
Abstract
The psychedelic psilocybin may have lasting therapeutic effects for patients with chronic pain syndromes. Some preclinical data suggest these putative benefits derive from direct analgesic effects; however, this possibility has not been comprehensively tested in preclinical models. Here, we evaluated the analgesic properties of a single exposure to psilocybin at acute and chronic time points in Complete Freund’s Adjuvant-induced inflammatory pain, spared nerve injury model of neuropathic pain, and acid-induced muscle pain. Across these models, we tested a range of doses (0.3, 2, and 10 mg/kg i.p.) in male and female mice using multiple behavioral assays evaluating sensory aspects (von Frey, cold plate, Hargreaves, thermal place preference, and muscle withdrawal threshold) and functional aspects of pain (marble burying). We further tested the effects of psilocybin on the affective dimension of pain in a surgical model of acute pain (mouse grimace scale). Except for cold sensitivity, we found no effect of psilocybin across pain models, behavioral assays, drug doses, or sex. The apparent reduction in cold sensitivity may be explained by profound hypothermia induced by psilocybin rather than true analgesia.
Similar content being viewed by others

Introduction
Psilocybin, the prototypical classic psychedelic, produces hallucinogenic activity in humans through activation of discrete 5-hydroxytryptamine receptor (5-HTR) subtypes. A resurgence of interest in the therapeutic potential of psychedelics, particularly psilocybin, has led to testing psilocybin-assisted therapy for a range of neuropsychiatric conditions, most notably depression and substance use disorders1,2,3,4. Early studies across clinical indications have shown large, rapid, and durable therapeutic effects after a single hallucinogenic dose of psilocybin, leading to broad optimism that psilocybin may have a role in other high-impact health conditions, such as chronic pain5.
Clinical studies performed in the 1960s and 1970s suggested that a hallucinogenic dose of a similar psychedelic, lysergic acid diethylamide (LSD), could reduce pain either by a direct analgesic effect or by globally improving emotional health5,6,7. Exploratory analyses from modern psilocybin trials performed in advanced-stage cancer patients with a variety of pain etiologies have shown improvements in several psychiatric conditions associated with chronic pain, such as anxiety and depression8,9,10,11, but have noted a heterogeneous analgesic effect of psilocybin10. Recent survey studies and pilot clinical trial data suggest that patients with various chronic pain syndromes can effectively manage pain symptoms with psychedelics over a range of doses11,12,13,14, even suggesting that psychedelics (i.e., LSD) may have a direct analgesic effect15, although changes in pain beliefs and pain acceptance are also likely to contribute. Modern randomized, placebo-controlled trials for various pain syndromes have yet to be completed.
Recently, a few studies in laboratory animals have suggested that single doses of psilocybin and other psychedelic compounds have potent and lasting analgesic effects in some models of chronic pain16,17,18,19,20. However, it is unclear whether these early results generalize across models of chronic pain that are widely used in the field, or whether these findings are dose- or model-specific. Accordingly, our aim was not to replicate any single prior protocol, but to test whether such effects generalize across standard mouse pain models and reasonable protocol variation.
To address this knowledge gap, we performed in vivo experiments to evaluate the antinociceptive (pain-relieving; analgesic) activity of psilocybin using a range of doses, in a range of experimental pain models in mice, and in each model performed multiple behavioral tests that assayed different sensory, functional, and affective aspects of pain. We chose a range of well-studied doses of psilocybin (0.3, 2, and 10 mg/kg) that have clear central nervous system activity21,22,23,24,25,26. Further, we tested each of these models, behavioral assays, and doses at acute and chronic time points in both male and female mice. Surprisingly, psilocybin had no analgesic properties in any of these tests, except for cold sensitivity in a neuropathic pain model. However, this result was fully accounted for by psilocybin-induced body temperature dysregulation.
Results
Psilocybin has dose-dependent effects on head twitch and rearing behavior
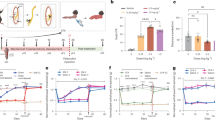
We confirmed that psilocybin was physiologically active over the dose range (0.3, 2, and 10 mg/kg) by observing a dose-dependent increase in head twitch response (HTR), a proxy for 5-HT2AR activity24,27 (Fig. 1A), and a decrease in rearing behaviors, a proxy for 5-HT1R activity28,29 (Fig. 1B). In both sexes, we observed dose dependent effects of psilocybin (Fig. S1A-D); however, these studies were not adequately powered to detect a meaningful sex differences.

A Psilocybin (PSI) at 0.3, 2, and 10 mg/kg is physiologically active as shown by quantification of Head Twitch Response (saline N = 10 [7 M/3 F]; 0.3 mg/kg N = 6 [3 M/3 F]; 2 mg/kg N = 10 [7 M/3 F]; 10 mg/kg N = 6 [3 M/3 F]). One-way ANOVA of total head twitches at the final time point. F(3,28) = 42.98, P < 0.000001. Post-hoc Tukey Test, *P < 0.05 versus saline. Mean ± SEM. B PSI reduces rearing bouts in a dose-dependent manner (saline N = 10 [7 M/3 F]; PSI 0.3 mg/kg N = 6 [3 M/3 F]; PSI 2 mg/kg N = 10 [7 M/3 F]; PSI 10 mg/kg N = 6 [3 M/3 F]). One-way ANOVA of total rearing bouts at the final time point. F(3,28) = 15.96, P = 0.000003. *Post-hoc Tukey Test, P < 0.05 versus saline. Mean ± SEM. PSI 2 mg/kg did not improve mechanical (C) or thermal hypersensitivity (D), whereas morphine (MOR 10 mg/kg) produced significant antinociception in both (N = 5/group, all M). Two-way repeated measures ANOVA (Mechanical: F(2.904, 23.2) = 103.0, P < 0.000001; Thermal: F(2.207,39.72) = 89.85, P < 0.00001). Post-hoc Bonferroni test, * P < 0.05 across pre-MOR vs 30 m post-MOR for mechanical and thermal tests. Mean ± SEM. E PSI 2 mg/kg reduces marble burying performance in CFA-treated mice vs control (CON) and CFA + buprenorphine (BUP 1 mg/kg). (CON N = 14 [7 M/7F]; CFA N = 14 [7 M/7 F]; CFA + BUP N = 16 [9 M/ 7F]; CFA + PSI 2 mg/kg N = 21 [10 M/11 F]). One-way ANOVA of buried marbles, F(5,90) = 83.07, P < 0.000001. Post-hoc Tukey Test, *P < 0.05 versus control; # P < 0.05 versus CFA and control. Mean ± SEM. F BUP 1 mg/kg, but not PSI 2 mg/kg, reduces mouse grimace scale score after laparotomy (SAL N = 12 [6 M/6 F]; BUP 1 mg/kg N = 11 [7 M/4 F]; PSI 2 mg/kg N = 14 [7 M/7 F]). One-way ANOVA of mouse grimace scale difference score (post-laparotomy minus baseline mouse grimace scale values), F(2,33) = 4.535, P = 0.0182. Post-hoc Tukey test, *P < 0.05 versus SAL and PSI 2 mg/kg. Mean ± SEM. Source data are provided as a Source Data file.
Psilocybin has no acute analgesic properties in reflexive, functional, and affective measures of pain in mice
In our initial survey of psilocybin’s sensory and analgesic properties, we tested a dose of psilocybin (2 mg/kg) that has broad effects on acute mouse behavior and immediate early gene expression21,22,25,30. At this dose, psilocybin had no effect on the cutaneous mechanical sensitivity of naive mice (Fig. S1E). We then compared the analgesic effects of psilocybin against mu-opioid receptor agonists (morphine 10 mg/kg or buprenorphine 1 mg/kg) in inflammatory and surgical pain models. In the inflammatory model, CFA was injected into the paw, resulting in mechanical and thermal hyperalgesia as well as decreased function on a marble burying task (Fig. 1C–E). Psilocybin had no effect on mechanical or thermal hyperalgesia (Fig. 1C, D) and further impaired performance on the marble burying task beyond the effect of CFA alone (Fig. 1E). The reduced marble burying effect of psilocybin returned to baseline by 24 hours (Fig. S1F). Consistent with their well-established analgesic properties, morphine significantly reduced mechanical and thermal hyperalgesia, while buprenorphine significantly increased marble burying (Fig. 1C–E). Similarly, psilocybin had no effect on a non-reflexive (affective) measure of pain, the mouse grimace scale31, measured acutely after a laparotomy. As expected, buprenorphine significantly reduced pain-related facial expressions in this assay32 (Fig. 1F). No sex differences were observed in the marble burying task after CFA injection or mouse grimace score after laparotomy (Fig. S1F, G).
Psilocybin has no immediate or prolonged effects in acute or chronic inflammatory, neuropathic, or centrally-mediated pain models
We next expanded our assessment of psilocybin’s analgesic properties across multiple pain models to include a 3-point dose-response curve at multiple time points (1, 4, and 24 hours after drug injection) during both acute and chronic pain phases (Fig. 2A). We tested multiple sensory modalities in each pain model. As expected, after SNI, mice demonstrated a significant decrease in withdrawal latency when exposed to radiant heat as well as a significant decrease in mechanical threshold, indicating the development of robust thermal (Fig. 2A) and mechanical (Fig. 2B) hypersensitivity at day 7 after injury. None of the psilocybin doses administered altered the established thermal or mechanical hypersensitivity at any of the acute time points tested. Two weeks later, a second dose of psilocybin was given, and again, no effect on thermal or mechanical hypersensitivity was observed. We did not observe any sex differences at any time point or dose for SNI (Fig. S2A–D). Together, these findings suggest that psilocybin does not have analgesic properties in a mouse neuropathic pain model.

A Experimental timeline, pain models, and behavioral assessments. Syringes indicate time of injection. All pain models were compared with a concurrent control (CON) group. B, C SNI (N = 12 [6 M/6 F] per group). Mechanical hyperalgesia showed significant effects of dose (F[4,450] = 553.42, P < 0.000001) and time point (F[8,450] = 121.31, P < 0.00001), but not sex (F[1,450] = 0.00, P = 0.960), with a significant dose × time point interaction (F[32,450] = 8.87, P < 0.000001). Thermal sensitivity showed significant effects of dose (F[4,450] = 382.52, P < 0.000001) and time point (F[8,450] = 95.65, P < 0.000001), but not sex (F[1,450] = 2.77, P = 0.097), with a significant dose × time point interaction (F[32,450] = 4.86, P < 0.000001). Three-way ANOVA. D, E AIMP (CON N = 14 [7 M/7 F]; AIMP + PSI 0.3 mg/kg N = 12 [6 M/6 F]; AIMP + PSI 2 mg/kg N = 12 [6 M/6 F]; AIMP + PSI 10 mg/kg N = 10 [5 M/5 F]). Mechanical muscle hyperalgesia showed significant effects of dose (F[4,450] = 494.16, P < 0.000001), time point (F[8,450] = 109.05, P < 0.000001), and sex (F[1,450] = 8.86, P = 0.003), with a significant dose × time point interaction (F[32,450] = 9.77, P < 0.000001). Cutaneous thermal hyperalgesia showed significant effects of dose (F[4,450] = 124.46, P < 0.001) and time point (F[8,450] = 23.37, P < 0.001), with significant dose × time point (F[32,450] = 2.54, P = 0.000013) and dose × sex interactions (F[4,450] = 3.56, P = 0.007). Three-way ANOVA. F, G CFA (N = 8 [4 M/4 F] per group). Thermal hyperalgesia showed significant effects of dose (F[4,270] = 152.68, P < 0.000001) and time point (F[8,270] = 25.16, P < 0.000001), but not sex (F[1,270] = 2.35, P = 0.126). Mechanical hyperalgesia showed significant effects of dose (F[4,270] = 274.38, P < 0.000001) and time point (F[8,270] = 66.33, P < 0.000001). Significant dose × time point interactions were observed for thermal (F[32,270] = 3.67, P < 0.000001) and mechanical (F[32,270] = 4.25, P < 0.000001) sensitivity. Three-way ANOVA. Post hoc Tukey tests: *P < 0.05 versus CON for dose × time point interactions. Data are mean ± SEM. Source data are provided as a Source Data file.
We next tested psilocybin on the acid-induced muscle pain (AIMP) mouse model of widespread musculoskeletal pain, which is used to mimic the widespread pain associated with fibromyalgia33. This model produced cutaneous thermal (Fig. 2C), muscle pressure (Fig. 2D), and cutaneous mechanical sensitivity (Fig. S2E). None of the psilocybin doses administered improved cutaneous thermal, muscle pressure, or cutaneous mechanical hypersensitivity in the AIMP model in the acute phase (Fig. 2C, D; Fig S2E. Two weeks later, a second dose of psilocybin was given; however, no effect on any of these behavioral measures was observed at any of the time points tested (Fig. 2C, D; Fig S2E). We did not observe any sex differences at any time point or dose for AIMP in tests of mechanical sensitivity of skin or muscle (Fig. S2F-I). We did observe a reduction in thermal sensitivity at a single dose and time point in females (0.3 mg/kg, 4 hours after psilocybin injection at the chronic time point, Fig S1K), otherwise psilocybin had no effect on thermal sensitivity. Together, these findings suggest that psilocybin does not have analgesic properties in this model of widespread musculoskeletal pain.
We tested a third model, CFA-induced inflammatory pain. CFA injected under the plantar surface of the hind paw produced thermal (Fig. 2E) and mechanical (Fig. 2F) hyperalgesia of the paw, as evidenced by decreased response latency and reduced paw withdrawal thresholds, respectively. In the acute phase, no dose of psilocybin had an effect on either thermal or mechanical hyperalgesia at any of the time points tested (Fig. 2E, F). Two weeks later, a second dose of psilocybin was administered and, again, we observed no effect at any of the time points tested (Fig. 2E, F). We did not observe any sex differences at any time point or dose for the CFA pain model (Fig. S2L–O). These findings suggest that psilocybin does not have analgesic properties in this mouse model of inflammatory pain.
Psilocybin drives thermal place preference and cold-insensitivity through hypothermia
We next tested the effect of psilocybin on hot and cold temperature aversion by performing thermal place preference testing in CFA-injected mice. Control mice had no preference for either the 30 °C or 40 °C surface while CFA-injected mice avoided the 40 °C surface (Fig. 3A). Morphine (10 mg/kg, i.p.) abolished this avoidance, while psilocybin-injected mice dose-dependently preferred the 40 °C surface (Fig. 3A). This preference was observed even in the absence of CFA injection (Fig. 3B). Further, psilocybin produced a dose-dependent avoidance of a 20 °C surface (Fig. 3C), suggesting that the psilocybin does not reduce temperature sensitivity, but rather induces a place preference for warmer temperatures. This effect appears stronger in female mice at the 2 mg/kg and 10 mg/kg doses. Similarly, acute psilocybin dose-dependently decreased acute formalin-induced cold sensitivity on the cold-plate test (Fig. 3D). Psilocybin had no significant effect on cold sensitivity in our acute neuropathic pain model (SNI), either at 1 h post-drug administration (Fig. 3E), later post-acute timepoints (Fig. 3F, G), or at any time point after injection in the chronic phase of SNI (Fig. 3H–J).

A–C Thermal Place Preference. Mice are given free access to 30 °C and either 40 C or 20 C surfaces. A CFA-treated, but not CON mice, avoid the 40 °C plate. MOR 10 mg/kg blocks the CFA effect. PSI dose-dependently produces a preference for the 40 °C plate in CFA mice. (CON N = 9 [5 M/4 F]; CFA N = 13 [7 M/6 F]; CFA + MOR N = 9 [5 M/4 F]; PSI 0.3 mg/kg, PSI 2 mg/kg, and PSI 10 mg/kg N = 8 [4 M/4 F]). One-way ANOVA. F(5,49) = 9.147, P = 0.000003. Post-hoc Tukey Test, * P < 0.05 versus CON. Mean ± SEM. B Without CFA treatment, PSI also dose-dependently produces preference for the 40 °C plate. (CON N = 6 [3 M/3 F]; PSI 0.3 mg/kg, PSI 2 mg/kg, and PSI 10 mg/kg N = 8 [4 M/4 F]). One-way ANOVA F(3,26) = 10.34, P = 0.000117. Post-hoc Tukey Test, * P < 0.001 versus CON. Mean ± SEM. C Without CFA treatment, PSI dose-dependently produces aversion for the 20 °C plate. (CON N = 8 [4 M/4 F]; PSI 0.3 mg/kg N = 10 [5 M/5 F]; PSI 2 mg/kg N = 11 [5 M/6 F]; PSI 10 mg/kg N = 8 [4 M/4 F]). One-way ANOVA. F(3,34) = 7.211, P = 0.0007. Post-hoc Tukey Test, *P < 0.001 versus CON. Mean ± SEM. D Formalin treatment + cold plate test (CON, FORM, FORM + PSI 0.3 mg/kg, FORM + PSI 10 mg/kg N = 12 [6 M/6 F]; FORM + PSI 2 mg/kg N = 11 [6 M/5 F] One-way ANOVA. F(5,53) = 8.311, P < 0.000001; Post-hoc Tukey Test *P < 0.05, versus SNI). Mean ± SEM. E–J SNI + cold plate test (N = 12 [6 M/6 F] per group) at acute (7 days after SNI) and chronic phases (21 days after SNI). Acute phase: No dose of PSI had an effect at E 1 hour, F 4 hours, or G 24 hours after injection (One-way ANOVA, 1 hour: F[4,55] = 4.840, P = 0.002, 4 hours: F[4, 55] = 5.571, P = 0.0008, 24 hours F[4,55] = 3.520, P = 0.0125; Post-hoc Tukey Test * P < 0.05, versus SNI). Chronic phase: No dose of PSI had an effect at H 1 hour, I 4 hours, or J 24 hours after injection (One-way ANOVA, 1 hour: F[4,55] = 5.370, P = 0.0010, 4 hours: F[4,55] = 5.571, P = 0.0008, 24 hours: F[4,55] = 6.190, P = 0.0003; Post-hoc Tukey Test *, P < 0.05 versus SNI). Source data are provided as a Source Data file. Mean ± SEM.
Using a thermal camera, we measured the body temperature of mice after injection with either saline or psilocybin and found a dose-dependent decrease in body temperature that emerged at 17 minutes and resolved by 70 minutes, with a nadir around 30 minutes (Fig. 4A–D), supporting the hypothesis that observed acute thermal analgesic effects of high-dose psilocybin are best explained by central hypothermia. These studies were not adequately powered to detect meaningful sex differences.

A PSI produces a dose-dependent decrease in body temperature (N = 4 [2 M/2 F] per group). Two-way repeated measures ANOVA showed main effects for Time × Treatment (F[330,1320] = 2.692, P < 0.000001), Time (F[110,1320] = 12.77, P < 0.000001), and Treatment (F[3,12] = 3.524, P = 0.0488). Thermal camera image of female mouse immediately after B, and 30 minutes after C PSI 10 mg/kg injection. Scale bar = 3 cm. D 30 minutes after injection, body temperature is significantly lower in PSI 10 mg/kg as compared to CON. One-way ANOVA (F[3,12] = 6.651, P = 0.0068). Post-hoc Tukey Test, * P < 0.05, different from control. Source data are provided as a Source Data file.
Discussion
The recent interest in psychedelics for the treatment of various psychiatric conditions has prompted speculation that it may also be efficacious for the treatment of chronic pain. Our experiments explore the latter possibility using multiple pain models, both sexes, and testing both reflexive and non-reflexive outcomes. Herein, we reproduce the finding that psilocybin is physiologically active, in a dose-dependent manner, from 0.3 to 10 mg/kg in HTR and rearing assays; however, we found that none of these doses produced measurable analgesia in any of the acute or chronic pain models tested. These results limit the generalizability of reported psilocybin analgesia under common mouse pain assay implementations, without invalidating any single optimized protocol in the growing preclinical literature. Accordingly, a recent effort to systematically evaluate psilocybin’s effects on mouse behavior across three institutions found several acute drug effects but suggested that psilocybin’s persistent effects in mice may be more modest and inconsistent than previously suggested30. Multi-laboratory, highly coordinated research efforts may serve as a model for addressing replicability and reducing efforts based on poorly generalizable results.
In head-to-head comparisons, we show that both morphine and buprenorphine reverse sensory, functional, and affective measures of pain, while psilocybin did not. We then expanded our survey to include both acute and chronic time points, multiple doses of psilocybin, and both sexes in three distinct pain models: spared nerve injury (neuropathic), acid-induced muscle pain (widespread musculoskeletal), or Complete Freund’s Adjuvant (inflammatory). We found that none of the three physiologically active doses possessed any measurable analgesic qualities at any time point in any of these models. We did observe, however, that psilocybin causes a dose-dependent decrease in body temperature, which likely explains the preference for warmer temperature environments in mice treated with psilocybin. This temperature dysregulation may also explain the acute effects of high-dose psilocybin on cold sensitivity.
The tested models have distinct underlying mechanisms providing broad potential to see analgesic effects for a variety of drug classes in current clinical use. Further, the range of behavioral assays used tests different features of the pain experience. Reflexive tests depend on sensory transduction at the peripheral tissue, which then engages a local reflex arc at the level of the spinal cord. While these tests provide valuable insight into peripheral sensitization and spinal cord mechanisms, they do not necessarily evaluate the unpleasantness or functional impairments caused by a pain state at the level of the brain. To address these aspects of pain, we included marble burying as an assay for functional impairment wherein the instinct to bury marbles is opposed by the hyperalgesia triggered by performing the task34. We also tested the effect of laparotomy on facial expressions using the PainFace automated scoring tool. Facial expressions reveal the spontaneous, ongoing affective component of pain31. This combination of sensory, functional, and affective measures gives a more complete understanding of the pain experience and suggests that psilocybin has no effect on these aspects of pain that can be measured in mice.
Our findings are consistent with existing pre-clinical literature on the complex role of serotonin in nociception and analgesia. Psilocin, the active metabolite of psilocybin, potently activates multiple 5-HTRs, which collectively have varied effects at peripheral nociceptors, spinal cord, and descending pain modulation pathways35. Psilocin most notably activates 5-HT2ARs, which mediate the hallucinogenic effect of virtually all serotonergic psychedelics24,29,36,37. However, 5-HT2AR agonists administered peripherally or intrathecally can worsen pain in animal models, while 5-HT2AR antagonists can enhance analgesia38. On the other hand, activation of intrathecal 5-HT1AR has been linked to stress-induced analgesia through an endocannabinoid-mediated mechanism39. Both 5HT1ARs and 5-HT2ARs are expressed in the descending pain modulation system projecting from RVM to spinal cord dorsal horn and have facilitatory and inhibitory effects on pain depending on the cell type and the animal’s conditions35. Notably, our results showed no change in mechanical sensitivity in either naïve or injury models, suggesting psilocybin may not affect pain thresholds regardless of this balance. Plasticity-related effects of psilocin and other psychedelics are not clearly associated with analgesia either: trkB activity, ostensibly stimulated by psilocin-induced release of Brain Derived Neurotrophic Factor36, is associated with worsened mechanical hypersensitivity40 and 5-HT2AR-mediated plasticity in the spinal cord may contribute to chronic neuropathic pain35.
During our screening for analgesic properties of psilocybin, we observed a dose-dependent decrease in body temperature. This finding corroborates previous work41 demonstrating a reduction in core body temperature similar to the time course in our study. Erkizia-Santamaria found a greater magnitude of effect at lower doses, likely due to the greater temperature sensitivity of invasive monitoring. These investigators also provide pharmacologic evidence that 5-HT2AR is necessary for the hypothermic effect41. Thus, behavioral assays, particularly those that rely on variations in temperature, may be confounded by the hypothermic effects of psilocybin. Further, disruption of thermal regulation may be more than an incidental finding. Alitalo et al. demonstrated that drug-induced hypothermia can alter signaling through TrkB, one of the putative mechanisms for psychedelic-induced neuroplasticity42. These data suggest that body temperature may be a key parameter that should be monitored and controlled for in future studies of psilocybin.
Our findings contradict recent pre-clinical studies testing various serotonergic psychedelics in other pain models. Kolbmann et al. showed that i.v. Administration of psilocin produced immediate and persistent reductions in mechanical and thermal hyperalgesia induced by formalin in rats16. This discrepancy may be explained by differences in the route of drug administration or the species used43. Lauria et al. found ayahuasca--an herbal brew most commonly made with material from B. caapi and P. viridis plants containing a range of psychoactive substances, including N, N-dimethyltriptamine (DMT) and multiple harmala alkaloids that act on the 5-HT2AR--had acute (on-drug) analgesic effects in inflammatory and neuropathic pain models, however, consistent with our findings, no analgesic effects were observed at later time points18,44. These plants also contain monoamine oxidase inhibitors, which alter the metabolism of DMT as well as endogenous monoamines and have analgesic properties themselves44,45. A recent preprint reports that psilocybin (both 0.3 mg/kg and 1 mg/kg) very modestly (i.e., visible with log-transformation) improved SNI-induced mechanical sensitivity for days to weeks after a single dose. 0.3 mg/kg psilocybin reduced cold sensitivity over a similar time course, but 1 mg/kg psilocybin appeared less effective19. Overall, preclinical studies from other groups have not consistently demonstrated post-acute analgesic effects of serotonergic psychedelics, and methodological differences or hypothermic effects may potentially explain differences observed at acute time points.
Human studies of the analgesic properties of psychedelics, however, suggest they may have some beneficial effects on the cognitive-evaluative aspects of pain. Ramaekers et al. found that low-dose lysergic acid diethylamide (LSD) improved cold tolerance and reduced the unpleasantness associated with the cold pressor task15. Notably, this effect was only observed at the highest dose, 20 µg, at which the subjects began reporting psychoactive effects from LSD. This finding is consistent with our pre-clinical data showing an acute reduction in cold sensitivity in mice. Other studies of LSD in cancer patients found the drug improved mood, anxiety, and perceived quality of life8,9. These dimensions of pain are outside the scope of rodent models used in our work and merit further evaluation in future studies. Beyond LSD, early-stage trials of psilocybin for headache showed some efficacy; however, the putative analgesic mechanisms for psilocybin may differ from those of other pain conditions. Specifically, activation of 5-HT1B and 5-HT1D receptors in cerebral vasculature may account for the apparent pain relief in certain headache disorders, rather than changes in pain modulation or neuronal plasticity as previously discussed46.
The contrasting results between the present study and data from human subjects highlight important limitations in the use of animal models, particularly mice, in studying the therapeutic efficacy of psychedelics. While animal models for depression and anxiety exist, other dimensions of the human experience may not be amenable to animal studies. Overall quality of life, meaningfulness of existence, optimism, positive attitudes towards life and oneself, and satisfaction with life are substantially improved by psilocybin in human subjects8; however, these characteristics cannot be readily measured in animals. Given the emerging safety data for psilocybin47, it may be prudent to focus future efforts in well-controlled human trials where these nuanced effects can be observed. Our data suggest that mice do not reflect direct analgesic effects of psilocybin in either sensory or affective components; should these properties emerge in clinical studies, mouse models may be challenging to reproduce reliably. The potential beneficial effects on anxiety, mood, and motivation may warrant further investigation. Rats or other, higher-order mammals may be more appropriate model systems.
Our study has several important limitations. First, we did not evaluate all clinically relevant pain models, most notably cancer pain, where psilocybin improves mood-related dimensions of pain, or models like complex regional pain syndrome48, which may have distinct underlying pain mechanisms. Further, our battery of behavioral tests focuses more on hypersensitivity measures with only limited measures of affect and spontaneous pain at a single time point. Second, our timeline is limited to approximately 14 days of chronic pain (21 days in the SNI model). It is possible that mechanisms of chronic pain differ across time, and changes in the CNS that occur later in chronic pain states may be more amenable to the effects of psilocybin. Third, some experiments were not powered to detect small but potentially meaningful sex differences, nor did we track the estrus cycle or perform manipulations of sex hormones. Fourth, our experiments only test a limited exposure to psilocybin without pharmacokinetic validation or repeated administration. While human studies of depression report long-lasting effects after a single therapy-supported dose2, chronic pain may be more responsive to repeated low-dose psychedelics (‘microdosing’), or repeated high-dose regimen49. Ultimately, the potential applications of psychedelics like psilocybin for the treatment of chronic pain will require well-controlled clinical trials conducted with sufficient rigor to establish efficacy, and capturing multidimensional outcome measures that reflect the biological, psychological, and social facets of chronic pain.
Methods
Animals
Male and female (10-16 wk) C57BL/6 J wild-type mice (The Jackson Laboratory, #000664) were housed 4-5 per cage in sterile ventilated cages by Innovive (San Diego, CA) containing Alpha-dri bedding, Enviro-dri enrichment material (Shepherd Specialty Papers), and ad libitum access to pre-filled acidified water bottles (Innovive) and irradiated 18% protein rodent diet (Teklad Global). Animals were maintained on a 12:12 light/dark cycle at 40–60% humidity and 20–24 °C in an SPF facility. They were allowed to habituate to the Stanford University Vivarium for at least 1 week prior to testing. All behavior testing took place during the light cycle. All procedures followed animal care guidelines approved by Stanford University’s Administrative Panel on Laboratory Animal Care (APLAC) and the recommendations of the International Association for the Study of Pain.
Experimenter details
Four experimenters performed the behavioral studies. A female (A.R.) performed the morphine control experiments (Fig. 1C, D) and reproduced the acute mechanical sensitivity experiment in CFA and SNI-treated mice (Fig. 2C, G). The remaining experiments were performed by males: head twitch and rearing by A.B.C.; marble burying, cold plate, body temperature by T.E.G.; grimace, time course experiments, thermal place preference, and cold plate by N.S.G. Behavioral testing and analysis were performed blindly such that the experimenter was unaware of the sham vs. active pain model group assignment, or the drug treatment group.
Compound administered
Psilocybin was obtained from the NIDA Drug Supply Program and reconstituted to a working concentration of 0.03, 0.2, or 1 mg/kg in sterile saline to keep injected volumes consistent across doses. Morphine sulfate (West-Ward, Tinton Falls, NJ, USA) was obtained at a stock concentration of 8 mg/ml and diluted with saline (0.9%) to obtain a working concentration of 1 mg/ml. Buprenorphine was obtained at a concentration of 0.3 mg/mL and diluted to 0.1 mg/mL working concentration (Par Pharmaceuticals, Chestnut Ridge, NY, USA). Mice were dosed intraperitoneally at 0.3, 2, or 10 mg/kg (psilocybin), 10 mg/kg (morphine), or 1 mg/kg (buprenorphine).
Head twitch response and rearing bouts
Before experimentation, animals were acclimatized for at least 90 minutes in a dimly lit room. Subjects were randomized, treated with 0.3, 2, or 10 mg/kg of psilocybin or saline (i.p.), and immediately placed in a transparent acrylic arena (68 × 22 x 26 cm) for observation over 20 minutes. A trained observer (A.B.C.) recorded head twitch responses and rearing behaviors in 2-minute intervals. A head twitch was identified as a rapid, back-and-forth rocking tic movement of the head. Rearing bouts were defined by an unsupported or supported upward movement of the rostral portion of the animal with its forepaws raised off the ground, independent of a grooming bout. After testing, each animal was placed back into its home cage.
Spared Nerve Injury (SNI) model of neuropathic pain50,51
Mice were anesthetized with isoflurane anesthesia (4% induction, 1.5% maintenance in oxygen), and the left hind leg was shaved and wiped clean with alcohol and betadine. A 1 cm incision was made in the skin of the upper thigh, approximately where the sciatic nerve trifurcates. The overlying biceps femoris muscles were retracted by blunt dissection, exposing the common peroneal, tibial, and sural branches of the sciatic nerve. Next, the common peroneal and tibial nerves were ligated with 5-0 sutures (Ethilon) and cut, with care not to distend the sural nerve. The skin was closed with surgical staples, followed by a Betadine application. During recovery from surgery, mice were placed on a warming pad until awake and achieved normal balanced movement. Mice were then returned to their home cage and closely monitored over the following week for well-being. All behavior tests were performed 7 days post-surgery. Following the SNI procedure, the lateral aspect of the hind paw (innervated by the sural nerve) displays a hypersensitive phenotype.
Formalin model of neuropathic pain
The formalin model was induced as previously described52. Briefly, the mice were anesthetized with isoflurane, and the plantar surface of the left hind paw was cleaned with an ethanol wipe. 10 µL of 4% formalin (Sigma-Aldrich) in saline was injected into the plantar surface of the paw using a 30 G needle. Animals in the control group received isoflurane anesthesia alone. Acute cold sensitivity was performed 30 minutes after formalin injection. Mechanical and thermal sensitivity experiments were performed starting 24 hours following formalin injection.
Complete Freund’s Adjuvant (CFA) model of inflammatory pain53
The mice were anesthetized with isoflurane (4% induction, 1.5% maintenance in oxygen), and the plantar surface of the left hind paw was cleaned with an ethanol wipe. 10 μl Complete Freund’s Adjuvant (F5881, Sigma-Aldrich) was injected under the epidermis into the plantar surface of the left hind paw using a 30 G needle. Animals in the control group received light isoflurane anesthesia alone. All behavior tests were performed starting 24 hours following CFA injection.
Acidic Saline-Induced Muscle Pain
Acid Saline-Induced Muscle Pain (AIMP) was induced using the repeated acidic saline injection model54. Specifically, on day 1, mice were lightly anesthetized with 1–2% isoflurane, and 20 µL normal saline adjusted to a pH of 4.0 + 0.1 was injected into the gastrocnemius muscle. This was repeated on day 5. Behavioral testing was then performed starting the next day.
Laparotomy model of surgical pain
The laparotomy model of surgical pain55 was induced by anesthetizing with isoflurane (4% induction, 1.5% maintenance in oxygen), the abdomen was shaved and wiped clean with alcohol and betadine. Each animal received 1 mL normal saline subcutaneously before a 1 cm incision was made through the skin and muscle at approximately the mid-level of the abdomen. A cotton-tipped applicator was brushed against the peritoneal lining for 30 seconds. Then, a 1 cm portion of the bowel was removed and clamped for 10 seconds before being returned to the abdominal cavity. The muscle layer was then closed with 5-0 sutures (Ethilon), and the skin was closed with Vetbond (3 M Company). Animals then received i.p. injection of their assigned experimental compound before being allowed to recover for 30 minutes on a warmed pad.
Mechanical Hypersensitivity
The von Frey filament test51 for mechanical hypersensitivity was performed be placing mice in plexiglass enclosures mounted onto a testing platform with a metal, perforated floor. Mice were acclimated to the testing chamber for 1–2 days prior to the start of the study (1 h/session). On each testing day, the animals were acclimated in the testing chamber for 1 h. Mechanical allodynia was assessed by applying von Frey filaments to the lateral plantar region of each hind paw for approximately 2 s per stimulus using calibrated filaments (Touch Test kit, NC12775-99, North Coast Medical). All trials proceeded using an up–down trial design starting with 0.16 g filament. A sudden paw withdrawal, flinching, or paw licking was regarded as a nocifensive response. A negative response was followed by the use of a larger filament, while a positive response was followed by the use of a lighter filament.
Thermal hypersensitivity
The plantar test (Hargreaves)53 to assess thermal hypersensitivity was performed by placing subjects in plexiglass enclosures on top of a glass platform. The animals were acclimatized to the testing environment for 1–2 days prior to the start of the study (1 h/session). On each testing day, the animals were acclimatized in the testing environment for 20 min, or until cessation of exploratory behavior. A radiant heat source (Plantar Test Analgesia Meter, Harvard Apparatus) was applied to the plantar surface of the hind paw, and the time to a nocifensive response was recorded. A cut-off time of 30 sec was enforced to avoid potential injury due to tissue damage. Two trials were performed on the left (ipsilateral) hind paw to obtain the average reaction time per paw and a third reaction time was obtained if the preceding two values differed by 2 sec or more.
Muscle Withdrawal Threshold (MWT)
Muscle sensitivity to mechanical stimuli was assessed using muscle withdrawal threshold33. Mice were acclimated to the behavior room and restraints in two 5-minute sessions over two days. Testing sessions consisted of three trials, which were averaged for each session. For each trial, the mouse was placed in a gardening glove, and the hind limb was exposed. A force-sensitive tweezer (Bioseb Rodent Pincher – Analgesia Meter, Bioseb, Pinellas Park, FL, USA) was then used to squeeze the belly of the gastrocnemius muscle in a steadily increasing manner such that a peak of 1600 g was achieved over 2 seconds. The trial was terminated when the animal attempted to withdraw the limb or achieved the peak value. Trials that deviated by more than 15% from the targeted rate of force application were discarded and repeated.
Thermal place preference
The affective component of thermal sensitivity was assessed using the thermal place preference test56. 24 h after CFA or saline injection, mice were injected with either saline, morphine 10 mg, or psilocybin (0.3, 2, or 10 mg/kg) 60 minutes before testing. Subjects were then placed on an apparatus consisting of adjacent temperature-controlled plates. For heat sensitivity, the plates were set to 30 °C (neutral) and 40 °C (hot). For cold sensitivity, the plates were set to 30 °C and 20 °C (cold). Mice were initially placed on the neutral plate and allowed to move freely for 5 minutes while a video was recorded. Animal movement was tracked using SLEAP version 1.3.357, and location was scored using SimBA version 1.93.458.
Marble burying
The marble burying test34 was performed to measure the effects of pain on repetitive activity in mice. For each trial, an empty standard habitat was filled with 5 cm of flat ⅛” corncob bedding, and twenty-five marbles were evenly spaced in rows in the enclosure. One mouse was then placed into each cage and left undisturbed for fifteen minutes. Once the mouse was removed, researchers quantified the number of marbles buried (number of marbles that were >⅔ covered in bedding). Marbles are cleaned with dish soap and water between each trial.
Body temperature
Body temperature was assessed by recording mice with a thermal camera (e53, FLIR Systems, Goleta, CA). Mice were injected with the drug and then immediately placed in a plastic enclosure for recording. The area of the body visible to the camera was tracked using SLEAP57. For each image frame, pixel intensities for the area identified as the body were averaged and converted to temperature using the calibration provided by the thermal camera. Frames were then averaged and reported as one-minute bins.
Mouse Grimace Scale
The Mouse Grimace Scale was assessed using the automated machine learning analysis tool PainFace31. Briefly, mice were placed in a plastic enclosure and recorded for 10 minutes to establish a baseline score. Then, 30 minutes after completing laparotomy and receiving saline or drug injection, mice were recorded for a second 10 minute session. These videos were then uploaded to the PainFace website (www.painface.net) where they were automatically scored using the default C57BL/6 model (c57bl6/pain-mgs/20221115-black). A difference score was calculated by subtracting the baseline score from the post-treatment score.
Euthanasia
Mice were euthanized with an APLAC-approved method after completing testing or if they met criteria for humane early euthanization.
Statistical methods
Cohort sizes were determined based on historical data from our laboratory using a power analysis to provide >80% power to discover 25% differences with P < 0.05 between groups, requiring a minimum of 4 animals per sex per group for all behavioral outcomes. Data are expressed as the mean +/− SEM. Statistical analysis was performed using the statsmodels (version 0.14.2) Python library (Three-way ANOVA) or GraphPad Prism version 10.1.0 (all other tests, GraphPad Software). Data were analyzed using One-way analysis of variance (ANOVA), Two-way ANOVA, Three-way ANOVA, or an unpaired t-test, as indicated in the figure captions, with post-hoc testing where appropriate.
For the three-way ANOVA analysis, a sequential approach was used. If drug dose × time or sex × time interaction alone was significant in three-way ANOVA testing, a one-way ANOVA with post-hoc Tukey testing was performed at each individual time point to identify significant differences. If both interactions were significant, then a two-way ANOVA (sex × dose) was performed at each time point. Further, if a significant interaction was identified at a given time point, unpaired t-tests were performed at each drug dose to identify doses with sex differences. The n for each individual experiment is listed in the figure legends.
All tests were two-sided.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Source data are provided with this paper. The individual sex data generated in this study are provided in Source Data 2. Source data are provided with this paper.
References
Raison, C. L. et al. Single-Dose Psilocybin treatment for major depressive disorder: a randomized clinical trial. JAMA 330, 843–853 (2023).
Goodwin, G. M. et al. Single-Dose Psilocybin for a treatment-resistant episode of major depression. N. Engl. J. Med 387, 1637–1648 (2022).
Bogenschutz, M. P. et al. Percentage of heavy drinking days following Psilocybin-Assisted Psychotherapy vs Placebo in the treatment of adult patients with alcohol use disorder: a randomized clinical trial. JAMA Psychiatry 79, 953–962 (2022).
von Rotz, R. et al. Single-dose psilocybin-assisted therapy in major depressive disorder: a placebo-controlled, double-blind, randomised clinical trial. eClinicalMedicine 56, 1–13 (2023).
Dworkin, R. H. et al. If the Doors of Perception Were Cleansed, Would Chronic Pain be Relieved? Evaluating the Benefits and Risks of Psychedelics. J. Pain. 23, 1666–1679 (2022).
Kast, E. C. & Collins, V. J. Study of Lysergic Acid Diethylamide as an Analgesic Agent. Anesth. Analges. 43, 285 (1964).
Grof, S., Goodman, L. E., Richards, W. A. & Kurland, A. A. LSD-Assisted Psychotherapy in Patients with Terminal Cancer. Int. Pharmacopsych. 8, 129–144 (1973).
Griffiths, R. R. et al. Psilocybin produces substantial and sustained decreases in depression and anxiety in patients with life-threatening cancer: A randomized double-blind trial. J. Psychopharmacol. 30, 1181–1197 (2016).
Ross, S. et al. Rapid and sustained symptom reduction following psilocybin treatment for anxiety and depression in patients with life-threatening cancer: a randomized controlled trial. J. Psychopharmacol. 30, 1165–1180 (2016).
Grob, C. S. et al. Pilot study of psilocybin treatment for anxiety in patients with advanced-stage cancer. Arch. Gen. Psychiatry 68, 71–78 (2011).
Aday, J. S. et al. Preliminary safety and effectiveness of psilocybin-assisted therapy in adults with fibromyalgia: an open-label pilot clinical trial. Front. Pain Res. 6, 1–15 (2025).
Bornemann, J., Close, J. B., Spriggs, M. J., Carhart-Harris, R. & Roseman, L. Self-medication for chronic pain using classic psychedelics: a qualitative investigation to inform future research. Front. Psychiatry 12, 735427 (2021).
Bonnelle, V. et al. Analgesic potential of macrodoses and microdoses of classical psychedelics in chronic pain sufferers: a population survey. Br. J. Pain. 16, 619–631 (2022).
Glynos, N. G., Pierce, J., Davis, A. K., McAfee, J. & Boehnke, K. F. Knowledge, Perceptions, and Use of Psychedelics among Individuals with Fibromyalgia. J. Psychoact. Drugs 55, 73–84 (2023).
Ramaekers, J. G. et al. A low dose of lysergic acid diethylamide decreases pain perception in healthy volunteers. J. Psychopharmacol. 35, 398–405 (2021).
Kolbman, N. et al. Intravenous psilocybin attenuates mechanical hypersensitivity in a rat model of chronic pain. Curr. Biol. 33, R1282–R1283 (2023).
Koseli, E. et al. IUPHAR Article: Psilocybin induces long-lasting effects via 5-HT2A receptors in mouse models of chronic pain. Pharmacol. Res. 215, 107699 (2025).
Lauria, P. S. S. et al. Ayahuasca and its major component harmine promote antinociceptive effects in mouse models of acute and chronic pain. J. Ethnopharmacol. 323, 117710 (2024).
Askey, T. et al. Psilocybin ameliorates neuropathic pain-like behaviour in mice and facilitates the gabapentin-mediated analgesia. Preprint at https://doi.org/10.21203/rs.3.rs-5026806/v1 (2024).
Hammo, A., Wisser, S. & Cichon, J. Single-dose psilocybin rapidly and sustainably relieves allodynia and anxiodepressive-like behaviors in mouse models of chronic pain. Nat. Neurosci. 1–11 https://doi.org/10.1038/s41593-025-02068-0 (2025).
Rijsketic, D. R. et al. UNRAVELing the synergistic effects of psilocybin and environment on brain-wide immediate early gene expression in mice. Neuropsychopharmacology 48, 1798–1807 (2023).
Davoudian, P. A., Shao, L.-X. & Kwan, A. C. Shared and Distinct Brain Regions Targeted for Immediate Early Gene Expression by Ketamine and Psilocybin. ACS Chem. Neurosci. 14, 468–480 (2023).
Nardou, R. et al. Psychedelics reopen the social reward learning critical period. Nature 618, 790–798 (2023).
Halberstadt, A. L., Koedood, L., Powell, S. B. & Geyer, M. A. Differential contributions of serotonin receptors to the behavioral effects of indoleamine hallucinogens in mice. J. Psychopharmacol. 25, 1548–1561 (2011).
Shao, L.-X. et al. Psilocybin induces rapid and persistent growth of dendritic spines in frontal cortex in vivo. Neuron 109, 2535–2544.e4 (2021).
Jones, N. T. et al. Transient Elevation of Plasma Glucocorticoids Supports Psilocybin-Induced Anxiolysis in Mice. ACS Pharmacol. Transl. Sci. 6, 1221–1231 (2023).
Shahar, O. et al. Role of 5-HT2A, 5-HT2C, 5-HT1A and TAAR1 Receptors in the Head Twitch Response Induced by 5-Hydroxytryptophan and Psilocybin: Translational Implications. Int J. Mol. Sci. 23, 14148 (2022).
Haberzettl, R., Fink, H. & Bert, B. Role of 5-HT(1A)- and 5-HT(2A) receptors for the murine model of the serotonin syndrome. J. Pharm. Toxicol. Methods 70, 129–133 (2014).
González-Maeso, J. et al. Hallucinogens recruit specific cortical 5-HT(2A) receptor-mediated signaling pathways to affect behavior. Neuron 53, 439–452 (2007).
Lu, O. D. et al. A multi-institutional investigation of psilocybin’s effects on mouse behavior. 2025.04.08.647810 Preprint at https://doi.org/10.1101/2025.04.08.647810 (2025).
McCoy, E. S. et al. Development of PainFace software to simplify, standardize, and scale up mouse grimace analyses. PAIN https://doi.org/10.1097/j.pain.0000000000003187 (2022).
Matsumiya, L. C. et al. Using the mouse grimace scale to reevaluate the efficacy of postoperative analgesics in laboratory mice. J. Am. Assoc. Lab. Anim. Sci. 51, 42–49 (2012).
Skyba, D. A., Radhakrishnan, R. & Sluka, K. A. Characterization of a method for measuring primary hyperalgesia of deep somatic tissue. J. Pain. 6, 41–47 (2005).
Ruskin, D. N., Sturdevant, I. C., Wyss, L. S. & Masino, S. A. Ketogenic diet effects on inflammatory allodynia and ongoing pain in rodents. Sci. Rep. 11, 725 (2021).
Heijmans, L., Mons, M. R. & Joosten, E. A. A systematic review on descending serotonergic projections and modulation of spinal nociception in chronic neuropathic pain and after spinal cord stimulation. Mol. Pain. 17, 17448069211043965 (2021).
Heifets, B. D. & Olson, D. E. Therapeutic mechanisms of psychedelics and entactogens. Neuropsychopharmacol 49, 104–118 (2024).
Glatfelter, G. C. et al. Structure–activity relationships for Psilocybin, Baeocystin, Aeruginascin, and related analogues to produce pharmacological effects in mice. ACS Pharmacol. Transl. Sci. 5, 1181–1196 (2022).
Sierra, S. et al. Sex-specific role for serotonin 5-HT2A receptor in modulation of opioid-induced antinociception and reward in mice. Neuropharmacology 209, 108988 (2022).
Mallet, C. et al. Endocannabinoid and serotonergic systems are needed for acetaminophen-induced analgesia. Pain 139, 190–200 (2008).
Wang, X., Ratnam, J., Zou, B., England, P. M. & Basbaum, A. I. TrkB signaling is required for both the induction and maintenance of tissue and nerve injury-induced persistent pain. J. Neurosci. 29, 5508–5515 (2009).
Erkizia-Santamaría, I., Alles-Pascual, R., Horrillo, I., Meana, J. J. & Ortega, J. E. Serotonin 5-HT2A, 5-HT2c and 5-HT1A receptor involvement in the acute effects of psilocybin in mice. In vitro pharmacological profile and modulation of thermoregulation and head-twitch response. Biomed. Pharmacother. 154, 113612 (2022).
Alitalo, O. et al. Linking Hypothermia and Altered Metabolism with TrkB Activation. ACS Chem. Neurosci. 14, 3212–3225 (2023).
Mogil, J. S. The translatability of pain across species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 374, 20190286 (2019).
Ayahuasca: A review of historical, pharmacological, and therapeutic aspects - Ruffell - 2023 - Psychiatry and Clinical Neurosciences Reports - Wiley Online Library. https://onlinelibrary-wiley-com.laneproxy.stanford.edu/doi/full/10.1002/pcn5.146.
Villarinho, J. G. et al. The antinociceptive effect of reversible monoamine oxidase-A inhibitors in a mouse neuropathic pain model. Prog. Neuropsychopharmacol. Biol. Psychiatry 44, 136–142 (2013).
Schindler, E. A. D. Psychedelics as preventive treatment in headache and chronic pain disorders. Neuropharmacology 215, 109166 (2022).
Jevotovsky, D. S. et al. Psilocybin and chronic neuropathic pain: a systematic review. Reg. Anesth. Pain Med. https://doi.org/10.1136/rapm-2024-105532 (2024).
Donovan, L. J. et al. Repopulated spinal cord microglia exhibit a unique transcriptome and contribute to pain resolution. Cell Rep. 43, 113683 (2024).
Cavarra, M. et al. Potential analgesic effects of psychedelics on select chronic pain conditions: A survey study. Eur. J. Pain. 28, 153–165 (2024).
Cichon, J., Sun, L. & Yang, G. Spared Nerve Injury Model of Neuropathic Pain in Mice. Bio Protoc. 8, e2777 (2018).
Wang, D. et al. Functional Divergence of Delta and Mu Opioid Receptor Organization in CNS Pain Circuits. Neuron 98, 90–108.e5 (2018).
Ruan, Y. et al. An effective and concise device for detecting cold allodynia in mice. Sci. Rep. 8, 14002 (2018).
Ram, A. et al. GPR171 Agonist Reduces Chronic Neuropathic and Inflammatory Pain in Male, But Not Female Mice. Front. Pain Res. 2, 1–13 (2021).
Sluka, K. A., Kalra, A. & Moore, S. A. Unilateral intramuscular injections of acidic saline produce a bilateral, long-lasting hyperalgesia. Muscle Nerve 24, 37–46 (2001).
Durst, M. S., Arras, M., Palme, R., Talbot, S. R. & Jirkof, P. Lidocaine and bupivacaine as part of multimodal pain management in a C57BL/6J laparotomy mouse model. Sci. Rep. 11, 10918 (2021).
Modi, A. D., Parekh, A. & Pancholi, Y. N. Evaluating pain behaviours: Widely used mechanical and thermal methods in rodents. Behav. Brain Res. 446, 114417 (2023).
Pereira, T. D. et al. SLEAP: A deep learning system for multi-animal pose tracking. Nat. Methods 19, 486–495 (2022).
Goodwin, N. L. et al. Simple Behavioral Analysis (SimBA) as a platform for explainable machine learning in behavioral neuroscience. Nat. Neurosci. 27, 1411–1424 (2024).
Acknowledgements
We thank the entire Heifets, Tawfik, and Malenka Labs for helpful discussions, and the NIDA Drug Supply Program for supplying psilocybin. This work was supported by a Mentored Research Award from the International Anesthesia Research Society (N.S.G.), NIH grant T32DA035165 (A.R.) NIH grant P50 DA042012 (B.D.H. and R.C.M.), funds from the Department of Anesthesiology, Perioperative & Pain Medicine at Stanford University (V.L.T and B.D.H.), and a grant from the Stanford University Wu Tsai Neurosciences Institute (R.C.M.).
Author information
Authors and Affiliations
Contributions
Nicholas S. Gregory (N.S.G.) designed and performed experiments (CFA, SNI, AIMP, laparotomy models; von Frey, cold plate, Hargreaves, thermal place preference, muscle withdrawal threshold, mouse grimace scale), analyzed the data, interpreted the results, and wrote the manuscript; Tyler E. Girard (T.E.G.) performed experiments (cold plate, marble burying, body temperature tracking), analyzed data, and contributed to the manuscript; Akila Ram (A.R.) performed experiments (morphine controls, von Frey, Hargreaves) and contributed to the manuscript; Austen B. Casey (A.B.C.) performed experiments (rearing, head twitch) and contributed to the manuscript; Robert C. Malenka (R.C.M.) analyzed data and interpreted results; Vivianne L. Tawfik (V.L.T.) conceived of the study, designed experiments, interpreted results, and revised the manuscript; Boris D. Heifets (B.D.H.) conceived of the study, designed experiments, analyzed data, interpreted results, and contributed to the manuscript.
Corresponding author
Ethics declarations
Competing interests
B.D.H. is on the scientific advisory boards of Journey Clinical and Osmind, and is a paid consultant to Arcadia Medicine, Inc, Tactogen, LLC, and Vida Ventures, LLC. R.C.M. is now on leave from Stanford, functioning as Chief Scientific Officer at Bayshore Global Management. R.C.M. is on the scientific advisory boards of MapLight Therapeutics, Bright Minds, MindMed, and Aelis Farma. The remaining authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Gregory, N.S., Girard, T.E., Ram, A. et al. No evidence of immediate or persistent analgesic effect from a single dose of psilocybin in three mouse models of pain. Nat Commun 17, 1916 (2026). https://doi.org/10.1038/s41467-026-68763-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-68763-z
