
Abstract
Temperature compensation stabilizes the speed of circadian clocks. Uncompensated molecular clock cycles would accelerate severalfold with each 10 °C increase, precluding reliable timekeeping. Despite such thermal buffering, some clock-controlled behavioral cycles complete by up to two hours earlier or later depending on environmental temperatures. We show that temperature-dependent changes in the speed of behavioral cycles can be explained by changes in the speed of the clock itself. Although the speed of all clocks is insensitive to thermal energy, we found that in neurons the clock speed is regulated by temperature information. When the threshold of ~26 °C is exceeded for ~24 h, a pathway mediated by the LIM-homeodomain transcription factor Lim1 instructs the clocks in the Drosophila brain to accelerate. Clock acceleration enables earlier morning awakening. This work suggests that modestly altering the clock speed enables behavioral thermoadaptation, via regulated steps that do not compromise the reliability of circadian timekeeping.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others


Data availability
Source data are provided with this paper.
Code availability
MATLAB programs used to (1) quantify calcium transients and (2) parse DAM activity data are available online at https://github.com/CrickmoreRoguljaLabs.
References
Tattersall, G. J. et al. Coping with thermal challenges: physiological adaptations to environmental temperatures. Compr. Physiol. 2, 2151–2202 (2012).
Miquel, J., Lundgren, P. R., Bensch, K. G. & Atlan, H. Effects of temperature on the life span, vitality and fine structure of Drosophila melanogaster. Mech. Ageing Dev. 5, 347–370 (1976).
Patke, A., Young, M. W. & Axelrod, S. Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 21, 67–84 (2020).
Pittendrigh, C. S. On temperature independence in the clock system controlling emergence time in Drosophila. Proc. Natl Acad. Sci. USA 40, 1018–1029 (1954).
Hastings, J. W. & Sweeney, B. M. On the mechanism of temperature independence in a biological clock. Proc. Natl Acad. Sci. USA 43, 804–811 (1957).
Lahiri, K. et al. Temperature regulates transcription in the zebrafish circadian clock. PLoS Biol. 3, e351 (2005).
Mehra, A. et al. A role for casein kinase 2 in the mechanism underlying circadian temperature compensation. Cell 137, 749–760 (2009).
Kidd, P. B., Young, M. W. & Siggia, E. D. Temperature compensation and temperature sensation in the circadian clock. Proc. Natl Acad. Sci. USA 112, E6284–E6292 (2015).
Zhou, M., Kim, J. K., Eng, G. W., Forger, D. B. & Virshup, D. M. A Period2 phosphoswitch regulates and temperature compensates circadian period. Mol. Cell 60, 77–88 (2015).
Rawson, K. S. Effects of tissue temperature on mammalian activity rhythms. Cold Spring Harb. Symp. Quant. Biol. 25, 105–113 (1960).
Aschoff, J. Exogenous and endogenous components in circadian rhythms. Cold Spring Harb. Symp. Quant. Biol. 25, 11–28 (1960).
Aschoff, J. Circadian rhythms: influences of internal and external factors on the period measured in constant conditions. Z. Tierpsychol. 49, 225–249 (1979).
Lee, T. M., Holmes, W. G. & Zucker, I. Temperature dependence of circadian rhythms in golden-mantled ground squirrels. J. Biol. Rhythms 5, 25–34 (1990).
Lehmann, M., Spoelstra, K., Visser, M. E. & Helm, B. Effects of temperature on circadian clock and chronotype: an experimental study on a passerine bird. Chronobiol. Int. 29, 1062–1071 (2012).
Rogulja, D. & Young, M. W. Control of sleep by cyclin A and its regulator. Science 335, 1617–1621 (2012).
Hobert, O. & Westphal, H. Functions of LIM-homeobox genes. Trends Genet. 16, 75–83 (2000).
Shirasaki, R. & Pfaff, S. L. Transcriptional codes and the control of neuronal identity. Annu. Rev. Neurosci. 25, 251–281 (2002).
Hatori, M. et al. Lhx1 maintains synchrony among circadian oscillator neurons of the SCN. eLife 3, e03357 (2014).
Bedont, J. L. et al. Lhx1 controls terminal differentiation and circadian function of the suprachiasmatic nucleus. Cell Rep. 7, 609–622 (2014).
Bedont, J. L. et al. An Lhx1-regulated transcriptional network controls sleep/wake coupling and thermal resistance of the central circadian clockworks. Curr. Biol. 27, 128–136 (2017).
Hamada, F. N. et al. An internal thermal sensor controlling temperature preference in Drosophila. Nature 454, 217–220 (2008).
Reinhard, N. et al. Synaptic connectome of the Drosophila circadian clock. Nat. Commun. 15, 10392 (2024).
Delventhal, R. et al. Dissection of central clock function in Drosophila through cell-specific CRISPR-mediated clock gene disruption. eLife 8, e48308 (2019).
Schlichting, M., Diaz, M. M., Xin, J. & Rosbash, M. Neuron-specific knockouts indicate the importance of network communication to Drosophila rhythmicity. eLife 8, e48301 (2019).
Takahashi, J. S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 18, 164–179 (2017).
Rosato, E. & Kyriacou, C. P. Analysis of locomotor activity rhythms in Drosophila. Nat. Protoc. 1, 559–568 (2006).
Bargiello, T. A., Jackson, F. R. & Young, M. W. Restoration of circadian behavioural rhythms by gene transfer in Drosophila. Nature 312, 752–754 (1984).
Reddy, P. et al. Molecular analysis of the period locus in Drosophila melanogaster and identification of a transcript involved in biological rhythms. Cell 38, 701–710 (1984).
Versteven, M., Ernst, K. M. & Stanewsky, R. A robust and self-sustained peripheral circadian oscillator reveals differences in temperature compensation properties with central brain clocks. iScience 23, 101388 (2020).
Dillon, M. E., Wang, G., Garrity, P. A. & Huey, R. B. Review: thermal preference in Drosophila. J. Therm. Biol. 34, 109–119 (2009).
Chen, C. et al. Drosophila ionotropic receptor 25a mediates circadian clock resetting by temperature. Nature 527, 516–520 (2015).
Gallio, M., Ofstad, T. A., Macpherson, L. J., Wang, J. W. & Zuker, C. S. The coding of temperature in the Drosophila brain. Cell 144, 614–624 (2011).
Ni, L. et al. A gustatory receptor paralogue controls rapid warmth avoidance in Drosophila. Nature 500, 580–584 (2013).
Budelli, G. et al. Ionotropic receptors specify the morphogenesis of phasic sensors controlling rapid thermal preference in Drosophila. Neuron 101, 738–747.e3 (2019).
Sweeney, S. T., Broadie, K., Keane, J., Niemann, H. & O’Kane, C. J. Targeted expression of tetanus toxin light chain in Drosophila specifically eliminates synaptic transmission and causes behavioral defects. Neuron 14, 341–351 (1995).
Zhong, L. et al. Thermosensory and nonthermosensory isoforms of Drosophila melanogaster TRPA1 reveal heat-sensor domains of a thermoTRP channel. Cell Rep. 1, 43–55 (2012).
Luo, J., Shen, W. L. & Montell, C. TRPA1 mediates sensation of the rate of temperature change in Drosophila larvae. Nat. Neurosci. 20, 34–41 (2017).
Tang, X. et al. The role of PDF neurons in setting the preferred temperature before dawn in Drosophila. eLife 6, e23206 (2017).
Lamaze, A. et al. Regulation of sleep plasticity by a thermo-sensitive circuit in Drosophila. Sci. Rep. 7, 40304 (2017).
Jin, X. et al. A subset of DN1p neurons integrates thermosensory inputs to promote wakefulness via CNMa signaling. Curr. Biol. 31, 2075–2087 (2021).
Alpert, M. H., Gil, H., Para, A. & Gallio, M. A thermometer circuit for hot temperature adjusts Drosophila behavior to persistent heat. Curr. Biol. 32, 4079–4087 (2022).
Collins, R. T. & Treisman, J. E. Osa-containing Brahma chromatin remodeling complexes are required for the repression of wingless target genes. Genes Dev. 14, 3140–3152 (2000).
Gao, X. J. et al. A transcriptional reporter of intracellular Ca2+ in Drosophila. Nat. Neurosci. 18, 917–925 (2015).
Flourakis, M. et al. A conserved bicycle model for circadian clock control of membrane excitability. Cell 162, 836–848 (2015).
Liang, X., Holy, T. E. & Taghert, P. H. Synchronous Drosophila circadian pacemakers display nonsynchronous Ca2+ rhythms in vivo. Science 351, 976–981 (2016).
Zhang, E. E. & Kay, S. A. Clocks not winding down: unravelling circadian networks. Nat. Rev. Mol. Cell Biol. 11, 764–776 (2010).
Zhang, L. et al. DN1(p) circadian neurons coordinate acute light and PDF inputs to produce robust daily behavior in Drosophila. Curr. Biol. 20, 591–599 (2010).
Noyes, M. B. et al. Analysis of homeodomain specificities allows the family-wide prediction of preferred recognition sites. Cell 133, 1277–1289 (2008).
Abruzzi, K. C. et al. RNA-seq analysis of Drosophila clock and non-clock neurons reveals neuron-specific cycling and novel candidate neuropeptides. PLoS Genet. 13, e1006613 (2017).
Vergara, C., Latorre, R., Marrion, N. V. & Adelman, J. P. Calcium-activated potassium channels. Curr. Opin. Neurobiol. 8, 321–329 (1998).
Abou Tayoun, A. N. et al. The Drosophila SK channel (dSK) contributes to photoreceptor performance by mediating sensitivity control at the first visual network. J. Neurosci. 31, 13897–13910 (2011).
Walcott, K. C. E., Mauthner, S. E., Tsubouchi, A., Robertson, J. & Tracey, W. D. The Drosophila small conductance calcium-activated potassium channel negatively regulates nociception. Cell Rep. 24, 3125–3132 (2018).
Gertner, D. M., Desai, S. & Lnenicka, G. A. Synaptic excitation is regulated by the postsynaptic dSK channel at the Drosophila larval NMJ. J. Neurophysiol. 111, 2533–2543 (2014).
Nitabach, M. N. et al. Electrical hyperexcitation of lateral ventral pacemaker neurons desynchronizes downstream circadian oscillators in the fly circadian circuit and induces multiple behavioral periods. J. Neurosci. 26, 479–489 (2006).
Hamasaka, Y. et al. Glutamate and its metabotropic receptor in Drosophila clock neuron circuits. J. Comp. Neurol. 505, 32–45 (2007).
Guo, F. et al. Circadian neuron feedback controls the Drosophila sleep–activity profile. Nature 536, 292–297 (2016).
Baines, R. A., Uhler, J. P., Thompson, A., Sweeney, S. T. & Bate, M. Altered electrical properties in Drosophila neurons developing without synaptic transmission. J. Neurosci. 21, 1523–1531 (2001).
Dissel, S. et al. The logic of circadian organization in Drosophila. Curr. Biol. 24, 2257–2266 (2014).
Green, E. W. et al. Drosophila circadian rhythms in seminatural environments: summer afternoon component is not an artifact and requires TrpA1 channels. Proc. Natl Acad. Sci. USA 112, 8702–8707 (2015).
Vanin, S. et al. Unexpected features of Drosophila circadian behavioural rhythms under natural conditions. Nature 484, 371–375 (2012).
Wheeler, D. A., Hamblen-Coyle, M. J., Dushay, M. S. & Hall, J. C. Behavior in light-dark cycles of Drosophila mutants that are arrhythmic, blind, or both. J. Biol. Rhythms 8, 67–94 (1993).
Buhr, E. D., Yoo, S. H. & Takahashi, J. S. Temperature as a universal resetting cue for mammalian circadian oscillators. Science 330, 379–385 (2010).
Majercak, J., Sidote, D., Hardin, P. E. & Edery, I. How a circadian clock adapts to seasonal decreases in temperature and day length. Neuron 24, 219–230 (1999).
Yadlapalli, S. et al. Circadian clock neurons constantly monitor environmental temperature to set sleep timing. Nature 555, 98–102 (2018).
Lamaze, A., Kratschmer, P., Chen, K. F., Lowe, S. & Jepson, J. E. C. A wake-promoting circadian output circuit in Drosophila. Curr. Biol. 28, 3098–3105 (2018).
Zhang, Y., Liu, Y., Bilodeau-Wentworth, D., Hardin, P. E. & Emery, P. Light and temperature control the contribution of specific DN1 neurons to Drosophila circadian behavior. Curr. Biol. 20, 600–605 (2010).
Guo, F., Holla, M., Diaz, M. M. & Rosbash, M. A circadian output circuit controls sleep-wake arousal in Drosophila. Neuron 100, 624–635.e4 (2018).
Patke, A. et al. Mutation of the human circadian clock gene CRY1 in familial delayed sleep phase disorder. Cell 169, 203–215.e13 (2017).
Kang, K. et al. Analysis of Drosophila TRPA1 reveals an ancient origin for human chemical nociception. Nature 464, 597–600 (2010).
Xiang, Y. et al. Light-avoidance-mediating photoreceptors tile the Drosophila larval body wall. Nature 468, 921–926 (2010).
He, H. et al. Neurodevelopmental role for VGLUT2 in pyramidal neuron plasticity, dendritic refinement, and in spatial learning. J. Neurosci. 32, 15886–15901 (2012).
Blau, J. & Young, M. W. Cycling vrille expression is required for a functional Drosophila clock. Cell 99, 661–671 (1999).
Titos, I. et al. A gut-secreted peptide suppresses arousability from sleep. Cell 186, 1382–1397.e21 (2023).
McGuire, S. E., Le, P. T., Osborn, A. J., Matsumoto, K. & Davis, R. L. Spatiotemporal rescue of memory dysfunction in Drosophila. Science 302, 1765–1768 (2003).
Tackenberg, M. C. & Hughey, J. J. The risks of using the chi-square periodogram to estimate the period of biological rhythms. PLoS Comput. Biol. 17, e1008567 (2021).
Schmid, B., Helfrich-Forster, C. & Yoshii, T. A new ImageJ plug-in ‘ActogramJ’ for chronobiological analyses. J. Biol. Rhythms 26, 464–467 (2011).
Geissmann, Q., Garcia Rodriguez, L., Beckwith, E. J. & Gilestro, G. F. Rethomics: an R framework to analyse high-throughput behavioural data. PLoS ONE 14, e0209331 (2019).
Hendricks, J. C. et al. Rest in Drosophila is a sleep-like state. Neuron 25, 129–138 (2000).
Nitz, D. A., van Swinderen, B., Tononi, G. & Greenspan, R. J. Electrophysiological correlates of rest and activity in Drosophila melanogaster. Curr. Biol. 12, 1934–1940 (2002).
Shaw, P. J., Cirelli, C., Greenspan, R. J. & Tononi, G. Correlates of sleep and waking in Drosophila melanogaster. Science 287, 1834–1837 (2000).
Long, X., Colonell, J., Wong, A. M., Singer, R. H. & Lionnet, T. Quantitative mRNA imaging throughout the entire Drosophila brain. Nat. Methods 14, 703–706 (2017).
Chen, T. W. et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 499, 295–300 (2013).
Feng, Y., Ueda, A. & Wu, C. F. A modified minimal hemolymph-like solution, HL3.1, for physiological recordings at the neuromuscular junctions of normal and mutant Drosophila larvae. J. Neurogenet. 18, 377–402 (2004).
Fogle, K. J., Parson, K. G., Dahm, N. A. & Holmes, T. C. CRYPTOCHROME is a blue-light sensor that regulates neuronal firing rate. Science 331, 1409–1413 (2011).
Carter, A. G. & Sabatini, B. L. State-dependent calcium signaling in dendritic spines of striatal medium spiny neurons. Neuron 44, 483–493 (2004).
Zhang, S. X., Rogulja, D. & Crickmore, M. A. Dopaminergic circuitry underlying mating drive. Neuron 91, 168–181 (2016).
Cavanaugh, D. J. et al. Identification of a circadian output circuit for rest: activity rhythms in Drosophila. Cell 157, 689–701 (2014).
Akerboom, J. et al. Optimization of a GCaMP calcium indicator for neural activity imaging. J. Neurosci. 32, 13819–13840 (2012).
Mukamel, E. A., Nimmerjahn, A. & Schnitzer, M. J. Automated analysis of cellular signals from large-scale calcium imaging data. Neuron 63, 747–760 (2009).
Grima, B., Chelot, E., Xia, R. & Rouyer, F. Morning and evening peaks of activity rely on different clock neurons of the Drosophila brain. Nature 431, 869–873 (2004).
Kaneko, M., Helfrich-Forster, C. & Hall, J. C. Spatial and temporal expression of the period and timeless genes in the developing nervous system of Drosophila: newly identified pacemaker candidates and novel features of clock gene product cycling. J. Neurosci. 17, 6745–6760 (1997).
Acknowledgements
We thank D. Anderson (California Institute of Technology), J. Blau (New York University), J. Botas (Baylor College of Medicine), P. Dolph (Dartmouth College), P. Hardin (Texas A&M University), F. Rouyer (CNRS Université Paris-Saclay), M. Rosbash (Brandeis University), G. Rubin (Janelia Research Campus), R. Stanewsky (University of Münster), the Vienna Drosophila Resource Center, TsingHua Fly Center and the Bloomington Stock Center for providing reagents. Enhanced Neuroimaging Core at the Harvard Neurodiscovery Center provided access to their confocal microscope during the initial stages of the project. B. Sabatini provided support and equipment for two-photon microscopy. We thank the members of the Rogulja and Crickmore labs, as well as M. Rosbash, M. Young and C. Weitz for comments on the manuscript and helpful discussions. This work was supported by the Whitehall Foundation and the Klingenstein-Simons Foundation (to D.R.) and the National Science Foundation of China (32571159 to Z.L.). Z.L. was an Edward R. and Anne G. Lefler Postdoctoral Fellow. S.X.Z. was a Stuart H.Q. and Victoria Quan Fellow. The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
Z.L. and D.R. designed the study and wrote the manuscript with input from all authors. Z.L., D.X., S.X.Z., W.C. and H.Z. performed the experiments. All authors analyzed the data.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Neuroscience thanks the anonymous reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Pan-neuronal versus clock cell-specific Lim1 depletion leads to distinct sleep phenotypes.
a, Left, daily sleep profiles of controls (nSyb-Gal4 and UAS-lim1-RNAi) and flies with Lim1 depleted from the nervous system (nSyb>lim1-RNAi) at 25 °C in 12-h light/12-h dark conditions (denoted by white–black bars on the bottom). Right, quantification of total daily sleep. n = 19 for nSyb-Gal4; 15 for UAS-lim1-RNAi; 15 for nSyb>lim1-RNAi, one-way ANOVA followed by Dunnett’s multiple comparisons test. b, Immunostaining for Lim1 in controls and flies with Lim1 depleted from the nervous system. Two different lim1-RNAi lines produced the same result. Scale bar = 50 µm. c, Quantification of Lim1 levels in controls (tim-Gal4) and flies with Lim1 depleted from clock neurons (tim>lim1-RNAi), at 25 °C and 30 °C. Sample sizes ranged from n = 10 to n = 24, two-way ANOVA followed by Tukey’s multiple comparisons test. d, Daily sleep profiles in 12-h light/12-h dark conditions (denoted by white–black bars on the bottom). Right, total sleep during the last 4 h of darkness (black bars in left panels). Flies depleted of Lim1 in clock neurons (tim>lim1-RNAi) are compared to the parental controls (tim-Gal4 and UAS-lim1-RNAi). n (25 °C, 30 °C): tim-Gal4 (32, 32); UAS-lim1-RNAi (32, 18); tim>lim1-RNAi (32, 66), two-way ANOVA followed by Tukey’s multiple comparisons test. e, Daily averaged locomotor activity profiles (top left) and sleep profiles (bottom left) in 12-h light/12-h dark conditions (denoted by white–black bars on the bottom). Flies were depleted of Lim1 in circadian clock neurons using a second RNAi line (tim>lim1-RNAi 2) and compared to the parental controls (tim-Gal4 and UAS-lim1-RNAi 2) at 25 °C and 30 °C. Right, quantification of the total locomotor activity (top right) and sleep time (bottom right) during the 4 h preceding lights-on (black bars in left panels). n (25 °C, 30 °C): tim-Gal4 (29, 16); UAS-lim1-RNAi 2 (32, 11); tim>lim1-RNAi 2 (32, 23), two-way ANOVA followed by Tukey’s multiple comparisons test. Quantification represents mean ± s.e.m. ***P < 0.001.
Extended Data Fig. 2 Warming accelerates the clock via Lim1.
a, Percentage of rhythmic animals when clock proteins PER or TIMELESS (TIM) are depleted from s-LNvs (PDF-Gal4), LNds (Mai179-Gal4;pdf-Gal80)90, or both (Mai179-Gal4), using CRISPR. Disrupting the molecular clock in both populations renders flies arrhythmic. b, PER oscillations in the brains of control flies (tim-Gal4) and flies with Lim1 depleted from circadian clock neurons (tim>lim1-RNAi) on day 6 in darkness, extra sum‑of‑squares F test. Data are replotted from Fig. 1b. c, Quantification of PER oscillation in nonpacemaking clock neurons (clock neurons other than s-LNv and LNd), in controls (tim-Gal4) and flies with Lim1 depleted from clock neurons (tim>lim1-RNAi). In controls, DN1ps had an advanced phase of PER oscillations at 30 °C compared to 25 °C, consistent with previous findings that s-LNvs pace the molecular clocks in DN1ps47. Additionally, DN3s displayed an advanced PER oscillation phase at 30 °C compared to 25 °C, likely because s-LNvs also synchronize DN3 clocks. Depleting Lim1 from clock neurons abolished the temperature-induced phase shift. l-LNvs showed no daily PER rhythmicity, mirroring previous reports of rapid PER oscillation decay in darkness90. DN2 neurons exhibited a ~12-h phase shift in PER oscillations relative to pacemaking neurons, consistent with previous reports91. n (25 °C, 30 °C): tim-Gal4 (6–14); tim>lim1-RNAi (6–15), extra sum‑of‑squares F test. Quantification represents mean ± s.e.m. ***P < 0.001.
Extended Data Fig. 3 TRPA1 is required for clock acceleration in warm conditions.
a, Quantification of temperature-dependent PER oscillations in common fly strains. n (25 °C, 30 °C): Canton-S (5–14); w1118 (8–16), extra sum‑of‑squares F test. b, Locomotor rhythm periodicity in common strains at 20 °C, 25 °C and 30 °C. n = 16 for Hikone-A-W; 24 for Canton-S; 21 for Florida-9; 17 for Oregon-R; 18 for Berlin-K at 20 °C, two-way ANOVA followed by Tukey’s multiple comparisons test. The data for 25 °C and 30 °C are the same as plotted in Fig. 2b. c, Left, schematic of fly thermosensors. Right, quantification of locomotor rhythm periodicity in controls (w1118), TRPA11 mutants, nocte1 mutants, and w1118 flies with aristae surgically removed. n (25 °C, 30 °C): Canton-S (16, 16); TRPA11 (17, 21); nocte1 (20, 15); flies with no aristae (12, 32), two-way ANOVA followed by Tukey’s multiple comparisons test. d, Left, schematic of the four TRPA1 isoforms and the three isoform-specific TRPA1 mutants. Gray bar: UTR; black bar: exon. TRPA1AB: isoforms A and B are deleted. TRPA1CD: isoforms C and D are deleted. TRPA1ACD: isoform A is mutated and isoforms C and D are deleted. Right, quantification of locomotor rhythm periodicity of isoform-specific TRPA1 mutants. n (25 °C, 30 °C): TRPA1AB (9, 10); TRPA1CD (23, 22); TRPA1ACD (18, 21), two-way ANOVA followed by Tukey’s multiple comparisons test. Quantification represents mean ± s.e.m. **P < 0.01, ***P < 0.001. Fruit fly diagram in c created with BioRender.com.
Extended Data Fig. 4 Warmth increases Lim1 levels in DN1ps in a TRPA1-dependent manner.
a, Levels of a pan-neuronal protein Osa (a subunit of the SWI/SNF chromatin remodeling complex) in DN1ps do not change with temperature. n = 11 and 10 at 25 °C and 30 °C, respectively, Student’s t-tests. b, Quantification of Lim1 levels in DN1p neurons of controls (w1118), TRPA11 and nocte1 mutants, and flies with aristae surgically removed. TRPA1 mutation, but not other manipulations, prevents Lim1 levels in DN1ps from increasing when temperature increases. n (25 °C, 30 °C): w1118 (32, 15); TRPA11 (32, 17); nocte1 (27, 17); flies with no aristae (33, 16), two-way ANOVA followed by Tukey’s multiple comparisons test. c, Immunostaining for Lim1 and CLK. Lim1 is expressed in DN1ps and DN2s. Lim1 signal is eliminated from DN1ps but not DN2s when lim1-RNAi is expressed with DN1-Gal4 during adulthood. Scale bar = 5 µm. d, Averaged daily locomotor activity (top) and sleep (bottom) profiles in 12-h light/12-h dark conditions (denoted by white–black bars on the bottom). Flies with Lim1 depleted from DN1 neurons during adulthood (DN1>lim1-RNAi;Tub-Gal80ts) are compared to the parental controls (DN1-Gal4 and UAS-lim1-RNAi;Tub-Gal80ts). Please note that in this experiment flies lost more sleep after shifting from 25 °C to 30 °C than in experiments shown in Figs. 1a and 7a, likely because they were raised at 18 °C; flies raised at lower temperature are more sensitive to warming (our observation). Although the combination of 18 °C rearing and 30 °C testing induced a significant baseline reduction in sleep, a statistically significant difference can be observed between Lim1 knockdown flies and controls during the predawn hours (black bars in left panels) within the 30 °C condition. n (25 °C, 30 °C): DN1-Gal4 (32, 35); UAS-lim1-RNAi;Tub-Gal80ts (13, 23); DN1>lim1-RNAi;Tub-Gal80ts (32, 28), two-way ANOVA followed by Tukey’s multiple comparisons test. Quantification represents mean ± s.e.m. ***P < 0.001.
Extended Data Fig. 5 LPN clock neurons are not required for temperature-dependent adjustment of clock speed.
Top, averaged locomotor activity for control flies (LPN-Gal4) and flies with LPN neurons silenced (LPN>Kir2.1 and LPN > TNT), at 25 °C and 30 °C. Bottom, quantification. n (25 °C, 30 °C): LPN-Gal4 (15, 13); UAS-Kir2.1 (25, 22); LPN>Kir2.1 (16, 14); UAS-TNT (25, 25); LPN > TNT (14, 21), two-way ANOVA followed by Tukey’s multiple comparisons test. Quantification represents mean ± s.e.m. ***P < 0.001.
Extended Data Fig. 6 Lim1 increases both the basal activity and the optogenetically induced calcium transients in DN1 neurons specifically during the morning.
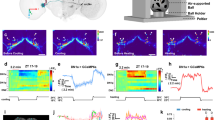
a, DN1 neurons can be activated by blue light via Cryptochrome-mediated excitation84. We used two-photon laser-scanning microscopy for calcium imaging to confirm the results obtained using regular fluorescence microscopy (Fig. 4b), which might be affected by experimental activation of Cryptochrome. Two-photon images of DN1 projections in the SMPp showing GCaMP6s fluorescence when excited at 910 nm (calcium-dependent) and 820 nm (calcium-independent). Depletion of Lim1 from DN1 neurons (DN1>lim1-RNAi + GCaMP6s) specifically abolishes the morning (ZT0-2) calcium signal compared to controls (DN1-Gal4). Each image is a maximum projection spanning ~45 µm along the anterior-posterior axis. See Methods for details. Scale bar = 5 µm. b, Left, normalized DN1 > GCaMP6s fluorescence (F910 nm/F820 nm) in the SMPp is higher in the morning (ZT0-2) than evening (ZT10-12), a difference that is abolished when Lim1 is depleted. Right, no difference was seen in the calcium-independent fluorescence (F820 nm) among different groups. n = 8 for all conditions, two-way ANOVA followed by Tukey’s multiple comparisons test. c, Optically triggered calcium transients in DN1 projections (DN1>CsChrimson + GCaMP6s) have a higher amplitude in the subjective morning (left, CT0-2) than evening (middle, CT10-12), a difference that is abolished when Lim1 is depleted from these cells (DN1>lim1-RNAi + CsChrimson + GCaMP6s). The GCaMP6s transient peaks are quantified on the right. All experiments were performed at 25 °C. n = 8 for all conditions, two-way ANOVA followed by Tukey’s multiple comparisons test. Quantification represents mean ± s.e.m.
Extended Data Fig. 7 Circadian period analysis and Lim1-dependent modulation of sleep timing under simulated seasonal light and temperature cycles.
a, Quantification of locomotor rhythm periodicity of Gal4 controls (DN1-Gal4, PDF-Gal4, and Mai179-Gal4;PDF-Gal80), UAS controls (UAS-VGlut-RNAi 2, UAS-mGluRA-RNAi 2), flies with VGlut depleted from DN1s using a second RNAi line (DN1>VGlut-RNAi 2), mGluRA depleted from LNvs (s-LNvs and l-LNvs) using a second RNAi line (PDF>mGluRA-RNAi 2), and mGluRA depleted from LNds using two independent RNAi lines (Mai179;pdf-Gal80>mGluRA-RNAi 1 and Mai179;pdf-Gal80>mGluRA-RNAi 2) in constant darkness at 25 °C or 30 °C. The data for DN1-Gal4 and PDF-Gal4 are the same as plotted in Fig. 6a. n (25 °C, 30 °C): DN1-Gal4 (27, 29); UAS-VGlut-RNAi 2 (31, 32); DN1>VGlut-RNAi 2 (27, 28); PDF-Gal4 (21, 31); UAS-mGluRA-RNAi 2 (14, 21); PDF>mGluRA-RNAi 2 (20, 26); Mai179-Gal4;PDF-Gal80 (20, 25); Mai179-Gal4;PDF-Gal80>mGluRA-RNAi 1 (8, 15); Mai179-Gal4;PDF-Gal80>mGluRA-RNAi 2 (8, 14), two-way ANOVA followed by Tukey’s multiple comparisons test. b, Left, averaged daily sleep profiles in 12-h light/12-h dark conditions (denoted by white–black bars on the bottom). Controls (DN1-Gal4) are compared to flies with Lim1 depleted from DN1 neurons (DN1>lim1-RNAi), and flies with Lim1 depleted and dominant-negative SK overexpressed in DN1 neurons (DN1>lim1-RNAi + SK-DN). Right, quantification of total sleep during the final 4 h of the dark phase (black bars in left panels), n (25 °C, 30 °C): DN1-Gal4 (18, 29); DN1>lim1-RNAi (15, 23); DN1>lim1-RNAi + SK-DN (32, 33), two-way ANOVA followed by Tukey’s multiple comparisons test. c, Locomotor activity in naturalistic conditions. Top, average daily locomotor activity profiles under spring-like (left) and summer-like conditions (right). Temperature fluctuations (background color gradient) and light cycle (color bar, peak ~380 lux) are indicated. Morning (M), afternoon (A), and evening (E) activity peaks are labeled. Bottom right, quantification of total locomotor activity within a 3-h window spanning the typical morning activity period (black bars in top panels; defined as 1 h before scheduled lights-on to 2 h after scheduled lights-on) to specifically assess changes in morning wake timing. n (spring, summer): DN1-Gal4 (27, 48); UAS-lim1-RNAi (32, 32); DN1>lim1-RNAi (48, 40), two-way ANOVA followed by Tukey’s multiple comparisons test. Quantification represents mean ± s.e.m. ***P < 0.001.
Extended Data Fig. 8 Lim1 does not regulate entrainment of the clock phase to temperature cycles.
Circadian clocks can be reset not only by light-dark cycles, but also by temperature cycles with an amplitude as low as ~2 °C61. DN1ps mediate entrainment to low-amplitude temperature cycles31; we asked whether this requires Lim1. Double-plotted actograms of control (DN1-Gal4) and Lim1-depleted (DN1>lim1-RNAi) flies. a, Flies were entrained to 12-h light/12-h dark (LD) cycles for 3 days before shifting to the first temperature cycle (TC1, 21 °C/25 °C, constant darkness, 6 h delayed relative to the LD cycle) for 5 days, followed by 5 days in the second temperature cycle (TC2, 21 °C/25 °C, constant darkness, 6 h delayed relative to TC1), finishing with 5 days in constant temperature and darkness. n = 31 and 30 for DN1-Gal4 and DN1>lim1-RNAi, respectively, Student’s t-tests. b, As in a, but with TC1 and TC2 at 25 °C/29 °C. n = 31 and 31 for DN1-Gal4 and DN1>lim1-RNAi, respectively, Student’s t-tests. c, Flies were entrained to LD cycle at 25 °C for 3 days before shifting to the first temperature cycle (TC, 25 °C/27 °C, constant darkness, 8 h advanced relative to the LD cycle) for 6 days, finishing with 8 days in constant temperature and darkness. n = 28 and 31 for DN1-Gal4 and DN1>lim1-RNAi, respectively, Student’s t-tests. d, Flies were kept in constant light (LL) for 3 days before shifting to the first temperature cycle (TC1, 25 °C/29 °C, constant light, 8 h advanced relative to the LD cycle) for 6 days, followed by 8 days in the second temperature cycle (TC2, 25 °C/29 °C, constant light, 6 h delayed relative to TC1). n = 28 and 30 for DN1-Gal4 and DN1>lim1-RNAi, respectively, Student’s t-tests. Quantification represents mean ± s.e.m.
Supplementary information
Supplementary Information (download PDF )
Supplementary Tables 1–5.
Source data
Source Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Fig. 3 (download XLSX )
Statistical source data for Fig. 3b–e.
Source Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Fig. 5 (download XLSX )
Statistical source data for Fig. 5a,c–e.
Source Data Fig. 6 (download XLSX )
Statistical source data for Fig. 6a.
Source Data Fig. 7 (download XLSX )
Statistical source data for Fig. 7a,b.
Source Data Extended Data Fig. 1 (download XLSX )
Statistical source data for Extended Data Fig. 1a,c–e.
Source Data Extended Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 4 (download XLSX )
Statistical source data for Extended Data Fig. 4a,b,d.
Source Data Extended Data Fig. 5 (download XLSX )
Statistical source data for Extended Data Fig. 5a.
Source Data Extended Data Fig. 6 (download XLSX )
Statistical source data for Extended Data Fig. 6b,c.
Source Data Extended Data Fig. 7 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 8 (download XLSX )
Statistical source data.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Liu, Z., Xie, D., Zhang, S.X. et al. Behavioral adaptation to warm conditions via Lim1-mediated acceleration of neuronal clocks. Nat Neurosci 29, 374–386 (2026). https://doi.org/10.1038/s41593-025-02139-2
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41593-025-02139-2
