
Abstract
Understanding how species are exposed to different types of extreme events is an emerging priority to inform biodiversity conservation under climate change. Using climate impact projections and species range data, we predict changes in exposure to droughts, heatwaves, river floods and wildfires for 33,936 terrestrial vertebrate species and 794 ecoregions. By 2050, under a medium–high emission scenario (SSP3–7.0), on average 74% of the area within species’ current geographic ranges are projected to be exposed to heatwaves, 16% to wildfires, 8% to droughts and 3% to river floods. These trends include species-rich areas in the Amazon basin, Africa and Southeast Asia. By 2050, 22 ecoregions, primarily in mid-latitudes, are estimated to have at least 50% of their area exposed to two or more types of extreme events, increasing to 236 ecoregions by 2085 (SSP3–7.0). By 2085, 36% of the area within species’ ranges are projected to be exposed to multiple event types (SSP3–7.0). These findings highlight the need for further research into species’ sensitivity and adaptive capacity to extreme events, and for conservation strategies that address the impacts of multiple extreme events.
Similar content being viewed by others


Main
Extreme climate events, such as heatwaves and wildfires, can have devastating impacts on terrestrial biodiversity. For example, the 2019/2020 heatwave in Australia killed more than 72,000 flying foxes1. In that same year, wildfires in the Pantanal killed an estimated 17 million vertebrates2. A review of 519 studies found that 57% of studies documented negative responses of species to extreme events, including 100 cases with a population decline of more than 25% and 31 records of local extirpations3.
Climate change has led to an increase in extreme climate events—a trend that is projected to intensify4. Vulnerability to climate change depends on three conditions: the degree to which species are exposed, their sensitivity to change and their ability to adapt5. One key reason extreme events are rarely integrated into conservation planning and species risk assessments is our limited understanding of the types of events to which species will be exposed most strongly across different geographic regions6. As a first step, we focus here on exposure, examining how species’ exposure to different types of extreme events and to multiple hazards is projected to change.
Most studies on species’ exposure to climate extremes have focused on events that can be derived directly from the output of global climate models, especially temperature extremes7,8 (but see ref. 9). However, published studies demonstrate that species’ sensitivity varies by event type, with different extremes affecting body condition, reproductive success and survival through distinct mechanisms (Table 1). In addition, the impact can be amplified when extreme events are compounded spatially and/or temporally10. Biodiversity research rarely incorporates output from climate impact models, that is, process-based models that translate climate data into climate impacts such as wildfires or floods (Methods). Yet, estimates of burned area from impact models, for example, are more relevant to species risks than fire weather indices derived from climate models alone. Investigating exposure to various types of extreme events can thus help identify which changes are most relevant for terrestrial biodiversity, and inform effective conservation planning.
We analysed projected changes in extreme event exposure globally for terrestrial vertebrates, that is, amphibians, birds, mammals and reptiles, using a dataset of four extreme climate events (drought, heatwave, river flood and wildfire; for event definition see Methods). We used climate impact simulations from the Inter-Sectoral Impact Model Intercomparison Project (ISIMIP Phase 3b11), which provides output from six climate impact models (Supplementary Table 1) forced by climate projections from five climate models of the Coupled Model Intercomparison Project (CMIP6) under three emission scenarios: low (SSP1–2.6), medium–high (SSP3–7.0) and high (SSP5–8.5). We defined extreme events relative to their historical distribution within each grid cell (at a resolution of 0.5°), thereby accounting for natural spatial variations. We analysed changes in extreme event exposure between a baseline period centred on 2000 (1985–2014) and future periods, using 30-year moving windows centred between 2030 and 2085. For each grid cell, we calculated the event frequency as the proportion of years with an event within the 30-year window. We also identified grid cells with high frequencies of multiple extreme events (≥0.33 year−1, corresponding to events every third year) and summed the number of different event types per cell.
To identify hotspots of terrestrial biodiversity exposure, we used species richness (all species (Fig. 1 and Extended Data Fig. 1d–f) and threatened species (Supplementary Fig. 1)) and rarity-weighted richness (Supplementary Fig. 2) data from the International Union for Conservation of Nature (IUCN) Red List of Threatened Species12. To quantify exposure for individual species, we used species range layers from the IUCN Red List of Threatened Species12, the Global Assessment of Reptile Distributions13 and BirdLife International14. Due to the spatial resolution, extreme event data are not available for small islands. Species restricted to small islands not represented in the ISIMIP grid were excluded resulting in a final dataset of 33,936 terrestrial vertebrate species (7,605 amphibians; 10,562 birds; 5,476 mammals and 10,293 reptiles). For each year, we quantified species exposure as the proportion of grid cells where extreme events occur, weighted by event frequency, grid cell area and the proportion of overlap between each cell and the species’ range. Using the same approach we also quantified exposure for 794 ecoregions15.

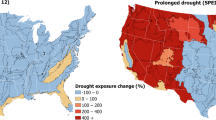
a–d, change in annual frequency of extreme events from 2000 to 2050 for SSP3–7.0 for heatwave (a), wildfire (b), drought (c) and river flood (d) for all four taxa combined based on species richness data from the IUCN Red List of Threatened Species. Icons from the Noun Project. Basemaps reproduced from ISIMIP Repository under a CC0 1.0 Universal Public Domain license; Lange, S., Büchner, M. https://doi.org/10.48364/ISIMIP.822294.
Results
Exposure to single event types
We found the strongest increase in extreme event exposure for heatwaves (Fig. 1). Under SSP1–2.6, aligned with the Paris Agreement of staying below 2° warming, on average 63% (model ensemble minimum to maximum: 45–81%; Methods) of the area within species’ ranges are projected to be exposed to heatwaves by 2050 (Supplementary Table 2). This corresponds to an increase by 45% (31–64%) compared with 2000 levels (Fig. 2a). Under SSP3–7.0, matching current developments of regional rivalry and high emissions more closely, 74% (59–90%) of the area within species’ ranges are projected to be exposed, corresponding to an increase by 56% (45–73%, Supplementary Table 2). By 2050, 9,434 (6,864–10,296) bird, 4,729 (3,302–5,299) mammal, 6,849 (5,089–7,384) amphibian and 9,155 (6,676–9,886) reptile species are projected to have at least 50% of their geographic range exposed to heatwaves (SSP3–7.0, Supplementary Tables 3–6). By 2085, 93% (85–99%) of the area within species’ ranges will be exposed to heatwaves for SSP3–7.0. Strong increases in event frequency are not only projected for species-rich areas (that is, Amazon basin, tropical Africa and Southeast Asia, Fig. 1a), but exposure increases by 50% (41–71%) across all ecoregions (SSP3–7.0; Extended Data Fig. 2a and Supplementary Table 7). Southeast Asia further emerges as a hotspot for threatened species exposure (Supplementary Fig. 1).

a–d, Change in proportion of geographic range exposed relative to year 2000 averaged for each taxon for three scenarios for heatwave (a), wildfire (b), drought (c) and river flood (d). Vertical lines show the range of model ensemble across all four taxa for the year 2085. Values for each taxon and each climate model–impact model combination are in Supplementary Figs. 13–16. Icons from the Noun Project.
Extreme wildfires are projected to be the second most prevalent event. By 2050, 16% (11–22%) of the area within species’ ranges are projected to be exposed for SSP3–7.0, reaching 25% (17–35%) by 2085 (Supplementary Table 2). Hotspots for increased wildfire frequency in species-rich areas are the Amazon basin, southern Africa and Southeast Asia (Fig. 1b). Mid-latitude ecoregions will be increasingly exposed, with 130 ecoregions projected to have at least 25% of the area exposed to wildfires by 2050 (SSP3–7.0; Extended Data Fig. 2).
Drought exposure, as defined in this study, remains relatively low: 8% (3–15%) of the area within species’ ranges is projected to be exposed in 2050 (SSP3–7.0), increasing to 14% (5–29%) by 2085 (Fig. 1c and Supplementary Table 2). Amphibian ranges will be exposed more strongly compared with other taxa, especially by 2085 and for SSP3–7.0 and SSP5–8.5 (Fig. 2c and Supplementary Fig. 3). Strong increases in flood exposure are projected only for localized areas in Central Africa, taiga and tundra, especially by 2085, and for SSP5–8.5, affecting small fractions of species’ ranges (Fig. 2d). By 2050, 3% (2–5%) of the area within species’ ranges is projected to be exposed to river floods, increasing to 5% (2–10%) by 2085 (SSP3–7.0; Supplementary Table 2).
Exposure to multiple event types
By 2050, 14% (10–18%) of the area within species’ ranges are projected to be exposed to at least two types of extreme events, increasing to 36% (26–45%) by 2085 (SSP3–7.0; Supplementary Table 2, breakdown by event type combination in Supplementary Table 8 and Supplementary Fig. 4), suggesting that terrestrial vertebrates could be exposed to different types of extreme events in the same year or in consecutive years (Fig. 3 and Extended Data Fig. 1). However, for 22 ecoregions, more than 50% of the area is exposed to at least two types of events already by 2050 (SSP3–7.0; Supplementary Table 7), with these ecoregions visible in mid-latitude regions in Fig. 3a. By 2085, this increases to 236 ecoregions (Fig. 3b). For SSP1–2.6, 9% (5–12%) of the area within species’ ranges is projected to be exposed to multiple events by 2085, but this increases to 44% (36–54%) under SSP5–8.5 (Supplementary Table 2).

a,b, Proportion of ecoregion projected to be exposed to at least two types of extreme events for SSP3–7.0 for the years 2050 (a) and 2085 (b). c, Proportion of geographic range exposed to at least two different types of extreme events for SSP3–7.0 for the year 2085. d, Proportion of range exposed against area of species range for different numbers of extreme event type for each species. Sensitivity of results for applying different thresholds to define multiple events are shown in Supplementary Figs. 10 and 11. Basemaps in a and b from the ISIMIP Repository under a CC0 1.0 license (https://doi.org/10.48364/ISIMIP.822294).
Discussion
Three key considerations are necessary to contextualize our findings. First, although extreme events can impact biodiversity negatively, they can also benefit certain species10 (Table 1). For example, the ornate chorus frog (Pseudacris ornate) experiences lower predation pressure during droughts16. Second, some species and ecosystems are adapted to, or even rely on, these disturbances. For example, the riffian skink (Chalcides colosii) was found only in forested areas in early postfire years, because it depends on open habitat17. As a result, regions with regular flooding or moderate-intensity wildfires often exhibit greater biodiversity17. Third, species exhibit behavioural and physiological plasticity to adapt to changing conditions. For example, gorillas (Gorilla beringei) have been shown to drink more frequently with increasing maximum daily temperatures18. However, adaptive behaviours may involve trade-offs. For example, although high temperatures can reduce risk of fungal infections in amphibians because these fungi grow more slowly, they do not actively seek out hotter areas, probably to avoid desiccation19.
However, climate change is intensifying extreme events beyond what many species are likely to be able to adapt to, especially within a short timeframe20. Species with restricted ranges face particularly severe risks21, as demonstrated by the Carnaby’s Black Cockatoo (Calyptorhynchus latirostris), which experienced a 60% population decline following the 2011 Western Australian heatwave22. Meta-analyses indicate native species typically show higher vulnerability to these events than non-native species23. Extreme events can also cause abrupt, widespread impacts across taxonomic groups simultaneously—the 2011 Australian heatwave not only led to a crash of the Black Cockatoo population, but also caused high shrub and tree mortality, altering the habitat quality for many species22. Thus, when extreme events exacerbate other stressors like habitat loss and disease, they create synergistic threats to biodiversity24. This compounding effect was shown in a study on the 2019–2020 Australian fire showing 27–40% greater declines across plant and animal species when fire was preceded by drought, highlighting the danger of multi-hazard extreme events25.
Although our analysis faces limitations from uncertainties in scenarios, data and modelling, and the coarse spatial and temporal resolution, it is a first step to integrate biodiversity data with harmonized multi-hazard data. The absence of small islands in our analysis may lead to an underestimation of exposure for island-endemic species, which are particularly vulnerable given their limited range size and dispersal options. We use current species distributions with known constraints26, without accounting for species range shifts in response to habitat destruction or climate change. Evidence shows that range shifts often follow complex, inconsistent patterns influenced by factors beyond climate alone27,28, making reliable predictions challenging across all terrestrial vertebrates. Our projections show substantial increases in extreme event exposure within relatively short timeframes (by 2050), during which many species will probably shift their ranges only partially. This temporal mismatch between rapid climate change and slower biological adaptation underscores the urgency of conservation interventions.
We quantify exposure to locally extreme conditions. Whether this exposure translates into impacts depends on population-level sensitivity and adaptive capacity5. Although there are studies that use a species-specific threshold to define an extreme event and thereby already include species sensitivity in their threshold definition, this implies a focus on a single mechanism of sensitivity (for example, exceedance of thermal tolerance limits29,30). Moreover, using a fixed sensitivity threshold implies a binary classification of impact (that is, within or outside thermal tolerance), whereas extreme event impact can be gradual (for example, by changing foraging activity or altering reproductive success well within physiological tolerance limits). Furthermore, thermal tolerance can vary substantially within species across their geographic range due to local adaptation, making it difficult to define a single biologically meaningful threshold applicable across an entire species’ distribution31,32. Our grid-based approach focuses on exposure only, but does not assume a single mechanism of sensitivity and instead captures exposure to conditions that can impact species through multiple pathways, such as body condition, behavioural changes or resource availability (Table 1) and allows for a consistent assessment across different event types. Therefore, our estimates are not directly comparable with studies that define extremes using species-specific physiological thresholds (for example, ref. 29).
Because studies differ in methodology (for example, extreme event definition, input data, spatial scale, species subsets), there are differences in projected exposure magnitude and spatial patterns. For example, our results show lower levels of projected drought exposure than other studies8, due to differences in species subsets and whether droughts were defined using climate data (for example, precipitation) or climate impact data (for example, soil moisture). Yet, consistent with previous work, we find that amphibians are likely to experience stronger drought exposure than other taxonomic groups7,33. Studies consistently show that species are likely to face increasing exposure to extreme events.
Our spatially explicit projections provide a baseline of exposure to extreme events across terrestrial vertebrates and highlight the need for finer-scale research on population-level sensitivity and adaptive capacity34, because knowledge gaps persist even for well-studied groups like great apes35. Research agendas on species’ vulnerability to increasing extreme events, as well as the subsequent decline in ecosystem services36, need to be developed collaboratively between scientists and conservation practitioners, integrating research with applied conservation practice and feasible management interventions. At the same time this study underlines that a low-emission scenario reduces the number of species facing significant changes in extreme event exposure, emphasizing the need for ambitious climate change mitigation.
Methods
Extreme event data
The extreme event dataset was generated following the methodology by ref. 37 and is described in ref. 38 (analytical workflow in Supplementary Fig. 5). It is based on climate and climate impact projections from ISIMIP Phase 3b11.
Climate models (also called general circulation models) simulate the physical climate system. They use the laws of physics to estimate how temperature, precipitation, wind patterns, ocean circulation and other climate variables change over time under different greenhouse gas scenarios. The CMIP provides a standardized framework for running and comparing simulations from several climate models under consistent scenarios and protocols. Results from climate models form the backbone of the IPCC Working Group I report, that is, the physical science of climate change.
Climate impact models take the output of climate models as inputs and simulate how these climate variables affect natural or human systems through dynamic, process-based representations of physical, biological, geological and chemical processes in combination with socio-economic boundary conditions (for example, land use). For example, hydrological models simulate the terrestrial water cycle and translate changes in temperature and precipitation into changes in soil moisture or river discharge, by simulating processes such as water storage, evapotranspiration and runoff39. Vegetation models simulate vegetation composition and distribution by representing processes such as photosynthesis, plant growth, fire disturbance and vegetation structure. ISIMIP provides a harmonized framework for running and comparing climate impact models across different sectors under consistent simulation protocols, forcing data and scenarios, similar to how CMIP coordinates climate models. Climate impact model results form an important basis for IPCC Working Group II assessments on climate change impacts and adaptation.
Details on the hydrological and vegetation models used in this study and references for the model description papers are in Supplementary Table 1. Impact models are forced by daily bias-adjusted climate data from five CMIP6 climate models: GFDL-ESM4, UKESM1-0-LL, IPSL-CM6A-LR, MPI-ESM1-2-HR and MRI-ESM2-0. The climate models were selected for the ISIMIP3b simulation round based on how the models performed in the historical period, their structural independence and the presentation of processes, and representing the range of equilibrium climate sensitivity of the CMIP6 model ensemble11. Simulations were run for a historical (1850–2014) and a future (2015–2100) period. Future simulations are based on three emission scenarios: low (SSP1–2.6), medium–high (SSP3–7.0) and high (SSP5–8.5). In addition, all models ran baseline simulations for stable pre-industrial climate conditions for 1850–2100, which we refer to here as the ‘pre-industrial control simulation’. These long simulations allow for a more robust identification of extreme events return periods as they include natural climate variability11,40. Spatial resolution is 0.5° latitude × 0.5° longitude (for consistency among different impact categories).
Exposure to extreme events was calculated as the area exposed annually37. This aggregation to an annual time scale allowed for an intercomparison of different extreme event types, while acknowledging that extreme climate impacts typically occur on smaller time scales38.
Various approaches exist for defining extreme events (for example, droughts41 and heatwaves42). We defined extreme events relative to the historical distribution within each grid cell. This approach quantifies conditions that deviate substantially from local historical patterns. A grid cell is classified as experiencing extreme conditions when patterns exceed the 97.5th percentile (2.5th for droughts) of the pre-industrial control simulation (1850–2100) for that cell following the approach established by ref. 37. Percentile-based definitions are standard in climate extremes research, with thresholds typically ranging from 90th to 99th percentile4. Our choice is conservative within this range and maintains consistency with previous assessments37. Recent biodiversity studies have applied similar grid-based percentile thresholds to quantify species exposure to extreme events8,33,43.
Drought exposure was derived from soil moisture simulated by three global hydrological models (H08, JULES-W2 and WaterGAP2-2e; Supplementary Table 1), that is, going beyond meteorological droughts41. The hydrological models simulate soil moisture at root level, at ~1-m depth, at daily temporal resolution, which is then aggregated to average monthly soil moisture. Unlike extreme events such as heatwaves and floods, which evolve within days and can be captured at daily resolution, soil moisture droughts develop over longer periods (months and years) and are therefore analysed at monthly time scale44. Soil moisture responds slowly to meteorological changes due to physical buffering of water in the soil, making monthly aggregation appropriate for drought assessment45. A grid cell is classified as drought-affected in a given year if soil moisture falls below the 2.5th percentile of the pre-industrial control simulation for three or more consecutive months37. A soil moisture mask excludes dry grid cells where annual mean discharge is below 0.1 mm per day.
Heatwaves are calculated from daily near-surface air temperature from climate models using a two-step approach38. First, we identify heatwave periods using the HeatWave Magnitude Index daily (HWMId)46. The HWMId quantifies the intensity of all heatwave periods occurring in a year. A heatwave period is defined as at least three consecutive days for which daily maximum temperature is higher than the 90th percentile of daily maximum temperatures under pre-industrial climate conditions, centred on a 31-day window37. The index produces a single value that increases with the severity and persistence of the heatwave. Because thresholds are calculated separately for each grid cell using a 31-day moving window centred on each day of the year, the method accounts for both spatial and seasonal temperature variation. Short heatwaves (3–4 days) split across the December–January boundary may be missed, but this affects pre-industrial, historical and future periods equally and would lead to underestimation rather than overestimation of exposure. Second, to identify extreme heatwaves for this study, we classified a grid cell as exposed to an extreme heatwave if the HWMId exceeded the 97.5th percentile of the pre-industrial control simulation for that grid cell.
River flood is based on daily runoff simulated by three global hydrological models (H08, JULES-W2 and WaterGAP2-2e; Supplementary Table 1). The river routing model CaMa-Flood47 redistributes runoff along prescribed river networks to determine outflow (detailed in ref. 48). A generalized extreme value distribution was fitted to the annual maxima time series of daily outflow of the pre-industrial control simulation. For each year, a grid cell is classified as flooded if the return period of maximum annual outflow exceeds 40 years, which corresponds to the 97.5th percentile. This is a binary classification. Based on outflow CaMa-Flood determines flood depth and flooded area. To exclude very dry areas, grid cells for which less than 1% of area is flooded are excluded and the soil moisture mask from above was applied. Note that flooded area is upscaled from 0.25° × 0.25° resolution (the original resolution of CaMa-Flood output) to 0.5° × 0.5° to ensure consistency between impact types.
Wildfire exposure was based on burned area simulated by three global vegetation models (CLASSIC, LPJmL5-7-10-fire, VISIT; Supplementary Table 1). The vegetation models simulate burned area at sub-annual time step (daily by CLASSIC and LPJmL5-7-10-fire, and monthly by VISIT). Monthly burned area output is commonly used in fire modelling49,50, although some models have moved to finer temporal resolution. Burned area is summed to annual totals for all models. A wildfire is considered an extreme event if the yearly burned area exceeds the 97.5th percentile of the respective pre-industrial control simulation, consistent with the other event types and ref. 37.
For a detailed discussion of the limitations of this dataset we refer to ref. 38. One limitation is the approach chosen to define extreme events, as alternative thresholds could yield different results. We implemented a sensitivity analysis using 95th, 97.5th and 99th percentile thresholds (5th, 2.5th and 1st for droughts) and show that the choice of percentile threshold introduces substantially less variability than climate model and impact model selection (Supplementary Figs. 6–9), which is line with previous assessments44. Furthermore, although we identify extreme values from impact model outputs, these models can have challenges simulating extreme events48. Specifically, the fire models do not fully capture the characteristics of megafires38 that have been documented to affect species in recent studies.
Change in extreme event exposure
For each 30-year period, we calculated the event frequency by dividing the number of years with events by 30. We then calculated the mean across all climate model–impact model combinations. Ensemble minimum refers to the minimum value across all model combinations and the ensemble maximum to the maximum value across all model combinations. Extreme event exposure was determined for two time periods. For the historical period, we calculated average exposure for a 30-year period centred on 2000 (1985–2014). For the future period, we calculated 30-year averages as a moving window centred on 2030–2085. The change in frequency was then determined as the difference between future and past periods. For example, a change in event frequency from 0 to 0.5 means that in a grid cell without any event in the historical period (frequency = 0), an event is projected to occur in 15 of the 30 years, that is, on average every second year (frequency = 0.5). Although we focus on event frequency to enable consistent comparison across event types, we acknowledge that intensity and duration are important for biological impacts29 and can vary independently of frequency.
To identify areas exposed to multiple extreme events, we first determined for each grid cell whether the frequency of each event type was ≥0.33 (corresponding to events occurring at least every third year on average). We then summed the number of different event types meeting this threshold per grid cell, for the years 2000, 2050 and 2085. We provide results for applying different thresholds ranging from 0.2 to 0.5 in Supplementary Figs. 10 and 11.
Species and ecoregion exposure to extreme events
For the hotspot maps, we used species richness layers from the IUCN Red List of Threatened Species (v.2023-1, accessed August 2024) for amphibians, birds, mammals and reptiles. For each taxon, we used three types of layers: number of species per grid cell for Fig. 1a–d (species richness), number of threatened species for Supplementary Fig. 1 and rarity weighted richness for Supplementary Fig. 2 (aggregate importance of each grid cell to the species occurring there, unitless). IUCN-aggregated richness patterns are consistent with those from individual species range layers (Supplementary Fig. 12). The original data in Mollweide projection at 30-km resolution were reprojected to WGS84 and resampled to match the 0.5-degree ISIMIP grid.
For all quantitative analysis, we used range layers for individual species. Species range data were obtained from multiple sources: the IUCN Red List of Threatened Species12 (terrestrial mammals and amphibians), the Global Assessment of Reptile Distributions13 (GARD, reptiles) and BirdLife International14 (birds). Following IUCN guidelines for extent of occurrence maps, we included polygons for extant species of native, reintroduced and assisted colonization origin and all seasonality codes. The initial dataset comprised 7,731 amphibians; 10,992 birds; 5,593 mammals and 10,914 reptiles. We mapped species ranges to the ISIMIP grid, by calculating for each grid cell the proportion of overlap with the species range. Due to the spatial resolution, small islands are not included in the ISIMIP grid and extreme event data are not available. Species restricted to small islands not represented in the ISIMIP grid were excluded (126 amphibians, 430 birds, 117 mammals and 621 reptiles), resulting in a final dataset of 33,936 terrestrial vertebrate species (7,605 amphibians; 10,562 birds; 5,476 mammals and 10,293 reptiles).
Ecoregion polygons were obtained from the Ecoregions 201715 dataset. The dataset includes 847 ecoregions, but 53 ecoregions had no overlap with ISIMIP land cells. We used the remaining 794 ecoregions for our analysis.
To quantify species exposure to extreme events, we calculated the proportion of grid cells exposed, and weighted each grid cell by how often it was exposed within a 30-year period, by grid cell area and the proportion of overlap between each cell and the species range. Weighting event exposure by how much each grid cell overlaps with the species range reduces overestimation for small-ranged species. Grid cell area in a 0.5-degree global grid vary due to the spherical geometry of Earth, necessitating area-based weighting. Similarly, for ecoregion exposure, we weighted grid cells by event frequency, grid cell area and the proportion of overlap between each cell and the ecoregion polygon. All analyses were conducted in R (v.4.1.0) using the packages ncdf4 (v.1.19), sf (v.1.0.8) and sp (v.1.5.0).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Climate projections, climate impact projections and extreme event data are available from the ISIMIP Repository (https://data.isimip.org/); specifically, heatwave and wildfire data can be downloaded from https://doi.org/10.48364/ISIMIP.920810 (ref. 51) and drought and river flood data from https://doi.org/10.48364/ISIMIP.792444 (ref. 52). Species richness layers were downloaded from the IUCN Red List of Threatened Species (https://www.iucnredlist.org/). Species range data were downloaded from the IUCN Red List of Threatened Species (for terrestrial mammals and amphibians), from the Global Assessment of Reptile Distributions (GARD, http://www.gardinitiative.org/) and BirdLife International (http://datazone.birdlife.org). Ecoregions were downloaded from https://ecoregions.appspot.com/.
Code availability
The code to reproduce the results of this study is available via Zenodo at https://doi.org/10.5281/zenodo.18861352 (ref. 53).
References
Mo, M. et al. Estimating flying-fox mortality associated with abandonments of pups and extreme heat events during the austral summer of 2019–20. Pac. Conserv. Biol. 28, 124–139 (2021).
Tomas, W. M. et al. Distance sampling surveys reveal 17 million vertebrates directly killed by the 2020’s wildfires in the Pantanal, Brazil. Sci. Rep. 11, 23547 (2021).
Maxwell, S. L. et al. Conservation implications of ecological responses to extreme weather and climate events. Divers. Distrib. 25, 613–625 (2019).
Seneviratne, S. I. et al. Weather and climate extreme events in a changing climate. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change (eds Masson-Delmotte, V. et al) Ch. 11 (Cambridge Univ. Press, 2021).
Foden, W. B. et al. Climate change vulnerability assessment of species. WIREs Clim. Change 10, e551 (2019).
Goymer, P. Age of extremes. Nat. Ecol. Evol. 8, 1381–1381 (2024).
Shi, H. et al. Terrestrial biodiversity threatened by increasing global aridity velocity under high-level warming. Proc. Natl Acad. Sci. USA 118, e2015552118 (2021).
van den Bosch, M., Costanza, J. K., Peek, R. A. & Steel, Z. L. Projected increases in climate extremes across global vertebrate diversity hotspots. Glob. Chang. Biol. 31, e70272 (2025).
Kropf, C. M., Vaterlaus, L., Bresch, D. N. & Pellissier, L. Tropical cyclone risk for global ecosystems in a changing climate. Nat. Clim. Chang. 15, 92–100 (2025).
Soifer, L. G. et al. Extreme events drive rapid and dynamic range fluctuations. Trends Ecol. Evol. 40, 862–873 (2025).
Frieler, K. et al. Scenario set-up and the new CMIP6-based climate-related forcings provided within the third round of the Inter-Sectoral Model Intercomparison Project (ISIMIP3b, group I and II). Preprint at https://doi.org/10.5194/egusphere-2025-2103 (2025).
International Union for Conservation of Nature. The IUCN Red List of threatened species. v.2024-1. The IUCN Red List of Threatened Species https://www.iucnredlist.org/resources (2024).
Roll, U. et al. The global distribution of tetrapods reveals a need for targeted reptile conservation. Nat. Ecol. Evol. 1, 1677–1682 (2017).
BirdLife International & Handbook of the Birds of the World. Bird species distribution maps of the world. http://datazone.birdlife.org/species/requestdis (2023).
Dinerstein, E. et al. An ecoregion-based approach to protecting half the terrestrial realm. BioScience 67, 534–545 (2017).
Davis, C. L. et al. Species interactions and the effects of climate variability on a wetland amphibian metacommunity. Ecol. Appl. 27, 285–296 (2017).
Santos, X., Chergui, B., Belliure, J., Moreira, F. & Pausas, J. G. Reptile responses to fire across the western Mediterranean Basin. Conserv. Biol. 39, e14326 (2025).
Wright, E. et al. Higher maximum temperature increases the frequency of water drinking in mountain gorillas (Gorilla beringei beringei). Front. Conserv. Sci. 3, 738820 (2022).
Beukema, W. et al. Microclimate limits thermal behaviour favourable to disease control in a nocturnal amphibian. Ecol. Lett. 24, 27–37 (2021).
Cunningham, C. X., Williamson, G. J. & Bowman, D. M. J. S. Increasing frequency and intensity of the most extreme wildfires on Earth. Nat. Ecol. Evol. 8, 1420–1425 (2024).
Gonçalves, F. et al. A global map of species at risk of extinction due to natural hazards. Proc. Natl Acad. Sci. USA 121, e2321068121 (2024).
Ruthrof, K. X. et al. Subcontinental heat wave triggers terrestrial and marine, multi-taxa responses. Sci. Rep. 8, 13094 (2018).
Gu, S., Qi, T., Rohr, J. R. & Liu, X. Meta-analysis reveals less sensitivity of non-native animals than natives to extreme weather worldwide. Nat. Ecol. Evol. 7, 2004–2027 (2023).
Luedtke, J. A. et al. Ongoing declines for the world’s amphibians in the face of emerging threats. Nature 622, 308–314 (2023).
Driscoll, D. A. et al. Biodiversity impacts of the 2019–2020 Australian megafires. Nature 635, 898–905 (2024).
Hurlbert, A. H. & Jetz, W. Species richness, hotspots, and the scale dependence of range maps in ecology and conservation. Proc. Natl Acad. Sci. USA 104, 13384–13389 (2007).
Gibson-Reinemer, D. K. & Rahel, F. J. Inconsistent range shifts within species highlight idiosyncratic responses to climate warming. PLoS ONE 10, e0132103 (2015).
Jensen, A. J. et al. Mammals on the margins: identifying the drivers and limitations of range expansion. Glob. Chang. Biol. 31, e70222 (2025).
Murali, G., Iwamura, T., Meiri, S. & Roll, U. Future temperature extremes threaten land vertebrates. Nature 615, 461–467 (2023).
Pottier, P. et al. Vulnerability of amphibians to global warming. Nature 639, 954–961 (2025).
Cabello-Vergel, J. et al. Seasonal and between-population variation in heat tolerance and cooling efficiency in a Mediterranean songbird. J. Therm. Biol. 125, 103977 (2024).
Cohen, J. M., Fink, D. & Zuckerberg, B. Spatial and seasonal variation in thermal sensitivity within North American bird species. Proc. Biol. Sci. 290, 20231398 (2023).
Wu, N. C. et al. Global exposure risk of frogs to increasing environmental dryness. Nat. Clim. Chang. 14, 1314–1322 (2024).
Des Roches, S. et al. The ecological importance of intraspecific variation. Nat. Ecol. Evol. 2, 57–64 (2018).
Kiribou, R. et al. Exposure of African ape sites to climate change impacts. PLoS Climate 3, e0000345 (2024).
Mahecha, M. D. et al. Biodiversity and climate extremes: known interactions and research gaps. Earth’s Future 12, e2023EF003963 (2024).
Lange, S. et al. Projecting exposure to extreme climate impact events across six event categories and three spatial scales. Earth’s Future 8, e2020EF001616 (2020).
Zantout, K. et al. Shifting dominant periods in extreme climate impacts under global warming. Nat Commun 16, 9746 (2025).
Müller Schmied, H. et al. Graphical representation of global water models. Geosci. Model Dev. 18, 2409–2425 (2025).
Grant, L. et al. Global emergence of unprecedented lifetime exposure to climate extremes. Nature 641, 374–379 (2025).
AghaKouchak, A. et al. Anthropogenic drought: definition, challenges, and opportunities. Rev. Geophys. 59, e2019RG000683 (2021).
Perkins, S. E. A review on the scientific understanding of heatwaves—their measurement, driving mechanisms, and changes at the global scale. Atmos. Res. 164–165, 242–267 (2015).
Twomey, E., Sylvester, F., Jourdan, J., Hollert, H. & Schulte, L. M. Quantifying exposure of amphibian species to heat waves, cold spells, and droughts. Conserv. Biol. 39, e70074 (2025).
Satoh, Y. et al. A quantitative evaluation of the issue of drought definition: a source of disagreement in future drought assessments. Environ. Res. Lett. 16, 104001 (2021).
Pokhrel, Y. et al. Global terrestrial water storage and drought severity under climate change. Nat. Clim. Chang. 11, 226–233 (2021).
Russo, S., Sillmann, J. & Fischer, E. M. Top ten European heatwaves since 1950 and their occurrence in the coming decades. Environ. Res. Lett. 10, 124003 (2015).
Yamazaki, D., Kanae, S., Kim, H. & Oki, T. A physically based description of floodplain inundation dynamics in a global river routing model. Water Res. Res. 47, W04501 (2011).
Heinicke, S. et al. Global hydrological models continue to overestimate river discharge. Environ. Res. Lett. 19, 074005 (2024).
Rabin, S. S. et al. The Fire Modeling Intercomparison Project (FireMIP), phase 1: experimental and analytical protocols with detailed model descriptions. Geosci. Model Dev. 10, 1175–1197 (2017).
Burton, C. et al. Global burned area increasingly explained by climate change. Nat. Clim. Chang. 14, 1186–1192 (2024).
Zantout, K. et al. Extreme event exposure data for crop failure, heatwave and wildfire (v.1.0). ISIMIP Repository https://data.isimip.org/10.48364/ISIMIP.920810 (2025).
Heinicke, S. et al. Extreme event exposure data for drought and river flood. Version 1.0. ISIMIP Repository https://doi.org/10.48364/ISIMIP.792444 (2026).
Heinicke, S. Code for land vertebrates exposure multiple extreme events. Zenodo https://doi.org/10.5281/zenodo.18861351 (2026).
Acknowledgements
This research has received funding from the European Union’s Horizon Europe research and innovation programme under grant no. 101056858. We gratefully acknowledge the Ministry of Research, Science and Culture (MWFK) of Land Brandenburg for supporting this project by providing resources on the high performance computer system at the Potsdam Institute for Climate Impact Research (grant no. 22-Z105-05/002/001). This work used resources of the Deutsches Klimarechenzentrum (DKRZ) granted by its Scientific Steering Committee (WLA) under project bb0820. A.I. was partly supported by the KAKENHI Fund (grant no. 21H95318).
Funding
Open access funding provided by Potsdam-Institut für Klimafolgenforschung (PIK) e.V.
Author information
Authors and Affiliations
Contributions
S. Heinicke conceptualized the study, developed methodology, analysed and visualized the data and wrote the original draft. H.S.K. and C.P.O.R. developed methodology. K.F. acquired funding and supervized the study. K.Z., S.Z., M.B., S.N.G., M.G., S. Hantson, A.I., S.K.-G., A.K., H.M.S., S.O., K.O. and Y.P. contributed data. B.M. provided analysis for an earlier version. All authors reviewed and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Ecology & Evolution thanks Laura Antão and Gopal Murali for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Exposure of species richness to multiple event types.
a-c, number of event types projected for SSP3 − 7.0 for the year (a) 2000, (b) 2050 and (c) 2085, d-f, exposure to multiple events mapped against species richness combined across all four taxa (from the IUCN Red List of Threatened Species) for the same three time periods.
Extended Data Fig. 2 Exposure to projected change in extreme event occurrences for ecoregions.
Change in frequency of extreme events from 2000 to 2050 for SSP3 − 7.0 for (a) heatwave, (b) wildfire, (c) drought, (d) river flood for 794 ecoregions. Values for each climate model – impact model combination are in Supplementary Fig. 17.
Supplementary information
Supplementary Information (download PDF )
Supplementary Tables 1–9 and Figs. 1–17.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Heinicke, S., Zantout, K., Kühl, H.S. et al. Land vertebrates increasingly exposed to multiple extreme events by 2085. Nat Ecol Evol 10, 854–863 (2026). https://doi.org/10.1038/s41559-026-03050-0
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41559-026-03050-0
