
Abstract
Metastatic brain disease occurs in up to 30% of patients with lung, melanoma and breast cancers, and the median survival time remains less than a year. Treating these patients is a challenge because surgical approaches are limited and most chemotherapeutic drugs and immunotherapies are ineffective at crossing the blood–brain barrier (BBB). Given the unique abilities of macrophages to cross the BBB and exert their phagocytic function on tumour cells, we genetically engineer macrophages that express a chimaeric antigen receptor (CAR) targeting mesothelin (MSLN). To specifically target metastatic brain tumours, we fused the cells with the immune signalling molecule MyD88. This chimaeric antigen receptor macrophage (CARMA) penetrates the BBB and decreases brain metastasis growth in a humanized mouse model. MSLN–CARMA shows antigen-specific phagocytosis activity against tumour cells and exhibits a bystander effect by releasing TNF to act on surrounding tumour cells lacking the tumour antigen. These features of CARMA represent advantages over other immune therapies and CARMA may serve as a promising therapeutic tool for the treatment of brain metastasis.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others

Data availability
The single-cell-sequencing dataset in this study has been previously reported92. Our generated single-cell RNA-sequencing data are available in the NCBI Gene expression Omnibus database (GEO) with accession number GSE234832 (ref. 92). The analysis codes for survival analysis are shared in GitHub at https://github.com/QSong-github/BrMet (ref. 93) and in Zenodo. To investigate the influence of MSLN on the progression of lung cancer, we retrieved data on the overall survival and MSLN expression for patients at stages 1–4 from the TCGA lung cancer cohort. Patients lacking overall survival or MSLN expression data in the cohort were excluded. These patients were subsequently grouped into MSLN-High and MSLN-Low cohorts based on the median MSLN expression. The overall survival data for these two groups were analysed using Prism 9.0. We employed Kaplan–Meier survival analysis to determine the relationship between overall survival and MSLN gene expression in lung cancer patients. The references for the Kaplan–Meier survival analysis used in the study can be accessed in public repositories on the following websites: https://www.ncbi.nlm.nih.gov/gds, https://www.tfcheckpoint.org and http://kmplot.com/analysis/index.php?p=service. We obtained gene expression profiles from GEO datasets89,90,91 (GSE123902, GSE137797, GSE200563) and analysed the expression of MSLN in lung cancer and its association with brain metastasis. For the tumour antigen analysis, the references used in the study are available in a public repository provided by the Human Protein Atlas data (https://www.proteinatlas.org). To analyse immune cell populations in brain metastasis and primary CNS tumours, we utilized data from the URECA and TIMER 2.0 datasets. These datasets can be accessed in public repositories on the following websites: http://kmplot.com/analysis/, http://ureca-singlecell.kr/ and http://timer.comp-genomics.org/. We downloaded all immune cell profiles from URECA and examined the proportions of different immune cell populations in lung cancer brain metastasis. Source data are provided with this paper.
References
Boire, A., Brastianos, P. K., Garzia, L. & Valiente, M. Brain metastasis. Nat. Rev. Cancer 20, 4–11 (2020).
Kerschbaumer, J. et al. Sector irradiation vs. whole brain irradiation after resection of singular brain metastasis—a prospective randomized monocentric trial. Front. Oncol. 10, 591884 (2020).
Gregory, G. P. et al. Pembrolizumab plus dinaciclib in patients with hematologic malignancies: the phase 1b KEYNOTE-155 study. Blood Adv. 6, 1232–1242 (2022).
Ramos, C. A. et al. Anti-CD30 CAR-T cell therapy in relapsed and refractory Hodgkin lymphoma. J. Clin. Oncol. 38, 3794–3804 (2020).
O’Byrne, K. et al. Long-term comparative efficacy and safety of nivolumab plus ipilimumab relative to other first-line therapies for advanced non-small-cell lung cancer: a systematic literature review and network meta-analysis. Lung Cancer 177, 11–20 (2023).
Perets, R. et al. Safety and efficacy of quavonlimab, a novel anti-CTLA-4 antibody (MK-1308), in combination with pembrolizumab in first-line advanced non-small-cell lung cancer. Ann. Oncol. 32, 395–403 (2021).
Adusumilli, P. S. et al. A phase I trial of regional mesothelin-targeted CAR T-cell therapy in patients with malignant pleural disease, in combination with the anti-PD-1 agent pembrolizumab. Cancer Discov. 11, 2748–2763 (2021).
Maude, S. L., Teachey, D. T., Porter, D. L. & Grupp, S. A. CD19-targeted chimeric antigen receptor T-cell therapy for acute lymphoblastic leukemia. Blood 125, 4017–4023 (2015).
Shalabi, H. et al. Systematic evaluation of neurotoxicity in children and young adults undergoing CD22 chimeric antigen receptor T-cell therapy. J. Immunother. 41, 350–358 (2018).
Karschnia, P. et al. Clinical presentation, management, and biomarkers of neurotoxicity after adoptive immunotherapy with CAR T cells. Blood 133, 2212–2221 (2019).
Morris, E. C., Neelapu, S. S., Giavridis, T. & Sadelain, M. Cytokine release syndrome and associated neurotoxicity in cancer immunotherapy. Nat. Rev. Immunol. 22, 85–96 (2022).
Ma, R. Y., Black, A. & Qian, B. Z. Macrophage diversity in cancer revisited in the era of single-cell omics. Trends Immunol. 43, 546–563 (2022).
Cheng, S. et al. A pan-cancer single-cell transcriptional atlas of tumor infiltrating myeloid cells. Cell 184, 792–809.e23 (2021).
Kohno, K. et al. Inflammatory M1-like macrophages polarized by NK-4 undergo enhanced phenotypic switching to an anti-inflammatory M2-like phenotype upon co-culture with apoptotic cells. J. Inflamm. 18, 2 (2021).
Ma, W., Liu, Y., Ellison, N. & Shen, J. Induction of C-X-C chemokine receptor type 7 (CXCR7) switches stromal cell-derived factor-1 (SDF-1) signaling and phagocytic activity in macrophages linked to atherosclerosis. J. Biol. Chem. 288, 15481–15494 (2013).
Wei, Y. et al. The local immune landscape determines tumor PD-L1 heterogeneity and sensitivity to therapy. J. Clin. Invest. 129, 3347–3360 (2019).
Min, H., Jang, Y. H., Cho, I. H., Yu, S. W. & Lee, S. J. Alternatively activated brain-infiltrating macrophages facilitate recovery from collagenase-induced intracerebral hemorrhage. Mol. Brain 9, 42 (2016).
Chen, Z. et al. Cellular and molecular identity of tumor-associated macrophages in glioblastoma. Cancer Res. 77, 2266–2278 (2017).
Feng, X. et al. Loss of CX3CR1 increases accumulation of inflammatory monocytes and promotes gliomagenesis. Oncotarget 6, 15077–15094 (2015).
Lin, C. M. et al. Distinct tumor microenvironment at tumor edge as a result of astrocyte activation is associated with therapeutic resistance for brain tumor. Front. Oncol. 9, 307 (2019).
Brouckaert, G. et al. Phagocytosis of necrotic cells by macrophages is phosphatidylserine dependent and does not induce inflammatory cytokine production. Mol. Biol. Cell 15, 1089–1100 (2004).
Budai, Z. et al. Macrophages engulf apoptotic and primary necrotic thymocytes through similar phosphatidylserine-dependent mechanisms. FEBS Open Bio 9, 446–456 (2019).
Underhill, D. M., Bassetti, M., Rudensky, A. & Aderem, A. Dynamic interactions of macrophages with T cells during antigen presentation. J. Exp. Med. 190, 1909–1914 (1999).
Kachala, S. S. et al. Mesothelin overexpression is a marker of tumor aggressiveness and is associated with reduced recurrence-free and overall survival in early-stage lung adenocarcinoma. Clin. Cancer Res. 20, 1020–1028 (2014).
Bera, T. K. & Pastan, I. Mesothelin is not required for normal mouse development or reproduction. Mol. Cell. Biol. 20, 2902–2906 (2000).
Kim, N. et al. Single-cell RNA sequencing demonstrates the molecular and cellular reprogramming of metastatic lung adenocarcinoma. Nat. Commun. 11, 2285 (2020).
Winnicka, B. et al. CD13 is dispensable for normal hematopoiesis and myeloid cell functions in the mouse. J. Leukoc. Biol. 88, 347–359 (2010).
Chen, M. et al. Twenty-four-color full spectrum flow cytometry panel for minimal residual disease detection in acute myeloid leukemia. Open Med. 18, 20230745 (2023).
Liu, Y., Xia, Y. & Qiu, C. H. Functions of CD169 positive macrophages in human diseases (Review). Biomed. Rep. 14, 26 (2021).
Gentles, A. J. et al. The prognostic landscape of genes and infiltrating immune cells across human cancers. Nat. Med. 21, 938–945 (2015).
Argani, P. et al. Mesothelin is overexpressed in the vast majority of ductal adenocarcinomas of the pancreas: identification of a new pancreatic cancer marker by serial analysis of gene expression (SAGE). Clin. Cancer Res. 7, 3862–3868 (2001).
Hassan, R. et al. Detection and quantitation of serum mesothelin, a tumor marker for patients with mesothelioma and ovarian cancer. Clin. Cancer Res. 12, 447–453 (2006).
Oakley, M. S. et al. TCRbeta-expressing macrophages induced by a pathogenic murine malaria correlate with parasite burden and enhanced phagocytic activity. PLoS ONE 13, e0201043 (2018).
Marr, K. A. et al. Differential role of MyD88 in macrophage-mediated responses to opportunistic fungal pathogens. Infect. Immun. 71, 5280–5286 (2003).
Rodriguez-Cruz, A. et al. CD3+ macrophages deliver proinflammatory cytokines by a CD3- and transmembrane TNF-dependent pathway and are increased at the BCG-infection site. Front. Immunol. 10, 2550 (2019).
Zhang, L. et al. Pluripotent stem cell-derived CAR-macrophage cells with antigen-dependent anti-cancer cell functions. J. Hematol. Oncol. 13, 153 (2020).
Daigneault, M., Preston, J. A., Marriott, H. M., Whyte, M. K. & Dockrell, D. H. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PLoS ONE 5, e8668 (2010).
Petersen, S. L. et al. Autocrine TNFα signaling renders human cancer cells susceptible to Smac-mimetic-induced apoptosis. Cancer Cell 12, 445–456 (2007).
Wang, L., Du, F. & Wang, X. TNF-α induces two distinct caspase-8 activation pathways. Cell 133, 693–703 (2008).
Yang, J., Zhang, L., Yu, C., Yang, X. F. & Wang, H. Monocyte and macrophage differentiation: circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomark. Res. 2, 1 (2014).
Adachi, O. et al. Targeted disruption of the MyD88 gene results in loss of IL-1- and IL-18-mediated function. Immunity 9, 143–150 (1998).
Warner, N. & Nunez, G. MyD88: a critical adaptor protein in innate immunity signal transduction. J. Immunol. 190, 3–4 (2013).
Guo, Q., Xiao, X. & Zhang, J. MYD88 is a potential prognostic gene and immune signature of tumor microenvironment for gliomas. Front. Oncol. 11, 654388 (2021).
Weber, K. & Schilling, J. D. Distinct lysosome phenotypes influence inflammatory function in peritoneal and bone marrow-derived macrophages. Int. J. Inflam. 2014, 154936 (2014).
House, I. G. et al. Macrophage-derived CXCL9 and CXCL10 are required for antitumor immune responses following immune checkpoint blockade. Clin. Cancer Res. 26, 487–504 (2020).
Chow, M. T. et al. Intratumoral activity of the CXCR3 chemokine system is required for the efficacy of anti-PD-1 therapy. Immunity 50, 1498–1512.e5 (2019).
Feng, M. et al. Phagocytosis checkpoints as new targets for cancer immunotherapy. Nat. Rev. Cancer 19, 568–586 (2019).
Cabrales, P. RRx-001 acts as a dual small molecule checkpoint inhibitor by downregulating CD47 on cancer cells and SIRP-alpha on monocytes/macrophages. Transl. Oncol. 12, 626–632 (2019).
Denninger, J. K., Smith, B. M. & Kirby, E. D. Novel object recognition and object location behavioral testing in mice on a budget. J. Vis. Exp. https://doi.org/10.3791/58593 (2018).
Klichinsky, M. et al. Human chimeric antigen receptor macrophages for cancer immunotherapy. Nat. Biotechnol. 38, 947–953 (2020).
Call, M. E., Pyrdol, J., Wiedmann, M. & Wucherpfennig, K. W. The organizing principle in the formation of the T cell receptor-CD3 complex. Cell 111, 967–979 (2002).
Narayanan, P. et al. A composite MyD88/CD40 switch synergistically activates mouse and human dendritic cells for enhanced antitumor efficacy. J. Clin. Invest. 121, 1524–1534 (2011).
Wang, X. et al. Inducible MyD88/CD40 synergizes with IL-15 to enhance antitumor efficacy of CAR-NK cells. Blood Adv. 4, 1950–1964 (2020).
Collinson-Pautz, M. R. et al. Constitutively active MyD88/CD40 costimulation enhances expansion and efficacy of chimeric antigen receptor T cells targeting hematological malignancies. Leukemia 33, 2195–2207 (2019).
Foster, A. E. et al. Regulated expansion and survival of chimeric antigen receptor-modified T cells using small molecule-dependent inducible MyD88/CD40. Mol. Ther. 25, 2176–2188 (2017).
Prinzing, B. et al. MyD88/CD40 signaling retains CAR T cells in a less differentiated state. JCI Insight https://doi.org/10.1172/jci.insight.136093 (2020).
Fremond, C. M. et al. Fatal Mycobacterium tuberculosis infection despite adaptive immune response in the absence of MyD88. J. Clin. Invest. 114, 1790–1799 (2004).
Rackov, G. et al. p21 mediates macrophage reprogramming through regulation of p50-p50 NF-kappaB and IFN-beta. J. Clin. Invest. 126, 3089–3103 (2016).
Wang, N., Liang, H. & Zen, K. Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front. Immunol. 5, 614 (2014).
Gao, X. R. et al. NF-kappaB/let-7f-5p/IL-10 pathway involves in wear particle-induced osteolysis by inducing M1 macrophage polarization. Cell Cycle 17, 2134–2145 (2018).
Suzuki, E. & Umezawa, K. Inhibition of macrophage activation and phagocytosis by a novel NF-kappaB inhibitor, dehydroxymethylepoxyquinomicin. Biomed. Pharmacother. 60, 578–586 (2006).
Zhu, F., Yue, W. & Wang, Y. The nuclear factor kappa B (NF-kappaB) activation is required for phagocytosis of Staphylococcus aureus by RAW 264.7 cells. Exp. Cell. Res. 327, 256–263 (2014).
Li, Q. et al. Enhanced NF-kappaB activation and cellular function in macrophages lacking IkappaB kinase 1 (IKK1). Proc. Natl Acad. Sci. USA 102, 12425–12430 (2005).
Wang, X. & Lin, Y. Tumor necrosis factor and cancer, buddies or foes? Acta Pharmacol. Sin. 29, 1275–1288 (2008).
Holler, N. et al. Fas triggers an alternative, caspase-8-independent cell death pathway using the kinase RIP as effector molecule. Nat. Immunol. 1, 489–495 (2000).
Degterev, A. et al. Identification of RIP1 kinase as a specific cellular target of necrostatins. Nat. Chem. Biol. 4, 313–321 (2008).
Kim, Y. S., Morgan, M. J., Choksi, S. & Liu, Z. G. TNF-induced activation of the Nox1 NADPH oxidase and its role in the induction of necrotic cell death. Mol. Cell 26, 675–687 (2007).
Perez, D. & White, E. TNF-alpha signals apoptosis through a bid-dependent conformational change in Bax that is inhibited by E1B 19K. Mol. Cell 6, 53–63 (2000).
Alvarez, S., Blanco, A., Fresno, M. & Munoz-Fernandez, M. A. TNF-alpha contributes to caspase-3 independent apoptosis in neuroblastoma cells: role of NFAT. PLoS ONE 6, e16100 (2011).
Komohara, Y., Fujiwara, Y., Ohnishi, K. & Takeya, M. Tumor-associated macrophages: potential therapeutic targets for anti-cancer therapy. Adv. Drug Deliv. Rev. 99, 180–185 (2016).
Neniskyte, U., Vilalta, A. & Brown, G. C. Tumour necrosis factor alpha-induced neuronal loss is mediated by microglial phagocytosis. FEBS Lett. 588, 2952–2956 (2014).
Bernardino, L. et al. Modulator effects of interleukin-1β and tumor necrosis factor-α on AMPA-induced excitotoxicity in mouse organotypic hippocampal slice cultures. J. Neurosci. 25, 6734–6744 (2005).
Olmos, G. & Llado, J. Tumor necrosis factor alpha: a link between neuroinflammation and excitotoxicity. Mediators Inflamm. 2014, 861231 (2014).
Bi, M. et al. Tau exacerbates excitotoxic brain damage in an animal model of stroke. Nat. Commun. 8, 473 (2017).
Chao, M. P., Weissman, I. L. & Majeti, R. The CD47-SIRPα pathway in cancer immune evasion and potential therapeutic implications. Curr. Opin. Immunol. 24, 225–232 (2012).
Koh, E. et al. Exosome-SIRPα, a CD47 blockade increases cancer cell phagocytosis. Biomaterials 121, 121–129 (2017).
Nelson, H. H. et al. The relationship between tumor MSLN methylation and serum mesothelin (SMRP) in mesothelioma. Epigenetics 6, 1029–1034 (2011).
Castelletti, L., Yeo, D., van Zandwijk, N. & Rasko, J. E. J. Anti-mesothelin CAR T cell therapy for malignant mesothelioma. Biomark. Res. 9, 11 (2021).
Tokatlian, T. et al. Mesothelin-specific CAR-T cell therapy that incorporates an HLA-gated safety mechanism selectively kills tumor cells. J. Immunother. Cancer https://doi.org/10.1136/jitc-2021-003826 (2022).
Klopp, C. T., Alford, T. C., Bateman, J., Berry, G. N. & Winship, T. Fractionated intra-arterial cancer; chemotherapy with methyl bis amine hydrochloride; a preliminary report. Ann. Surg. 132, 811–832 (1950).
Boockvar, J. A. et al. Safety and maximum tolerated dose of superselective intraarterial cerebral infusion of bevacizumab after osmotic blood-brain barrier disruption for recurrent malignant glioma. Clinical article. J. Neurosurg. 114, 624–632 (2011).
Riina, H. A. et al. Short-term clinico-radiographic response to super-selective intra-arterial cerebral infusion of Bevacizumab for the treatment of vestibular schwannomas in neurofibromatosis type 2. Interv. Neuroradiol. 18, 127–132 (2012).
Riina, H. A. et al. Superselective intraarterial cerebral infusion of bevacizumab: a revival of interventional neuro-oncology for malignant glioma. J. Exp. Ther. Oncol. 8, 145–150 (2009).
Shin, B. J., Burkhardt, J. K., Riina, H. A. & Boockvar, J. A. Superselective intra-arterial cerebral infusion of novel agents after blood-brain disruption for the treatment of recurrent glioblastoma multiforme: a technical case series. Neurosurg. Clin. N. Am. 23, 323–329 (2012).
Westbury, G. Treatment of advanced cancer by extracorporeal perfusion and continuous intra-arterial infusion. Proc. R. Soc. Med. 55, 643–646 (1962).
Nguyen, D. X. et al. WNT/TCF signaling through LEF1 and HOXB9 mediates lung adenocarcinoma metastasis. Cell 138, 51–62 (2009).
Gill, S. et al. Preclinical targeting of human acute myeloid leukemia and myeloablation using chimeric antigen receptor-modified T cells. Blood 123, 2343–2354 (2014).
Song, Q. et al. Dissecting intratumoral myeloid cell plasticity by single cell RNA-seq. Cancer Med. 8, 3072–3085 (2019).
Laughney, A. M. et al. Regenerative lineages and immune-mediated pruning in lung cancer metastasis. Nat. Med. 26, 259–269 (2020).
Schulz, M. et al. Cellular and molecular changes of brain metastases-associated myeloid cells during disease progression and therapeutic response. iScience 23, 101178 (2020).
Zhang, Q. et al. The spatial transcriptomic landscape of non-small cell lung cancer brain metastasis. Nat. Commun. 13, 5983 (2022).
Song, Q. et al. Single-cell sequencing reveals the landscape of the human brain metastatic microenvironment. Commun. Biol. 6, 760 (2023).
Song, Q. QSong-github/BrMet: single cell sequencing reveals the landscape of the brain metastatic microenvironment (v1.0.0). GitHub https://github.com/QSong-github/BrMet (2023).
Acknowledgements
We thank J. Massagué for providing the H2030- and PC9-BrM cell lines. This work was supported by the National Institutes of Health grants RO1CA173499, R01CA185650 and R01CA205067, and by Department of Defense grant W81XWH2110075 (to K. Watabe). This study also utilized the Cancer Center Shared Resources, including Cancer Genomics, Tumor Tissue and Pathology, Cell Engineering, Flow Cytometry, and Biostatistics and Bioinformatics, which are supported by the Comprehensive Cancer Center of Wake Forest University NCI, National Institutes of Health Grant (P30CA012197). W.Z. was supported by the Hanes and Willis Family Professorship in Cancer. Additional support for Q.S. and W.Z. were provided by a Fellowship to W.Z. from the National Foundation for Cancer Research. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Cancer Institute. S.-Y.W. was supported by the T32 training grant (T32CA247819) and the NIH Pathway to Independence Award (K99CA276895).
Author information
Authors and Affiliations
Contributions
S.-Y.W. and K. Watabe designed the study and wrote the manuscript. S.-Y.W., S.S. and K. Wu conducted experiments and acquired, analysed and interpreted the data. S.S., A.T. and K. Wu performed animal experiment. S.-Y.W. and K. Wu performed intracardiac injections. J.-S.L., E.C.S., D.Z., R.P.D., J.W.K. and F.X. performed clinical sample collection and analysis. S.-Y.W., B.-S.P. and H.-K.L. designed and performed the CRISPER KO TNF in monocytes. S.-Y.W., A.P., F.X., M.C., M.H.S., J.R. and Y.L. performed single-cell sequencing. S.-Y.W., Q.S., J.C., W.Z., L.D.M., A.P. and M.C. analysed the results of single-cell sequencing. S.-Y.W., S.S., K. Wu, A.T., D.Z., R.P.D., E.C.S. and K. Watabe reviewed and edited manuscript and interpreted the data. S.-Y.W. and K. Watabe supervised the study.
Corresponding authors
Ethics declarations
Competing interests
H.-K.L. is a consultant for Stablix, Inc. and Chang Gung University of Science and Technology.
Peer review
Peer review information
Nature Biomedical Engineering thanks Saar Gill and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Macrophage infiltration and the expression of MSLN in lung cancer.
a, Analysis of epithelial and immune cell population in brain metastasis of lung cancer using the single cell sequencing data of URECA (http://ureca-singlecell.kr/). b, Population of different immune cells in brain tumors was analyzed using the PRECOG dataset30. Population analysis of different immune cells in brain tumors [Astrocytoma (n = 341), glioblastoma (n = 697), meningioma (n = 68), and oligodendrogliomas (n = 276)] was performed using the PRECOG dataset30. c, The expression levels of CD14+ monocyte/macrophage and Reg-TAMs markers (CD163, Arg1, Arg2) were investigated in the tumor adjacent normal brain (Non-tumor) and brain metastatic (BM) lesions of lung cancer patients using the TIMER2.0 dataset. Unpaired two-tailed t-test: *, p = 0.04. (n = 5 / group). d, Analysis of mRNA expression of MSLN in different brain regions and in distant organs using Human Protein Atlas data (https://www.proteinatlas.org/). e, Analysis of the expression of MSLN and other lung cancer tumor antigens in the brain metastatic samples using single-cell RNA sequencing. The data are presented as the mean ± SD.
Extended Data Fig. 2 Cytotoxic effect of CARMA on tumor cell growth.
a, Protein expressions of CD3 and CD28 in THP-1 human monocyte cell line and PBMC-derived primary human monocytes (Mono). b, The procedure of CARMA generation. The details of the procedure were provided in the Methods section. Image was created with BioRender.com. c, The expression of CD74+ Inflam-TAMs and CD206+ Reg-TAMs before or after CD14 microbeads isolation. (n = 5 / group). d, mRNA and protein expression of INF-γ, TNF-α and IL-1β on monocyte before or after CD14 microbeads isolation. (n = 5 / group). e, Efficiency of virus infection of Flag-labeled anti-MSLN-CAR constructs to the human primary (Pri) macrophage (MΦ) and N-CARMA and anti-MSLN-CARMA were examined by FACS. Significance was calculated using One-way ANOVA with Tukey’s multiple comparisons test: (e) ***, p = 0.0001 (n = 4 / group). f, The Flag protein expressions on the primary T, primary macrophage, Anti-MSLN-CAR-T, and anti-MSLN-CARMA (MyD-CARMA) cells were examined by western blot. g, MSLN binding ability in control N-CARMA and anti-MSLN-MyD-CARMA. h, Morphological changes of monocyte/macrophage that were treated with PBS (Veh), 1 nM CSF1, GM-CSF, or PMA. i, Cell viability of primary macrophage (Mϕ) and different CARMA cells after the treatment with PBS, CSF1 (1 nM), GM-CSF (1 nM) or PMA (1 nM) for 24 h. i, We calculated cell viability by dividing the absorbance of the wells containing cells treated with various cytokines by the average absorbance of the control wells. (n = 5 / group). j, Expression of Inflam-TAMs/Reg-TAMs phenotype on CARMA with PBS, CSF1, GM-CSF, or PMA treatment for 24 h. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 4 / group). k, The expression of MSLN in the tumor and CARMA. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 5 / group). l, The MyD-CAR construct in lentivirus was introduced into the PBMC-derived primary human monocyte by viral infection, and respective CAR-monocyte cells were generated. MyD-monocytes were treated with PBS, CSF1, PMA, or GM-CSF. After 24 h, the cells were washed with PBS and they were co-cultured with luciferase-labeled H2030BrM cells (E/T ratio: 30:1) followed by assaying cell viability by IVIS. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 3 / group). m, Real-Time Cytotoxicity Assay (RTCA) was used to evaluate cytotoxic function of CARMA. Target cells (H2030BrM) were first added to the culture wells, and cell index was recorded. After 25 h, effector cells (N-, CD3-, and MyD-CARMA) were added at a 30:1 (Effector: Target cells) ratio. The percentage of specific cell lysis was shown in the right panel. One-way ANOVA with Tukey’s multiple comparisons test: **, p = 0.001. (n = 4 / group). n, Primary N-, CD3-, or MyD-CARMA were cultured with luciferase-labeled H2030BrM cells (E/T ratio: 10:1) followed by assaying long-tern effect of cell viability by IVIS. Two-way ANOVA with Tukey’s multiple comparisons test: **, p = 0.0063 and ***, p = 0.0001. (n = 4 / group). The data are presented as the mean ± SD.
Extended Data Fig. 3 Effect of CARMA on brain metastasis.
a, Expressions of Inflam-TAMs/Reg-TAM markers were examined in isolated N-, CD3- or MyD-CARMA cells after co-culturing with PC9BrM (E/T ratio: 30:1). The value of qRT-PCR in each figure was normalized using actin as a control. The y-axis indicates arbitrary unit. Significance was calculated using One-way ANOVA with Tukey’s multiple comparisons test: CD74: *, p = 0.012; ***, p = 0.0001; CD80: *, p = 0.023; ***, p = 0.0001; CD86: ***, p = 0.0001; ***, p = 0.0001; HLA-B: *, p = 0.041; ***, p = 0.0001; HLA-C: ***, p = 0.0001; ***, p = 0.0001; CD163: ***, p = 0.0001; ***, p = 0.0001; CD204: ***, p = 0.0001; ***, p = 0.0001; CD206: ***, p = 0.0001; ***, p = 0.0001; RHEB: **, p = 0.007; APOE: **, p = 0.0013. (n = 5 / group). b, c, After 24 tumor co-culture of CARMA and PC9BrM cells, CARMA were isolated from the co-culture using CD14 microBeads. The macrophage polarization of CARMA cells was examined by FACS using antibody to CD74+ (Inflam-TAMs) and CD206+ (Reg-TAMs). One-way ANOVA with Tukey’s multiple comparisons test: (b) *, p = 0.03; ***, p = 0.0001; (c) *, p = 0.04; ***, p = 0.0001. (n = 5 / group). d, CARMA cells were co-cultured with PC9BrM for 24 h followed by examining the mRNA expression of Inflam-TAMs-related cytokines (INF-γ, TNF-α, and IL-1β). One-way ANOVA with Tukey’s multiple comparisons test: INF-γ: ***, p = 0.0001; TNF-α: *, p = 0.01; **, p = 0.001; ***, p = 0.0001; IL-1β: ***, p = 0.0001. (n = 5 / group). e, N-, CD3-, and MyD-CARMA cells were incubated with PC9BrM for 24 h and the protein level of TNF-α in conditioned medium was measured by ELISA. One-way ANOVA with Tukey’s multiple comparisons test: *, p = 0.03; ***, p = 0.0001. (n = 5 / group). f, CARMA cells were incubated with PKH26-labeled PC9BrM cells for 24 h, followed by measurement of the monocyte and macrophage phagocytic activity by FACS. One-way ANOVA with Tukey’s multiple comparisons test: *, p = 0.014; ***, p = 0.0001; *, p = 0.02; ***, p = 0.0001 (From left to right). (n = 5 / group). g-j, Scheme of the in vivo experiment (g). H2030BrM cells were transplanted intracranially into the nude mice. After 7 days of tumor transplantation, the mice received 1 ×105 human primary N(MyD)-CAR (N-CAR) monocyte (h), primary MyD-CAR monocyte (i), or primary CAR-T (j) that were all luciferase-labeled, by intracardiac injection, and the mice were scarified and checked for the luciferase intensity at the endpoint (Day 35). k, ex vivo BLI signals were examined for various organs at the endpoint after injection of luciferase-labeled CAR-T, N(MyD)-CARMA (N-CARMA), or MyD-CARMA. One-way ANOVA with Tukey’s multiple comparisons test: Brain: ***, p = 0.0001; ***, p = 0.0001; Liver: **, p = 0.002; **, p = 0.003; Spleen: ***, p = 0.0001; ***, p = 0.0001 (From left to right). (n = 3 / group). l, Population of microglia (Tmem119+/CD45−) and circulating macrophage (CD14+/CD45+) in brain metastatic region. t-test: ***, p = 0.001. (n = 4 / group). m, After one week of H2030BrM transplantation, mice received N-CAR-monocyte (N-CARMA), CD3-CAR-monocyte (CD3-CARMA), or MyD-CAR-monocyte (MyD-CARMA) by intracardiac injection. At the endpoint (Day 35), the brains were removed from the mice injected with or without CARMA. The cells were dissociated, and they were isolated using human CD14+ microbeads. They were then stained with human CD14 and Flag antibodies followed by analysis of FACS. One-way ANOVA with Tukey’s multiple comparisons test: *, p = 0.019; **, p = 0.0028; ***, p = 0.0001. (n = 5 / group). n, Human primary monocytes were transduced with the Flag-conjugated CAR construct. After 24 h, the CAR-transduced primary monocyte cells were washed with PBS and cultured in fresh regular medium for 3 days. The primary Flag+ CAR-monocytes (CARMA) were isolated by FACS. To test the efficacy of CARMA, Luciferase labeled-tumor cells (H2030BrM cells) were transplanted into nude mice by intracranial injection. After one week of tumor transplantation, the mice were treated with different concentrations of N-CARMA, CD3-CARMA or MyD-CARMA cells every two weeks by intracardiac injection. Quantitative data of in vivo brain tumor. Two-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 9 / group). o, Ex vivo BLI signals in the brain at the endpoint. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 8 / group). p, Percentage of brain metastases of the mice that received different doses of primary N-CARMA, CD3-CARMA, or MyD-CARMA. Chi Square test: ***, p = 0.0001. (n = 8 / group). q, Metastatic lesions in the brain were removed from the mice used in (n) and the protein expression of TNF-α in the brain of each group was measured by ELISA. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001 (n = 8 / group). The data are presented as the mean ± SD.
Extended Data Fig. 4 Infiltration of PBMC-derived CARMA into the brain metastasis region.
a, Experimental procedure to test anti-tumor effect of PBMC-derived CARMA in the humanized mouse model. Mice were engrafted with human PBMC cells. After human myeloid engraftment was confirmed, H2030BrM cells were transplanted into the brain by intracranial injection. After seven days, 1 × 105 PBMC-derived N-CARMA or MyD-CARMA were injected intracardially every 2 weeks until the endpoint. Brain tumors were harvested at the endpoint, and they were dissociated, and single cell RNA sequencing was performed. Scale bar: 100 µm. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 3 / group). b, Population of human CD45+ cells in humanized mice one month after receiving PBMC engraftment. c, The positive signal intensity of different circulating human immune cells in the blood of humanized mouse. d, The human CD45+/Flag+ cell were detected by FACS. Unpaired two-tailed t-test: ***, p = 0.0004. (n = 5 / group). e, Mice were engrafted with human PBMC cells. After the confirmation of human myeloid engraftment, H2030BrM cells were transplanted into the brain by intracranial injection. After seven days, 1 × 105 cells of PBMC-derived primary monocyte (Pri. MΦ), PBMC-derived N-CAR monocyte (N-CARMA), or PBMC-derived MyD-CAR monocyte (MyD-CARMA) were injected intracardially every 2 weeks until the endpoint. Upper panels: BLI images of representative mice from each experimental group at the endpoint. Lower panels: ex vivo image of brain metastasis. f, g, Quantitative data of in vivo (f) and ex vivo (g) brain tumor. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 6 / group). h, Percentage of brain metastasis in each group was measured. Chi Square test: ***, p = 0.00001. (n = 9 / group). i, j, Percentage of Reg-TAMs (i) and Inflam-TAMs (j) in the tumor was measured. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 9 / group). k, The expression of TNF-α in brain metastasis in the mice that were treated with primary monocyte, N-, or MyD-monocyte was measured. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 9 / group). l, Expression of Flag in the tumor brain of humanized mouse implanted with CARMA. The data are presented as the mean ± SD.
Extended Data Fig. 5 Effect of CAR on cell viability and polarization of CARMA and T cell trafficking.
a, b, Proliferation rate of Ki67+/cFMS+ cells (a) and cell viability (b) of the primary macrophage and different CARMA were examined using FACS and MTS. (n = 4 / group). c, Infiltration index of primary macrophage or different CARMA was measured using the boyden chamber. The CARMA were seeded on the upper chamber and H2030BrM cells were seeded on the lower chamber (E/T ratio: 30:1). After 72 h of incubation, the macrophages in the lower chamber were examined by FACS. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 5 / group). d, Expression of lysosome capacity in primary macrophage and CARMA. e, Polarization phenotype in N- and MyD-CARMA before or after CAR insertion. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 4 / group). f, Expressions of TNF-α, IL-1β, and INF-γ in primary macrophage, N-CARMA and MyD-CARMA cells that were co-cultured with H2030BrM were quantified by ELISA. One-way ANOVA with Tukey’s multiple comparisons test: **, p = 0.002; *, p = 0.018; *, p = 0.013. (n = 6 / group). g-i, Expressions of TNF-α, IL-1β, and INF-γ in MyD-CARMA cells before or after CAR insertion and CARMA-H2030BrM co-culture were quantified by ELISA. One-way ANOVA with Tukey’s multiple comparisons test: *, p = 0.03; ***, p = 0.001 (n = 5 / group). j, k, H2030BrM cells were co-cultured with PBMC-derived CARMA (E/T ratio: 30:1) for 72 h. The primary CARMA cells were then isolated from the co-culture and the expressions of CXCL9 (j) and CXCL10 (k) of the isolated CARMA cells were examined by qRT-PCR. One-way ANOVA with Tukey’s multiple comparisons test: (j) *, p = 0.03; **, p = 0.0012; ***, p = 0.0001; (k) ***, p = 0.0001 (n = 5 / group). l, Primary T cells were co-cultured with H2030BrM-pretreated primary CARMA (E/T ratio: 30:1) for 24 h. The T cells were then isolated from the co-culture, and the expression of CXCR3 of the T cells was examined by qRT-PCR. One-way ANOVA with Tukey’s multiple comparisons test: *, p = 0.03; ***, p = 0.0001. (n = 5 / group). m, H2030BrM cells were co-cultured with primary CARMA (E/T ratio: 30:1) for 72 h. CARMA cells were then separated from the co-culture and incubated with primary T cells that were labeled with cell proliferation dye. The T cell proliferation rate was then examined by FACS. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 5 / group). n, H2030BrM cells were co-cultured with primary CARMA (E/T ratio: 30:1) for 24 h. CARMA cells were then separated from the co-culture and seeded in the lower chamber, and the primary T cells were seeded in the upper chamber. After 3 days of co-culture, the primary T cells were isolated from the lower chamber and examined for the chemotactic index. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 5 / group). o, The luciferins-labeled H2030BrM cells were co-cultured with ‘IL-4-activated primary T cell only (Tumor + pri T)’ or ‘CARMA plus IL-4-activated primary T cells (Tumor + CARMA + pri. T)’ for 0, 2, 5 days, and the tumor cell growth was examined by IVIS. One-way ANOVA with Tukey’s multiple comparisons test: Day 2: **, p = 0.015; **, p = 0.014; Day 5: *, p = 0.02; **, p = 0.009; ***, p = 0.0001. (n = 5 / group). The data are presented as the mean ± SD.
Extended Data Fig. 6 Bystander effect of CARMA.
a, N-CARMA or MyD-CARMA cells were cultured with H2030BrM (E/T ratio: 30:1) for 24 h. CM was collected and analyzed by using the cytokine/growth factor array (Raybio). b, Expressions of Inflam-TAMs-related cytokines in CM were examined by ELISA after the co-culture of primary CARMA and H2030BrM cells. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0002; ***, p = 0.00023; **, p = 0.0021; *, p = 0.03; **, p = 0.0001. (n = 3). c and d, The expression of TNF-α, IL-1β, or INF-γ receptor on tumor population in human brain metastatic sample (c) and humanized brain metastatic mouse sample (d). e, mRNA expression of TNF-α, IL-1β or INF-γ receptor on H2030BrM cell. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.001. (n = 5). f, Cell viability of H2030BrM after one week of recombinant TNF-α, IL-1β or INF-γ treatment. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.001. (n = 5). g, The CM derived from the co-culture of MyD-CARMA and H2030BrM cells was treated with anti-TNF-α, anti-IL-1β, or anti-INF-γ antibody to deplete each cytokine in CM. H2030BrM cells were then treated with the cytokine-depleted CM followed by assaying the colony-forming ability. One-way ANOVA with Tukey’s multiple comparisons test: **, p = 0.001; ***, p = 0.0001. (n = 4). h, Scheme of experimental design. NCI-H522MSLN+/GFP+ and NCI-H522MSLN-/RFP+ cells were mixed and intracranially transplanted into mice. After 7 days, the mice received 1 ×105 PBMC-derived primary N-, CD3-, or MyD-CAR monocyte by intracardiac injection every 2 weeks for 35 days. i, Quantitative data of in vivo brain metastasis of lung cancer. Two-way ANOVA with Tukey’s multiple comparisons test: ##, p = 0.01; ###, p = 0.001; **, p = 0.01. (n = 6-8 / group). j, Ex vivo BLI signals in the brain at the endpoint. One-way ANOVA with Tukey’s multiple comparisons test: *, p = 0.05; ***, p = 0.0001. (n = 9 / group). k, Kaplan–Meier analysis of brain tumor free survival was performed. Two-way ANOVA with Tukey’s multiple comparisons test: *, p = 0.05; ***, p = 0.001. (n = 7-9 / group). l, The metastatic lesions in the mouse brain in Extended Data Fig. 6h were sectioned and subjected to immunohistochemical analysis for GFP (MSLN+) and RFP (MSLN−). Scale bar: 100 μm. m-o, Metastatic brain tumors in Extended Data Fig. 6h were isolated from the brain and examined by FACS for the population of CD45+ macrophage (m), Reg-TAMs (n), and Inflam-TAMs (o). One-way ANOVA with Tukey’s multiple comparisons test: (m) *, p = 0.04; ***, p = 0.0001; (n) **, p = 0.01; ***, p = 0.0001; (o) ***, p = 0.0001. (n = 6 / group). p, Metastatic tumors in the brain were removed from the mice in Extended Data Fig. 6h. The expression of TNF-α in the dissociated mouse brain from the primary N-, CD3-, or MyD-CARMA treated groups were measured by ELISA. One-way ANOVA with Tukey’s multiple comparisons test: *, p = 0.012; ***, p = 0.0001. (n = 6 / group). The data are presented as the mean ± SD.
Extended Data Fig. 7 Effect of knockdown of TNF-α on the growth of NCI-H522MSLN+/GFP+ and NCI-H522MSLN-/RFP+ cells.
a, Efficiency of the knockout (KO) of TNF-α in CARMA cells. CRISPR/Cas9 KO plasmids consist of scrambled or 3 sets of TNF-α-specific guide RNA sequences and GFP reporter. The plasmid was transduced to CAR-THP-1 human monocyte cells. The scrambled GFP+ and KO TNF-α GFP+ cell were isolated by FACS. b, The transfection efficiency of isolated CAR-THP-1 in Extended Data Fig. 7a was examined by measuring GFP-positive signal (Right panel). Scale bar: 100 µm. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 4 / group). c, The expression of TNF-α was measured by qRT-PCR. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 5 / group). d, Scheme of testing the effect of KO of TNF-α on bystander killing effect. MSLN+ cells were co-cultured with scramble or KO TNF-α CARMA (E/T ratio: 30:1) for 72 h, and CARMA CM was collected. MSLN-negative cells were then treated with the CM. e-g, The MSLN-negative cells (NCI-H522) were treated with the CM that were collected from Scrambled or KO TNF-α CARMA cells that were co-cultured with MSLN-positive tumor cells (NCI-H522MSLN+), followed by assaying the cell viability (e), colony formation ability (f), and cancer stem cell population (g) of the MSLN-negative cells. One-way ANOVA with Tukey’s multiple comparisons test: (e) ***, p = 0.0001; (f) *, p = 0.02; ***, p = 0.0001; (g) *, p = 0.03; ***, p = 0.0001. (n = 5 / group). h, NCI-H522MSLN+ cells were labeled with GFP while NCI-H522MSLN− cells were labeled with RFP. Tumor cells were then co-cultured with primary MyD-CARMA with or without knockdown of TNF-α at a 30:1 effector/target cells ratio for 72 h. Growth of the NCI-H522MSLN+/GFP+ and NCI-H522MSLN-/RFP+ cells was observed by microscopy, and representative images were shown. Scale bar: 100 µm. i-k, Percentage of NCI-H522MSLN+/GFP+ and NCI-H522MSLN-/RFP+ cells were examined by flow cytometry. One-way ANOVA with Tukey’s multiple comparisons test: (i) ***, p = 0.0001. (n = 5 / group); (j) ***, p = 0.0001. (n = 6 / group). The data are presented as the mean ± SD.
Extended Data Fig. 8 Expression of TNF-α and bystander effect of CARMA and CAR-T.
a, Kaplan–Meier analysis was performed for overall survival of lung cancer patients in relation to CD47 (left panel) and SIRPα (right panel) expression using the TCGA dataset. b, Expression of CD47 on primary and metastatic brain tumor from lung adenocarcinoma patients was examined using the GSE123902 dataset89. Unpaired two-tailed t-test: ***, p = 0.0001. [Primary (n = 40); BM (n = 19)]. c, Expression of SIRPα on primary N-, CD3-, and MyD-CARMA was examined by qRT-PCR. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0005; ***, p = 0.0001. (n = 3 / group). d, Expression of SIRPα on CD45+ macrophages was examined in the regions of brain metastasis and adjacent normal areas of lung cancer patients. Scale bar: 200 μm. e, Luciferase-labeled H2030BrMMSLN+ cells [Target cells (T)] were co-cultured with the different ratios of CAR-T or MyD-CARMA [Effector cells (E)] for one or three days and the cell viability was measured by IVIS bioluminescence imaging. One-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 3 / group). f, g, Expressions of TNF-α gene and TNF-α protein were examined by qRT-PCR (f) and ELISA (g) after the co-culture of primary T cells, primary macrophage (Mϕ), CAR-T, and MyD-CARMA with or without H2030BrM cells (E/T ratio: 30:1) for 24 h. One-way ANOVA with Tukey’s multiple comparisons test: **, p = 0.01; ***, p = 0.001. (n = 5 / group). h-j, Scheme of testing the effect of CAR-T and CARMA on bystander killing effect (h). NCI-H522MSLN− cells were labeled with RFP (NCI-H522MSLN-/RFP+). NCI-H522MSLN− cells that have ectopic overexpression of MSLN and were labeled with GFP and designated as NCI-H522MSLN+/GFP+. The NCI-H522MSLN+/GFP+ and NCI-H522MSLN-/RFP+ cells were then co-cultured (1:1 ratio). The primary T cells, primary macrophage, CAR-T, and MyD-CARMA cells were incubated with the mixed-culture of NCI-H522MSLN+/GFP+ and NCI-H522MSLN-/RFP+ for 72 h, and the percentage of surviving NCI-H522MSLN+/GFP+ (i) and NCI-H522MSLN-/RFP+ (j) was analyzed by FACS. One-way ANOVA with Tukey’s multiple comparisons test: *, p = 0.04; **, p = 0.01; ***, p = 0.0001. (n = 4 / group). The data are presented as the mean ± SD.
Extended Data Fig. 9 Brain MRI demonstrates minimum neurotoxicity of CARMA.
a, T2-weighted coronal images of a mouse brain at multiple planes. Diagram showing the planes of imaging in a mouse brain, starting with the striatum and continuing through the substantia nigra. T2 analysis for measuring volume and microstructure of brain. ROIs indicate the various brain regions for quantitative analysis. Yellow circle: tumor location. Red areas: hemorrhage region. White arrow: abnormal space. The table shows brain volume in different regions with or without tumor transplantation or CARMA supplement. (n = 5 / group). b, Color images were generated from post-processed imaging data from (a). They provide a visual representation of the quantitative. c, Diffusion Tensor Imaging (DTI) for analysis of axonal organization in the brain after CARMA injection. Red: transvers fibers. Green: anteroposterior fibers. Blue: Craniocaudal fibers. (n = 5 / group). **, p = 0.01: compared to non-tumor transplantation group. d-g, CAR-T and CARMA cells were generated using the second-generation CAR construct. Each H23030BrM transplanted animal in primary CAR-T, primary N-CARMA, and primary MyD-CARMA group was subjected to the object recognition test, and the percentages of investigation time of moved (d) and novel (e) object were measured. f,g, Body weight (f) and liver function (g) of the mice were measured at the end points in each group. Unpaired two-tailed t-test: (d) **, p = 0.006; **, p = 0.003; **, p = 0.002. (n = 6 / group); (g) *, p = 0.03; *, p = 0.03 (From left to right). (n = 4 / group). h, Expression of MSLN in brain metastatic tumor region and tumor-adjacent bronchi tissues. (n = 10 / group). Scale bar: 200 µm. i, Morphology of lung tissue in N-, CD3- or MyD-CARMA injected mice. Scale bar: 200 µm. The data are presented as the mean ± SD.
Extended Data Fig. 10 Effect of mouse CARMA on brain metastasis in male and female wild-type mouse models.
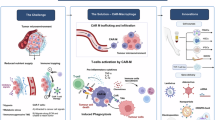
a, Mouse primary monocytes were transduced with the Flag-conjugated CAR construct. After 24 h, the CAR-transduced primary monocyte cells were washed with PBS and cultured in fresh regular medium for 3 days. To test the efficacy of CARMA, Luciferase labeled-mouse lung cancer tumor cells (LL2-BrM cells) were transplanted into male C57BL/6 mice by intracranial injection. After one week of tumor transplantation, the mice were treated with N-CARMA or MyD-CARMA cells every two weeks by intracardiac injection. Image was created with BioRbender.com. b, Quantitative data of in vivo male brain tumor. Two-way ANOVA with Tukey’s multiple comparisons test: ***, p = 0.0001. (n = 6 / group). c-f Each LL2-BrM transplanted animal in N-CARMA and MyD-CARMA group was subjected to the object recognition test, and the percentages of investigation time of moved (c) and novel (d) object were measured. (e) Liver function and (f) body weight of the mice were measured at the end points in each group. ***, p = 0.0001. (n = 6 / group). g, Luciferase labeled-mouse lung cancer tumor cells (LL2-BrM cells) were transplanted into female C57BL/6 mice by intracranial injection. After one week of tumor transplantation, the mice were treated with N-CARMA or MyD-CARMA cells every two weeks by intracardiac injection. ***, p = 0.0001. (n = 6 / group). h-k Each LL2-BrM transplanted female mice in N-CARMA and MyD-CARMA groups were subjected to the object recognition test, and the percentages of investigation time of moved (h) and novel (i) object were measured. Body weight (j) and liver function (k) of the mice were measured at the end points in each group. ***, p = 0.0001. (n = 6 / group). The data are presented as the mean ± SD. l, Morphology of lung tissue in mouse N- and MyD-CARMA injected wild-type mice. Scale bar: 100 µm. m, Genetically engineered macrophages (CARMA) that express tumor-specific antibody fused with MyD88 drastically decreased brain metastasis growth by showing antigen-specific phagocytosis activity against tumor cells. CARMA also showed bystander killing effect on antigen-negative-tumor cells by secreting cytokine TNF-α. Image was created with BioRender.com.
Supplementary information
Supplementary Information (download PDF )
Supplementary methods.
Source data
Source Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Fig. 7 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 7 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 8 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 9 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 10 (download XLSX )
Statistical source data.
Source Data Fig. 2 (download PDF )
Unprocessed western blots.
Source Data Extended Data Fig. 2 (download PDF )
Unprocessed western blots.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Wu, SY., Tyagi, A., Wu, K. et al. MyD88-mediated chimaeric antigen receptor macrophages suppress brain metastasis using target-specific phagocytosis. Nat. Biomed. Eng (2026). https://doi.org/10.1038/s41551-026-01613-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41551-026-01613-x
