
Abstract
Decapping is a critical step in mRNA decay, but the mechanisms regulating human decapping enzyme DCP2 remain poorly understood. Here, we reconstitute the human decapping network using full-length recombinant proteins and compare it to the yeast system. Unlike in yeast, we find that the C-terminal region of human DCP2 is not autoinhibitory. RNA-binding residues of yeast Dcp2 are not conserved in the human homolog, and we find instead that a charged C-terminal region mediates substrate recognition. Human DCP1 does not stably interact with or directly stimulate DCP2, but mediates activation by the enhancer PNRC2. We also demonstrate that decapping enhancer EDC4 forms tetramers through an extended coiled-coil region, and that both DCP1 and EDC4 homomeric species can further assemble into higher-order oligomers. Furthermore, structural predictions incorporating these findings suggest a model for DCP2 recruitment by EDC4 tetramers. These findings reveal key mechanistic differences between human and yeast decapping regulation and provide insight into the molecular architecture underlying mRNA decay.
Similar content being viewed by others


Introduction
Gene expression levels depend on a careful balance between mRNA synthesis and decay. Steps in mRNA degradation provide critical regulatory points in cell division1 and differentiation2 and allow for dynamic responses to external stimuli3,4 and viral infection5. Removing the 5′ cap structure, or decapping, is a decisive and penultimate step in the 5′-to-3′ mRNA decay pathway. Decapping commits transcripts to rapid degradation by the exoribonuclease XRN16,7. In eukaryotes, this decapping step is carried out by the Nudix hydrolase DCP28,9,10. Unlike many cap-binding factors, which directly recognize the 5′ cap structure, such as eIF4E11, DCP2 relies on interactions with the RNA body rather than the cap itself for substrate recognition9,10.
Given the decisive nature of decapping in mRNA decay, it is a tightly regulated step. A network of decapping enhancers mediates this regulation through direct modulation of DCP2 catalytic activity and recruits it to target transcripts12,13,14,15. A key component of this enhancer network is DCP1, which interacts tightly with DCP2 in fission yeast and mediates stimulation by enhancers such as Edc1 and Edc212,16. Yeast Dcp1 consists solely of a structured EVH1 domain, which binds stably to the N-terminal domain of Dcp2 and interacts with decapping enhancers such as Edc112,16 (Fig. 1a). Metazoan DCP1, however, consists of a homologous EVH1 domain as well as an extended C-terminal sequence (Fig. 1a). This C-terminal extension is predicted to be largely disordered, except for a helical motif shown to bind the decapping enhancer EDC3 in Drosophila melanogaster14, and a small helical bundle at the C-terminus which is thought to mediate homotrimerization17. Evidence suggests that a stable interaction between DCP1 and DCP2 may not be preserved in metazoa, although DCP1 retains a vital role in decapping regulation16,18. Current understanding of the DCP1–DCP2 interaction is based largely on studies of colocalization and the activity of immunopurified complexes, which could potentially include unidentified factors.

a Schizosaccharomyces pombe Dcp1 (Sp Dcp1) consists solely of an EVH1 domain (green). Human DCP1 (Hs DCP1) retains this EVH1 domain but is followed by a long intrinsically disordered region (IDR, light gray) that contains a short helical-leucine motif (HLM; black bar) and ends in a trimerization domain (TD; green). Hs DCP1(1–135) construct was truncated to include only the N-terminal EVH1 domain. b Yeast and human decapping enzymes share conserved N-terminal regulatory (NRD, cyan) and catalytic Nudix (dark blue) domains, followed by species-specific C-terminal IDRs that harbor short linear motifs: multiple HLMs in Sp Dcp2 (black bars) and a phenylalanine-rich EDC4-binding (FEB) motif in Hs DCP2 (dark gray bar). Constructs include Sp Dcp2(1–242) (structured core only), Sp Dcp2(1–504) (core plus partial IDR), full-length Hs DCP2 (1–420), and the core construct Hs DCP2(1–245). c The intrinsically disordered activator Sp Edc1 and its functional analog Hs PNRC2 each contain a single decapping-activator motif (DAM; magenta bar). d Human and Caenorhabditis elegans EDC4 share an N-terminal WD40 β-propeller domain (orange) followed by an IDR (gray) and a C-terminal α-helical region that can be divided into proximal (yellow) and distal (red) segments. The nematode protein has a shorter central IDR but preserves the same overall organization. Residue numbers above schematics denote domain boundaries; numbers at the right indicate the final residue (length) of each full-length protein or truncation construct.
While the N-terminal helical and Nudix domains of DCP2 are conserved between yeast and human homologs, the regions C-terminal to the Nudix domain differ significantly (Fig. 1b). Both yeast and human C-terminal segments are predicted to be intrinsically disordered regions (IDRs) containing short functional motifs (Fig. 1b). The C-terminal segment of yeast Dcp2 contains binding elements for decapping enhancers such as Edc3 and Scd614 and for substrate targeting via Upf1 and Pat115. While these motifs are not conserved in the human DCP2 ortholog, some of these functions are likely retained by other components of the human decapping network14,19. In yeast Dcp2, autoinhibitory motifs have also been identified in the C-terminal IDR20,21.
Yeast Edc1 and Edc2 are known stimulators of decapping activity22,23. Dcp1 mediates this stimulation by binding a proline-rich sequence (PRS) in Edc1/2 through its EVH1 domain12,24,25. Dcp1 can also recruit other non-stimulatory decay factors containing similar PRS motifs24,26. While there are no known orthologs of Edc1/2 in metazoans, the vertebrate-specific nonsense-mediated decay (NMD) factor PNRC2 (Fig. 1c) contains a PRS that binds yeast and human DCP1 in a manner analogous to the yeast Dcp1:Edc1 interaction27,28,29. PNRC2 is thought to couple NMD and decapping based on the observation that tethering PNRC2 to reporter mRNA induces decay only when PNRC2 is capable of interacting with DCP127,28. Although a truncated mutant of PNRC2 can stimulate the activity of C-terminally truncated DCP2 in the presence of the DCP1 EVH1 domain28, it is not known whether this synergy occurs with full-length wild-type proteins.
The metazoan-specific decapping enhancer EDC4 is thought to serve as a binding scaffold for DCP2 and decapping regulators13,30,31 and to facilitate 5′-to-3′ degradation, the final decay step, by recruiting XRN118,26,31. EDC4 is also a critical component of P-bodies, cytoplasmic condensates containing mRNA decay factors as well as translationally repressed transcripts32. EDC4 consists of an N-terminal WD40 domain and a C-terminal helical region connected by an extended low complexity region (Fig. 1d). The C-terminal helical region of EDC4 mediates self-association and interactions with XRN1 and DCP218,30,33,34. A phenylalanine-rich EDC4-binding (FEB) motif in DCP2’s C-terminal IDR mediates interaction with EDC418,34,35, but the mechanism by which EDC4 affects decapping activity remains unknown.
In this work, we investigate the mechanisms of mRNA decapping regulation using a reconstituted system of recombinant purified components to study the eukaryotic decapping network. This system includes several key decapping factors, characterized for the first time in their full-length recombinant forms. Our findings reveal a divergence in the function of the DCP2 C-terminal sequence between yeast and human homologs, with no direct stimulation of DCP2 by DCP1 observed. However, the role of DCP1 as a mediator of enhancer-driven stimulation is conserved, with full-length oligomeric DCP1 facilitating the stimulation of DCP2 by full-length PNRC2. Additionally, we demonstrate that EDC4 forms coiled-coil-based tetramers that can self-associate into megadalton-scale oligomers and provide a model for DCP2 recruitment consistent with the tetrameric state of EDC4.
Results
Divergent roles of human and yeast DCP2 C-terminal regions
To explore the roles of divergent DCP2 C-terminal sequences, we compared full-length human, Hs (Homo sapiens), DCP2 with a yeast, Sp (Schizosaccharomyces pombe), Dcp2 construct containing known autoinhibitory motifs. Because S. pombe Dcp2 is considerably larger and contains extensive low-complexity regions that limit recombinant expression and sample homogeneity, we performed cross-species comparisons using a large S. pombe Dcp2 fragment encompassing the conserved catalytic core and a segment of the C-terminal low-complexity region (residues 1–504), which retains decapping activity in vitro and has been used previously for biochemical studies21; full-length human DCP2 is comparatively tractable and was therefore used throughout. Unlike previous studies, we directly assessed both homologs under uniform conditions without decapping enhancers. Decapping activity was measured using an established in vitro assay that detects the cleavage of radiolabeled cap from an in vitro transcribed RNA substrate (Supplementary Table 1). We found that human DCP2 exhibited higher basal activity than the yeast homolog (Fig. 2a). Autoinhibitory motifs in the yeast Dcp2 C-terminal sequence were previously identified but have only been studied in the context of copurified Dcp1/2 complexes21. To test if autoinhibition occurs in the absence of Dcp1, we compared the activity of yeast Dcp2(1–504) with a C-terminally truncated construct, Dcp2(1–242). Consistent with the Dcp1/2 complex, deleting the yeast Dcp2 C-terminal region increased activity significantly (Fig. 2b), reaching levels comparable to full-length human DCP2.

a In vitro decapping assays were performed with full-length Hs DCP2 and Sp Dcp2(1–504). Aliquots were quenched at the indicated times (0–60 min), products and substrate were separated by PEI-cellulose TLC (left), and the percent decapped RNA was quantified (right). Data points represent the mean ± SEM of three independent experiments; error bars that are smaller than the symbols are obscured. b Decapping activity of Sp Dcp2(1–504) (structured core + partial IDR) was compared with the catalytic core Sp Dcp2(1–242). Assays and quantification were carried out as in panel (a) (n = 3). c Full-length Hs DCP2 was assayed alongside the truncated core construct Hs DCP2(1–245). Reaction conditions and analysis were identical to those in panel (a) (n = 3). In all panels, solid lines show single-exponential fits to the averaged data, and filled circles mark substrate (●) or m⁷GDP product (○) positions on TLC plates.
We next tested whether removing the C-terminal region of human DCP2 would similarly enhance activity. Contrary to expectations, C-terminal truncation of human DCP2 significantly decreased activity compared to the full-length enzyme (Fig. 2c). These results indicate a fundamental divergence in the roles of DCP2 C-terminal regions.
Eukaryotic DCP2 orthologs cannot bind and hydrolyze cap structures directly and instead rely on recognition of the adjacent RNA moiety9,10,36,37,38. Substrate binding by yeast Dcp2 has been attributed to a series of critical lysines on the BoxB helix near the C-terminal end of the Nudix domain16,25, but these lysines are not conserved in human DCP2 (Fig. 3a). Observing that the human C-terminal region is enriched in positively charged residues (Supplementary Fig. 1a), we wondered whether the reduced activity of truncated human DCP2 could be a result of decreased interaction with substrate. To explore the involvement of C-terminal IDRs in substrate recognition, we tested the RNA-binding activity of the DCP2 constructs used in the initial decapping experiments using an electrophoretic mobility shift assay (EMSA) (substrate in Supplementary Table 1) (Fig. 3b). Removing the C-terminal region of yeast Dcp2 did not reduce RNA binding, indicating that the structured N-terminal domains are sufficient for substrate recognition. The shift in migration of bound RNA seen after removing the C-terminal IDR represents a reduced portion of the protein-RNA complex retained in the well. In contrast, truncating human DCP2 nearly eliminated RNA-binding activity, suggesting that the charge-enriched IDR plays a critical role in substrate recognition. The C-terminally truncated Hs DCP2 preserves the Nudix catalytic/regulatory core needed for transient 5′-cap engagement but lacks the positively charged C-terminal tail that confers high-affinity RNA-body binding. This may explain why this construct fails to form the stable complexes detected by EMSA yet remains capable of cap cleavage in decapping assays.

a Left, crystal structure of the Sp Dcp2 Nudix domain (PDB 2QKM) with the four lysines that mediate Box B RNA binding (Lys230, Lys231, Lys234, Lys235) shown as blue sticks. Right, AlphaFold model of the human DCP2 Nudix domain (AF-Q8IU60-F1) with the residues occupying the equivalent positions (Arg236, Asp237, Ser240, Arg241) rendered as sticks. Backbones are colored slate in both views. b Electrophoretic mobility-shift assays (EMSA) were performed with a 5′-labeled Rrp41 2 × DE RNA probe and increasing concentrations (0, 0.25, 1.0 µM) of the indicated DCP2 constructs. Full-length Sp Dcp2(1–504) and human DCP2 produce robust shifts, whereas their catalytic cores, lacking most or all of the C-terminal IDR, Sp Dcp2(1–242) and Hs DCP2(1–245), display markedly reduced binding. The gel shown is representative of three independent experiments.
We next used the IDR sequence analysis tool flDPnn39 to predict RNA-binding propensity along the length of the human DCP2 C-terminal IDR (Supplementary Fig. 1b). This analysis identified several segments of the DCP2 C-terminal IDR with high propensities for RNA binding. This finding marks a significant divergence in the basic mechanisms underlying decapping activity, suggesting the human C-terminal IDR has an emergent role in substrate recognition. To further explore how the human C-terminal region might facilitate substrate recognition, we used RoseTTAFold2NA40 to generate structural predictions of full-length human DCP2 in complex with RNA substrate (Supplementary Fig. 1c–e). The highly charged IDR was consistently predicted to interact with RNA, which is sandwiched between the charged IDR and a surface formed by both the Nudix and NRD domains (Supplementary Fig. 1f).
Trimerization dynamics and interaction specificity of human DCP1
To further investigate the mechanisms of decapping regulation, we produced recombinant human DCP1 in its full-length form for the first time. Previous work reported trimerization of an isolated peptide sequence from the C-terminal region of DCP1, termed the trimerization domain (TD)17 (Fig. 1a). To determine whether trimerization also occurs in the context of full-length DCP1, we used mass photometry, which directly estimates the molecular mass of individual protein particles in solution. DCP1 particles were primarily ~190 kDa, consistent with the theoretical mass of trimers (Fig. 4a, upper). To test whether the previously described TD was sufficient to mediate trimerization, we generated a DCP1 construct with mutations that prevent trimerization of the TD peptide17. Mass photometry measurements of the DCP1 TD mutant (TDm) indicated a primarily monomeric form (Fig. 4a, lower), confirming the importance of TD interactions in DCP1 trimerization. For this mutant, we also detected a minor but distinct population of particles matching the mass of DCP1 dimers (~130 kDa), suggesting homomeric interactions distinct from trimerization may occur. Size-exclusion chromatography revealed that wild-type DCP1 eluted as a single peak at 118–120 ml, indicative of an exclusive trimeric species. In contrast, the trimer-disrupting mutant (DCP1 TDm) produced an additional significant peak at 145 ml, consistent with a substantial fraction of the species shifting to a monomeric state (Supplementary Fig. 2a, b). Fractions across both peaks were analyzed by SDS-PAGE to verify the protein integrity. We examined earlier-eluting gel filtration fractions to test for the presence of additional oligomers for DCP1. Within an earlier-eluting wild-type DCP1 fraction, we found populations matching the molecular weights expected for trimers, hexamers, and nonamers (Fig. 4b), indicating that DCP1 trimers can further associate to form higher-order oligomers. An earlier-eluting DCP1-TDm fraction contained monomers, dimers, and a small population of trimers (Fig. 4c).

a Mass photometry histograms show that wild-type Hs DCP1 (red) is almost exclusively trimeric, with a major peak at 187 kDa (gray band, “Trimer”) and negligible monomer signal. In contrast, the TD mutant (teal) shifts to a predominant monomer peak at 59 kDa (gray band, “Monomer”) plus a minor dimer shoulder at ~129 kDa. Representative replicate from n = 3 independent measurements. Peak labels denote estimates of apparent masses and may deviate modestly from exact integer multiples due to calibration/peak-picking. b Expanding the mass window reveals additional peaks at ~188 kDa (trimer), 387 kDa (hexamer), 587 kDa (nonamer), and 786 kDa (dodecamer), indicating stepwise self-association of trimers. Representative replicate, n = 3. c The mutant displays peaks at ~60 kDa (monomer), 130 kDa (dimer), and 205 kDa (trimer), but lacks species above 300 kDa, confirming that disruption of the TD suppresses higher-order assembly. Representative replicate, n = 3. In all panels, histograms are plotted as relative frequency versus calculated molecular weight, and shaded gray regions mark the expected mass windows for monomers and trimers.
In contrast to the stable complex formed between yeast Dcp2 and Dcp116,25, immunoprecipitation (IP) experiments suggest the interaction between human DCP2 and DCP1 is relatively weak13,18. Structural characterization of the yeast Dcp2:Dcp1 heterodimer16 revealed residues critical to the interface, which are not conserved in human DCP1 (Supplementary Fig. 3). To understand how DCP1 influences DCP2 in the human system, we directly compared the yeast and human interactions in the absence of enhancers. We used a streptavidin-based pulldown assay to immobilize yeast and human DCP2 constructs and tested their ability to capture purified yeast and human DCP1, respectively. As expected, yeast Dcp2 efficiently captured Dcp1, with both truncated (1–242) and extended (1–504) constructs capturing Dcp1 to a similar extent (Fig. 5a). Consistent with prior IP results and the lack of interface conservation, neither full-length nor truncated (1–242) human DCP2 captured DCP1 (Fig. 5b). Known interactor PNRC227,28 efficiently captured DCP1, confirming the integrity of purified DCP1 (Fig. 5b).

a StrepII-tagged Sp Dcp2(1–504), Sp Dcp2(1–242), or maltose-binding protein (MBP) were immobilized on StrepTactin resin and incubated with purified Sp Dcp1. Bound material was eluted, resolved by SDS-PAGE, and visualized with Coomassie blue. The input lane shows Sp Dcp1 used as prey (representative gel from n = 3). b Pulldowns were performed as in (a) using StrepII-tagged full-length Hs DCP2, the catalytic core Hs DCP2(1–245), MBP, or MBP-Hs PNRC2 as bait and purified Hs DCP1 as prey (representative gel, n = 3). c Mass photometry of Sp Dcp2(1–504) alone (top, orange) reveals a major monomer peak (101 kDa) and a minor dimer shoulder. Addition of equimolar Sp Dcp1 (bottom, navy) shifts the distribution to slightly higher masses (119 kDa, 240 kDa), consistent with formation of a heterodimer of one Dcp2 and one Dcp1. Shaded regions mark expected monomer and dimer windows (representative trace, n = 3). d Mass photometry histograms of Hs DCP1 alone (top, red) show a single trimeric species (~184 kDa). Hs DCP2 alone (middle, orange) is monomeric (~91 kDa). A 1:1 mixture (bottom, navy) yields two populations: the DCP2 monomer and a species at ~180 kDa (pink shading) that matches the DCP1 trimer; no discrete DCP1–DCP2 complex is detected (representative trace, n = 3). Because each trimeric DCP1 particle contains three polypeptides, its particle count is lower than the equimolar DCP2 monomer. e Decapping assays were performed with full-length Hs DCP2 in the absence (blue) or presence (magenta) of equimolar Hs DCP1. Reactions were quenched at the indicated times (0–60 min), products were resolved by TLC, and the percentage of mRNA decapped was quantified. Data are mean ± SEM of three independent experiments (n = 3) with single-exponential fits. f Decapping kinetics of Sp Dcp2(1–242) (cyan) and Sp Dcp2(1–504) (light red) were measured with or without equimolar Sp Dcp1 (dotted lines). Plots show mean ± SEM of three independent experiments (n = 3) with single-exponential fits.
We further examined these interactions using mass photometry. In isolation, yeast Dcp2(1–504) existed primarily as a monomer, but a small population of homodimers was also detected (Fig. 5c, upper). Although yeast Dcp1 is below the detection limit when measured alone, combining yeast Dcp1 with Dcp2(1–504) led to an increase in mass consistent with heterodimer formation (Fig. 5c, lower). Masses of Dcp2(1–504) monomers and homodimers increased by masses consistent with one and two Dcp1 molecules, indicating that both monomeric and homodimeric forms of yeast Dcp2 are capable of binding Dcp1. In isolation, human DCP1 and DCP2 were in trimeric and monomeric forms, respectively (Fig. 5d, upper and middle). In contrast to the yeast proteins, no shift in mass was observed upon combining human DCP1 and DCP2 (Fig. 5d, lower), indicating that no heterocomplexes were formed. Consistent with our pulldown results, these data confirm that recombinant human DCP1 and DCP2 do not form a stable complex.
We next tested whether human DCP1 directly stimulates decapping activity by adding it to human DCP2 in decapping assays. We found that human DCP1 did not increase decapping beyond the basal activity of human DCP2 (Fig. 5e, Supplementary Fig. 4a). Neither impairing trimerization nor truncating DCP1 down to the EVH1 domain (1–135) altered the effect of DCP1 on DCP2 (Supplementary Fig. 4b, c). Similarly, adding yeast Dcp1 caused no increase in the activities of yeast Dcp2(1–242) or Dcp2(1–504) (Fig. 5f, Supplementary Fig. 4d, e). Additionally, activities of copurified yeast Dcp1/2 complexes were no higher than the basal activities of the corresponding Dcp2 constructs (Supplementary Fig. 4f).
To determine whether Sp Dcp1-mediated stimulation of Sp Dcp2 is metal-dependent, we removed MnCl₂ and retained MgCl₂ as the sole divalent cation in the reaction buffer. We next performed the assays in Mg²⁺-only buffer to see whether the higher baseline activity with Mn²⁺ had masked any Dcp1-specific stimulation. As expected, the overall decapping rate reduced in Mg²⁺, prompting further investigation of whether the Mn²⁺-enhanced turnover could be concealing a stimulatory effect of Dcp1 on Dcp2 (Supplementary Fig. 5a). We further lowered the Dcp2 concentration to 100 nM, quenched reactions at earlier time points, and supplemented Dcp1 at either equimolar or 10-fold molar excess. Although overall turnover decreased in Mg²⁺, Dcp1 again produced no stimulation in equimolar ratio or excess (Supplementary Fig. 5b). Thus, any catalytic enhancement conferred by Dcp1 under these conditions appears to be minimal, and its absence is not an artifact of Mn²⁺-dependent acceleration.
DCP1 mediates stimulation of human DCP2 by PNRC2
Since DCP1 does not directly activate human DCP2, we explored whether it mediates DCP2 stimulation by decapping enhancers. We produced recombinant full-length PNRC2, which was proposed to function analogous to yeast enhancers Edc1 and Edc228, and assessed its effect on DCP2 activity with and without full-length DCP1 (Fig. 6a, Supplementary Fig. 6a). In the presence of DCP1, PNRC2 strongly stimulated decapping activity. However, adding PNRC2 in the absence of DCP1 decreased activity. Since PNRC2 directly interacts with RNA (Supplementary Fig. 6b), this reduction may result from RNA sequestration by PNRC2.

a Decapping reactions were performed with Hs DCP2 and equimolar Hs DCP1, Hs PNRC2, or both cofactors. Aliquots were quenched at 0–60 min, products were resolved by TLC, and the percentage of substrate decapped was quantified. Plotted values are mean ± SEM of three independent experiments (n = 3); solid lines are single-exponential fits. b An alignment of the Edc1 activation segment (residues 145–188) and the corresponding PNRC2 region (residues 80–125) highlights the conserved YAGxxF decapping-activator motif (DAM, orange bar) and an adjacent proline-rich segment (PRS, maroon bar). Strictly conserved residues are shown in dark gray; functionally conserved are in light gray. c Decapping kinetics were measured for Hs DCP2 + DCP1 alone (baseline) and with either full-length Hs PNRC2, a synthetic DAM peptide, or a synthetic PRS peptide. Data were collected and fitted as in panel (a).
In yeast, the association of Edc1 with Dcp1 is necessary but not sufficient to stimulate the Dcp2:Dcp1 complex. The full decapping activating motif (DAM) of Edc1 includes a proline-rich sequence (PRS), which mediates the interaction with Dcp1, as well as a short segment N-terminal to the PRS. This segment contains a ‘YAG’ motif which binds the groove between the Dcp2 NRD and Nudix domains and is thought to stabilize an active conformation which positions the 5′-cap optimally in the active site25,41,42. Human PNRC2, as well as PNRC1, both contain a DAM that closely resembles the Edc1 DAM, including a YAG motif N-terminal to the PRS43 (Fig. 6b).
To determine whether PNRC2 stimulates human DCP2 in a manner mirroring the yeast system, we compared the effect of PNRC2 PRS and DAM peptides to stimulation by full-length PNRC2 (Fig. 6c, Supplementary Fig. 6c). The effect of the PNRC2 DAM peptide matched that of full-length PNRC2, whereas stimulation by the PRS peptide lacking the YAG motif was impaired. These findings demonstrate that DCP1 can mediate DCP2 stimulation by decapping enhancers and that the mechanism of stimulation by PNRC2 in humans mirrors that of Edc1 in yeast.
The C-terminal region of EDC4 forms a tetrameric coiled-coil interface for DCP2
EDC4, a critical component of P-bodies, is thought to regulate the recruitment of DCP2 and other decay factors, such as the 5′-to-3′ exonuclease XRN130,33,44. The C-terminal region of EDC4 contains an extended α-helical region necessary for self-association in cells30,33. The C-terminal ‘distal’ region of the Drosophila melanogaster EDC4 homolog has been structurally characterized33, assuming a globular fold which consists of a bundle of α-helices and resembles ARM- and HEAT-repeat proteins. The ‘proximal’ region of EDC4, located N-terminal to the distal region, has not been structurally characterized.
To better understand how EDC4 influences mRNA decay, we recombinantly expressed and purified the C-terminal region (residues 520–843) of the Ce (Caenorhabditis elegans) EDC4 homolog. Initial mass photometry measurements revealed that this C-terminal region exists primarily in a tetrameric state (Fig. 7a). Prompted by the observation that a small population consistent with the mass of octamers was also visible in initial measurements at 25 nM, increasing the concentration to 300 nM revealed the presence of higher order oligomers including 8-, 12-, 16-, 20-, 24-, and 28-mers (Fig. 7b). These results indicate that EDC4 exists in a base-level tetrameric state which is capable of assembling into larger oligomeric complexes. To explore the basis of tetramer formation, we used AlphaFold2-Multimer45 to predict the structure of C. elegans EDC4 C-terminal region tetramers (Fig. 7c). Consistent with prior structural work, each distal portion was predicted to form a globular bundle of α-helices. The proximal regions, however, were predicted to assemble into an extended parallel tetrameric coiled-coil (Fig. 7d). Similar predictions for human and Arabidopsis thaliana homologs (Supplementary Fig. 7a, b) suggest that coiled-coil formation by C-terminal proximal regions could provide a conserved mechanism for EDC4 tetramer formation. The structure prediction of full-length C. elegans EDC4 revealed that WD40 domains are connected via flexible linkers to the coiled-coil region and are likely to be dynamically flexible with respect to the C-terminal region (Supplementary Fig. 7c).

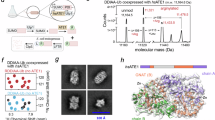
a, b Mass photometry histograms for purified Ce EDC4 residues 520–843 reveal a single species at ~315 kDa at 25 nM (a), consistent with a tetramer, and successive, equally spaced peaks beginning at ~303 kDa when the concentration is increased to 300 nM (b), indicating higher-order oligomers that grow in multiples of the tetrameric unit. Representative traces from three independent data sets are shown. c Residues 520–843 form an extended parallel four-helix bundle. The proximal stalk (rainbow-colored from green to yellow) is continuous with the distal α-helical bundle (magenta and cyan). The crystallized segment analyzed in panels (d–f) is bracketed (520–552). d Top, the 33-residue peptide (520–552) with residues resolved in the crystal structure colored blue; the canonical coiled-coil heptad positions (a–g) are annotated below. Bottom, helical-wheel diagrams illustrate the packing of hydrophobic a/d residues (gray) within a parallel tetramer. e Two orthogonal views show the crystal structure of the segment of the parallel four-helix bundle; a/d layer residues are depicted as sticks. f Superposition of the crystal structure (blue) onto the corresponding region of the AlphaFold-Multimer model (rainbow) demonstrates near-perfect register and side-chain packing. g Representative micrograph of negatively stained MBP-tagged Ce EDC4(520–843) with boxed inset (arrows mark individual particles); scale bar, 50 nm. The experiment was repeated independently three times with similar results. h Selected 2D class averages (particle counts and estimated resolutions indicated; scale bar, 51 nm) reveal elongated rod-shaped molecules terminating in a globular MBP density, consistent with a long coiled coil capped by the distal bundle.
To experimentally validate this prediction, we crystallized a peptide from the proximal region of Ce EDC4 (residues 520–552). Molecular replacement revealed four α-helices arranged in a parallel tetrameric coiled-coil configuration, and the structure was refined to 1.4 Å resolution (Fig. 7e). The tetrameric assembly determined by crystallography superimposes well with the AlphaFold2-Multimer prediction (Fig. 7f, Supplementary Fig. 7d, e). To further characterize the architecture of the C-terminal region, we used negative stain electron microscopy to visualize the recombinant C-terminal construct. Micrographs showed extended particles with long filamentous regions flanked by globular domains (Fig. 7g). 2D classification revealed a central filamentous region ~25 nm long and an overall architecture consistent with the predicted tetrameric structure (Fig. 7h). Notably, numerous ‘bent’ particles in the micrographs suggest that the coiled-coil region is flexible.
Previous work has shown that DCP2 associates with the proximal C-terminal region of EDC4 via a phenylalanine-rich EDC4 binding (FEB) motif in the DCP2 C-terminal IDR18. To explore how EDC4 tetramers might interact with DCP2, we used AlphaFold2-Multimer to predict the structure of the EDC4 C-terminal region in complex with the C-terminal IDR of DCP2. Predictions for both Hs and Ce homologs showed the FEB motif of DCP2 binding along the surface of the EDC4 tetramer (Fig. 8a, b). These predictions locate key FEB motif phenylalanines within a cleft formed by EDC4 tetramerization. To test this interaction, we conducted a pulldown assay using the C-terminal region of Ce DCP2 (residues 745–786), predicted to mediate binding. This peptide successfully captured the C-terminal α-helical region of EDC4 (Fig. 8c), supporting the notion that the FEB motif of DCP2 interacts with the EDC4 tetrameric coiled-coil. Together, these findings suggest a conserved mechanism where the FEB motif binds into a groove assembled by the tetrameric proximal α-helical coiled-coil, explaining the critical role of FEB phenylalanines in the DCP2-EDC4 interaction.

a An AlphaFold-Multimer was used to place the C-terminal IDR of Hs DCP2 (residues 393–420; red) onto the tetrameric coiled coil of the Hs EDC4 C-terminal α-helical region (residues 945–1250; four protomers colored cyan, yellow, magenta, and orange). The boxed region is enlarged (right) and rendered as a molecular surface (left inset) to highlight an extended phenylalanine-rich stretch in DCP2 (F399–L420) that packs against the hydrophobic groove formed by two adjacent EDC4 helices. b A corresponding AlphaFold-Multimer prediction shows the Ce DCP2 C-terminal fragment (residues 745–786; red) bound to the Ce EDC4 coiled coil (residues 520–697). The inset (left) reveals a similar array of bulky hydrophobics anchoring the interaction, consistent with the mechanism inferred from the human model. c StrepII-tagged Ce DCP2(745–786)-SUMO or a SUMO control was incubated with MBP-Ce EDC4(520–843). Pulldown eluates were analyzed by Coomassie-stained SDS-PAGE. MBP-Ce EDC4 co-purifies only with the DCP2 bait, confirming a direct interaction mediated by the predicted FEB segment (representative of n = 3).
It has been hypothesized that EDC4 enhances decapping by binding both DCP1 and DCP2, facilitating DCP1 stimulation of DCP218. To test this, we purified recombinant full-length human EDC4 and conducted decapping assays. Contrary to the hypothesis, EDC4 did not stimulate DCP2 activity, whether DCP1 was present or not (Supplementary Fig. 8a, b). Instead, EDC4 addition to the decapping reaction consistently decreased activity. We then tested whether EDC4 can potentiate the DCP1/PNRC2-mediated stimulation of DCP2. Under conditions where PNRC2 robustly stimulates DCP2/DCP1, we observed that EDC4 does not provide additional catalytic enhancement (Supplementary Fig. 8c, d).
In conclusion, we identified significant functional differences between yeast and human DCP2, particularly regarding the roles of the C-terminal IDRs. The human C-terminal region is crucial for RNA substrate binding, whereas the yeast C-terminal region contains autoinhibitory motifs. Additionally, DCP1 does not directly stimulate DCP2 in either species but mediates stimulation by decapping enhancers. Furthermore, we demonstrated using the Ce ortholog that EDC4 forms a tetrameric structure that interacts with DCP2’s C-terminal FEB motif, suggesting a conserved decapping regulatory mechanism among metazoans. These findings underscore the importance of species-specific adaptations in decapping regulation.
Discussion
Decapping is a crucial regulatory step in 5′-to-3′ mRNA decay, controlled by enhancers that modulate DCP2 activity. While much of our understanding of DCP2 regulation is derived from studies of the yeast system, research on human homologs has largely depended on immunopurified complexes and truncated recombinant proteins. In this study, we produced full-length recombinant human decapping factors, enabling direct in vitro examination of regulatory mechanisms in a compositionally defined system and allowing for comparisons with their yeast counterparts.
Consistent with previous studies, we observe that the yeast Dcp2 C-terminal region exhibits autoinhibitory activity independently of Dcp1, whereas the human DCP2 C-terminal region does not. These findings suggest evolutionary shuffling of functional motifs within the decapping network. For example, while yeast Dcp2 contains an Edc3-binding motif, the human network utilizes a similar motif on DCP1 to recruit EDC346. This redistribution reflects a reorganization of regulatory interactions in higher eukaryotes. Accordingly, other human decapping regulators may possess trans-acting motifs that inhibit DCP2 activity, compensating for the lack of autoinhibitory sequences in human DCP2.
Unlike yeast, we show that the C-terminal region of human DCP2 is essential for full basal decapping activity and RNA substrate interaction. While yeast Dcp2 uses BoxB residues for RNA binding16,25, these residues are absent in human DCP2, and the charged C-terminal IDR appears to compensate. RNA recognition via disordered sequences has been previously observed47, and this mode of interaction could contribute to the generality of DCP2 substrates. Further studies are needed to identify specific regions within the DCP2 IDR involved in RNA binding. Enhancer binding to motifs in this region may regulate substrate recognition, indicating a potential regulatory role. Unexpectedly, EDC4 reduced DCP2 activity, suggesting that EDC4 binding to the DCP2 IDR may interfere with substrate recognition. Consistent with this, EDC4 does not enhance catalysis even when PNRC2 is present, implying its primary role is likely architectural or regulatory rather than catalytic.
Previous studies of recombinant human DCP1 have primarily utilized C-terminally truncated constructs containing only the conserved EVH1 domain. In contrast, we produced full-length recombinant human DCP1 and discovered that it exists predominantly in a trimeric state. This trimerization is mediated by a specific motif known to form trimers in peptide form and to facilitate homomeric interactions in vivo17. Additionally, we observed the formation of higher-order multimers composed of DCP1 trimers and identified homomeric interactions independent of the trimerization motif, which may further contribute to oligomerization. This propensity for oligomerization could play a significant role in the function of DCP1 within P-bodies, as multivalency is a crucial factor in condensation processes, and yeast Dcp1 was recently shown to form condensates48,49. DCP1b, a paralog of DCP1 (alternatively referred to as DCP1a), was recently shown to link the decapping network to protein degradation and translation50. Given the functional non-redundancy of these paralogs, it will be interesting to explore whether the biochemistry of DCP1b mirrors our findings.
In yeast, Dcp1 and Dcp2 form a stable complex, but the interface residues essential for this interaction are poorly conserved in human DCP1. Our experiments confirmed that recombinant human DCP1 and DCP2 do not form a stable complex and showed that DCP1 does not directly stimulate DCP2 activity in vitro. In our reconstituted S. pombe system, Dcp1 alone also does not stimulate Dcp2 with magnesium as a co-factor, which contrasts with the potent activation mediated by Edc1/2. However, because we used MBP-tagged constructs for biochemical tractability, we cannot rule out that the MBP tag alters DCP2’s responsiveness to DCP1. These findings align with immunoprecipitation studies indicating that additional enhancers are necessary to facilitate the in vivo association of DCP1 with DCP218.
Despite the lack of direct interaction, we confirmed that the role of DCP1 as a mediator of DCP2 stimulation by enhancers is conserved in humans. Specifically, we found that PNRC2 stimulates DCP2 activity in a DCP1-dependent manner, mirroring the stimulatory effect of Edc1 observed in yeast. This suggests that PNRC2, which is known to recruit the decapping machinery to NMD targets27, could enhance targeted transcript degradation by stimulating decapping activity. Further analysis revealed that full-length PNRC2 directly interacts with RNA, whereas the DAM motif alone is sufficient to stimulate decapping activity. Although PNRC2’s RNA-binding ability is not necessary for the stimulation of DCP2, it may play a role in other stages of mRNA recruitment or decay processes. This indicates that PNRC2 and PNRC1 may facilitate decapping through multiple mechanisms, contributing to the regulation of mRNA turnover.
EDC4 is an essential component of P-bodies and the decapping network13,30,31,35,44. While it is known that EDC4 self-associates within cells, the specific nature of these interactions has remained unclear. Our biochemical experiments reveal that EDC4 forms tetrameric structures through coiled-coil interactions in the proximal C-terminal α-helical region. Additionally, these tetramers associate to create higher-order oligomers. We detected soluble multimers with megadalton masses, and given the limitations of our detection method, it is plausible that even larger assemblies are formed. These insights advance our understanding of EDC4’s structural organization and its function within the decapping network.
The C-terminal α-helical region of EDC4 is essential for its localization to P-bodies30,51, indicating that coiled-coil interactions play a central role in P-body formation. Electron microscopy analysis has characterized P-bodies as clusters of electron-dense fibrils52. Given the coiled-coil structure and extensive multimerization observed in our studies, these fibrils likely represent higher-order EDC4 assemblies. Coiled-coil-mediated higher-order oligomerization is a known mechanism for facilitating functions in membraneless organelles53. Condensate dynamics are determined by a complex interplay of biophysical properties and therefore can be highly tuned and context dependent54. The wide range of self-associating species we observed suggests EDC4 could play a role in fine-tuning condensate-cytoplasm exchange. These higher-order EDC4 oligomers could also provide a multivalent platform for recruiting other mRNA decay factors, thereby regulating decay by increasing local concentrations or affecting activity through phase separation.
Furthermore, conserved residues within the DCP2 C-terminal FEB motif are crucial for interacting with EDC418,35. Our findings suggest that the aromatic residues of the FEB motif bind within an interface formed by EDC4 tetramerization. Each EDC4 coiled-coil tetramer can theoretically accommodate four FEB motifs, thereby enhancing the multivalency of the EDC4 scaffold and potentially facilitating more efficient recruitment of mRNA decay factors.
To conclude, our study reveals significant differences in decapping regulation between human and yeast systems, particularly concerning the roles of DCP2’s C-terminal region and DCP1. Additionally, we characterized the tetrameric structure of EDC4 and its capacity for higher-order assembly, providing valuable insights into conserved regulatory mechanisms of mRNA decay across metazoan species. Collectively, these findings underscore the importance of species-specific adaptations in decapping regulation and open new avenues for investigating the control of mRNA decay.
Methods
Cloning
Plasmids used for protein expression are detailed in Supplementary Table 2. Fragments generated by PCR with Phusion polymerase (NEB) and/or restriction digest were assembled using standard isothermal assembly procedures, and assembled products were propagated in DH5ɑ Escherichia coli.
Baculovirus production
pLIB vectors described in Supplementary Table 2 were transformed into chemically competent DH10EmbacY cells (Geneva Biotech, Switzerland). Positive integrands were selected using blue/white screening, and bacmid DNA was purified as previously described55. 1–5 μg of bacmid DNA was transfected into 1.0 × 106 Sf21 cells (a kind gift from Imre Berger, University of Bristol) using Fugene HD (Promega) and supernatant containing V0 low-titer baculovirus was collected once at least 50% of cells were positive for YFP marker expression. Suspension cultures at 1.0 × 106 cells/mL were infected with 10% (v/v) V0 baculovirus stock and high-titer V1 baculovirus stocks were collected 24 h following proliferation arrest.
Protein expression and purification
Full-length human DCP2 with an N-terminal MBP fusion and C-terminal His6 tag was expressed in BL21 Star (DE3) E. coli, induced with 0.5 mM IPTG for 3 h at 30 °C. Cells were lysed by sonication in a buffer containing 50 mM HEPES/NaOH pH 7.0, 300 mM NaCl, 2.5 mM CHAPS, and 5% (v/v) glycerol, and lysate was clarified by centrifugation. Affinity capture was performed with amylose resin (NEB) and then IMAC using a HisTrap HP column (Cytiva), followed by size exclusion chromatography on a Superdex 200 26/600 column (Cytiva) equilibrated in lysis buffer. Purified protein was concentrated with a 30 K MWCO centrifugal filter unit (MilliporeSigma), flash-frozen, and stored at −80 °C. For pulldowns, full-length human DCP2 with an N-terminal MBP fusion and C-terminal 2xStrepII tag was purified using the same procedure with the IMAC step omitted.
C-terminally-truncated human DCP2 (residues 1–245) with an N-terminal MBP fusion was expressed in BL21 Star (DE3) E. coli and induced with 0.5 mM IPTG for 3 h at 30 °C. Cells were lysed by sonication in a buffer containing 50 mM HEPES/NaOH pH 7.0, 300 mM NaCl, and 5% (v/v) glycerol, and lysate was clarified by centrifugation. Amylose resin (NEB) was used for affinity purification, followed by size exclusion chromatography on a Superdex 200 26/600 column (Cytiva) equilibrated in lysis buffer. Purified protein was concentrated with a 30 K MWCO centrifugal filter unit (MilliporeSigma), flash-frozen, and stored at −80 °C.
Sp Dcp2 including a portion of C-terminal IDR sequence (residues 1–504) with a 3C-cleavable N-terminal MBP fusion and C-terminal 2xStrepII tag was expressed in Sf21 cells. Cells were infected with recombinant Sp Dcp2(1–504) baculovirus stock (1:100 v/v) and harvested 48 h following proliferation arrest. Cells were lysed by sonication in a buffer containing 50 mM Tris/HCl pH 7.5, 300 mM NaCl, 2.5 mM CHAPS, 5% (v/v) glycerol, and 1 mM TCEP. Lysate was clarified by centrifugation and filtered through 0.45 μm syringe-driven filters (MilliporeSigma) before affinity purification with a StrepTrap HP column (Cytiva). Protein was eluted in lysis buffer supplemented with 2.5 mM desthiobiotin, concentrated with a 30 K MWCO centrifugal filter unit (MilliporeSigma), flash-frozen, and stored at −80 °C. C-terminally truncated Sp Dcp2 (residues 1–242) with an N-terminal MBP fusion and C-terminal 2xStrepII tag was expressed in BL21 Star (DE3) E. coli, induced with 1 mM IPTG for 16 h at 20 °C. Sp Dcp2(1–242) was purified using the same procedure as Sp Dcp2(1–504) with the concentration step omitted.
Sp Dcp1 with a C-terminal His6 tag was expressed in BL21 Star (DE3) E. coli, induced with 0.5 mM IPTG for 3 h at 30 °C. Cells were lysed by sonication in a buffer containing 50 mM HEPES/NaOH pH 7.0, 500 mM NaCl, 2.5 mM CHAPS, 5% (v/v) glycerol, and 20 mM imidazole. Lysate was clarified by centrifugation and filtered through 0.45 μm syringe-driven filters (MilliporeSigma) before IMAC purification with a HisTrap HP column (Cytiva). Following elution, a HiPrep 26/10 desalting column (Cytiva) was used for buffer exchange into storage buffer containing 50 mM HEPES/NaOH, pH 7.0, 500 mM NaCl, 2.5 mM CHAPS, and 5% (v/v) glycerol before the protein was flash-frozen and stored at −80 °C.
Copurified complexes Sp Dcp1:Sp Dcp2(1–242) and Sp Dcp1:Sp Dcp2(1–504) were produced by co-expression of Sp Dcp1 with each Sp Dcp2 construct in BL21 Star (DE3) E. coli, induced with 0.5 mM IPTG for 16 h at 16 °C. Cells were lysed by sonication in a buffer containing 50 mM HEPES/NaOH, pH 7.5, 300 mM NaCl, 2.5 mM CHAPS, 5% (v/v) glycerol, 20 mM imidazole, and 1 mM TCEP. Lysate was clarified by centrifugation and filtered through 0.45 μm syringe-driven filters (MilliporeSigma) before IMAC purification with a HisTrap HP column (Cytiva). Complexes were further purified by size exclusion chromatography using a Superdex 200 26/600 column (Cytiva) equilibrated in storage buffer containing 50 mM HEPES/NaOH, pH 7.5, 300 mM NaCl, 2.5 mM CHAPS, 5% (v/v) glycerol, and 1 mM TCEP before protein was flash-frozen and stored at −80 °C.
Full-length Hs DCP1 (alternatively referred to as DCP1a) with an N-terminal His6 tag was expressed in Sf21 cells. Cells were infected with recombinant DCP1 baculovirus stock (1:100 v/v) and harvested 48 h following proliferation arrest. Cells were lysed by sonication in a buffer containing 50 mM Tris/HCl, pH 8.0, 500 mM NaCl, 2.5 mM CHAPS, 5% (v/v) glycerol, and 20 mM imidazole. Lysate was clarified by centrifugation and filtered through 0.45 μm syringe-driven filters (MilliporeSigma) before IMAC purification with a HisTrap HP column (Cytiva). Protein was further purified by size exclusion chromatography using a Superdex 200 26/600 column (Cytiva) equilibrated in storage buffer containing 50 mM HEPES/NaOH, pH 7.0, 300 mM NaCl, 5% (v/v) glycerol, and 2 mM TCEP before flash-freezing and storage at −80 °C. The DCP1-TDm mutant was expressed and purified using the same procedure as the wild-type construct.
C-terminally-truncated Hs DCP1 (residues 1–135) with a 3 C protease-cleavable N-terminal MBP fusion was expressed in BL21 Star (DE3) E. coli, induced with 1 mM IPTG for 3 h at 30 °C. Cells were lysed by sonication in a buffer containing 50 mM HEPES/NaOH, pH 7.5, 500 mM NaCl, 2.5 mM CHAPS, 5% (v/v) glycerol, and 1 mM TCEP, and the lysate was clarified by centrifugation. Affinity purification was carried out using amylose resin (NEB) and followed by 3C protease cleavage, incubating with 1:30 (w/w) 3C at 4 °C for 16 h. Cleaved DCP1(1–135) was separated from MBP by size exclusion chromatography using a Superdex 200 26/600 column (Cytiva) equilibrated in lysis buffer. Purified protein was concentrated with a 10 K MWCO centrifugal filter unit (MilliporeSigma), flash-frozen, and stored at −80 °C.
Full-length Hs PNRC2 with an N-terminal MBP fusion was expressed in BL21 Star (DE3) E. coli, induced with 1 mM IPTG for 4 h at 30 °C. Cells were lysed by sonication in a buffer containing 50 mM HEPES/NaOH, pH 7.0, 500 mM NaCl, and 5% (v/v) glycerol, and the lysate was clarified by centrifugation. Affinity purification was carried out using amylose resin (NEB) and followed by size exclusion chromatography using a Superdex 200 26/600 column (Cytiva) equilibrated in lysis buffer. Purified protein was concentrated with a 30 K MWCO centrifugal filter unit (MilliporeSigma), flash-frozen, and stored at −80 °C.
Full-length Hs EDC4 with a C-terminal 2xStrepII tag was expressed in Sf21 cells. Cells were infected with recombinant EDC4 baculovirus stock (1:100 v/v) and harvested 72 h following proliferation arrest. Cells were lysed by sonication in a buffer containing 50 mM HEPES, pH 7.0, 300 mM NaCl, 2.5 mM CHAPS, and 5% (v/v) glycerol. Lysate was clarified by centrifugation and filtered through 0.45 μm syringe-driven filters (MilliporeSigma) before affinity purification with a StrepTrap HP column (Cytiva). Protein was eluted in lysis buffer supplemented with 2.5 mM desthiobiotin. Purified protein was flash-frozen, and stored at −80 °C.
N-terminally truncated Ce EDC4 (residues 520–843) with an N-terminal MBP fusion and C-terminal His6 tag was expressed in BL21 Star (DE3) E. coli, induced with 0.5 mM IPTG for 16 h at 16 °C. Cells were lysed by sonication in a buffer containing 50 mM HEPES/NaOH, pH 7.0, 500 mM NaCl, 2.5 mM CHAPS, 10% (v/v) glycerol, 1 mM TCEP, and 25 mM imidazole. Lysate was clarified by centrifugation and filtered through 0.45 μm syringe-driven filters (MilliporeSigma) before IMAC purification with a HisTrap HP column (Cytiva). Following elution, a HiPrep 26/10 desalting column (Cytiva) was used for exchange into storage buffer containing 50 mM HEPES/NaOH, pH 7.0, 500 mM NaCl, 2.5 mM CHAPS, and 10% (v/v) glycerol before the protein was flash-frozen and stored at −80 °C. For electron microscopy, a construct with the His6 tag replaced by a C-terminal 2xStrepII tag was expressed, lysed, and clarified in the same manner as above, but with imidazole omitted from the lysis buffer. Affinity purification was performed using a StrepTrap XT column (Cytiva), eluted in lysis buffer supplemented with 50 mM biotin, flash-frozen, and stored at −80 °C.
In all protein constructs that were produced as fusions with MBP, except for Hs DCP1(1–135), the N-terminal MBP tags were retained.
Decapping assay
The RNA body was produced by in vitro transcription using T7 RNA polymerase (NEB) from a PCR-amplified DNA substrate using the manufacturer’s protocol. The 144 nt product (Supplementary Table 1) was purified by phenol:chloroform extraction followed by ethanol precipitation. Cap was added using Vaccinia Capping Enzyme and mRNA Cap 2′-O-Methyltransferase (NEB) with [α−32P]GTP (PerkinElmer) following the manufacturer’s ‘one-step capping and 2′-O-methylation reaction’ protocol to produce a cap-1 structure with 32P at the γ position of the cap’s triphosphate linkage. Unincorporated nucleotides were removed using Sephadex G-50 spin columns (MilliporeSigma), and the capped substrate was further purified by phenol:chloroform extraction followed by LiCl precipitation. The capped RNA substrate was at ~40 nM (assuming 100% labeling efficiency) in all assays.
Proteins were pre-diluted and, in cases with multiple proteins, combined to 2 μM in 1X decapping buffer consisting of 50 mM Tris/HCl pH 7.5, 50 mM ammonium sulfate, 0.1% (w/v) BSA, and 5 mM MgCl2 and incubated at 4 °C for 15 min. Reactions also contained 5 mM MnCl2 (or MgCl2 where indicated) and were initiated by adding proteins to a 200 nM final concentration (or as indicated). Reactions were carried out at 10 °C, and samples were taken at indicated time points and quenched by adding EDTA to 83 mM final concentration. Before sample application, PEI Cellulose F plates (MilliporeSigma) were pre-run in water and scored in 1 cm increments. 1 μl of quenched timepoint samples were applied, and plates were developed in 0.75 M LiCl. Minor differences in the insertion depth of each plate (±1–2 mm) altered the total solvent-front migration, resulting in a slight apparent offset in band position when the plates were scanned together.
TLC plates were exposed to BAS-IP MS storage phosphor screens (Cytiva) overnight, and screens were imaged using an Amersham Typhoon (Cytiva). Image quantification was performed in ImageJ (NIH), using the Gel Analyzer tool to plot signal intensity by lane. Authentic radiolabeled standards previously run in parallel confirm that the sole product comigrates with m⁷GDP, not m⁷GMP. This is also as expected for DCP2, which cleaves the β-γ phosphoanhydride bond to release m⁷GDP. The product signal from t = 0 lanes was subtracted from the product signal of all lanes, and the percent decapped was calculated relative to the total signal from the product and intact substrate. Percents decapped were plotted by time point, and curves were produced in Prism 9 (GraphPad Software, Inc.) using nonlinear regression with least squares fitting using the ‘One phase exponential association’ equation.
Electrophoretic mobility shift assay
RNA substrate for EMSA experiments consisted of a 60 nt sequence (Supplementary Table 1) from the 5′ UTR of the Rrp41 gene, which was previously found to interact with DCP256,57 and was synthesized commercially with a 3′ 6-FAM label (IDT). Proteins were pre-diluted in EMSA buffer consisting of 20 mM HEPES/NaOH, pH 7.5, 50 mM KCl, and 10% (v/v) glycerol. Binding reactions were conducted for 30 min at 4 °C with 10 nM RNA and 0.25–1.0 μM protein in EMSA buffer supplemented with 5 mM MgCl2, 0.1 mg/mL BSA, and 0.1% (v/v) IGEPAL CA-630. 4% acrylamide (75:1 acrylamide:bisacrylamide) gels were cast and run in a buffer composed of 5 mM disodium borate decahydrate, pH 8.0 and 0.1% (v/v) IGEPAL CA-630. Binding reactions were loaded directly onto gels, run at 150 V for 5 min, and imaged using the Cy2 channel of an Amersham Typhoon (Cytiva).
Pulldown assay
Bait proteins MBP-Sp Dcp2(1–242), MBP-Hs DCP2, MBP-Hs DCP2(1–245), MBP-Hs PNRC2, SUMO, and Ce DCP2(745–786)-SUMO fused to 2xStrepII tags were produced in BL21 Star (DE3) E. coli grown in auto-induction medium overnight at 37 °C. 2xStrepII-tagged MBP-Sp Dcp2(1–504) was expressed in the baculovirus system as described in the section ‘Protein Expression and Purification’. Bacterial pellets from MBP-Sp Dcp2(1–242), MBP-Hs DCP2(1–245), and MBP-Hs PNRC2 expression were lysed by sonication in pulldown buffer containing 50 mM HEPES/NaOH, pH 7.5, 300 mM NaCl, and 2.5 mM CHAPS, and the lysate was clarified by centrifugation. Bait was loaded directly from lysate for all proteins other than 2xStrepII-tagged MBP and MBP-Hs DCP2, where purified proteins were diluted into pulldown buffer. 50 μl of StrepTactin Sepharose resin slurry was added to lysates or diluted bait solutions, and incubated for 1 h at 4 °C before washing three times with pulldown buffer. Purified Sp Dcp1 (40 μg) or Hs DCP1 (80 μg) was diluted in pulldown buffer and incubated with bait-loaded beads for 1 h at 4 °C before washing three times with pulldown buffer. Proteins were eluted with 50 μl of pulldown buffer supplemented with 50 mM biotin for 1 h at 4 °C and resolved by SDS-PAGE. Assays examining the interaction between Ce DCP2 and Ce EDC4 were performed as described above with a pulldown buffer containing 50 mM HEPES/NaOH, pH 7.0, 300 mM NaCl, 1 mM TCEP, and 0.03% (v/v) Tween 20, applying 80 μg of MBP-Ce EDC4(521–843) prey.
Mass photometry
Measurements were acquired with the TwoMP mass photometer (Refeyn), using Refeyn AcquireMP software and calibrated as per the manufacturer’s protocol using β-amylase and thyroglobulin. For measurements of DCP1 oligomerization, proteins were pre-diluted into and measured in a buffer containing 50 mM HEPES/NaOH, pH 7.0, 300 mM NaCl, 5% (v/v) glycerol, and 2 mM TCEP. To examine trimerization of wild-type and TDm mutant DCP1, purified protein stocks were diluted to 100 nM immediately before use, and 5 μl of 100 nM stock was added to 15 μl buffer droplets for measurement. To examine high molecular weight species isolated during size exclusion chromatography, 0.5 μl of void peak samples were diluted directly into 18 μl buffer droplets for measurement. For measurements of DCP2–DCP1 complexes, the buffer contained 50 mM HEPES/NaOH, pH 7.0 and 300 mM NaCl. Binding reactions were prepared in this buffer with proteins at 2 μM and incubated for 30 min at 4 °C. 1 μl of binding reactions were diluted directly into 19 μl buffer droplets for measurement. MBP-Ce EDC4(520–843)-His6 was diluted into and measured in a buffer containing 50 mM HEPES/NaOH, pH 7.0 and 500 mM NaCl. For measurements at 25 nM, 5 μl of freshly diluted 100 nM protein solution was diluted into 15 μl droplets. To measure at 300 nM, 6 μl of 1 μM protein solution was diluted into 14 μl droplets. Frequency distributions were created from event counts in Prism 9 (GraphPad Software, Inc.), tabulating relative frequency with a bin width of 6. Molecular masses shown above peaks in frequency distribution plots were generated through analysis with Refeyn DiscoverMP software.
Structural predictions
The AlphaFold prediction of human DCP2 (AF-Q8IU60-F1) was obtained from the AlphaFold Protein Structure Database and was created with the AlphaFold Monomer v2.0 pipeline58,59. Structural predictions of protein complexes were generated using AlphaFold2-Multimer, and each prediction shown is representative of five or more replicates45. Predictions of DCP2–RNA substrate complexes were generated using RoseTTAFold2NA40. We modeled the RNA substrate using a fragment of Me31B RNA, the substrate used in decapping assays, and Rrp41 2xDE RNA, the substrate used in gel shift assays (sequence provided in Supplementary Table 1), as well as a randomly generated 40-nt RNA sequence. The Ce EDC4 crystal structure alignment with the AlphaFold Multimer prediction was performed using the cealign command in PyMOL (The PyMOL Molecular Graphics System, Version 3.0, Schrödinger, LLC.). Structural images were prepared with UCSF ChimeraX60.
Bioinformatic analysis
Protein sequences were obtained from UniProt. The EMBOSS v6.5.7 tool ‘charge’ was used to calculate charge values with a window size of one. Protein disorder was predicted using IUPred2A61, and the RNA-binding propensity of disordered regions was predicted with flDPnn39. Plots of amino acid charge and disorder score were generated with second-order smoothing and a sliding window size of 30. All plots were generated in Prism 9 (GraphPad Software, Inc.).
Crystallization and structure determination
Crystals of Ce EDC4 (UniProt ID: Q8ITV7) C-terminal proximal region residues 520–552 were obtained at 18 °C using the sitting-drop vapor diffusion method two days after mixing 10 mg/mL peptide solution with crystallization solution containing 3 M ammonium sulfate and 10% glycerol. Crystals were cryoprotected in mother liquor supplemented with 4 M sodium formate and flash-cooled in liquid nitrogen. Data was collected at 100 K on a PILATUS 6 M detector at the PII beamline at the PETRAIII synchrotron in Hamburg, Germany. Diffraction data were processed with XDS and scaled using XSCALE62. Phases were obtained by molecular replacement using PHASER63. An idealized polyalanine helix was used as a search model with an asymmetric unit containing two copies of the model. The molecular replacement solution was used to rebuild the initial model using the PHENIX AutoBuild wizard64. To complete the structure, iterative cycles of model building and refinement were performed with COOT65 and PHENIX66, respectively. Stereochemical properties were verified with MOLPROBITY67, and structural images were prepared with UCSF ChimeraX60. Diffraction data and refinement statistics are summarized in Supplementary Table 3.
Negative-stain electron microscopy
MBP-Ce EDC4(520–843)−2xStrepII was diluted to 25 μg/mL in a buffer containing 50 mM HEPES pH 7.0 and 500 mM NaCl, and 5 μl was applied to freshly glow-discharged 400-mesh copper grids with carbon support (Electron Microscopy Sciences) and incubated at room temperature for 30 s. Grids were rinsed briefly twice, first with water and then 2% (w/v) uranyl acetate (UA), stained with two rounds of 2% (w/v) UA, first for 30 s and then for 1 min, then blotted and air-dried for at least 30 min before imaging. Images were acquired on a Talos L120C TEM with a 4k x 4k Ceta CMOS camera (Thermo Fisher Scientific). Data were acquired using SerialEM at a defocus of −5 μm at 57,000x magnification, corresponding to a pixel size of 4.83 Å/pixel.
All image processing was performed using cryoSPARC on a dataset of 98 micrographs. Blob Picker was used to pick circular and elliptical blobs with a 30–300 Å diameter. Micrographs were extracted with a 256-pixel box size, 148,073 particles were used for 2D classification and poor particles were removed, yielding 30,333 particles in 11 classes. Of these, six representative classes are shown.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Atomic coordinates and structure factors for the reported crystal structure have been deposited with the Protein Data Bank under accession code 9E7Q. Source data for the figures and Supplementary Figs are provided as a Source Data file. The corresponding authors will provide the raw data, additional information, and materials upon request and subject to a completed Materials Transfer Agreement. Source data are provided with this paper.
References
Krenning, L., Sonneveld, S. & Tanenbaum, M. E. Time-resolved single-cell sequencing identifies multiple waves of mRNA decay during the mitosis-to-G1 phase transition. Elife 11, 1–26 (2022).
Battich, N. et al. Sequencing metabolically labeled transcripts in single cells reveals mRNA turnover strategies. Science 367, 1151–1156 (2020).
Rabani, M. et al. Metabolic labeling of RNA uncovers principles of RNA production and degradation dynamics in mammalian cells. Nat. Biotechnol. 29, 436–442 (2011).
Vijjamarri, A. K. et al. Decapping factor Dcp2 controls mRNA abundance and translation to adjust metabolism and filamentation to nutrient availability. Elife 12, e85545 (2023).
Abernathy, E. & Glaunsinger, B. Emerging roles for RNA degradation in viral replication and antiviral defense. Virology 479-480, 600–608 (2015).
Muhlrad, D., Decker, C. J. & Parker, R. Deadenylation of the unstable mRNA encoded by the yeast MFA2 gene leads to decapping followed by 5’–>3' digestion of the transcript. Genes Dev. 8, 855–866 (1994).
Nagarajan, V. K., Jones, C. I., Newbury, S. F. & Green, P. J. XRN 5’→3’ exoribonucleases: structure, mechanisms and functions. Biochim. Biophys. Acta 1829, 590–603 (2013).
Lykke-Andersen, J. Identification of a human decapping complex associated with hUpf proteins in nonsense-mediated decay. Mol. Cell. Biol. 22, 8114–8121 (2002).
van Dijk, E. et al. Human Dcp2: a catalytically active mRNA decapping enzyme located in specific cytoplasmic structures. EMBO J 21, 6915–6924 (2002).
Wang, Z., Jiao, X., Carr-Schmid, A. & Kiledjian, M. The hDcp2 protein is a mammalian mRNA decapping enzyme. Proc. Natl. Acad. Sci. USA 99, 12663–12668 (2002).
Matsuo, H. et al. Structure of translation factor eIF4E bound to m7GDP and interaction with 4E-binding protein. Nat. Struct. Biol. 4, 717–724 (1997).
Borja, M. S., Piotukh, K., Freund, C. & Gross, J. D. Dcp1 links coactivators of mRNA decapping to Dcp2 by proline recognition. RNA 17, 278–290 (2011).
Fenger-Grøn, M., Fillman, C., Norrild, B. & Lykke-Andersen, J. Multiple processing body factors and the ARE binding protein TTP activate mRNA decapping. Mol. Cell 20, 905–915 (2005).
Fromm, S. A. et al. The structural basis of Edc3- and Scd6-mediated activation of the Dcp1:Dcp2 mRNA decapping complex. EMBO J. 31, 279–290 (2012).
He, F., Wu, C. & Jacobson, A. Dcp2 C-terminal cis-binding elements control selective targeting of the decapping enzyme by forming distinct decapping complexes. Elife 11, 1–34 (2022).
She, M. et al. Structural basis of dcp2 recognition and activation by dcp1. Mol. Cell 29, 337–349 (2008).
Tritschler, F. et al. DCP1 forms asymmetric trimers to assemble into active mRNA decapping complexes in metazoa. Proc. Natl. Acad. Sci. USA 106, 21591–21596 (2009).
Chang, C.-T., Bercovich, N., Loh, B., Jonas, S. & Izaurralde, E. The activation of the decapping enzyme DCP2 by DCP1 occurs on the EDC4 scaffold and involves a conserved loop in DCP1. Nucleic Acids Res. 42, 5217–5233 (2014).
Tritschler, F. et al. A divergent Sm fold in EDC3 proteins mediates DCP1 binding and P-body targeting. Mol. Cell. Biol. 27, 8600–8611 (2007).
He, F. & Jacobson, A. Control of mRNA decapping by positive and negative regulatory elements in the Dcp2 C-terminal domain. RNA 21, 1633–1647 (2015).
Paquette, D. R., Tibble, R. W., Daifuku, T. S. & Gross, J. D. Control of mRNA decapping by autoinhibition. Nucleic Acids Res. 46, 6318–6329 (2018).
Dunckley, T., Tucker, M. & Parker, R. Two related proteins, Edc1p and Edc2p, stimulate mRNA decapping in Saccharomyces cerevisiae. Genetics 157, 27–37 (2001).
Schwartz, D., Decker, C. J. & Parker, R. The enhancer of decapping proteins, Edc1p and Edc2p, bind RNA and stimulate the activity of the decapping enzyme. RNA 9, 239–251 (2003).
Wurm, J. P., Overbeck, J. & Sprangers, R. T. heS. pombe mRNA decapping complex recruits cofactors and an Edc1-like activator through a single dynamic surface. RNA 22, 1360–1372 (2016).
Valkov, E. et al. Structure of the Dcp2-Dcp1 mRNA-decapping complex in the activated conformation. Nat. Struct. Mol. Biol. 23, 574–579 (2016).
Braun, J. E. et al. A direct interaction between DCP1 and XRN1 couples mRNA decapping to 5’ exonucleolytic degradation. Nat. Struct. Mol. Biol. 19, 1324–1331 (2012).
Cho, H., Kim, K. M. & Kim, Y. K. Human proline-rich nuclear receptor coregulatory protein 2 mediates an interaction between mRNA surveillance machinery and decapping complex. Mol. Cell 33, 75–86 (2009).
Lai, T. et al. Structural basis of the PNRC2-mediated link between mRNA surveillance and decapping. Structure 20, 2025–2037 (2012).
Mugridge, J. S., Ziemniak, M., Jemielity, J. & Gross, J. D. Structural basis of mRNA-cap recognition by Dcp1-Dcp2. Nat. Struct. Mol. Biol. 23, 987–994 (2016).
Xu, J., Yang, J.-Y., Niu, Q.-W. & Chua, N.-H. Arabidopsis DCP2, DCP1, and VARICOSE form a decapping complex required for postembryonic development. Plant Cell 18, 3386–3398 (2006).
Brothers, W. R., Ali, F., Kajjo, S. & Fabian, M. R. The EDC4-XRN1 interaction controls P-body dynamics to link mRNA decapping with decay. EMBO J. 42, e113933 (2023).
Hubstenberger, A. et al. P-body purification reveals the condensation of repressed mRNA regulons. Mol. Cell 68, 144–157.e5 (2017).
Jinek, M. et al. The C-terminal region of Ge-1 presents conserved structural features required for P-body localization. RNA 14, 1991–1998 (2008).
Bloch, D. B., Nobre, R. A., Bernstein, G. A. & Yang, W.-H. Identification and characterization of protein interactions in the mammalian mRNA processing body using a novel two-hybrid assay. Exp. Cell Res. 317, 2183–2199 (2011).
Erickson, S. L. et al. Competition between decapping complex formation and ubiquitin-mediated proteasomal degradation controls human Dcp2 decapping activity. Mol. Cell. Biol. 35, 2144–2153 (2015).
Deshmukh, M. V. et al. mRNA decapping is promoted by an RNA-binding channel in Dcp2. Mol. Cell 29, 324–336 (2008).
Piccirillo, C., Khanna, R. & Kiledjian, M. Functional characterization of the mammalian mRNA decapping enzyme hDcp2. RNA 9, 1138–1147 (2003).
Steiger, M., Carr-Schmid, A., Schwartz, D. C., Kiledjian, M. & Parker, R. Analysis of recombinant yeast decapping enzyme. RNA 9, 231–238 (2003).
Hu, G. et al. flDPnn: accurate intrinsic disorder prediction with putative propensities of disorder functions. Nat. Commun. 12, 4438 (2021).
Baek, M. et al. Accurate prediction of protein-nucleic acid complexes using RoseTTAFoldNA. Nat. Methods 21, 117–121 (2024).
Mugridge, J. S., Tibble, R. W., Ziemniak, M., Jemielity, J. & Gross, J. D. Structure of the activated Edc1-Dcp1-Dcp2-Edc3 mRNA decapping complex with substrate analog poised for catalysis. Nat. Commun. 9, 1152 (2018).
Wurm, J. P., Holdermann, I., Overbeck, J. H., Mayer, P. H. O. & Sprangers, R. Changes in conformational equilibria regulate the activity of the Dcp2 decapping enzyme. Proc. Natl. Acad. Sci. USA. 114, 6034–6039 (2017).
Gaviraghi, M. et al. Tumor suppressor PNRC1 blocks rRNA maturation by recruiting the decapping complex to the nucleolus. EMBO J. 37, e99179 (2018).
Vidya, E. et al. EDC-3 and EDC-4 regulate embryonic mRNA clearance and biomolecular condensate specialization. Cell Rep 43, 114781 (2024).
Evans, R. et al. Protein complex prediction with AlphaFold-Multimer. bioRxiv https://doi.org/10.1101/2021.10.04.463034 (2022).
Jonas, S. & Izaurralde, E. The role of disordered protein regions in the assembly of decapping complexes and RNP granules. Genes Dev 27, 2628–2641 (2013).
Zeke, A. et al. Deep structural insights into RNA-binding disordered protein regions. Wiley Interdiscip. Rev. RNA 13, e1714 (2022).
Gomes, E. & Shorter, J. The molecular language of membraneless organelles. J. Biol. Chem. 294, 7115–7127 (2019).
Currie, S. L. et al. Quantitative reconstitution of yeast RNA processing bodies. Proc. Natl. Acad. Sci. USA 120, e2214064120 (2023).
Vukovic, I. et al. Non-redundant roles for the human mRNA decapping cofactor paralogs DCP1a and DCP1b. Life Sci. Alliance 7, e202402938 (2024).
Yu, J. H., Yang, W.-H., Gulick, T., Bloch, K. D. & Bloch, D. B. Ge-1 is a central component of the mammalian cytoplasmic mRNA processing body. RNA 11, 1795–1802 (2005).
Yang, Z. et al. GW182 is critical for the stability of GW bodies expressed during the cell cycle and cell proliferation. J. Cell Sci. 117, 5567–5578 (2004).
Lee, M. et al. The structure of human SFPQ reveals a coiled-coil-mediated polymer essential for functional aggregation in gene regulation. Nucleic Acids Res. 43, 3826–3840 (2015).
Folkmann, A. W., Putnam, A., Lee, C. F. & Seydoux, G. Regulation of biomolecular condensates by interfacial protein clusters. Science 373, 1218–1224 (2021).
Raisch, T. et al. Reconstitution of recombinant human CCR4-NOT reveals molecular insights into regulated deadenylation. Nat. Commun. 10, 3173 (2019).
Li, Y., Song, M.-G. & Kiledjian, M. Transcript-specific decapping and regulated stability by the human Dcp2 decapping protein. Mol. Cell. Biol. 28, 939–948 (2008).
Li, Y., Ho, E. S., Gunderson, S. I. & Kiledjian, M. Mutational analysis of a Dcp2-binding element reveals general enhancement of decapping by 5’-end stem-loop structures. Nucleic Acids Res. 37, 2227–2237 (2009).
Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. Nature 596, 583–589 (2021).
Varadi, M. et al. AlphaFold Protein Structure Database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res 50, D439–D444 (2022).
Pettersen, E. F. et al. UCSF ChimeraX: structure visualization for researchers, educators, and developers. Protein Sci 30, 70–82 (2021).
Erdős, G. & Dosztányi, Z. Analyzing protein disorder with IUPred2A. Curr. Protoc. Bioinformatics 70, e99 (2020).
Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 66, 125–132 (2010).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Terwilliger, T. C. et al. Iterative model building, structure refinement and density modification with the PHENIX AutoBuild wizard. Acta Crystallogr. D Biol. Crystallogr. 64, 61–69 (2008).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 66, 486–501 (2010).
Afonine, P. V. et al. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 68, 352–367 (2012).
Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 66, 12–21 (2010).
Acknowledgments
Initial experiments were done in Elisa Izaurralde’s former laboratory at the Max Planck Institute for Developmental Biology, Tübingen, Germany, and we gratefully acknowledge Elisa’s support and encouragement during the project’s initial phase. We acknowledge DESY (Hamburg, Germany) for the provision of experimental facilities at beamline P11, PETRA III. We also thank Sergey Tarasov and Marzena Dyba for support with biophysical measurements, Dan Shi for advice and guidance on electron microscopy, and Catrin Weiler for excellent technical assistance with cloning and insect cell culture. We are also grateful to our colleagues at the RNA Biology Laboratory for support and advice. This study was supported by the Intramural Research Program of the National Institutes of Health (project number 1ZIABC011977 to E.V.) and the Max Planck Society (E.V. and S.M.). The contributions of the NIH authors are considered works of the United States Government. The findings and conclusions presented in this paper are those of the authors and do not necessarily reflect the views of the NIH or the U.S. Department of Health and Human Services.
Author information
Authors and Affiliations
Contributions
E.A.J.S., S.M., T.M.M., and A.L.V. performed the experiments, S.M., E.A.J.S., and E.V. designed the project, E.V. supervised it, and E.A.J.S. wrote the initial draft. All authors contributed to manuscript preparation.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Agnieszka Tudek and the other anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Simko, E.A.J., Muthukumar, S., Myers, T.M. et al. Conserved and divergent features of human mRNA decapping revealed by biochemical reconstitution. Nat Commun 17, 3697 (2026). https://doi.org/10.1038/s41467-026-72177-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-72177-2
