
Abstract
Peripheral nerve injuries beyond 5 cm lack effective treatments. Functional nerve guidance conduits (NGCs) have emerged as transformative tools orchestrating regeneration by leveraging Schwann cell (SC)-centric mechanisms. This review comprehensively analyzes how NGC designs modulate SC behavior through three synergistic axes: physical cues, biochemical signaling, and bioelectric regulation. By enhancing microenvironmental regulation, next-generation NGCs aim to surpass autograft efficacy, offering scalable solutions for functional nerve recovery and improved patient outcomes.
Similar content being viewed by others
Introduction
Peripheral nerve injury (PNI) imposes a substantial clinical burden, with over 200,000 annual cases in Europe and the United States alone1. While peripheral nerves possess intrinsic regenerative capacity, functional recovery in critical gaps (>5cm) remains unpredictable, frequently leading to permanent motor and sensory dysfunction. Current gold-standard treatments—direct end-to-end suturing and autograft transplantation—are limited by donor site morbidity, size mismatch, and graft scarcity2. These shortcomings highlight the pressing demand for innovative strategies to bridge nerve defects and restore functional connectivity.
Polymer-based nerve guiding conduits (NGCs) provide a promising alternative for repairing large areas of nerve defects. NGCs have been extensively used in clinical practice in the last few decades. Studies have found that functional recovery has significantly increased in cases where conduits are used3. By bridging the proximal and distal ends of nerve defects, NGCs not only guide the orderly growth of axon sprouts but also prevent the invasion of harmful cells, such as fibroblasts4. Clinically deployed NGCs are categorized into non-degradable (e.g., silicone) and biodegradable conduits (e.g., collagen, PLGA), with the latter dominating current designs due to their tunable resorption. For example, clinical practice has widely used collagen-based commercial conduits such as NeuroFlex, NeuroMatrix, and NeuroWrap5. The first and second generation NGCs are only designed as tubular structures, and their function is limited to bridging nerve injury gaps and physically guiding axonal growth6. Structurally, the hollow single-lumen tube is sufficient to meet the needs of short gaps (<3cm). However, this type of nerve conduit is ineffective for lesions larger than or equal to 3 cm, as most of them only connect the injured stump without a promising stimulus for nerve regeneration, leading to permanent nerve function impairment7. These conventional NGCs often inadequately replicate the dynamic microenvironment essential for robust regeneration, particularly in modulating Schwann cell (SC) behavior—the master regulators of peripheral nerve repair. As the predominant glial population (90% of nucleated cells) in the peripheral nervous system, SCs orchestrate post-injury repair through phenotypic reprogramming, including debris clearance, neurotrophic factor secretion, and Büngner band formation to guide regenerating axons7,8,9. Although their plasticity in transitioning between repair-supportive and myelinating states is critical for regeneration, conduit designs often fail to exploit this capability10 entirely.
Recent advances in bioengineering have shifted the paradigm toward the third-generation NGCs, NGCs that are multifunctional and actively regulate SC behavior through tailored physical, biochemical, and electrical cues. Innovations such as topographically patterned surfaces, stiffness-tunable matrices, and bioelectric scaffolds now aim to mimic the native nerve extracellular matrix (ECM), enhancing SC adhesion, migration, and axon-glia crosstalk11,12. Emerging ‘smart’ NGCs incorporate electroconductive polymers (e.g., PPy) or stimuli-responsive hydrogels to regulate Schwann cell behavior. Complementary strategies, including decellularized ECM integration, spatiotemporal growth factor delivery, and conductive material incorporation, further synergize SC-mediated repair mechanisms13,14,15. Despite preclinical progress, critical challenges persist, including insufficient spatiotemporal control of bioactive cues, long-term biocompatibility concerns, and limited clinical translation.
This review critically examines the interplay between NGC design and SC functionality through three interconnected axes: (1) Physical cues (surface topography, mechanical stiffness, porous architecture) that direct SC alignment and Büngner band organization; (2) Biochemical modulation via ECM components, decellularized matrices, and controlled growth factor delivery to enhance SC proliferation and neurotrophic signaling; and (3) Bioelectric regulation using conductive polymers and piezoelectric materials to replicate endogenous electrical signaling and synchronize SC-axon interactions (Fig. 1). By integrating these perspectives, we highlight how multifunctional NGCs can amplify SC-driven repair while addressing unresolved challenges in clinical translation. Ultimately, this synthesis aims to bridge interdisciplinary innovations in material science, mechanobiology, and neuroengineering, providing a roadmap for next-generation conduits to restore functional precision in nerve repair.

Physical cues: Aligned topographies (nanofibers, grooves) provide contact guidance for SC migration and Büngner band formation via mechanotransduction pathways (e.g., YAP/TAZ). Stiffness gradients instruct SC phenotypic fate (softness promotes repair state; stiffness promotes redifferentiation). Porous architectures facilitate cell infiltration and nutrient exchange. Biochemical Cues: Spatially controlled delivery of extracellular matrix (ECM) proteins (e.g., Laminin for adhesion/myelination, Fibronectin for migration), decellularized ECM (dECM), and neurotrophic factors (e.g., NGF, GDNF) provides chemotactic signals and enhances SC proliferation, migration, and myelination through receptors such as integrins and Trk. Conductive polymers (e.g., PPy) and piezoelectric materials (e.g., PVDF) transmit endogenous or exogenous electrical signals. Electrical stimulation (ES) directs SC migration (electrotaxis), accelerates Wallerian degeneration, enhances neurotrophic secretion, and promotes remyelination by modulating Ca²⁺ signaling and purinergic pathways. The synergistic integration of these multimodal cues within a single NGC creates a biomimetic microenvironment that orchestrates the entire SC repair program, aiming to overcome the limitations of autografts for critical nerve gap repair.
Temporal dynamics of SC behavior in nerve regeneration
In the PNS, mature SCs radially ensheathe axons with myelin to facilitate rapid signal conduction. Following PNI, this structural integrity is disrupted, necessitating SC-mediated repair to restore axonal connectivity. The temporal progression of SC behavior post-injury—spanning Wallerian degeneration, inflammatory activation, regenerative niche formation, and functional reintegration—serves as a blueprint for designing NGCs. This section delineates the dynamic interplay between SC phenotypic transitions and nerve regeneration (Fig. 2), emphasizing key checkpoints for NGC intervention.

(1) Pro-inflammatory phase (0–3 days): SCs dedifferentiate (↑ c-Jun/Sox2, ↓Krox20/MPZ), activate myelinophagy (MMP-9/lysosomal pathways), and recruit macrophages via CCL2/C3 opsonization. (2) Pro-regenerative phase (3–7 days): SCs align into Büngner bands (Rac1/Cdc42-driven cytoskeletal reorganization) and secrete neurotrophic factors (NGF, BDNF, GDNF) to guide axonal regrowth. Metabolic coupling via MCT1-mediated lactate shuttling supports axonal energy demands. (3) Remyelination phase (weeks–months): SCs redifferentiate via NRG1-ErbB signaling and myelinate axons through α6β4 integrin-laminin interactions.
Pro-Inflammatory Reprogramming: Transition from Myelinating to Repair Phenotype
SCs undergo rapid phenotypic reprogramming to initiate repair within hours to days post-injury. Mature myelinating and non-myelinating SCs downregulate myelin-associated genes (e.g., MPZ, MAG) and adopt a repair state marked by pro-inflammatory signaling. This transformation is driven by transcriptional rewiring, including upregulation of c-Jun and Sox2, which license SCs to re-enter the cell cycle and secrete cytokines (e.g., TNF-α, IL-1α) to recruit macrophages10,16. Concurrently, SCs activate myelinophagy—a lysosome-dependent selective autophagy process that degrades their myelin sheaths17,18. Myelin debris clearance is further amplified by phosphatidylserine “eat-me” signals and complement opsonization (e.g., C3), creating a permissive environment for axonal regrowth19,20. Critically, residual myelin proteins (e.g., Nogo-A, MAG) act as potent inhibitors of axon extension, underscoring the necessity of efficient debris removal for successful regeneration.
Pro-regenerative niche establishment: büngner bands as axonal guidance tracks
By 3-7 days post-injury, repair SCs coalesce into longitudinally aligned cellular scaffolds termed Büngner bands within preserved basal lamina tubes. These bands provide physical guidance tracks for regenerating axons while secreting neurotrophic factors (e.g., GDNF, BDNF, artemin) to sustain neuronal survival and stimulate axonal elongation21,22,23,24. Simultaneously, SCs deposit ECM proteins (e.g., laminin, collagen IV) to stabilize the regenerative niche and direct growth cone navigation. After nerve injury, SC metabolism rapidly tilts towards glycolysis in the hypoxic injury environment to sustain its survival. Glycolysis produces ATP necessary for actin polymerization, which drives SC migration towards the Büngner band formation25.
Meanwhile, SCs protect injured axons by a dramatic glycolytic upregulation that arises in SCs as an inherent adaptation to axon injury. This glycolytic response, paired with enhanced axon–glia metabolic coupling, supports the survival of axons. Lactate exported via monocarboxylate transporter 1 (MCT1) is shuttled to regenerating axons as an energy substrate, sustaining growth cone motility26,27.
Axon-SC reciprocal remodeling: from guidance to functional reinnervation
Over weeks to months, bidirectional SC-axon interactions refine functional recovery. SCs re-establish a 1:1 relationship with large-caliber axons under neuregulin-1 (NRG1) type III signaling, though remyelination yields thinner sheaths and shorter internodes than developmental myelination. This structural adaptation is modulated by mechanosensitive pathways (e.g., YAP/TAZ) and reduced mechanical tension in regenerating nerves28,29,30,31. Concurrently, SCs reassemble Ranvier nodes through the secretion of gliomedin and NrCAM, which cluster axonal sodium channels to restore saltatory conduction32. At neuromuscular junctions, terminal SCs extend processes to bridge denervated endplates, while CXCL12α-CXCR4 signaling enhances motor axon reinnervation, ensuring synaptic specificity33,34.
The temporal dynamics of SC behavior-from dedifferentiation to Büngner band formation and eventual remyelination-highlight their intrinsic capacity to orchestrate nerve repair. However, this endogenous regenerative program is often disrupted in clinical settings such as critical nerve gaps, leading to incomplete functional recovery. That is to say, although the peripheral nerve can achieve partial regeneration, the repair process has inherent limitations and is often incomplete, leading to poor functional recovery25. Bioactive NGCs must therefore provide a suitable dynamic microenvironment to support the spatiotemporal dynamics of SC behavior.
Functional modulation of SCs by NGCs
NGCs are pivotal in bridging nerve defects by providing structural support and bioactive cues to direct SC-mediated regeneration. While conventional NGCs offer basic guidance, their clinical efficacy lags behind autografts due to insufficient mimicry of the native nerve microenvironment. To bridge this gap, advanced NGC designs now prioritize active modulation of SC behavior through tailored physical, biochemical, and electrical cues. This section examines strategies to enhance NGC functionality by targeting SC adhesion, migration, alignment, and phenotypic plasticity. Their design and SCs activation strategies could provide guidance and perspective for the future development of NGCs.
Physical cues in NGC design
Despite their potential, clinical outcomes of conventional NGCs often lag behind autografts due to insufficient mimicry of the native nerve’s physical microenvironment, a critical determinant of SC behavior. Emerging evidence demonstrates that SCs dynamically sense physical cues from NGCs, such as surface topography, mechanical stiffness, and porous architecture. NGCs can precisely regulate SC behavior by optimizing these parameters to improve regenerative outcomes.
Surface topography-guided SC alignment
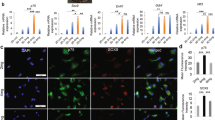
The surface topography of NGCs provides critical directional cues for SC alignment and migration, mimicking the anisotropic structure of native nerves. At the microscale (10-100 μm), aligned grooves or ridges replicate the longitudinal orientation of endoneurial tubes, providing contact guidance for SC migration. For instance, chitosan scaffolds with 30/30 μm ridge/groove patterns enhance SC alignment by 2.1-fold compared to smooth surfaces, accelerating gap bridging in 10 mm rat sciatic nerve defects13. Additionally, Polycaprolactone (PCL) and polylactic acid (PLA) films with different groove shapes (sloped walls, V-shaped, and square-shaped) improve SC attachment, proliferation, and orientation. Sloped wall grooves showed the best results in guiding nerve regeneration in vivo14. Zhen Zhang et al. explored the effects of submicron groove size (width/depth) on SC behavior (morphology, proliferation, migration, gene expression, membrane potential) (Fig. 3A). The results confirmed that submicron grooves (especially 800/400 nm) can effectively guide the directional arrangement and migration of SCs (depth dependent), unregulate key genes for axonal regeneration and myelin formation (MBP/Smad6), alter cell membrane potential, but do not affect cell cycle growth.15. Furthermore, nanoscale grooves engraved on electrospun microfibers significantly enhanced SC migration35. Such nanotextured surfaces upregulate pro-migratory genes (Rac1, Cdc42), enabling SCs to traverse gaps faster than on planar substrates.

A The fabrication process of submicron-grooved films can be utilized to direct the alignment of SCs. Reproduced with permission15. Copyright 2023, MDPI. B The expression of YAP (green) in SCs after 3 days of incubation in aligned methacrylated silk fibroin fibers (ASFFs) and the possible mechanism of RGD@ASFFs on regulating SC behavior. Reproduced with permission39. Copyright 2022, Elsevier. C Expression and localization of pro-myelinating transcription factors in SCs modulated by substrate stiffness. Reproduced with permission43. Copyright 2022, John Wiley and Sons. D Expression of pro-regenerating/repair transcription factor in SCs is modulated by substrate stiffness. Reproduced with permission43. Copyright 2022, John Wiley and Sons.
Aligned electrospun nanofibers (diameter 200–800 nm) emulate the size and alignment of collagen fibrils in the endoneurium. SCs cultured on aligned PCL nanofibers adopt elongated morphologies and secrete elevated levels of neurotrophic factors (NGF, BDNF) compared to random fiber scaffolds36,37. In vivo studies demonstrate higher myelinated axon density in aligned nanofiber NGCs versus smooth conduits38. The superior myelinated axon density observed in aligned nanofiber NGCs versus smooth conduits arises from topographically driven SC reprogramming. Xiaoli Chen et al.39 found that aligned nanofibers could promote SCs’ myelination by increasing gene expressions of Krox20, Pmp22, and MBP, essential in the SCs’ myelination process. They confirmed that the aligned topology of fibers can induce the orientation and differentiation of SCs via mechanical stimulation, activating YAP nuclear translocation to promote myelination and neurotrophin secretion function (Fig. 3B).
Moreover, advanced 3D topographical structures have shown promise in guiding SC behavior. Core-sheath nanoyarn scaffolds (PCL/gelatin) combine macroscale alignment for migration with nanoscale roughness for integrin binding, achieving dual-scale guidance40. Surfaces with hierarchical topographies that combine micro- and nanoscale features provide SCs with multiple cues, enhancing cell adhesion, migration, and differentiation41. This combination of features more closely mimics the ECM, thus creating an environment that is highly conducive to nerve regeneration. Using multi-scale surface topographies allows for a more effective and versatile approach in supporting SC behavior and improving the outcomes of NGCs in nerve repair.
In conclusion, surface topography, particularly micro/nanoscale anisotropic cues like parallel microgrooves, aligned fibers, or channels, profoundly directs SC alignment through contact guidance and mechanical biological mechanisms (YAP pathway). The development from single scale to micro-nano multi-level composite design can accurately simulate the anisotropy of neural ECM and provide a core strategy for constructing efficient NGCs. While topographical cues direct SC alignment, substrate stiffness further fine-tunes their mechanoresponsive behavior, as discussed below.
Stiffness gradients for mechanoresponsive SC regulation
SCs are inextricably connected with their extracellular environment, and there is growing evidence that SCs are sensitive to changes in the mechanical properties of the surrounding environment42. SCs mechanically sense the extracellular matrix’s stiffness, and SC mechanosensitivity modulates their morphology and migratory behavior43. Their cell morphology and migratory phenotype can be altered in response to a stiffness gradient. Notably, it exhibits remarkable mechanosensitivity through durotaxis, demonstrating directional migration along stiffness gradients independent of gradient steepness44. Thus, embedding stiffness cues into an NGC presents another promising strategy for controlling SC migration following injury and ultimately promoting successful nerve regeneration.
Substrate stiffness exerts biphasic effects on SC phenotypic regulation. Compliant matrices (1.1 kPa PAAm) induce dedifferentiation characterized by elevated c-Jun expression, establishing a repair-permissive cellular state through mechanical reprogramming. Conversely, rigid substrates (27.7 kPa) promote redifferentiation and myelination competence, evidenced by Krox20 upregulation—a master regulator of myelinogenesis (Fig. 3C, D)43. Intermediate stiffness regimes (7.45 kPa) demonstrate optimal support for fundamental cellular processes, enhancing SC elongation, focal adhesion formation, and proliferation in 2D culture systems45. Intriguingly, 3D microenvironments reveal distinct mechanical preferences, with maximal proliferation observed in 0.53 kPa scaffolds46, highlighting the necessity of dimensional considerations in scaffold design. Silk fibroin scaffolds with β-sheet domains enhance SC migration and proliferation by balancing mechanical support and elasticity47.
Molecular mechanotransduction pathways mediating SC stiffness responses are beginning to be elucidated. The PIEZO1/YAP/TAZ axis emerges as a key regulatory system, with soft substrates (0.9–2.9 kPa) promoting mesenchymal stem cell transdifferentiation into SC-like phenotypes through this pathway48,49. These findings underscore the importance of mechanobiological investigations for developing next-generation nerve conduits. Strategic manipulation of scaffold stiffness gradients, informed by SC mechanosensitivity patterns, may significantly enhance directional migration and regenerative performance—a promising approach for improving peripheral nerve repair outcomes.
Porous architectures for 3D microenvironment control
Porous structures represent a fundamental three-dimensional morphological feature of NGCs, playing a critical role in facilitating cell infiltration, nutrient diffusion, and molecular signaling while allowing the removal of metabolic waste. Studies have demonstrated that NGCs with a porous architecture exhibit enhanced nerve conduction velocity and more myelinated axons, indicating superior regenerative outcomes50. It promotes the initial adhesion, spreading, proliferation, and differentiation of SCs, ultimately forming Büngner bands. Moreover, such a structure supports vascularization and minimizes fibrous scar formation.
The porosity and interconnectivity of NGCs are essential for SC migration, which is crucial for establishing a supportive microenvironment for regenerating axons51. Scaffolds with an approximately 70–90% porosity have been shown to support SC viability and proliferation. The size and morphology of the pores significantly influence SC adhesion, migration, and proliferation. An appropriately tuned pore size can enhance the surface roughness of NGCs, thereby providing anchoring points for early SC adhesion52. The porous and permeable sidewalls of NGCs allow endogenous SCs to infiltrate the injury site, aiding ECM reconstruction and nerve repair. Scaffolds with 25–50 μm pores maximize SC migration and Büngner band formation. Electrospun Antheraea pernyi silk fibroin (RASF) conduits with 25 μm pores significantly enhance SC infiltration53.
The three-dimensional microporous architecture within NGCs also plays a pivotal role in regulating SC behavior54. A directionally aligned chitosan/collagen filler, fabricated using a directional freezing technique with a stiffness comparable to natural tissue (2.0 ± 0.4 kPa), has been incorporated into a porous electrospun PCL sheath. This biocompatible filler, featuring a 20–50 μm grid-like pattern, facilitates SC migration and axonal extension within the conduit. The longitudinally aligned pores create a spatial environment mimicking the endoneurium, providing physical support and guidance cues for SC migration and axonal growth55.
Recent advances in 3D-bioprinted hydrogels have enabled precise integration of porosity, anisotropic architecture, and cell density to mimic native nerve microenvironments. Lee et al.56 engineered porous PLCL conduits with swellable gelatin-methacryloyl (GelMA) microgrooves, where controlled swelling (15% volume increase) generates interconnected 40-60 μm pores that support >90% SC viability at optimal cell densities (2 × 10⁶ cells/mL). Swellable microgrooves (50 μm width) in GelMA hydrogels undergo hydration-triggered expansion, compressing embedded SCs to form aligned cellular bands that secrete 3.1-fold higher laminin than random scaffolds—effectively emulating Büngner bands in vitro. Yoo et al.57 combined 3D-printed collagen hydrogel cores (20 μm longitudinal channels) with porous PLCL sheaths (70% porosity), creating dual-scale architectures that enhance SC migration velocity by 2.4-fold compared to isotropic controls. The collagen channels direct SC alignment, while PLCL pores facilitate vascular ingrowth. Current printing resolution (>50 μm) limits replication of endoneurial-scale topographies (<5 μm), constraining SC-axon contact guidance. Uniform cell loading fails to mimic injury-site hypercellularity, and emerging inkjet-printing of SC spheroids may enable zonal density control.
Porous architectures with specific biological functions are crucial for successful peripheral nerve repair. However, the design of porous structures presents particular challenges. Pore sizes exceeding 30 μm may permit fibrous tissue infiltration, which can hinder axonal regeneration58. Therefore, optimizing pore size is essential to balance structural integrity while promoting efficient SC migration and axonal guidance. Both excessively large and overly small pores can compromise conduit performance. Moreover, increased porosity (>70%) improves permeability but reduces structural integrity, necessitating balanced designs. Asymmetric conduits with selective permeability, featuring smaller pores on the inner surface and larger pores on the outer surface, offer an effective strategy for nerve regeneration. Studies suggest that nano-sized pores (~50 nm) on the inner surface effectively block fibrous tissue infiltration, whereas larger pores (~ 50 μm) on the outer surface support vascular ingrowth and nutrient diffusion59.
Biochemical strategies for SC regulation
Beyond physical and topographical optimization, biochemical functionalization of NGCs with ECM components, decellularized matrices, and neurotrophic factors establishes a biomimetic signaling milieu to regulate SCs’ plasticity. This section examines strategies to enhance SC-driven regeneration through targeted biochemical cues.
ECM-derived bioactive matrices: collagen, fibronectin, and laminin
ECM proteins provide structural and biochemical signals to direct SC behavior. Collagen emerges as a multifunctional scaffold material, demonstrating unique capabilities in guiding SC behavior. Type I collagen coatings enhance SC migration velocity compared to unmodified surfaces, with aligned fibrillar networks providing contact guidance for directional axonal growth60,61. Beyond structural roles, collagen-integrin interactions activate MAPK and YAP/TAZ mechanotransduction pathways49,62, driving SC differentiation and basal lamina formation essential for myelination63,64. Notably, collagen-modified scaffolds upregulate neurotrophins (BDNF, NGF) and immunomodulatory factors65, establishing paracrine signaling fields that support axonal survival. Aligned fibrillar collagen networks further guide directional axonal growth11,66.
Fibronectin induces 2.1-fold greater SC proliferation than collagen coatings67,68. Its migratory guidance capability is equally remarkable, with SCs demonstrating faster movement on fibronectin than on type IV collagen substrates (Fig. 4A)69. This migratory effect is mediated through interactions with specific integrins, particularly alpha-v integrins, which facilitate SC movement on fibronectin substrates70. Fibronectin enhances muscle reinnervation efficiency and axonal density through synergistic mechanochemical effects when engineered into aligned topographic patterns or integrated with chitosan conduits/alginate hydrogels71,72,73—these properties position fibronectin as the preferred coating for applications requiring rapid SC expansion and directed migration.

A Effects of ECM proteins (laminin, fibronectin, and type IV collagen) on the biological behavior of SCs. Reproduced with permission69. Copyright 2023, Frontiers. a Adhesion and cell morphology of SCs in early 2D culture conditions. Axial pseudopodia (red arrow) and radial pseudopodia (green arrow). b The proliferation of SCs under 2D culture conditions (EdU staining). c Transwell migration assay of SCs and SCs morphology under 3D culture conditions. B Effects of dECM on SC morphology and gene expression. Reproduced with permission89. Copyright 2025, Springer. C Potential molecular mechanism by which NGF regulates myelin clearance and axon regeneration following PNI. Reproduced with permission100. Copyright 2020, Ivyspring International Publisher.
Laminin, a key neural ECM component, exhibits unparalleled bioactivity in regulating SC behavior. Studies have shown that SCs exhibit higher attachment rates on laminin-coated surfaces than on other ECM proteins like collagen I74. SCs exhibit higher proliferation rates on laminin than on other ECM molecules such as fibronectin and collagen. Furthermore, laminin supports SC differentiation by regulating cytoskeletal rearrangements and cell polarity75,76. It is more effective in promoting myelination than fibronectin and collagen type IV69,77. Laminin, particularly its γ1-chain isoforms, serves as the molecular linchpin for SC-ECM adhesion and myelination competence; its laminin globular (LG) domains bind integrin α6β1 and dystroglycan on SCs, triggering two critical cascades: (1) γ1-integrin engagement activates PI3K, phosphorylating Akt to suppress SC apoptosis in hypoxic injury sites, and (2) γ1-dystroglycan binding recruits utrophin to anchor F-actin, polarizing SCs along axons—a prerequisite for spiral myelination. Critically, γ1-chain deficiency disrupts these interactions, with knockout models showing >80% reduction in myelinated axons due to failed SC-axon contact stabilization (impairing Krox20 nuclear translocation) and collapsed basal lamina architecture (causing aberrant Büngner band formation)77,78. Laminin-derived peptides (YIGSR, IKVAV) also accelerated nerve regeneration by inducing oriented SC migration and remyelination79. A new class of self-assembling peptides (SAPs) with designer motifs has also been widely explored as potential scaffolds for nerve repair80. A peptide (SAP RADA-16-I-BMPH1) co-spun scaffold demonstrated better cellular infiltration and significantly higher gene expression of neural cell adhesion molecule, glial fibrillary acidic protein (GFAP), and peripheral myelin protein 22 (PMP22)81.
The differential bioactivities of these ECM components highlight the importance of strategic molecular selection. Collagen excels in creating mechanically robust scaffolds with neurotrophic support82, while fibronectin dominates in proliferative induction and migration guidance. Laminin, with its unique adhesive specificity and myelination competence, proves indispensable for functional recovery. Recent advances in peptide engineering allow selective amplification of these ECM functions-YIGSR for directional migration83, IKVAV for remyelination84—providing modular solutions for clinical nerve repair. This evolving understanding of ECM-SC interactions underscores the potential of hybrid matrices combining multiple bioactive elements to address the complex demands of peripheral nerve regeneration.
Decellularized ECM: preserving native signaling complexity
The inherent complexity of native SC microenvironments poses significant challenges for synthetic scaffold biofunctionalization85. While single-component ECM coatings provide partial biological activity, they fail to recapitulate the synergistic signaling architecture of natural neural matrices. Decellularized extracellular matrix (dECM), created through decellularization technology that removes immunogenic cell components from human or animal tissues, while preserving physicochemical signals and biological attributes, is particularly beneficial for PNI regeneration86. This bioactive reservoir enables coordinated SC proliferation, migration, and differentiation regulation through multidimensional signaling integration87,88.
A composite NGC that combined a poly(lactic-co-glycolic acid) (PLGA) shell and a dECM of rat sciatic seeded with SCs has been explored. It reported that dECM enhanced the proliferation, survival, and neurotrophic factor secretion of SCs by upregulating the key components of the PI3K/Akt pathway (Fig. 4B)89. The in vivo testing further demonstrated that this PLGA/dECM-SC conduit significantly improved nerve regeneration, showing enhanced axonal growth, myelination, and motor function restoration comparable to autografts. An injectable hydrogel derived from decellularized porcine peripheral nerves has been developed to efficiently deliver SCs while maintaining higher cellular viability and protecting SCs from an astrocyte-mediated inflamed microenvironment90. Similarly, a spinal cord dECM-incorporated conduit has been reported to improve peripheral nerve regeneration in vivo by promoting the reprogramming of SCs to promote axon regeneration and remyelination91. Moreover, the reprogrammed SCs and the NGCs had anti-inflammatory effects and altered the activation state of macrophages to M2 phenotypes to enhance nerve regeneration. Another in vivo study reveals enhanced functionality of dECM-engineered conduits with accelerating SCs migration velocity through chemotactic guidance by residual CXCL12 chemokines92. These effects are mediated through preserved integrin-binding domains in dECM components that activate YAP/TAZ mechanosensing pathways, creating a self-reinforcing loop between matrix remodeling and SC differentiation. However, challenges remain in standardizing decellularization protocols (e.g., detergent concentration gradients, sonication parameters) to balance antigen removal with bioactive component retention. Emerging techniques like supercritical CO2 decellularization combined with ECM photopatterning show promise in creating spatially organized dECM scaffolds with region-specific bioactivity for precision nerve repair.
Furthermore, in parallel to extensively studied tissue-derived ECMs, cell-derived ECMs can also partially recapitulate the complicated regenerative microenvironment of native nerve tissues93. Cell-derived ECM has been used to provide mechanical support and adhesion receptors for cell adhesion, proliferation, and differentiation94,95. Cell sheet technology has developed a novel scaffold enriched with cell-derived ECM. SCs grown on decellularized sheets showed upregulation of genes and proteins associated with myelination, such as Erbb2 and myelin protein zero, and increased expression of laminin chains96. PCL nanofibrous scaffolds have been surface-coated with a cell-derived ECM of SCs, positively influencing the SCs differentiation and maturation to myelin-promoting cells97. In another study, PCL nanofibrous scaffolds were coated with SCs-derived ECM to provide the microenvironment and biochemical cues to promote cell adhesion, proliferation, and remyelination98. SCs-derived ECM fulfilled the role of laminin in promoting cell adhesion and, eventually, PMP22 gene expression99. Current research has confirmed that cell-derived ECM can promote nerve regeneration after PNI, demonstrating significant clinical potential. However, transferring from the bench to the bedside is still far. Several challenges remain to be further addressed, such as deepening the mechanistic understanding of cell-derived ECM nerve regeneration, optimizing decellularization by exploring apoptosis-based methods to preserve bioactive ECM integrity, and diversifying applications beyond coatings (e.g., powders, hydrogels, 3D-bioprinted bioinks) for spatial nerve reconstruction93.
Spatiotemporal control of neurotrophic factor delivery
The therapeutic potential of neurotrophic factors lies in their ability to recapitulate the dynamic biochemical interplay governing SC plasticity during nerve regeneration. Nerve growth factor (NGF) orchestrates SC behavior through multimodal signaling cascades, most notably via its interaction with the p75 neurotrophin receptor (p75NTR). This binding triggers AMP-activated protein kinase (AMPK) phosphorylation, suppressing mTOR activity, inducing autophagosome formation, and accelerating myelin debris clearance, key to reducing nerve remodeling time (Fig. 4C)100. Concurrently, NGF-mediated focal adhesion kinase (FAK) phosphorylation enhances SC migration by upregulating matrix metalloproteinase-2 (MMP-2) secretion, enabling efficient ECM remodeling and directional movement101. Remarkably, exogenous NGF administration creates a self-reinforcing regenerative niche, elevating endogenous glial cell line-derived neurotrophic factor (GDNF) secretion, thereby amplifying neurotrophic support for axonal regrowth102.
GDNF stimulated the migration of SCs and enhanced myelination in neuron-SC cocultures. Phosphorylating extracellular signal-regulated kinases (ERK1/2) boosts Krox20 expression, while protein kinase A (PKA)-dependent CREB activation upregulates myelin basic protein synthesis. Simultaneously, protein kinase C membrane localization stabilizes SC-axon contact structures, ensuring proper myelination103.
Although exogenously administered neurotrophic factors can enhance beneficial SC functions, their in vivo application is limited by high costs and rapid degradation. Embedding growth factors into NGCs to form a sustained release system effectively improves their effectiveness. Advanced encapsulation techniques, such as microsphere- or hydrogel-based systems, stabilize neurotrophic factors against enzymatic degradation and enable sustained release, extending their bioactivity from hours to weeks. Spatial control strategies, such as 3D-printed conduits with GDNF gradients, caused an increase in the SC migration velocity104. Combinatorial strategies such as co-delivering GDNF and NGF have improved nerve regeneration rates compared to single-factor approaches105. Despite these advances, challenges persist in achieving spatiotemporal precision. The regulatory balance between growth factors is equally critical: platelet-derived growth factor (PDGF) enhances SC phagocytosis via TAM receptor (Tyro3/Axl)-mediated Erk/Rac1 activation, whereas vascular endothelial growth factor (VEGF) suppresses phagocytic efficiency through VEGFR-1/PI3K-induced RhoA inhibition106. This antagonism underscores the necessity for temporally controlled delivery systems to optimize regenerative phases. Future efforts must prioritize scalable biomaterial designs, precision in growth factor spatiotemporal dynamics, and rigorous clinical validation to address complex nerve injuries effectively.
Bioelectric modulation of SC behavior
The native PNS relies on bioelectric signaling for action potential propagation and cellular communication. Capitalizing on this intrinsic property, electrical modulation strategies, including exogenous electrical stimulation (ES) and conductive biomaterials, emerged as powerful tools to enhance SC functionality and accelerate nerve repair. These approaches synchronize SC-axon interactions and microenvironmental remodeling by mimicking endogenous electrophysiological dynamics, offering a promising avenue to bridge the efficacy gap between synthetic NGCs and autografts.
Electrical stimulation for SC activation
ES has been widely used as an effective physical method to promote nerve repair in pre-clinical and clinical settings107. Direct currents have been shown to enhance the differentiation, alignment, migration, protein secretion, and proliferation of several cell types involved in PNS repair, such as fibroblasts, induced pluripotent stem cells, and SCs9. SCs are reprogrammed into a repair phenotype under ES, which secretes neurotrophins and supports axonal regeneration (Fig. 5A)107A recent study demonstrated that ES significantly promoted the degeneration and clearance of axons and myelin, and the dedifferentiation of SCs to promote nerve regeneration108. ES also increased calcium influx in SCs, promoting their proliferation and transition toward a pro-repair phenotype109. Another study found that SCs migrated anodally in an applied electric field. While the directedness and displacement of this migration significantly increased as the electric field strength rose from 50 mV/mm to 200 mV/mm, the electric field strength did not substantially affect their migration speed110. Concurrently, ES upregulates neurotrophic factor secretion, creating a pro-regenerative niche that supports axonal survival and growth cone navigation111,112. At the myelination stage, ES enhanced SC-axon metabolic coupling through purinergic signaling. In rodent models, sciatic nerve stimulation elevates extracellular ATP levels, activating P2Y receptors on SCs and triggering Ca²⁺ oscillations (3–5 Hz) via IP3-dependent pathways113. These Ca²⁺ transients promote exosome release—vesicles enriched with miR-21 and laminin, enhancing neuronal survival and guiding remyelination114. ES also modulates local SCs to increase exosome release, which promotes a macrophage phenotype shift from pro-inflammatory M1 to pro-repair M2, ultimately downregulating pro-inflammatory cytokines, promoting pro-repair factors, expediting myelin debris clearance and remyelination, and priming the microenvironment for optimal functional recovery115,116,117. ES synchronized SC differentiation by activating MAPK/ERK and IL-17 pathways, increasing Krox20 expression and myelin thickness in regenerated nerves118.

A SCs are reprogrammed into a repair phenotype under ES, which secretes neurotrophins and supports axonal regeneration. Reproduced with permission107. Copyright 2025, Frontiers. B In vitro gene and protein expression levels in the SCs of the PPy/PDA/PLLA scaffold. Reproduced with permission122. Copyright 2023, American Chemical Society. C The cell viability and expression of nerve regeneration-related factors of SCs cultured with extracts from chitosan/ GO films. Reproduced with permission126. Copyright 2023, Wolters Kluwer Medknow. D Gene expression of SCs and signaling pathways expressed on piezoelectric topological scaffolds that may be associated with myelination and nerve repair. Reproduced with permission148. Copyright 2024, Elsevier.
Conductive materials
Conductive scaffold materials hold potential for creating a pro-regenerative microenvironment by replicating the electrophysical properties of native tissue. Additionally, electrically conductive materials are necessary for applying electrostimulation to cells. The neuro-conductive property can be imparted by blending the biodegradable, bioactive polymers with conductive polymers. This will provide additional features, i.e., electrical cues to the existing topographical and bioactive cues, to make it a more multifaceted neuroregenerative approach119. Recent studies have increasingly focused on integrating conductive materials into NGCs to harness the electroactive properties of these materials for improving nerve regeneration outcomes. Conductive polymers and carbon-based materials are the primary materials for fabricating conductive NGCs.
Polypyrrole (PPy) is a well-studied conductive polymer widely utilized in nerve conduit applications due to its excellent biocompatibility, electroactivity, and ease of synthesis. PPy’s reversible redox activity and tunable conductivity (10⁻³–10² S/cm) make it ideal for interfacing with SCs. SC culture confirmed that the silk fibroin/PPy (SF/PPy) nanofibrous mats improved cell proliferation and adhesion compared to pure SF mats120. Another conductive SF/PPy composite scaffold has been found to enhance the proliferation, migration, and neurotrophic factor secretion ability of SCs under ES118. It also effectively promotes axonal regeneration and remyelination in vivo. The synergy between gelatin and PPy enhanced conductivity, increasing myelination gene expression and nerve growth factor release of SCs121. A biomimetic PPy/ polydopamine (PDA)/poly-L-lactic acid (PLLA) electrospun fibrous scaffold was confirmed to directly activate SCs through electrical activity, promoting their proliferation, differentiation, and secretion of neurotrophic factors (Fig. 5B). The Calcium-AMPK signal axis was a key pathway for this scaffold to regulate SCs’ function and provide energy and metabolic support for nerve regeneration122. While PPy exhibits excellent conductivity, its long-term cytotoxicity remains a concern due to potential oxidative degradation.
Polyaniline (PANi) is another important and widely used material due to its low cost and easy conductivity manipulation123. Developing pure PANi into an NGC is challenging because of its brittleness and rigidity. It is commonly used in polymer composites and different biocompatible polymers. PANI/cellulose hydrogels have been fabricated for nerve regeneration124. In this work, the cellulose hydrogel was used as a template to in situ synthesize PANi via the limited interfacial polymerization method, leading to one conductive side in the polymer. The conductive side containing PANi significantly promoted adhesion, proliferation, and pseudopod extension of SCs. PANi played an indispensable role in the peripheral nerve regeneration. The hierarchical micronanostructure and electrical conductivity of PANi could remarkably induce the adhesion and guiding extension of neurons. A novel water-soluble conductive material has been synthesized by grafting polyaniline (PANI) onto carboxymethyl chitosan (CMCS)125. An electroconductive hydrogel (ACCP) with injectable and self-healing properties was further fabricated by adding CMCS-PANi (CP) to the dynamic gel network generated by the Schiff base reaction between aldehyde-based hyaluronic acid (ALHA) and CMCS. The in vitro experiments indicated that ACCP3 (3 wt% CP) promoted the proliferation and migration of the SCs, and upregulated the gene expression of MBP and NGF. In vivo experiments further confirmed that this conductive hydrogel promoted nerve conduction velocity, enhanced the expression of axon-specific proteins in neurons, induced axonal extension and myelin regeneration, and prevented muscle atrophy.
Graphene oxide (GO) has gained increasing interest in nerve regeneration applications due to its large surface area, unique nanostructure, good mechanical properties, and hydrophilic functional groups suitable for cell attachment. GO and reduced graphene oxide (rGO) are two primary carbon-based forms applied for NGCs. SCs exhibited elongated spindle shapes and upregulated expression of nerve regeneration-related factors such as Krox20 (a key myelination factor), Zeb2 (essential for SC differentiation, myelination, and nerve repair), and transforming growth factor β (a cytokine with regenerative functions) on chitosan/ GO films at ≤1 mg/mL concentrations (Fig. 5C)126. Hybrid nanofibers with low concentrations of rGO (0.25 and 0.5 wt%) significantly improved the proliferation of SCs. More importantly, rGO/GelMA/PCL hybrid nanofibers could activate the epithelial–mesenchymal transition (EMT)-related gene expression of SCs127. The filler in hollow NGC, chitosan/oxidized hydroxyethyl cellulose (CS/OHEC) hydrogel loaded with asiaticoside liposome and the conductive rGO was developed and used to reform the microenvironment for peripheral nerve regeneration128. The application of ES after adding rGO promoted the differentiation and proliferation of nerve cells, accelerating nerve regeneration. The scaffolds may present altered mechanical properties with GO incorporation, contributing to SC fate determination. A recent study found that matrix stiffness primarily governed the survival, proliferation, and morphology of SCs, while conductivity synergistically enhanced SC functions, such as neurotrophic factor secretion129. It has been reported that conductive biomaterials could reduce intracellular Ca2+ levels by inhibiting calcium-sensitive receptors and phospholipase C-β (PLCβ) pathways, thereby inducing SCs to secrete neurotrophic factors130.
Meanwhile, the decrease in intracellular Ca2+ levels may also lead to the inactivation of protein kinase C (PKC), which may affect cell differentiation, migration, and secretion through the mitogen-activated protein kinase (MAPK) pathway131. According to previous reports, GO may exhibit certain cytotoxicity due to hydrophobic interactions with cell membranes and subsequent extraction of membrane cholesterol and phospholipids132. It has also been reported to stimulate pro-inflammatory responses and cause the recruitment of immune cells133. Concentration-dependent cytotoxicity necessitates precise dosing (5 mg/mL GO), which induces apoptosis via ROS overproduction, whereas ≤1 mg/mL maintains 90% SC viability134.
Carbon nanotubes (CNTs) have attracted increasing attention in peripheral nerve tissue engineering due to their unique structural and physical characteristics135,136. Studies have shown that CNT-based electrospun scaffolds can mimic the electrophysiological properties of native peripheral nerve tissue, which is beneficial for neuronal differentiation and neurite elongation137,138. A fibrous scaffold, fully assimilating polyurethane (PU) and silk fibroin associated with functionalized multi-walled carbon nanotubes (MWCNTs) has been developed by the electrospinning technique139, in vitro tests revealed that this PU/Silk-MWCNTs scaffold significantly stimulated the growth and proliferation of SCs, together with the differentiation and spontaneous neurite outgrowth of rat pheochromocytoma (PC12) cells that were particularly guided along the axis of fiber alignment. In another study, composite nanoyarns based on poly(p-dioxanone) (PPDO) biopolymer and different concentrations of CNTs were manufactured by utilizing a modified electrospinning apparatus140. The biological tests revealed that the addition of CNTs better maintained the phenotype of SCs, and significantly enhanced the differentiation capability of human adipose-derived mesenchymal stem cells (hADMSCs) into SC-like cells by notably upregulating the expression levels of SC myelination-associated gene markers and increasing the growth factor secretion when combined with ES. Although initial safety concerns about the toxicity of independent CNTs limited their use in neuronal applications, growing research suggests that these issues can be bypassed by controlling the firm attachment of CNTs to the substrate141. Further nanotoxicological and pharmacokinetic studies are also required, and more studies should be carried out on the relationship between toxicity and shape, size, functional groups, and release of CNT when integrated into the substrate. As more control over CNT behavior is gained, a better comprehension of their interactions with nerve cells will be attained. This will facilitate the more effective integration of CNTs into NGCs for application in nerve regeneration.
Piezoelectric materials
Conventional ES is usually performed by invasive implanted electrodes, which display continuous contact with neural tissue, easily causing secondary damage and inflammatory responses. It will inevitably cause mobility difficulties, limiting its clinical application142. Developing an effective route of non-invasive remote ES is necessary for better tissue regeneration. Due to the electromechanical coupling effect, piezoelectric materials can convert their mechanical energy into electrical signals via a ‘non-invasive’ style, thus showing the potential of application in nerve regeneration143,144.
Piezoelectric materials enable passive energy harvesting by converting physiological mechanical stimuli (e.g., muscle contraction or conduit deformation) into endogenous bioelectric signals. Piezoelectric materials generate electrical signals through the induction of mechanical strains and modulate cellular behaviors. Studies have demonstrated that polyvinylidene fluoride (PVDF) and its copolymer with trifluoroethylene (PVDF-TrFE 70/30) are biocompatible and support SC proliferation and myelination by providing a conducive environment145,146. A piezoelectric zinc oxide nanogenerator scaffold was found to supply SCs with an adhesive, proliferative, and angiogenic interface, as is reflected by higher expression of functional proteins including NGF and VEGF147. A biomimetic-inspired piezoelectric topological ovalbumin/BaTiO3 scaffold was constructed by combining piezoelectric BaTiO3 nanoparticles and surface microtopography. The results revealed that the piezoelectric topological scaffolds up-regulated the gene expression referring to myelination and axon growth, which facilitated the local enrichment of Ca2+ and myelination of nerve fibers and played an important role in electrical stimulation-induced nerve regeneration (Fig. 5D)148. A SF/poly(vinylidene fluoride-co-hexafluoropropylene)/Ti3C2Tx (SF/PVDF-HFP/MXene) composite scaffold with piezoelectricity has been found to induce the proliferation of SCs, enhance the elongation of axons, and promote axonal myelination149.
Hybrid designs (PVDF/poly (D, L-lactide-co-caprolactone) (PLCL)/PEDOT) combined piezoelectricity with conductivity, regulating the immune microenvironment by activating the PI3K/AKT-Nrf2 signaling pathway, thereby promoting macrophage polarization toward M2 anti-inflammatory phenotype. This subsequently facilitated SCs’ recruitment and myelin formation by mediating the macrophage paracrine system150. Another hybrid design was a nanopatch consisting of an oriented barium titanate (BTO)-incorporated PCL nanofiber membrane for electricity generation and a layer of GO-doped GelMA for neural interaction151. With the assistance of ultrasound, some mechanical energy was converted into electricity by this nanopatch to accelerate nerve regeneration by promoting the proliferation of SCs and the axonal outgrowth of neurons.
Hydrogel-based piezoelectric devices have recently attracted intensive attention because of their unique piezoelectric performance and conductive hydrogel configuration152. Piezoelectric hydrogel has been reported to produce localized electrical charges under external ultrasound (US) to promote cell migration, attract host cells, and induce endogenous growth factors like TGF-β1153. A US-responsive composite hydrogel composed of a gelatin/polyvinyl alcohol (PVA) interpenetrating polymer network incorporated with KNN nanocrystals and rGO has been designed to realize sonoelectric conversion to accelerate diabetic wound healing by facilitating local neurovascular regeneration154. These hydrogels, combined with US-responsive electrical stimulation, activated cellular synthesis and secretion processes of SCs via the PPAR signaling pathway. Under ES, Ca2+ flowed through voltage-gated calcium channels (VGCC) to activate the AMPK signaling pathway, which regulated ATP synthesis to support transmembrane transport of vesicles containing neurotrophic factors such as NGF and BDNF.
When subjected to deformation or vibration, Piezoelectric materials can generate mechanical forces transmitted to cells, thereby inducing changes in cell morphology, alignment, and cytoskeletal organization. These mechanical cues are sensed by cells through mechanotransduction pathways, which convert mechanical stimuli into biochemical signals, ultimately modulating cellular behavior and function. Moreover, the electric charges generated by piezoelectric materials can directly influence cellular activities by interacting with membrane receptors and ion channels sensitive to electrical stimuli. This interaction can regulate key cellular processes, including proliferation, migration, differentiation, and gene expression, further highlighting the multifaceted role of piezoelectric materials in cell modulation155.
Despite promising preclinical outcomes, challenges persist in long-term stability and biomechanical compatibility. Due to the stable crystallinity and intrinsic structural characteristics of most existing piezoelectric polymers and their hydrophobic nature (e.g., PVDF), these materials exhibit poor biodegradability or are even resistant to degradation. These inherent limitations significantly restrict their broader application in the clinic. Future research should prioritize the development of piezoelectric materials that balance tunable degradation rates with sustained piezoelectric activity, thereby addressing the dynamic demands of nerve regeneration.
Multimodal synergy in nerve regeneration
While preceding sections delineate how isolated physical, biochemical, and bioelectric cues modulate Schwann cell behavior, their compartmentalized application in conventional NGCs fails to recapitulate the dynamic reciprocity of native nerve regeneration, where spatial, molecular, and electrophysiological signals converge to orchestrate SC plasticity. Recent studies have also revealed that the synergetic effects of different cues could promote nerve repair156. Combining two or even multiple cues can potentially encourage neurite eruption, accelerate nerve elongation, and enhance functional recovery more than an individual cue157. Here, we review the use of multimodal synergistic NGCs to enhance SC-mediated nerve repair, overcoming the limitations of single-cue approaches.
Physical-biochemical crosstalk
Combining biomimetic structure and biochemistry guidance cues has more potential to promote nerve regeneration than an individual cue. Shuhui Yang et al.158 fabricated an aligned fibrin/functionalized self-assembling peptide (AFG/fSAP) interpenetrating nanofiber hydrogel that exerts synergistic topographical and biochemical cues for peripheral nerve regeneration via electrospinning and molecular self-assembly. The aligned fibers were superior in inducing cell orientation that promotes SC maturation and myelination. Additionally, the hydrogels were gradually degraded with the growth of SCs. The cells not only adhered to the surface of the hydrogel but also degraded the hydrogel underneath and extended inside, contributing to forming an SC band. The aligned fibrin fiber also enhanced the secretion function of SCs. Promoted remyelination was found via the synergistic effect of aligned fibrin fibers and functional motifs. The aligned hydrogel upregulates regeneration-related genes and activates the PI3K/Akt and MAPK signaling pathways. It represented a promising approach for peripheral nerve repair by integrating structural guidance and biochemical stimulation. Deteng Zhang et al.159 prepared micropatterns and peptide gradients on the inner wall of a PLCL NGCs (Fig. 6A). After linear ridges/grooves of 20/40 μm in width are created on the PLCL film, its surface is aminolyzed in a kinetically controlled manner to obtain the continuous gradient of amino groups, which are then transferred to CQAASIKVAV peptide (active fragment derived from laminin) density gradient via covalent coupling of glutaraldehyde. The NGCs with these features could guide the migration of SCs directionally in vitro, and enhance the regeneration of sciatic nerves in rats in vivo. On the linear ridge/groove micro pattern with a width of 20/40 μm, SCs are arranged and extended along the stripe direction through contact guidance. Micropatterns promote the formation of a spindle or oval shape of SCs and enhance the directional extension of lamellipodia/filopodia. High peptide density regions significantly enhanced the number and migration rate of SCs. Compared with a single cue, the synergistic effect of the two cues significantly increased the elongation ratio of SCs and the efficiency of directional migration (80% of cells migrated to the high peptide density region). Micro patterns provide spatial topology guidance, while peptide gradients provide chemical chemotactic signals, jointly promoting the directed migration of SCs and the ordered extension of axons. Remote high peptide density design accelerates the migration of SCs to the distal end of injury and shortens nerve regeneration time. The directional migration of SCs enhances the secretion of NGF, promotes the directional arrangement of vascular endothelial cells, and forms a microenvironment of “neural vascular synergistic regeneration. Integrating spatial topography features and biosignals can effectively mediate and improve the interaction between cells and medical devices, which may provide a preferable microenvironment for tissue regeneration.

A Physical-Biochemical crosstalk: micropatterned PLCL film incorporated with a CQAASIKVAV peptide density gradient, which is manufactured into a conduit for peripheral nerve regeneration. Reproduced with permission159. Copyright 2022, Elsevier. B Physical-Electro coupling: the fabrication of NGCs by incorporating small amounts of single-layer GO and nanodiamond into nanogrooved PCL fibers for repairing peripheral nerve injury. Reproduced with permission160. Copyright 2025, John Wiley and Sons. C Biochemical –Electro combination: the fabrication of dECM-doped conductive hydrogel-filled PVA conduit for peripheral nerve regeneration. Reproduced with permission92. Copyright 2024, John Wiley and Sons.
Physical-electro coupling
Adequate biophysical cues on nerve scaffolds that resemble the mechanical and bioelectrical microenvironments represent an advanced technique for realizing desirable neural interfaces. Considering that SCs and axons are surrounded by a unique mechanical microenvironment and the electrically sensitive nature of peripheral nerve, Lei Zhan et al.160 designed a novel NGCs by incorporating single-layer GO and nanodiamond (ND) into nanogrooved PCL fibers. Combining these nanomaterials with the anisotropic topography of the fibrous conduit showed extraordinary synergy in enhancing the nerve regeneration process (Fig. 6B). It successfully triggered the myelinating capacity of SCs via Piezo1 signaling and further enabled the concurrent activation of NFAT and Krox-20 molecules. Combining nanomaterials with anisotropic topography can enable the myelinating capacity of SCs, thus offering a platform strategy toward fabricating a desirable microenvironment for peripheral nerve regeneration. A PDA-decorated PLCL NGCs has been fabricated to induce a synergetic effect of electrical stimulation and topological morphology for peripheral nerve regeneration161. This conduit with ES effectively stimulated the directional cell migration, adhesion, and elongation, and enhanced neuronal expression of SCs. The rat sciatic nerve crush model demonstrated that the conductive micropatterned conduit with ES promotes myelin sheath growth, faster nerve regeneration, and 20-fold functional recovery in vivo. These discoveries proved that physical-electro coupling is a promising tool for PNI treatment by providing the functional integration of physical guidance and bioelectrical stimulation.
Biochemical–Electro combination
Current clinical treatments using NGCs still face challenges in providing a biomimetic microenvironment to promote nerve repair. Lizhao Yan et al.92 pioneered the dECM conductive hydrogel composite NGCs, which realized the coordinated regulation of nerve regeneration by biochemical and electrical signals for the first time (Fig. 6C). ECM provides a bionic microenvironment that directly promotes SCs migration/proliferation. Conductive hydrogels enhance SCs’ activity by transmitting electrical signals to activate endogenous electric field-dependent pathways, such as ERK1/2 phosphorylation. RNA sequencing confirmed that the MAPK/PI3K Akt pathway was significantly concentrated under the synergistic effect of the two cues, driving SCs’ proliferation and axon myelination. This work demonstrated the synergistic effects of extracellular matrix (biochemical) and electrical cues in enhancing peripheral nerve regeneration. Delivering growth factors or cells with scaffolds can stimulate nerve regeneration processes, but chemical and electrical stimulation may have similar and additional benefits with fewer limitations. Rosalie Bordett et al.162 developed ionically conductive (IC) NGCs with synergistic electrical and chemical cues for extensive peripheral nerve defect regeneration. These chitosan-based NGCs released 4-aminopyridine (4-AP) from embedded halloysite nanotubes, and the NGC’s IC properties enabled transcutaneous ES without invasive electrodes. SCs are key effector cells for peripheral nerve regeneration, and this study focuses on verifying their functional changes under the synergistic stimulation of electrochemical signals. The combination of ES and 4-AP significantly increased the adhesion, proliferation and survival rate of SCs on scaffolds. The secretion of neurotrophic factors and their receptors by SCs under combined ES and 4-AP treatment was significantly higher than with 4-AP or ES alone, confirmed at both gene and protein levels. The combined treatment also enhanced myelin sheath expression and neovascularization in vivo, with elevated levels of neurotrophic genes (NGF, NT-3, BDNF, GDNF), their receptors (TRKA, TRKB, TRKC), and the endothelial marker CD31. ES and 4-AP form a powerful combination for nerve regeneration, leveraging neurotrophic factor-driven pathways and vascular support to maximize functional recovery.
Physical-biochemical-electro integration
Incorporating multiple cues within a single NCG can maximize the impact of guidance signaling and hold promise for repairing significant nerve gaps. However, integrating various guiding cues into a conduit remains a considerable challenge. At the developing phase (multifactor NGC), numerous element combinations exist for accurate function recovery with the growing applications of GO, 3D printing, and targeting ES163. A novel poly(3,4-ethylenedioxythiophene) (PEDOT)-integrated fish swim bladder (FSB) has been presented as a conductive nerve conduit with ordered topology and electrical stimulation to promote nerve regeneration164. The adhesive peptides (IKVAV) were also successfully incorporated onto the decellularized FSB substrate through pre-coating with PDA. The obtained PEDOT/IKVAV-integrated FSB substrate exhibited multiple clues of nerve regeneration induced by topology (physical signal), conductivity (electrical signal), and bioactive peptide (biochemical signal). The multimodal synergy significantly promoted the directional extension of neurites and SC-mediated myelination. Yuting Cai et al.165 further proposed a triple-guided multichannel hydrogel conduit, which synergistically enhanced peripheral nerve repair through the whole topology guidance (multichannel structure formed by directional freezing), conductive gradient (dopamine reduction of GO), and biochemical gradient (NGF concentration gradient) (Fig. 7). SCs were aligned along the direction of the anisotropic conduit. The cell density increased with the increase in guiding clues. Conductance gradient and NGF gradient enhance SCs migration. Conductance gradient may also promote SCs to wrap axons and form myelin sheath by regulating the cell membrane potential. This work found that the presence of a conductivity gradient could enhance the regeneration of peripheral nerves. However, the mechanism was unclear, and with limited relative reports, deeper research was necessary for validation. It also confirmed that the synergistic effect of multiple guidance cues significantly enhanced peripheral nerve regeneration compared to the use of a single cue. This inspired one of the research and development strategies of NGCs to integrate more guiding clues (such as stiffness gradient clues) into a single conduit to obtain the treatment effect similar to that of autologous transplantation. In addition, the potential mechanism of its synergistic effect deserves further study, especially the possibility of antagonism between different cues.

Physical-Biochemical-Electro integration: the fabrication of triple-cue-guided multichannel hydrogel conduit to synergistically enhance peripheral nerve repair. Reproduced with permission165. Copyright 2025, American Chemical Society.
Conclusion and future perspectives
Functional NGCs have demonstrated significant potential in orchestrating SC-mediated regeneration for critical PNIs. By integrating multimodal strategies—physical guidance (aligned nanofibers, stiffness gradients, porous architectures), biochemical signaling (ECM proteins, dECM, neurotrophic gradients), and bioelectric regulation (conductive polymers, piezoelectric materials)—NGCs can dynamically modulate SC plasticity across regenerative phases. Therefore, incorporating multiple cues within a single NCG can maximize the impact of guidance signaling and hold promise for repairing significant nerve gaps. NGCs are developing towards multifunctional composite scaffolds that integrate biomaterials, drug delivery systems, cell loading, and dynamic regulatory mechanisms. These advancements enable precise control over SC dedifferentiation, Büngner band formation, and remyelination, achieving preclinical outcomes comparable to autografts in short-to-moderate nerve gaps.
Despite significant progress in NGC research, challenges exist, particularly in addressing long-distance neural defects. Many NGC strategies are still in the experimental stage. Clinical translation remains hindered by unresolved challenges: insufficient spatiotemporal control of bioactive cues, limited scalability for defects exceeding 5 cm, and incomplete understanding of SC mechanotransduction pathways. Furthermore, the interplay between SCs and ancillary cells (e.g., macrophages, fibroblasts) within the regenerative niche necessitates deeper exploration to optimize multicellular coordination. Further evaluation and validation of the role of NGCs in repairing extended nerve defects can provide data support for clinical applications. In addition, it is necessary to address issues such as manufacturing complexity, cost, repeatability, and safety to promote the clinical application of NGCs. Multidisciplinary collaboration and technological innovation can drive breakthroughs in neural conduit manufacturing, aiming to go beyond autologous nerve grafts and provide efficient and intelligent clinical solutions.
To advance the field, future efforts must prioritize the integration of multimodal cues into dynamically responsive NGCs. Hierarchical architectures combining aligned nanofibers, tunable stiffness gradients, and electroconductive hydrogels could emulate the native nerve’s electroactive, anisotropic microenvironment. For instance, piezoelectric PVDF scaffolds may synergize with decellularized ECM to deliver simultaneous mechanical, biochemical, and electrical signals, amplifying SC-driven repair. More precise designs for simulating nerve structures are needed. 3D bioprinting offers easy and fast manufacturing of complex NGCs structures166. It allows for the manufacture of NGCs with multiple branches, cavities, gradients, and carrier functions, as well as NGCs with even more stimulus responses (such as electrical reactions)167. Using its full clinical applicability requires a lot of research and innovation. Addressing challenges related to the selection of bio-ink, mechanical properties, and bioprinter parameters will help improve the effectiveness of bioprinting NGCs. Recent advances in artificial intelligence (AI)-driven NGC optimization demonstrate how machine learning can predict scaffold porosity and growth factor release kinetics, enabling patient-specific conduit designs168. Machine learning and deep learning algorithms can optimize biomaterial properties, such as predicting the ideal methacrylation degree of poly(glycerol sebacate) (PGS) to balance degradation rates and mechanical strength with patient-specific nerve gap requirements. AI-driven computer vision analyzes high-dimensional imaging data to quantify neurite outgrowth patterns and align them with topographical or electrical cues in conduit microstructures, enhancing axon guidance169.
The next frontier in peripheral nerve repair lies in transcending static NGC designs toward adaptive, “smart” systems that dynamically engage SC biology. By converging advances in material science, mechanobiology, and neuroengineering, next-generation conduits may ultimately achieve functional parity with autografts, offering scalable solutions to restore sensorimotor precision and improve patient quality of life.
Data availability
No datasets were generated or analyzed during the current study.
References
Houshyar, S., Bhattacharyya, A. & Shanks, R. Peripheral nerve conduit: materials and structures. ACS Chem. Neurosci. 10, 3349–3365 (2019).
Ray, W. Z. & Mackinnon, S. E. Management of nerve gaps: autografts, allografts, nerve transfers, and end-to-side Neurorrhaphy. Exp. Neurol. 223, 77–85 (2010).
Rizwana, N. et al. Insights on advances in 3D bioprinting for peripheral nerve regeneration. Adv. Therapeutics 8 (2025).
Yu, L., Bennett, C. J., Lin, C.-H., Yan, S. & Yang, J. Scaffold design considerations for peripheral nerve regeneration. J. Neural Eng. 21, 041001 (2024).
Jin, B. et al. Combining a density gradient of biomacromolecular nanoparticles with biological effectors in an electrospun fiber-based nerve guidance conduit to promote peripheral nerve repair. Adv. Sci. 10 (2022).
Mankavi, F., Ibrahim, R. & Wang, H. Advances in biomimetic nerve guidance conduits for peripheral nerve regeneration. Nanomaterials 13, 2528 (2023).
Zhou, W. et al. Perspectives on the novel Multifunctional Nerve Guidance Conduits: from specific regenerative procedures to motor function rebuilding. Adv. Mater. 36 (2024).
Nocera, G. & Jacob, C. Mechanisms of schwann cell plasticity involved in peripheral nerve repair after injury. Cell. Mol. Life Sci. 77, 3977–3989 (2020).
Smith, C. S., Orkwis, J. A., Bryan, A. E., Xu, Z. & Harris, G. M. The impact of physical, biochemical, and electrical signaling on Schwann cell plasticity. Eur. J. Cell Biol. 101, 151277 (2022).
Arthur-Farraj, P. J. et al. C-jun reprograms Schwann cells of injured nerves to generate a repair cell essential for regeneration. Neuron 75, 633–647 (2012).
Wang, Y. et al. Orientated guidance of peripheral nerve regeneration using conduits with a microtube array sheet (mtas). ACS Appl. Mater. Interfaces 7, 8437–8450 (2015).
Li, J. & Gao, W. Fabrication and characterization of 3D microtubular collagen scaffolds for peripheral nerve repair. J. Biomater. Appl. 33, 541–552 (2018).
Li, G. et al. Porous chitosan scaffolds with surface micropatterning and inner porosity and their effects on Schwann cells. Biomaterials 35, 8503–8513 (2014).
Mobasseri, A. et al. Polymer scaffolds with preferential parallel grooves enhance nerve regeneration. Tissue Eng. A 21, 1152–1162 (2015).
Zhang, Z. et al. Submicron-grooved films modulate the directional alignment and biological function of Schwann cells. J. Funct. Biomater. 14, 238 (2023).
Jessen, K. R. & Mirsky, R. The repair Schwann cell and its function in regenerating nerves. J. Physiol. 594, 3521–3531 (2016).
Gomez-Sanchez, J. A. et al. Schwann cell autophagy, myelinophagy, initiates myelin clearance from injured nerves. J. Cell Biol. 210, 153–168 (2015).
Jang, S. Y. et al. Autophagic myelin destruction by Schwann cells during Wallerian degeneration and segmental demyelination. Glia 64, 730–742 (2015).
Klein, D. & Martini, R. Myelin and macrophages in the PNS: An intimate relationship in trauma and disease. Brain Res. 1641, 130–138 (2016).
Brosius Lutz, A. et al. Schwann cells use TAM receptor-mediated phagocytosis in addition to autophagy to clear myelin in a mouse model of nerve injury. Proc. Natil Acad. Sci. 114 (2017).
Gomez-Sanchez, J. A. et al. After nerve injury, lineage tracing shows that myelin and remak Schwann cells elongate extensively and branch to form repair Schwann cells, which shorten radically on Remyelination. J. Neurosci. 37, 9086–9099 (2017).
Fontana, X. et al. C-jun in Schwann cells promotes axonal regeneration and motoneuron survival via paracrine signaling. J. Cell Biol. 198, 127–141 (2012).
Rosenberg, A. F., Isaacman-Beck, J., Franzini-Armstrong, C. & Granato, M. Schwann cells and deleted in colorectal carcinoma direct regenerating motor axons towards their original path. J. Neurosci. 34, 14668–14681 (2014).
Feltri, M. L., Poitelon, Y. & Previtali, S. C. How Schwann cells sort axons. Neuroscientist 22, 252–265 (2015).
Sundaram, V. K. et al. Adipo-glial signaling mediates metabolic adaptation in peripheral nerve regeneration. Cell Metabolism 35 (2023).
Babetto, E., Wong, K. M. & Beirowski, B. A glycolytic shift in Schwann cells supports injured axons. Nat. Neurosci. 23, 1215–1228 (2020).
Jha, M. K. & Morrison, B. M. Glia-neuron energy metabolism in health and diseases: New insights into the role of Nervous System Metabolic Transporters. Exp. Neurol. 309, 23–31 (2018).
Michailov, G. V. et al. Axonal neuregulin-1 regulates myelin sheath thickness. Science 304, 700–703 (2004).
Stassart, R. M. et al. A role for Schwann cell–derived neuregulin-1 in remyelination. Nat. Neurosci. 16, 48–54 (2012).
Fernando, R. N. et al. Optimal myelin elongation relies on yap activation by axonal growth and inhibition by CRB3/Hippo pathway. Nat. Commun. 7 (2016).
Tricaud, N. Myelinating Schwann cell polarity and mechanically-driven myelin sheath elongation. Front. Cell. Neurosci. 11 (2018).
Amor, V. et al. Long-term maintenance of Na+Channels at nodes of Ranvier depends on glial contact mediated by Gliomedin and NRCAM. J. Neurosci. 34, 5089–5098 (2014).
Negro, S. et al. cxcl12α/sdf-1 from Perisynaptic Schwann cells promotes regeneration of injured motor axon terminals. EMBO Mol. Med. 9, 1000–1010 (2017).
Perez-Gonzalez, A. P. et al. Functional adaptation of glial cells at neuromuscular junctions in response to injury. Glia 70, 1605–1629 (2022).
Wu, T., Xue, J. & Xia, Y. Engraving the surface of electrospun microfibers with nanoscale grooves promotes the outgrowth of neurites and the migration of Schwann cells. Angew. Chem. 132, 15756–15762 (2020).
Ren, Y.-J. et al. Enhanced differentiation of human neural crest stem cells towards the Schwann cell lineage by aligned electrospun fiber matrix. Acta Biomaterialia 9, 7727–7736 (2013).
Gnavi, S. et al. The effect of electrospun gelatin fibers alignment on Schwann cell and axon behavior and organization in the perspective of artificial nerve design. Int. J. Mol. Sci. 16, 12925–12942 (2015).
Wang, W. et al. Effects of schwann cell alignment along the oriented electrospun chitosan nanofibers on nerve regeneration. J. Biomed. Mater. Res. Part A 91A, 994–1005 (2008).
Chen, X. et al. Silk-inspired fiber implant with multi-cues enhanced bionic microenvironment for promoting peripheral nerve repair. Biomater. Adv. 135, 112674 (2022).
Wu, S. et al. Guiding mesenchymal stem cells into myelinating Schwann cell-like phenotypes by using electrospun core–sheath nanoyarns. ACS Biomater. Sci. Eng. 5, 5284–5294 (2019).
Li, G. et al. Bionic microenvironment-inspired synergistic effect of anisotropic micro-nanocomposite topology and biology cues on peripheral nerve regeneration. Sci. Adv. 7 (2021).
Manganas, P. et al. The role of mechanobiology on the Schwann Cell response: a tissue engineering perspective. Front. Cell. Neurosci. 16 (2022).
Rosso, G. et al. Matrix stiffness mechanosensing modulates the expression and distribution of transcription factors in Schwann cells. Bioeng. Transl. Med. 7 (2021).
Evans, E. B., Brady, S. W., Tripathi, A. & Hoffman-Kim, D. Schwann cell durotaxis can be guided by physiologically relevant stiffness gradients. Biomater. Res. 22 (2018).
Gu, Y. et al. The influence of substrate stiffness on the behavior and functions of Schwann cells in culture. Biomaterials 33, 6672–6681 (2012).
Zhou, W., Stukel, J. M., Cebull, H. L. & Willits, R. K. Tuning the mechanical properties of poly(ethylene glycol) microgel-based scaffolds to increase 3D Schwann cell proliferation. Macromol. Biosci. 16, 535–544 (2016).
Naghilou, A. et al. Insights into the material properties of dragline spider silk affecting Schwann cell migration. Int. J. Biol. Macromol 244, 125398 (2023).
Gao, S. et al. 3D-bioprinted Gelma nerve guidance conduits promoted peripheral nerve regeneration by inducing trans-differentiation of MSCs into SCLCs via Piezo1/YAP axis. Mater. Today Adv. 17, 100325 (2023).
Acheta, J. et al. Piezo channels contribute to the regulation of myelination in Schwann cells. Glia 70, 2276–2289 (2022).
Chang, J. et al. Highly permeable genipin-cross-linked gelatin conduits enhance peripheral nerve regeneration. Artif. Organs 33, 1075–1085 (2009).
Johnson, L. D. V., Aleemardani, M., Atkins, S., Boissonade, F. M. & Claeyssens, F. Emulsion templated composites: porous nerve guidance conduits for peripheral nerve regeneration. Mater. Des. 239, 112779 (2024).
Wan, T. et al. The porous structure of peripheral nerve guidance conduits: Features, fabrication, and implications for peripheral nerve regeneration. Int. J. Mol. Sci. 24, 14132 (2023).
Zou, S. et al. Electrospun regenerated antheraea pernyi silk fibroin scaffolds with improved pore size, mechanical properties and cytocompatibility using mesh collectors. J. Mater. Chem. B 9, 5514–5527 (2021).
Yin, K., Divakar, P. & Wegst, U. G. Plant-derived nanocellulose as structural and mechanical reinforcement of freeze-cast chitosan scaffolds for biomedical applications. Biomacromolecules 20, 3733–3745 (2019).
Huang, L. et al. A compound scaffold with uniform longitudinally oriented guidance cues and a porous sheath promotes peripheral nerve regeneration in vivo. Acta Biomaterialia 68, 223–236 (2018).
Lee, H. S. et al. Development of a regenerative porous PLCL nerve guidance conduit with swellable hydrogel-based microgrooved surface pattern via 3D printing. Acta Biomaterialia 141, 219–232 (2022).
Yoo, J. et al. Augmented peripheral nerve regeneration through elastic nerve guidance conduits prepared using a porous PLCL membrane with a 3D printed collagen hydrogel. Biomater. Sci. 8, 6261–6271 (2020).
Bhatnagar, D. et al. Fibrin Glue as a stabilization strategy in peripheral nerve repair when using porous nerve guidance conduits. J. Mater. Sci.: Mater. Med. 28 (2017).
Oh, S. H. et al. Peripheral nerve regeneration within an asymmetrically porous PLGA/pluronic F127 nerve guide conduit. Biomaterials 29, 1601–1609 (2008).
Pinzon-Herrera, L., Magness, J., Apodaca-Reyes, H., Sanchez, J. & Almodovar, J. Surface modification of nerve guide conduits with ECM Coatings and investigating their impact on Schwann Cell response. Adv. Healthcare Mater. 13 (2024).
Klein, S., Prantl, L., Vykoukal, J., Loibl, M. & Felthaus, O. Differential effects of coating materials on viability and migration of Schwann cells. Materials 9, 150 (2016).
Han, S. et al. Chiral hydrogel nerve conduit boosts peripheral nerve regeneration via regulation of Schwann Cell reprogramming. ACS Nano 18, 28358–28370 (2024).
Chen, P., Cescon, M. & Bonaldo, P. The role of collagens in peripheral nerve myelination and function. Mol. Neurobiol. 52, 216–225 (2014).
Chernousov, M. A. et al. Glypican-1 and α4(V) collagen are required for Schwann cell myelination. J. Neurosci. 26, 508–517 (2006).
Li, G. et al. Hierarchically aligned gradient collagen micropatterns for rapidly screening Schwann cells behavior. Colloids Surf. B: Biointerfaces 176, 341–351 (2019).
Schuh, C. M. A. P., Day, A. G. E., Redl, H. & Phillips, J. An optimized collagen-fibrin blend engineered neural tissue promotes peripheral nerve repair. Tissue Eng. A 24, 1332–1340 (2018).
Vleggeert-Lankamp, C. L. et al. Adhesion and proliferation of human Schwann cells on adhesive coatings. Biomaterials 25, 2741–2751 (2004).
Chen, W. et al. Schwann cell-like cells derived from human amniotic mesenchymal stem cells promote sciatic nerve repair through an exosome-induced Sox2/FN1 pathway in vitro. Int. J. Mol. Med. 49 (2022).
Yu, P. et al. Effects of ECM proteins (laminin, fibronectin, and type IV collagen) on the biological behavior of Schwann cells and their roles in the process of remyelination after peripheral nerve injury. Front. Bioeng. Biotechnol. 11 (2023).
Milner, R. et al. Division of labor of Schwann cell integrins during migration on peripheral nerve extracellular matrix ligands. Dev. Biol. 185, 215–228 (1997).
Gonzalez-Perez, F. et al. Stabilization, rolling, and addition of other extracellular matrix proteins to collagen hydrogels improve regeneration in chitosan guides for long peripheral nerve gaps in rats. Neurosurgery 80, 465–474 (2017).
Mosahebi, A., Wiberg, M. & Terenghi, G. Addition of fibronectin to alginate matrix improves peripheral nerve regeneration in tissue-engineered conduits. Tissue Eng. 9, 209–218 (2003).
Mukhatyar, V. J. et al. Role of fibronectin in topographical guidance of neurite extension on electrospun fibers. Biomaterials 32, 3958–3968 (2011).
Armstrong, S. J., Wiberg, M., Terenghi, G. & Kingham, P. J. ECM molecules mediate both Schwann cell proliferation and activation to enhance neurite outgrowth. Tissue Eng. 13, 2863–2870 (2007).
Feltri, M. L. & Wrabetz, L. Laminins and their receptors in Schwann cells and hereditary neuropathies. J. Peripheral Nerv. Syst. 10, 128–143 (2005).
Chernousov, M. A., Yu, W., Chen, Z., Carey, D. J. & Strickland, S. Regulation of Schwann cell function by the extracellular matrix. Glia 56, 1498–1507 (2008).
McKee, K. K. et al. Schwann cell myelination requires integration of laminin activities. J. Cell Sci. https://doi.org/10.1242/jcs.107995 (2012).
Yu, W.-M., Feltri, M. L., Wrabetz, L., Strickland, S. & Chen, Z.-L. Schwann cell-specific ablation of laminin γ1 causes apoptosis and prevents proliferation. J. Neurosci. 25, 4463–4472 (2005).
Itoh, S. et al. Hydroxyapatite-coated tendon chitosan tubes with adsorbed laminin peptides facilitate nerve regeneration in vivo. Brain Res. 993, 111–123 (2003).
Nune, M., Subramanian, A., Krishnan, U. M., Kaimal, S. S. & Sethuraman, S. Self-assembling peptide nanostructures on aligned poly(lactide-co-glycolide) nanofibers for the functional regeneration of sciatic nerve. Nanomedicine 12, 219–235 (2017).
Nune, M., Subramanian, A., Krishnan, U. M. & Sethuraman, S. Peptide nanostructures on nanofibers for peripheral nerve regeneration. J. Tissue Eng. Regenerative Med. https://doi.org/10.1002/term.2860 (2019).
Li, X. et al. The application of collagen in the repair of peripheral nerve defect. Front. Bioeng. Biotechnol. 10 (2022).
Gao, H. et al. Surface topologized ovalbumin scaffolds containing YIGSR peptides for modulating Schwann Cell behavior. Int. J. Biol. Macromol. 253, 127015 (2023).
Lopez-Silva, T. L. et al. Self-assembling multidomain peptide hydrogels accelerate peripheral nerve regeneration after Crush Injury. Biomaterials 265, 120401 (2021).
Jiang, M., Chen, M. & Liu, N. Interactions between Schwann cell and extracellular matrix in peripheral nerve regeneration. Front. Neurol. 15 (2024).
Fan, N. et al. E-jet 3D printed aligned nerve guidance conduits incorporated with decellularized extracellular matrix hydrogel encapsulating extracellular vesicles for peripheral nerve repair. Acta Biomaterialia 194, 122–139 (2025).
Jin, B. et al. Microtubes with gradient decellularized porcine sciatic nerve matrix from microfluidics for sciatic nerve regeneration. Bioact. Mater. 21, 511–519 (2023).
Deng, R. et al. Decellularized extracellular matrix containing electrospun fibers for nerve regeneration: A comparison between core–shell structured and preblended composites. Adv. Fiber Mater. 4, 503–519 (2022).
Xue, Z. et al. Composite poly(lactic-co-glycolic acid) and a decellularized extracellular matrix conduit with Schwann cells for enhanced nerve regeneration. Bio-Des. Manuf. 8, 359–374 (2025).
Agarwal, G. et al. Decellularized porcine peripheral nerve based injectable hydrogels as a Schwann cell carrier for injured spinal cord regeneration. J. Neural Eng. 21, 046002 (2024).
Wang, Y. et al. Biodegradable polyurethane-incorporating decellularized spinal cord matrix scaffolds enhance Schwann cell reprogramming to promote peripheral nerve repair. J. Mater. Chem. B 11, 2115–2128 (2023).
Yan, L. et al. Constructing nerve guidance conduit using decm-doped conductive hydrogel to promote peripheral nerve regeneration. Adv. Funct. Mater. 34 (2024).
Xu, Y. et al. Engineering cell-derived extracellular matrix for peripheral nerve regeneration. Mater. Today Bio 27, 101125 (2024).
Aksel, H., Sarkar, D., Lin, M. H., Buck, A. & Huang, G. T.-J. Cell-derived extracellular matrix proteins in colloidal Microgel as a self-assembly hydrogel for Regenerative Endodontics. J. Endod. 48, 527–534 (2022).
Assunção, M. et al. Cell-derived extracellular matrix for tissue engineering and Regenerative Medicine. Front. Bioeng. Biotechnol. 8 (2020).
Junka, R. & Yu, X. Novel acellular scaffold made from decellularized Schwann cell sheets for peripheral nerve regeneration. Regenerative Eng. Transl. Med. 1, 22–31 (2015).
Nune, M., Bhat, M. & Nagarajan, A. Design of ECM functionalized polycaprolactone aligned nanofibers for peripheral nerve tissue engineering. J. Med. Biol. Eng. 42, 147–156 (2022).
Nagarajan, A. et al. Polycaprolactone/graphene oxide/acellular matrix nanofibrous scaffolds with antioxidant and promyelinating features for the treatment of peripheral demyelinating diseases. J. Mater. Sci.: Mater. Med. 34 (2023).
Al-Hadeethi, Y. et al. Schwann cell-matrix coated PCL-MWCNT multifunctional nanofibrous scaffolds for neural regeneration. RSC Adv. 13, 1392–1401 (2023).
Li, R. et al. Nerve growth factor activates autophagy in Schwann cells to enhance myelin debris clearance and to expedite nerve regeneration. Theranostics 10, 1649–1677 (2020).
QIN, J. et al. Concentrated growth factor promotes Schwann cell migration partly through the integrin β1-mediated activation of the focal adhesion kinase pathway. Int. J. Mol. Med. 37, 1363–1370 (2016).
QIN, J. et al. Concentrated growth factor increases Schwann cell proliferation and neurotrophic factor secretion and promotes functional nerve recovery in vivo. Int. J. Mol. Med. 37, 493–500 (2015).
Iwase, T., Jung, C. G., Bae, H., Zhang, M. & Soliven, B. Glial cell line-derived neurotrophic factor-induced signaling in Schwann cells. J. Neurochem. 94, 1488–1499 (2005).
Johnson, B. N. et al. 3D printed anatomical nerve regeneration pathways. Adv. Funct. Mater. 25, 6205–6217 (2015).
Madduri, S., di Summa, P., Papaloïzos, M., Kalbermatten, D. & Gander, B. Effect of controlled co-delivery of synergistic neurotrophic factors on early nerve regeneration in rats. Biomaterials 31, 8402–8409 (2010).
Basu, S. et al. In vitro modulation of Schwann cell behavior by VEGF and PDGF in an inflammatory environment. Sci. Rep. 12 (2022).
Thakkar, V., Mehdipour, M. & Chang, S. Unlocking nerve regeneration: Electrical stimulation and bioscaffolds to enhance peripheral nerve regeneration. Front. Neurosci. 19 (2025).
Li, X. et al. Electrical stimulation accelerates Wallerian degeneration and promotes nerve regeneration after sciatic nerve injury. Glia 71, 758–774 (2022).
Izhiman, Y. & Esfandiari, L. Emerging role of extracellular vesicles and exogenous stimuli in molecular mechanisms of peripheral nerve regeneration. Front. Cell. Neurosci. 18 (2024).
Yao, L., Li, Y., Knapp, J. & Smith, P. Exploration of molecular pathways mediating electric field-directed Schwann cell migration by RNA-seq. J. Cell. Physiol. 230, 1515–1524 (2015).
Kim, I. S. et al. Biphasic electrical targeting plays a significant role in Schwann Cell activation. Tissue Eng. A 17, 1327–1340 (2011).
Ong Sio, L. C., Hom, B., Garg, S. & Abd-Elsayed, A. Mechanism of action of peripheral nerve stimulation for chronic pain: a narrative review. Int. J. Mol. Sci. 24, 4540 (2023).
Ino, D. et al. Neuronal regulation of Schwann cell mitochondrial ca2+ signaling during myelination. Cell Rep. 12, 1951–1959 (2015).
Hu, M. et al. Electrical stimulation enhances neuronal cell activity mediated by Schwann cell derived exosomes. Sci. Rep. 9 (2019).
Hardy, P. B., Wang, B. Y., Chan, K. M., Webber, C. A. & Senger, J. B. The use of electrical stimulation to enhance recovery following peripheral nerve injury. Muscle Nerve 70, 1151–1162 (2024).
Zhao, Y. et al. Peripheral nerve injury repair by electrical stimulation combined with graphene-based scaffolds. Front. Bioeng. Biotechnol. 12 (2024).
Ni, L. et al. Electrical stimulation therapy for peripheral nerve injury. Front. Neurol. 14 (2023).
Zhao, Y. et al. Application of conductive ppy/SF composite scaffold and electrical stimulation for neural tissue engineering. Biomaterials 255, 120164 (2020).
Nune, M., Manchineella, S. & TG, K. S. N. Melanin incorporated electroactive and antioxidant silk fibroin nanofibrous scaffolds for nerve tissue engineering. Mater. Sci. Eng. C 94, 17–25 (2019).
Lu, N. et al. Fabrication of the conductive polypyrrole/silk fibroin nanofibrous mats and its biodegradable activity. Bull. Mater. Sci. 48 (2025).
Jiang, L. et al. The role of gel-ppy-modified nerve conduit on the repair of sciatic nerve defect in rat model. FASEB J. 37 (2023).
Xiong, F. et al. Aligned electroactive electrospun fibrous scaffolds for peripheral nerve regeneration. ACS Appl. Mater. Interfaces 15, 41385–41402 (2023).
Yi, H. et al. Conducting polymer-based scaffolds for neuronal tissue engineering. J. Mater. Chem. B 11, 11006–11023 (2023).
Xu, D. et al. Micro-nanostructured polyaniline assembled in cellulose matrix via interfacial polymerization for applications in nerve regeneration. ACS Appl. Mater. Interfaces 8, 17090–17097 (2016).
Yi, Z. et al. An electroconductive hydrogel with injectable and self-healing properties accelerates peripheral nerve regeneration and motor functional recovery. Chem. Eng. J. 478, 147261 (2023).
Zheng, Y. et al. Electrodeposition of chitosan/graphene oxide conduit to enhance peripheral nerve regeneration. Neural Regeneration Res. 18, 207 (2023).
Fang, X. et al. Reduced graphene oxide–gelma–PCL hybrid nanofibers for Peripheral Nerve Regeneration. J. Mater. Chem. B 8, 10593–10601 (2020).
Zheng, F. et al. The electrostimulation and scar inhibition effect of chitosan/oxidized hydroxyethyl cellulose/reduced graphene oxide/asiaticoside liposome based hydrogel on peripheral nerve regeneration in vitro. Mater. Sci. Eng. C 109, 110560 (2020).
Zhao, Y. et al. Conductive biocomposite hydrogels with multiple biophysical cues regulate Schwann cell behaviors. J. Mater. Chem. B 10, 1582–1590 (2022).
Wu, Y., Wang, L., Guo, B., Shao, Y. & Ma, P. X. Electroactive biodegradable polyurethane significantly enhanced Schwann cells myelin gene expression and neurotrophin secretion for peripheral nerve tissue engineering. Biomaterials 87, 18–31 (2016).
Han, H. J. et al. Uric acid inhibits renal proximal tubule cell proliferation via at least two signaling pathways involving PKC, MAPK, CPLA2, and NF-ΚB. Am. J. Physiol.-Renal Physiol. 292 (2007).
Ajiteru, O. et al. A 3D printable electroconductive biocomposite Bioink based on silk fibroin-conjugated graphene oxide. Nano Lett. 20, 6873–6883 (2020).
Ma, J. et al. Crucial role of lateral size for graphene oxide in activating macrophages and stimulating pro-inflammatory responses in cells and animals. ACS Nano 9, 10498–10515 (2015).
Wang, K. et al. Biocompatibility of graphene oxide. Nanoscale Res. Lett. 6 (2010).
Li, X. et al. CNT/sericin conductive nerve guidance conduit promotes functional recovery of transected peripheral nerve injury in a rat model. ACS Appl. Mater. Interfaces 12, 36860–36872 (2020).
Ghosh, S., Dhiman, M., Chauhan, S., Roy, P. & Lahiri, D. Dual functional electroconductive biofortified electrospun scaffold functionalized with MWCNTs and Bacopa Monnieri for accelerated peripheral nerve regeneration. Small 21, (2025).
Huang, Z. et al. Tracing carbon nanotubes (CNTS) in rat peripheral nerve regenerated with conductive conduits composed of poly(lactide-co-glycolide) and fluorescent CNTS. ACS Biomater. Sci. Eng. 6, 6344–6355 (2020).
Zhang, J. et al. Conductive composite fiber with optimized alignment guides neural regeneration under electrical stimulation. Adv. Healthcare Mater. 10 (2020).
Shrestha, S. et al. A conducting neural interface of polyurethane/silk-functionalized multiwall carbon nanotubes with enhanced mechanical strength for neuroregeneration. Mater. Sci. Eng. C 102, 511–523 (2019).
Wu, S. et al. Electrospun conductive nanofiber yarns for accelerating mesenchymal stem cells differentiation and maturation into Schwann cell-like cells under a combination of electrical stimulation and chemical induction. Acta Biomaterialia 139, 91–104 (2022).
Redondo-Gómez, C. et al. Recent advances in carbon nanotubes for nervous tissue regeneration. Adv. Polym. Technol. 2020, 1–16 (2020).
Ma, Y., Wang, H., Wang, Q., Cao, X. & Gao, H. Piezoelectric conduit combined with multi-channel conductive scaffold for peripheral nerve regeneration. Chem. Eng. J. 452, 139424 (2023).
Lv, Y. et al. Piezoelectric polyvinylidene fluoride-trifluoroethylene/reduced graphene oxide/polycaprolactone fiber material: Modulating neutrophil extracellular traps and reshaping the immune microenvironment in peripheral nerves. Adv. Fiber Mater. 7, 645–663 (2025).
Wang, X., Stefanello, S. T., Shahin, V. & Qian, Y. From mechanoelectric conversion to tissue regeneration: Translational progress in Piezoelectric Materials. Adv. Mater. 37 (2025).
Cheng, Y. et al. 3D structured self-powered PVDF/PCL scaffolds for Peripheral Nerve Regeneration. Nano Energy 69, 104411 (2020).
Wu, S. et al. Aligned fibrous PVDF-TrFE scaffolds with Schwann cells support neurite extension and myelination in vitro. J. Neural Eng. 15, 056010 (2018).
Qian, Y. et al. Mechano-informed biomimetic polymer scaffolds by incorporating self-powered zinc oxide nanogenerators enhance motor recovery and neural function. Small 16 (2020).
Gao, H. et al. Biomimetic-inspired piezoelectric ovalbumin/BATIO3 scaffolds synergizing with anisotropic topology for modulating Schwann Cell and DRG behavior. Int. J. Biol. Macromol. 271, 132394 (2024).
Zhang, H. et al. Electrospun piezoelectric scaffold with external mechanical stimulation for promoting regeneration of peripheral nerve injury. Biomacromolecules 24, 3268–3282 (2023).
Wang, Q. et al. Engineered PVDF/PLCL/pedot dual electroactive nerve conduit to mediate peripheral nerve regeneration by modulating the immune microenvironment. Adv. Funct. Mater. 34. (2024).
Liu, Y. et al. An easy nanopatch promotes peripheral nerve repair through wireless ultrasound-electrical stimulation in a band-aid-like way. Adv. Funct. Mater. 34 (2024).
Du, Y. et al. Recent progress on hydrogel-based piezoelectric devices for biomedical applications. Micromachines 14, 167 (2023).
Vinikoor, T. et al. Injectable and biodegradable piezoelectric hydrogel for osteoarthritis treatment. Nat. Commun. 14 (2023).
Wang, F. et al. An ultrasound-responsive hydrogel with piezoelectric-enhanced electrokinetic effect accelerates neurovascular regeneration for diabetic wound healing. Mater. Today 84, 48–64 (2025).
Wu, L. et al. Piezoelectric materials for neuroregeneration: a review. Biomater. Sci. 11, 7296–7310 (2023).
Xu, D. et al. Ultrasound-responsive aligned piezoelectric nanofibers derived hydrogel conduits for peripheral nerve regeneration. Adv. Mater. 36 (2024).
Vijayavenkataraman, S. Nerve guide conduits for Peripheral Nerve Injury Repair: a review on design, materials and fabrication methods. Acta Biomaterialia 106, 54–69 (2020).
Yang, S. et al. Aligned Fibrin/functionalized self-assembling peptide interpenetrating nanofiber hydrogel presenting multi-cues promotes peripheral nerve functional recovery. Bioact. Mater. 8, 529–544 (2022).
Zhang, D. et al. Micropatterns and peptide gradient on the inner surface of a guidance conduit synergistically promotes nerve regeneration in vivo. Bioact. Mater. 9, 134–146 (2022).
Zhan, L. et al. Anisotropic single-layer graphene/Nanodiamond loaded PCL conduits provide biophysical cues to manipulate nerve biomechanics and bioelectric function in the restoration of nerve microenvironment. Adv. Funct. Mater. 35 (2025).
Lu, S. et al. Polydopamine-decorated PLCL Conduit to induce synergetic effect of electrical stimulation and topological morphology for peripheral nerve regeneration. Small Methods 7 (2023).
Bordett, R. et al. Synergistic effects of electrical and chemical cues with biodegradable scaffolds for large peripheral nerve defect regeneration. Bioact. Mater. 49, 586–607 (2025).
Zhang, C. et al. Three potential elements of developing nerve guidance conduit for peripheral nerve regeneration. Adv. Funct. Mater. 33 (2023).
Zhang, H. et al. Pedot-integrated fish swim bladders as conductive nerve conduits. Advanced Science 11, (2024).
Cai, Y. et al. Triple-cue-guided multichannel hydrogel conduit to synergistically enhance peripheral nerve repair. ACS Nano 19, 22163–22178 (2025).
Rizwana, N. et al. Dual crosslinked antioxidant mixture of poly(vinyl alcohol) and cerium oxide nanoparticles as a Bioink for 3D bioprinting. ACS Appl. Nano Mater. 7, 18177–18188 (2023).
Joshi, A. et al. 4D printed programmable shape-morphing hydrogels as intraoperative self-folding nerve conduits for sutureless Neurorrhaphy. Adv. Healthcare Mater. 12 (2023).
Hassani, F. et al. Intelligent design of nerve guidance conduits: An artificial intelligence-driven fluid structure interaction study on modelling and optimization of nerve growth. Can. J. Chem. Eng. 103, 1978–2005 (2024).
Stewart, C. E. et al. Machine intelligence for nerve conduit design and production. J. Biol. Eng. 14 (2020).
Acknowledgements
This work was supported by the Science and Technology Program of Ningbo (Grant No. 2023Z187), the Medicine, Health, Science and Technology Program of Zhejiang Province (Grant No. 2024KY386), and the Ningbo Top Medical and Health Research Program (Grant No. 2022020506).
Author information
Authors and Affiliations
Contributions
K.Y. conducted an in-depth literature review, synthesized the structural framework of the manuscript, and wrote the initial draft. S.Y. contributed to the conceptual design of the review, curated recent advances in Schwann cell modulation, and revised the manuscript critically for intellectual content. X.T. collected and organized data regarding physical and bioelectrical cues in nerve guidance conduits and assisted in figure creation. X.H. focused on the biochemical aspects of nerve regeneration, including ECM signaling and decellularized matrix applications, and contributed to literature curation. T.S. reviewed clinical challenges of autografts, helped design the summary tables, and assisted in manuscript editing. H.C. supervised the entire project, integrated feedback from all co-authors, ensured scientific coherence across sections, and was the primary contributor in revising and finalizing the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yang, K., Yang, S., Teng, X. et al. Modulating Schwann cell behavior via functional nerve guidance conduits for enhanced peripheral nerve regeneration. npj Regen Med 10, 54 (2025). https://doi.org/10.1038/s41536-025-00443-w
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41536-025-00443-w
