
Abstract
Mammalian reproduction requires substantial immune adaptations to safeguard reproductive success and to ultimately shape the evolutionary trajectories of a species. Systemic and placental immunity shift towards tolerance during pregnancy1,2; however, how maternal immunity adapts in barrier tissues—which are sites of frequent infection and inflammation—from pregnancy until the postpartum lactation period remains poorly understood. Here we report a previously unrecognized role for eosinophils, a type of granulocyte typically associated with allergies and helminth infections3,4, in remodelling the intestinal barrier during reproduction. Beginning in pregnancy and peaking during lactation, eosinophils accumulate in the small intestine in the absence of infection or inflammation. Using genetic and pharmacological perturbations, organoid cultures and single-cell and spatial transcriptomics, we show that eosinophils promote goblet cell differentiation in a stem-cell-intrinsic manner that leads to increased mucus production. This remodelling culminates during lactation and limits pathogen entry and dissemination to confer broad innate protection against enteric bacterial infections. Moreover, in mice, intestinal remodelling and innate defence persist weeks after lactation cessation. Our findings demonstrate that despite a general trend towards systemic immune modulation during reproduction, the maternal intestine undergoes remodelling to strengthen innate defence, a mechanism that may have evolved to protect mothers and offspring in pathogen-rich environments. More broadly, we establish a framework for studying tissue-specific immune adaptation across the reproductive cycle and highlight that tissues can retain changes following physiological reproduction, with lasting implications for host defence and women’s health.
Similar content being viewed by others

Main
During mammalian reproduction from pregnancy until lactation, mothers undergo substantial immunological adaptations to tolerate semi-allogeneic offspring while navigating constant environmental exposures to pathogens. Maintaining this delicate balance between supporting offspring and defending against infection is essential for the survival and health of both mother and child, which ultimately shapes the fitness and evolutionary trajectories of a species.
During pregnancy, maternal immunity shifts towards higher tolerance in blood and reproductive tissues to safeguard fetal development1,2,5,6. However, first-line immune defence is primarily orchestrated at barrier tissues, particularly the gut, lungs and skin, which are constantly in contact with environmental challenges that range from beneficial microbiota to pathogens. Human and mouse studies have revealed that immune cells in barrier tissues differ markedly from those in the circulation and in lymphoid organs. For example, tissue-resident immune cells have specialized roles in local defences and in maintaining tissue homeostasis7,8,9,10. Yet, whether and how maternal immunity adapts in barrier tissues remains largely unexplored. Moreover, maternal immunity has been primarily studied in the three trimesters of pregnancy, with attention shifting to the newborn immediately after delivery. As a result, maternal immunity during the postpartum lactation period, a defining feature of all mammals, remains poorly understood.
One of the most notable physiological changes that occurs during reproduction is intestinal expansion, a phenomenon that has been documented across species11,12,13. Emerging evidence suggests that this growth enhances nutrient absorption to meet heightened energy demands to support offspring development14,15. Here we demonstrate that in addition to intestinal expansion, eosinophils accumulate throughout reproduction in the maternal intestine. These eosinophils promote goblet cell differentiation to enhance innate protection against enteric pathogens. Moreover, this intestinal immune adaptation and enhanced innate defence persists long after lactation. Increased circulating eosinophils have also been observed in postpartum humans16, which suggests that this eosinophil-driven barrier remodelling in reproduction may be conserved across mammals. We propose that this intestinal-specific immune adaptation in reproduction strengthens innate defences at a time of systemic immune modulation to support maternal and offspring survival and contributes to species fitness in pathogen-rich environments.
Gut eosinophils increase during reproduction
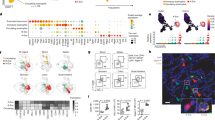
To investigate how maternal immunity adapts at barrier tissues during reproduction, we used a hormone-induced timed-pregnancy mouse model, focusing on day 12 of pregnancy and day 12 of lactation, which coincides with peak breastmilk production17 (Fig. 1a). We examined the small intestine, a barrier tissue crucial for nutrient absorption and for initiating immune defences against foodborne pathogens. We isolated immune cells from the small intestinal lamina propria and found a significant increase in total immune cell numbers in pregnant and lactating mice compared with control mice (Fig. 1b). To determine whether this increase reflected expansion of specific immune populations, we used multiparameter flow cytometry to quantify major innate and adaptive immune cell subsets (Extended Data Fig. 1a,b). Most subsets exhibited coordinated increases across reproductive stages, each showing varying degrees of induction. This result demonstrates that there is a dynamic shift in intestinal immunity during reproduction (Extended Data Fig. 1c). Notably, eosinophils exhibited the highest and most sustained increase in frequency during pregnancy and lactation (Fig. 1c). Measurements of the canonical eosinophil markers CD11b and siglec-F highlighted a progressive accumulation of eosinophils. Specifically, there was a twofold increase in frequency and up to a fourfold increase in total cell number during lactation compared with controls (Fig. 1d). Given their marked increase and abundance during lactation, we focused our subsequent analyses on intestinal eosinophils during this reproductive stage.

a, Timeline of the experimental model. Small intestinal tissues were collected from timed-pregnant dams on gestational day 12 and on day 12 of lactation. Age-matched nulliparous female mice served as controls. hCG, human chorionic gonadotropin; PMSG, pregnant mare serum gonadotropin. b, Absolute numbers of live CD45+ immune cells in the small intestine lamina propria (siLP) from control (n = 16), pregnant (n = 15) or lactating (n = 15) mice. c, Fold change in frequency of various immune cell populations in the siLP of pregnant mice and lactating mice compared with nulliparous controls (n = 4). d, Representative contour plots (left) and bar graphs (right) showing eosinophil frequency and absolute numbers in the siLP from control (n = 16), pregnant (n = 15) or lactating (n = 15) mice. e, Representative confocal images of proximal small intestine stained for eosinophils (EPX) and epithelial cells (EPCAM). Scale bars, 50 μm. f, Fold change in eosinophil numbers across bone marrow, spleen, lung, skin and siLP in lactating mice compared with nulliparous controls (n = 4). g, Representative contour plots (left) and bar graphs (right) showing frequency and absolute numbers of eosinophil precursors (Lineage–SCA1–KIT+CD34+CD16+CD32+CD125+) in the bone marrow from control (n = 16) and lactating (n = 13) mice. h, siLP eosinophil frequency and absolute numbers in distinct small intestinal regions: proximal, middle and distal (n = 5). Data are representative or pooled from at least two independent experiments. Each dot represents an individual mouse. Error bars and numbers in representative flow plots indicate the mean ± s.d. Significance was calculated using one-way analysis of variance (ANOVA) (b,d), two-tailed unpaired Student’s t-test (g) or two-way ANOVA (h). Fold changes were calculated relative to the mean of the control group, statistical analyses were performed on raw values using an unpaired Student’s t-test (c,f). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Icons in a and c created in BioRender; Huang, C. https://BioRender.com/pmrvxbl (2026).
Intestinal eosinophils are heterogeneous and adapt to the tissue microenvironment in response to local cues such as dietary factors18, endogenous molecules19,20 and the neuropeptide neuromedin U21. Activated intestinal eosinophils are characterized by PD-L1 expression22, whereas villus-resident eosinophils are marked by CD22 expression18,23. We observed a modest but significant increase in frequency of PD-L1+ and CD22+ eosinophils in the small intestine during lactation (Extended Data Fig. 2a). Imaging analyses confirmed an increase in eosinophil peroxidase-positive (EPX+) cells distributed throughout the villi, with enrichment near the villus base and comparable densities between control and lactating mice (Fig. 1e and Extended Data Fig. 2b–d). Despite the upregulation of activation markers, we did not observe differences in granularity (SSC-A) or secretory activity (CD9, CD63 and CD107a) between eosinophils from control and lactating mice (Extended Data Fig. 2e,f). Moreover, eosinophils from both groups produced comparable levels of canonical eosinophil-associated cytokines ex vivo, including IL-4, IL-10, IL-13 and IL-18 (Extended Data Fig. 2g). These findings indicate that eosinophils accumulate in increased numbers and adopt an activated phenotype during lactation, whereas their secretory function remains comparable to nulliparous control mice.
Next, we sought to determine whether eosinophil accumulation during reproduction is confined to the intestines or occurs systemically. To this end, we quantified eosinophils across lymphoid and barrier tissues. We observed a twofold increase in eosinophil numbers in the bone marrow of lactating mice, whereas eosinophil levels in the spleen, lung and skin remained unchanged (Fig. 1f). The number and frequency of eosinophil progenitors were elevated in bone marrow during lactation, which suggested that there was enhanced eosinophil production followed by migration to the intestines (Fig. 1g and Extended Data Fig. 2h). Increases in eosinophil production in the bone marrow and their accumulation in the intestine occurred during natural reproduction, which indicated that this phenomenon is independent of exogenous hormone treatment (Extended Data Fig. 2i). Notably, germ-free lactating mice also exhibited increased eosinophil numbers in the bone marrow but not in the small intestine. This result suggests that the microbiota may contribute to eosinophil accumulation in the intestine during reproduction (Extended Data Fig. 2j).
The small intestine exhibits zonation in epithelial composition, transcriptomic landscapes and physiological functions along the proximal-to-distal axis24,25,26,27. To determine whether eosinophil accumulation occurs uniformly along this axis, we quantified eosinophils in three regions of the small intestine. Specifically, we defined the first 10 cm adjacent to the stomach as proximal, the last 10 cm as distal and the intervening segment as middle. Eosinophil numbers and frequency increased most prominently in the proximal small intestine, exhibited a moderate rise in the middle and remained unchanged in the distal region. Moreover, during lactation, eosinophils accounted for up to 40% of immune cells in the proximal small intestine (Fig. 1h). This marked accumulation of eosinophils in the absence of infection or inflammation is unexpected given their classical roles as effector cells in type 2 inflammatory responses, such as during allergen exposure or helminth infection3,4,28,29. To explore whether this eosinophil enrichment is accompanied by type 2 immune activation, we next assessed cytokine production. Although we observed a significant increase in intestinal group 2 innate lymphoid (ILC2) cells during lactation, T helper 2 (TH2) cells remained unchanged in number (Extended Data Fig. 1c). Moreover, ILC2 cells exhibited a reduced capacity to produce type 2 cytokines, including IL-5 and IL-13, during lactation (Extended Data Fig. 2k). Similarly, although the number of other lymphocyte populations, including ILC1, ILC3, TH1, TH17 and cytotoxic CD8 T cells, remained stable or showed modest induction (Extended Data Fig. 1c), their cytokine production, including IFNγ and IL-17A, was significantly reduced during lactation (Extended Data Fig. 2k–m). These data show that canonical cytokine production in lymphocytes is reduced during lactation.
Together, these findings demonstrate that the maternal intestinal immune landscape undergoes significant adaptations during reproduction that is marked by both a general increase in immune cell numbers and suppressed cytokine production by lymphocytes. Among these changes, the striking enrichment of eosinophils in the proximal small intestine, in the absence of infection or inflammation, points to a physiological immune adaptation tailored to the reproductive state.
Lactation induces goblet cells
Recent studies have shown that eosinophils in the small intestine contribute to tissue remodelling30 by shaping intestinal architecture at homeostasis31,32, promoting goblet cell differentiation21 and degrading extracellular matrix structures during helminth infection19. Building on our observation of increased eosinophil accumulation in the proximal small intestine during lactation, we next investigated tissue remodelling during reproduction. In line with recent reports14,15, the small intestines of pregnant and lactating mice expanded significantly in length, increasing by up to 30% during lactation compared with nulliparous controls (Extended Data Fig. 3a). Notably, total immune cells and eosinophils were proportionally maintained relative to tissue mass. However, when accounting for intestinal length, myeloid cells continued to expand from pregnancy to lactation, whereas type 2 lymphocytes peaked only during pregnancy. This result highlights the dynamic remodelling of intestinal immunity during reproduction (Extended Data Fig. 3b,c). Both the crypt and villus expanded in length during lactation, which led to a significant increase in the absorptive surface area of the proximal small intestine (Extended Data Fig. 3d,e). These findings indicate that similar to other segments of the small intestine14,15, the proximal small intestine expands to meet the increased nutrient demands of reproduction.
To examine whether such morphological alterations are accompanied by shifts in epithelial cellular composition and transcriptomic profiles, we isolated intestinal epithelial cells (IECs) from the proximal small intestine of nulliparous control mice, pregnant mice and lactating mice and performed single-cell transcriptomic analyses. Through unsupervised clustering, we identified six major IEC subtypes (Fig. 2a, Extended Data Fig. 4a and Supplementary Table 1). Chi-square tests revealed progressive changes across reproductive stages, with the most marked alterations occurring during lactation. Specifically, stem cell frequency decreased from 20% to 6%, whereas goblet cells increased from 31% to 42% of IECs during lactation (Fig. 2b,c). Immunofluorescence staining further confirmed reduced numbers of LGR5+ stem cells per crypt and an increase in UEA1+ goblet cells along villi compared with controls (Fig. 2d,e and Extended Data Fig. 4b,c).

a, Uniform manifold approximation and projection (UMAP) plots of scRNA-seq data showing the six IEC subsets and signature genes of each subset. Stem and goblet cell clusters are indicated by dashed circles. b, Heatmap of significant changes in IEC subset proportions during pregnancy and lactation compared with nulliparous controls. c, Changes in frequencies of each IEC subset. d, Left, representative confocal images of stem cells in duodenal crypts. Scale bars, 100 μm. Right, average count of stem cells per crypt (n = 3). e, Left, representative confocal images of goblet cells in duodenal villi. Scale bars, 50 μm. Right, average count of UEA1+ goblet cells per villus for control (n = 5) and lactating (n = 4) mice. f, Volcano plot of differentially expressed genes in stem cells. Green, goblet cell signatures; pink, nutrient absorption; purple, stemness signatures. g, Left, representative bright-field images of day 8 organoids derived from single LGR5+ stem cells. Scale bars, 200 μm. Right, clonogenicity and average organoid area for control (n = 8) and lactating (n = 6) mice. h, Left, representative confocal images of day 8 organoids, with staining of goblet cells (UEA1) and epithelial cells (EPCAM). Scale bars, 50 μm. Right, average UEA1+ goblet cell counts in organoids (n = 4). i, Genes differentially overexpressed in goblet cells. Green, goblet cell signatures. For d, e, g and h, data are representative or pooled from at least three independent experiments. Each dot represents an individual mouse. Error bars represent the mean ± s.d. Significance was calculated using Chi-square tests (b) or two-tailed unpaired Student’s tests (d,e,g,h). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
We next investigated whether alterations in stem cell and goblet cell frequencies during lactation are accompanied by transcriptional changes. Comparative single-cell transcriptomic analyses of stem cells from control and lactating mice revealed two distinct subclusters (Extended Data Fig. 4d). Notably, stem cells from lactating mice exhibited downregulation of stemness markers (Lgr5, Olfm4 and Stmn1), coupled with an upregulation of genes associated with nutrient absorption (S100g, Fabp1, Fabp2 and Prap1) and goblet cell differentiation (Zg16, Tff3 and Spink4) (Fig. 2f). These transcriptomic alterations suggest that intestinal stem cells undergo transcriptional reprogramming during lactation to favour goblet cell differentiation over self-renewal. To test this hypothesis, we isolated single LGR5+ intestinal epithelial stem cells from control and lactating mice and assessed their clonogenicity, growth and lineage commitment in ex vivo organoid cultures (Extended Data Fig. 4e). Stem cells from both groups exhibited comparable organoid-forming efficiency. However, organoids derived from stem cells of lactating mice were significantly larger, which indicated their enhanced differentiation capacity (Fig. 2g). Moreover, goblet cell density was substantially higher in organoids from lactating mice than controls. This result supports the idea that lactation intrinsically reprograms intestinal stem cells towards goblet cell fate (Fig. 2h and Extended Data Fig. 4f).
Goblet cells are specialized secretory cells that produce antimicrobial peptides and mucus, which form a protective physical barrier between the intestinal epithelium and luminal microbes. Comparative transcriptomic analyses of goblet cells from lactating mice revealed significant upregulation of several key mucus components, including Muc2, Muc13, Clca1 and Fcgbp, and of antimicrobial peptides, such as Reg1 and Reg4, compared with controls (Fig. 2i). In line with these findings, we observed a substantial increase in MUC2-producing goblet cells specifically in the proximal small intestine, with no differences in distal intestinal regions of lactating mice (Extended Data Fig. 4g,h).
Together, our integrated analyses demonstrate that lactation drives substantial remodelling of the proximal small intestinal epithelium that is marked by coordinated shifts in stem and goblet cell populations, a result that highlights the region-specific nature of this adaptation. Notably, the concurrent increase in eosinophils and goblet cells was observed in allogeneic reproduction, which indicates that these adaptations are not restricted to syngeneic mating (Extended Data Fig. 4i,j).
Eosinophils induce goblet cells in lactation
In the context of helminth infection and type 2 inflammation, eosinophils promote goblet cell hyperplasia as part of the host defence response33,34. Building on this established role, we sought to determine whether eosinophils similarly contribute to goblet cell induction during reproduction. To this end, we pharmacologically targeted CCR3, a key chemokine receptor, to block eosinophil accumulation in the intestine. Notably, both nulliparous and lactating mice exhibited comparable levels of eosinophils in the blood, which constituted over 80% of CCR3+ cells in both conditions (Extended Data Fig. 5a,b). In the bone marrow, CCR3 was expressed by around 27% of total eosinophils in nulliparous mice, and this frequency significantly increased during lactation (Extended Data Fig. 5c). To directly validate the efficacy of CCR3 blockade, we performed chemotaxis assays and found that anti-CCR3 treatment reduced bone marrow eosinophil migration towards eotaxin-1 and eotaxin-2 in vitro (Extended Data Fig. 5d). Accordingly, CCR3 blockade did not influence eosinophil numbers in the bone marrow but substantially reduced eosinophil numbers in the blood and abolished eosinophils in the small intestine. By contrast, other intestinal immune populations remained largely unchanged in both nulliparous and lactating mice (Extended Data Fig. 5e,f). Anti-CCR3 treatment did not affect pregnancy outcomes, intestinal elongation, villus length or maternal or pup bodyweight (Extended Data Fig. 6a–d). However, blocking eosinophil migration significantly reduced goblet cell numbers in the proximal small intestine of lactating mice (Fig. 3a and Extended Data Fig. 6e). To further investigate whether eosinophils act directly on intestinal epithelial stem cells to promote goblet cell differentiation, we generated organoid cultures using proximal small intestinal stem cells from lactating mice in which eosinophils had been temporarily depleted through CCR3 blockade. Compared with isotype controls, organoids derived from eosinophil-depleted lactating mice exhibited markedly reduced goblet cell differentiation to levels comparable to those of nulliparous mice (Fig. 3b and Extended Data Fig. 6f). Together, these data provide support that eosinophils have a direct role in driving goblet cell fate during reproduction.

a,b, Timed-pregnant dams were injected with isotype or anti-CCR3 antibodies. a, Left, representative images of villus-resident UEA1+ goblet cells (indicated by white arrowheads) from lactating mice. Scale bars, 70 μm. Right, average count of goblet cells per villus in the proximal small intestine of control mice (n = 3) and mice given the isotype (n = 6) or anti-CCR3 (n = 5) antibody. b, Left, representative images of organoids derived from single LGR5+ intestinal stem cells. Scale bars, 50 μm. Right, average counts of UEA1+ goblet cells in organoid culture from control (n = 3) mice and mice given the isotype (n = 4) or anti-CCR3 (n = 3) antibody. c, Average count of UEA1+ goblet cells per villus in the proximal small intestine (n = 3). d–j, Xenium analysis of the first 10 cm of the proximal small intestine. d, Representative views of cell segmentation and annotation along the crypt–villus structure. Scale bars, 100 μm. NK cell, natural killer cell. e, Absolute number of stem cells, goblet cells and enterocytes detected in samples (n = 4). f,g, Mean kernel density of cell counts along the proximal–distal intestinal length, with shaded s.d. in each condition for goblet cells (f) and eosinophils (g). h, Differentially overexpressed genes that are increased in eosinophils in lactating mice compared with control mice. i, Gene expression (z-scored) within eosinophil subclusters. j, Left, mean kernel density of eosinophil subsets across the normalized crypt–villus longitude, with s.d. indicated. Right, representative views of eosinophil subsets along the crypt–villus structure. Scale bars, 100 μm. For a–c, data are pooled or representative of at least two independent experiments. Each dot represents an individual mouse. For d–j, n = 4 biological replicates pooled from two independent experiments. Error bars represent the mean ± s.d. Significance was calculated using one-way ANOVA (a,b), two-way ANOVA (c) or two-tailed unpaired Student’s t-test (e). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, NS, not significant. Icons in a created in BioRender; Huang, C. https://BioRender.com/pmrvxbl (2026).
We next compared eosinophil-deficient ΔdblGATA mice to their wild-type littermates during nulliparous and lactation periods35. ΔdblGATA mice exhibited considerable ablation in intestinal eosinophils without major alterations in other immune populations (Extended Data Fig. 7a,b). Notably, ΔdblGATA dams delivered comparable numbers of pups and had intestinal lengths similar to wild-type dams, which indicated that eosinophil deficiency does not impair pregnancy outcomes or intestinal growth (Extended Data Fig. 7c,d). However, lactating ΔdblGATA dams exhibited significantly reduced goblet cell numbers in the proximal small intestine compared with wild-type lactating controls. By contrast, no differences were observed in goblet cell numbers between nulliparous wild-type and ΔdblGATA mice. These data indicate that eosinophils are dispensable for goblet cell differentiation and maintenance under homeostatic conditions but are required specifically during reproduction (Fig. 3c).
Having established that eosinophils are required for goblet cell induction during reproduction, we next sought to spatially resolve eosinophil localization and transcriptional profiles in situ. To minimize transcriptional degradation, we used a spatial transcriptomic approach with the 10x Genomics Xenium platform, which enabled the detection of more than 5,000 genes. We analysed the first 10 cm of small intestine with preserved tissue architecture from control and lactating mice (Extended Data Fig. 8a). As expected, the same anatomical segment from lactating mice had an increased total cell number compared with nulliparous controls (Extended Data Fig. 8b). Using multimodel cell segmentation and unsupervised transcriptomic clustering, we identified major epithelial and immune cell clusters across the tissue (Extended Data Fig. 8c,d and Supplementary Table 2). Spatial visualization of these clusters revealed substantial regionalization, with EPCAM+ cells forming the single-layer epithelial barrier and CD45+ immune cells residing in the lamina propria (Fig. 3d). Although the absolute number of stem cells remained unchanged, we observed an expansion of differentiated epithelial cells, particularly enterocytes and goblet cells (Fig. 3e). Analyses along the proximal–distal longitudinal axis revealed that goblet cell induction occurred throughout the proximal small intestine (Fig. 3f). This spatial approach captures the complete villus–crypt architecture and therefore complements our single-cell RNA sequencing (scRNA-seq) analysis, which used a crypt-enrichment strategy to ensure representation of all epithelial subsets and report their frequencies in that compartment (Fig. 2a–c). Together, these complementary approaches support the idea that although the stem cell pool is maintained during lactation, intestinal remodelling favours goblet cell differentiation.
We next focused on eosinophils, which were identified on the basis of high expression of the bona fide markers Ccr3 and Il5ra in the myeloid cluster (Extended Data Fig. 8d). Consistent with this transcriptomic profile, flow cytometry analysis confirmed that all intestinal eosinophils expressed CCR3, albeit at variable levels, whereas other myeloid populations showed minimal expression (Extended Data Fig. 8e). We acknowledge that spatial transcriptomics may not capture the full extent of eosinophil abundance in the tissue. However, we observed a trend towards increased eosinophil numbers across the proximal small intestine, paralleling the overall pattern of goblet cell induction (Fig. 3g and Extended Data Fig. 8f). Notably, differential gene expression analysis revealed that eosinophils from lactating mice upregulated the expression of extracellular matrix components, including collagens (Col6a2, Col18a1 and Col4a1), laminins (Hspg2) and fibronectin (Fn1), which suggests that eosinophils have a role in active tissue remodelling during lactation. We also observed upregulated expression of Nmur1 (Fig. 3h), which encodes a factor that has been previously implicated in goblet cell induction in the context of helminth infection21. Neighbouring cell analysis recapitulated expected spatial relationships, with stem cells positioned adjacent to transit-amplifying and Paneth cells, a result that provided support for the robustness of our spatial data (Extended Data Fig. 8g). Eosinophils were located near stromal cells but did not show spatial proximity to stem or goblet cells. This result suggests that their effects on goblet cell differentiation are unlikely to occur through direct cell–cell interactions (Extended Data Fig. 8h). Notably, eosinophils in the proximal small intestine displayed transcriptomic heterogeneity and formed four main subclusters: a Ccr3hi cluster; a Il5rahi cluster; a population resembling previously described active eosinophils marked by Cd80 and Cd274 expression20; and basal eosinophils characterized by the relatively low expression of these markers (Fig. 3i and Supplementary Table 3). All four subsets were detected along the villus–crypt axis without a distinct zonation pattern and exhibited similar spatial organization between control and lactating mice. Among these subsets, Ccr3hi and active eosinophils were the most abundant, but only active eosinophils increased significantly during lactation (Fig. 3j and Extended Data Fig. 8i). Active eosinophils upregulated genes related to extracellular matrix and immune activation, which suggests that they represent a key effector population that drives intestinal remodelling during reproduction (Extended Data Fig. 8j and Supplementary Table 3). Together, our data demonstrate that eosinophils exhibit distinct transcriptional signatures during lactation and are positioned to support intestinal remodelling in the proximal small intestine.
Eosinophils promote innate defence in lactation
Under homeostatic conditions, goblet cells are most abundant in the distal gut, where they form a protective mucus layer that limits direct epithelial contact with dense microbial populations36,37. During lactation, we observed augmented eosinophil accumulation and goblet cell expansion in the proximal small intestine, which raised the possibility that this adaptation enhances resistance to enteric pathogens. To test this hypothesis, we used Yersinia pseudotuberculosis, a foodborne Gram-negative bacterium that targets the small intestine and rapidly disseminates to systemic organs38. Following oral infection, lactating mice exhibited significantly lower faecal pathogen burden than nulliparous controls. This effect began as early as 6 h after infection and persisted throughout the innate phase of infection (Fig. 4a,b). Furthermore, although baseline mucus deposition was comparable between groups, lactating mice mounted a robust increase in mucus production in response to infection, which indicated that there was enhanced inducible mucus barrier defence during lactation (Extended Data Fig. 9a). Nulliparous controls showed a trend towards increased eosinophil and neutrophil numbers after infection. By contrast, myeloid cell numbers, which are already elevated during lactation, did not increase further following infection (Extended Data Fig. 9b). Moreover, eosinophils in control and lactating mice displayed comparable granularity and PD-L1 expression before and after infection, which suggested that there was no major shift in activation state (Extended Data Fig. 9c). Correspondingly, bacterial dissemination was significantly reduced, with only 55% of lactating mice exhibiting splenic Y. pseudotuberculosis colonization compared with 88% in nulliparous control mice, with a 1–2 log reduction in splenic bacterial load (Fig. 4c). Although lactating mice experienced mild bodyweight loss following infection, pup growth during this period was unaffected (Extended Data Fig. 9d). Together, these results reveal that contrary to the prevailing view that reproduction increases infection susceptibility, lactation is associated with enhanced innate defence in the gut and enables effective restriction of pathogen colonization and limits systemic dissemination.

a, Schematic of the experiment of oral infection of lactating dams and controls with foodborne pathogens. b,c, Infection with 2 × 107 Y. pseudotuberculosis. b, Left, faecal burden at 6 h for control (n = 15) and lactating (n = 23) mice. Right, kinetics for control (n = 13, 12, 12 for days 1, 2, 3) and lactating (n = 13, 13, 20 for days 1, 2, 3) mice. c, Dissemination at day 3. Top, frequency of mice with detectable pathogen. Bottom, splenic burden. n = 24 control mice; n = 20 lactating mice. d,e, Infection with 106 L. monocytogenes. d, Left, faecal burden at 6 h (n = 9). Right, kinetics for control (n = 9, 8, 9 for days 1, 2, 3) and lactating (n = 9) mice. e, Dissemination at day 3. Top, frequency of mice with detectable pathogen. Bottom, pathogen burden (n = 9). f–h, Infection with 106 S. Typhimurium. f, Left, faecal burden at 6 h (n = 15). Right, kinetics for control (n = 9, 8, 10 for days 1, 2, 5) and lactating (n = 6, 6, 12 for days 1, 2, 5) mice. g, Dissemination at day 5 for control (n = 15) and lactating (n = 14) mice. h, Survival curves. i–m, Eosinophil depletion using anti-CCR3 in timed-pregnant dams followed by infection with Y. pseudotuberculosis (j,k) or S. Typhimurium during lactation (l,m). j, Left, faecal burden at 6 h for control mice (n = 10) and mice given an isotype (n = 11) or anti-CCR3 (n = 12) antibody. Right, kinetics for control mice (n = 10, 5, 8 for days 1, 2, 3) and mice given an isotype (n = 11) or anti-CCR3 (n = 12, 13, 12 for days 1, 2, 3) antibody. k, Dissemination at day 3. Top, frequency of mice with detectable pathogen. Bottom, splenic burden. n = 19 for control, n = 18 for isotype, n = 24 for anti-CCR3. l, Faecal burden at 6 h for control mice (n = 15) and mice given an isotype (n = 15) or anti-CCR3 (n = 16) antibody. m, Dissemination at day 3 for control mice (n = 15) and mice given an isotype (n = 15) or anti-CCR3 (n = 16) antibody. Data are pooled from at least two independent experiments. Each dot represents an individual mouse (except kinetics). Error bars are the mean ± s.d. Significance analysis: two-tailed unpaired Student’s t-test (b–h), one-way ANOVA (j–m) or Fisher’s exact test for frequency (c). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Icons in a and i created in BioRender; Huang, C. https://BioRender.com/pmrvxbl (2026).
To determine whether lactation-associated innate protection extends to additional enteric pathogens, we orally infected mice with representative foodborne pathogens that primarily colonize the small intestine. Following Listeria monocytogenes infection, lactating mice exhibited a 1–2 log reduction in faecal pathogen burden during the innate phase of infection, which was accompanied by substantially reduced systemic dissemination. Indeed, around 80% of mice did not show detectable burden in the spleen (Fig. 4d,e). Similarly, lactating mice displayed significantly reduced bacterial shedding following Salmonella enterica subsp. enterica serovar Typhimurium infection as early as 6 h after infection, with a 1–2 log reduction compared with controls, and this effect persisted throughout the innate phase of infection (Fig. 4f). Moreover, although S. Typhimurium infection is typically lethal in mice39, reduced intestinal colonization in lactating mice translated into reduced dissemination and significantly improved survival following infection (Fig. 4g,h). Together, these findings demonstrate that lactation confers broad-spectrum innate protection against enteric pathogens by limiting early colonization and dissemination to distal organs.
Having established that eosinophils drive goblet cell induction during lactation, we next sought to determine whether this adaptation is important for the enhanced innate protection we observed. To this end, we administered anti-CCR3 antibodies to pregnant and lactating mice to block eosinophil migration to the intestine followed by oral Y. pseudotuberculosis challenge (Fig. 4i). Anti-CCR3-treated lactating mice exhibited increased faecal pathogen burden and elevated systemic dissemination, with higher frequencies of splenic colonization and elevated bacterial loads (Fig. 4j,k). Similarly, after S. Typhimurium infection, eosinophil depletion through anti-CCR3 abrogated lactation-associated protection, which led to a modest induction in pathogen burden and a significant increase in systemic dissemination (Fig. 4l,m). Together, these findings indicate that intestinal eosinophils orchestrate maternal innate defence during lactation through two coordinated mechanisms. First, reinforcement of goblet cell-derived mucus barrier that limits pathogen entry and then restriction of dissemination to distal tissues.
To examine whether intestinal eosinophil-driven goblet cell induction and innate defence are specific to lactation or reflect pregnancy-associated changes, we compared pregnant and postpartum non-lactating mice with nulliparous controls and lactating dams. In postpartum non-lactating mice, eosinophil numbers in the bone marrow showed a modest increase relative to controls, whereas intestinal eosinophil levels had returned to baseline (Extended Data Fig. 9e). Goblet cell numbers in both pregnant and postpartum non-lactating mice were significantly increased compared with controls yet remained lower than actively lactating mice (Extended Data Fig. 9f). Functionally, neither pregnant nor postpartum non-lactating mice displayed enhanced innate defence to foodborne pathogens, and displayed faecal pathogen burden and splenic dissemination levels comparable to controls (Extended Data Fig. 9g–j). Taken together, these findings indicate that eosinophils progressively accumulate in the intestine and promote goblet cell induction across reproduction, whereas effective innate defence to foodborne pathogens only culminates during active lactation.
Reproduction reshapes gut immunity
Last, to determine whether these reproduction-induced adaptations persist beyond the reproductive period, we examined intestinal immunity at 12 weeks after lactation (Fig. 5a), approximately 4 times the duration of the lactation period40. Eosinophils and other myeloid populations, including neutrophils, macrophages and dendritic cells, remained elevated compared with age-matched nulliparous controls (Fig. 5b), albeit at levels lower than those observed during active reproduction (Extended Data Fig. 10a). By contrast, the increase in lymphocyte numbers and reduction in cytokine production observed during reproduction had fully resolved (Extended Data Fig. 10b,c). Post-lactating mice retained significantly elongated small intestines, despite the villus length returning to baseline (Extended Data Fig. 10d,e). When immune cell numbers were normalized to intestinal length, eosinophils, monocytes and macrophages and dendritic cells remained elevated, which indicated that these effects were due to immune remodelling rather than a consequence of increased tissue size (Extended Data Fig. 10f). Notably, goblet cell numbers in the proximal small intestine remained elevated, which was accompanied by sustained mucus production (Fig. 5c,d and Extended Data Fig. 10g,h). Functionally, post-lactating dams exhibited enhanced innate defence against a foodborne pathogen, with reduced pathogen burden and limited systemic dissemination compared with age-matched nulliparous controls (Fig. 5e–g). However, this residual innate defence was dose-dependent and attenuated, as post-lactating mice did not control infection at the higher inoculum used during lactation (Fig. 5h). Notably, intestinal permeability was unchanged during lactation and post-lactation, which provided support that the integrity of the barrier was preserved (Extended Data Fig. 10i).

a, Timed-pregnant female mice were analysed 12 weeks after lactation. b, Absolute numbers of major myeloid populations in the siLP of control (n = 11) and post-lactation (n = 10) mice. c, Left, representative confocal images of the proximal small intestine, with staining for villus-resident goblet cells (UEA1), epithelial cells (EPCAM) and nuclei (DAPI). Scale bars, 70 μm. Right, average count of UEA1+ goblet cells per villus (n = 8). d, Left, representative confocal images of the small intestine, with staining for MUC2+ goblet cells. Scale bars, 50 μm. Right, average count of MUC2+ goblet cells per villus from control (n = 4) and post-lactation (n = 3) mice. e–g, Mice 12 weeks post-lactation and age-matched nulliparous controls were orally infected with 105 S. Typhimurium. e, Schematic of the infection model. f, Left, faecal pathogen burden at 6 h after infection in control (n = 18) and post-lactation (n = 17) mice. Right, kinetics of faecal pathogen burden in control (n = 12, 9 and 16 for day 1, 3 and 5, respectively) and post-lactation (n = 9, 12 and 15 for days 1, 3 and 5, respectively) mice. g, Pathogen dissemination at day 5 after infection. Top, frequency of mice with detectable S. Typhimurium in the spleen. Bottom, pathogen burden in the spleen for control (n = 15) and post-lactation (n = 16) mice. h, Pathogen burden in faeces at 6 h after infection with 106 S. Typhimurium in control (n = 9) and post-lactation (n = 10) mice. Data are representative or pooled from at least two independent experiments. Each dot represents an individual mouse except for the kinetic plot. Error bars represent the mean ± s.d. Significance was calculated using two-tailed unpaired Student’s t-test (b–d,f–h). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Icons in a and e created in BioRender; Huang, C. https://BioRender.com/pmrvxbl (2026).
Discussion
Collectively, our findings demonstrated that intestinal immunity undergoes transformative and lasting remodelling in response to reproduction. During reproduction, in the absence of infection or inflammation, eosinophils accumulate in the small intestine where they program goblet cell fate and enhance innate defences against enteric pathogens. These results challenge the prevailing view that reproduction is associated solely with heightened susceptibility to infection41,42. Instead, we uncovered a previously unrecognized intestine-specific adaptation that strengthens barrier immunity and limits pathogen entry and dissemination during this critical reproductive period.
Reproduction is orchestrated through substantial and intricate immune adaptations, many of which have been well characterized in the circulation and placenta during pregnancy. Progesterone-induced thymic regulatory T cells promote fetal tolerance43, whereas oestrogen-driven haematopoietic stem cell expansion supports erythropoiesis and prevents anaemia44. Furthermore, uterine natural killer cells remodel spiral arteries to enable implantation45, and pregnancy alters antibody glycosylation to enhance neonatal protection46. Our findings align with the broader view that reproduction is accompanied by systemic immune modulation, as evidenced by reduced cytokine production from lymphocytes. We also discovered that such immune modulation occurs in parallel with an enhancement of innate defence in the maternal gut. In the proximal small intestine, a primary site for nutrient uptake, an increase in eosinophil numbers drives goblet cell differentiation and mucus production, which fortifies the mucosal barrier and ultimately enhances innate defence. By limiting pathogen entry at the barrier, this adaptation may reduce the need for energy-demanding immune activation and thereby preserve maternal resources during reproduction.
Although features of maternal intestinal immune adaptations during lactation, including eosinophil accumulation, goblet cell expansion and epithelial remodelling, echo hallmarks of gastrointestinal helminth infections, the responses we uncovered occur in a fundamentally distinct context. That is, a physiological alliance between mother and offspring aimed at ensuring species survival rather than a defensive response to host–parasite confrontation. Notably, we observed heightened eosinophil activation and increased expression of Nmur1, a mediator of goblet cell induction during helminth infection21. These observations suggest that eosinophils co-opt conserved effector programs to induce goblet cells during both parasitic infection and reproduction. Indeed, our findings regarding eosinophil function in reproduction align with the ‘local immunity and remodelling’ hypothesis, which posits that eosinophils are intrinsically homeostatic cells that regulate immunity and drive tissue remodelling28,47. Our study supports this framework, establishing that eosinophils, in addition to propagating inflammatory responses during helminth infection, use similar effector mechanisms to enhance tissue integrity during physiological reproduction. However, the upstream signals that initiate and sustain eosinophil output and control how they are selectively recruited to the intestine during reproduction remain unclear. Elucidating these mechanisms will shed light on how physiological states dynamically reprogram local immunity to balance the needs for defence against pathogens and for immune tolerance during reproduction.
We also observed remodelling of both immune and epithelial compartments in the maternal intestine, with long-lasting innate defence evident even 12 weeks after cessation of lactation, a time frame nearly four times the length of the nursing period. This prolonged alteration suggests that sustained tissue remodelling can be imprinted not only by pathological exposures such as infection and inflammation48,49 but also by physiological reproduction. The observation that circulating eosinophils are elevated in postpartum women suggests that this immune adaptation may be conserved across mammals16. However, the duration and magnitude of such effects in longer-lived species, including humans, remain unknown. Longitudinal epidemiological studies examining gastrointestinal infection susceptibility in postpartum women will be important to determine whether similar physiological remodelling persists across reproductive cycles.
We propose that reproduction acts as a powerful conditioning event that reshapes maternal gut immunity. By priming tissue for heightened innate immunity, this sustained intestinal remodelling probably represents an evolutionarily selected strategy that enhances maternal resilience in pathogen-rich environments, as faced by wild animals and humans throughout evolutionary history, thus ensuring the survival of both mother and offspring.
Methods
Mice
All animal experiment were performed in accordance with protocol approved by the Institutional Animal Care and Use Committee and Institutional Biosafety Committee at Princeton University. C57BL/6 (stock no. 000664), Lgr5eGFP−IRES-creERT2 (stock no. 008875), BALB/cJ (stock no. 000651) and ΔdblGATA (stock no. 033551) mice were purchased from The Jackson Laboratory. Germ-free C57BL/6 mice were purchased from Taconic Biosciences. For timed pregnancy, females were intraperitoneally injected with 2 IU PMSG followed by 2 IU hCG 46–48 h later and then co-housed with male breeder mice. This hormone-induced timed pregnancy model was used for all experiments unless otherwise specified. For all experiments, C57BL/6 male breeder mice were used unless otherwise specified. Tissue collection and analyses were performed at embryonic day 12.5 for pregnant mice or 12 days postpartum for lactating mice unless otherwise specified. For long-term experiments, pups were weaned at 3 weeks and female mice were maintained for 12 weeks post-lactation. Age-matched mice that were administrated with hormones and co-housed with male mice but did not get pregnant were used in each experiment as nulliparous controls. Animals were randomly assigned to experimental groups whenever possible, and sample processing and outcome assessment were performed in a blinded manner where feasible.
Bacteria and infections
Wild-type Y. pseudotuberculosis IP32777 was grown overnight in 2× YT medium (Sigma-Aldrich) at 25 °C with shaking at 200 rpm. A nalidixic-acid-resistant strain of S. Typhimurium, IR715, was cultured in LB at 37 °C with shaking at 200 rpm. L. monocytogenes strain 10403s was cultured overnight in brain heat infusion broth (Teknova) at 37 °C with shaking at 200 rpm. Overnight cultures were centrifuged at 4,000g, resuspended in PBS and adjusted to a density (absorbance at 600 nm) of 1.0 in PBS. Mice fasted overnight (12–16 h) in new cages were orally administered 200 μl of the indicated pathogen suspension. To determine bacterial burden in the maternal spleen and faecal samples, spleens were homogenized through 70 μm filters with PBS, whereas faecal samples were collected in pre-weighed tubes and homogenized in PBS. For Y. pseudotuberculosis, homogenates were serially diluted and plated onto Yersinia-selective agar with Yersinia-selective supplement (Millipore), and colonies were counted after incubation at 25 °C for 48 h. For S. Typhimurium, homogenates were serially diluted and plated onto LB agar with 50 mg ml–1 nalidixic acid and incubated overnight at 37 °C. For L. monocytogenes, homogenates were serially diluted and plated onto brain heart infusion agar (Sigma Aldrich) with 100 mg ml–1 streptomycin and incubated overnight at 37 °C. All lactating dams were infected between day 5 and 8 postpartum period.
Tissue processing
Mice were euthanized with CO2. Tissues (small intestine, bone marrow, lung, skin and spleen) were collected into complete medium (RPMI 1640 supplemented with 20 mM HEPES, 2 mM l-glutamine, 1 mM sodium pyruvate, 1X nonessential amino acids, 55 μM β-mercaptoethanol, 100 U ml−1 penicillin and 100 μg ml–1 streptomycin) with 3% FBS on ice until further processing. For compartmentalization analysis, we defined the first 10 cm adjacent to the stomach as proximal, the last 10 cm as distal and the intervening segment as middle. For cell isolation from the small intestinal lamina propria, Peyer’s patches were removed and tissues were opened longitudinally, washed with cold PBS twice, cut into 1 cm segments and placed immediately into 3% complete medium containing 5 mM EDTA and 0.154 mg ml−1 (1 mM) dithiothreitol. Tissues were then incubated for 20 min at 37 °C with continuous stirring at 400 rpm, vigorously shaken in RPMI containing 20 mM HEPES, 2 mM EDTA, 100 U ml–1 penicillin and 100 μg ml–1 streptomycin for 1 min 3 times to remove any remaining mucus and further digested with 10 ml of 0% medium containing 0.5 mg ml–1 DNase I (Sigma-Aldrich) and 0.1 mg ml–1 Liberase TL (Roche) for 25 min at 37 °C with constant stirring at 400 rpm. Lungs were diced and incubated in 2 ml of prewarmed medium containing 0.5 mg ml−1 DNase I and 0.1 mg ml–1 collagenase IV for 35 min at 37 °C. To isolate cells from the skin, ear pinnae were excised and separated into ventral and dorsal sheets, digested by placing the dermal side down in medium containing 0.5 mg ml–1 DNase I with 0.25 mg ml−1 Liberase TL and incubated for 90 min at 37 °C. Digested intestines, lung and skin were passed through 70 μm cell strainers and centrifuged at 1,500 rpm for 5 min. Leukocytes were enriched by resuspending the samples in 4 ml of 37.5% Percoll and centrifuged at 1,800 rpm for 5 min. For blood analysis, 200 μl blood was added with 1 ml of ACK lysing buffer and incubate on ice for 10 min with occasional shaking. The reaction was then quenched with 5 ml of 3% medium followed by spinning at 1,800 rpm for 5 min. The pellet was then resuspended in 3% medium. Cells were washed with HBSS once before downstream staining and analyses.
Flow cytometry analysis
Cells isolated from tissues were plated into 96-well plates with counting beads. To measure cytokine production potential, cells were re-stimulated in complete IMDM supplemented with 10% FBS, 50 ng ml–1 PMA, 5 μg ml–1 ionomycin and 3 μg ml–1 BFA for 2.5 h at 37 °C. Cells were washed twice with cold HBSS before staining. Surface staining was performed on ice for 20 min, followed by washing with HBSS once and 10 min of Live/Dead staining. Cells were then washed twice with HBSS and fixed for 1 h at room temperature using either eBioscience intracellular fixation buffer (00-5123-43, Invitrogen) if intracellular staining was needed or BD Cytofix Fixation buffer (554722, BD Biosciences) for surface staining only. For intracellular staining, fixed cells were washed twice with the corresponding permeabilization buffer and intracellular staining was performed overnight at 4 °C. Before flow analysis, cells were washed twice and resuspended in 200 μl cold PBS. Flow cytometry analysis was performed on a FACSymphony A3.
The following antibodies were used for flow cytometry: anti-CD90-2 (FITC; 1:200; 53-2.1), anti-CD90-2 (BV786; 1:400;3 0-H12), anti-MHCII (BV650; 1:500; M5/114.15.2), anti-CD64 (BV605; 1:100; X54-5/7.1), anti-CD103 (BV510; 1:100; 2E7), anti-Ly6C (APCCy7; 1:200; HK1.4), anti-F4/80 (AF700; 1:200; BM8), anti-PD-L1 (PE; 1:100; 10 F.9G2), anti-TCRγ/δ (BV650; 1:200; GL3), anti-IFNγ (BV605; 1:200; XMG1.2), anti-CD4 (BV510; 1:200; RM4-5), anti-IL17A (BV421; 1:200; TC11-18H10.1), anti-T-bet (PECy7; 1:200; 4B10), anti-CD48 (BV786; 1:200; HM48-1), anti-CD45R/B220 (BV421; 1:400; RA3-6B2), anti-CD135 (APC; 1:200; A2F10), anti-CD150 (PECF594; 1:200; TC15-12F12.2), anti-FcεRIα (AF700; 1:200; MAR-1), anti-CCR3 (PECy7; 1:200; J073E5), anti-cKit (PECy7; 1:200; 2B8) and anti-CD200R3 (PE; 1:100; Ba13) were from BioLegend; anti-CD11c (BV786; 1:200; N418), anti-B220 (BUV805/PerCPe710; 1:200; RA3-6B2), anti-CD11b (BUV737; 1:100; M1/70), anti-CD45 (BUV395/APCCy7; 1:200/1,000; 30-F11), anti-FOXP3 (FITC; 1:400; FJK-16s), anti-TCRb (BUV737; 1:100; H57-597), anti-CD44 (AF700; 1:100; IM7), anti-GATA3 (APC; 1:200; TWAJ), anti-IL-5 (PE; 1:200; TRFK5), anti-IL-13 (PE; 1:200; eBio13A), anti-cKit (PerCPe710; 1:200; 2B8), anti-CD34 (AF488; 1:100; RAM34), anti-SCA1 (BV605; 1:400; D7), anti-CD16/CD32 (AF700; 1:200; 93) and anti-CD127 (PECy7; 1:200; A7R34) were from eBioscience; and anti-Ly6G (BV650/APC; 1:200; 1A8), anti-Siglec-F (PECF594/BV510; 1:200; E50-2440), anti-CD8b (BUV395; 1:400; H35-17.2), anti-RORγt (PECF594; 1:400; Q31-378), anti-CD125 (PE; 1:100; T21) and anti-CD90-2 (BUV805; 1:200; 30-H12) were from BD Bioscience.
Imaging of intestinal tissues
The small intestine was removed from the animal immediately after CO2 euthanasia and the first 10 cm was sectioned. The tissue was flushed with cold PBS using a gavage needle attached to a 20 ml syringe to remove all faeces and excess liquid was removed from inside. The tissue was dangled over a fix tray and 3 ml of fix (1 volume of Cytofix in 2 volumes of PBS) was flushed through the tissue. The tissue was kept on ice for 1 h and then laid straight on a blotting paper soaked with cold PBS. An L-shaped scissor was used to cut the tissue open lengthwise. The two ends of the tissue were sliced off, and the remaining tissue was rolled into a tight spiral with the distal end inside. The roll was then pierced with a 19 G needle and fixed with 2 ml cold fix in a 14-ml Falcon tube overnight at 4 °C. The next day, the roll was transferred to 30% sucrose in PBS and stored overnight at 4 °C. For embedding, the roll was transferred into a 14-ml Falcon tube with OCT compound and shaken at 200 rpm at room temperature for 4–8 h. The tissues were then removed from the needle and placed into moulds filled with OCT so that the entire tissue was covered with OCT and the upward face was a spiral. The mould was placed on a pre-chilled metal tray on dry ice to freeze immediately. After complete freezing, the blocks were kept at –80 °C for long-term storage. Sections were cut at 20 μm using a cryostat at –20 °C and stored at –80 °C for at least 24 h before staining.
For immunofluorescence staining, slides were equilibrated to room temperature and fixed in pre-chilled acetone for 10 min at –20 °C. An ImmEdge Hydrophobic Barrier PAP Pen (Vector Laboratory) was used to draw squares around the tissues and the slides were air dried at room temperature for 1 h. The tissues were rehydrated with cold PBS 3 times for 5 min each and blocked with base block buffer (1% BSA and 0.24% Triton X-100 in PBS) for 1 h at room temperature. Primary antibodies were diluted in the base block buffer and staining was performed either for 4 h at room temperature or overnight at 4 °C. The tissue samples were then washed with PBS twice, 5 min each, stained with DAPI in PBS for 30 min at room temperature and washed again with PBS for 3 times. Excess liquid was carefully removed from the slides using a Kimwipe. The slides were mounted with Prolong Gold Antifade reagent (ThermoFisher) and dried overnight before imaging on a Nikon CSU-W1. For measurement of goblet cell counts and villus and crypt lengths, at least ten crypt–villus units were measured for each mouse. For LGR5+ stem cell imaging, Lgr5eGFP-IRES-creERT2 mice were used. For MUC2 staining, 1.5 cm of the proximal and distal small intestine was cut off and put into Carnoy’s fixative at 4 °C overnight. The next day, tissue samples were transferred to 70% ethanol and placed at 4 °C for at least 24 h. Tissue samples were then processed on an ASP300S tissue processor and embedded in paraffin for long-term storage. Slides were sectioned at 7 μm. Slides were deparaffinized and incubated in 1× antigen retrieval buffer at 95 °C for 30 min and washed twice with PBS for 5 min each. Tissues were then permeabilized with 0.1% Triton X-100 in PBS for 10 min and blocked with blocking buffer (3% BSA and 0.1% Tween-20 in PBS) for 1 h at room temperature. Anti-MUC2 primary antibody was diluted in blocking buffer and applied to tissues overnight at 4 °C. The next day, tissue samples were washed with PBS twice, 5 min each, stained with fluorochrome-conjugated secondary antibody and DAPI in blocking buffer for 1 h at room temperature and washed again with PBS for 3 times. Mounting and imaging were performed as described above.
The following antibodies were used for confocal imaging. Anti-mouse antibodies were anti-EPCAM (AF647; 1:1,000; 118212; BioLegend), anti-MUC2[C3] (1:200; GTX100664; Genetex), anti-EPX (MM25-82.2.1; 1:1,000; by E. A. Jacobsen from Mayo Clinic). Other antibodies were anti-GFP (AF488; 1:400; A21311; Invitrogen) and UEA1 (Rhodamine; 1:200; RL-1062; Vector Laboratory). Secondary antibodies were goat anti-rabbit IgG H&L (AF594; 1:500; ab150080; Abcam).
Intestinal epithelial cell sorting
Small intestines were collected from CO2-euthanized animals and the first 10 cm was sectioned. The tissues were opened longitudinally, washed 2 times with 5% gut wash buffer (HBSS supplemented with 25 mM HEPES, 2 mM l-glutamine, 1 mM sodium pyruvate, 1X nonessential amino acids, 100 U ml–1 penicillin and 100 μg ml–1 streptomycin with 5% FBS) and cut into 2–5 mm pieces into 10 ml gut wash buffer containing 2 mM DTT. The tissue was incubated in a 37 °C water bath for 10 min, briefly shaken every 2 min and passed through a 100 μm filter after vigorous shaking 15 times. The tissue was then transferred to a new conical tube containing 10 ml gut wash buffer with 5 mM EDTA and incubated at 37 °C for 8 min with brief shaking every 2 min. The supernatant fraction was collected on ice by passing through a 100 μm filter and the tissue was transferred to a new conical tube to repeat the EDTA wash step for a total of 3 times. The second and third fractions were collected in the same tube as the first one. Cells were centrifuged at 1,000 rpm for 5 min at 4 °C and washed once with 0% DMEM (supplemented the same as gut wash buffer but with different FBS concentration, referred to as DMEM below). The cell pellet was incubated in 10 ml 0% DMEM supplemented with 0.1 U ml–1 Dispase and 0.1 mg ml–1 DNase in a 37 °C water bath for 8 min, with brief shaking every 2 min. After neutralization with 5 ml 2.5% DMEM, the cells were centrifuged at 1,000 rpm for 5 min and washed once with the staining buffer (same as the 5% gut wash buffer). Antibodies diluted in the staining buffer were added and staining was performed on ice for 20 min. The cells were washed with 10 volumes of 10% DMEM and resuspended in 10% DMEM for sorting. All cell sorting was performed on a Sony MA900 sorter. Before loading onto the sorter, the cell suspension was filtered using a 5 ml Falcon tube with a cell strainer cap and DAPI was added at a 1:400 concentration. Alive EPCAM+ (anti-EPCAM (AF594; 1:1,000; G8.8; BioLegend)) epithelial cells were sorted directly into 10% DMEM.
scRNA-seq and analysis
EPCAM+ IECs were sorted from 12–15-week-old nulliparous, pregnant or lactating mice as described in the previous section. For each group, cells were pooled from 3 mice and a total number of 25,000 cells per lane were loaded onto a Chromium Single Cell Controller. Libraries were prepared using Chromium Next GEM Single Cell 3′ Reagent kits v.3.1 (10x Genomics) following the manufacturer’s instructions and sequenced on a NovaSeq 6000 System. Cell by gene count matrices were generated for each sample using the count function from CellRanger v.7.1.0 (10x Genomics).
Data were analysed using Scanpy (v.1.8.2). Count matrices from the three samples were concatenated then filtered and processed together. Data were filtered to keep only cells with at least 10 expressed genes, 100 total counts and less than 15% of total counts coming from mitochondrial genes. Genes were kept if they were expressed in at least 1% of filtered cells (211 cells) and had at least 250 total counts across cells.
For clustering and cell-type assignment, the count matrix was normalized by Pearson residuals (θ = 1) and then used for principal component analysis (100 components), k-nearest neighbour graph construction (k = 30, number of principal components = 50) and Leiden clustering. Clusters were annotated on the basis of the expression of known marker genes. In some cases, clusters were further subclustered and then annotated by cell-type identity. Cells that were potentially doublets (expressing marker genes from multiple cell types) or unknown were removed before downstream analyses.
For differential gene expression, the raw count matrix was normalized by library size and then log-transformed. For each cell type, differential expression was performed between nulliparous versus pregnancy and nulliparous versus lactating conditions. Genes were filtered out if they were not expressed by at least 5% of cells in either condition. For remaining genes, log fold change values were calculated as the difference between the mean expression values of log-library size normalized counts for the compared conditions. P values were calculated using a Mann–Whitney U-test and corrected for multiple hypothesis with the Benjamini–Hochberg procedure.
Spatial transcriptomic analysis using Xenium Prime Mouse 5K
The first 10 cm of the small intestine was collected from mice, fresh-frozen with isopentane and embedded in OCT. Tissues were cryosectioned at 10 μm, mounted on Xenium slides and processed according to the Xenium Prime 5K protocol (10x Genomics). Xenium datasets were generated using a Xenium Analyzer (instrument software v.3.1.0.0 and analysis v.xenium-3.1.0.4). Tissue slides were also stained with Xenium MultiTissue Stain mix for multimodal cell segmentation, and this segmentation from Xenium Analyzer was used for all analyses. Post-Xenium Analyzer haematoxylin and eosin staining was performed according to the official demonstrated protocol.
Transcripts for each cell were filtered to keep only transcripts overlapping the nucleus segmentation for each cell. Cells from all samples were concatenated and the subsequent count matrix was further filtered to keep only cells with counts from at least ten genes and with exactly one segmented nucleus. Data were analysed using Scanpy (v.1.10.4). The count matrix was normalized by library size, log-transformed and then z-scored across genes. Normalized data were used for principal component analysis (50 components), k-nearest neighbour graph construction (k = 15) and Leiden clustering. Each cluster was then separately clustered into more fine-grained clusters. A count matrix for the same cells but for all transcripts, including non-nuclear transcripts, was also generated and normalized as described above. These counts were used to confirm the expression patterns for the nuclear-only transcript counts.
Clusters and subclusters were manually annotated on the basis of the expression of known marker genes. Cluster-specific differentially expressed genes were identified by calculating log fold change values of the mean expression between normalized counts for cells included and excluded in each cluster. For this analysis, counts were normalized by library size only. P values were calculated using a Mann–Whitney U-test and corrected for multiple hypothesis testing with the Benjamini–Hochberg procedure. Myeloid clusters were identified on the basis of Cd45 and Cd11b expression, and eosinophils were further labelled on the basis of Il5ra and Ccr3 expression in the myeloid component. Differentially expressed genes between nulliparous and lactating conditions for eosinophils were identified using DESeq2 with a pseudobulk approach50. 10x Genomics Xenium Explorer (v.3.2.0) was used to visualize cell and transcript location and to generate spatial images of cell-type identity overlaying DAPI staining.
Swiss rolls were computationally unrolled into the proximal-distal axis and normalized crypt–villus axis using a previously described approach51. In brief, for each sample, the base of the small intestine roll was manually annotated, and the nearest location on the base spiral was calculated for each cell. The relative position of this nearest location was used as the proximal–distal position and the distance to this nearest location was used as the crypt–villi position, normalized by local maxima to be between 0 and 1. For plotting kernel density estimates of the distribution of cell counts along the proximal–distal axis and crypt–villus axis, we applied a Gaussian KDE (bandwidth determined using Scott’s rule). The KDE was scaled so that the total area under the curve equals the total cell count using a grid size of 500 bins. An individual KDE was calculated for each sample, and the mean and standard deviation of these KDE curves are shown. Neighbouring cell analysis was performed by identifying the directly adjacent spatial neighbours for each cell by using cell segmentation boundaries within 2 μm of each other.
Organoid culture and imaging
Epithelial cells were isolated and sorted from Lgr5eGFP-IRES-creERT2 mice using a MA900 cell sorter as described in the previous section. Alive EPCAM+LGR5+ cells were gated as shown in Extended Data Fig. 5b and directly sorted into cold complete organoid growth medium (OGM) and centrifuged at 400g for 8 min. The cell pellet was then resuspended in complete OGM at a concentration of 40 cells per μl and mixed with 1 volume of undiluted Matrigel matrix. Next, 10 μl of the mixture was quickly pipetted into the centre of each well of a 96-well plate. The plate was immediately placed into a 37 °C incubator for 10 min for the Matrigel to set, and 100 μl pre-warmed complete OGM supplemented with 10 μM Y-27632 was added to each well. The medium was changed every other day and Y-27632 was only supplemented for the first 3 days. On day 8, plates were centrifuged at 3,000 rpm for 10 min at 10 °C to spin the organoids down. Organoids were then fixed in 4% paraformaldehyde and PBS overnight at 4 °C and washed with PBS 3 times before staining. Organoids were dehydrated with 30% sucrose in PBS for 30 min at room temperature and permeabilized with permeabilization buffer (0.1% Triton X-100 and 3% BSA in PBS) for 1 h at room temperature. Primary staining was performed using the permeabilization buffer either for 4 h at room temperature or overnight at 4 °C. Organoids were washed twice with cold PBS, stained with DAPI for 30 min at room temperature and then washed again with PBS 3 times. Next, 50 μl CytoVista 3D Cell Culture Clearing reagent (Invitrogen) was added to each well and the plate was placed at 4 °C for 10 min before imaging on a Nikon CSU-W1. Clonogenicity was assessed on day 8 by quantifying the percentage of organoid formed out of the number of LGR5+ intestinal stem cell initial seeded.
Eosinophil isolation and ex vivo culture
Intestinal lamina propria was prepared as described in the previous section. Alive CD45+CD11b+Siglec-F+ eosinophils (anti-Siglec-F (PE; 1:200; E50-2440; BD), anti-CD11b (AF647; 1:100; M1/70; BioLegend) and anti-CD45 (APC-eF780; 1:200; 30-F11; eBioscience)) were directly sorted into cold complete RPMI medium supplemented with 20% FBS. Cells were centrifuged at 400g for 8 min and counted so that 30,000 eosinophils were resuspended in 65 μl 20% RPMI and plated in each well of a 96-well plate. Next, 65 μl of 20% RPMI supplemented with 20 ng ml–1 IL-5 was overlaid, and the plate was gently swirled before being placed into an incubator set at 37 °C with 5% CO2. After 48 h, the plate was centrifuged at 1,600 rpm for 5 min at 4 °C. A volume of 120 μl supernatant was collected from each well and stored at –80 °C until analysis. Cytokine and growth factors were measured using multiplex assay kits.
Blockade of eosinophil migration using anti-CCR3 antibody
Timed-pregnant mice were retro-orbitally injected with 1 mg of anti-mouse CCR3 antibody (BE0316; BioXCell) or rat IgG2b isotype control weekly, beginning from gestational day 10.5 until day 10 postpartum for a total of 4 doses and analysed at day 12 postpartum. For infection analysis, mice received a total of 3 doses before the analysis. Small intestine samples were collected and analysed by flow cytometry to confirm successful blockade of eosinophils.
Eosinophil chemotaxis assay using Transwell
Mice were treated with 1 mg isotype or anti-CCR3 antibody. At 24 h after treatment, alive CD45+CD11b+Siglec-F+ eosinophils were isolated from bone marrow and resuspended at a concentration of 2 × 105 cells per ml in 20% RPMI medium without rmIL-5. In vitro chemotaxis assays were performed in HTS Transwell plates (Corning) with a 5 µm pore size. Recombinant mouse eotaxin-1 and eotaxin-2 (R&D) were prepared in a concentration of 100 ng ml–1 using the same medium. For cell migration assays, 100 µl cell suspension was added to the upper well and 100 µl medium with or without eotaxins was added to the bottom well, followed by 1 h of incubation at 37 °C. Migrated eosinophils were collected from the bottom compartment, analysed by flow cytometric counting and normalized to no-eotaxin control.
Morphometric analysis for the mucus layer
For the quantitative analysis of the mucus area present in the duodenum tissue, a previously described method52 was used. PAS and Alcian Blue-stained histological sections were visualized and scanned using a Scan Aperio slide scanner (Leica Biosystems) at ×40 magnification. For each mouse, the entire duodenum mucosa was analysed for the quantification of the mucin area (PAS and Alcian Blue-positive) with the aid of QuPath software (https://qupath.github.io)53. The total length of each section was manually measured in micrometres to determine the actual proportion of the duodenal surface analysed for each animal. Then, the mucus area for each animal was normalized to the respective duodenum mucosa length. The methodology used for imaging and morphometric conditions was adapted from a previously published method54.
Measurement of gut permeability
Intestinal permeability was measured by performing oral gavage of 4 kDa FITC-labelled dextran (Sigma-Aldrich) and measuring fluorescent translocation into the plasma. Mice were fasted for 4 h before being orally gavaged with 200 μl of 60 mg ml–1 (12 mg) FITC-dextran. Two hours later, blood was retro-orbitally collected and centrifuged at room temperature at 12,000g for 15 min to obtain the plasma. Fluorescence intensity was then measured on fluorescence plates using an excitation wavelength of 485 nm and an emission wavelength of 535 nm.
Statistical analysis
Two-tailed unpaired Student’s t-tests, one-way ANOVA and two-way ANOVA were performed to compare the results using GraphPad Prism (v.10). No samples were excluded from analyses. We treated P < 0.05 as significantly different, with *P < 0.05, **P < 0.01, ***P < 0.001 and ****P < 0.0001. Details regarding the exact sample size for each experimental group and condition and representative data are presented in the figure legends.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Single-cell and spatial datasets generated during this study have been deposited in the Gene Expression Omnibus with the accession numbers GSE296052 (scRNA-seq) and GSE296053 (Xenium). Source data are provided with this paper.
Code availability
The code used to process and analyse the scRNA and Xenium data is available at GitHub (https://github.com/pritykinlab/eosinophil-intestinal-remodeling-analysis).
References
Somerset, D. A., Zheng, Y., Kilby, M. D., Sansom, D. M. & Drayson, M. T. Normal human pregnancy is associated with an elevation in the immune suppressive CD25+ CD4+ regulatory T-cell subset. Immunology 112, 38–43 (2004).
Samstein, R. M., Josefowicz, S. Z., Arvey, A., Treuting, P. M. & Rudensky, A. Y. Extrathymic generation of regulatory T cells in placental mammals mitigates maternal–fetal conflict. Cell 150, 29–38 (2012).
Klion, A. D. & Nutman, T. B. The role of eosinophils in host defense against helminth parasites. J. Allergy Clin. Immunol. 113, 30–37 (2004).
Arnold, I. C. & Munitz, A. Spatial adaptation of eosinophils and their emerging roles in homeostasis, infection and disease. Nat. Rev. Immunol. 24, 858–877 (2024).
Abu-Raya, B., Michalski, C., Sadarangani, M. & Lavoie, P. M. Maternal immunological adaptation during normal pregnancy. Front. Immunol. 11, 575197 (2020).
Aghaeepour, N. et al. An immune clock of human pregnancy. Sci. Immunol. 2, eaan2946 (2017).
Li, J., Xiao, C., Li, C. & He, J. Tissue-resident immune cells: from defining characteristics to roles in diseases. Signal Transduct. Target. Ther. 10, 12 (2025).
Niec, R. E., Rudensky, A. Y. & Fuchs, E. Inflammatory adaptation in barrier tissues. Cell 184, 3361–3375 (2021).
Farber, D. L. Tissues, not blood, are where immune cells function. Nature 593, 506–509 (2021).
Weisberg, S. P., Ural, B. B. & Farber, D. L. Tissue-specific immunity for a changing world. Cell 184, 1517–1529 (2021).
Pelletier, G., de Passillé, A. M., Cardou, M. B. & Morisset, J. Influence of pregnancy, lactation, litter size and diet energy density on the pancreas of sows. J. Nutr. 116, 1480–1488 (1986).
Fell, B. F., Campbell, R. M. & Boyne, R. Observations on the morphology and nitrogen content of the alimentary canal in breeding hill sheep. Res. Vet. Sci. 5, 175–189 (1964).
Johnson, M. L., Saffrey, M. J. & Taylor, V. J. Gastrointestinal capacity, gut hormones and appetite change during rat pregnancy and lactation. Reproduction 157, 431–443 (2019).
Onji, M. et al. RANK drives structured intestinal epithelial expansion during pregnancy. Nature 637, 156–166 (2025).
Ameku, T. et al. Growth of the maternal intestine during reproduction. Cell 188 2738–2756.e22 (2025).
Marcus, M. B., Cibley, L. J., Brandt, M. L., Millman, L. & Barlas, D. Circulating eosinophils in labor and puerperium. Am. J. Obstet. Gynecol. 75, 11–15 (1958).
Knight, C. H., Maltz, E. & Docherty, A. H. Milk yield and composition in mice: effects of litter size and lactation number. Comp. Biochem. Physiol. A Comp. Physiol. 84, 127–133 (1986).
Kutyavin, V. I., Korn, L. L. & Medzhitov, R. Nutrient-derived signals regulate eosinophil adaptation to the small intestine. Proc. Natl Acad. Sci. USA 121, e2316446121 (2024).
Diny, N. L. et al. The aryl hydrocarbon receptor contributes to tissue adaptation of intestinal eosinophils in mice. J. Exp. Med. 219, e20210970 (2022).
Wang, W. L. et al. The aryl hydrocarbon receptor instructs the immunomodulatory profile of a subset of Clec4a4+ eosinophils unique to the small intestine. Proc. Natl Acad. Sci. USA 119, e2204557119 (2022).
Li, Y. et al. Neuromedin U programs eosinophils to promote mucosal immunity of the small intestine. Science 381, 1189–1196 (2023).
Gurtner, A. et al. Active eosinophils regulate host defence and immune responses in colitis. Nature 615, 151–157 (2023).
Wen, T. et al. The pan-B cell marker CD22 is expressed on gastrointestinal eosinophils and negatively regulates tissue eosinophilia. J. Immunol. 188, 1075–1082 (2012).
Agace, W. W. & McCoy, K. D. Regionalized development and maintenance of the intestinal adaptive immune landscape. Immunity 46, 532–548 (2017).
Mowat, A. M. & Agace, W. W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 14, 667–685 (2014).
Esterházy, D. et al. Compartmentalized gut lymph node drainage dictates adaptive immune responses. Nature 569, 126–130 (2019).
Zwick, R. K. et al. Epithelial zonation along the mouse and human small intestine defines five discrete metabolic domains. Nat. Cell Biol. 26, 250–262 (2024).
Lee, J. J., Jacobsen, E. A., McGarry, M. P., Schleimer, R. P. & Lee, N. A. Eosinophils in health and disease: the LIAR hypothesis. Clin. Exp. Allergy 40, 563–575 (2010).
Gazzinelli-Guimaraes, P. H., Jones, S. M., Voehringer, D., Mayer-Barber, K. D. & Samarasinghe, A. E. Eosinophils as modulators of host defense during parasitic, fungal, bacterial, and viral infections. J. Leukoc. Biol. 116, 1301–1323 (2024).
Gurtner, A., Crepaz, D. & Arnold, I. C. Emerging functions of tissue-resident eosinophils. J. Exp. Med. 220, e20221435 (2023).
Ignacio, A. et al. Small intestinal resident eosinophils maintain gut homeostasis following microbial colonization. Immunity 55, 1250–1267 (2022).
Jung, Y. et al. IL-1β in eosinophil-mediated small intestinal homeostasis and IgA production. Mucosal Immunol. 8, 930–942 (2015).
Marillier, R. G. et al. IL-4/IL-13 independent goblet cell hyperplasia in experimental helminth infections. BMC Immunol. 9, 11 (2008).
Herbert, D. R. et al. Intestinal epithelial cell secretion of RELM-β protects against gastrointestinal worm infection. J. Exp. Med. 206, 2947–2957 (2009).
Yu, C. et al. Targeted deletion of a high-affinity GATA-binding site in the GATA-1 promoter leads to selective loss of the eosinophil lineage in vivo. J. Exp. Med. 195, 1387–1395 (2002).
Gustafsson, J. K. & Johansson, M. E. V. The role of goblet cells and mucus in intestinal homeostasis. Nat. Rev. Gastroenterol. Hepatol. 19, 785–803 (2022).
Nyström, E. E. L. et al. An intercrypt subpopulation of goblet cells is essential for colonic mucus barrier function. Science 372, eabb1590 (2021).
Sorobetea, D. et al. Inflammatory monocytes promote granuloma control of Yersinia infection. Nat. Microbiol. 8, 666–678 (2023).
Velazquez, E. M. et al. Endogenous Enterobacteriaceae underlie variation in susceptibility to Salmonella infection. Nat. Microbiol. 4, 1057–1064 (2019).
Canul-Medina, G. & Fernandez-Mejia, C. Morphological, hormonal, and molecular changes in different maternal tissues during lactation and post-lactation. J. Physiol. Sci. 69, 825–835 (2019).
Sappenfield, E., Jamieson, D. J. & Kourtis, A. P. Pregnancy and susceptibility to infectious diseases. Infect. Dis. Obstet. Gynecol. 2013, 752852 (2013).
Billington, W. D. The immunological problem of pregnancy: 50 years with the hope of progress. A tribute to Peter Medawar. J. Reprod. Immunol. 60, 1–11 (2003).
Paolino, M. et al. RANK links thymic regulatory T cells to fetal loss and gestational diabetes in pregnancy. Nature 589, 442–447 (2021).
Nakada, D. et al. Oestrogen increases haematopoietic stem-cell self-renewal in females and during pregnancy. Nature 505, 555–558 (2014).
Robson, A. et al. Uterine natural killer cells initiate spiral artery remodeling in human pregnancy. FASEB J. 26, 4876–4885 (2012).
Erickson, J. J. et al. Pregnancy enables antibody protection against intracellular infection. Nature 606, 769–775 (2022).
Shah, K., Ignacio, A., McCoy, K. D. & Harris, N. L. The emerging roles of eosinophils in mucosal homeostasis. Mucosal Immunol. 13, 574–583 (2020).
Naik, S. & Fuchs, E. Inflammatory memory and tissue adaptation in sickness and in health. Nature 607, 249–255 (2022).
Ordovas-Montanes, J., Beyaz, S., Rakoff-Nahoum, S. & Shalek, A. K. Distribution and storage of inflammatory memory in barrier tissues. Nat. Rev. Immunol. 20, 308–320 (2020).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Reina-Campos, M. et al. Tissue-resident memory CD8 T cell diversity is spatiotemporally imprinted. Nature 639, 483–492 (2025).
Sardinha-Silva, A. et al. Giardia-induced type 2 mucosal immunity attenuates intestinal inflammation caused by co-infection or colitis in mice. Nat. Microbiol. 10, 1886–1901 (2025).
Bankhead, P. et al. QuPath: open source software for digital pathology image analysis. Sci. Rep. 7, 16878 (2017).
Oliveira, F. M. et al. Susceptibility to Entamoeba histolytica intestinal infection is related to reduction in natural killer T-lymphocytes in C57BL/6 mice. Infect. Dis. Rep. 4, e27 (2012).
Acknowledgements
We thank members of the Lim Laboratory for discussion and constructive feedback; staff at Princeton University for their continuous assistance and staff at the Molecular Biology Flow Cytometry Resource Facility, confocal imaging facility and the Lewis-Sigler Institute Genomics Core Facility and Laboratory Animal Resources for technical support. Flow data were acquired using a BD Biosciences FACSymphony A3 flow cytometer, purchased with NIH S10 award 1S10OD028592-01A1. This project is partially funded by the Princeton Catalysis Initiative. A.I.L. is a Freeman Hrabowski Scholar from the Howard Hughes Medical Institute and is supported by the Searle Scholars Program and The Branco Weiss Fellowship. C.H. is supported by a fellowship provided by the China Scholarship Council, based on the April 2015 Memorandum of Understanding between the CSC and Princeton University. J.R. received a Cancer Research Institute Irvington postdoctoral fellowship. K.H.A.F. received a Pew Latin American Fellowship funded by The Pew Charitable Trust and the Princeton Molecular Biology Postdoctoral Research Fellowship. T.R.C. was supported by MLMB training grant T32GM148739. Y.P. was supported by NIH grant DP2AI171161 and the Ludwig Institute for Cancer Research.
Author information
Authors and Affiliations
Contributions
A.I.L. conceived the project, secured funding and is the corresponding author. C.H. and A.I.L. led the design of all experiments, assembled the figures, analysed the data and wrote the manuscript with input from all authors. C.H. performed all the experiments with help from J.R., J.R.d.S., T.R.C. and K.H.A.F. A.S. performed scRNA-seq and spatial transcriptomic computational analyses under supervision by Y.P., with help from Y.Q. P.G.-G. and F.M.S.O. provided expertise in eosinophils and assisted with experiments.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Rodney Newberry, Sing Sing Way and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Reproduction induces alternations in myeloid and lymphoid populations.
a,b, Flow cytometry plots depict the gating scheme of myelocyte (a) and lymphocyte (b) subsets in siLP and are applicable to other organs including bone marrow, spleen, lung, and ear skin. c, Absolute numbers of myelocyte and lymphocyte subsets in siLP of female nulliparous, pregnant and lactating mice; n = 4. Data are representative of 3 independent experiments. Each dot represents one individual mouse. Error bars represent mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001 (one-way ANOVA).
Extended Data Fig. 2 Changes in immune functional profiles during lactation.
a, Representative histogram plots showing the expression level of PD-L1 and CD22 in eosinophils from siLP of nulliparous and lactating mice. Dashed lines show the gating threshold for positive populations; numbers stand for the frequency of PD-L1+ and CD22+ subsets. b, Single-channel grey-scale images for representative confocal images of proximal small intestine stained for eosinophils (EPX) and epithelial cells (EPCAM). Scale bar = 50 μm. c, Average distribution of EPX+ eosinophils along the crypt-villus axis; Control (n = 3), Lactating (n = 4). d, Counts of EPX+ eosinophils normalized by lamina propria area in small intestines; Control (n = 3), Lactating (n = 4). (e,f), Representative histogram plots showing the expression level of functional markers in eosinophils from siLP. e, Granularity (SSC-A). f, Markers for secretory activities (CD9, CD63 and CD107a). g, Eosinophils were isolated from nulliparous and lactating females and cultured ex vivo for 48 h. Plot showing levels of canonical eosinophil-associated cytokines measured from the cell culture supernatant; Control (n = 14), Lactating (n = 7 (IL-13) and 8 (IL-4/18/10)). h, Flow cytometry plots depict the gating scheme of eosinophil precursors (Live Lin- Sca-1− c-Kit+ CD34+ CD16/32+ CD125+) in bone marrow. i, Absolute eosinophil numbers in the bone marrow and siLP of nulliparous or lactating mice that reproduce without any administration of exogenous hormones; Control (n = 4), Lactating (n = 7). j, Absolute eosinophil numbers in the bone marrow and siLP of germ-free nulliparous or lactating mice; Control (n = 4), Lactating (n = 5 (bone marrow) and 6 (siLP)). (k–m), Representative histogram plots showing expression levels of canonical cytokines in ILCs, T helper cells and CD8+ T cells from the siLP of nulliparous and lactating females. a, IFN-γ production in ILC1, IL-5/IL-13 production in ILC2, and IL-17A production in ILC3. b, IFN-γ production in Th1, IL-5/IL-13 production in Th2, and IL-17A production in Th17. c, IFN-γ production in CD8+ T cells. Dashed lines show gating threshold for cytokine-producing populations; numbers stand for their frequency. Data are representative or pooled from at least two independent experiments. Each dot represents one individual mouse. Error bars and numbers in representative flow plots indicate mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001 (two-tailed unpaired Student’s t test).
Extended Data Fig. 3 Intestinal growth during reproduction.
a, Left: representative image of small intestine from female nulliparous, pregnant, and lactating mice. Right: bar graph showing length of small intestines; Control (n = 40), Pregnant (n = 24), Lactating (n = 31). b, Absolute numbers of total immune cells and eosinophils in siLP of nulliparous, pregnant and lactating mice, normalized by tissue weight; Control (n = 9), Pregnant (n = 5), Lactating (n = 8). c, Absolute numbers of total immune cells and various cell types in siLP of nulliparous, pregnant and lactating mice, normalized by small intestinal length; Control (n = 8), Pregnant (n = 7), Lactating (n = 7). d, Representative confocal image of views of individual villus stained for nuclei (DAPI) and epithelial cells (EPCAM) in the proximal small intestine. Scale bar = 50 μm. e, Bar graph showing average length of intestinal crypt and villus within the proximal small intestine. At least 30 villi/crypts were measured per mouse; n = 3. Data are representative or pooled from at least two independent experiments. Each dot represents one individual mouse. Error bars represent mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001 (one-way ANOVA).
Extended Data Fig. 4 Altered intestinal epithelium during lactation.
a, Heatmap showing the six major Epcam+ epithelial cell type annotations from the first 10 cm of proximal small intestine. b, Single-channel grey-scale images for the representative confocal images of stem cells in duodenal crypts. Scale bar = 100 μm. c, Single-channel grey-scale images for the representative confocal images of goblet cells in duodenal villi. Scale bar = 50 μm. d, UMAP analysis showing intestinal stem cells from the nulliparous and lactating females forming two distinct clusters. e, Representative contour plots showing gating scheme for EpCAM+Lgr5+ intestinal stem cells from the proximal small intestine of nulliparous and lactating heterozygous Lgr5EGFP-IRES-CreERT2 female mice. f, Single-channel grey-scale images for the representative confocal images of day-8 organoids, with staining of Goblet cells (UEA1) and epithelial cells (EPCAM). Scale bar = 50 μm. g, Left: representative confocal images of proximal intestinal villi, stained for goblet cells (MUC2) and epithelial cells (EPCAM). Scale bar = 50 μm. Right: bar graph showing the average count of MUC2+ goblet cells per villus within the proximal small intestine; n = 3. h, Left: representative confocal images of distal intestinal villi, stained for goblet cells (MUC2) and epithelial cells (EPCAM). Scale bar = 50 μm. Right: bar graph showing the average count of MUC2+ goblet cells per villus within the distal small intestine; Control (n = 4), Lactating (n = 3). i, Absolute numbers of eosinophils in the small intestines of nulliparous or lactating B6 female mice mated with B6 (syngeneic) or BALB/c (allogeneic) males; Control (n = 3), Syngeneic (n = 4), Allogeneic (n = 6). j, Average count of MUC2+ goblet cells per villus within the proximal small intestine of nulliparous or lactating B6 female mice mated with B6 (syngeneic) or BALB/c (allogeneic) males; Control (n = 3), Syngeneic (n = 3), Allogeneic (n = 5). For (g–j), each dot represents one individual mouse. At least 10 villi were measured per mouse. Error bars represent mean ± SD. *P < 0.05; **P < 0.01; ****P < 0.0001. For (g,h), two-tailed unpaired Student’s t test was used; for (i-j), one-way ANOVA was used.
Extended Data Fig. 5 Anti-CCR3 treatment abolished eosinophil accumulation in small intestine.
a, Absolute numbers of eosinophils in 100 μl blood from nulliparous and lactating mice; Control (n = 6), Lactating (n = 7). b, Representative contour plots showing the frequencies of Siglec-F+CCR3+ cells among all alive CD45+ cells in the blood of nulliparous and lactating mice. c, Left: representative contour plots showing CCR3+ eosinophil frequencies in the bone marrow. Right: bar plots showing CCR3+ eosinophil frequencies; Control (n = 8), Lactating (n = 9). d, Bone marrow eosinophils isolated from isotype- or anti-CCR3-treated mice were used to measure chemotaxis towards eotaxin-1 and -2 in vitro. Left: schematic illustration of the experimental design. Right: bar plots showing the migration of eosinophils in the presence of a mouse eotaxin-1 and -2 gradient, determined as the fold change in migrated cell numbers over control (no eotaxin); n = 4. (e,f), Timed-pregnant dams were injected with isotype control or 1 mg of anti-CCR3 weekly, starting from E11.5 and analyzed at day 12 of lactation. e, Bar graphs showing eosinophil absolute numbers in bone marrow, blood and siLP; Nulliparous isotype (n = 3), Nulliparous α-CCR3 (n = 5), Lactating isotype (n = 4), Lactating α-CCR3 (n = 5). f, Absolute numbers of other major myeloid cell populations and lymphoid cells in siLP; Nulliparous isotype (n = 3), Nulliparous α-CCR3 (n = 5), Lactating isotype (n = 4), Lactating α-CCR3 (n = 5). Data are representative or pooled from at least two independent experiments. Each dot represents one individual mouse. Error bars and numbers in representative flow plots indicate mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001 (one-way ANOVA). For (c,d), two-tailed unpaired Student’s t test was used; for (e,f), one-way ANOVA was used. Icons in d created in BioRender; Huang, C. https://BioRender.com/xo3xcgc (2026).
Extended Data Fig. 6 Anti-CCR3 treatment have minimal influence on pregnancy outcomes, but reduced goblet cell induction in the small intestine.
a, Left: images of fetus at ~E18.5 in isotype- or anti-CCR3-treated pregnant mice. Red arrows indicate absorbed fetus. Right: Bar plots showing fetal numbers; n = 4. b, Small intestinal length; Control (n = 3), Isotype (n = 6), α-CCR3 (n = 5). c, Villus length; Control (n = 3), Isotype (n = 6), α-CCR3 (n = 6). d, Dam and pup weight curves. e, Single-channel grey-scale images for the representative confocal images of villus-resident UEA1+ goblet cells. Scale bar = 70 μm. f, Single-channel grey-scale images for the representative confocal images of organoids derived from single Lgr5+ ISCs. Scale bar = 50 μm. Data are representative or pooled from at least two independent experiments. Each dot represents one individual mouse except for kinetic plot (d). Error bars represent mean ± SD. *P < 0.05 (one-way ANOVA).
Extended Data Fig. 7 Intestinal immune landscape of ΔdblGATA mice.
a, Absolute numbers of major myeloid cell populations in siLP; WT Nulliparous (n = 3), WT Lactating (n = 3), ΔdblGATA Nulliparous (n = 5), ΔdblGATA Lactating (n = 4). b, Absolute numbers of major lymphoid cell populations in siLP; WT Nulliparous (n = 3), WT Lactating (n = 3), ΔdblGATA Nulliparous (n = 5), ΔdblGATA Lactating (n = 4). c, Small intestinal length of wild type and ΔdblGATA mice under nulliparous or lactating status; WT Nulliparous (n = 3), WT Lactating (n = 3), ΔdblGATA Nulliparous (n = 5), ΔdblGATA Lactating (n = 5). d, Pup numbers of wild type and ΔdblGATA mice; WT (n = 3), ΔdblGATA (n = 5). c, Data are representative or pooled from at least two independent experiments. Each dot represents one individual mouse. Error bars represent mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001 (two-tailed unpaired student’s t test).
Extended Data Fig. 8 Spatial transcriptomic analysis revealed diverse epithelial and immune populations across the proximal small intestine.
a, Overview of the rolled first 10 cm of small intestine samples with H&E staining. Scale bar = 1000 μm. b, Left: Total cell numbers detected in the tissue samples; Right: UMAP showing the four categories of Cd45+ immune cells, Epcam+ epithelial cells, stromal cells and other cells. c, Left: UMAP plot showing clustering within the Epcam+ epithelial cells, color-coded by cell types; Right: Heatmap showing z-score normalized mean gene expression of cell type annotations within the Epcam+ epithelial cells. d, Left: UMAP plot showing clustering within the Cd45+ immune cells, color-coded by cell types; Right: Heatmap showing z-score normalized mean gene expression of cell type annotations within the Cd45+ cells. e, Representative flow histograms of CCR3 expression levels in CD11b+SiglecF+ eosinophils and CD11b+SiglecF− counterparts under the Live CD45+ gating. f, Total eosinophil numbers detected in the tissue samples. g, Bar plots showing the top 5 cell types frequently in direct proximity to stem cells. h, Bar plots showing the top 5 cell types frequently in direct proximity to eosinophils. i, Left: Mean distribution of four eosinophil subsets plotted across 500 bins of the normalized crypt-villus longitude, with shaded standard deviation within each condition; Right: absolute cell numbers of each subcluster in control and lactating samples. j, Heatmap showing some immune-regulatory and collagen genes enriched in active eosinophils. N = 4 biological replicates pooled. Error bars represent mean ± SD. For (b, i), two-tailed unpaired student’s t test was used. * P < 0.05; *** P < 0.001.
Extended Data Fig. 9 Protection against enteric infections is established during active lactation.
a–d, 5-day postpartum lactating dams and age-matched nulliparous controls were orally infected with Y. pseudotuberculosis (Yptb). a, Left: Representative pseudo-colored images showing mucus (red) detected by AB-PAS-stained proximal small intestine. Scale bar = 50 μm. Right: Bar blots showing normalized mucus area in the first 1.5 cm of proximal small intestine; PBS Control (n = 3), PBS Lactating (n = 4), Yptb Control (n = 4), Yptb Lactating (n = 4). b, Absolute cell numbers of major myeloid cell populations in siLP; PBS Control (n = 4), PBS Lactating (n = 3), Yptb Control (n = 5), Yptb Lactating (n = 7). c, Representative histograms showing expression levels of PD-L1 and granularity (SSC-A) in siLP eosinophils. d, Dams’ and pups’ bodyweight curve following Y. pseudotuberculosis (Yptb) infection; Dam Control (n = 3), Dam Infected (n = 5), Pup Control (n = 11), Pup Infected (n = 12). e, Absolute numbers of bone marrow and siLP eosinophils in age-matched nulliparous, pregnant, lactating and non-lactating mice; Bone Marrow Nulliparous (n = 12), Pregnant (n = 10), Lactating (n = 10), Non-lactating (n = 15); siLP Nulliparous (n = 12), Pregnant (n = 10), Lactating (n = 9), Non-lactating (n = 15). f, Average counts of MUC2+ goblet cells per villus in the four groups; Nulliparous (n = 4), Pregnant (n = 5), Lactating (n = 4), Non-lactating (n = 4). g,h, Pregnant mice at gestational day 12 and age-matched nulliparous controls were orally infected with Y. pseudotuberculosis. g, Left: fecal pathogen burden at 6 h post-infection; Control (n = 12), Pregnant (n = 11). Right: kinetics of fecal pathogen burden; Control (n = 12, 11, 10 (day1-3)), Pregnant (n = 11 (day1-3)). h, Pathogen dissemination at day 3 post-infection; Control (n = 11), Pregnant (n = 10). i,j, Non-lactating mice 12 days post-postpartum (pups removed from day 1) and age-matched nulliparous controls were orally infected with Y. pseudotuberculosis. i, Left: fecal pathogen burden at 6 h post-infection; Control (n = 10), Non-lactating (n = 12). Right: kinetics of fecal pathogen burden; Control (n = 9, 8, 8 (day1-3)), Non-lactating (n = 10, 7, 8 (day1-3)). j, Pathogen dissemination at day 3 post-infection; Control (n = 9), Non-lactating (n = 8). Data are representative or pooled from at least two independent experiments. Each dot represents one individual mouse except for kinetic plots. Error bars represent mean ± SD. For (a,b,d,g–j), two-tailed unpaired student’s t test was used; for (e-f), one-way ANOVA was used. *P < 0.05; **P < 0.01; ***P < 0.001.
Extended Data Fig. 10 Cellular and functional profiles in post-lactational small intestines.
Timed-pregnant female mice were analyzed 12 weeks after pups were weaned. a, Fold change in cell number of major myeloid cell types in siLP of pregnant, lactating and post-lactating mice compared with aged-matched nulliparous controls; Pregnant (n = 4), Lactating (n = 4), Post-lactation (n = 10). b, Fold change in cell number of various lymphocytes in siLP of post-lactating mice compared with aged-matched nulliparous controls; n = 6. c, Frequency of cytokine-producing lymphocytes. d, Small intestinal length; Control (n = 11), Post-lactation (n = 10). e, Villus length; Control (n = 3), Post-lactation (n = 6). f, Absolute cell numbers of total immune cells and major myeloid cell types in siLP, normalized by small intestinal length; Control (n = 11), Post-lactation (n = 10). g, Single-channel grey-scale images for the representative confocal images of proximal small intestine, with staining for epithelial cells (EPCAM), villus-resident goblet cells (UEA1) and nuclei (DAPI). Scale bar = 70 μm. h, Single-channel grey-scale images for the representative confocal images of small intestine, with staining for MUC2+ goblet cells. Scale bar = 50 μm. i, Intestinal permeability in lactating, post-lactating mice and their respective age-matched nulliparous controls, measured by FITC-dextran translocation into the circulation following oral gavage; Control (n = 10), Lactating (n = 8), Control (n = 9), Post-lactation (n = 10). Data are representative or pooled from at least two independent experiments. Each dot represents one individual mouse. Error bars and numbers in representative flow plots indicate mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001 (two-tailed unpaired student’s t test).
Supplementary information
Supplementary Table 1 (download XLSX )
Single-cell RNA-seq of crypt-enriched intestinal epithelium.
Supplementary Table 2 (download XLSX )
Xenium-based spatial profiling of intestinal epithelium and immune cell populations.
Supplementary Table 3 (download XLSX )
Eosinophil subsets identified from Xenium spatial transcriptomic analysis of the intestine.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Huang, C., Sun, A., Reyes, J. et al. Eosinophils drive intestinal remodelling and innate defence in reproduction. Nature (2026). https://doi.org/10.1038/s41586-026-10531-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41586-026-10531-6
