
Abstract
Once tau aggregates are formed, their neurotoxicity significantly contributes to neuronal death and cognitive decline in tauopathies, with Alzheimer’s disease being the most well-known example. Despite its central pathogenic role, however, effective therapeutic strategies targeting the neurotoxicity of tau remain poor. Here we demonstrate the pathogenic role of neuronal cell death in tau-related neurodegeneration (PS19 mouse model). Tau-expressing neurons undergo cell death through Z-DNA-binding protein 1 (ZBP1) activation triggered by endogenous Z-RNAs. These Z-RNAs are derived from reactivated transposable elements that are typically silenced within heterochromatin. Tau aggregates show a strong affinity for H3K9me3-modified chromatin, effectively sequestering these epigenetic marks from heterochromatin protein 1 (HP1), thereby disrupting the condensation of constitutive heterochromatin. Clinically, an inverse correlation between ZBP1 expression levels in excitatory neurons and cognitive performance in individuals with Alzheimer’s disease was observed. Importantly, Zbp1 haploinsufficiency significantly ameliorated cognitive deficits in aged (24-month-old) tau-transgenic mice, highlighting the therapeutic potential of ZBP1 inhibition to combat neurodegeneration in tauopathies.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others
Data availability
The source data supporting the findings of this study are available within the paper. All source data for lncRNA-seq (GSE306753), bulk RNA-seq (GSE306761), ChIP–seq (GSE306751) and RIP-seq (GSE306739) have been deposited in the Gene Expression Omnibus (GEO) and are publicly available. Source data are provided with this paper.
Code availability
No custom software code was used in the data analysis for this paper.
References
Creekmore, B. C., Watanabe, R. & Lee, E. B. Neurodegenerative Disease Tauopathies. Annu. Rev. Pathol. 19, 345–370 (2024).
Braak, H., Alafuzoff, I., Arzberger, T., Kretzschmar, H. & Del Tredici, K. Staging of Alzheimer disease-associated neurofibrillary pathology using paraffin sections and immunocytochemistry. Acta Neuropathol. 112, 389–404 (2006).
Ghetti, B. et al. Invited review: frontotemporal dementia caused by microtubule-associated protein tau gene (MAPT) mutations: a chameleon for neuropathology and neuroimaging. Neuropathol. Appl. Neurobiol. 41, 24–46 (2015).
Kovacs, G. G. et al. Distribution patterns of tau pathology in progressive supranuclear palsy. Acta Neuropathol. https://doi.org/10.1007/s00401-020-02158-2 (2020).
Zhang, W. et al. Novel tau filament fold in corticobasal degeneration. Nature 580, 283–287 (2020).
Jin, Y. et al. Amyloid-β-targeting immunotherapies for Alzheimer’s disease. J. Control. Release 375, 346–365 (2024).
Wilson, D. M. et al. Hallmarks of neurodegenerative diseases. Cell 186, 693–714 (2023).
Hu, J. et al. Microglial Piezo1 senses Aβ fibril stiffness to restrict Alzheimer’s disease. Neuron https://doi.org/10.1016/j.neuron.2022.10.021 (2023).
Yoshiyama, Y. et al. Synapse loss and microglial activation precede tangles in a P301S tauopathy mouse model. Neuron 53, 337–351 (2007).
Merchán-Rubira, J., Sebastián-Serrano, Á, Díaz-Hernández, M., Avila, J. & Hernández, F. Peripheral nervous system effects in the PS19 tau transgenic mouse model of tauopathy. Neurosci. Lett. 698, 204–208 (2019).
Zhang, T. et al. Influenza virus Z-RNAs induce ZBP1-mediated necroptosis. Cell https://doi.org/10.1016/j.cell.2020.02.050 (2020).
Caccamo, A. et al. Necroptosis activation in Alzheimer’s disease. Nat. Neurosci. 20, 1236–1246 (2017).
Balusu, S. et al. MEG3 activates necroptosis in human neuron xenografts modeling Alzheimer’s disease. Science 381, 1176–1182 (2023).
Mila-Aloma, M. et al. Plasma p-tau231 and p-tau217 as state markers of amyloid-β pathology in preclinical Alzheimer’s disease. Nat. Med. 28, 1797–1801 (2022).
Prissette, M. et al. Disruption of nuclear envelope integrity as a possible initiating event in tauopathies. Cell Rep. 40, 111249 (2022).
Manczak, M. & Reddy, P. H. Abnormal interaction of oligomeric amyloid-β with phosphorylated tau: implications to synaptic dysfunction and neuronal damage. J. Alzheimers Dis. 36, 285–295 (2013).
Rauch, J. N. et al. LRP1 is a master regulator of tau uptake and spread. Nature 580, 381–385 (2020).
Mellone, M. et al. Tau pathology is present in vivo and develops in vitro in sensory neurons from human P301S tau transgenic mice: a system for screening drugs against tauopathies. J. Neurosci. 33, 18175–18189 (2013).
Wang, R. et al. Gut stem cell necroptosis by genome instability triggers bowel inflammation. Nature 580, 386–390 (2020).
Yang, Z.-H. et al. ZBP1 senses splicing aberration through Z-RNA to promote cell death. Mol. Cell https://doi.org/10.1016/j.molcel.2025.03.023 (2025).
Cai, Z.-Y. et al. A ZBP1 isoform blocks ZBP1-mediated cell death. Cell Rep. 43, 114221 (2024).
Maelfait, J. et al. Sensing of viral and endogenous RNA by ZBP1/DAI induces necroptosis. EMBO J. 36, 2529–2543 (2017).
Sanders, D. W. et al. Distinct tau prion strains propagate in cells and mice and define different tauopathies. Neuron 82, 1271–1288 (2014).
Riegerová, P. et al. Expression and Localization of AβPP in SH-SY5Y cells depends on differentiation state. J. Alzheimers Dis. 82, 485–491 (2021).
Khlistunova, I. et al. Inducible expression of Tau repeat domain in cell models of tauopathy: aggregation is toxic to cells but can be reversed by inhibitor drugs. J. Biol. Chem. 281, 1205–1214 (2006).
Wischik, C. M. et al. Isolation of a fragment of tau derived from the core of the paired helical filament of Alzheimer disease. Proc. Natl Acad. Sci. USA 85, 4506–4510 (1988).
Styren, S. D., Hamilton, R. L., Styren, G. C. & Klunk, W. E. X-34, a fluorescent derivative of Congo red: a novel histochemical stain for Alzheimer’s disease pathology. J. Histochem. Cytochem. 48, 1223–1232 (2000).
DeVos, S. L. et al. Tau reduction prevents neuronal loss and reverses pathological tau deposition and seeding in mice with tauopathy. Sci. Transl. Med. https://doi.org/10.1126/scitranslmed.aag0481 (2017).
Krall, J. B., Nichols, P. J., Henen, M. A., Vicens, Q. & Vögeli, B. Structure and formation of Z-DNA and Z-RNA. Molecules https://doi.org/10.3390/molecules28020843 (2023).
Wang, A. J. et al. Left-handed double helical DNA: variations in the backbone conformation. Science 211, 171–176 (1981).
Zhang, H. et al. Nuclear lamina erosion-induced resurrection of endogenous retroviruses underlies neuronal aging. Cell Rep. 42, 112593 (2023).
Reilly, M. T., Faulkner, G. J., Dubnau, J., Ponomarev, I. & Gage, F. H. The role of transposable elements in health and diseases of the central nervous system. J. Neurosci. 33, 17577–17586 (2013).
Towbin, B. D. et al. Step-wise methylation of histone H3K9 positions heterochromatin at the nuclear periphery. Cell 150, 934–947 (2012).
Padeken, J., Methot, S. P. & Gasser, S. M. Establishment of H3K9-methylated heterochromatin and its functions in tissue differentiation and maintenance. Nat. Rev. Mol. Cell Biol. 23, 623–640 (2022).
Siegenfeld, A. P. et al. Polycomb-lamina antagonism partitions heterochromatin at the nuclear periphery. Nat. Commun. 13, 4199 (2022).
Harr, J. C. et al. Directed targeting of chromatin to the nuclear lamina is mediated by chromatin state and A-type lamins. J. Cell Biol. 208, 33–52 (2015).
Xiong, X. et al. Epigenomic dissection of Alzheimer’s disease pinpoints causal variants and reveals epigenome erosion. Cell https://doi.org/10.1016/j.cell.2023.08.040 (2023).
Wang, L. et al. Histone modifications regulate chromatin compartmentalization by contributing to a phase separation mechanism. Mol. Cell 76, 646–659.e646 (2019).
Déléris, A., Berger, F. & Duharcourt, S. Role of polycomb in the control of transposable elements. Trends Genet. 37, 882–889 (2021).
Waheed, Z. et al. The role of tau proteoforms in health and disease. Mol. Neurobiol. 60, 5155–5166 (2023).
Ayalon, G. et al. Antibody semorinemab reduces tau pathology in a transgenic mouse model and engages tau in patients with Alzheimer’s disease. Sci. Transl. Med. https://doi.org/10.1126/scitranslmed.abb2639 (2021).
Galpern, W. R. et al. Phase 1 studies of the anti-tau monoclonal antibody JNJ-63733657 in healthy participants and participants with Alzheimer’s disease. J. Prev. Alzheimers Dis. 11, 1592–1603 (2024).
Zhang, Y. et al. A unique death pathway keeps RIPK1 D325A mutant mice in check at embryonic day 10.5. PLoS Biol. 19, e3001304 (2021).
Zhao, Y. et al. β2-Microglobulin coaggregates with Aβ and contributes to amyloid pathology and cognitive deficits in Alzheimer’s disease model mice. Nat. Neurosci. 26, 1170–1184 (2023).
Chen, X. et al. Microglia-mediated T cell infiltration drives neurodegeneration in tauopathy. Nature 615, 668–677 (2023).
Dejanovic, B. et al. Complement C1q-dependent excitatory and inhibitory synapse elimination by astrocytes and microglia in Alzheimer’s disease mouse models. Nat. Aging 2, 837–850 (2022).
Malin, S. A., Davis, B. M. & Molliver, D. C. Production of dissociated sensory neuron cultures and considerations for their use in studying neuronal function and plasticity. Nat. Protoc. 2, 152–160 (2007).
Gibson, B. A. et al. Organization of chromatin by intrinsic and regulated phase separation. Cell 179, 470–484.e21 (2019).
Mathys, H. et al. Single-cell atlas reveals correlates of high cognitive function, dementia, and resilience to Alzheimer’s disease pathology. Cell 186, 4365–4385.e4327 (2023).
Schneider, J. A., Arvanitakis, Z., Bang, W. & Bennett, D. A. Mixed brain pathologies account for most dementia cases in community-dwelling older persons. Neurology 69, 2197–2204 (2007).
Acknowledgements
We thank the National Health and Disease Human Brain Tissue Resource Center for providing the brain material. We are grateful to X. Wang, N. Liu, G. Li and B. Zhu for providing the experimental resources. The study was supported by the Zhejiang Provincial Natural Science Foundation of China (LHZSD25C070001 to W.M.), the National Natural Science Foundation of China (82530046, 82588302, 32225016 to W.M.; 82388201 to J.H.; 32170751 to Z.-H.Y.; 82203426, 82472742 to L.S.), the National Key R&D Program of China (2024YFA1306400 and 2021YFA1101401 to W.M.; 2020YFA0803500 to J.H.), the ‘Pioneer’ and ‘Leading Goose’ R&D Program of Zhejiang (2025C02110 to W.M. and S.N.), the CAMS Innovation Fund for Medical Sciences (CIFMS) (2019-I2M-5-062 to J.H.), the Fujian Province Central to Local Science and Technology Development Special Program (2022L3079 to J.H.), the Fu-Xia-Quan Zi-Chuang District Cooperation Program (3502ZCQXT2022003 to J.H.) and the Noncommunicable Chronic Diseases-National Science and Technology Major Project (2024ZD0524900 to Z.-H.Y.).
Author information
Authors and Affiliations
Contributions
W.M. and J.H. conceptualized and supervised the study. S.-A.W., Q.C., B.-X.Z, S.-H.G., L.L., W.S. and Z.-H.Y. curated the data. X.X., J.N., J.W., L.Z., P.L., Z.-Y.C., H.-F.Y., S.Z., S.N., B.L., D.W., P.C., X.Q., Z.-H.Y., J.H. and W.M. conducted formal analysis. S.-A.W. and Z.-H.Y. devised the methodology. D.F., W.C., Y.Z. and J.H. obtained resources. W.M., Z.-H.Y. and Q.C. wrote the original draft of the manuscript; J.H., Q.C., Z.-H.Y. and W.M. contributed to review and editing. S.-A.W. and L.L. identified that neuronal cell death was pathogenic for neurodegeneration in AD. Q.C., W.L., S.-H.G. and S.-A.W. performed most of the immunofluorescence, immunohistochemistry and behavioral testing experiments on the mouse model. B.-X.Z. performed electrophysiological experiments. S.-H.G. and B.-X.Z. performed experiments on patient samples. W.S. and P.L. performed LLPS experiments. J.N. and X.X. uncovered the negative correlation of ZBP1 levels with the cognitive diagnosis of patients with AD. Z.-H.Y. performed cellular experiments and in vitro biochemical experiments. B.L., S.N. and W.C. provided pathological diagnoses.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Neuroscience thanks the anonymous reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Neuron necroptosis is essential for Tau-induced neurodegeneration.
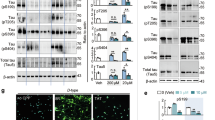
a. Representative images of TUNEL staining and quantification of TUNEL/DAPI on hippocampus sections of mice with indicated genotypes and on spinal cord (SC) section of PS19 mice (WT: 9-month-old, 5xFAD: 7-month-old and PS19: 9-month-old). Scale bar: 50 μm. n = 10 mice. Selected areas are shown at higher magnification. b. Schematic of classical necroptosis and apoptosis pathway, created in BioRender; Qian, Chen. https://biorender.com/ip4hf6s (2026). c. Representative images of TUNEL staining and quantification of TUNEL/DAPI on hippocampus sections from mice with indicated genotypes (9-month-old). Scale bar: 50 μm. n = 10 mice. d. RT–qPCR analysis of relative Ripk3 mRNA expression level in hippocampus of mice with indicated genotypes. n = 3 mice. e. RIPK3 and p-Tau (AT8) staining on brain sections of mice with indicated genotypes. Selected areas are shown at higher magnification. Scale bar: 5 μm. f. Representative images of pMLKL co-stained with NeuN on brain sections of mice with indicated genotypes at 9-month-old. Selected areas are shown magnified to the left of each image. Scale bar: 20 μm. g. Representative images of pRIPK3 co-stained with p-Tau (AT8) and NeuN on brain sections of mice with indicated genotypes at 9-month-old. Selected areas are shown magnified to the right of each image. Scale bar: 10 μm. Significance between two groups is determined by two-tailed unpaired t-test (a). Significance between three groups is determined by one-way ANOVA test (a, c, d). Data are mean ± s.e.m.
Extended Data Fig. 2 Characterization of pathological Tau in mouse model.
a. Schematic drawing of tau acceleration model, created in BioRender; Qian, C. https://BioRender.com/3hrufq2(2026). b. Left: representative images of staining of various phospho-Tau epitope on brain sections from PS19 mice treated with mutant Tau. Selected areas are shown magnified to the left of each image. Scale bar: 100 μm. Right: quantification analysis of the percent of each phospho-Tau epitope in AT8-positive cells. n = 10 mice. 13w: 13-week-old. c. Left: representative images of staining of various phospho-Tau epitope on brain sections from 9-month-old (9 M) PS19 mice. Selected areas are shown magnified to the left of each image. Scale bar: 50 μm. Right: quantification analysis of the percent of each phospho-Tau epitope in AT8-positive cells. n = 10 mice. d. Upper: representative images of X34 and AT8 staining on brain sections from PS19 mice treated with mutant Tau. Scale bar: 50 μm. Lower: quantification analysis of the percent of X34 in AT8-positive cells. n = 10 mice. e. Upper: representative images of X34 and AT8 staining on brain sections from 9-month-old PS19 mice. Scale bar: 50 μm. Lower: Quantification analysis of the percent of X34 in AT8-positive cells. n = 10 mice. f. Upper: representative images of pMLKL and NeuN staining on brain sections from PS19 mice treated with mutant Tau. Scale bar: 10 μm. Lower: quantification analysis of the percent of pMLKL-positive neurons. n = 10 mice. g. Left: representative images of IBA1 and GFAP staining on brain sections from PS19 mice treated with mutant Tau. Scale bar: 300 μm. Selected areas are shown at higher magnification. Right: quantification of relative intensity of IBA1 and GFAP. Sham PS19: n = 8 mice, Tau injected PS19: n = 10 mice. Significance between two groups is determined by two-tailed unpaired t-test (f, g). Data are mean ± s.e.m.
Extended Data Fig. 3 Tau aggregate induces ZBP1-dependent neuronal necroptosis.
a. Left: representative images of Tuj1, IBA1 and GFAP staining and bright field of primary cortex neurons cultured in vitro. Scale bar: 20 μm. Right: quantification analysis of percent of Tuj1, IBA1 and GFAP positive cells. n = 3 independent experiments. b. Left: representative images of X34, PicoGreen staining and bright field of primary cortex neurons cultured in vitro. Scale bar: 20 µm. Right: quantification analysis of the percentage of X34 nuclear positive cells. n = 3 independent experiments. c. Left: representative images of Tuj1, IBA1 and GFAP staining and bright filed of primary DRG neurons cultured in vitro. Scale bar: 20 μm. Right: quantification analysis of percent of Tuj1, IBA1 and GFAP positive cells. n = 3 independent experiments. d. Left: representative images of pMLKL, Tuj1, IBA1 and GFAP staining and bright field of primary PS19 DRG neurons treated with recombinant WT or mutant Tau protein. Scale bar: 20 μm. Right: quantification analysis of pMLKL-positive cells. n = 3 independent experiments. e. Schematic drawing of extracellular signal-driven cell death (left) and intracellular signal-driven cell death (right), created in BioRender; Qian, C. https://BioRender.com/ip4hf6s (2026). f. Left upper: schematic drawing of PS19 DRG neuron culture with treatment of TNFα antibody. Left lower: representative images of pMLKL and AT8 staining and bright filed of primary DRG neurons treated recombinant WT or mutant Tau protein with or without TNF antibody. Scale bar: 20 μm. Right upper: the efficiency of TNF antibody was tested by examining the cell viability of L929 treated with TNFα with or without TNF antibody. Right lower: quantification of pMLKL-positive cells treated with TNF antibody or not was performed. n = 3 independent experiments. g. Global transcriptome analysis of hippocampus from 9-month-old mice (PS19 versus WT). h. Global transcriptome analysis of ileal crypts (SETDB1 KO versus WT) from published work (Wang et al., Nature, 2020). i. Upper: representative images of ZBP1, NeuN, IBA1 and GFAP staining on brain sections of WT and PS19 mice (9-month-old). Scale bar: 50 μm. Lower: quantification of intensity of ZBP1 in NeuN+, IBA1+ and GFAP+ cells. n = 5 mice. j. Left: Schematic drawing of two isoforms of human ZBP1. Right: Immunoblotting analysis of ZBP1 protein level in H929 cells transduced with Lenti-CRISPR/Cas9-gRNA construct targets targeting ZBP1 or Ctrl. k. Immunoblotting and quantification of ZBP1 protein level in the hippocampal protein samples of AD patients at various stage as indicated. Sample from IFN-treated SH-SY5Y cells was used as a positive control (PC). l. Left: representative images of pMLKL (red) and AT8 (green) staining and bright filed of primary DRG neurons from PS19 and PS19, Zbp1−/− mice cultured in vitro. Scale bar: 20 μm. Right: quantification analysis of percent of pMLKL-positive cells. n = 3 independent experiments. m-o. Representative images and quantification analysis of percent of neuronal death (TUNEL-positive) in Tau acceleration model with indicated genotypes (Sham PS19: n = 5 mice, PS19: n = 10 mice; PS19, Zbp1−/−: n = 7 mice; PS19, Ripk3−/−: n = 6 mice; PS19, Zbp1Za1,2Mut/Za1,2Mut: n = 6 mice). Selected areas are shown magnified to the right of each image. Scale bar: 20 μm. Significance between two groups is determined by two-tailed unpaired t-test (d, i, l). Significance between three or more groups is determined by one-way ANOVA test (a-c, f, k, o). Data are mean ± s.e.m. NS, not significant.
Extended Data Fig. 4 Characterization of pathological Tau in SH-SY5Y (Tau-4R-YFP) cells.
a. Schematic drawing of the tau seeding in SH-SY5Y-Tau 4 R cells (SH-SY5Y stably expressed with Tau-4R-YFP). b. SH-SY5Y-Tau 4 R cells were treated with brain lysates from rTg4510 or WT mice. Upper: representative images of phospho-Tau (S356) signal (red) in tau puncta (green) containing cells. Scale bar: 20 μm. Lower: quantification analysis of percent of S356-positive cells in Tau puncta containing cells. n = 3 independent experiments. c. SH-SY5Y-Tau 4 R cells were treated with brain lysates from rTg4510 or WT mice. Upper: representative images of phospho-Tau (S356) signal (red) with X34 in tau puncta (green) containing cells. Scale bar: 20 μm. Lower: quantification analysis of percent of X34-positive cells in Tau puncta containing cells. n = 3 independent experiments. d. SH-SY5Y-Tau 4 R cells were treated with brain lysates from rTg4510 or WT mice. Upper: representative images of phospho-Tau (S356) signal (red) with Thio-S on tau puncta (green) containing cells. Scale bar: 20 μm. Lower: quantification analysis of percent of Thios-positive cells in Tau puncta containing cells. n = 3 independent experiments. e. SH-SY5Y-Tau 4 R cells were treated with brain lysates from rTg4510 or WT mice. Upper: representative images of pMLKL signal (red) on tau puncta (green) containing cells. Scale bar: 20 μm. Lower: quantification analysis of percent of pMLKL-positive cells in Tau puncta containing cells. n = 3 independent experiments. Significance between two groups is determined by two-tailed unpaired t-test (b-e). Data are mean ± s.e.m.
Extended Data Fig. 5 Tau aggregates produced Z-RNAs from reactivated TEs.
a. dsRNA staining of intestinal organoid from mice with indicated genotypes. Scale bar: 20 μm. b. dsRNA staining of MEF cell transfected with poly(I:C). Scale bar: 20 μm. c. Upper: representative images of dsRNA staining in SY5Y-Tau 4 R cells treated with Tau seeds or control. The dsRNA signal was further confirmed with RNase or DNase treatment. Lower: quantification of dsRNA signal co-localized with Tau puncta. n = 3 independent experiments. d. Upper: dsRNA staining on brain sections from mice with indicated genotypes at 9-month-old. Scale bar: 20 μm. Lower: quantification of dsRNA-positive cells. n = 10 mice per group. e. Upper: representative images of ZNA staining on brain sections from PS19 mice at 9-month-old with RNase, DNase or PBS treatment. Scale bar: 20 μm. Lower: quantification of area of ZNA on brain sections. n = 10 mice per group. f. Representative images of ZNA (with RNase or DNase treatment) and AT8 staining on brain sections from AD patients at indicated stage. Scale bar: 20 μm. g. Upper: working flow of RIP-seq. Lower: Immunoblotting analysis of ZBP1-IP efficiency as indicated. Significance between two groups is determined by two-tailed unpaired t-test (c, d). Significance between three or more groups is determined by one-way ANOVA test (e). Data are mean ± s.e.m.
Extended Data Fig. 6 Pathogenic Tau deforms heterochromatin condensation.
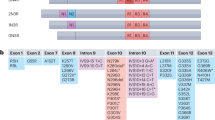
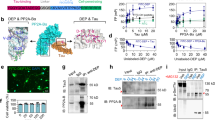
a. Immunoblotting analysis of SETDB1 in hippocampus of WT and PS19 mice (9-month-old). b. Left: representative images of Lamin B1 staining on SH-SY5Y (Tau-4R-YFP) cells treated with Tau seed from rTg4510 or WT mice. Scale bar: 20 μm. Selected areas are shown magnified to the right. Right: quantification of percent of cells with normal/abnormal nuclear lamina in Tau puncta-positive SH-SY5Y (Tau-4R-YFP) cells. n = 3 independent experiments. c. Left: representative images of H3K9me3 (red) and HP1α (green) staining on SH-SY5Y (Tau-4R-YFP) cells treated with Tau seed from rTg4510 or WT mice. Scale bar: 20 μm. Selected areas are shown magnified to the right. Middle: quantification of percent of cells with HP1α and H3K9me3 overlap signal in Tau puncta-positive Tau-4R-YFP cells was performed. Right: quantification of the intensity of H3K9me3 and HP1α signal in tau puncta containing cells. n = 3 independent experiments. d. (Top) Quality assessment by Coomassie blue staining of the recombinant H3K9me3-nucleosomes used in the pull-down assay. (Bottom) Pull-down assay demonstrates a direct interaction between H3K9me3-modified nucleosomes and the reader protein HP1α. e. WT or mutant tau protein was incubated with His-tagged HP1α and H3K9me3-marked nucleosome as indicated. The inputs and anti-HP1α immunoprecipitates were analyzed by immunoblotting as indicated. f. ChIP-qPCR analysis of (Top) H3K9me3 and (Bottom) pTau occupancy at transposable element (TE) loci in the hippocampus of 9-month-old (9 M) PS19 mice. The specificity of pTau antibody was validated in 2-month-old (2 M) PS19 mice, which show no detectable pTau signal at this age. The Actb gene, which is not expected to bind these factors, was included as a negative control. n = 4 mice. g. (Top) Quality assessment by Coomassie blue staining of the recombinant H3K27me3-nucleosomes. (Bottom) Pull-down assay confirms the specific binding of CBX2 to H3K27me3-modified nucleosomes. h. Recombinant mutant tau protein (20 nM) was incubated with His-tagged unmodified, H3K9me3-marked or H3K27me3-marked nucleosome respectively. Inputs and Ni-NTA pull-down samples were analyzed by immunoblotting as indicated. Note: the histone 3 in the unmodified and H3K27me3-marked nucleosome included an additional Flag tag. i. Left: representative images of p-Tau (T181), HP1αand NeuN staining on hippocampus sections of AD patients at various stage as indicated. Scale bar: 20 μm. Selected areas are shown at higher magnification. Right: quantification analysis of intensity of HP1α signal on hippocampus of AD patients. Non-AD: n = 3, Stage I: n = 3, Stage IV + V: n = 5. Significance between two groups is determined by two-tailed unpaired t-test (c). Significance between three or more groups is determined by one-way ANOVA test (i) or two-way ANOVA test (f). Data are mean ± s.e.m.
Extended Data Fig. 7 Zbp1 haploinsufficiency effectively rescues the neurodegenerative phenotypes in PS19 mice.
a. The cognitive function of indicated groups was evaluated by the Y-maze test for measuring spontaneous arm alternation (left) and total arm entries (right) in mice with the indicated genotypes at 10-month-old. WT: n = 10 mice, PS19: n = 10 mice, PS19, Ripk3+/−: n = 16 mice. b. Quantification analysis of the grip strength in mice with indicated genotypes at 10-month-old. WT: n = 10 mice, PS19: n = 10 mice and PS19, Ripk3+/−: n = 16 mice. c. Left: super-resolution images of Synapsin i (red) and PSD95 (green) immunoreactive puncta in the hippocampus of mice with the indicated genotypes at 10-month-old. Scale bar: 5 μm. Right: quantification of SYN+PSD95+ puncta within groups. n = 10 mice. d. Left: Nissl staining on brain sections of mice with indicated genotypes at 10-month-old. Scale bar: 700 μm. Lower: quantification analysis of relative neuron number in CA1 and CA3. n = 10 mice. e. Representative images and quantification of area of CD68, IBA1 and GFAP on brain sections of indicated genotypes at 10-month-old. Scale bar: 300 μm. n = 10 mice. f. Immunoblotting of ZBP1 protein level in hippocampus of PS19 and PS19, Zbp1+/− mice at 10-month-old. n = 3 mice. g. Left: representative images of pMLKL /NeuN staining on hippocampus of mice with indicated genotypes at 10-month-old. Scale bar:10 μm. Right: quantification analysis percent of pMLKL-positive neurons. n = 10 mice. h. Left: representative images of TUNEL staining on hippocampus of indicated genotypes at 10-month-old. Scale bar: 50 μm. Right: quantification analysis of percent of TUNEL-positive cells. n = 10 mice. i. Left: representative images of pMLKL /NeuN staining on spinal cord of mice with indicated genotypes at 10-month-old. Scale bar: 20 μm. Right: quantification analysis of percent of pMLKL-positive neurons and neuron numbers. n = 10 mice. j. Relative mRNA expression of Synaptophysin, PSD95, Slc1a1 and Neurexin 1 in hippocampus of mice with indicated genotypes at 10-month-old. n = 4 mice. k. Heat map analysis of type I interferon-related genes in the hippocampus from mice with indicated genotypes. n = 3 mice l. Heat map analysis of TEs in the hippocampus from mice with indicated genotypes. n = 3 mice. Significance between three or more groups is determined by one-way ANOVA test (a-e, g-j). Data are mean ± s.e.m.
Extended Data Fig. 8 Zbp1 deletion rescues the neurodegenerative phenotypes in PS19 mice.
a. Quantification analysis of body weight of PS19, Zbp1−/− and WT mice at 12-month-old, and PS19 mice at 10-month-old. n = 10 mice. b. The cognitive function evaluated by the Y-maze test for measuring total arm entries (right) and spontaneous arm alternation (left) in PS19, Zbp1−/− and WT mice at 12-month-old, and PS19 mice at 10-month-old. n = 10 mice. c. Quantification analysis of the grip strength in PS19, Zbp1−/− and WT mice at 12-month-old, and PS19 mice at 10-month-old. n = 10 mice. d. LTP recording for continuous 60 min in the hippocampal Schaffer collateral region of mice with indicated genotypes at 12-month-old. Averaged potentiation (mean ± s.e.m.) of baseline normalized fEPSP in the indicated groups was calculated WT: n = 8 slices from 3 mice, PS19: n = 4 slices from 3 mice, PS19, Zbp1−/−: n = 13 slices from 3 mice. Significance between three or more groups is determined by one-way ANOVA test (a-d). Data are mean ± s.e.m.
Extended Data Fig. 9 ZBP1 is not the direct upstream of Tau aggregates.
a. Left: immunoblotting analysis of human Tau in the hippocampus from mice with indicated genotypes at 10-month-old. Right: quantification of human Tau protein level within indicated groups. n = 3 mice. b. Upper: representative images of Tau puncta in SY5Y-Tau 4 R cells treated with brain lysates from rTg4510 or WT mice with or without interferon stimulation (20 ng/ml IFNβ). Scale bar: 20 μm. Lower: quantification of Tau puncta within indicated groups. n = 3 independent experiments. c. Upper: representative images of S356-positive cells treated with brain lysates from rTg4510 or WT mice with or without interferon stimulation (20 ng/ml IFNβ). Scale bar: 20 μm. Lower: quantification of S356-positive cells within indicated groups. n = 3 independent experiments. d. Upper: representative images of Thio-S-positive cells treated with brain lysates from rTg4510 or WT mice with or without interferon stimulation (20 ng/ml IFNβ). Scale bar: 20 μm. Lower: quantification of Thio-S-positive cells within indicated groups. n = 3 independent experiments. e. Upper: representative images of X34-positive cells treated with brain lysates from rTg4510 or WT mice with or without interferon stimulation (20 ng/ml IFNβ). Scale bar: 20 μm. Lower: quantification of X34-positive cells within indicated groups. n = 3 independent experiments. Significance between three or more groups is determined by one-way ANOVA test (a, e). Data are mean ± s.e.m.
Supplementary information
Supplementary Table 1 (download XLSX )
Table for key resources
Source data
Source Data Figs. 1–6 and Extended Data Figs .1–9 (download XLSX )
Statistical source data
Source Data (download PDF )
Unprocessed western blots and/or gels.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Liu, W., Wu, SA., Zhang, BX. et al. Tau aggregates cause reactivation of transposable DNA elements, leading to Z-RNA–ZBP1-mediated neuronal death. Nat Neurosci (2026). https://doi.org/10.1038/s41593-026-02299-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41593-026-02299-9
