
Abstract
Nucleic acid degradation is a common strategy for prokaryotic anti-phage systems, as exemplified by the CRISPR-Cas system. The PD-(D/E)-XK nucleases constitute a widely distributed family in these defenses. Notably, most members exhibit a single nuclease domain, while variants containing dual nuclease domains within a single polypeptide remain underexplored, and their molecular mechanisms largely obscure. Here, we biochemically and functionally study a single-protein system containing an uncharacterized PD-(D/E)-XK defense protein (Upx). As revealed by single-particle electron cryo-microscopy (cryo-EM) structure, the C-terminal domain (CTD) harboring the conserved PD-(D/E)XK catalytic core is buttressed by the N-terminal domain (NTD) and the middle domain (MD). Functional assays demonstrate that the nucleic acid binding capability of the CTD is enhanced by the MD. The NTD also displays a noncanonical, basal exonuclease activity that is auto-inhibited by MD. IP-MS experiments identify Upx-interacting phage proteins, and substrate profiling defines its physiological preferences, collectively pointing to its potential physiological targets. Notably, the phage protein gp16 was found to relieve MD-mediated inhibition of the NTD, suggesting a virus-triggered mechanism for activating Upx’s dual nuclease activity. Together, these findings establish Upx as a single-protein dual-nuclease anti-phage system, expanding our understanding of bacterial immunity and informing antiviral strategy development.
Similar content being viewed by others


Introduction
Bacteria and bacteriophages participate in an ongoing coevolutionary arms race, driving the development of sophisticated strategies for phage exploitation and host manipulation1,2,3. In response, bacteria have evolved a diverse arsenal of immune defenses, collectively known as anti-phage defense systems4,5,6,7. These anti-phage defense systems include innate mechanisms like restriction-modification systems, as well as adaptive strategies such as CRISPR-Cas6,8,9,10. These systems were historically considered the primary lines of bacterial defense; recent genomic and functional analyses have uncovered an extensive array of anti-phage strategies clustered within genomic “defense islands”3,11,12. These discoveries have expanded the known bacterial immune landscape to over 154 distinct defense systems, many of which remain mechanistically uncharacterized.
Among these, the PD-(D/E)XK superfamily of nucleases, defined by a conserved catalytic core and a bipartite PD-(D/E)XK motif, constitutes a major class of phage defense systems13,14,15,16,17. These nucleases typically function as single-nuclease domain enzymes that cleave nucleic acids with high specificity15,18. However, their functional diversity, regulatory mechanisms, and evolutionary adaptations remain poorly understood. Recently, Upx has been identified as a putative single-gene defense factor within the PD-(D/E)XK family11. While its C-terminal domain (CTD) was predicted to adopt a canonical PD-(D/E)XK fold, the functional role of its N-terminal domain (NTD) remained enigmatic due to the absence of recognizable catalytic motifs11. The lack of structural and biochemical characterization has further obscured its underlying molecular mechanism. Notably, phylogenetic and genomic analyses place Upx within a clade of single-protein defense systems, raising fundamental questions about how a solitary protein can mediate robust phage resistance19.
Here, we show Upx as an example of a single-polypeptide anti-phage system that integrates two distinct nuclease domains within a unified architecture. Using Cryo-EM, we resolve the Upx structure and reveal a tripartite organization comprising an auto-inhibited N-terminal nuclease (NTD), a central regulatory middle domain (MD), and a C-terminal PD-(D/E)XK nuclease domain (CTD)20. Biochemical characterization demonstrates that the CTD functions as a magnesium-dependent, nonspecific 3′–5′ exonuclease targeting both ssDNA and ssRNA, while the NTD—previously unrecognized as a nuclease—harbors a minimal PD-(D/E)XK-like catalytic core with specificity for ssDNA degradation. Notably, the MD acts as a regulatory hub, suppressing NTD activity while enhancing CTD efficiency. Our IP-MS analysis revealed that the phage protein gp16 interacts with Upx and functionally relieves the MD-mediated autoinhibition of the NTD. The phylogenetic restriction of gp16 to ΦV-1 and a narrow set of related phages provides the molecular basis for Upx’s narrow antiphage spectrum. Substrate profiling further defines Upx’s physiological preferences, providing insights into its potential in vivo targets and the functional relevance of its dual nuclease activities. Functional assays confirm that both nuclease domains cooperate to disrupt phage replication, thereby conferring resistance against bacteriophage ФV-1. Our findings redefine the functional landscape of PD-(D/E)XK nucleases by demonstrating that single-protein defense systems can evolve dual enzymatic activities through domain fusion and regulatory specialization. This work expands the mechanistic repertoire of bacterial immunity. It provides a framework for understanding how modular domain architectures drive the evolution of sophisticated anti-phage strategies within compact genetic systems.
Results
Phylogenetic analysis of Upx as a member of PD-(D/E)XK superfamily in bacterial defense system
The PD-(D/E)XK superfamily is defined by a conserved structural core known as the restriction endonuclease-like fold13,14. Among the 154 prokaryotic defense systems included in the DefenseFinder Wiki (https://defensefinder.mdmlab.fr/wiki/), 13 defense systems are part of this superfamily. Notably, 8 of these, including Upx, are made up of a single gene, suggesting that they may function as solitary nuclease proteins and directly target nucleic acids in their anti-phage defense activity.
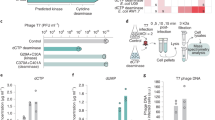
To explore the functional diversity of PD-(D/E)XK superfamily members in bacterial immunity, we aligned 1000 representative protein sequences using MAFFT. The phylogenetic tree was generated using the IQ-TREE server and then visualized with FigTree. The analysis indicated that the 13 defense systems in PD-(D/E)XK superfamily collected in DefenseFinder span in seven clades based on sequence similarity, with Upx closely related to AbiU-1 (Fig. 1a). Previous studies have shown that AbiU-1 inhibits phage DNA transcription in infected bacterial cells16, suggesting that Upx may confer phage resistance through a similar mechanism.

a Phylogenetic analysis of Upx as a member of PD-(D/E)XK superfamily. b Phage challenge assays with Upx or the E1176A/K1178A mutants. Empty vector control, Upx or the E1176A/K1178A mutant were expressed in E. coli MG1655, which were then challenged with phages vB, Mu, SP, T6, T1, P1, and φV-1 at serial dilutions. c Gel filtration analysis of Upx protein using Superdex 200. Source data are provided as a Source Data file. d Peak fraction of Upx protein analyzed by SDS-PAGE. All assays were performed in triplicate.
Upx protects bacterial cells from bacteriophage infections
To evaluate the phage defense capability of Upx, we challenge cells expressing Upx in plaque assays with E. coli phages representing seven distinct genera (Fig. 1b). Upx gene (NCTC6026_01102, protein SQI81149.1) from Salmonella enterica NCTC6026 was cloned into the pBAD LIC cloning vector (8 A) and transformed into E. coli MG1655. Solutions comprising seven distinct bacteriophages were prepared and serially diluted 10-fold in LB medium. An aliquot of 2.5 µL from each dilution was applied onto bi-layer agar plates that had been pre-inoculated with E. coli MG1655. The assays were performed in triplicates, with the following groups: blank control (empty vector), negative control (Upx inactivating mutant E1176A/K1178A), and experimental (Upx), and the representative results are shown in Fig. 1b.
The results indicated that Upx expression conferred significant resistance to phage ФV-1 infection in E. coli MG1655 but did not provide substantial protection against the other six types of bacteriophages. However, the precise molecular mechanisms underlying this resistance remain unclear and require further investigation.
The Cryo-EM structure of Upx
To elucidate the molecular mechanisms underlying the anti-phage capability of Upx, we expressed and purified the Upx protein from Escherichia coli BL21(DE3). Results of the Gel filtration chromatography experiment using a Superdex 200 column showed an eluting peak at 14.3 mL, suggesting that Upx likely exists as a monomer in solution (Fig. 1c), a conclusion further supported by analytical ultracentrifugation (AUC) experiments (Supplementary Fig. 1a). The result of the SDS-PAGE assay showed that the molecular weight of Upx is approximately 143 kDa, with sufficient purity for subsequent experiments (Fig. 1d).
The structural characteristics of Upx were analyzed using cryo-EM, resulting in a structure with a high resolution of 3.01 Å (Supplementary Fig. 1b-d, Supplementary Fig. 2; Supplementary Table 1). The cryo-EM structure confirmed that Upx adopts a symmetric monomeric conformation, consistent with the Gel filtration chromatography and AUC results (Fig. 2b, c; Supplementary Fig. 1a). A DALI search against the PDB database yielded no significant structural homologs, indicating that Upx adopts a fold that is distinct from those of previously characterized proteins (Supplementary Fig. 3). Upx consists of three distinct domains: the N-terminal domain (NTD, residues 1–515), the middle domain (MD, residues 516–714), and the C-terminal domain (CTD, residues 715–1272), arranged in a fusiform shape with the NTD and CTD at opposite ends and the MD in the center (Fig. 2a-c). Although the three domains are relatively distinct, they are structurally interconnected and likely function cooperatively.

a Domain organization of Upx. b Cryo-EM density map of Upx. The density corresponding to the NTD, MD, and CTD is colored lightpink, paleyellow, and palegreen, respectively. c Cryo-EM structures of Upx. The cartoon representation of the three distinct domains is colored according to (b).
Upx is a magnesium-dependent 3’−5’ exonuclease targeting ssDNA
Having resolved the structure of Upx, we next sought to characterize its nuclease activity. To this end, we performed cleavage assays using single-stranded DNA (ssDNA) and RNA (ssRNA) substrates labeled with Cy3 fluorophores at either the 5’ or 3’ ends. Upx efficiently degraded 5’ Cy3-labeled ssDNA into progressively shorter fragments, producing a degradation pattern characteristic of exonuclease activity (Fig. 3a). In contrast, no cleavage was detected for the 3’ Cy3-labeled ssDNA substrate (Supplementary Fig. 4a), suggesting that the bulky Cy3 moiety at the 3’ terminus prevented enzymatic access and thereby revealed the 3′−5′ directionality of nuclease activity. Similarly, while Upx only weakly acted upon 5’ Cy3-labeled ssRNA, it showed no activity 3’-labeled ssRNA, further supporting its role as a 3′−5′ exonuclease (Fig. 3b; Supplementary Fig. 4b). To validate this observation, we introduced three phosphorothioate linkages at the 3’ end of ssDNA, a modification known to block exonuclease cleavage. As anticipated, Upx completely lost nuclease activity against this modified substrate (Fig. 3c). By contrast, no detectable cleavage was observed on double-stranded DNA (dsDNA), indicating that Upx selectively targets single-stranded nucleic acids (Fig. 3d).

a The 5’-Cy3-labeled 60 nt ssDNA was incubated with Upx, and cleavage assays were performed over a time course ranging from 10 to 30 min. b The 5’-Cy3-labeled 45 nt ssRNA was incubated with Upx, and cleavage assays were performed over a time course ranging from 10 to 30 min. c Cy3-labeled ssDNA substrates (5’ end labeled) with phosphorothioate (PT) modifications at the three terminal nucleotides of the 3’ end are resistant to cleavage by full-length Upx. d Cleavage assays were performed using dsDNA substrates bearing a 5’ Cy3 label on one strand and a 5’ Cy5 label on the complementary strand at a protein-to-substrate molar ratio of 20:1. e Cleavage of ssDNA substrates of different sequences and lengths (all labeled with Cy3 at the 5’ end) by Upx. f Metal ion dependency of Upx nuclease activity assayed with 5’ Cy3-labeled ssDNA (60 nt) in the presence of 0.2 mM Mg²⁺, Fe²⁺, Ni²⁺, Ca²⁺, Co²⁺, or Zn²⁺. g Cleavage assays of 5’ Cy3-labeled ssDNA (60 nt) by full-length Upx and its truncated variants, including NTD, NTD-MD, MD, CTD, and MD-CTD. All cleavage products were analyzed by 13% denaturing urea-PAGE and visualized by fluorescence imaging. Each assay was performed in triplicate. h Cleavage assays of 5’ Cy3-labeled ssRNA (60 nt) by full-length Upx and its truncated variants, including NTD, NTD-MD, MD, CTD, and MD-CTD. i Structural features of the CTD catalytic core. The four β-strands are shown in lime and the two α-helices in marine. Key residues (E1176, D1161, K1178) are highlighted in red. j Metal ion dependency of CTD nuclease activity assayed with 5’ Cy3-labeled ssDNA (60 nt) in the presence of 0.2 mM Mg²⁺, Fe²⁺, Ni²⁺, Ca²⁺, Co²⁺, or Zn²⁺. All cleavage products were analyzed by 13% denaturing urea-PAGE and visualized by fluorescence imaging. Each assay was performed in triplicate.
We next investigated whether Upx exhibits sequence or length selectivity. Substrates of different lengths (60 nt, 50 nt, and 30 nt) and different sequences were all efficiently degraded, indicating that Upx functions as a sequence- and length-independent exonuclease (Fig. 3e). Similarly, ssRNA substrates of varying lengths (45 nt and 60 nt) were also cleaved, albeit with markedly lower efficiency compared to ssDNA (Fig. 3b; Supplementary Fig. 4c). Finally, we examined the requirement for divalent metal ions. To explore the metal ion dependency of Upx’s nuclease activity, we examined seven divalent metal ions: magnesium (Mg²⁺), iron (Fe²⁺), nickel (Ni²⁺), cerium (Ce²⁺), cobalt (Co²⁺), zinc (Zn²⁺), and Manganese (Mn2+). The findings revealed that the degradation of ssDNA by Upx and CTD was facilitated by magnesium (Mg²+). Manganese (Mn2+) also supported substantial activity, while iron (Fe²⁺) permitted only trace cleavage. The remaining cations did not support significant activity (Fig. 3f; Supplementary Fig. 5a).
The CTD harbors a canonical PD-(D/E)XK catalytic core
Given that Upx is composed of three domains (NTD, MD, and CTD), we next sought to determine which domain is responsible for its nuclease activity. To this end, we purified truncation constructs corresponding to individual domains (NTD, CTD) and paired domains (NTD–MD, MD–CTD) and evaluated their nuclease activity. The MD–CTD truncation exhibited cleavage activity comparable to that of full-length Upx, whereas CTD alone retained partial activity on both ssDNA, but not dsDNA (Fig. 3d, g). Interestingly, the NTD construct displayed detectable nuclease activity, but this activity was abolished when fused with the MD (Fig. 3g-h), suggesting that MD may exert a regulatory effect on NTD-mediated catalysis, which we investigated in detail in a later section.
The PD-(D/E)XK family members share a common structural core, comprising four or five β-sheets flanked on both sides by α-helices. This core is often embedded within highly variable peripheral regions. The central β-sheet serves as a scaffold for a weakly conserved active site, typically comprising two or three acidic residues (Asp or Glu) along with a Lys residue, collectively forming the characteristic bipartite catalytic motif PD-(D/E)XK, where X is any amino acid21,22. To definitively classify the CTD, we performed a structural alignment with the DALI server, which identified it as a canonical member of the PD-(D/E)XK superfamily (Fig. 3i; Supplementary Fig. 4d-i). The core domain (residues 1121–1272) forms the characteristic fold of two α-helices and four β-sheets, featuring a sharply kinked α2 helix that, together with α1, defines the active site pocket (Fig. 3i). The conserved active site is defined by key residues D1161 in β2 and E1176/K1178 in β3, resembling the active site architecture of the restriction enzyme FokI and other members of the PD-(D/E)XK superfamily (Supplementary Fig. 4d-i). This structural evidence further confirms that Upx is a representative member of the PD-(D/E)XK superfamily from a structural biology perspective.
To explore the metal ion dependency of CTD’s nuclease activity, we examined six divalent metal ions: magnesium (Mg²⁺), iron (Fe²⁺), nickel (Ni²⁺), cerium (Ce²⁺), cobalt (Co²⁺), and zinc (Zn²⁺). The findings revealed that the degradation of ssDNA by CTD was facilitated by magnesium (Mg²+) and, to a lesser extent, iron (Fe²+). At the same time, the other tested divalent cations did not significantly affect cleavage activity (Fig. 3j).
The NTD contains an atypical simplified PD-(D/E)XK catalytic core
Cryo-EM analysis revealed that the NTD harbors a distinct, negatively charged pocket consistent with a potential nuclease active site (Fig. 4a, b). Biochemical assays confirmed that the NTD functions as a magnesium-dependent 3′–5′ exonuclease selectively targeting ssDNA, whereas no cleavage was detected toward ssRNA or dsDNA substrates (Fig. 4c; Supplementary Fig. 4c). Structure-guided mutagenesis identified five charged residues—D122, D127, R129, E147, and D173—as critical for catalysis, since their substitution markedly impaired or abolished nuclease activity (Fig. 4d; Supplementary Fig. 5b-e). Structural inspection showed that D127 and R129 form a DXR motif on β2, analogous to the canonical DXK catalytic triad of PD-(D/E)XK nucleases, while D122, E147, and D173 reside on β1, α1, and α2, respectively (Fig. 4b, g). Unlike the classical PD-(D/E)XK fold comprising four or five β-strands, the NTD active site is formed by only two β-strands and two flanking helices, representing a simplified yet catalytically competent variant of this nuclease scaffold. Domain topology comparison further revealed that while the Upx CTD adopts the canonical PD-(D/E)XK architecture, the NTD constitutes a noncanonical, minimalist version, serving as a structurally convergent counterpart (Fig. 4e-g). Collectively, these findings identify a previously unrecognized nuclease topology, expanding the structural and functional diversity of the PD-(D/E)XK superfamily.

a Electrostatic surface representation of Upx-NTD showing the catalytic pocket (blue circle). b Close-up view of the catalytic core, with key residues shown as sticks. c Metal ion dependency of NTD nuclease activity assayed with 5’ Cy3-labeled ssDNA (60 nt) in the presence of 0.2 mM Mg²⁺, Fe²⁺, Ni²⁺, Ca²⁺, Co²⁺, or Zn²⁺. All cleavage products were analyzed by 13% denaturing urea-PAGE and visualized by fluorescence imaging. Each assay was performed in triplicate. d Effects of mutating key catalytic residues (D122A/R129A, D127A, E147A, D173A, K111A/K134A, D135A/N138A) on nuclease activity, assayed using 5’ Cy3-labeled ssDNA (60 nt) at a protein-to-substrate ratio of 20:1. All cleavage products were analyzed by 13% denaturing urea–PAGE and visualized by fluorescence imaging. Each assay was performed in triplicate. e–g Topological comparison of the classical PD-(D/E)XK fold with the catalytic cores of Upx-CTD and Upx-NTD. Key residues were annotated as asterisks.
The MD structurally integrates and functionally regulates the NTD and CTD
The three domains of Upx form an integrated structural unit, with the middle domain (MD) serving as a central hub that physically connects the NTD and CTD (Fig. 5a). This architecture provides the structural basis for its regulatory role. Detailed analysis shows distinct interaction patterns at the two interfaces: the MD–NTD interface is dominated by electrostatic interactions involving residue pairs D122–R517, R183–E562, R183–D514, and D514–R530 (Fig. 5b), while the MD–CTD interface employs a more diverse stabilization strategy through hydrogen bonds (R595–L820, N705–G819, R707–L818), electrostatic interactions (R541–E824), and π–π stacking (F536–W810) (Fig. 5c). Functionally, the MD exerts differential regulation on the two nuclease domains. It enhances CTD activity—while the isolated CTD exhibits only weak cleavage activity, fusion with the MD (MD–CTD) significantly improves nuclease efficiency on ssDNA (Fig. 3g). This enhancement is corroborated by EMSA, where CTD alone fails to produce a detectable shift with ssDNA, likely due to transient binding, while the MD–CTD construct yields a clear shift, indicating that the MD promotes stable substrate engagement (Fig. 5d).

a Domain organization of Upx showing NTD, MD, and CTD. b–c Interaction assays between MD and NTD (b) or MD and CTD (c). d Electrophoretic mobility shift assays (EMSA) of 5’ Cy3-labeled ssDNA (60 nt) incubated with full-length Upx or its truncated variants. All assays were performed in triplicate. e Pulldown assay showing interaction between Upx and gp16.Each assay was performed in triplicate. f gp16 relieves MD-mediated inhibition of NTD nuclease activity, assayed using 5’ Cy3-labeled ssDNA (60 nt). All cleavage products were analyzed by 13% denaturing urea-PAGE and visualized by fluorescence imaging. Each assay was performed in triplicate. g–k Cleavage activity of Upx on different structured DNA substrates, including 5’ overhang DNA, 3’ overhang DNA, partial duplex DNA, and forked DNA. Asterisks denote the Cy5-labeled strand. All cleavage products were analyzed by 13% denaturing urea-PAGE and visualized by fluorescence imaging. Each assay was performed in triplicate. l Cleavage assays were performed using the unpaired 3’ single-stranded regions within both D-loop and R-loop structures. Asterisks denote the fluorescently labeled strands (Cy3, green; Cy5, magenta). Each assay was performed in triplicate. m Cleavage assays were performed using D-loop structures labeled with 5’ Cy3 at the terminus. The cleavage products were analyzed by 6% native urea–PAGE and visualized by fluorescence imaging. Each assay was performed in triplicate.
Conversely, the MD inhibits the activity of NTD. Although the isolated NTD cleaves ssDNA efficiently, fusion with the MD (NTD–MD) abolishes this activity (Fig. 3g). Structural analysis shows that the MD partially occludes the NTD catalytic pocket (Fig. 5a), restricting substrate access. Consistently, EMSA revealed that the isolated NTD binds ssDNA effectively, whereas the NTD–MD construct shows no mobility shift, confirming that the MD suppresses both substrate binding and cleavage by the NTD (Fig. 5d).
To explore the physiological regulation of Upx, we performed MBP-tagged IP–MS under phage infection conditions (Supplementary Fig. 6a). This identified several phage-derived interactors, and pull-down assays confirmed direct binding between Upx and the phage protein gp16 as well as DNA polymerase (Fig. 5e; Supplementary Fig. 6b; Supplementary Table 2). Functional assays showed that gp16—a large multimeric virion structural protein that forms part of the phage injection apparatus and engages the host during the earliest stage of infection—specifically relieves MD-mediated inhibition and restores NTD activity in both the NTD-MD fragment and full-length Upx, whereas DNA polymerase showed no such activating effect (Fig. 5f; Supplementary Fig.6c,f). Pull-down mapping revealed that gp16 binds broadly to multiple Upx domains (NTD, CTD, NTD–MD, and MD–CTD), but not to the MBP control (Supplementary Fig. 6d, e), suggesting a potential activation mechanism in which gp16 counteracts MD repression to unlock NTD nuclease function during infection. Phylogenetic analysis shows that gp16 is uniquely present in ΦV-1 and a narrow set of related phages, but absent in all Upx-insensitive phages tested, including T6, Mu, P1, SP, T1, and vB (Supplementary Fig. 7; Supplementary Fig. 8). This restricted distribution provides a mechanistic explanation for the narrow antiphage spectrum of Upx, linking its activation to a phage-encoded specificity determinant.
Substrate profiling reveals Upx’s preference for phage replication intermediates
Since ΦV-1 is a dsDNA phage yet Upx lacks dsDNA cleavage activity, its physiological substrates are likely single-stranded nucleic acid intermediates generated during essential processes such as replication, transcription, or recombination. To delineate the substrate selectivity of Upx, we examined its activity toward a panel of structured nucleic acids mimicking physiological intermediates, including 5’ overhangs, 3’ overhangs, fork structures, D-loops, and R-loops (Fig. 5g-m; Supplementary Table 3, 4). Full-length Upx preferentially cleaved substrates bearing unpaired 3’ ssDNA termini, such as 3’ overhangs and fork structures (Fig. 5g, j, k), but showed no activity against 5’ overhangs or 3’ ends blocked with a Cy3 moiety. Given that DNA-RNA hybrids and D-loop intermediates frequently arise during phage transcription, replication, and host recombination, we next asked whether Upx could process these complex substrates. Notably, Upx exhibited robust activity against D-loop structures but only trace activity against R-loops (Fig. 5l). To further dissect its cleavage specificity, we designed additional D-loop variants and found that Upx only cleaved those possessing a 3’ overhang (Fig. 5m), consistent with its identified 3’−5’ exonuclease activity. In parallel, IP-MS suggested a possible interaction between Upx and phage DNA polymerase, which was subsequently confirmed by pull-down assays (Fig. 5e).
Collectively, these results support a model in which Upx targets single-stranded nucleic acid intermediates—such as those generated during phage genome replication, transcription, or host recombination—to exert anti-phage activity.
Upx suppresses phage genome accumulation and reprograms host transcription during infection
To directly assess the impact of Upx on phage genome dynamics during infection, we performed whole-genome sequencing at early (5 min) and late (20 min) time points following phage challenge in E. coli strains expressing wild-type Upx, a catalytically inactive Upx mutant (Upx mutation), or an empty vector control. In both intervals, normalized relative DNA abundance of phage genomes was markedly reduced in the Upx-expressing cells compared with either the vector control or the nuclease-inactive mutant (Fig. 6a), demonstrating that Upx efficiently cleaves incoming phage DNA immediately upon infection and continues to suppress its accumulation through later stages.

a Normalized read depth (RD) of phage ΦV-1 genomes at 5 min (left) and 20 min (right) post-infection in cells expressing wild-type Upx, a catalytically inactive mutant (Upx-mut), or an empty vector control. RD values were obtained from a single sequencing dataset per condition and normalized to the sample specific genomic average RD of the combined E. coli–ΦV-1 reference genome. Each condition represents an independently prepared sample. b Hierarchical clustering of differentially expressed genes (DEGs, adjusted p-value < 0.05) between Upx-expressing and non-expressing cells following phage infection (n = 3 biologically independent samples). Red and blue indicate up- and downregulation, respectively. Statistical analysis was performed using the Wald test within the DESeq2 framework. P-values were two-sided and adjusted for multiple comparisons using the Benjamini-Hochberg (BH) method to control the false discovery rate (FDR). c Vocanal plot of expression levels of ΦV-1 phage genes. The x-axis represents Log2Fold Change (log₂FC) of gene expression, and the y-axis represents -log₁₀ of adjusted P-value (padj). Dashed vertical lines indicate the log₂FC threshold ( | log₂FC | = 1), and the dashed horizontal line indicates the significance threshold (padj = 0.05). Blue dots represent significantly downregulated genes of phage ФV-1 (log₂FC < -1 and padj <0.05). Statistical analysis was performed using the Wald test within the DESeq2 framework on data from n = 3 biologically independent samples. P-values were two-sided and adjusted for multiple comparisons using the Benjamini-Hochberg (BH) method to control the false discovery rate (FDR). d Gene Ontology (GO) enrichment analysis of downregulated DEGs from (b). Enriched terms in Biological Process (BP), Cellular Component (CC), and Molecular Function (MF) categories are shown. Dot size represents the number of genes per term; color corresponds to the GO Category. Functional enrichment was assessed using the hypergeometric test (via the clusterProfiler package). The p-values represent one-sided tests (testing for over-representation). Adjustments for multiple testing were made using the Benjamini-Hochberg (BH) method (analysis based on n = 3 biological replicates).
To determine how Upx interferes with the transcriptional progression of ΦV-1 infection, we performed RNA-seq on infected cells with or without Upx expression. Hierarchical clustering revealed a strong separation between the two groups (Fig. 6b), indicating that Upx profoundly reshapes the transcriptional landscape during phage attack. Direct analysis of phage-derived transcripts showed that every detectable ΦV-1 gene was markedly downregulated in the presence of Upx (Fig. 6c), demonstrating that Upx effectively suppresses phage transcription after infection. This result is fully consistent with Upx-mediated degradation of incoming phage genomes, which prevents the initiation of viral gene expression. GO analysis of the downregulated genes further revealed that Upx blocks multiple cellular processes required for phage propagation (Fig. 6d). Pathways associated with the viral life cycle—including “viral process”, “viral life cycle”, “virus tail”, and other virion structural components—were among the most strongly suppressed. In addition, Upx reduced expression of structural and motility pathways such as bacterial-type flagellum assembly, cell projection organization, and periplasmic or membrane-associated components, all of which are known to be extensively remodeled during productive phage infection.
Together, these results show that Upx imposes a phage-restrictive state at both the DNA and RNA levels. Upx prevents effective amplification of the phage genome, leading to a marked reduction in viral DNA accumulation. In the meantime, Upx imposes a phage-restrictive transcriptional state: by destroying phage DNA, Upx eliminates viral transcripts at their source and prevents the activation of both viral gene expression programs and the host pathways that ΦV-1 normally exploits during its replication cycle.
Anti-phage function requires cooperation of NTD and CTD
To directly assess the contribution of each domain to phage defense, we compared the anti-phage activity of full-length Upx with truncation and point-mutant variants. Deletion of either the NTD or the CTD completely abolished phage resistance, demonstrating that both nuclease domains are indispensable for defensive function (Fig. 7a). Consistently, expression of individual domains or partial constructs failed to confer measurable protection, underscoring that the cooperative action of all domains is required (Fig. 7a). Furthermore, alanine substitutions of residues critical for catalysis within either the NTD (D127A/R129A) or the CTD (E1176A/K1178A) also eliminated phage resistance, confirming that the enzymatic activities of both domains are essential in vivo (Fig. 7a). We also introduced alanine substitutions at residues located at the MD–NTD or MD–CTD interfaces (e.g., R530A/E562A/R517A or R707A/R595A/R541A/N705A). Interestingly, none of these mutations impaired the anti-phage activity of Upx, highlighting an unexpected layer of complexity in its regulation (Fig. 7b). These findings establish that Upx-mediated defense relies on the coordinated contributions of the NTD and CTD, integrated through MD-dependent regulation. To elucidate the system-level defense strategy of Upx, we investigated whether it functions via an abortive-infection or direct immunity mechanism. Following the classification criteria described previously12, we performed phage-infection assays across a range of multiplicities of infection (MOIs). In E. coli strains overexpressing Upx, rapid bacterial growth was observed consistently at both low (MOI = 0.1) and high (MOI = 3) infection multiplicities (Fig. 7c). This growth pattern, where robust proliferation occurs regardless of phage dose, demonstrates that Upx provides direct immunity to infected cells rather than employing an abortive-infection strategy that sacrifices individual cells. In conclusion, Upx represents a previously uncharacterized dual-nuclease domain defense protein, functioning as a magnesium-dependent, non-sequence-specific 3′−5′ exonuclease active on ssDNA and ssRNA. The NTD emerges as a previously uncharacterized nuclease domain whose activity is autoinhibited by the MD, while the CTD displays robust nuclease activity that is potentiated by the MD. The ability of phage protein gp16 to relieve MD-mediated inhibition of the NTD highlights a dynamic regulatory mechanism that may be engaged during infection. Collectively, our study defines Upx as a previously unrecognized single-protein dual-nuclease defense system, expanding the functional and mechanistic repertoire of bacterial immunity (Fig. 7d).

a, b Antiphage activity of Upx and its mutant or truncated variants against phage infection. c Growth curves of E. coli MG1655 carrying either the control plasmid or Upx cloned into the pBAD LIC vector (8 A), assessed under uninfected conditions or following ΦV-1 infection at MOIs of 0.1 or 3. Data represent the mean ± s.d. of three independent biological replicates. Source data are provided as a Source Data file. d Proposed working model of the Upx-mediated antiphage defense system. In the absence of phage, Upx exhibits basal nuclease activity. Upon phage infection, the phage-encoded protein gp16 enhances Upx function, promoting the degradation of phage-derived single-stranded DNA intermediates, thereby blocking phage replication and conferring antiphage protection.
Discussion
Bacteria have developed a variety of complex and advanced mechanisms to protect themselves from viral infections, which are collectively referred to as the bacterial immune system23,24. While the PD-(D/E)XK superfamily represents a major class of these defense nucleases, they are typically single-domain enzymes23,24. Here, we report the identification and mechanistic characterization of Upx, a single-protein anti-phage system that integrates two distinct nuclease domains within a unified polypeptide. Our study reveals that Upx’s N-terminal domain (NTD) represents a previously unrecognized, minimalist variant of the PD-(D/E)XK fold with specific ssDNA exonuclease activity, while the C-terminal domain (CTD) functions as a canonical PD-(D/E)XK nuclease targeting both ssDNA and ssRNA. These two activities are structurally integrated and functionally regulated by a central middle domain (MD), which represses the NTD while enhancing CTD efficiency. This multi-domain architecture embodies a sophisticated regulatory strategy, enabling a single protein to exert potent anti-phage activity while minimizing collateral damage to the host, representing a significant expansion of the functional repertoire within this nuclease superfamily.
Our functional assays firmly establish that the cooperative action of both nuclease domains is essential for antiviral defense. In vivo plaque assays demonstrated that deletion of either the NTD or CTD, or the loss of catalytic activity in either domain, completely abolishes phage resistance. This unequivocally confirms that Upx operates as a bimodal nuclease system in a physiological context. The indispensability of both domains suggests a synergistic mechanism where the combined degradation of different nucleic acid substrates creates a formidable barrier against phage propagation. The observation that alanine substitutions at the MD-NTD or MD-CTD interfaces did not impair defense, despite their regulatory roles in vitro, hints at a robust and potentially redundant regulatory network in vivo that ensures functionality, a complexity that merits further investigation.
A central question was how an ssDNA/ssRNA-targeting nuclease could effectively defend against a dsDNA phage like ΦV-1. Our comprehensive substrate profiling provides a clear answer: Upx does not target the packaged dsDNA genome but rather preferentially cleaves transient single-stranded intermediates critical for phage replication and recombination. The enzyme shows a marked preference for structures like 3’ overhangs, replication forks, and most notably, D-loops, while its activity on R-loops is comparatively weaker. This substrate preference, coupled with our IP-MS and pull-down data identifying a direct interaction between Upx and the phage DNA polymerase, strongly supports a model wherein Upx is recruited to the sites of active phage genome replication. While our in vitro assays detected trace RNA cleavage activity, this activity is significantly weaker than DNA cleavage and lacks in vivo evidence for functional relevance during phage infection. Therefore, we consider DNA substrates—particularly replication intermediates like D-loops and 3’ overhangs—to be the primary physiological targets of Upx, with RNA cleavage likely representing a secondary biochemical property rather than a core defense mechanism. There, it disrupts the infection cycle by degrading essential ssDNA intermediates, effectively halting replication. This defines the physiological battlefield of Upx and moves beyond mere in vitro substrate characterization to propose a coherent model of its in vivo function. Beyond this direct nucleic acid targeting, our transcriptomic data reveal a broader anti-phage strategy: Upx expression globally suppresses the phage-induced transcriptional reprogramming of the host. This includes dampening the activation of host metabolic pathways and downregulating key viral processes. This dual mechanism—direct degradation of replication intermediates and disruption of essential phage-driven transcriptional responses—collectively undermines the infection cycle.
In conclusion, Upx exemplifies a sophisticated evolutionary solution for compact yet potent antiviral defense: the integration of dual nuclease activities into a single protein, governed by an autoinhibitory domain and activated by a specific phage trigger. Notably, gp16—a large multimeric phage structural protein involved in DNA injection and delivered early during phage entry—binds Upx and likely relieves MD-mediated autoinhibition, thereby activating the NTD specifically during early infection. This mechanism provides a clear rationale for how Upx detects incoming phage particles and triggers nuclease activation at the earliest stage of infection, which may underlie the narrow phage specificity of this defense system. Our findings redefine the functional landscape of PD-(D/E)XK nucleases by demonstrating that domain fusion and regulatory specialization can give rise to complex sensor-effector systems within minimal genetic footprints. The discovery of the minimalist NTD fold also expands the structural diversity of this superfamily, suggesting that such simplified yet functional nuclease domains may be more widespread in defense systems than previously appreciated.
Methods
Phylogenetic analysis of PD-D (E)XK nuclease
The phylogenetic analysis of the PD-(D/E)XK superfamily was performed utilizing a dataset comprising 83,737 protein sequences obtained from UniProt. In order to improve data integrity and minimize redundancy, proteins of eukaryotic origin or those with fewer than 100 amino acids were excluded from the analysis, and then de-duplication was carried out to eliminate sequences with more than 70% similarity. This process resulted in a final dataset consisting of 1000 representative sequences. Multiple sequence alignment was performed using the MAFFT (version 7)25,26, while phylogenetic reconstruction was executed using the IQ-TREE (v3.0.1)27, which incorporated standard model selection. The generated phylogenetic tree was visualized using FigTree software (v1.4.4).
Plasmid construction
The genes of interest were amplified utilizing PrimeSTAR® HS DNA Polymerase (Takara), employing primers that possess 5’ ends complementary to the ends of the plasmid backbone. The linear vector backbone was generated through PCR amplification or restriction enzyme digestion at multiple cloning sites (MCS) locations. The assembly of plasmids was conducted using the In-Fusion® HD Cloning Kit (Clontech). Upx and mutant constructs were assembled into the pLIB vector, respectively, with an N-terminal TEV cleavable His-ZZ-tag, for downstream purification via ZZ-tag affinity. Genes inserted into pLIB vector were checked via Sanger sequencing (Source Bioscience, Cambridge) before downstream steps.
Protein expression and purification
The genes of interest were successfully cloned into the pET-28-MKH8SUMO vector, which features an N-terminal 8×His-SUMO tag. The resultant plasmids were transformed into E. coli BL21 (DE3) cells to facilitate protein expression in LB medium. Protein expression was achieved by adding 0.2 mM IPTG at 16 °C overnight. After the protein expression induction period, the cells were harvested and then lysed via sonication in binding buffer (25 mM Tris–HCl, 500 mM NaCl, 5% glycerol, 5 mM imidazole, and, 2 mM β-mercaptoethanol, pH 8.0) on ice. After centrifugation, the supernatants were incubated with the Ni-NTA resin (Qiagen) for 1 h and subsequently washed thoroughly with washing buffer (25 mM Tris–HCl, 500 mM NaCl, 5% glycerol, 15 mM imidazole, and 2 mM β-mercaptoethanol, pH 8.0). The His-tagged target proteins were then eluted using an elution buffer (25 mM Tris–HCl, 300 mM imidazole, 500 mM NaCl, 5% glycerol, and 2 mM β-mercaptoethanol, at pH 8.0). The eluted samples were subsequently applied to a heparin column (GE Healthcare) and eluted using a linear NaCl gradient, followed by size-exclusion chromatography utilizing a Superdex 200 column (GE Healthcare), the buffer consisting of 25 mM Tris–HCl, 2 mM DTT, 150 mM NaCl, at pH 8.0. Protein peak fractions were collected and concentrated, the protein concentration were determined for subsequent experiments.
The N-terminal His-SUMO tag of Upx and truncation mutants protein can be cleaved by adding the TEV protease at the molar ratio of 1:10, and the tag can be removed by re-incubating with Ni-NTA resin. The resulting tag-free Upx and truncation mutant protein can then undergo further purified using HiTrap Heparin HP column (Cytiva) then Superdex 200 column (Cytiva). The protein samples were subsequently analyzed using 10% SDS-PAGE. The purified protein samples that are not immediately utilized would be stored at −80 °C.
Plaque assays and bacteriophage cultivation
The bacteriophages vB, Mu, SP, T6, T1, P1, and ФV-1, along with the host E. coli strain MG1655 (ATCC 25404), were ordered from the American Type Culture Collection (ATCC). The E. coli MG1655 strain was cultured in LB medium until OD600 reached 0.4. Subsequently, the bacteriophage was introduced into the host strain in a 1:2 ratio and incubated for 5–8 h. Upon observing a transition of the culture medium from turbidity to clarity. The lysate was subjected to centrifugation at 4696 × g for 15 min to eliminate cellular debris. Subsequently, the resultant supernatant was filtered using a 0.22 µm filter.
The phage titer was assessed by the small-drop plaque assay method28. For the plaque assays, freshly cultured E. coli MG1655 (OD₆₀₀ = 0.8) transformed with different constructs was mixed with semi-solid medium containing 50 mg/L ampicillin and 200 mg/L arabinose at a 1:100 (v/v) ratio. This mixture was then poured onto a 15 cm LB agar plate and allowed to stand for 1 h to become solid. Phage solutions ranging from 106 to 1011 PFU/mL were diluted with LB in a 10-fold series and then dropped onto a bilayer solid agar plate at 2.5 μL per drop and repeated in triplicate. The plates were allowed to dry for 30 min before being incubated for 12 h at 37 °C.
In vitro DNA/RNA cleavage assays
The DNA/RNA substrates were incubated with purified Upx at 37 °C for 30 min in a 20 μL DNA cleavage reaction buffer containing 25 mM Tris at pH 8.0, 2 mM DTT, 150 mM KCl, 2 mM MgCl2, and 5% Glycerol unless otherwise stated. After incubation, reactions were stopped with DNA loading buffer containing 60 mM EDTA and 5 μL was analyzed on a 13% Urea-PAGE gel, which was run at 240 V for 5 min followed by 150 V for 1 h. The gel was then analyzed on ChemiDoc MP Imaging System. To test activity in the presence of different divalent cations (Mg²⁺, Mn²⁺, Fe²⁺, Zn²⁺, Ni²⁺, Co²⁺, or Ce²⁺), Upx (or its truncations) was treated with 10 mM EDTA on ice for 30 min and then exchanged into metal‑free buffer using a desalting column. Cleavage reactions were carried out as described above, except that the reaction buffer was replaced with 20 mM Tris-HCl (pH 8.0) and 50 mM NaCl, supplemented with 0.2 mM of the indicated metal chloride or sulfate salt.
Cryo-EM sample preparation and data collection
For the Upx protein sample, 4 μL of 1 mg/mL was applied to a glow-discharged Quantifoil holey carbon grid (Au, R1.2/1.3, 300 mesh). The grids were subjected to blotting with a force of 2 for 2 seconds, after which they were rapidly plunged into liquid ethane using a Vitrobot Mark IV (Thermo Scientific). Cryo-EM grids were screened using a Talos Arctica microscope (ThermoScientific) operating at 200 kV, and the final map was collected on a Titan Krios G4 (FEI) operating at 300 kV. Images were captured through the Smart EPU software (v2.8.1) at a nominal magnification of 165, 000×, resulting in a calibrated physical pixel size of 0.692 Å/pixel, with a defocus range from −1.2 μm to −2.2 μm. Imaging was performed in super-resolution mode on a K3 summit electron direct detector, positioned at the output of a GIF-Quantum energy filter set to a slit width of 20 eV. Data were collected at a frame rate of 32 frames per second. The total electron dose was 50 e-/Å2. In total, 6,685 movie stacks were collected.
Cryo-EM image processing
The movie frames were processed using cryoSPARC (v4.7.1)29, aligned with MotionCor2 (v1.6.4)30 at a binning factor of 2, and underwent real-time contrast transfer function (CTF) estimation using Gctf (v1.06)31. A total of 1,025,867 particles were automatically identified using the topaz picking (v0.2.5)22, and 615,173 particles were retained for further analysis after 2D classification to eliminate poor-quality particles. Following the ab-initio reconstruction, a final set of 204, 860 particles was selected for non-uniform (NU) refinement, resulting in a resolution of 3.01 Å.
Model building and refinement
The atomic model of Upx, as predicted by AlphaFold3, was utilized as our initial template. We proceeded to segment this preliminary model according to its domains and subsequently docked these segments into the cryo-electron microscopy (cryo-EM) map using ChimeraX (v1.9)32,33. The comprehensive model was then subjected to manual refinement in Coot (v0.9.7)34. Following this, we performed iterative real-space refinements utilizing Phenix.real_space_refine (v1.20.1-4487)35. To evaluate the quality of the model, we utilizedMolProbity within the Phenix suite (v1.20.1-4487)36, and detailed refinement statistics are available in Table S1.
Electrophoretic mobility shift assay (EMSA)
The EMSA was conducted in a cleavage buffer with the addition of 10 mM EDTA, and incubated at 37 °C for 1 h. Purified Upx or different protein constructs were mixed with ssDNA in a 40:1 molar ratio. The ssDNA sequences utilized for the EMSA were nonspecific and labeled with 5’ Cy3. The reaction mixture was examined using a 6% native PAGE gel, which was run for 1 h at 120 V at 4 °C. The fluorescence signal was captured with the Tanon 5200 imaging system.
Co-immunoprecipitation coupled with mass spectrometry (Co-IP–MS)
There were three experimental groups: MBP (Blank control), MBP + ΦV-1 infected (control), and MBP–Upx + ΦV-1 infection, with two biological replicates per group. E. coli MG1655 cells overexpressing MBP or MBP–Upx were grown to an OD600 of 0.3and either infected with phage ΦV-1 or left uninfected as indicated. Cells were cultured for an additional 20 min following infection, harvested by centrifugation and lysed in Co-IP buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1 mM EDTA), supplemented with protease inhibitors. The lysates were incubated with MBP beads for 1 h at 4 °C, followed by extensive washing to remove nonspecific proteins.
Beads were washed twice with 50 mM NH₄HCO₃ and proteins were reduced with 5 mM dithiothreitol at 56 °C for 1 h, alkylated with 15 mM iodoacetamide at room temperature for 45 min in the dark, and quenched with 30 mM cysteine. Proteins were digested on beads with trypsin (Promega) overnight at 37 °C. Peptides were collected, dried, and desalted using μ-C18 ZipTips (Millipore).
Peptides were resuspended in 0.1% formic acid and analyzed by nanoLC–MS/MS using an EASY-nLC 1200 system coupled to an Orbitrap Q Exactive mass spectrometer (Thermo Fisher Scientific). Peptides were separated on a C18 column at 300 nL/min using a linear acetonitrile gradient. MS data were acquired in data-dependent mode with a Top15 method. Full MS scans were acquired at a resolution of 70,000 (m/z 350–1750), and MS/MS scans at a resolution of 17,500.
Raw data were processed with Proteome Discoverer (v3.0) and searched against the UniProtKB/Swiss-Prot phage ΦV-1proteome (November 6, 2023). Trypsin specificity with up to two missed cleavages was used. Carbamidomethylation of cysteine was set as a fixed modification, and methionine oxidation and N-terminal acetylation as variable modifications. Protein identifications were filtered at an FDR ≤ 0.01. Label-free quantification was performed using MS1 intensities, requiring at least two peptides per protein. Proteins with a fold change >2 and p < 0.01 were considered significantly altered. Three biological replicates were analyzed per condition.
Sample preparation for RNA sequencing
E. coli MG1655 cultures were grown in LB at 37 °C with shaking (200 rpm) to an OD₆₀₀ of 0.5 and subsequently infected with Escherichia phage phiV-1 at an MOI of 3. After infected for 20 min, culture samples were rapidly quenched and collected by centrifugation at 4 °C, and stored at −80 °C. Total RNA was isolated using the TRIzol methods, including chloroform phase separation, isopropanol precipitation, ethanol washing, and TURBO DNase treatment. RNA quality (RIN > 6.5) was identified prior to bacterial rRNA depletion and preparation of strand-specific sequencing libraries. Libraries were sequenced on an Illumina platform, and trimmed reads were aligned to a combined E. coli MG1655 + phiV-1 reference genome for gene-level quantification and differential expression analysis.
Bioinformatic analysis for RNA-seq data
Differential expression analysis used the DESeq2 R package (v1.34.0), a model based on the negative binomial distribution. After adjusted by Benjamini and Hochberg’s approach for controlling the false discovery rate, Padj of genes were setted <0.05 to detect differential expressed ones. Significantly differentially expressed genes (DEGs) were defined as if the genes met the criteria of Padj <0.05 and absolute log2 fold change ( | log2FC | ) > 1.
The genome sequences of Escherichia phage phiV-1 were aligned against the Swiss-Prot database, and the corresponding GO terms were then obtained based on the mapping relationship between gene IDs and GO IDs. We merged the gene ID-to-GO ID mappings of Escherichia coli and Escherichia phage phiV-1 to facilitate the subsequent construction of a customized OrgDb package. AnnotationForge R package (v1.36.0) was used building customized SQLite-based annotation OrgDb package. ClusterProfiler (v4.2.2) R package was used identifying Gene Ontology (GO) terms that annotate a list of significantly DEGs. Significantly enriched terms were filtered to retain those with q-value < 0.05.
Sample preparation for DNA-seq data (WGS data)
E. coli MG1655 cultures harboring the indicated plasmids were grown in LB medium supplemented with 0.2% (w/v) L‑arabinose to an OD600 of 0.5 and then infected with Escherichia phage ΦV‑1 at a MOI of 3. After 5 or 20 min of infection, cultures were rapidly chilled on ice, harvested by centrifugation at 4 °C, and stored at ‑80 °C. Total genomic DNA was extracted using the Bacterial Genomic DNA Extraction Kit (DP302, TIANGEN). DNA concentration and purity were measured with a NanoDrop 2000 spectrophotometer.
Library construction and sequencing
For DNA sequencing, genomic DNA (200 ng) was fragmented to ~300–350 bp using a Covaris ultrasonicator. Fragmented DNA was end‑repaired, 5’‑phosphorylated, 3’‑A‑tailed, and ligated to sequencing adaptors. Libraries were size‑selected using DNA cleanup beads and amplified by PCR for eight cycles. Library quality was assessed on an Agilent 2100 Bioanalyzer. Qualified libraries were sequenced in paired‑end mode (PE150) on Illumina platforms (Novaseq6000).
Bioinformatic processing
Raw sequencing reads were processed with fastp (v0.23.0) to remove adaptor sequences, PCR primers, reads containing >14 ambiguous bases (N), and low‑quality reads with >40% of bases below Q20. Clean reads were aligned to a combined reference genome consisting of the E. coli MG1655 chromosome (GenBank accession: U00096.3) and the complete phage ΦV‑1 genome (NC_001426.1) using the Sentieon pipeline (v202112.02). Duplicate reads were removed, and single‑nucleotide variants (SNVs) and insertions/deletions (InDels) were called using Sentieon’s variant‑calling modules.
Read‑depth (RD) analysis and normalization
To determine the relative abundance of phage DNA, we performed read‑depth (RD) analysis following principles established for copy‑number variation (CNV) detection37. The analysis was carried out with CNVnator (v0.3.1) using a bin size of 100 bp. A sample‑specific genomic baseline was established by analyzing the global distribution of RD across the entire combined reference genome. The RD for each genomic segment was then normalized against this internal baseline. Phage genome abundance was quantified as the mean normalized RD across all bins spanning the combined reference genome. This value represents the relative copy number of phage DNA per host‑genome equivalent, where a value of 1.0 corresponds to one phage genome per bacterial genome. The normalized RD values reflect the relative ratio of phage‑to‑host DNA in each sample.
Growth curve assays
Overnight bacterial cultures were diluted 1:100 in LB and grown to an OD₆₀₀ of 0.3. Subsequently, 180 μl of each culture was dispensed into a 96-well plate preloaded with 20 μl of either LB (uninfected control) or phage lysate to achieve final MOIs of 3 or 0.1. Plates were incubated at 25 °C with shaking in microplate reader, and OD₆₀₀ readings were recorded every 15 min. Infections were set up in three independent biological replicates. Three wells containing only medium served as blanks, and their readings were subtracted from all sample OD600 measurements.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The atomic coordinates data generated in this study have been deposited in the Protein Data Bank under accession code 9WUF, and the electron microscopy map has been deposited in the Electron Microscopy Data Bank under accession code EMD-66260. The RNA sequencing data have been deposited to Gene Expression Omnibus (GEO) database under accession code GSE311613 (RNA-seq profiling of E. coli and their overexpression derivatives (UPX overexpressed) infected with Escherichia phage ΦV-1. Samples were collected after infected for 20 min). The whole-genome resequencing data of E. coli and phage samples generated in this study have been deposited in the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) database. Corresponding BioSample accessions are SAMN53654015 (Control groups at 5 min after ΦV-1 infection), SAMN53654016 (Control groups at 20 min after ΦV-1 infection), SAMN53654017 (UPX mutation groups at 5 min after ΦV-1 infection), SAMN53654018 (UPX mutation groups at 20 min after ΦV-1 infection), SAMN53654019 (UPX wild-type groups at 5 min after ΦV-1 infection), and SAMN53654020 (UPX wild-type groups at 20 min after ΦV-1 infection). The sequence data submissions are linked to these BioSample accessions, and all data can be accessed via the NCBI SRA portal (https://www.ncbi.nlm.nih.gov/sra) or the associated BioProject page (accession number: PRJNA1370017 [https://www.ncbi.nlm.nih.gov/bioproject/ PRJNA1370017]). Source data are provided with this paper.
References
Yirmiya, E. et al. Phages overcome bacterial immunity via diverse anti-defence proteins. Nature 625, 352–359 (2024).
Antine, S. P. et al. Structural basis of Gabija anti-phage defence and viral immune evasion. Nature 625, 360–365 (2024).
Stokar-Avihail, A. et al. Discovery of phage determinants that confer sensitivity to bacterial immune systems. Cell 186, 1863–1876 e16 (2023).
Tesson, F. et al. A comprehensive resource for exploring antiphage defense: defensefinder webservice, wiki and databases. Peer Community J. 4, e91 (2024).
Georjon, H. & Bernheim, A. The highly diverse antiphage defence systems of bacteria. Nat. Rev. Microbiol. 21, 686–700 (2023).
Millman, A. et al. An expanded arsenal of immune systems that protect bacteria from phages. Cell Host Microbe 30, 1556–1569 e5 (2022).
Smith, W. P. J. et al. Bacterial defences: mechanisms, evolution and antimicrobial resistance. Nat. Rev. Microbiol 21, 519–534 (2023).
Tock, M. R. & Dryden, D. T. The biology of restriction and anti-restriction. Curr. Opin. Microbiol. 8, 466–472 (2005).
Marraffini, L. A. CRISPR-Cas immunity in prokaryotes. Nature 526, 55–61 (2015).
Swarts, D. C. et al. DNA-guided DNA interference by a prokaryotic Argonaute. Nature 507, 258–261 (2014).
Gao, L. et al. Diverse enzymatic activities mediate antiviral immunity in prokaryotes. Science 369, 1077–1084 (2020).
Vassallo, C. N. et al. A functional selection reveals previously undetected anti-phage defence systems in the E. coli pangenome. Nat. Microbiol. 7, 1568–1579 (2022).
Knizewski, L. et al. Realm of PD-(D/E)XK nuclease superfamily revisited: detection of novel families with modified transitive meta profile searches. BMC Struct. Biol. 7, 40 (2007).
Steczkiewicz, K. et al. Sequence, structure and functional diversity of PD-(D/E)XK phosphodiesterase superfamily. Nucleic Acids Res. 40, 7016–7045 (2012).
Yang, X. Y. et al. DdmDE eliminates plasmid invasion by DNA-guided DNA targeting. Cell 187, 5253–5266 e16 (2024).
Dai, G. et al. Molecular characterization of a new abortive infection system (AbiU) from Lactococcus lactis LL51-1. Appl. Environ. Microbiol. 67, 5225–5232 (2001).
Rostol, J. T. et al. The Card1 nuclease provides defence during type III CRISPR immunity. Nature 590, 624–629 (2021).
Loeff, L. et al. DNA end sensing and cleavage by the Shedu anti-phage defense system. Cell 188, 721–733 e17 (2025).
Tuck, O. T. et al. Genome integrity sensing by the broad-spectrum Hachiman antiphage defense complex. Cell 187, 6914–6928 e20 (2024).
Yip, K. M. et al. Atomic-resolution protein structure determination by cryo-EM. Nature 587, 157–161 (2020).
Gu, Y. et al. Functional characterization of Mrr-family nuclease SLL1429 involved in MMC and phage resistance. Microbiol. Res. 296, 128123 (2025).
Bepler, T. et al. Positive-unlabeled convolutional neural networks for particle picking in cryo-electron micrographs. Nat. Methods 16, 1153–1160 (2019).
Finlay, B. B. & McFadden, G. Anti-immunology: evasion of the host immune system by bacterial and viral pathogens. Cell 124, 767–782 (2006).
Koonin, E. V., Makarova, K. S. & Wolf, Y. I. Evolutionary genomics of defense systems in archaea and bacteria. Annu. Rev. Microbiol. 71, 233–261 (2017).
Katoh, K., Rozewicki, J. & Yamada, K. D. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 20, 1160–1166 (2019).
Kuraku, S. et al. aLeaves facilitates on-demand exploration of metazoan gene family trees on MAFFT sequence alignment server with enhanced interactivity. Nucleic Acids Res. 41, W22–W28 (2013).
Minh, B. Q. et al. IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 37, 1530–1534 (2020).
Mazzocco, A. et al. Enumeration of bacteriophages using the small drop plaque assay system. Methods Mol. Biol. 501, 81–85 (2009).
Punjani, A. et al. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290 (2017).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Zhang, K. Gctf: Real-time CTF determination and correction. J. Struct. Biol. 193, 1–12 (2016).
Goddard, T. D. et al. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. 27, 14–25 (2018).
Meng, E. C. et al. UCSF ChimeraX: Tools for structure building and analysis. Protein Sci. 32, e4792 (2023).
Casanal, A., Lohkamp, B. & Emsley, P. Current developments in Coot for macromolecular model building of electron cryo-microscopy and crystallographic data. Protein Sci. 29, 1069–1078 (2020).
Afonine, P. V. et al. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr D. Struct. Biol. 74, 531–544 (2018).
Williams, C. J. et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 27, 293–315 (2018).
Abyzov, A. et al. CNVnator: an approach to discover, genotype, and characterize typical and atypical CNVs from family and population genome sequencing. Genome Res. 21, 974–984 (2011).
Acknowledgements
We thank the staff members from the Cryo-Electron Microscopy Facility of Hubei University for their help with data collection and computation. This work was supported by National Natural Science Foundation of China (32201010, 22577133 to Y.W.), and National Key R&D Program of China (2022YFC3400400 to Y.W.).
Author information
Authors and Affiliations
Contributions
S.Z. (Shengkai Zuo), H.Y. and Y.W. designed the research. R.Z., Y.L., Q.Z., C.Z., L.Z., Y.Z., W.C., X.L., Z.L. (Zhikun Liu), and Z.Y. carried out the experimental work. Data analysis was performed by R.Z., X.L., S.Z. (Shuqin Zhang), N.J., H.Z., Z.L. (Zhuang Li), H.Y., J.T., and Y.W.; R.Z., H.Y and Y.W. wrote the paper with help from Y.L. and H.Z.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Matthias Bochtler, James Eaglesham, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhou, R., Liu, Y., Zhang, Q. et al. Structural and mechanistic insights into the dual-nuclease defense protein Upx as an anti-phage system. Nat Commun 17, 3692 (2026). https://doi.org/10.1038/s41467-026-70435-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-70435-x
