
Abstract
Tissue-resident stem cells play an essential role in repairing barrier tissues subjected to frequent insults. However, the local cues that coordinate successful barrier repair or lead to tissue remodeling are largely unknown. Here we use murine models of airway injury, fate mapping, and null strains to identify a role for rare tuft epithelial cells in signaling to submucosal stem cells through the generation of cysteinyl leukotrienes (CysLTs) and activation of the CysLT receptor OXGR1. This results in mobilization of SOX9+ submucosal gland progenitors, aberrant repair of the surface airway epithelium, and durable features of airway remodeling including submucosal gland hyperplasia and collagen deposition. Remarkably, selective deletion of SOX9 from the airway stem compartment allows epithelial restoration and prevents tissue remodeling. These findings demonstrate a tuft cell- OXGR1- and SOX9- circuit that remodels the airway after injury and is detected in the human sinus mucosa.
Similar content being viewed by others

Introduction
Damage to epithelial barrier tissues is associated with the rapid mobilization of tissue-resident stem cells to aid in repair. In the airway, basal cells (BCs) are the dominant progenitor of differentiated ciliated, secretory, and specialized epithelial cells (EpCs)1,2, although significant epithelial damage can elicit the de-differentiation of secretory EpCs to acquire progenitor capacity3,4, and/or the mobilization of accessory progenitor cells from submucosal glands (SMGs)5,6. In chronic airway diseases such as asthma, disruption of the epithelial barrier is associated with goblet cell metaplasia in the epithelium and with remodeling in the submucosa that includes the deposition of extracellular matrix proteins and hypertrophy of submucosal glands7,8,9. Despite the high frequency with which these features are detected, the pathways that prevent normal repair and lead to chronically remodeled tissue are poorly understood.
Tuft cells are chemosensory EpCs that serve a sentinel role, recognizing diverse microbes including helminths, protists, bacteria, and fungi10,11,12,13,14. On activation, tuft cells rapidly generate pro-inflammatory mediators including IL-2510,11,12, acetylcholine (Ach)14,15,16, cysteinyl leukotrienes (CysLTs)13,17,18, prostaglandin D219 and prostaglandin E220. Tuft cell mediators activate type 2 innate lymphoid cells (ILC2s) to generate IL-13 and facilitate goblet cell metaplasia (GCM) and mucus release for the elimination of intestinal pathogens10,11,12,18. In the airway, tuft cells are also activated in response to common protease-containing aeroallergens such as house dust mite and the fungus Alternaria alternata (ALT). Here they elicit pathologic type 2 inflammation through their generation of IL-2521 and CysLTs17,22. Tuft cell recognition of allergens happens, at least in part, indirectly through the damage-associated release of ATP and the activation of P2Y213. Thus, tuft cells are also sentinel detectors of tissue damage.
Tuft cells are rare EpCs, but their numbers increase in the setting of inflammation, injury, and tissue repair. Tuft cell hyperplasia is detected in the sinonasal mucosa of patients with chronic rhinosinusitis with nasal polyposis (CRSwNP)20,23, in covid-infected human lung24, in murine models of influenza- or bleomycin-elicited lung damage25,26,27, and in allergen-elicited airway damage13,21. As such, several groups have speculated that they may have a role in tissue repair or in the dysplastic remodeling seen in those settings, but thus far no reparative function has been demonstrated, and no mechanism by which tuft cells might promote dysplasia has been identified.
Here we show that rare tuft epithelial cells orchestrate airway repair by engaging submucosal gland stem cells through a CysLT–OXGR1 signaling axis. This interaction drives the activation of SOX9⁺ glandular progenitors, leading to defective restoration of the surface airway epithelium (SAE) and persistent features of airway remodeling, including glandular hyperplasia and extracellular matrix (ECM) deposition. Disruption of SOX9 within the airway stem cell compartment restores epithelial integrity and prevents pathologic tissue remodeling. Together, these findings define a tuft cell–OXGR1–SOX9 regulatory circuit that governs injury-induced airway remodeling and is conserved in the human sinonasal mucosa.
Results
Tuft cells are associated with chronic injury in the SAE
To better understand whether tuft cells contribute to tissue repair or remodeling, we exploited a murine model of airway injury and repair elicited by repetitive intranasal exposure to Alternaria (ALT)28, a common protease-containing mold aeroallergen21. As expected, ALT inhalation elicited acute epithelial damage with sloughing of the apical epithelium in both WT and tuft cell-deficient Pou2f3-/- mice (Supplementary Fig. 1a, d and Fig. 1a–d). Epithelial DNA damage (assessed by p-γ-H2A.X) and epithelial apoptosis (assessed by TUNEL staining) was detected in both WT and Pou2f3-/- mice (Supplementary Fig. 1e). With repetitive ALT challenge, both WT and Pou2f3-/- mice demonstrated repair of the surface airway epithelium (SAE) with the appearance of a multilayered epithelial barrier at day 14 (Fig. 1c–e), and WT mice developed tuft cell hyperplasia over the course of 14 days that was absent, as expected, in Pou2f3-/- mice (Supplementary Fig. 2a–e). However, further assessment of the epithelium at day 14 demonstrated that WT mice had evidence of ongoing injury with DNA damage (assessed by p-γ-H2A.X) and epithelial apoptosis (assessed by TUNEL staining) that was remarkably reduced in Pou2f3-/- mice (Fig. 1e, f). Moreover, the epithelium of ALT-challenged WT mice had increased epithelial permeability compared to Pou2f3-/- mice, with increased FITC-Dextran detected in the submucosa (Fig. 1g, h and Supplementary Fig. 3a). By contrast, epithelial integrity, as assessed by expression of the tight junction protein claudin-1 (CLDN1) was robustly detected in the regenerating epithelium of ALT-challenged Pou2f3-/- mice, compared to ALT-challenged WT (Fig. 1i, j and Supplementary Fig. 3b). Taken together, these data indicate that tuft cells prevent optimal barrier repair.

a Experimental schema. b Representative images of H&E staining in tracheal sections from the indicated strains at day 1. c, d Quantification of DAPI+ cells (c) and epithelial height (d) from the WT and Pou2f3-/- at the indicated time points. Group sizes: n = 3, 4, 12, 11, 6, 7 mice, from left to right. Two-tailed unpaired Mann-Whitney U test. e Representative images of TUNEL or p-γ-H2A.X staining in tracheal sections from the indicated strains at day 14. f Quantification of TUNEL+ (top panel) or p-γ-H2A.X+ (bottom panel) cells from the WT and Pou2f3-/- at the indicated time points. Group sizes: n = 3, 4, 12, 11, 6, 7 mice, from left to right. Kruskal–Wallis test followed by Dunn’s multiple-comparisons test with Holm–Bonferroni correction. g, h Representative images (g) and quantification (h) of FITC-dextran in tracheal sections from the WT and Pou2f3-/- at day 11. Group sizes: n = 6, 7, 6 mice, from left to right. Two-tailed unpaired Mann-Whitney U test. i, j Representative images (i) and quantification (j) of CLDN1 staining in tracheal sections from the WT and Pou2f3-/- at day 14. Group sizes: n = 6, 7, 6 mice, from left to right. Two-tailed unpaired Mann-Whitney U test. k Experimental schema. l–n Representative images (l), and quantification of TUNEL+ (m) and p-γ-H2A.X+ (n) from the indicated strains. Group sizes: n = 4, 7 mice, from left to right. Two-tailed unpaired Mann-Whitney U test. Scale bars: 50 μm (b, e, g, i, l). The dashed line marks the border between surface airway epithelium (SAE) and submucosa (SM). Box plots (c, d, f, h, j, m, n) show median (center line), 25th–75th percentiles (box), and min–max values (whiskers). Each data point represents the average of 3–4 images from one mouse, except FITC intensity (h), which represents total intensity per trachea measured by tiled imaging. See “Methods”. Panels a and k were created in BioRender. Lee, M. (https://BioRender.com/dg3i7rm). Source data are provided as a Source Data file.
Tuft cells secrete several products including pro-inflammatory CysLTs (LTC4, LTD4, LTE4). To understand whether tuft cell-derived CysLTs contribute to the poor epithelial repair observed in WT mice, we deleted the CysLT biosynthetic enzyme LTC4S in tuft cells (Supplementary Fig. 4a, b) and subjected them to the same protocol (Fig. 1k). Epithelial damage was reduced in Pou2f3CreLtc4sfl/fl mice, as compared to controls (Fig. 1l-n), demonstrating a role for tuft cell-derived CysLTs in preventing productive airway barrier repair.
Tuft cells promote airway remodeling and regeneration of the SAE from submucosal gland (SMG) progenitors
Histochemical staining of the trachea in ALT-challenged WT mice demonstrated that airway repair was accompanied by an increase in subepithelial collagen deposition, as compared to PBS controls (Fig. 2a and Supplementary Fig. 5a, b). This deposition was reduced in ALT-challenged mice deficient in either tuft cells or tuft cell-derived CysLTs (Fig. 2a, b). Similarly, submucosal glands (SMGs), typically found only around the proximal C1 cartilage in naïve mice were increased in number and detected down to the C5 cartilage in ALT-challenged WT mice, but SMG hyperplasia was reduced in ALT-challenged mice deficient in either tuft cells or tuft cell-derived CysLTs (Fig. 2a, c). To understand whether these marked changes persist, we challenged mice and then rested them for an additional month before analysis. While epithelial damage detected in ALT-challenged WT mice resolved over 1 month of rest, approaching the levels seen in ALT-challenged Pou2f3-/- mice (Supplementary Fig. 5c–e), SMG hyperplasia and collagen deposition were not reduced (Fig. 2d, e and Supplementary Fig. 5f), demonstrating that tuft cells orchestrate airway remodeling, producing durable changes in tissue architecture.

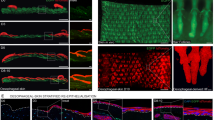
a Representative Masson’s Trichrome staining in tracheal sections at day 14. Scale bars: 200 μm. Cartilage rings are labelled C1–C6. b–e Quantification of collagen deposition over WT PBS d14 (b) or over WT ALT d14 (d), and gland numbers from C1-C7 cartilages (c, e). Group sizes: n = 7, 7, 5, 4, 7 (b, c) and n = 4, 5, 3 (d, e) mice, from left to right. f Experimental schema. g Representative images of lineage-labelled Sox9-tdT+ cells among BCAM+ BCs on the SAE at day 14. Scale bar: 200 μm. Areas outlined (a, b) are shown below (Scale bars: 50 μm). Cartilage rings are labelled C1–C7. CC Cricoid cartilage. h, i Quantification of Sox9-tdT+BCAM+ cells in the SAE per high power field (HPF) (h) and as a percentage of total BCAM+ cells (i) in ALT-injured trachea at day 14. Group sizes: n = 7, 15, 6 mice, from left to right. j, k Quantification of Sox9-tdT+BCAM+ cells in the SAE per HPF (j) and as a percentage of total BCAM+ cells (k) in influenza-injured trachea at 7 days post infection. Group sizes: n = 3, 6, 8 mice, from left to right. l, m Representative images (l) and quantification (m) of BCAM and SOX9 staining in tracheal sections at day 14. Scale bars: 20 μm. Group sizes: n = 5, 6, 4 mice, from left to right. Two-tailed unpaired Mann-Whitney U test (b–e, m). Kruskal–Wallis test followed by Dunn’s multiple-comparisons test with Holm–Bonferroni correction (h–k). The dashed outline (g, i) marks the border between SAE and SM. Box plots (b–e, h–k, m) show median (center line), 25th–75th percentiles (box), and min–max values (whiskers). Each data point represents the average of 3–4 images from one mouse, except for the glandular number (c, e), which represents the total glands per trachea measured by tiled imaging. See “Methods”. Panel f was created in BioRender. Lee, M. (https://BioRender.com/dg3i7rm). Source data are provided as a Source Data file.
Prior studies have demonstrated that SMG stem cells, marked by expression of the pioneer transcription factor SOX9, act as accessory stem cells, repairing the SAE after injury6,29. Given our findings for tuft cells driving SMG hyperplasia, we next asked whether tuft cells control SOX9+ SMG stem cell-dependent repair of the SAE. To assess this, we crossed Sox9 CreERT2R26tdT mice to Pou2f3-/- mice, or littermate controls, and performed fate-labelling experiments (Fig. 2f). In naïve trachea, rare expression of lineage-traced Sox9-tdT+ cells were detected in the SAE, and high numbers of Sox9-tdT+ cells were detected in the cartilage and SMGs, as previously reported6,30 (Supplementary Fig. 6a, b). In vehicle-challenged WT mice, low numbers of Sox9-tdT+ cells were again detected in the SAE (Supplementary Fig. 6c). However, in ALT-challenged WT mice, lineage-traced Sox9-tdT+ cells were readily detected in the SAE and frequently expressed basal cell adhesion molecule (BCAM) (Fig. 2g), identifying them as BCs. In total, 16.3 ± 3.9% of all BCs were derived from Sox9-tdT SMG progenitors (Fig. 2g–i). This regeneration of the SAE stem cell compartment from SMG progenitors was significantly reduced in ALT-challenged Pou2f3-/- mice (Fig. 2g–i and Supplementary Fig. 6d). As expected, newly arriving Sox9-tdT SMG progenitors on the SAE were largely protected from damage with few lineage-traced cells expressing p-γ-H2A.X or exhibiting TUNEL staining (Supplementary Fig. 7a–d). To understand whether reduced SMG progenitor mobilization in the Pou2f3-/- mice might reflect a direct effect of Pou2f3 function in SMG progenitors, we turned to a Pou2f3 reporter strain. Assessment of Pou2f3 expression in both vehicle- and ALT-challenged Pou2f3CreERT2R26tdT mice demonstrated no expression in SMGs (Supplementary Fig. 8a, b). These results indicate that the decreased number of lineage-traced Sox9-tdT+ cells observed in the SAE of Pou2f3-/- mice is not likely to result from a loss of Pou2f3 function in SMG progenitors, but rather results from the loss of tuft cells on the SAE.
Alternaria is rich in serine protease activity, which contributes airway inflammation31,32,33. Accordingly, we asked whether exposure to heat-inactivated allergen, which lacks protease activity, is sufficient to promote a Sox9-tdT SMG progenitor cell mobilization. With repeated injections of heat-inactivated ALT over 2 weeks, no Sox9⁺ progenitors were observed in the SAE, although some progenitors were observed in the submucosa and several Sox9⁺ glands were identified beneath the C1 cartilage (Supplemental Fig. 9a). After a single injection of heat-inactivated ALT, there was no evidence of SAE injury (Supplementary Fig. 9b), suggesting that protease activity is required for the full phenotype.
To understand whether tuft cells direct SMG progenitors in response to diverse environmental insults, we used a previously reported model of mouse adapted H1N1 influenza that elicits tracheal damage and repair6 (Supplementary Fig. 10a). After lineage labeling Sox9-tdT+ SMG progenitors and influenza infection, we again saw regeneration of the SAE stem cell compartment from Sox9-tdT+ SMG progenitors (Supplementary Fig. 10b). In WT mice 17.4 ± 3.9 % of all BCs were derived from Sox9-tdT+ SMG progenitors, and this was again significantly reduced in Pou2f3-/- mice (Fig. 2j, k and Supplementary Fig. 10b–d).
Finally, we assessed SOX9 protein expression on the SAE in mice with tuft cell-specific deletion of LTC4S. Here we found that SOX9 expression in BCAM+BCs on the SAE was reduced in ALT-challenged Pou2f3CreLtc4sfl/fl mice (Fig. 2l, m), compared to littermate controls, indicating that tuft cell generation of CysLTs promotes regeneration of the SAE stem cell compartment from this accessory SMG stem cell.
OXGR1 on SMG progenitors drives their regeneration of the SAE
Having established a role for CysLTs in the mobilization of SMG progenitors, we next asked which, if any, of the established CysLT receptors may mediate this and performed Alternaria challenge in mice with targeted deletions of CysLT1, CysLT2, or OXGR1 (also called GPR99 and CysLT3). This demonstrated that only OXGR1-deficient (Oxgr1-/-) mice had reduced SOX9 staining on the SAE (Supplementary Fig. 11a, b). To confirm this finding, we crossed Sox9CreERT2R26tdT mice to the Oxgr1-/- background. After tamoxifen treatment and ALT-challenge, lineage-labelled Sox9-tdT+ cells appeared on the SAE of WT mice but were reduced in Oxgr1-/- mice (Fig. 3a, b). We then looked at features of airway remodeling in ALT-challenged Oxgr1-/- mice, finding that Oxgr1-/- mice were not protected from acute epithelial injury (Supplementary Fig. 12a–e), but they were protected from persistent epithelial damage (Supplementary Fig. 12f) and submucosal remodeling including subepithelial collagen formation and SMG hyperplasia (Supplementary Fig. 12g, h), similar to Pou2f3-/- mice. To understand whether OXGR1 controls SMG stem cell repair of the SAE in other settings, Sox9 CreERT2R26tdT mice on the WT and Oxgr1-/- background were treated with tamoxifen and then infected with influenza (Supplementary Fig. 12i–k). Similar to the Pou2f3-/- strain, Oxgr1-/- mice had reduced regeneration of the SAE stem cell compartment from fate-labelled Sox9-tdT+ SMG stem cells. These results demonstrate that tuft cells elicit the activation of OXGR1-expressing cells to direct regeneration of the SAE and remodeling of the submucosa.

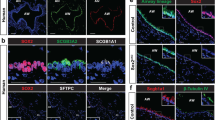
a Representative images of DCLK1 and BCAM staining in tracheal sections from the indicated Sox9-tdT reporter strains at day 14. Sox9-tdT+BCAM+ cells (white arrow). Scale bars: 50 µm. b Quantification of Sox9-tdT+BCAM+ cells in the SAE from the indicated strains at day 14. Group sizes: n = 5, 6, 12, 5 mice, from left to right. Two-tailed unpaired Mann-Whitney U test. c, d Combined in situ hybridization (ISH) for Oxgr1, Krt5 and Krt8, and immunofluorescence for SOX9 and α-SMA in tracheal sections from WT mice with or without ALT treatment (Krt5 for BCs, Krt8 for differentiated cells, SOX9 for SMG progenitor cells, and α-SMA for SMG myoepithelial cells). Oxgr1+SOX9+ cells (green arrows). Scale bars: 20 μm. SMG Submucosal gland. (e) Quantification of Oxgr1+ cells in the specified EpC subsets. Group sizes: n = 7, 16, 8, 13, 6, 10, 7, 8 mice, from left to right. Kruskal–Wallis test followed by Dunn’s multiple-comparisons test with Holm–Bonferroni correction. f Quantification of Oxgr1+ cells among the Krt5+ or Krt8+ cells. Group sizes: n = 12, 11, 12, 7 mice, from left to right. Two-tailed unpaired Mann-Whitney U test. g Experimental schema. h, i Representative images (h) and quantification (i) of BCAM and SOX9 staining in tracheal sections from the indicated strains at day 14. Scale bars: 20 μm. Group sizes: n = 5, 8, 7, 5 mice, from left to right. Kruskal–Wallis test followed by Dunn’s multiple-comparisons test with Holm–Bonferroni correction. The dashed line marks the border between SAE and SM. Box plots (b, e, f, i) show median (center line), 25th–75th percentiles (box), and min–max values (whiskers). Each data point represents the average of 3–4 images from one mouse. See “Methods”. Panel g was created in BioRender. Lee, M. (https://BioRender.com/dg3i7rm). Source data are provided as a Source Data file.
OXGR1 expression has been poorly understood21,34,35. RNA probes to assess Oxgr1 (Supplementary Fig. 13a, b) demonstrated expression in Krt5+ BCs in the basal layers of the SAE, Krt8+ BCs in the apical layers of the SAE, and in SOX9+ progenitor cells in the SMG of naïve mice (Fig. 3c–e). There was little detection in other EpC subsets (Fig. 3e and Supplementary Fig. 13c), but some detection in endothelial cells (Supplementary Fig. 13c). With ALT delivery, there was an increase in Oxgr1 expression in Krt5+ BCs (Fig. 3d, f), but no increase in Oxgr1 expression in either Krt8+ cells (Fig. 3f) or in SMG cells (Supplementary Fig. 13d, e).
To understand how OXGR1 on these cells contributes to regeneration of the SAE stem cell compartment, we generated Oxgr1fl/fl mice and crossed them to KRT5CreERT2 (targeting both BCs and SMG cells) and to Sox9CreERT2 strains (targeting SMG progenitors). Additionally, because SOX9 expression has also been reported in tuft cells36, we generated Pou2f3CreERT2Oxgr1fl/fl for a tuft cell-specific Oxgr1 deletion. ALT-challenged mice (Fig. 3g) had an Oxgr1 deletion efficiency of ~80% in each null strain, as compared to controls (Supplementary Fig. 14a–c). Consistent with our lineage tracing experiments in the Oxgr1-/- strain, deletion of OXGR1 specifically on SMG progenitors reduced expression of SOX9 on the SAE (Fig. 3g–i). Deletion of Oxgr1 from both SOX9+ SMG progenitors and SAE BCs (Krt5CreERT2Oxgr1fl/fl mice) had no additional decrement, and deletion of Oxgr1 from tuft cells (Pou2f3CreERT2Oxgr1fl/fl) had no effect on SOX9 expression in the SAE (Fig. 3g–i). Taken together, these results demonstrate that tuft cells signal to SOX9+ SMG stem cells to drive their regeneration of the SAE.
Deletion of Sox9 restores ciliary differentiation and prevents airway remodeling
To understand the distinctions between BCs derived from SOX9+ SMG progenitors (SOX9+ BCs) and conventional SOX9- BCs, we sorted these populations from lineage-traced Sox9-tdT reporter mice after ALT-challenge and performed low-input bulk RNA-seq (Fig. 4a and Supplementary Fig. 15a). Principal component analysis (PCA) demonstrated that genotype corresponded to PC1 (Fig. 4b). Dot plots showed high relative expression of Sox9 in lineage-labelled Sox9-tdT+ BCs (Fig. 4c), validating the sorting. There were 1443 differentially expressed genes (DEGs; padj<0.05 and |log2FoldChange | >0.58) between the groups, 702 genes with increased expression in SOX9+BCs and 741 genes with increased expression in conventional BCs (Fig. 4d and Supplementary Dataset 1). Over-representation analysis (ORA) of DEGs with increased expression in SOX9+ BCs revealed an increase in Hallmark pathway genes for TNFA SIGNALING VIA NFKB and EPITHELIAL MESENCHYMAL TRANSITION (Supplementary Fig. 15b), and gene set enrichment confirmed the same pathways (Fig. 4e). Among DEGs upregulated in Sox9-tdT+ BCs were many chemokines, alarmins, and Ager, encoding the RAGE receptor for the alarmin HMGB1A1 (Fig. 4f)37. A second group of genes upregulated in Sox9-tdT+ BCs encoded diverse collagen components (Fig. 4g). By contrast, Sox9-tdT+ BCs had markedly lower levels of genes involving ciliogenesis (Fig. 4h and Supplementary Fig. 15c). To understand whether SOX9+ SMG progenitors did indeed have reduced ciliogenesis compared to KRT5+ BCs, we subjected Krt5-tdT reporters and Sox9-tdT reporters to the ALT protocol and assessed the frequency with which each cell type gave rise to Ac-tub+ ciliated cells at day 14. While 20.5 ± 7% of Krt5-tdT reporters gave rise to ciliated cells, this was reduced to 5.4 ± 3% in Sox9-tdT reporters (Fig. 4i-j, Supplementary Fig. 16). These findings suggest that SOX9+ BCs have the potential to promote both inflammation and fibrosis, and that they are not poised to generate ciliated cells.

a Experimental schema. b Principal-component analysis (PCA) of the top 500 most variable genes expressed in the two groups. Numbers in parentheses indicate the percent variance captured by each PC. c Relative expression of Sox9 and canonical BC genes. d Heatmap of 1443 differentially expressed genes (DEGs) following unsupervised hierarchical clustering, defined by |log2FoldChange | > 0.58 and adjusted p-values < 0.05 with multiple testing correction using the Benjamini–Hochberg method (e) GSEA enrichment plots for two of the most overrepresented gene sets. Gene set enrichment analysis (GSEA) was performed using a weighted Kolmogorov–Smirnov–like statistic to calculate enrichment scores. Statistical significance was assessed by permutation testing, and nominal P values were derived from the permutation-based null distribution. Normalized enrichment scores (NES) were calculated to account for differences in gene set size, and multiple hypothesis testing was controlled using the Benjamini–Hochberg false discovery rate (FDR) correction. f–h Heatmaps of DEGs related to alarmins and inflammatory genes (f), collagen component genes (g), and ciliogenesis and cilia assembly genes (h). i, j Representative images (i) and quantification (j) of acetylated tubulin (Ac-tub) staining on tracheal sections from the indicated strains at day 14. Scale bars: 20 μm. The dashed line marks the border between SAE and SM. Group sizes: n = 7, 8 mice, from left to right. Two-tailed unpaired Mann-Whitney U test. Box plots (j) show median (center line), 25th–75th percentiles (box), and min–max values (whiskers). Each data point represents the average of 3–4 images from one mouse. See Methods. Panel a was created in BioRender. Lee, M. (https://BioRender.com/dg3i7rm). Source data are provided as a Source Data file.
To better understand the function of SOX9 in airway stem cells, we deleted Sox9 from SMG cells and from BCs on the SAE by generating Krt5CreERT2Sox9fl/fl mice and treated them with ALT (Fig. 5a). Immunofluorescence staining revealed a deletion efficiency of ~80% in each cell type (Supplementary Fig. 17a, b). Deletion of Sox9 was associated with significant reductions in epithelial damage (Fig. 5b, c), collagen deposition in the submucosa (Fig. 5d) and SMG hyperplasia (Fig. 5e). Remarkably, while the epithelium of ALT-challenged Krt5CreERT2 mice had poor expression of acetylated α-tubulin (Ac-tub) that marks the apical border of well-differentiated ciliated cells, low expression of the epithelial marker E-Cadherin, and high expression of the mesenchymal marker α-smooth muscle actin (α-SMA), deletion of Sox9 restored ciliated cell differentiation (Fig. 5f, g), restored epithelial E-Cadherin expression (Fig. 5h, i), and reduced α-SMA expression across the epithelium (Fig. 5h, i). Collectively, our findings demonstrate that SOX9 expression in airway stem cells prevents normal barrier repair and ciliated epithelial cell differentiation and drives mesenchymal changes in the epithelium and submucosa.

a Experimental schema. b–e Quantification of p-γ-H2A.X+ (b), TUNEL+ (c), collagen deposition (d), and gland numbers (e) from the indicated strains at day 14. Group sizes: n = 4, 7 mice, from left to right. Two-tailed unpaired Mann-Whitney U test. f, g Representative images (f) and quantification (g) of acetylated tubulin (Ac-tub) staining on tracheal sections from the indicated strains at day 14. Scale bars: 200 μm. Areas outlined are shown on the right (Scale bars: 50 μm). Group sizes: n = 6, 8 mice, from left to right. Two-tailed unpaired Mann-Whitney U test. h, i Representative images (h) and quantification (i) of E-Cadherin and α-SMA staining in tracheal sections from the indicated strains at day 14. Scale bars: 50 μm. Group sizes: n = 7, 7 mice, from left to right. Two-tailed unpaired Mann-Whitney U test. The dashed line marks the border between SAE and SM. Box plots (b–e, g, i) show median (center line), 25th–75th percentiles (box), and min–max values (whiskers). Each data point represents the average of 3–4 images from one mouse, except for glandular number (e), which represents the total glands per trachea measured by tiled imaging. See “Methods”. Panel a was created in BioRender. Lee, M. (https://BioRender.com/dg3i7rm). Source data are provided as a Source Data file.
Tuft cells and SOX9 expression are correlated with mesenchymal features in chronic rhinosinusitis with nasal polyps (CRSwNP)
Finally, we applied our mechanistic insights to CRSwNP, an airway disease characterized by tuft cell hyperplasia, increased CysLT generation, and impaired barrier function20,38,39. We first looked to assess tuft cell numbers in CRSwNP, as compared to CRS without nasal polyps (CRSsNP) and healthy controls (HCs). As expected, the patients with CRSwNP exhibited an increased number of POU2F3+ tuft cells (Fig. 6a, b), and had decreased expression of E-Cadherin levels, as compared to CRSsNP or HC patients (Fig. 6a, c)20. Further analysis confirmed that E-Cadherin expression was negatively correlated with POU2F3+ tuft cell numbers (Fig. 6d). As a reduction in E-Cadherin expression might reflect barrier damage, and not mesenchymal change, we also assessed the expression of SOX9 and α-SMA. Histologic analysis demonstrated an increased frequency of SOX9+ cells, α-SMA+ cells, and stellate shaped SOX9+ α-SMA+ cells in the sinonasal mucosa of patients with CRSwNP, compared to CRSsNP and HC (Fig. 6e, f). Further analysis in patients with CRSwNP also demonstrated an inverse correlation between the level of E-Cadherin expression and the frequency of SOX9+ or SOX9+α-SMA+ cells (Fig. 6g-i). These results demonstrate mesenchymal changes in the mucosa of CRSwNP. To better determine whether BCs from CRSwNP express a SOX9-dependent mesenchymal program or simply express SOX9, we assessed whether genes upregulated in murine SOX9+ BCs (Fig. 4d) were overrepresented in BCs from a recently reported single cell RNA-seq dataset on CRSwNP. Module scoring for the SOX9+ BC signature was upregulated in BC clusters from CRSwNP, as compared to CRSsNP and HCs (Fig. 6j). These findings indicate that a tuft cell- and SOX9-driven mesenchymal change is expressed in the human sinonasal mucosa.

a Combined ISH for Pou2f3, and immunofluorescence for E-Cadherin in human sinonasal tissue from healthy control (HC) and CRSwNP. Pou2f3+ cells (yellow arrow). Scale bars: 20 µm. b, c Quantification of Pou2f3+ tuft cells (b) and E-Cadherin staining (c) in the SAE across human sinonasal tissues. Group sizes: n = 6, 11, 27 independent patients, from left to right. Two-tailed unpaired Mann-Whitney U test (b). Kruskal–Wallis test followed by Dunn’s multiple-comparisons test with Holm–Bonferroni correction (c). d Spearman’s correlation analysis (two-sided) between E-Cadherin levels and Pou2f3+ tuft cell numbers in patients with CRSwNP (n = 27, independent patients). No multiple comparison adjustment was applied. e Representative images of E-Cadherin, α-SMA, and SOX9 staining in human sinonasal mucosa from CRS patients and healthy control (HC). SOX9+α-SMA+ cells (yellow arrow). Scale bars: 20 µm. f Quantification of SOX9+ and/or α-SMA+ cells in the SAE from the indicated groups. Group sizes: n = 6, 11, 27 independent patients, from left to right. Kruskal–Wallis test followed by Dunn’s multiple-comparisons test with Holm–Bonferroni correction. g, h Spearman’s correlation analysis (two-sided) between SOX9+ cell numbers and E-Cadherin intensity (g) and α-SMA+ cell numbers (h) in patients with CRSwNP (n = 27 independent patients). No multiple comparison adjustment was applied. i Spearman’s correlation analysis (two-sided) between E-Cadherin levels and SOX9+α-SMA+ cell numbers in patients with CRSwNP (n = 27 independent patients). No multiple comparison adjustment was applied. The Spearman r values and the corresponding P values were calculated (g–i). j SOX9 progenitor cell signature module score in basal EpCs from the Wang scRNA-seq dataset (HRA000772) across the patients. One-sided Wilcoxon rank sum test was used. No multiple comparison adjustment was applied. The dashed line marks the border between SAE and SM. Box plots (b, c, f) show median (center line), 25th–75th percentiles (box), and min–max values (whiskers). Each data point represents the average of 3–4 images from one patient. See “Methods”. Source data are provided as a Source Data file.
Discussion
Tuft epithelial cells provide well-defined sentinel and innate immune functions in the respiratory tract, triggering neural sensory networks14,15,40, activating local ILC2s17,21, stimulating ciliary beat16,41, and promoting anti-microbial peptide release in a paracrine fashion42. Prior studies in the respiratory tract have identified that tuft cells also sense tissue damage, and do so through activation of the P2Y2 receptor for extracellular nucleotides13. Moreover, they are detected at an increased frequency in the setting of injury and repair24,25,27,43, but a functional role in tissue repair or remodeling has not been clearly defined. In this study, we used models of airway injury elicited by influenza and by protease allergen to demonstrate that tuft cell-dependent CysLTs are critical for activating OXGR1-expressing SOX9+ accessory stem cells in SMGs. Activated SMG progenitors regenerate the SAE stem compartment, but bring a pro-inflammatory and mesenchymal program to the SAE and reduce ciliated cell differentiation. This leads not to optimal epithelial repair, but to impaired tight junction protein expression and barrier leak. Furthermore, activation of SMG progenitors leads to durable increases in submucosal collagen deposition and glandular hyperplasia. These findings are prevented with deletion of tuft cells, Oxgr1, or Sox9. In the human sinonasal mucosa, we find that the numbers of tuft cells and of SOX9+α-SMA+ cells are elevated in patients with CRSwNP, and that their frequencies correlate with regions of disrupted epithelium. These findings demonstrate a link between tuft cell sensing of tissue damage and the initiation of wound healing programs that are detected in the remodeled human airway (Fig. 7).

(Left) In homeostasis, basal cells are the dominant progenitor of all ciliated and secretory epithelial cells on the SAE. (Right) In the setting of tissue damage elicited by allergens or by influenza (1), tuft cells generate CysLTs (2). Leukotriene E4 (LTE4) activates OXGR1 (also known as CysLTR3 and GPR99) on SOX9+ SMG progenitors (3). This results in airway remodeling including SMG hyperplasia (4) and submucosal collagen deposition (5). SOX9+ progenitors regenerate the SAE but allow persistent barrier leak (6), bringing a program that is rich in mesenchymal genes and deficient in ciliogenesis genes. This leads to an SAE with reduced ciliated cells, reduced E-Cadherin, and increased α-SMA (7). This figure was created in BioRender. Barrett, N. (https://BioRender.com/d9cr3ur).
The coordination of tissue maintenance and repair in adult organs is facilitated by stem cell niches2,44. In the epithelium, niche elements tailor stem cell programs, licensing stem cells to migrate to damaged tissue, and instructing them to exhibit characteristics of the local neighborhood4,45,46,47,48,49,50. Prior studies have shown that SOX9+ SMG progenitor cells can act as reserve stem cells in the setting of severe airway injury6,29,51,52,53,54, but the local cues guiding SMG progenitors have not been well-defined. Here we used models of moderate airway injury, allowing us to identify a surprising role for SMG progenitors responding to CysLT generation from tuft cells in the SAE. Previous studies have implicated tuft cells in epithelial repair and/or remodeling in diverse settings. In the olfactory mucosa, they drive proliferation of KRT5+ BCs55, whereas in the small intestine, tuft cells promote both goblet cell metaplasia10,11,12 and paneth cell hyperplasia56. Additionally, a recent study using clonogenic assays identified that human intestinal tuft cells were radio-resistant and capable to reconstitute epithelial lineages ex vivo57. Thus, further identification of the tuft cell products that elicit autocrine and paracrine mobilization of epithelial stem cell programming is warranted. Notably, while tuft cells are not normally detected in the murine lung parenchyma, they are newly generated from dysplastic KRT5+ cells several weeks after viral or chemical lung injury25,27,58. In this context, studies have shown a limited effect of tuft cells on tissue remodeling, presumptively due to the absence of a tuft cell-activating ligand at late timepoints and/or a remodeling process that is already well-established before tuft cell differentiation25,58.
We found that tuft cell generation of CysLTs was important for OXGR1- and SOX9-dependent repair of the SAE in response to both a protease allergen and influenza. Tuft cell CysLT generation can be elicited by other pathogens associated with tissue damage such as N. brasiliensis helminth infection59 and protease-containing house dust mite aeroallergens13, suggesting additional settings where tuft cell-elicited remodeling may be germane. Notably, CysLTs can also be generated in activated immune cells such as mast cells, eosinophils, dendritic cells, and macrophages that infiltrate the mucosa in chronic inflammatory diseases such as CRSwNP, asthma, or ulcerative colitis60,61, and our study does not exclude a role for tuft cells in recruiting CysLT-generating inflammatory cells and thereby amplifying a remodeling circuit. Taken together, these data suggest that OXGR1- and SOX9-dependent remodeling is likely to be detected in the context of chronic type 2 or even non-type 2 inflammation.
Previous reports found that SMG progenitors could reconstitute both ciliated and secretory epithelial cells on the cell surface, maintain expression of barrier function proteins such as E-Cadherin and ZO-1, and respond appropriately to subsequent injury5,6. However, we found that SMG progenitors were imperfect substitutes for resident BCs leading to poor ciliated cell differentiation, a leaky barrier, and an increase in mesenchymal programs detected on the SAE. This is in keeping with findings for SOX9 promoting EMT in mammary, oral, and colorectal epithelial cells62,63,64. Thus, in some contexts, SOX9+ SMG progenitors are likely to lower the threshold for EMT in response to injury. Interestingly, while lineage tracing studies and antibody staining demonstrated that injury-elicited SOX9+ cells on the SAE were a minority of EpCs, changes in E-Cadherin and α-SMA expression were far more prevalent, suggesting a potential paracrine effect of SMG progenitors on surrounding BC-derived EpCs. Alternately, the extensive expression of α-SMA may represent expansion of myoepithelial cells, another submucosal progenitor population previously reported to migrate to the SAE upon injury5,6. Notably, although SMGs are primarily restricted to the proximal trachea of mice65,66, they are present throughout the cartilaginous airways in human lung. Thus, our mechanistic findings for SMG progenitors may play a more significant role in the human airway.
In the submucosa, activation of SMG progenitors was associated with durable airway remodeling including the submucosal deposition of collagen. This finding was reduced with deletion of Sox9, consistent with recent papers demonstrating that deletion of Sox9 from endothelial cells reduces fibrosis of the heart, lung, and liver67 and deletion from type 2 alveolar cells reduces lung fibrosis68. Whether SOX9 promotes fibrosis solely through an autocrine effect as a pioneer transcription factor regulating stem cell fate69 or whether it also regulates a secretory program with paracrine effects on surrounding immune cells or fibroblasts70 is not yet clear.
In sum, our findings demonstrate a direct association between tissue damage, tuft epithelial cell CysLT generation, and activation of a SMG stem cell program that leads to aberrant repair in the airway SAE and remodeling in the airway submucosa. These findings provide an understanding of how innate immune repair programs can be coopted to lead not to successful repair, but to remodeling. Ultimately, while SMG stem cells may provide a rapidly mobilized stem function to quickly repair an injured barrier, our findings suggest that optimal regeneration of the injured airway should be left to ‘professional’ BCs on the SAE.
Methods
All research conducted in this study complied with all relevant ethical regulations. This study was approved by the Brigham and Women’s Hospital Institutional Review Board under protocol number 2003P002088. Animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) at Brigham and Women’s Hospital under protocol number 2016N000296. For a complete list of antibodies and clone numbers, please see Supplemental Table 1. For a list of chemical reagents and peptides, See Supplemental Table 2. For a list of strains, see Supplemental Table 3. For a list of software and versions, see Supplemental Table 4.
Animal studies
All laboratory mice were maintained under standard husbandry and housing conditions approved by the Brigham and Women’s Hospital-specific pathogen-free animal facility in accordance with the guidelines established by the Institutional Animal Care and Use Committee (IACUC) and Laboratory Animal Resource Center (protocol number 2016N000296). Pups were weaned between 21 and 28 days old. Purchased mice were used for experiments after at least 2 weeks in the same mouse facility. Animals were maintained on a 12 h light/dark cycle and provided with food and water ad libitum in individually ventilated units. All experiments were performed during the day. Animals were genotyped at weaning using earhole punches.
Female (12 weeks- to 14 weeks) and male (10 weeks- to 12 weeks) mice of the following genotypes and strains were used: C57BL/6 (Charles River Laboratories, CRL# 027), Oxgr1-/- (KOMP, stock number 048933-UCD), Pou2f3-/- (The Jackson Laboratory, stock number 037040), Cysltr1-/- (kindly provided by Dr. Lora Bankova, BWH, Boston), Cysltr2-/- (kindly provided by Dr. Joshua Boyce, BWH, Boston), Krt5CreERT2 (The Jackson Laboratory, stock number 029155), Pou2f3CreERT2 (The Jackson Laboratory, stock number 037511), Sox9CreERT2 (The Jackson Laboratory, stock number 035092), Sox9 fl/fl (The Jackson Laboratory, stock number 013106), Oxgr1fl/fl (Barrett Lab), Ltc4sfl/fl(kindly provided by Dr. Lora Bankova, BWH, Boston), and Ai9 (RCL-tdT) (The Jackson Laboratory, stock number 007909). Gene-specific reporter mice were generated by crossing Ai9 (RCL-tdT) mice to Pou2f3CreERT2, Krt5CreERT2 and Sox9CreERT2, respectively. Oxgr1 conditional knock out mice were obtained by crossing Oxgr1fl/fl animals to Krt5CreERT2, Pou2f3CreERT2, and Sox9CreERT2, respectively. Pou2f3CreERT2::Ltc4sfl/fl mice were generated by crossing Pou2f3CreERT2 mice with Ltc4sfl/fl mice. To evaluate the SOX9 progenitor cell regeneration of the SAE upon Oxgr1 and Pou2f3 deletion, Sox9CreERT2::R26R tdT mice were additionally bred with either Oxgr1-/- or Pou2f3-/- mice, respectively. Sox9 fl/fl mice were crossed with Krt5CreERT2 to evaluate the effect of Sox9 deletion on epithelial barrier dysfunction and remodeling.
Animals were assigned randomly to experimental groups and studies were performed as blinded. However, age- and sex-matched, and whenever possible, heterozygous littermates (Cre +/-) were used for each experiment. In murine studies, both males and females were used. After reviewing damage to the epithelium (Fig. 1f) and SOX9-dependent regeneration (Fig. 2h, i) and finding no differences between males and females (see bottom of source data table for these panels), we no longer disaggregated by sex. Experiments were initiated using mice that had no known health/immune concerns, were not involved in previous procedures, and were otherwise drug/test naive. For lineage tracing and gene-specific deletion experiments, Cre-mediated recombination was induced in mice by i.p. injection of tamoxifen (1 mg/20 gm body weight) every 24 h for a total of 5 consecutive days. Mice were allowed to recover for indicated time points between tamoxifen treatment and ALT administration. For the lineage tracing experiments, tamoxifen was administered when the mice were 5–6 weeks old, while for the gene-specific deletion experiments, it was given at 9–10 weeks of age.
For all in vivo allergen challenge experiments, mice were given intranasal inhalations of Alternaria alternate (ALT) culture filtrate (Greer Laboratories) after full sedation with an intraperitoneal injection of ketamine (10 mg/kg) and xylazine (20 mg/kg). Mice received an intranasal application of 15 µg of ALT culture filtrate antigen in 20 µl of sterile HBSS (without calcium, magnesium, and phenol red, Corning). This was repeated for the indicated number of weeks (the length of time is indicated in each figure and figure legend). After each intranasal application the mice were observed on a heating pad until they had recovered from anesthesia. Mice were euthanized at the indicated time points with ketamine overdose, and tissues were collected for histology or digested for flow cytometry and sequencing. Mice of a given genotype were randomized to treatment dose, and challenges were performed in groups organized by genotype and treatment dose.
For influenza-induced injury experiments, the dose of Influenza virus A/Puerto Rico/8/34 (PR8) (ATCC) was determined based on titration experiments. Mice were anesthetized with ketamine (10 mg/kg) and xylazine (20 mg/kg) and administered 103 PFU of the virus through intranasal inhalation, as previously described6. The animals were monitored daily, and they were euthanized on days 0 and 7 post-infection (7 DPI), after which the tracheas were collected for histological analysis.
Assessment of epithelial tight junctions and permeability
After the final ALT challenge, the mice were sedated with an intraperitoneal injection of ketamine (10 mg/kg) and xylazine (20 mg/kg), then treated intranasally with FITC-4kDa Dextran (10 mg/ml; Sigma). 1 h later, the mice were euthanized, and their tracheas were collected, thoroughly rinsed with HBSS, and processed for confocal scanning. To further evaluate epithelial integrity, conventional immunofluorescence (IF) staining was performed using a Claudin-1 antibody (1:200, Rb, Invitrogen). Detailed steps for tissue processing and IF staining are outlined below.
Histopathology assessment
The murine tracheas were harvested and processed as described below. H&E staining and masson’s trichrome (MT) staining were conducted using H&E and MT staining kits from Abcam, following the manufacturer’s instructions with minor modifications. The slides were dehydrated and mounted using VectaMount® Express Mounting Medium (Vector). All images were taken on a Leica DMi8 microscope with Leica Application Suite X (LAS X) software, equipped with a Leica DMC5400 color camera for histological samples.
TUNEL assay
To obtain cryosections (5 µm), the murine tracheas were processed as described below. Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay with immunofluorescence staining against p-γ-H2A.X (Abcam) were performed using the Click-iT™ Plus TUNEL Assay Kits for In Situ Apoptosis Detection (Invitrogen) according to the manufacturer’s instructions. The tissues were mounted with VECTASHIELD Vibrance Antifade Mounting medium with DAPI (Vector) and subjected to imaging. Fluorescent images were taken with a Zeiss LSM 880 with an Airyscan confocal system on a Zeiss Axio Observer Z1 inverted microscope equipped with a 20× Zeiss (0.8 NA) objective.
RNA in situ hybridization (RNAscope) and RNA-protein co-detection
The mouse trachea and human sinus tissues were fixed in 4% PFA and dehydrated in 30% sucrose before being sectioned for OCT embedding. The tissue samples were sliced at a thickness of 5–7 µms and attached on Superfrost plus Microscope slides (Thermo Fisher Scientific). The RNAscope Multiplex Fluorescent v2 Assay combined with immunofluorescence – Integrated Co-Detection Workflow (ICW) (Advanced Cell Diagnostics) was used following the manufacturer’s protocol. Briefly, the tracheal sections were washed with PBS for 10 min at room temperature, baked for 30 min at 60 °C in the HybEZ Oven, and post-fixed in 4% PFA for 15 min at 4 °C. RNAscope hydrogen peroxide was used to block endogenous peroxidase activity for 10 min at room temperature, followed by RNAscope target retrieval ~100 °C for 10 min. After a 5 min of wash with 1X PBST, the slides were incubated overnight at 4 °C with the indicated primary antibodies: anti-DCLK1 (1:100, Rabbit, Abcam), anti-SOX9 (1:50, Goat, R&D Systems), anti-SOX9 (1:50, Rabbit, Abcam), anti-α-SMA (1:100, Rabbit, Abcam), anti-Uteroglobin (1:100, Rabbit, Abcam), anti-KRT5 (1:200, Rabbit, Abcam), anti-KRT5 (1:200, Chicken, Biolegend), anti-BCAM (1:100, Goat, R&D Systems), anti-E-Cadherin (1:200, Rabbit, Cell Signaling), and anti-E-Cadherin (1:200, Mouse, Invitrogen). The primary antibodies were post-fixed in 10% Neutral Buffered Formalin (NBF) for 30 min, followed by incubation with RNAscope Protease III for 20 min at 40 °C in the HybEZ Oven before probe incubation. The standard amount of the probe mix for mOxgr1 (Cat. No.1077911-C4), mKrt5 (Cat. No.415041-C2), mKrt8 (Cat. No. 424521-C1), mLtc4s (Cat. No.1046751-C3), mPou2f3 (Cat. No. 539211-C1), hPou2f3 (Cat. No. 554051-C3), mDclk1 (Cat. No. 476631-C1), mCFTR (Cat. No. 1095651-C1), mCGRP (Cat. No. 578771-C3), and mPecam1 (Cat. No. 316721-C3) was denatured for 15 min at 40 °C, added to the slides, and incubated for 2-3 h at 40 °C in the HybEZ Oven. The hybridization was carried out for AMP1, AMP2, and AMP3, thereafter the HRP-C1, C2, C3, and C4 signals were developed sequentially according to the manufacturer’s instructions. Each channel was stained with specific Opal dyes (Opal 520, 620, 690; Akoya Biosciences). Next, the slides were incubated with appropriate secondary antibodies for 40 min at room temperature: donkey anti-rabbit AF488, AF594, AF680 (1:200, Invitrogen), donkey anti-mouse AF680 (1:200, Invitrogen), and donkey anti-goat AF488, AF680 (1:200, Invitrogen). Finally, the slides were mounted using VECTASHIELD Vibrance Antifade Mounting medium with DAPI (Vector) and scanned with a Zeiss LSM 880 with Airyscan confocal system on a Zeiss Axio Observer Z1 inverted microscope with 10× Zeiss [0.30 numerical aperture (NA)], 20× Zeiss (0.8 NA), and a 63× Zeiss oil (1.4 NA) objectives.
Mouse tracheal epithelial cell isolation and sorting
For SOX9-tdT+ cell isolation, tracheas were collected and connective tissue was discarded. Tracheas were opened longitudinally and incubated in pre-warmed HBSS solution containing dispase (16 U/ml; Gibco) and deoxyribonuclease I (DNase I; 20 µg/ml; Roche) for 30 min at room temperature with gentle nutation. The first step of digestion was stopped by adding cold RPMI 1640 medium + 5% fetal bovine serum (FBS; Sigma-Aldrich), and then the tracheal epithelium was peeled off with a scalpel under a dissecting microscope. The epithelium (for Sox9-tdT+ cells in the SAE) was incubated in Hepes-Tyrode’s buffer without calcium (Boston BioProducts) containing 15 µl of papain (28 U/mg; Sigma-Aldrich) per milliliter of buffer and 10 µl L-cysteine (25 mg/ml; Sigma-Aldrich) per milliliter of buffer for 20 min at 37 °C on a shaker set to 200 rpm. Papain digestion was terminated with Tyrode’s solution with calcium (Boston BioProducts) including 4 µl leupeptin (5 mg/ml; Sigma-Aldrich) per milliliter of buffer. The digested tissue was vortexed for 10 s and thoroughly triturated using a 21-gauge needle 10 times. All cell suspensions were combined, passed through a 40-µm cell strainer (Thermo Fisher Scientific), and centrifuged at 1000 g for 10 min. Cell pellets were re-suspended with cold FACS buffer before staining.
Single-cell suspensions from the trachea of SOX9-tdT mice were blocked with Fc-blocker for 10 min on ice, then the following anti-mouse antibodies were utilized for FACS: CD31-BV421 (1:100, Biolegend), CD45-BV421 (1:100, Biolegend), CD90.2-BV421 (1:100, Biolegend), EpCAM-FITC (1:100, Biolegend), BCAM (1:100, MBL International). For cell sorting, the following populations were identified: Lin-EpCAM+BCAM+ tdT+ (for SOX9-tdT+ progenitor cells) and Lin-EpCAM+ BCAM+ tdT- (for conventional BCs). Dead cells were excluded using a Ghost fixable dye (Tonbo bioscience). Cell sorting was performed at the Brigham and Women’s Human Immunology Flow Core using a BD FACSAria Fusion cell sorter.
Immunofluorescence staining and microscopy
For whole trachea mounts, tracheas were harvested, cleaned of connective tissue, longitudinally opened, and subjected to overnight fixation in 4% PFA (Boston Bioproducts) at 4 °C. The tissues were then washed in 1X PBS (Boston Bioproducts) for 30 min at room temperature, followed by permeabilization in a 1X blocking buffer (Abcam) containing 0.2% Triton X-100 (Sigma-Aldrich), 0.1% Tween 20 (Sigma-Aldrich), and 5% normal donkey serum (Jackson Immuno Research Laboratories) for at least 2 h at room temperature. The tracheas were incubated overnight at 4 °C for 48 h with the specified primary antibodies: anti-DCLK1 (1:100, Rabbit, Abcam), and anti-GNAT3 (1:100, Goat, MyBioscience). The tracheas were rinsed with washing buffer (PBS containing 0.2% Triton X-100 and 0.1% Tween 20) for 1-2 h at 4 °C, followed by incubation with the appropriate combinations of secondary antibodies along with DAPI (1:500, Sigma-Aldrich): donkey anti-rabbit AF488 (1:200, Invitrogen), and donkey anti-goat AF594 (1:200, Invitrogen). Finally, the tracheas were longitudinally split into two halves, with the epithelial surface facing upward, and embedded using VECTASHIELD Vibrance Antifade Mounting medium with DAPI (Vector).
For sectioned slides, mouse trachea or human sinus tissues were fixed in 4% PFA at 4 °C overnight and then washed in PBS for 10 min at room temperature, followed by overnight incubation in 30% sucrose at 4 °C. The tissues were then embedded in an O.C.T Embedding Compound (Thermo Fisher Scientific) and cryosectioned (5-7 µms) for histological analysis. The frozen tissue sections were rinsed in PBS for 10 min at room temperature and then permeabilized in a 1X blocking buffer (Abcam) containing 0.2% Triton X-100, 0.1% Tween 20, and 5% normal donkey serum for at least 30 min at room temperature. The slides were incubated with the indicated primary antibodies at 4 °C overnight: anti-DCLK1 (1:100, Rabbit, Abcam), anti-p-γ-H2A.X (1:100, Rabbit, Abcam), anti-Claudin-1 (1:200, Rb, Invitrogen), anti-GNAT3 (1:100, Goat, MyBioscience), anti-BCAM (1:100, Goat, R&D Systems), anti-Ac-tubulin (1:200, Mouse, Sigma), anti-SOX9 (1:50, Goat, R&D Systems), anti-SOX9 (1:50, Rabbit, Abcam), anti-α-SMA (1:100, Rabbit, Abcam), anti-α-SMA (1:100, Mouse, Sigma Aldrich), anti-E-Cadherin (1:200, Goat, R&D Systems), anti-E-Cadherin (1:200, Mouse, Invitrogen), anti-KRT5 (1:200, Chicken, Biolegend), and anti-KRT5 (1:200, Rabbit, Abcam). Afterward, the slides were rinsed with washing buffer and incubated with the appropriate secondary antibodies: donkey anti-rabbit AF488, AF594, AF680 (1:200, Invitrogen), donkey anti-mouse AF488, AF594 (1:200, Invitrogen), and donkey anti-goat AF488, AF594, AF680 (1:200, Invitrogen). Finally, the tissues were mounted with VECTASHIELD Vibrance Antifade Mounting medium with DAPI (Vector) and subjected to imaging. Fluorescent images were acquired with a Zeiss LSM 880 with Airyscan confocal system on a Zeiss Axio Observer Z1 inverted microscope equipped with 10× Zeiss [0.30 numerical aperture (NA)], 20× Zeiss (0.8 NA), and a 63× Zeiss oil (1.4 NA) objectives.
Image processing and quantification of cell numbers
Confocal images were obtained using a Zeiss LSM 880 with Airyscan confocal system on a Zeiss Axio Observer Z1 inverted microscope, utilizing 10× Zeiss [0.30 numerical aperture (NA)], 20× Zeiss (0.8 NA), or a 63× Zeiss oil (1.4 NA) objectives for quantification. For some experiments, assessments were conducted using tile-scanned images which contain 5-7 cartilages starting from the proximal to the distal region and then normalized to the length of the basement membrane (BM). Large panels (tile scanned images) shown in Figures were generated by aligning and stitching individual pictures using tile imaging. The confocal and wide-field images were processed using Zen software (Carl Zeiss) and Leica Application Suite X (LAS X) software, respectively.
Detailed quantification methods are provided below. The raw data for every sample is provided in the Source Data file.
Epithelial denuding was assessed by measuring and averaging the epithelial height and DAPI+ cell numbers per high power field at three randomly selected regions for each mouse trachea. To evaluate acute and chronic epithelial damage, TUNEL+ cells and p-γ-H2A.X+ cells were manually counted and averaged from three randomly selected regions per mouse trachea.
To assess epithelial barrier leakage, the intensity of FITC-Dextran in the sub-epithelium was measured using tile-scanned images and then normalized to the length of the basement membrane for each mouse trachea. To evaluate the intensity of Claudin-1, the intensity of Claudin-1 was measured and averaged from three randomly selected regions per mouse trachea. To compensate for variations in total cell density, the expression levels were normalized to the corresponding number of SAE DAPI+ cells.
To quantify collagen deposition, whole mount images were taken to quantify collagen in the intercartilagenous regions from C1 to C6. After color deconvolution in Image J as previously described71 (Supplementary Fig. 5a), the intensity of collagen fibers in the sub-epithelium was measured and averaged from three randomly selected regions per trachea (Supplementary Fig. 5a) and then normalized to the length of the basement membrane. The relative levels of collagen deposition are shown as fold changes.
To quantify gland hyperplasia, whole mount images were taken, stained with Masson Trichrome, and the number of glands located in the subepithelium from C1 to C7 was counted. Each dot represents the sum of glands from the C1 to the C7 cartilage.
To quantify the recruitment of SOX9+ progenitor cells to the SAE in Sox9CreERT2::R26R TdT mice, the number of Sox9-tdT+BCAM+ cells per HPF was manually counted and averaged from three randomly selected regions per sample (day 14 of ALT) or from four randomly selected regions per sample (7 days post-influenza infection). For quantifications in the Sox9CreERT2::R26R tdT mice bred with either Oxgr1-/- or Pou2f3-/-, the number of Sox9-tdT+BCAM+ cells was manually counted and averaged from four randomly selected regions per mouse trachea. To calculate the percentage of Sox9-tdT+BCAM+ cells, the total number of BCAM+ cells was manually counted and averaged from the same regions and used as the denominator. For the tuft cell-specific Ltc4s deletion mice and the cell-specific Oxgr1 deficient mice, Sox9+BCAM+ cells were manually counted and averaged from three randomly selected regions per mouse trachea.
To quantify Oxgr1-expressing cells within a subset of EpCs, the Oxgr1+ cells that co-expressed each representative EpC marker were counted and averaged from three randomly selected regions per mouse trachea. Oxgr1 was quantified using automated imaging software (QuPath), with the following parameters: pixel size of 1.0 μm, background radius of 8.0 μm, minimum area of 10 μm², maximum area of 30 μm², intensity threshold of 10, and cell expansion size of 1.
To evaluate the intensity of Ac-tubulin, the intensity of Ac-tubulin was measured and averaged from three randomly selected regions per mouse trachea. To compensate for variations in total cell density, the expression levels were normalized to the corresponding number of SAE DAPI+ cells.
To determine the proportion of lineage-traced Krt5+ reporter cells that became acetylated tubulin–positive (Ac-tub⁺) ciliated cells, the number of Krt5-tdT⁺Ac-tub⁺ cells was counted and averaged from 3 regions per mouse trachea and divided by the total number of Krt5-tdT⁺ cells in the same regions. The same calculation was performed to determine the proportion of lineage-traced Sox9-tdT⁺ cells that became ciliated cells. To evaluate the E-Cadherin and α-SMA expression levels in mouse tracheal tissues, these markers were measured using Image J software and averaged from three randomly selected regions per mouse trachea. To compensate for variations in total cell density, the expression levels were normalized to the corresponding number of SAE DAPI+ cells.
To quantify tuft cells in whole trachea mounts, the numbers of Gustducin+ and/or DCLK1+ cells were manually counted and averaged from three randomly selected areas per mouse trachea.
To assess the deletion efficiency of Oxgr1 and Sox9 following tamoxifen delivery, either Oxgr1+ or Sox9+ cell numbers were manually counted and averaged from three randomly selected regions per mouse trachea and then normalized to the count of the indicated cells (e.g., KRT5-, Pou2f3-, or Sox9-positive cells), respectively. For Ltc4s deletion efficiency, Pou2f3+ cells expressing Ltc4s were manually counted and averaged from three different regions per mouse trachea. Oxgr1 was quantified using automated imaging software (QuPath) using the same criteria as described above.
For Oxgr1 probe validation, Oxgr1+ cells were counted and averaged from three different randomly selected regions per mouse trachea in both the WT and Oxgr1-/- strains. Oxgr1 was quantified using automated imaging software (QuPath) using the same criteria as described above.
To identify Pou2f3-expressing cells in response to the ALT challenge, Pou2f3CreERT2::R26R tdT mice were challenged with ALT, and Pou2f3-tdT+ cells were manually counted and averaged per sample, specifically focusing on the proximal part, where SMG cells are typically clustered.
To quantify the recruitment of Sox9+ cells to the SAE after ALT injury in different mouse strains, Sox9+ cell numbers were manually counted and averaged from three randomly selected regions per mouse trachea.
To quantify the SOX9+ and α-SMA+ in the human sinus tissues, each marker was manually counted and averaged in three randomly selected regions per patient sample. To account for differences in the density of total cells, the numbers of target cells were normalized to the corresponding SAE DAPI+ cell numbers. For Pou2f3+ cells, the number of Pou2f3+ were manually counted and averaged from four randomly selected regions per patient sample.
To quantify the E-Cadherin levels in human sinus tissues, E-Cadherin expressions were measured utilizing Image J software in three randomly selected regions and averaged per patient sample, specifically focusing on the SAE layer. To compensate for differences in the density of total cells, the expression levels were normalized to the corresponding SAE DAPI+ cell numbers.
To assess the correlation between E-Cadherin intensity and POU2F3+ tuft cells, a correlation analysis was performed across 108 tissue sections from 27 CRSwNP patients.
Patient characterization, sinus tissue procurement, and preparation
Male and female subjects were recruited from the Brigham and Women’s Hospital (Boston, MA) Allergy and Immunology clinics and Otolaryngology clinics between Jun 2021 and May 2022. The local Institutional Review Board approved the study (protocol number: 2003P002088), and all subjects provided written informed consent. Sinus tissues were obtained at the time of elective endoscopic sinus (ESS) surgery from CRSsNP and CRSwNP patients with the diagnosis based on established guidelines72. The CRSwNP group comprised patients with and without aspirin-exacerbated respiratory disease, although no differentiation was made between these specific subtypes for the analyses. Non-CRS control patients were undergoing sinus surgery to address anatomical abnormalities through the removal of concha bullosa (CB). Subjects with known cystic fibrosis, allergic fungal rhinosinusitis, and unilateral polyps were excluded from the study. Following surgical dissection, the sinus specimens were promptly transferred to cold RPMI 1640 medium containing 10% FBS. Upon reaching the laboratory, a tissue chunk was immediately fixed in 4% PFA at 4 °C overnight. It was then washed in PBS at room temperature for 10 min, followed by incubation in 30% sucrose at 4 °C overnight. The tissue was subsequently embedded in an OCT compound and stored at −80 °C until sectioning.
Low input RNA-sequencing and data analysis
Single-cell suspensions from mouse trachea were prepared as described above. For the preparation of BCs from the mouse trachea, each population was sorted with a minimum of 600 and a maximum of 800 cells. Due to the small number of SOX9+ BCs for bulk RNA-seq, SOX9+ BCs were sorted from several mice and then pooled together into one sample until there were enough cells for sequencing. All samples were mixed directly with 5 µL TCL buffer (Qiagen) and β-mercaptoethanol (Sigma-Aldrich) and stored at −80 °C. Libraries were prepared using Smart-Seq2 and 38 bp paired-end sequencing was conducted by the Genomics Platform of the Broad Institute of MIT and Harvard. Sequencing quality was assessed with FastQC (v0.12.0). Reads were pseudo-aligned and transcript expression was quantified with Kallisto (v0.51.1) using GRCm39 (GCA_000001635.9) for mouse sample. Quantification files were processed to obtain raw counts using the Tximport-DESeq2 pipeline (v1.40.2). Genes with fewer than 5 raw counts in 20% of samples were excluded.
Principal components analysis (PCA) was performed using the top 500 most variable genes (after variance stabilizing transformation of size factor-normalized counts). Significantly differential gene expression was determined based on a fold change ( | log2FoldChange | >0.58) and an adjusted P-value of <0.05, with the Benjamini-Hochberg method used to correct for multiple comparisons. Over-representation analysis (ORA) was performed using the clusterProfiler package (v4.8.3) with the gene ontology biological process (GOBP) and hallmark gene set databases from the mouse molecular signatures database (MSigDB). Gene set enrichment analysis (GSEA) was performed across the Hallmark gene sets using the fgsea package (v.1.26.0). Gene set enrichment analysis (GSEA) was performed using a weighted Kolmogorov–Smirnov–like statistic to calculate enrichment scores. Statistical significance was assessed by permutation testing, and nominal P values were derived from the permutation-based null distribution. Normalized enrichment scores (NES) were calculated to account for differences in gene set size, and multiple hypothesis testing was controlled using the Benjamini–Hochberg false discovery rate (FDR) correction.
To evaluate the SOX9 progenitor cell signature in the publicly available human data set, we first manually generated a SOX9 progenitor cell signature gene set consisting of genes differentially expressed ( | log2 fold change | > 0.58, p.adj <0.05) between purified Sox9+BCs and BCs from mice as described above. Differential expression testing was performed using DESeq2 (v1.40.2). These gene sets were then converted from mouse to human gene symbols via referencing the Jackson Laboratory’s database of mouse-human gene homologs. Metadata and scRNA-seq FASTQ files containing epithelial and immune cells from healthy control (n = 5), CRSsNP (n = 5), and CRSwNP (n = 11) were downloaded from the Genome Sequence Archive [HRA000772] and aligned to Gencode GRCh38 using 10x Genomics Cell Ranger (v6.1.2) with default parameters. Cells with >12,000 UMIs, >15% mitochondrial reads, or <500 genes detected were filtered out, as they were considered to be either fragments or doublets. We then processed the data using Seurat (v5.3.1) and Harmony (v1.2.4) to cluster and identify basal cells. Seurat was used to get module scores on a per sample (not a per cell) basis. The module score from each sample was averaged for each disease group and a one-sided wilcoxon rank sum test was used to determine the significance.
Quantification and statistical analysis
The sizes of the experimental groups were determined based on the results of preliminary experiments, and mice were randomly assigned to the groups. The number of animals per group is indicated in the figure legends as “n = x mice per group,” and multiple measurements (at least 3) were taken for each animal. All experiments were performed at least 3 times. The assignment of animals to experimental groups was random, and the studies were blinded. Statistical analyses were performed using GraphPad Prism (v10) and R (RStudio). Two-tailed unpaired Mann–Whitney U tests were conducted for pairwise comparisons. For multiple-group comparisons, Kruskal–Wallis tests followed by Dunn’s multiple-comparisons tests with Holm–Bonferroni correction were performed. The experiments were independently replicated, and representative data were presented. Statistical significance was evaluated using the unpaired two-tailed Mann-Whitney U test, the Kruskal-Wallis test, or the Spearman correlation test, as described in figure legends. Significance is reported as exact P values unless otherwise indicated.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The reanalyzed human scRNA-seq dataset is from the Genome Sequence Archive HRA000772. The murine bulk RNA-seq data of sorted epithelial subsets generated in this study have been deposited in the Gene Expression Omnibus (GEO) under accession number GSE280283. The processed data are available in Supplementary Dataset 1. All other data generated or analyzed in this study are included in the article and its Supplementary Information. Source data are provided with this paper.
References
Rock, J. R. et al. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proc. Natl. Acad. Sci. USA 106, 12771–12775 (2009).
Hogan, B. L. et al. Repair and regeneration of the respiratory system: complexity, plasticity, and mechanisms of lung stem cell function. Cell Stem Cell 15, 123–138 (2014).
Rawlins, E. L. et al. The role of Scgb1a1+ Clara cells in the long-term maintenance and repair of lung airway, but not alveolar, epithelium. Cell Stem Cell 4, 525–534 (2009).
Tata, P. R. et al. Dedifferentiation of committed epithelial cells into stem cells in vivo. Nature 503, 218–223 (2013).
Lynch, T. J. et al. Submucosal gland myoepithelial cells are reserve stem cells that can regenerate mouse tracheal epithelium. Cell Stem Cell 22, 779 (2018).
Tata, A. et al. Myoepithelial cells of submucosal glands can function as reserve stem cells to regenerate airways after injury. Cell Stem Cell 22, 668–683.e666 (2018).
Elias, J. A., Zhu, Z., Chupp, G. & Homer, R. J. Airway remodeling in asthma. J. Clin. Invest 104, 1001–1006 (1999).
Hough, K. P. et al. Airway remodeling in asthma. Front Med (Lausanne) 7, 191 (2020).
Varricchi G., Brightling C. E., Grainge C., Lambrecht B. N., Chanez P. Airway remodelling in asthma and the epithelium: on the edge of a new era. Eur. Respir. J. 63, 2301619 (2024).
von Moltke, J., Ji, M., Liang, H. E. & Locksley, R. M. Tuft-cell-derived IL-25 regulates an intestinal ILC2-epithelial response circuit. Nature 529, 221–225 (2016).
Howitt, M. R. et al. Tuft cells, taste-chemosensory cells, orchestrate parasite type 2 immunity in the gut. Science 351, 1329–1333 (2016).
Gerbe, F. et al. Intestinal epithelial tuft cells initiate type 2 mucosal immunity to helminth parasites. Nature 529, 226–230 (2016).
Ualiyeva S. et al. Airway brush cells generate cysteinyl leukotrienes through the ATP sensor P2Y2. Sci. Immunol. 5, 2301619 (2020).
Krasteva, G. et al. Cholinergic chemosensory cells in the trachea regulate breathing. Proc. Natl. Acad. Sci. USA 108, 9478–9483 (2011).
Saunders, C. J., Christensen, M., Finger, T. E. & Tizzano, M. Cholinergic neurotransmission links solitary chemosensory cells to nasal inflammation. Proc. Natl. Acad. Sci. USA 111, 6075–6080 (2014).
Perniss, A. et al. Chemosensory cell-derived acetylcholine drives tracheal mucociliary clearance in response to virulence-associated formyl peptides. Immunity 52, 683–699.e611 (2020).
Ualiyeva, S. et al. Tuft cell-produced cysteinyl leukotrienes and IL-25 synergistically initiate lung type 2 inflammation. Sci. Immunol. 6, eabj0474 (2021).
Keshavarz, M. et al. Cysteinyl leukotrienes and acetylcholine are biliary tuft cell cotransmitters. Sci. Immunol. 7, eabf6734 (2022).
Xiong, Z. et al. Intestinal Tuft-2 cells exert antimicrobial immunity via sensing bacterial metabolite N-undecanoylglycine. Immunity 55, 686–700.e687 (2022).
Kotas M. E. et al. IL-13-programmed airway tuft cells produce PGE2, which promotes CFTR-dependent mucociliary function. JCI Insight 7, e159832 (2022).
Bankova L. G. et al. The cysteinyl leukotriene 3 receptor regulates expansion of IL-25-producing airway brush cells leading to type 2 inflammation. Sci. Immunol. 3, eaat9453 (2018).
Ualiyeva, S., Hallen, N., Kanaoka, Y., Barrett, N. A. & Bankova, L. Nasal epithelial brush cells generate cysteinyl leukotrienes in response to aeroallergens and stress signals. J. Allergy Clin. Immunol. 143, AB89 (2019).
Kohanski, M. A. et al. Solitary chemosensory cells are a primary epithelial source of IL-25 in patients with chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 142, 460–469.e467 (2018).
Melms, J. C. et al. A molecular single-cell lung atlas of lethal COVID-19. Nature 595, 114–119 (2021).
Barr J. et al. Injury-induced pulmonary tuft cells are heterogenous, arise independent of key Type 2 cytokines, and are dispensable for dysplastic repair. Elife 11, e78074 (2022).
Huang H. et al. Contribution of Trp63(CreERT2)-labeled cells to alveolar regeneration is independent of tuft cells. Elife 11, e78217 (2022).
Rane, C. K. et al. Development of solitary chemosensory cells in the distal lung after severe influenza injury. Am. J. Physiol. Lung Cell Mol. Physiol. 316, L1141–L1149 (2019).
Wang X. et al. Type 2 inflammation drives an airway basal stem cell program through insulin receptor substrate signaling. J. Allergy Clin. Immunol. 151, 1536–1549 (2023).
Lynch, T. J. et al. Submucosal gland myoepithelial cells are reserve stem cells that can regenerate mouse tracheal epithelium. Cell Stem Cell 22, 653–667.e655 (2018).
Wright, E. et al. The Sry-related gene Sox9 is expressed during chondrogenesis in mouse embryos. Nat. Genet 9, 15–20 (1995).
Boitano, S. et al. Alternaria alternata serine proteases induce lung inflammation and airway epithelial cell activation via PAR2. Am. J. Physiol. Lung Cell Mol. Physiol. 300, L605–L614 (2011).
Snelgrove, R. J. et al. Alternaria-derived serine protease activity drives IL-33-mediated asthma exacerbations. J. Allergy Clin. Immunol. 134, 583–592.e586 (2014).
Yee, M. C. et al. Protease-activated receptor-2 signaling through beta-arrestin-2 mediates Alternaria alkaline serine protease-induced airway inflammation. Am. J. Physiol. Lung Cell Mol. Physiol. 315, L1042–L1057 (2018).
Zeng Y. R. et al. The immunometabolite itaconate stimulates OXGR1 to promote mucociliary clearance during the pulmonary innate immune response. J. Clin. Invest. 133, e160463 (2023).
Bankova, L. G. et al. Leukotriene E4 elicits respiratory epithelial cell mucin release through the G-protein-coupled receptor, GPR99. Proc. Natl. Acad. Sci. USA 113, 6242–6247 (2016).
Montoro, D. T. et al. A revised airway epithelial hierarchy includes CFTR-expressing ionocytes. Nature 560, 319–324 (2018).
Sims, G. P., Rowe, D. C., Rietdijk, S. T., Herbst, R. & Coyle, A. J. HMGB1 and RAGE in inflammation and cancer. Annu Rev. Immunol. 28, 367–388 (2010).
Schleimer, R. P. Immunopathogenesis of chronic rhinosinusitis and nasal polyposis. Annu Rev. Pathol. 12, 331–357 (2017).
Steinke, J. W. et al. Cysteinyl leukotriene expression in chronic hyperplastic sinusitis-nasal polyposis: importance to eosinophilia and asthma. J. Allergy Clin. Immunol. 111, 342–349 (2003).
Tizzano, M. et al. Nasal chemosensory cells use bitter taste signaling to detect irritants and bacterial signals. Proc. Natl. Acad. Sci. USA 107, 3210–3215 (2010).
Perniss, A. et al. A succinate/SUCNR1-brush cell defense program in the tracheal epithelium. Sci. Adv. 9, eadg8842 (2023).
Lee, R. J. et al. Bitter and sweet taste receptors regulate human upper respiratory innate immunity. J. Clin. Invest 124, 1393–1405 (2014).
Roach, S. N. et al. Respiratory influenza virus infection causes dynamic tuft cell and innate lymphoid cell changes in the small intestine. J. Virol. 96, e0035222 (2022).
Visvader, J. E. & Clevers, H. Tissue-specific designs of stem cell hierarchies. Nat. Cell Biol. 18, 349–355 (2016).
Rajagopal, J. & Stanger, B. Z. Plasticity in the adult: how should the Waddington diagram be applied to regenerating tissues? Dev. Cell 36, 133–137 (2016).
Donati, G. et al. Wounding induces dedifferentiation of epidermal Gata6(+) cells and acquisition of stem cell properties. Nat. Cell Biol. 19, 603–613 (2017).
van Es, J. H. et al. Dll1+ secretory progenitor cells revert to stem cells upon crypt damage. Nat. Cell Biol. 14, 1099–1104 (2012).
Stange DE et al. Differentiated Troy+ chief cells act as reserve stem cells to generate all lineages of the stomach epithelium. Cell 155, 357–368 (2013).
Blanpain, C. & Fuchs, E. Stem cell plasticity. plasticity of epithelial stem cells in tissue regeneration. Science 344, 1242281 (2014).
Tetteh, P. W., Farin, H. F. & Clevers, H. Plasticity within stem cell hierarchies in mammalian epithelia. Trends Cell Biol. 25, 100–108 (2015).
Hegab, A. E. et al. Novel stem/progenitor cell population from murine tracheal submucosal gland ducts with multipotent regenerative potential. Stem Cells 29, 1283–1293 (2011).
Lynch, T. J. & Engelhardt, J. F. Progenitor cells in proximal airway epithelial development and regeneration. J. Cell Biochem 115, 1637–1645 (2014).
Lynch, T. J. et al. Wnt signaling regulates airway epithelial stem cells in adult murine submucosal glands. Stem Cells 34, 2758–2771 (2016).
Xie, W. et al. CGRP induction in cystic fibrosis airways alters the submucosal gland progenitor cell niche in mice. J. Clin. Invest 125, 2179 (2015).
Ualiyeva, S. et al. A nasal cell atlas reveals heterogeneity of tuft cells and their role in directing olfactory stem cell proliferation. Sci. Immunol. 9, eabq4341 (2024).
Fung, C. et al. Tuft cells mediate commensal remodeling of the small intestinal antimicrobial landscape. Proc. Natl. Acad. Sci. USA 120, e2216908120 (2023).
Huang L. et al. Tuft cells act as regenerative stem cells in the human intestine. Nature 634, 929–935 (2024).
Li Y. H. et al. G protein subunit Ggamma13-mediated signaling pathway is critical to the inflammation resolution and functional recovery of severely injured lungs. Elife 12, RP92956 (2024).
McGinty, J. W. et al. Tuft-cell-derived leukotrienes drive rapid anti-helminth immunity in the small intestine but are dispensable for anti-protist immunity. Immunity 52, 528–541.e527 (2020).
Rosen, M. J. et al. Mucosal expression of type 2 and type 17 immune response genes distinguishes ulcerative colitis from colon-only Crohn’s disease in treatment-naive pediatric patients. Gastroenterology 152, 1345–1357.e1347 (2017).
Maspero J. et al. Type 2 inflammation in asthma and other airway diseases. ERJ Open Res 8, 00576–2021 (2022).
Guo, W. et al. Slug and Sox9 cooperatively determine the mammary stem cell state. Cell 148, 1015–1028 (2012).
Hirano, T., Saito, D., Yamada, H., Ishisaki, A. & Kamo, M. TGF-beta1 induces N-cadherin expression by upregulating Sox9 expression and promoting its nuclear translocation in human oral squamous cell carcinoma cells. Oncol. Lett. 20, 474–482 (2020).
Choi, B. J., Park, S. A., Lee, S. Y., Cha, Y. N. & Surh, Y. J. Hypoxia induces epithelial-mesenchymal transition in colorectal cancer cells through ubiquitin-specific protease 47-mediated stabilization of Snail: a potential role of Sox9. Sci. Rep. 7, 15918 (2017).
Liu, X. & Engelhardt, J. F. The glandular stem/progenitor cell niche in airway development and repair. Proc. Am. Thorac. Soc. 5, 682–688 (2008).
Borthwick, D. W. et al. Murine submucosal glands are clonally derived and show a cystic fibrosis gene-dependent distribution pattern. Am. J. Respir. Cell Mol. Biol. 20, 1181–1189 (1999).
Trogisch, F. A. et al. Endothelial cells drive organ fibrosis in mice by inducing expression of the transcription factor SOX9. Sci. Transl. Med 16, eabq4581 (2024).
Cai, X. T. et al. IL-4-induced SOX9 confers lineage plasticity to aged adult lung stem cells. Cell Rep. 43, 114569 (2024).
Yang, Y. et al. The pioneer factor SOX9 competes for epigenetic factors to switch stem cell fates. Nat. Cell Biol. 25, 1185–1195 (2023).
Fan, Y. et al. Epithelial SOX9 drives progression and metastases of gastric adenocarcinoma by promoting immunosuppressive tumour microenvironment. Gut 72, 624–637 (2023).
Chen, Y., Yu, Q. & Xu, C.-B. A convenient method for quantifying collagen fibers in atherosclerotic lesions by ImageJ software. Int. J. Clin. Exp. Med. 10, 14904–14910 (2017).
Meltzer, E. O. et al. Rhinosinusitis: establishing definitions for clinical research and patient care. J. Allergy Clin. Immunol. 114, 155–212 (2004).
Acknowledgements
No AI-assisted technologies were used in preparation of this manuscript. National Institutes of Health R01AI134989 (NAB) National Institutes of Health K24HL179901 (NAB). National Institutes of Health U19AI095219, Project 2 (NAB). National Institutes of Health T32AI007306 (JB and NAB).
Author information
Authors and Affiliations
Contributions
Conceptualization: M.K.L., and N.A.B. Methodology: M.K.L., and X.W. Formal Analysis: M.K.L., G.X.H., M.V.M., and N.R.H. Investigation: M.K.L., and X.W. Resources: M.K.L., T.M.L., J.A.B., Q.Y and L.C. Writing – Original Draft: M.K.L. and N.A.B. Writing – Review & Editing: M.K.L., and N.A.B. Visualization: M.K.L., and N.A.B. Funding Acquisition: N.A.B. Supervision: N.A.B.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lee, M., Wang, X., Ye, Q. et al. Tuft cells shape airway remodeling by eliciting OXGR1- and SOX9-dependent stem cell programs. Nat Commun 17, 4356 (2026). https://doi.org/10.1038/s41467-026-70763-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-70763-y
