 (2014).
Al-Yozbaki, M., Acha-Sagredo, A., George, A., Liloglou, T. & Wilson, C. M. Balancing neurotrophin pathway and sortilin function: Its role in human disease. Biochim. Biophys. Acta BBA Rev. Cancer 1874, 188429 (2020).
Malik, A. R. & Willnow, T. E. VPS10P domain receptors: sorting out brain health and disease. Trends Neurosci. 43, 870–885 (2020).
Huang, B. et al. Designed endocytosis-inducing proteins degrade targets and amplify signals. Nature 638, 796–804 (2025).
Ahn, G. et al. LYTACs that engage the asialoglycoprotein receptor for targeted protein degradation. Nat. Chem. Biol. 17, 937–946 (2021).
Gustafsen, C. et al. Reshaping the progranulin/sortilin interaction for targeted degradation of extracellular proteins. Preprint at bioRxiv https://doi.org/10.1101/2025.03.03.641244 (2025).
Trabjerg, E. et al. Investigating the conformational response of the sortilin receptor upon binding endogenous peptide- and protein ligands by HDX-MS. Structure 27, 1103–1113.e3 (2019).
Berndorfer, U., Wilms, H. & Herzog, V. Multimerization of thyroglobulin (TG) during extracellular storage: isolation of highly cross-linked TG from human thyroids. J. Clin. Endocrinol. Metab. 81, 1918–1926 (1996).
Stark, H. GraFix: stabilization of fragile macromolecular complexes for single particle cryo-EM. Methods Enzymol. 481, 109–126 (2010).
Abramson, J. et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 630, 493–500 (2024).
Lee, J., Di Jeso, B. & Arvan, P. The cholinesterase-like domain of thyroglobulin functions as an intramolecular chaperone. J. Clin. Investig. 118, 2950–2958 (2008).
Quistgaard, E. M. et al. Revisiting the structure of the Vps10 domain of human sortilin and its interaction with neurotensin. Protein Sci. 23, 1291–1300 (2014).
Schrøder, T. J. et al. The identification of AF38469: an orally bioavailable inhibitor of the VPS10P family sorting receptor sortilin. Bioorg. Med. Chem. Lett. 24, 177–180 (2014).
Varadi, M. et al. AlphaFold protein structure database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 50, D439–D444 (2021).
Yu, D., Chojnowski, G., Rosenthal, M. & Kosinski, J. AlphaPulldown—a Python package for protein–protein interaction screens using AlphaFold-multimer. Bioinformatics 39, btac749 (2023).
Butkinaree, C. et al. Amyloid precursor-like protein 2 and sortilin do not regulate the PCSK9 convertase-mediated low density lipoprotein receptor degradation but interact with each other*. J. Biol. Chem. 290, 18609–18620 (2015).
Krystkowiak, I., Manguy, J. & Davey, N. E. PSSMSearch: a server for modeling, visualization, proteome-wide discovery and annotation of protein motif specificity determinants. Nucleic Acids Res. 46, W235–W241 (2018).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132 (2004).
Mazella, J. Sortilin/neurotensin receptor-3: a new tool to investigate neurotensin signaling and cellular trafficking? Cell Signal. 13, 1–6 (2001).
Nielsen, M. S. et al. The sortilin cytoplasmic tail conveys golgi-endosome transport and binds the VHS domain of the GGA2 sorting protein. EMBO J. 20, 2180–2190 (2001).
Burd, C. G. Physiology and pathology of endosome-to-golgi retrograde sorting. Traffic Cph. Den. 12, 948–955 (2011).
Takamura, A. et al. Sortilin is required for toxic action of Aβ oligomers (AβOs): extracellular AβOs trigger apoptosis, and intraneuronal AβOs impair degradation pathways. Life Sci. 91, 1177–1186 (2012).
Herda, S. et al. The sorting receptor sortilin exhibits a dual function in exocytic trafficking of interferon-γ and granzyme A in T cells. Immunity 37, 854–866 (2012).
Shi, J. & Kandror, K. V. Sortilin is essential and sufficient for the formation of Glut4 storage vesicles in 3T3-L1 adipocytes. Dev. Cell 9, 99–108 (2005).
Chen, Z.-Y. et al. Sortilin controls intracellular sorting of brain-derived neurotrophic factor to the regulated secretory pathway. J. Neurosci. Off. J. Soc. Neurosci. 25, 6156–6166 (2005).
Nilsson, S. K. et al. Endocytosis of apolipoprotein A-V by members of the low density lipoprotein receptor and the Vps10p domain receptor families*. J. Biol. Chem. 283, 25920–25927 (2008).
Canuel, M., Lefrancois, S., Zeng, J. & Morales, C. R. AP-1 and retromer play opposite roles in the trafficking of sortilin between the Golgi apparatus and the lysosomes. Biochem. Biophys. Res. Commun. 366, 724–730 (2008).
Mari, M. et al. SNX1 defines an early endosomal recycling exit for sortilin and mannose 6-phosphate receptors. Traffic 9, 380–393 (2008).
Almoyad, M. A. A., Wahab, S., Mohanto, S. & Khan, N. J. Repurposing drugs to modulate sortilin: structure-guided strategies against atherogenesis, coronary artery disease, and neurological disorders. ACS Omega 9, 18438–18448 (2024).
Al-Akhrass, H. et al. Sortilin limits EGFR signaling by promoting its internalization in lung cancer. Nat. Commun. 8, 1182 (2017).
Marechal, N., Serrano, B. P., Zhang, X. & Weitz, C. J. Formation of thyroid hormone revealed by a cryo-EM structure of native bovine thyroglobulin. Nat. Commun. 13, 2380 (2022).
Kim, K. et al. The structure of natively iodinated bovine thyroglobulin. Acta Crystallogr. Sect. Struct. Biol. 77, 1451–1459 (2021).
Adaixo, R. et al. Cryo-EM structure of native human thyroglobulin. Nat. Commun. 13, 61 (2022).
Andreoli, M., Sena, L., Edelhoch, H. & Salvatore, G. The noncovalent subunit structure of human thyroglobulin. Arch. Biochem. Biophys. 134, 242–248 (1969).
Braverman, L. E. & Cooper, D. S. Werner & Ingbar’s The Thyroid: A Fundamental and Clinical Text (Lippincott Williams & Wilkins, 2012).
Salvatore, G., Sena, L., Viscidi, E. & Salvatore, M. The thyroid iodoproteins 12S, 19S and 27S in various animal species and their physiological significance. Curr. Top. Thyroid Res. 193, 206 (1965).
Schneider, A. B., Bornet, H. & Edelhoch, H. Properties of thyroglobulin. XX. The biosynthesis of thyroglobulin. Effect of temperature on subunit species. J. Biol. Chem. 245, 2673–2678 (1970).
Dunn, J. T. & Ray, S. C. Changes in the structure of thyroglobulin following the administration of thyroid-stimulating hormone. J. Biol. Chem. 250, 5801–5807 (1975).
Yao, Y. et al. Extracellular phase separation mediates storage and release of thyroglobulin in the thyroid follicular lumen. Commun. Biol. 8, 1–9 (2025).
Herzog, V., Berndorfer, U. & Saber, Y. Isolation of insoluble secretory product from bovine thyroid: extracellular storage of thyroglobulin in covalently cross-linked form. J. Cell Biol. 118, 1071–1083 (1992).
Citterio, C. E. et al. Structural features of thyroglobulin linked to protein trafficking. Protein Sci. Publ. Protein Soc. 32, e4784 (2023).
Doğru, A. G., Rehders, M. & Brix, K. Investigations on primary cilia of Nthy-ori 3-1 cells upon cysteine cathepsin inhibition or thyrotropin stimulation. Int. J. Mol. Sci. 24, 9292 (2023).
Lisi, S. et al. Absence of A thyroid phenotype in sortilin-deficient mice. Endocr. Pract. 21, 981–985 (2015).
Marinò, M., Zheng, G. & McCluskey, R. T. Megalin (gp330) is an endocytic receptor for thyroglobulin on cultured Fisher rat thyroid cells. J. Biol. Chem. 274, 12898–12904 (1999).
Montuori, N. et al. The rat asialoglycoprotein receptor binds the amino-terminal domain of thyroglobulin. Biochem. Biophys. Res. Commun. 268, 42–46 (2000).
Consiglio, E. et al. Thyroglobulin interactions with thyroid membranes. Relationship between receptor recognition of N-acetylglucosamine residues and the iodine content of thyroglobulin preparations. J. Biol. Chem. 256, 10592–10599 (1981).
Citterio, C. E., Rivolta, C. M. & Targovnik, H. M. Structure and genetic variants of thyroglobulin: pathophysiological implications. Mol. Cell. Endocrinol. 528, 111227 (2021).
Tosatto, L. & Coscia, F. A glance at post-translational modifications of human thyroglobulin: potential impact on function and pathogenesis. Eur. Thyroid J. 11, e220046 (2022).
Kim, P. S. et al. A single amino acid change in the acetylcholinesterase-like domain of thyroglobulin causes congenital goiter with hypothyroidism in the cog/cog mouse: a model of human endoplasmic reticulum storage diseases. Proc. Natl. Acad. Sci. USA 95, 9909–9913 (1998).
Sparks, R. P. et al. An allosteric binding site on sortilin regulates the trafficking of VLDL, PCSK9, and LDLR in hepatocytes. Biochemistry 59, 4321–4335 (2020).
Banchenko, S. et al. Structural insights into cullin4-RING ubiquitin ligase remodelling by Vpr from simian immunodeficiency viruses. PLOS Pathog. 17, e1009775 (2021).
Kolbowski, L., Mendes, M. L. & Rappsilber, J. Optimizing the parameters governing the fragmentation of cross-linked peptides in a tribrid mass spectrometer. Anal. Chem. 89, 5311–5318 (2017).
Chambers, M. C. et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 30, 918–920 (2012).
Mendes, M. L. et al. An integrated workflow for crosslinking mass spectrometry. Mol. Syst. Biol. 15, e8994 (2019).
Cox, J. & Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 26, 1367–1372 (2008).
Iacobucci, C. et al. Carboxyl-photo-reactive MS-cleavable cross-linkers: unveiling a hidden aspect of diazirine-based reagents. Anal. Chem. 90, 2805–2809 (2018).
Meng, E. C. et al. UCSF ChimeraX: tools for structure building and analysis. Protein Sci. 32, e4792 (2023).
Madeira, F. et al. The EMBL-EBI job dispatcher sequence analysis tools framework in 2024. Nucleic Acids Res. 52, W521–W525 (2024).
Geourjon, C. & Deléage, G. SOPMA: significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Comput. Appl. Biosci. CABIOS 11, 681–684 (1995).
Elfmann, C. & Stülke, J. PAE viewer: a webserver for the interactive visualization of the predicted aligned error for multimer structure predictions and crosslinks. Nucleic Acids Res. 51, W404–W410 (2023).
Bokori-Brown, M. et al. Cryo-EM structure of lysenin pore elucidates membrane insertion by an aerolysin family protein. Nat. Commun. 7, 11293 (2016).
Kimanius, D. et al. Data-driven regularization lowers the size barrier of cryo-EM structure determination. Nat. Methods 21, 1216–1221 (2024).
Rohou, A. & Grigorieff, N. CTFFIND4: fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 192, 216–221 (2015).
Punjani, A., Zhang, H. & Fleet, D. J. Non-uniform refinement: adaptive regularization improves single-particle cryo-EM reconstruction. Nat. Methods 17, 1214–1221 (2020).
Afonine, P. V. et al. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr. Sect. Struct. Biol. 74, 531–544 (2018).
Croll, T. I. ISOLDE: a physically realistic environment for model building into low-resolution electron-density maps. Acta Crystallogr. Sect. Struct. Biol. 74, 519–530 (2018).
Deutsch, E. W. et al. The ProteomeXchange consortium at 10 years: 2023 update. Nucleic Acids Res. 51, D1539–D1548 (2023).
Perez-Riverol, Y. et al. The PRIDE database at 20 years: 2025 update. Nucleic Acids Res. 53, D543–D553 (2025).
Acknowledgements
We thank all members of Human Technopole facilities and in particular: Paolo Swuec, Gaetano D’Urso, Simona Sorrentino of the cryo-EM unit of the National Facility for Structural Biology for technical support and assistance with cryo-EM sample preparation and data collection, we thank Eugenia Cammarota from the bioimage analysis infrastructure unit of the national facility for data handling and analysis, the National Facility for Light Imaging for the assistance with image acquisition, the Biomass production Unit, the National Facility for Structural Biology–IU3 Biophysics Unit. We thank Daniele Colombo from the IT department for pivotal help with software installation and debugging. We thank Prof Luca Persani for donating thyroid cells. This research was supported by the European Research Council (ERC THYROMOL – GA 101041298).
Author information
Authors and Affiliations
Contributions
F.C.: conceptualisation; I.B., C.S.: biochemical and biophysical experiments; G.T.: cell biology assays and image analysis; I.B.: EM data collection and analysis; I.B., F.C.: model building, I.B., A.G.: multi-sequence alignments, AlphaFold and AlphaPulldown analysis; A.G., A.D.I.: mass spectrometry data collection and analysis. M.N.: Optimisation of binding experiments by MST of sortilin to TGpep and other peptides and MST analysis. F.C. and I.B. drafted the manuscript. All authors contributed to edit the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Cintia Citterio, Oliver Clarke, Nils Marechal, Juan Nicola and the other anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Boniardi, I., Tanzi, G., Di Ianni, A. et al. Molecular recognition of thyroglobulin by sortilin. Nat Commun 17, 2004 (2026). https://doi.org/10.1038/s41467-026-68658-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-68658-z
